Plant Science 224 (2014) 86–94 Contents lists available at ScienceDirect Plant Science j ourna l ho me pa g e: www.elsevier.com/locate/plantsci Phytoalexin biosynthesis genes are regulated and involved in plant response to Ralstonia solanacearum infection Yu-Mei Lin a,1 , Shu-Lein Shih a,1 , Wan-Chi Lin c , Jia-Wei Wu a , Ya-Ting Chen a , Chi-Ying Hsieh a , Li-Ching Guan a , Lu Lin a , Chiu-Ping Cheng a,b,∗ a Graduate Institute of Plant Biology, College of Life Science, National Taiwan University, Taipei 106, Taiwan, ROC b Department of Life Science, College of Life Science, National Taiwan University, Taipei 106, Taiwan, ROC c Institute of Plant and Microbial Biology, Academia Sinica, Taipei, Taiwan a r t i c l e i n f o Article history: Received 24 February 2014 Received in revised form 10 April 2014 Accepted 11 April 2014 Available online 18 April 2014 Keywords: EAS GT-1 PAD TPS PAMP a b s t r a c t Genes encoding phytoalexin biosynthesis enzymes are transcriptionally regulated and are required for defense against fungi and oomycetes. Here, we studied the regulation of tobacco 5-epi-aristolochene synthase 4 (EAS4) promoters on bacterial infection and investigated the roles of tomato and Arabidopsis phytoalexin biosynthesis genes in defense against pathogenic bacteria. Our results showed that the Nico- tiana glutinosa EAS4 (NgEAS4) promoter was significantly induced by treatments with several bacteria in treated and systemic leaves of transgenic plants. This promoter was also partially induced by treatments with type-III secretion-deficient mutants and total lysate of R. solanacearum (Rs), revealing that both bacterial pathogen-associated molecular patterns (PAMPs) and effectors are involved in R. solanacearum- induced local and systemic activation of the NgEAS4 promoter. Furthermore, the absence of a cis-element GT-1 box in the NgEAS4 promoter abolished systemic activation in non-treated leaves, whereas disrup- tion of the GT-1 box of the N. tabacum EAS4 (NtEAS4) promoter led to constitutive expression. Moreover, silencing of tomato sesquiterpene synthase genes and disruption of the key gene PAD3 for Arabidop- sis camalexin biosynthesis resulted in decreased tolerance to R. solanacearum. These results together, reveal the varied function of GT-1 boxes in regulating tobacco EAS4 promoters and the involvement of phytoalexin-biosythesis genes in plant defense against R. solanacearum. © 2014 Elsevier Ireland Ltd. All rights reserved. Abbreviations: BW, bacterial wilt; DIG, digoxigenin; DPI, days post inoc- ulation; EAS, 5-epi-aristolochene synthases; EF1a, ELONGATION FACTOR 1a; ET, ethylene; ETI, effector-triggered immunity; GFP, green fluorescence protein; GluA, class II acidic extracellular ˇ-1,3-GLUCANASE; GluB, class I basic intracellular ˇ-1,3- GLUCANASE; GUS, -glucuronidase; H7996, Hawaii7996; H2O2, hydrogen peroxide; MAMPs, microbe-associated molecular patterns; 4-MUG, 4-methylumbelliferyl - D-glucuronide; pad3, phytoalexin deficient 3; PAMPs, pathogen-associated molecular patterns; Pcc, Pectobacterium crotovorum subsp. crotovorum; PR, pathogenesis-related gene; PTI, PAMP-triggered immunity; ROS, reactive oxygen species; Rs, Ralstonia solanacearum; SA, salicylic acid; SD, standard deviation; SE, standard error; T3SS, type III secretion system; TPS, terpene synthase; TRV, Tobacco rattle virus; VIGS, virus-induced gene silencing; Xeu, Xanthomonas euvesicatoria; X-gluc, 5-bromo-4- chloro-3-indolyl -D-glucuronic acid. ∗ Corresponding author at: Graduate Institute of Plant Biology, College of Life Science, National Taiwan University, Taipei 106, Taiwan, ROC. Tel.: +886 2 3366 3793; fax: +886 2 2391 8940. E-mail addresses: [email protected], [email protected] (C.-P. Cheng). 1 These authors contributed equally to this work. Introduction Plants have developed subtle pre-existing and induced defense mechanisms to fend off pest and pathogen attacks. The inducible defense mechanisms mainly consist of two intercon- nected branches: pathogen/microbe-associated molecular patterns (PAMPs or MAMPs) triggered immunity (PTI), a conserved innate defense response, and effector-triggered immunity (ETI), an evolved defense response induced by recognizing specific pathogen effector proteins [1,2]. Both PTI and ETI activate certain common defense networks mediated by hormones, such as salicylic acid (SA) and ethylene (ET), reactive oxygen species (ROS), kinases and transcription factors [3–6]. In addition, through the signaling path- ways mediated by hormones, plants can also provoke systemic acquired resistance and induced systemic resistance to achieve a long-distance and wide-spectrum defense responses against dif- ferent pathogens [7,8]. Phytoalexins are low-molecular-weight antimicrobial com- pounds that accumulate in plants in response to pathogen infection or abiotic stress [9]. Induction of these compounds is important http://dx.doi.org/10.1016/j.plantsci.2014.04.008 0168-9452/© 2014 Elsevier Ireland Ltd. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pr
YCa
b
c
a
ARRAA
KEGPTP
uecGMDpgstvc
ST
(
h0
Plant Science 224 (2014) 86–94
Contents lists available at ScienceDirect
Plant Science
j ourna l ho me pa g e: www.elsev ier .com/ locate /p lantsc i
hytoalexin biosynthesis genes are regulated and involved in plantesponse to Ralstonia solanacearum infection
u-Mei Lina,1, Shu-Lein Shiha,1, Wan-Chi Linc, Jia-Wei Wua, Ya-Ting Chena,hi-Ying Hsieha, Li-Ching Guana, Lu Lina, Chiu-Ping Chenga,b,∗
Graduate Institute of Plant Biology, College of Life Science, National Taiwan University, Taipei 106, Taiwan, ROCDepartment of Life Science, College of Life Science, National Taiwan University, Taipei 106, Taiwan, ROCInstitute of Plant and Microbial Biology, Academia Sinica, Taipei, Taiwan
r t i c l e i n f o
rticle history:eceived 24 February 2014eceived in revised form 10 April 2014ccepted 11 April 2014vailable online 18 April 2014
eywords:AST-1ADPS
a b s t r a c t
Genes encoding phytoalexin biosynthesis enzymes are transcriptionally regulated and are required fordefense against fungi and oomycetes. Here, we studied the regulation of tobacco 5-epi-aristolochenesynthase 4 (EAS4) promoters on bacterial infection and investigated the roles of tomato and Arabidopsisphytoalexin biosynthesis genes in defense against pathogenic bacteria. Our results showed that the Nico-tiana glutinosa EAS4 (NgEAS4) promoter was significantly induced by treatments with several bacteria intreated and systemic leaves of transgenic plants. This promoter was also partially induced by treatmentswith type-III secretion-deficient mutants and total lysate of R. solanacearum (Rs), revealing that bothbacterial pathogen-associated molecular patterns (PAMPs) and effectors are involved in R. solanacearum-induced local and systemic activation of the NgEAS4 promoter. Furthermore, the absence of a cis-elementGT-1 box in the NgEAS4 promoter abolished systemic activation in non-treated leaves, whereas disrup-
AMP tion of the GT-1 box of the N. tabacum EAS4 (NtEAS4) promoter led to constitutive expression. Moreover,silencing of tomato sesquiterpene synthase genes and disruption of the key gene PAD3 for Arabidop-sis camalexin biosynthesis resulted in decreased tolerance to R. solanacearum. These results together,reveal the varied function of GT-1 boxes in regulating tobacco EAS4 promoters and the involvement ofphytoalexin-biosythesis genes in plant defense against R. solanacearum.
© 2014 Elsevier Ireland Ltd. All rights reserved.
Abbreviations: BW, bacterial wilt; DIG, digoxigenin; DPI, days post inoc-lation; EAS, 5-epi-aristolochene synthases; EF1a, ELONGATION FACTOR 1a; ET,thylene; ETI, effector-triggered immunity; GFP, green fluorescence protein; GluA,lass II acidic extracellular ˇ-1,3-GLUCANASE; GluB, class I basic intracellular ˇ-1,3-LUCANASE; GUS, �-glucuronidase; H7996, Hawaii7996; H2O2, hydrogen peroxide;AMPs, microbe-associated molecular patterns; 4-MUG, 4-methylumbelliferyl �--glucuronide; pad3, phytoalexin deficient 3; PAMPs, pathogen-associated molecularatterns; Pcc, Pectobacterium crotovorum subsp. crotovorum; PR, pathogenesis-relatedene; PTI, PAMP-triggered immunity; ROS, reactive oxygen species; Rs, Ralstoniaolanacearum; SA, salicylic acid; SD, standard deviation; SE, standard error; T3SS,ype III secretion system; TPS, terpene synthase; TRV, Tobacco rattle virus; VIGS,irus-induced gene silencing; Xeu, Xanthomonas euvesicatoria; X-gluc, 5-bromo-4-hloro-3-indolyl �-D-glucuronic acid.∗ Corresponding author at: Graduate Institute of Plant Biology, College of Lifecience, National Taiwan University, Taipei 106, Taiwan, ROC.el.: +886 2 3366 3793; fax: +886 2 2391 8940.
E-mail addresses: [email protected], [email protected]. Cheng).
1 These authors contributed equally to this work.
ttp://dx.doi.org/10.1016/j.plantsci.2014.04.008168-9452/© 2014 Elsevier Ireland Ltd. All rights reserved.
Introduction
Plants have developed subtle pre-existing and induceddefense mechanisms to fend off pest and pathogen attacks. Theinducible defense mechanisms mainly consist of two intercon-nected branches: pathogen/microbe-associated molecular patterns(PAMPs or MAMPs) triggered immunity (PTI), a conserved innatedefense response, and effector-triggered immunity (ETI), anevolved defense response induced by recognizing specific pathogeneffector proteins [1,2]. Both PTI and ETI activate certain commondefense networks mediated by hormones, such as salicylic acid(SA) and ethylene (ET), reactive oxygen species (ROS), kinases andtranscription factors [3–6]. In addition, through the signaling path-ways mediated by hormones, plants can also provoke systemicacquired resistance and induced systemic resistance to achieve along-distance and wide-spectrum defense responses against dif-
ferent pathogens [7,8].Phytoalexins are low-molecular-weight antimicrobial com-pounds that accumulate in plants in response to pathogen infectionor abiotic stress [9]. Induction of these compounds is important

Scien
fbcfAtfpdg
pipcEp(aEP[gtsi
pfIs(eibitisR
M
P
Agcp3bsvXt5pflbPp(p
Y.-M. Lin et al. / Plant
or establishing disease resistance to pathogens and pests and cane regulated by plant hormones, mitogen-activated protein kinaseascades and transcription factors. Unfortunately, reports mostlyocus on the regulation and function of phytoalexin biosynthesis inrabidopsis thaliana [9,10]. It is currently assumed that camalexin,he main phytoalexin of Arabidopsis, is effective against pathogenicungi and oomycetes but not against bacteria [10–12]. Althoughhytoalexins are suggested to be involved in bean and potatoefense response to the infection of pathogenic bacteria [13,14],enetic evidence is still lacking.
Capsidiol is the major sesquiterpenoid phytoalexin produced inepper (Capsicum annuum) and tobacco (Nicotiana tabacum) and
ts accumulation can be induced by fungi, oomycetes and variousathogen-derived elicitors [9]. Capsidiol biosynthesis involves aommitted step triggered by 5-epi-aristolochene synthases (EAS,C 4.2.3.61), which are encoded by a gene family and regulatedrimarily at the transcriptional level [15–17]. Previously, TEAS4NtEAS4) was demonstrated to be pathogen- and elicitor-induciblet, or near, the elicited sites [18,19]. Expression of a chili pepperAS gene (PEAS1) can be induced by fungal PAMPs and avirulentseudomonas syringae pv. syringae in both local and systemic leaves20]. Furthermore, ROS is suggested to mediate the induction of EASene expression and capsidiol accumulation [18,21,22]. However,he key cis-elements of the TEAS4 and PEAS1 promoters for theirtress responsiveness and systemic inducibility have not yet beennvestigated.
This study aimed to investigate the regulation of tobacco EAS4romoters in response to bacterial infection and to determine theunctionality of their cis-element GT-1 boxes in this regulation.n addition, we investigated the roles of phytoalexin biosynthe-is genes in tomato and Arabidopsis tolerance to R. solanacearumRs), a soil-borne vascular bacterium that causes lethal wilt dis-ases of a wide range of crops worldwide. Our results reveal thenducibility of the NgEAS4 promoter by several phytopathogenicacteria and the involvement of both bacterial PAMPs and effectors
n R. solanacearum-induced activation of this promoter. In addi-ion, a previously unreported role of the GT-1 box in the systemicnducibility of the NgEAS4 promoter is disclosed. Furthermore, ourtudy points to a clear role of phytoalexins in plant defense against. solanacearum.
aterials and methods
lant materials, growth conditions and stress treatments
The wild-type and phytoalexin deficient 3 (pad3-1) mutant ofrabidopsis thaliana ecotype Col-0 were grown for four weeks inrowth chambers at 22 ◦C under a 16 h-light/8 h-dark cycle. Solana-eous plants were grown in growth chambers under a 12-h lighthotoperiod at 25 ◦C. Transgenic N. tabacum cultivar Wisconsin8 (W38) plants were generated as described previously [23] (seeelow). The pathogens used in this study included R. solanacearumtrain Pss190 (phylotype I, biovar 4) and Pss4 (phylotype I, bio-ar 3), Pectobacterium crotovorum subsp. Crotovorum (Pcc) andanthomonas euvesicatoria (Xeu). For the preparation of bacterialotal lysates, an overnight culture of R. solanacearum grown in23 liquid medium [24] was plated on 523 agar plates and thelates were incubated overnight at 28 ◦C. The bacteria were washedrom the plates with sterile water and adjusted to OD600 = 10, fol-owed by boiling for 10 min. The content of total proteins in theacterial total lysates was then determined using the Pierce BAC
rotein Assay Kit (Thermo scientific, USA) and the lysate sam-les were prepared to the indicated concentration in MES bufferpH 5.6). Bacteria (OD600 = 0.3), bacterial total lysates (3 �g totalrotein/�L) or flagellin-related synthetic peptide (Flg22) (10 �M)ce 224 (2014) 86–94 87
were infiltrated into leaves of transgenic plants or used for feedingexperiments on detached leaves or seedling roots.
Transcriptional analyses
For northern blot analysis, at least three leaves from each of atleast three plants were used to collect leaf discs (1 cm in diame-ter) for stress treatments. Leaf discs were floated on 10 mM MESbuffer containing H2O2 (10 mM) or R. solanacearum suspensions(OD600 = 0.3) in Petri dishes at room temperature for 24 h, or floatedon water containing 150 mM sodium chloride (NaCl) for 3 h. Foreach treatment in every trial, eight leaf discs were combined forRNA extraction using TRIzol® Reagent (Invitrogen, USA) or theRNeasy Plant Mini Kit (Qiagen, Germany), followed by a treatmentwith RNase-free DNase I (Promega, USA). Northern blot analy-sis was performed as described previously with high-stringencywashes in 0.5× SSC containing 0.1% SDS at 68 ◦C [25]. The NtEAS4probe was prepared from N. tabacum cDNA using the primerslisted in Table S1 and was labeled with digoxigenin (DIG) (Roche,Germany).
For real-time RT-PCR assays, at least four leaves were collectedfrom two plants for each treatment and were then combinedfor RNA extraction. First-strand cDNA was synthesized from 1 �gtotal RNA with oligo (dT) primer and AMV reverse transcriptase(Promega), followed by PCR reaction with the gene-specific primerslisted in Table S1 using the KAPA SYBR® FAST qPCR Kit (KAPABiosystems, USA) on a Bio-Rad-Real-Time PCR Detection System(BIO-RAD MyiQTM). ELONGATION FACTOR 1a (EF1a) of the ana-lyzed plant species or cultivars was used as the internal control fornormalization. The data were analyzed by Bio-Rad iQTM5 Opticalsystem version 2.0.
Cloning of promoters and construction of expression vectors
A 634-bp fragment of the NtEAS4 promoter was isolated fromN. tabacum W38 by PCR with the NtEAS4-promoter primers (TableS1). The PCR product was digested with KpnI and XmaI andthen cloned into the pBSks+-NOS vector, a kind gift of Dr. Ming-Tsair Chan (Agricultural Biotechnology Research Center, AcademiaSinica, Taipei, Taiwan). In vitro site-directed mutagenesis was car-ried out by recombinant PCR using the QuikChangeTM Site-DirectedMutagenesis Kit (Stratagene, USA) with specific mutagenesisoligonucleotide primers (NtEAS4-OZZ, NtEAS4-XOX, NtEAS4-ZXO;Table S1). The mutated NtEAS4 promoter fragments were thenamplified using NtEAS4-promoter primers, digested with KpnI andNcoI and ligated into the pCAMBIA1301 vector to replace thecauliflower mosaic virus (CaMV) 35S promoter. The resultant con-structs were then transferred into Agrobacterium tumefaciens strainLBA4404 and the transformed bacteria were used for agroinfiltra-tion of N. tabacum (cv. W38) leaves as described previously [26].Vector pCAMBIA1381Z and pCAMBIA1301 were used as negativeand positive controls, respectively.
For the NgEAS4 promoter, the 5′-flanking region of NgEAS4 wasisolated using the Universal GenomeWalkerTM Kit (Clontech Labo-ratories, CA). Genomic DNA isolated from N. glutinosa was digestedwith five restriction enzymes (DraI, EcoRV, PvuII, ScaI, and StuI,respectively) to create blunt-end fragments that were then lig-ated to specific adaptors to produce five respective GenomeWalkerlibraries. The NgEAS4-specific primers NgEAS4-GST1 and NgEAS4-GST2 (Table S1) were used for the PCR reactions and the majorPCR products were purified and cloned into pCR®II-TOPO® vector(Invitrogen) for sequencing. Fragments of NgEAS4 promoter carry-
ing different numbers of GT-1 boxes were obtained by PCR usingprimers listed in Table S1. The PCR products were cloned into thepCR®8/GW/TOPO® vector (Invitrogen) and then into the pHGWFS7vector (Invitrogen). The resultant constructs were then transferred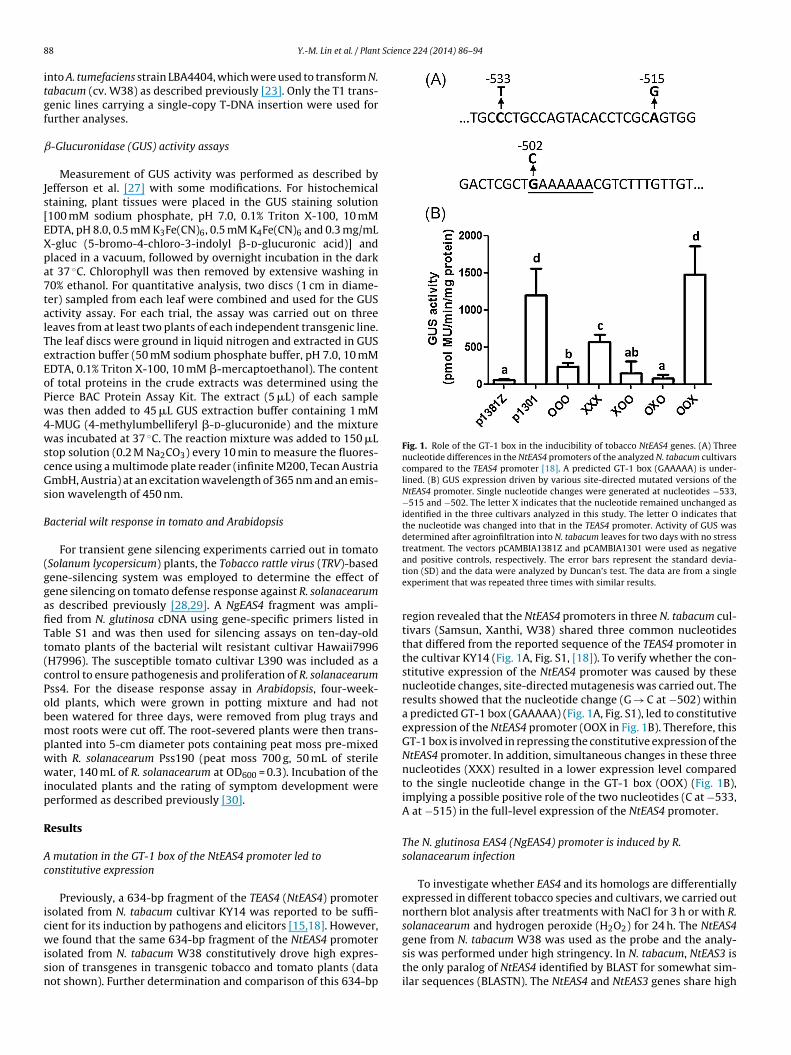
8 Science 224 (2014) 86–94
itgf
ˇ
Js[EXpa7talTeEoPw4wscGs
B
(ggafiTt(cPobmpwwip
R
Ac
icwisn
Fig. 1. Role of the GT-1 box in the inducibility of tobacco NtEAS4 genes. (A) Threenucleotide differences in the NtEAS4 promoters of the analyzed N. tabacum cultivarscompared to the TEAS4 promoter [18]. A predicted GT-1 box (GAAAAA) is under-lined. (B) GUS expression driven by various site-directed mutated versions of theNtEAS4 promoter. Single nucleotide changes were generated at nucleotides −533,−515 and −502. The letter X indicates that the nucleotide remained unchanged asidentified in the three cultivars analyzed in this study. The letter O indicates thatthe nucleotide was changed into that in the TEAS4 promoter. Activity of GUS wasdetermined after agroinfiltration into N. tabacum leaves for two days with no stresstreatment. The vectors pCAMBIA1381Z and pCAMBIA1301 were used as negative
8 Y.-M. Lin et al. / Plant
nto A. tumefaciens strain LBA4404, which were used to transform N.abacum (cv. W38) as described previously [23]. Only the T1 trans-enic lines carrying a single-copy T-DNA insertion were used forurther analyses.
-Glucuronidase (GUS) activity assays
Measurement of GUS activity was performed as described byefferson et al. [27] with some modifications. For histochemicaltaining, plant tissues were placed in the GUS staining solution100 mM sodium phosphate, pH 7.0, 0.1% Triton X-100, 10 mMDTA, pH 8.0, 0.5 mM K3Fe(CN)6, 0.5 mM K4Fe(CN)6 and 0.3 mg/mL-gluc (5-bromo-4-chloro-3-indolyl �-d-glucuronic acid)] andlaced in a vacuum, followed by overnight incubation in the darkt 37 ◦C. Chlorophyll was then removed by extensive washing in0% ethanol. For quantitative analysis, two discs (1 cm in diame-er) sampled from each leaf were combined and used for the GUSctivity assay. For each trial, the assay was carried out on threeeaves from at least two plants of each independent transgenic line.he leaf discs were ground in liquid nitrogen and extracted in GUSxtraction buffer (50 mM sodium phosphate buffer, pH 7.0, 10 mMDTA, 0.1% Triton X-100, 10 mM �-mercaptoethanol). The contentf total proteins in the crude extracts was determined using theierce BAC Protein Assay Kit. The extract (5 �L) of each sampleas then added to 45 �L GUS extraction buffer containing 1 mM
-MUG (4-methylumbelliferyl �-d-glucuronide) and the mixtureas incubated at 37 ◦C. The reaction mixture was added to 150 �L
top solution (0.2 M Na2CO3) every 10 min to measure the fluores-ence using a multimode plate reader (infinite M200, Tecan AustriambH, Austria) at an excitation wavelength of 365 nm and an emis-ion wavelength of 450 nm.
acterial wilt response in tomato and Arabidopsis
For transient gene silencing experiments carried out in tomatoSolanum lycopersicum) plants, the Tobacco rattle virus (TRV)-basedene-silencing system was employed to determine the effect ofene silencing on tomato defense response against R. solanacearums described previously [28,29]. A NgEAS4 fragment was ampli-ed from N. glutinosa cDNA using gene-specific primers listed inable S1 and was then used for silencing assays on ten-day-oldomato plants of the bacterial wilt resistant cultivar Hawaii7996H7996). The susceptible tomato cultivar L390 was included as aontrol to ensure pathogenesis and proliferation of R. solanacearumss4. For the disease response assay in Arabidopsis, four-week-ld plants, which were grown in potting mixture and had noteen watered for three days, were removed from plug trays andost roots were cut off. The root-severed plants were then trans-
lanted into 5-cm diameter pots containing peat moss pre-mixedith R. solanacearum Pss190 (peat moss 700 g, 50 mL of sterileater, 140 mL of R. solanacearum at OD600 = 0.3). Incubation of the
noculated plants and the rating of symptom development wereerformed as described previously [30].
esults
mutation in the GT-1 box of the NtEAS4 promoter led toonstitutive expression
Previously, a 634-bp fragment of the TEAS4 (NtEAS4) promotersolated from N. tabacum cultivar KY14 was reported to be suffi-ient for its induction by pathogens and elicitors [15,18]. However,
e found that the same 634-bp fragment of the NtEAS4 promotersolated from N. tabacum W38 constitutively drove high expres-ion of transgenes in transgenic tobacco and tomato plants (dataot shown). Further determination and comparison of this 634-bp
and positive controls, respectively. The error bars represent the standard devia-tion (SD) and the data were analyzed by Duncan’s test. The data are from a singleexperiment that was repeated three times with similar results.
region revealed that the NtEAS4 promoters in three N. tabacum cul-tivars (Samsun, Xanthi, W38) shared three common nucleotidesthat differed from the reported sequence of the TEAS4 promoter inthe cultivar KY14 (Fig. 1A, Fig. S1, [18]). To verify whether the con-stitutive expression of the NtEAS4 promoter was caused by thesenucleotide changes, site-directed mutagenesis was carried out. Theresults showed that the nucleotide change (G → C at −502) withina predicted GT-1 box (GAAAAA) (Fig. 1A, Fig. S1), led to constitutiveexpression of the NtEAS4 promoter (OOX in Fig. 1B). Therefore, thisGT-1 box is involved in repressing the constitutive expression of theNtEAS4 promoter. In addition, simultaneous changes in these threenucleotides (XXX) resulted in a lower expression level comparedto the single nucleotide change in the GT-1 box (OOX) (Fig. 1B),implying a possible positive role of the two nucleotides (C at −533,A at −515) in the full-level expression of the NtEAS4 promoter.
The N. glutinosa EAS4 (NgEAS4) promoter is induced by R.solanacearum infection
To investigate whether EAS4 and its homologs are differentiallyexpressed in different tobacco species and cultivars, we carried outnorthern blot analysis after treatments with NaCl for 3 h or with R.solanacearum and hydrogen peroxide (H2O2) for 24 h. The NtEAS4
gene from N. tabacum W38 was used as the probe and the analy-sis was performed under high stringency. In N. tabacum, NtEAS3 isthe only paralog of NtEAS4 identified by BLAST for somewhat sim-ilar sequences (BLASTN). The NtEAS4 and NtEAS3 genes share high
Y.-M. Lin et al. / Plant Scien
Fig. 2. Inducibility of the NgEAS4 promoter by R. solanacearum infection in trans-genic plants. (A) Schematic structure of pNg1011. In this construct, a NgEAS4fragment containing the 28-bp coding sequence and the 983-bp upstream sequencewas used to drive the expression of the GFP-GUS fusion. The PLACE (http://www.dna.affrc.go.jp/PLACE/) and PlantCare databases (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) were used to identify possible cis-elements in the NgEAS4promoter. (B) Assay for GUS activity in leaves of three independent transgenic N.tabacum lines (T1 generation) after infiltration with R. solanacearum (OD600 = 0.3) for24 h. Vector represents transgenic plants containing the empty vector (pHGWFS7).The error bars indicate the SD of the mean of three plants. Pair-wise comparisonswts
sccBtSaictctcw
iDtol2ceo(mptt(stp
ere made between the water- and bacterium-treated samples using the Student’s-test. An asterisk represents significant difference (p < 0.05). The data are from aingle experiment that was repeated three times with similar results.
equence homology, with an identity of 96% in the protein-codingDNA sequences and 90% at the amino acid level; however, their 3′
DNA and N-terminal protein sequences are dissimilar (Fig. S2A and). In contrast, NtEAS4 and NgEAS4 share high sequence homology inheir 5′ protein-coding regions (cDNA sequence identity = 98%) (Fig.2C). Therefore, the use of NtEAS4 as a probe could detect NtEAS3nd NgEAS4 even under high stringency. The results showed thatn three N. tabacum cultivars (Samsun, Xanthi, W38) under controlonditions, EAS4 and its paralog(s) together displayed a constitu-ive expression pattern and the expression levels varied in theseultivars (Fig. S3). In contrast, the overall expression of NgEAS4,ogether with other possible paralog(s), remained very low underontrol conditions but was significantly enhanced by treatmentsith R. solanacearum and H2O2 (Fig. S3).
To study further NgEAS4 regulation in response to bacterialnfection, a 1011-bp NgEAS4 fragment was isolated using genomicNA walking (see Materials and Methods for details). Part of
he obtained sequence matched the 5′-terminal cDNA sequencef the NgEAS4-coding region (Fig. S2C), confirming that the iso-ated sequence was from NgEAS4. This fragment contains the8-bp coding sequence and the 983-bp upstream sequence, whichontains two GT-1 boxes and one W box (Fig. S4). A reporterxpression cassette (pNg1011) was prepared for the productionf a reporter fusion composed of the green fluorescence proteinGFP) and GUS under the control of the 1011-bp NgEAS4 pro-
oter (Fig. 2A). Transgenic N. tabacum W38 plants containingNg1011 were generated and were used to study the features ofhe NgEAS4 promoter. The results showed that GUS activity in theransgenic plants containing the empty vector was largely very low
0.9469 ± 0.3917 pmol MU/min/mg protein for the water-treatedamples and was 1.1969 ± 0.6781 pmol MU/min/mg protein forhe bacterium-treated samples). GUS activity in independentNg1011-transgenic lines was detected at low levels after waterce 224 (2014) 86–94 89
treatment but was clearly enhanced by R. solanacearum inocula-tion (Fig. 2B), indicating the inducibility of the NgEAS4 promoter bythis bacterium.
To investigate the tissue specificity of the NgEAS4 promoter, spa-tial GFP and GUS expression patterns were examined in leaves androots of pNg1011-transgenic plants. The results revealed that theNgEAS4 promoter was active in leaf veins (Fig. 3) and roots (Fig. S5)upon R. solanacearum inoculation.
The NgEAS4 promoter is induced by treatments withphytopathogenic bacteria and PAMPs
Further analysis was carried out to determine whether theNgEAS4 promoter could be induced by other phytopathogenicbacteria and bacterial PAMPs. As shown in Fig. 4. GUS activityin pNg1011-transgenic plants was also significantly induced byinfection by P. crotovorum subsp. crotovorum, X. euvesicatoria andanother R. solanacearum strain (Pss190). In addition, type III secre-tion system (T3SS)-defective hrpG::Tn5 mutants of R. solanacearum[30] were used to test whether the NgEAS4 promoter could beinduced by bacterial PAMPs and therefore, whether T3SS-secrectedeffectors have an effect on NgEAS4 promoter induction. The resultsshowed that the hrpG::Tn5 mutants of R. solanacearum strain Pss4and Pss190 could partially induce GUS expression in pNg1011-transgenic plants compared to the wild-type strains (Fig. 4).However, the treatment of Flg22, which is a synthetic peptide of22 amino acids representing a conserved PAMP present in flag-ellins of a wide range of bacteria, only induced a very low levelof GUS expression (Fig. 3). Therefore, the NgEAS4 promoter can beinduced by the infection of different phytopathogenic bacteria andboth bacterial PAMPs and effectors are required for the full level ofR. solanacearum-induced NgEAS4 promoter activation.
A GT-1 box is required for systemic inducibility of the NgEAS4promoter
To determine whether the GT-1 boxes were involved in reg-ulating NgEAS4 expression, we generated transgenic N. tabacumW38 plants carrying GFP-GUS expression cassettes under the con-trol of the NgEAS4 promoter fragments containing one (pNg727)or no GT-1 box (pNg709) (Fig. 5A) and analyzed their GUS expres-sion patterns. In addition to the hrpG::Tn5 mutants, lysates of R.solanacearum were also used to determine the effect of bacterialPAMPs [24]. The results showed that the GUS induction patternsin pNg727- and pNg709-transgenic plants after treatment with R.solanacearum wild-type strain, hrpG::Tn5 mutant and lysate werenot significantly different from those in pNg1011-transgenic plants(Fig. 5B). In addition, R. solanacearum lysate triggered a partialinduction of the promoter activity similar to that in the hrpG::Tn5mutant. These results reveal that GT-1 boxes do not play a sig-nificant role in the regulation of NgEAS4 promoter activation inresponse to R. solanacearum infection.
Next, we asked whether the NgEAS4 promoter could be syste-mically induced in non-treated tissues by monitoring GUS activityin systemic leaves immediately adjacent to the treated ones. Theresults showed that treatments with R. solanacearum and its lysatecould both lead to the clear activation of NgEAS4 promoter inthe treated and the non-treated systemic leaves of pNg1011- andpNg727-transgenic plants (Fig. 5C). However, GUS activity in sys-temic leaves of pNg709-transgenic plants after infiltration withR. solanacearum or its lysate was constantly lower than 10 pmol
MU/min/mg protein) (Fig. 5C), revealing that a similar systemicinduction was not effective in pNg709-transgenic plants. Therefore,the GT-1 box located at −697 is required for the systemic inductionof the NgEAS4 promoter.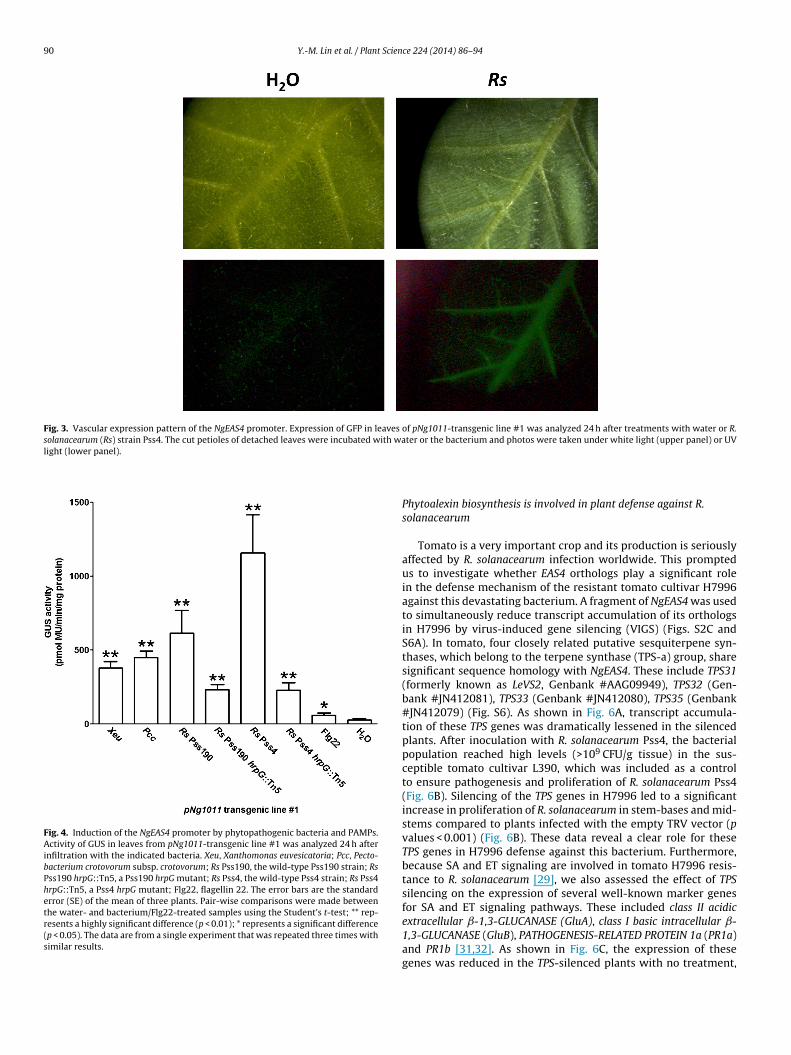
90 Y.-M. Lin et al. / Plant Science 224 (2014) 86–94
Fig. 3. Vascular expression pattern of the NgEAS4 promoter. Expression of GFP in leaves of pNg1011-transgenic line #1 was analyzed 24 h after treatments with water or R.solanacearum (Rs) strain Pss4. The cut petioles of detached leaves were incubated with water or the bacterium and photos were taken under white light (upper panel) or UVlight (lower panel).
Fig. 4. Induction of the NgEAS4 promoter by phytopathogenic bacteria and PAMPs.Activity of GUS in leaves from pNg1011-transgenic line #1 was analyzed 24 h afterinfiltration with the indicated bacteria. Xeu, Xanthomonas euvesicatoria; Pcc, Pecto-bacterium crotovorum subsp. crotovorum; Rs Pss190, the wild-type Pss190 strain; RsPss190 hrpG::Tn5, a Pss190 hrpG mutant; Rs Pss4, the wild-type Pss4 strain; Rs Pss4hrpG::Tn5, a Pss4 hrpG mutant; Flg22, flagellin 22. The error bars are the standarderror (SE) of the mean of three plants. Pair-wise comparisons were made betweenthe water- and bacterium/Flg22-treated samples using the Student’s t-test; ** rep-resents a highly significant difference (p < 0.01); * represents a significant difference(p < 0.05). The data are from a single experiment that was repeated three times withsimilar results.
Phytoalexin biosynthesis is involved in plant defense against R.solanacearum
Tomato is a very important crop and its production is seriouslyaffected by R. solanacearum infection worldwide. This promptedus to investigate whether EAS4 orthologs play a significant rolein the defense mechanism of the resistant tomato cultivar H7996against this devastating bacterium. A fragment of NgEAS4 was usedto simultaneously reduce transcript accumulation of its orthologsin H7996 by virus-induced gene silencing (VIGS) (Figs. S2C andS6A). In tomato, four closely related putative sesquiterpene syn-thases, which belong to the terpene synthase (TPS-a) group, sharesignificant sequence homology with NgEAS4. These include TPS31(formerly known as LeVS2, Genbank #AAG09949), TPS32 (Gen-bank #JN412081), TPS33 (Genbank #JN412080), TPS35 (Genbank#JN412079) (Fig. S6). As shown in Fig. 6A, transcript accumula-tion of these TPS genes was dramatically lessened in the silencedplants. After inoculation with R. solanacearum Pss4, the bacterialpopulation reached high levels (>109 CFU/g tissue) in the sus-ceptible tomato cultivar L390, which was included as a controlto ensure pathogenesis and proliferation of R. solanacearum Pss4(Fig. 6B). Silencing of the TPS genes in H7996 led to a significantincrease in proliferation of R. solanacearum in stem-bases and mid-stems compared to plants infected with the empty TRV vector (pvalues < 0.001) (Fig. 6B). These data reveal a clear role for theseTPS genes in H7996 defense against this bacterium. Furthermore,because SA and ET signaling are involved in tomato H7996 resis-tance to R. solanacearum [29], we also assessed the effect of TPSsilencing on the expression of several well-known marker genesfor SA and ET signaling pathways. These included class II acidicextracellular ˇ-1,3-GLUCANASE (GluA), class I basic intracellular ˇ-
1,3-GLUCANASE (GluB), PATHOGENESIS-RELATED PROTEIN 1a (PR1a)and PR1b [31,32]. As shown in Fig. 6C, the expression of thesegenes was reduced in the TPS-silenced plants with no treatment,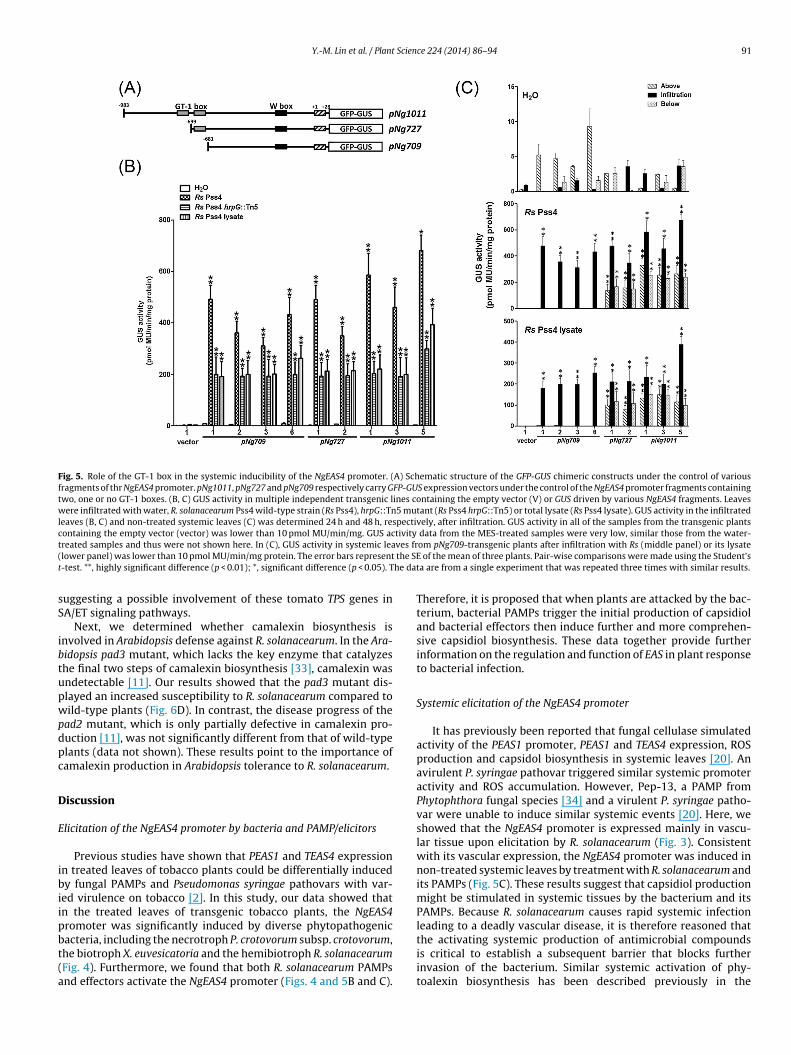
Y.-M. Lin et al. / Plant Science 224 (2014) 86–94 91
Fig. 5. Role of the GT-1 box in the systemic inducibility of the NgEAS4 promoter. (A) Schematic structure of the GFP-GUS chimeric constructs under the control of variousfragments of thr NgEAS4 promoter. pNg1011, pNg727 and pNg709 respectively carry GFP-GUS expression vectors under the control of the NgEAS4 promoter fragments containingtwo, one or no GT-1 boxes. (B, C) GUS activity in multiple independent transgenic lines containing the empty vector (V) or GUS driven by various NgEAS4 fragments. Leaveswere infiltrated with water, R. solanacearum Pss4 wild-type strain (Rs Pss4), hrpG::Tn5 mutant (Rs Pss4 hrpG::Tn5) or total lysate (Rs Pss4 lysate). GUS activity in the infiltratedleaves (B, C) and non-treated systemic leaves (C) was determined 24 h and 48 h, respectively, after infiltration. GUS activity in all of the samples from the transgenic plantscontaining the empty vector (vector) was lower than 10 pmol MU/min/mg. GUS activity data from the MES-treated samples were very low, similar those from the water-t ves fr( the SEt he dat
sS
ibtupwpdpc
D
E
ibiipbt(a
reated samples and thus were not shown here. In (C), GUS activity in systemic lealower panel) was lower than 10 pmol MU/min/mg protein. The error bars represent-test. **, highly significant difference (p < 0.01); *, significant difference (p < 0.05). T
uggesting a possible involvement of these tomato TPS genes inA/ET signaling pathways.
Next, we determined whether camalexin biosynthesis isnvolved in Arabidopsis defense against R. solanacearum. In the Ara-idopsis pad3 mutant, which lacks the key enzyme that catalyzeshe final two steps of camalexin biosynthesis [33], camalexin wasndetectable [11]. Our results showed that the pad3 mutant dis-layed an increased susceptibility to R. solanacearum compared toild-type plants (Fig. 6D). In contrast, the disease progress of the
ad2 mutant, which is only partially defective in camalexin pro-uction [11], was not significantly different from that of wild-typelants (data not shown). These results point to the importance ofamalexin production in Arabidopsis tolerance to R. solanacearum.
iscussion
licitation of the NgEAS4 promoter by bacteria and PAMP/elicitors
Previous studies have shown that PEAS1 and TEAS4 expressionn treated leaves of tobacco plants could be differentially inducedy fungal PAMPs and Pseudomonas syringae pathovars with var-
ed virulence on tobacco [2]. In this study, our data showed thatn the treated leaves of transgenic tobacco plants, the NgEAS4romoter was significantly induced by diverse phytopathogenic
acteria, including the necrotroph P. crotovorum subsp. crotovorum,he biotroph X. euvesicatoria and the hemibiotroph R. solanacearumFig. 4). Furthermore, we found that both R. solanacearum PAMPsnd effectors activate the NgEAS4 promoter (Figs. 4 and 5B and C).om pNg709-transgenic plants after infiltration with Rs (middle panel) or its lysate of the mean of three plants. Pair-wise comparisons were made using the Student’sa are from a single experiment that was repeated three times with similar results.
Therefore, it is proposed that when plants are attacked by the bac-terium, bacterial PAMPs trigger the initial production of capsidioland bacterial effectors then induce further and more comprehen-sive capsidiol biosynthesis. These data together provide furtherinformation on the regulation and function of EAS in plant responseto bacterial infection.
Systemic elicitation of the NgEAS4 promoter
It has previously been reported that fungal cellulase simulatedactivity of the PEAS1 promoter, PEAS1 and TEAS4 expression, ROSproduction and capsidol biosynthesis in systemic leaves [20]. Anavirulent P. syringae pathovar triggered similar systemic promoteractivity and ROS accumulation. However, Pep-13, a PAMP fromPhytophthora fungal species [34] and a virulent P. syringae patho-var were unable to induce similar systemic events [20]. Here, weshowed that the NgEAS4 promoter is expressed mainly in vascu-lar tissue upon elicitation by R. solanacearum (Fig. 3). Consistentwith its vascular expression, the NgEAS4 promoter was induced innon-treated systemic leaves by treatment with R. solanacearum andits PAMPs (Fig. 5C). These results suggest that capsidiol productionmight be stimulated in systemic tissues by the bacterium and itsPAMPs. Because R. solanacearum causes rapid systemic infectionleading to a deadly vascular disease, it is therefore reasoned that
the activating systemic production of antimicrobial compoundsis critical to establish a subsequent barrier that blocks furtherinvasion of the bacterium. Similar systemic activation of phy-toalexin biosynthesis has been described previously in the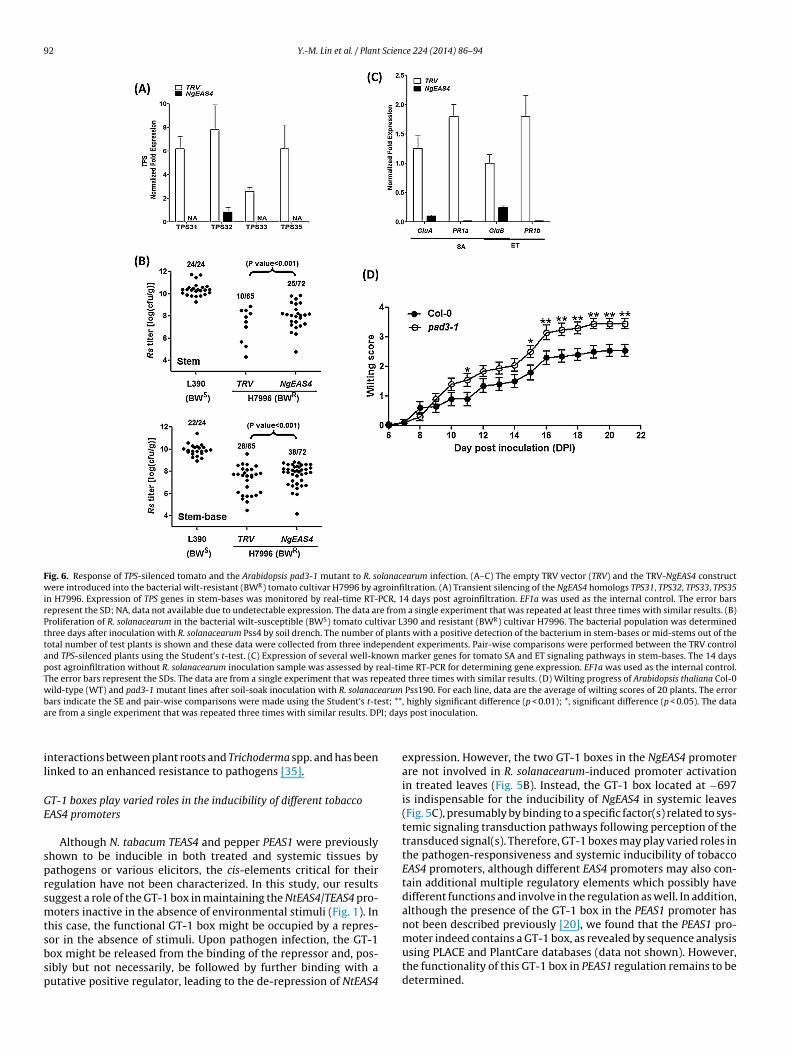
92 Y.-M. Lin et al. / Plant Science 224 (2014) 86–94
Fig. 6. Response of TPS-silenced tomato and the Arabidopsis pad3-1 mutant to R. solanacearum infection. (A–C) The empty TRV vector (TRV) and the TRV-NgEAS4 constructwere introduced into the bacterial wilt-resistant (BWR) tomato cultivar H7996 by agroinfiltration. (A) Transient silencing of the NgEAS4 homologs TPS31, TPS32, TPS33, TPS35in H7996. Expression of TPS genes in stem-bases was monitored by real-time RT-PCR, 14 days post agroinfiltration. EF1a was used as the internal control. The error barsrepresent the SD; NA, data not available due to undetectable expression. The data are from a single experiment that was repeated at least three times with similar results. (B)Proliferation of R. solanacearum in the bacterial wilt-susceptible (BWS) tomato cultivar L390 and resistant (BWR) cultivar H7996. The bacterial population was determinedthree days after inoculation with R. solanacearum Pss4 by soil drench. The number of plants with a positive detection of the bacterium in stem-bases or mid-stems out of thetotal number of test plants is shown and these data were collected from three independent experiments. Pair-wise comparisons were performed between the TRV controland TPS-silenced plants using the Student’s t-test. (C) Expression of several well-known marker genes for tomato SA and ET signaling pathways in stem-bases. The 14 dayspost agroinfiltration without R. solanacearum inoculation sample was assessed by real-time RT-PCR for determining gene expression. EF1a was used as the internal control.The error bars represent the SDs. The data are from a single experiment that was repeated three times with similar results. (D) Wilting progress of Arabidopsis thaliana Col-0wild-type (WT) and pad3-1 mutant lines after soil-soak inoculation with R. solanacearum Pss190. For each line, data are the average of wilting scores of 20 plants. The errorb st; **a I; day
il
GE
sprsmtsbsp
ars indicate the SE and pair-wise comparisons were made using the Student’s t-tere from a single experiment that was repeated three times with similar results. DP
nteractions between plant roots and Trichoderma spp. and has beeninked to an enhanced resistance to pathogens [35].
T-1 boxes play varied roles in the inducibility of different tobaccoAS4 promoters
Although N. tabacum TEAS4 and pepper PEAS1 were previouslyhown to be inducible in both treated and systemic tissues byathogens or various elicitors, the cis-elements critical for theiregulation have not been characterized. In this study, our resultsuggest a role of the GT-1 box in maintaining the NtEAS4/TEAS4 pro-oters inactive in the absence of environmental stimuli (Fig. 1). In
his case, the functional GT-1 box might be occupied by a repres-
or in the absence of stimuli. Upon pathogen infection, the GT-1ox might be released from the binding of the repressor and, pos-ibly but not necessarily, be followed by further binding with autative positive regulator, leading to the de-repression of NtEAS4, highly significant difference (p < 0.01); *, significant difference (p < 0.05). The datas post inoculation.
expression. However, the two GT-1 boxes in the NgEAS4 promoterare not involved in R. solanacearum-induced promoter activationin treated leaves (Fig. 5B). Instead, the GT-1 box located at −697is indispensable for the inducibility of NgEAS4 in systemic leaves(Fig. 5C), presumably by binding to a specific factor(s) related to sys-temic signaling transduction pathways following perception of thetransduced signal(s). Therefore, GT-1 boxes may play varied roles inthe pathogen-responsiveness and systemic inducibility of tobaccoEAS4 promoters, although different EAS4 promoters may also con-tain additional multiple regulatory elements which possibly havedifferent functions and involve in the regulation as well. In addition,although the presence of the GT-1 box in the PEAS1 promoter hasnot been described previously [20], we found that the PEAS1 pro-
moter indeed contains a GT-1 box, as revealed by sequence analysisusing PLACE and PlantCare databases (data not shown). However,the functionality of this GT-1 box in PEAS1 regulation remains to bedetermined.
Scien
Ps
pIiasdaTHcitibadtphus
hmtigibmsasRr
lptliisrimes
A
RtVttoUpo
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
Y.-M. Lin et al. / Plant
hytoalexin biosynthesis is important for plant defense against R.olanacearum
Accumulated studies have demonstrated the contribution ofhytoalexins to plant resistance against fungi and oomycetes [10].
n previous studies, it was concluded that Arabidopsis phytoalex-ns are not effective against pathogenic bacteria, such as P. syringaend Xanthomonas campestris [10–12]. However, our study demon-trates the importance of camalexin biosynthesis in Arabidopsisefense against R. solanacearum (Fig. 6D). Consistently, the properccumulation of transcripts encoding sequiterpene synthase TPS31,PS32, TPS33 and TPS35 is important for the expression of tomato7996 resistance to R. solanacearum (Fig. 6B), which implicates theontribution of phytoalexin production to tomato defense responsen vivo. These results support a positive role of phytoalexin biosyn-hesis in plant tolerance to R. solanacearum and agree with themplicated importance of phytoalexins in the defense response ofean and potato to pathogenic bacteria [13,14]. However, the exactntibacterial mechanisms of their actions remained to be eluci-ated. In addition, our study, together with other reports, revealshat Solanaceae EAS gene expression can be activated by diverseathogens, PAMPs/elicitors and abiotic stresses. Future studies willelp to clarify whether EAS4 orthologs and their phytoalexin prod-cts have a wide-spectrum of functions in the defense responses ofolanaceous plants against biotic and abiotic stresses.
This study shows that silencing TPS31, TPS32, TPS33 and TPS35as a negative effect on the expression of several well-knownarker genes for SA and ET signaling pathways (Fig. 6C). In addi-
ion, known ET- or SA-responsive cis-elements were also identifiedn the sequences upstream of the coding regions of these four TPSenes (data not shown). In line with our findings, previous stud-es have shown ET-mediated regulation of EAS expression in N.enthamiana [36] and the presence of putative ET-responsive ele-ents in the promoters of TEAS4 and PEAS1 [20]. These together
uggest a possible cross-talk between phytoalexin biosynthesisnd SA/ET defense signaling pathways. In addition, SA- and ET-ignaling pathways are involved in tomato H7996 resistance to. solanacearum [29]. Further investigation will help to clarify theelationship among these pathways.
Regulating the biosynthesis of phytoalexins is crucial for estab-ishing plant disease resistance. Here, we report that the NgEAS4romoter can be induced by diverse phytopathogenic bacteria andhat R. solanacearum PAMPs and effectors are both involved inocal and systemic NgEAS4 induction. Our study also highlights var-ed roles of GT-1 boxes in the regulation of NtEAS4 and NgEAS4n response to bacterial infection. Moreover, this study demon-trates the importance of phytoalexin biosynthesis for the defenseesponse of tomato and Arabidopsis against R. solanacearum. Suchnducible promoters and the information provided in this study
ight be valuable in engineering crops with tightly controllednhanced resistance to commercially important pathogens [37],uch as R. solanacearum.
cknowledgements
We thank Dr. Ming-Tsair Chan (Agricultural Biotechnologyesearch Center, Academia Sinica, Taipei, Taiwan) for providinghe pBSks+-NOS plasmid, Dr. Jaw-Fen Wang (AVRDC-The Worldegetable Center, Taiwan) for providing the phytopathogens (Pec-
obacterium crotovorum subsp. crotovorum, Xanthomonas euvesica-oria and R. solanacearum) and Dr. Laurent Zimmerli (Department
f Life Science and Institute of Plant Biology, National Taiwanniversity, Taipei, Taiwan) for providing the Arabidopsis thalianaad3-1 mutant. We appreciate the excellent technical assistancef Technology Commons, College of Life Science, National Taiwan[
ce 224 (2014) 86–94 93
University. This work was supported by grants from the Council ofAgriculture (99-1.1.1-Z1(z)) and National Science of Council (NSC101-2324-B-002-019) to C.-P. Cheng.
Appendix A. Supplementary data
Supplementary material related to this article can be found,in the online version, at http://dx.doi.org/10.1016/j.plantsci.2014.04.008.
References
[1] J.D. Jones, J.L. Dangl, The plant immune system, Nature 444 (2006) 323–329.[2] M. Bernoux, J.G. Ellis, P.N. Dodds, New insights in plant immunity signaling
activation, Curr. Opin. Plant Biol. 14 (2011) 512–518.[3] B.N. Kunkel, D.M. Brooks, Cross talk between signaling pathways in pathogen
defense, Curr. Opin. Plant Biol. 5 (2002) 325–331.[4] T. Eulgem, Regulation of the Arabidopsis defense transcriptome, Trends Plant
Sci. 10 (2005) 71–78.[5] K.F. Pedley, G.B. Martin, Role of mitogen-activated protein kinases in plant
immunity, Curr. Opin. Plant Biol. 8 (2005) 541–547.[6] A. Koornneef, C.M. Pieterse, Cross talk in defense signaling, Plant Physiol. 146
(2008) 839–844.[7] J. Shah, Plants under attack: systemic signals in defence, Curr. Opin. Plant Biol.
12 (2009) 459–464.[8] C.M. Pieterse, A. Leon-Reyes, S. Van der Ent, S.C. Van Wees, Networking
by small-molecule hormones in plant immunity, Nat. Chem. Biol. 5 (2009)308–316.
[9] I. Ahuja, R. Kissen, A.M. Bones, Phytoalexins in defense against pathogens,Trends Plant Sci. 17 (2012) 73–90.
10] P. Bednarek, Chemical warfare or modulators of defence responses – the func-tion of secondary metabolites in plant immunity, Curr. Opin. Plant Biol. 15(2012) 407–414.
11] J. Glazebrook, F.M. Ausubel, Isolation of phytoalexin-deficient mutants ofArabidopsis thaliana and characterization of their interactions with bacterialpathogens, Proc. Natl. Acad. Sci. U. S. A. 91 (1994) 8955–8959.
12] J. Glazebrook, M. Zook, F. Mert, I. Kagan, E.E. Rogers, I.R. Crute, E.B. Holub, R.Hammerschmidt, F.M. Ausubel, Phytoalexin-deficient mutants of Arabidopsisreveal that PAD4 encodes a regulatory factor and that four PAD genes contributeto downy mildew resistance, Genetics 146 (1997) 381–392.
13] N.T. Kenn, B.W. Kenndey, Hydroxyphaseollin and relatec isoflavonoids in thehypersensitive resistance response of soybeans against Pseudomonas glycinea,Physiol. Plant Pathol. 4 (1974) 173–185.
14] G.D. Lyon, C.E. Bayliss, The effect of rishitin on Erwinia carotovora var. atrosepticaand other bacteria, Physiol. Plant Pathol. 6 (1975) 177–186.
15] P.J. Facchini, J. Chappell, Gene family for an elicitor-induced sesquiterpenecyclase in tobacco, Proc. Natl. Acad. Sci. U. S. A. 89 (1992) 11088–11092.
16] K. Back, S. Yin, J. Chappell, Expression of a plant sesquiterpene cyclase gene inEscherichia coli, Arch. Biochem. Biophys. 315 (1994) 527–532.
17] K. Back, J. Nah, S.B. Lee, J.H. Song, D.H. Shin, H.Y. Kim, Cloning of a sesquiterpenecyclase and its functional expression by domain swapping strategy, Mol. Cells10 (2000) 220–225.
18] S. Yin, L. Mei, J. Newman, K. Back, J. Chappell, Regulation of sesquiterpenecyclase gene expression. Characterization of an elicitor- and pathogen-inducible promoter, Plant Physiol. 115 (1997) 437–451.
19] A. Ghannam, A. Jacques, P. De Ruffray, F. Baillieul, S. Kauffmann, Identificationof tobacco ESTs with a hypersensitive response (HR)-specific pattern of expres-sion and likely involved in the induction of the HR and/or localized acquiredresistance (LAR), Plant Physiol. Biochem. 43 (2005) 249–259.
20] L.D. Maldonado-Bonilla, M. Betancourt-Jimenez, E. Lozoya-Gloria, Local andsystemic gene expression of sesquiterpene phytoalexin biosynthetic enzymesin plant leaves, Eur. J. Plant Pathol. 121 (2008) 439–449.
21] S.T. Perrone, K.L. McDonald, M.W. Sutherland, D.I. Guest, Superoxide release isnecessary for phytoalexin accumulation in Nicotiana tabacum cells during theexpression of cultivar-race and non-host resistance towards Phytophthora spp,Physiol. Mol. Plant Pathol. 62 (2003) 127–135.
22] A.C. Araceli, C.M. Elda, L.G. Edmundo, G.P. Ernesto, Capsidiol production in pep-per fruits (Capsicum annuum L.) induced by arachidonic acid is dependent of anoxidative burst, Physiol. Mol. Plant Pathol. 70 (2007) 69–76.
23] I.S. Curtis, J.B. Power, M.R. Davey, NPTII assays for measuring gene expressionand enzyme activity in transgenic plants, in: Methods in Molecular Biology,Springer, New Jersey, 1995, pp. 149–159.
24] R. Takabatake, T. Mukaihara, Extracts from Ralstonia solanacearum induce effec-tive resistance to the pathogen in both Arabidopsis and solanaceous plants, J.Gen. Plant Pathol. 77 (2011) 33–42.
25] W.C. Lin, C.F. Lu, J.W. Wu, M.L. Cheng, Y.M. Lin, N.S. Yang, L. Black, S.K. Green, J.F.
Wang, C.P. Cheng, Transgenic tomato plants expressing the Arabidopsis NPR1gene display enhanced resistance to a spectrum of fungal and bacterial diseases,Transgenic Res. 13 (2004) 567–581.26] Y.N. Yang, R.G. Li, M. Qi, In vivo analysis of plant promoters and transcriptionfactors by agroinfiltration of tobacco leaves, Plant J. 22 (2000) 543–551.

9 Scien
[
[
[
[
[
[
[
[
[
[
4 Y.-M. Lin et al. / Plant
27] R.A. Jefferson, T.A. Kavanagh, M.W. Bevan, Gus fusions: �-glucuronidase as asensitive and versatile gene fusion marker in higher plants, EMBO J. 6 (1987)3901–3907.
28] F.I. Ho, Y.Y. Chen, Y.M. Lin, C.P. Cheng, J.F. Wang, A tobacco rattle virus-induced gene silencing system for a soil-borne vascular pathogen Ralstoniasolanacearum, Bot. Stud. 50 (2009) 413–424.
29] Y.Y. Chen, Y.M. Lin, T.C. Chao, J.F. Wang, A.C. Liu, F.I. Ho, C.P. Cheng, Virus-induced gene silencing reveals the involvement of ethylene-, salicylic acid- andmitogen-activated protein kinase-related defense pathways in the resistanceof tomato to bacterial wilt, Physiol. Plant. 136 (2009) 324–335.
30] Y.M. Lin, I.C. Chou, J.F. Wang, F.I. Ho, Y.J. Chu, P.C. Huang, D.K. Lu, H.L. Shen,M. Elbaz, S.M. Huang, C.P. Cheng, Transposon mutagenesis reveals differentialpathogenesis of Ralstonia solanacearum on tomato and Arabidopsis, Mol. PlantMicrobe Interact. 21 (2008) 1261–1270.
31] J.A.L. Van Kan, T. Cozijnsen, N. Danhash, P.J. De Wit, Induction of tomato stress
protein mRNAs by ethephon, 2,6-dichloroisonicotinic acid and salicylate, PlantMol. Biol. 27 (1995) 1205–1213.32] A. Block, E. Schmelz, P.J. O’Donnell, J.B. Jones, H.J. Klee, Systemic acquired tol-erance to virulent bacterial pathogens in tomato, Plant Physiol. 138 (2005)1481–1490.
[
ce 224 (2014) 86–94
33] C. Bottcher, L. Westphal, C. Schmotz, E. Prade, D. Scheel, E. Glaw-ischnig, The multifunctional enzyme CYP71B15 (PHYTOALEXIN DEFI-CIENT3) converts cysteine-indole-3-acetonitrile to camalexin in the indole-3-acetonitrile metabolic network of Arabidopsis thaliana, Plant Cell 21 (2009)1830–1845.
34] F. Brunner, S. Rosahl, J. Lee, J.J. Rudd, C. Geiler, S. Kauppinen, G. Rasmussen,D. Scheel, T. Nurnberger, Pep-13, a plant defense-inducing pathogen-associated pattern from Phytophthora transglutaminases, EMBO J. 21 (2002)6681–6688.
35] G.E. Harman, C.R. Howell, A. Viterbo, I. Chet, M. Lorito, Trichoderma species– opportunistic, avirulent plant symbionts, Nat. Rev. Microbiol. 2 (2004)43–56.
36] Y. Shibata, K. Kawakita, D. Takemoto, Age-related resistance of Nicotiana ben-thamiana against hemibiotrophic pathogen Phytophthora infestans requiresboth ethylene- and salicylic acid-mediated signaling pathways, Mol. Plant
Microbe Interact. 23 (2010) 1130–1142.37] P.M. Bleeker, R. Mirabella, P.J. Diergaarde, A. VanDoorn, A. Tissier, M. Prins, M. deVos, M.A. Haring, R.C. Schuurink, Improved herbivore resistance in cultivatedtomato with the sesquiterpene biosynthetic pathway from a wild relative, Proc.Natl. Acad. Sci. U. S. A. 109 (2012) 20124–20129.
Related Documents











