Phylogenetic analysis of Sesuvioideae (Aizoaceae) inferred from nrDNA internal transcribed spacer (ITS) sequences and morphological data N. S. Hassan 1 , J. Thiede 2 , and S. Liede-Schumann 3 1 Botany Department, Faculty of Science, Assiut University, Assiut, Egypt 2 Botanical Institute & Garden, Hamburg University, Hamburg, Germany 3 Department of Plant Systematics, University of Bayreuth, Bayreuth, Germany Received March 1, 2004; accepted May 22, 2005 Published online: August 8, 2005 ȑ Springer-Verlag 2005 Abstract. The phylogeny of the four genera of Aizoaceae subfamily Sesuvioideae (Sesuvium, Cypselea, Trianthema and Zaleya) is elucidated employing internal transcribed spacer (ITS) sequences and 23 morphological characters. Phy- logenetic analysis based on ITS sequences and a combined molecular-morphological analysis pro- vide largely congruent results. The monophyly of Sesuvioideae and its close relationship to Aizooi- deae s.l. and Mesembryanthemoideae is con- firmed. Zaleya is placed within Trianthema and Cypselea within Sesuvium by ITS analysis, but in the combined analysis, Zaleya forms an unre- solved polytomy with the two Trianthema clades and Cypselea, as well as two Sesuvium species, remain unresolved. Sesuvium sesuvioides and S. hydaspicum, previously treated as synonyms, are closely related, but molecular data do not support conspecifity. The Trianthema triquetra complex needs further intensive study because the African T. triquetra sample is closer to the NE African-Arabian T. sheilae than to Australian samples of T. triquetra. The close relationship of four species of Trianthema (T. patellitecta, T. rhynchocalyptra, T. megasperma, and T. pilosa) from Australia based on molecular data is sup- ported morphologically by the exclusive posses- sion of a well-developed indumentum in these taxa. Key words: Aizoaceae, Cypselea, Evolution, ITS, Morphology, Parsimony, Sesuvioideae, Sesuvium, Trianthema, Zaleya. The subfamily Sesuvioideae Lindley (1846) represents one of four subfamilies constituting the family Aizoaceae s. str. (Klak et al. 2003, Thiede 2004) as circumscribed by Bittrich and Hartmann (1988) within the order Caryophyll- ales (Cue´noud 2003). This subfamily is mor- phologically characterized by the possession of circumscissile capsules and leaf petioles with membranous sheaths or stipule-like append- ages at the base; in addition, bracteate inflo- rescences, perigynous flowers, a perianth mostly colored inside and with subapical dorsal appendages, arillate seeds and a basic chromosome number of n = 8 are frequently found (Bittrich and Hartmann 1988, Bittrich 1990). Sesuvioideae are distributed in tropical and subtropical regions all over the world and are widely found in salty soil, at the edges of Pl. Syst. Evol. 255: 121–143 (2005) DOI 10.1007/s00606-005-0357-x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Phylogenetic analysis of Sesuvioideae (Aizoaceae) inferred
from nrDNA internal transcribed spacer (ITS) sequences
and morphological data
N. S. Hassan1, J. Thiede
2, and S. Liede-Schumann
3
1Botany Department, Faculty of Science, Assiut University, Assiut, Egypt2Botanical Institute & Garden, Hamburg University, Hamburg, Germany3Department of Plant Systematics, University of Bayreuth, Bayreuth, Germany
Received March 1, 2004; accepted May 22, 2005Published online: August 8, 2005� Springer-Verlag 2005
Abstract. The phylogeny of the four genera ofAizoaceae subfamily Sesuvioideae (Sesuvium,Cypselea, Trianthema and Zaleya) is elucidatedemploying internal transcribed spacer (ITS)sequences and 23 morphological characters. Phy-logenetic analysis based on ITS sequences and acombined molecular-morphological analysis pro-vide largely congruent results. The monophyly ofSesuvioideae and its close relationship to Aizooi-deae s.l. and Mesembryanthemoideae is con-firmed. Zaleya is placed within Trianthema andCypselea within Sesuvium by ITS analysis, but inthe combined analysis, Zaleya forms an unre-solved polytomy with the two Trianthema cladesand Cypselea, as well as two Sesuvium species,remain unresolved. Sesuvium sesuvioides andS. hydaspicum, previously treated as synonyms,are closely related, but molecular data do notsupport conspecifity. The Trianthema triquetracomplex needs further intensive study becausethe African T. triquetra sample is closer to the NEAfrican-Arabian T. sheilae than to Australiansamples of T. triquetra. The close relationshipof four species of Trianthema (T. patellitecta,T. rhynchocalyptra, T. megasperma, and T. pilosa)from Australia based on molecular data is sup-ported morphologically by the exclusive posses-
sion of a well-developed indumentum in thesetaxa.
Key words: Aizoaceae, Cypselea, Evolution, ITS,Morphology, Parsimony, Sesuvioideae, Sesuvium,Trianthema, Zaleya.
The subfamily Sesuvioideae Lindley (1846)represents one of four subfamilies constitutingthe family Aizoaceae s. str. (Klak et al. 2003,Thiede 2004) as circumscribed by Bittrich andHartmann (1988) within the order Caryophyll-ales (Cuenoud 2003). This subfamily is mor-phologically characterized by the possession ofcircumscissile capsules and leaf petioles withmembranous sheaths or stipule-like append-ages at the base; in addition, bracteate inflo-rescences, perigynous flowers, a perianthmostly colored inside and with subapicaldorsal appendages, arillate seeds and a basicchromosome number of n = 8 are frequentlyfound (Bittrich and Hartmann 1988, Bittrich1990). Sesuvioideae are distributed in tropicaland subtropical regions all over the world andare widely found in salty soil, at the edges of
Pl. Syst. Evol. 255: 121–143 (2005)DOI 10.1007/s00606-005-0357-x
Used Distiller 5.0.x Job Options
This report was created automatically with help of the Adobe Acrobat Distiller addition "Distiller Secrets v1.0.5" from IMPRESSED GmbH. You can download this startup file for Distiller versions 4.0.5 and 5.0.x for free from http://www.impressed.de. GENERAL ---------------------------------------- File Options: Compatibility: PDF 1.2 Optimize For Fast Web View: Yes Embed Thumbnails: Yes Auto-Rotate Pages: No Distill From Page: 1 Distill To Page: All Pages Binding: Left Resolution: [ 600 600 ] dpi Paper Size: [ 595.276 785.197 ] Point COMPRESSION ---------------------------------------- Color Images: Downsampling: Yes Downsample Type: Bicubic Downsampling Downsample Resolution: 150 dpi Downsampling For Images Above: 225 dpi Compression: Yes Automatic Selection of Compression Type: Yes JPEG Quality: Medium Bits Per Pixel: As Original Bit Grayscale Images: Downsampling: Yes Downsample Type: Bicubic Downsampling Downsample Resolution: 150 dpi Downsampling For Images Above: 225 dpi Compression: Yes Automatic Selection of Compression Type: Yes JPEG Quality: Medium Bits Per Pixel: As Original Bit Monochrome Images: Downsampling: Yes Downsample Type: Bicubic Downsampling Downsample Resolution: 600 dpi Downsampling For Images Above: 900 dpi Compression: Yes Compression Type: CCITT CCITT Group: 4 Anti-Alias To Gray: No Compress Text and Line Art: Yes FONTS ---------------------------------------- Embed All Fonts: Yes Subset Embedded Fonts: No When Embedding Fails: Warn and Continue Embedding: Always Embed: [ ] Never Embed: [ ] COLOR ---------------------------------------- Color Management Policies: Color Conversion Strategy: Convert All Colors to sRGB Intent: Default Working Spaces: Grayscale ICC Profile: RGB ICC Profile: sRGB IEC61966-2.1 CMYK ICC Profile: U.S. Web Coated (SWOP) v2 Device-Dependent Data: Preserve Overprint Settings: Yes Preserve Under Color Removal and Black Generation: Yes Transfer Functions: Apply Preserve Halftone Information: Yes ADVANCED ---------------------------------------- Options: Use Prologue.ps and Epilogue.ps: No Allow PostScript File To Override Job Options: Yes Preserve Level 2 copypage Semantics: Yes Save Portable Job Ticket Inside PDF File: No Illustrator Overprint Mode: Yes Convert Gradients To Smooth Shades: No ASCII Format: No Document Structuring Conventions (DSC): Process DSC Comments: No OTHERS ---------------------------------------- Distiller Core Version: 5000 Use ZIP Compression: Yes Deactivate Optimization: No Image Memory: 524288 Byte Anti-Alias Color Images: No Anti-Alias Grayscale Images: No Convert Images (< 257 Colors) To Indexed Color Space: Yes sRGB ICC Profile: sRGB IEC61966-2.1 END OF REPORT ---------------------------------------- IMPRESSED GmbH Bahrenfelder Chaussee 49 22761 Hamburg, Germany Tel. +49 40 897189-0 Fax +49 40 897189-71 Email: [email protected] Web: www.impressed.de
Adobe Acrobat Distiller 5.0.x Job Option File
<< /ColorSettingsFile () /AntiAliasMonoImages false /CannotEmbedFontPolicy /Warning /ParseDSCComments false /DoThumbnails true /CompressPages true /CalRGBProfile (sRGB IEC61966-2.1) /MaxSubsetPct 100 /EncodeColorImages true /GrayImageFilter /DCTEncode /Optimize true /ParseDSCCommentsForDocInfo false /EmitDSCWarnings false /CalGrayProfile () /NeverEmbed [ ] /GrayImageDownsampleThreshold 1.5 /UsePrologue false /GrayImageDict << /QFactor 0.9 /Blend 1 /HSamples [ 2 1 1 2 ] /VSamples [ 2 1 1 2 ] >> /AutoFilterColorImages true /sRGBProfile (sRGB IEC61966-2.1) /ColorImageDepth -1 /PreserveOverprintSettings true /AutoRotatePages /None /UCRandBGInfo /Preserve /EmbedAllFonts true /CompatibilityLevel 1.2 /StartPage 1 /AntiAliasColorImages false /CreateJobTicket false /ConvertImagesToIndexed true /ColorImageDownsampleType /Bicubic /ColorImageDownsampleThreshold 1.5 /MonoImageDownsampleType /Bicubic /DetectBlends false /GrayImageDownsampleType /Bicubic /PreserveEPSInfo false /GrayACSImageDict << /VSamples [ 2 1 1 2 ] /QFactor 0.76 /Blend 1 /HSamples [ 2 1 1 2 ] /ColorTransform 1 >> /ColorACSImageDict << /VSamples [ 2 1 1 2 ] /QFactor 0.76 /Blend 1 /HSamples [ 2 1 1 2 ] /ColorTransform 1 >> /PreserveCopyPage true /EncodeMonoImages true /ColorConversionStrategy /sRGB /PreserveOPIComments false /AntiAliasGrayImages false /GrayImageDepth -1 /ColorImageResolution 150 /EndPage -1 /AutoPositionEPSFiles false /MonoImageDepth -1 /TransferFunctionInfo /Apply /EncodeGrayImages true /DownsampleGrayImages true /DownsampleMonoImages true /DownsampleColorImages true /MonoImageDownsampleThreshold 1.5 /MonoImageDict << /K -1 >> /Binding /Left /CalCMYKProfile (U.S. Web Coated (SWOP) v2) /MonoImageResolution 600 /AutoFilterGrayImages true /AlwaysEmbed [ ] /ImageMemory 524288 /SubsetFonts false /DefaultRenderingIntent /Default /OPM 1 /MonoImageFilter /CCITTFaxEncode /GrayImageResolution 150 /ColorImageFilter /DCTEncode /PreserveHalftoneInfo true /ColorImageDict << /QFactor 0.9 /Blend 1 /HSamples [ 2 1 1 2 ] /VSamples [ 2 1 1 2 ] >> /ASCII85EncodePages false /LockDistillerParams false >> setdistillerparams << /PageSize [ 576.0 792.0 ] /HWResolution [ 600 600 ] >> setpagedevice

mangroves and near roadsides (Hartmann2001a, b), rarely on rocky ground. The sub-family Sesuvioideae included four genera,Cypselea Turp. (3 spp.), Sesuvium L. (ca. 22spp.), Trianthema L. (ca. 28 spp.) and ZaleyaBurm. f. (ca. 7 spp.) (Hartmann 2001a, b). Afifth genus, Tribulocarpus S. Moore which wasformerly placed in subfamily Tetragonioideae,was added recently and takes a sistergroupposition to the four traditional Sesuvioideaegenera (Klak et al. 2003, Thiede 2004).
The position of Sesuvioideae and theirrelationships with the other subfamilies ofAizoaceae are a matter of debate among manyauthors and need further study to come to awell-based conclusion. Horaninow (1834) wasthe first author to use Sesuviaceae instead ofFicoideae (= Aizoaceae) in which he includedthe following genera: Aizoon L., Mesembryan-themum L. (Aptenia N. E. Br. and Ruschiagroup), Tetragonia L., Sesuvium L., Trianthe-ma L., Glinus L.; Reaumuria L. and NitrariaL. (Hartmann 2001a, b). Eckardt (1964)included the four genera of the traditionalSesuvioideae in subfamily Aizooideae Spreng.ex Arn. Hofmann (1973) took a different pointof view and and excluded the Sesuvium groupfrom Aizoaceae s. str. suggesting also that theSesuvium group should be transferred to thefamily Molluginaceae because of some simi-larities in morphological characters such as:inflorescences with distinct small bracts, sta-mens epi- and alternisepalous, plicate sectionof carpels well-developed, and arillate seeds.However, the phylogenetic status (apo- versusplesiomorphic) of the characters employed byHofmann (1973) remains uncertain, and sinceher results (the Sesuvium-group shares 4 char-acters with Aizoaceae and 6 with Mollugina-ceae) are not clear, her treatment did notacquire attention from other scholars. Ehren-dorfer (1976) proposed family rank for theSesuvium group and Cronquist (1981) alsoconsidered Sesuviaceae a distinct family in hisclassification. An important step forward wasprovided by the cladistic considerations ofBittrich and Hartmann (1988), Bittrich andStruck (1989) and Bittrich (1990). Bittrich and
Hartmann (1988) could not find any synapo-morphy for Molluginaceae plus Aizoaceae andthus rejected the hypothesis of a close rela-tionship between both families. In contrast,monophyly of the Aizoaceae was establishedbased mainly on the the synapomorphy ofbladder cell idioblasts in the epidermis, and thefamily was circumscribed as to include fivesubfamilies: Aizooideae, Sesuvioideae Lindley,Tetragonioideae Lindley, Mesembryanthemoi-deae Burnett, and Ruschioideae Schwantes inIhlenfeldt, Schwantes and Straka.
Recent rearrangements by Klak et al.(2003) and Thiede (2004), based on moleculardata, support unanimously the inclusion ofTetragonia in the Aizooideae and the transferof Tribulocarpus to the Sesuvioideae (fordetails see Thiede 2004). The latter was,however, not included in this study, whichwas concerned with the relationships amongthe four traditional Sesuvioideae genera. Nei-ther Bittrich and Hartmann (1988), nor Klaket al. (2003), nor Thiede (2004) were able todetermine synapomorphies for the subfamilyAizooideae. For the other subfamilies, mono-phyly is supported by at least one synapomor-phy, and the Sesuvioideae – Aizooideae andMesembryanthemoideae – Ruschioideae wereregarded as more closely related to each otherthan to the remaining ones (Bittrich andHartmann 1988). The Sesuvium-group sharesthe above mentioned synapomorphy of theAizoaceae and shares more features with thesubfamily Aizooideae than with Mollugina-ceae, and was consequently included in Bit-trich and Hartmann’s (1988) concept ofAizoaceae. This treatment was widely followedand has been confirmed by the mentionedrecent studies in principal (Klak et al. 2003,Thiede 2004).
Within Aizoaceae, Sesuvioideae areassumed to hold a basal position, eitherrepresenting a sister group to all other sub-families which still form a monophyleticgroup, or representing the sister-clade to thenewly defined Aizooideae, which in turn areboth sister to Mesembryanthemoideae plusRuschioideae (Bittrich and Struck 1989,
122 N. S. Hassan et al.: Sesuvioideae

Bittrich 1990). The former assumption issupported by the following characters 1) theanther wall in Sesuvioideae is composed of fivelayers (Raghavan and Srinivasan 1940), but offour in the remaining subfamilies (Schmid1925). However, according to Bittrich (1990)this character has been examined in few speciesonly and needs more investigation; 2) theprimordia of stamens originate both oppositeand alternate of the perianth lobes in Sesuvi-oideae but only alternate in the remainingsubfamilies (e.g. Hofmann 1973). Based onBittrich (1990), the following differences couldbe added: 3) circumscissile vs. hygrochasticcapsules; 4) epicuticular rodlet layer absent vs.present.
The latter assumption is likewise well-supported with morphological characters:Sesuvioideae and Aizooideae share the pres-ence of calcium oxalate druses, perigynousflowers with a simple perianth which is sepal-oid outside and petaloid inside, and a basicchromosome number of n = 8, as opposed tothe presence of calcium oxalate raphides, semi-epigynous or epigynous flowers with a doubleperianth with petaloid staminodes, and a basicchromosome number of n = 9 in Mesembry-anthemoideae and Ruschioideae (Bittrich andStruck 1989, Bittrich 1990).
Due to the lack of a suitable sister- (out-)group for the Aizoaceae and insufficientknowledge of the polarity of many characterstates within Caryophyllales, no decisionbetween both hypotheses could be made, butthe first one was regarded as more likely(Bittrich and Struck 1989, Bittrich 1990).
Sesuvioideae share with Molluginaceaearillate seeds which were regarded as a prob-ably primitive feature by Corner (1976), whosuggested that seeds of Aizoaceae ancestorswere covered with an aril. However, only fewtaxa within Caryophyllales exhibit arillateseeds [Stegnosperma Benth. (Phytolaccaceae;Hofmann 1977), Macarthuria Hugel ex Endl.(Molluginaceae; Muller 1909)]. Arils may,therefore, probably represent an apomorphicfeature (Bittrich and Hartmann 1988) whichmay have arisen independently in Sesuvioideae
and Molluginaceae. If the presence of an aril isconsidered an important synapomorphy forSesuvioideae, the position of subfamily Sesu-vioideae as part of Aizoaceae is stabilized(Hartmann 2001a). The adaptive value con-nected with arillate seeds is seen in myxo-spermy which occurs in at least some species ofTrianthema (Bittrich 1990).
Within Sesuvioideae, the inter- andintra-relationships of the four traditional gen-era were and still are a matter of debate. Forinstance, Zaleya and Trianthema were con-fused in floristic treatments (e.g. Oliver 1871,Muschler 1912, Backer 1954, Hutchinson andDalziel 1956, Tackholm 1956, Zohary 1966)even though they differ clearly in their mor-phological characters (Melville 1952, Hart-mann 2001), which caused Jeffrey (1960) tore-establish Zaleya. In her key, Hartmann(2001a) separated Sesuvium and Zaleya onone hand by the possession of at least twostigmas, and added that many seeds in axileplacentation are the most useful character todistinguish Sesuvium from Trianthema andZaleya, which are both few-seeded.
Anatomical information on the group isscarce. Bhambie et al. (1977) reported thattrilacunar and unilacunar leaf traces arefound in Trianthema, but only trilacunar onesin Zaleya. According to Landrum (2001)Sesuvioideae is devoid of wide-band tracheidslike all other taxa of Aizoaceae except forRuschioideae tribe Ruschieae, which possesswell-developed wide-band tracheids (Klaket al. 2003). Kranz cells are present in allfour genera of subfamily Sesuvioideae studiedhere, but not in all species of Sesuvium; forexample, they are found in S. sesuvioides butnot in S. portulacastrum (e.g. Bittrich 1990).Molluginaceae likewise possess some taxawith Kranz anatomy (Endress and Bittrich1993), but to use this feature to establish arelationship with Sesuvioideae is doubtfulsince C4 photosynthesis is of manyfold origin(Sage 2001) and perhaps more of ecologicalthan systematic significance. Biochemically,free amino and organic acids have been usedto separate two Trianthema species; in
N. S. Hassan et al.: Sesuvioideae 123

T. portulacastrum L-tyrosine is present, inT. triquetra it is absent (Singh 1992). How-ever, no further information on this characteris yet available.
A number of molecular studies haveelucidated the phylogeny of Caryophyllaleslately. In most of these studies, Aizoaceaecame out in a terminal clade within the highercore Caryophyllales together with Nyctagina-ceae, Phytolaccaceae, Petiveriaceae and somesmaller taxa; Molluginaceae were placed at adistance from this clade (e.g. Cuenoud et al.2002). The loss of the chloroplast rpl2 intronhas been established as a new synapomorphyfor the order Caryophyllales (Downie et al.1991, Downie and Palmer 1994). First molec-ular rbcL sequence data (Manhart and Rettig1994) grouped Trianthema (Sesuvioideae)separate from Lithops (Ruschioideae), but withvery low bootstrap support. Sequence datafrom the rpl16 intron indicated monophyly ofAizoaceae, with Sesuvioideae appearing assister to the rest of Aizoaceae (Wallace 1998,abstract only). In a chloroplast matK analysis,Cuenoud et al. (2002) included six genera ofAizoaceae representing three subfamilies(Aizooideae, Mesembryanthemoideae andRuschioideae). According to their results(Cuenoud et al. 2002), the subfamily Aizooi-deae (Galenia and Plinthus) is sister toMesembryanthemoideae (Phyllobolus) andRuschioideae (Delosperma, Ruschia andTrichodiadema) which are in turn sister to eachother. Klak et al. (2003) in an analysis of fourcpDNA regions of 91 Aizoaceae species andThiede (2004) in an analysis of ITS sequencesof 59 Aizoaceae included two samples ofSesuvioideae (Sesuvium and Trianthema) eachin their analyses. Both studies yielded coinci-dent results: the four genera constituting sub-family Sesuvioideae according to Bittrich andHartmann (1988) is monophyletic and sister toTribulocarpus which was formerly placed insubfamily Tetragonioideae; this clade is sisterto the remainder of the Aizoaceae. Conse-quently, the concept of Sesuvioideae wasbroadened as to include Tribulocarpus (Klaket al. 2003, Thiede 2004) and Klak et al. (2003)
carried out the formal transfer of Tribulocar-pus to Sesuvioideae.
The present study focusses on the subfam-ily Sesuvioideae in its previous circumscrip-tion. Monophyly of Sesuvioideae is stronglyindicated by morphological character statesand the studies of Klak et al. (2003) andThiede (2004), but a molecular study involvingall genera and a reasonable number of specieswas yet lacking. ITS (nrDNA) sequences havebeen shown to constitute a useful tool forphylogenetic tests at generic and specific level(e.g. Baldwin et al. 1995) and were usedsuccessfully within Caryophyllales for a studyof the Portulacaceae clade (Hershkowitz andZimmer 1997) and for Aizoaceae (Thiede2004). The aim of the present paper is there-fore, 1) to establish the monophlyly of Sesu-vioideae with a broad sample; and 2) toinvestigate the phylogenetic relations andmonophyly of the four traditionally includedgenera.
Materials and Methods
Plant material. Twenty-two species representing allfour genera in subfamily Sesuvioideae s. str. wereincluded in the present study (see Table 1, forauthors of species). One species of Aizooideae,Aizoanthemum dinteri, two of Mesembryanthemoi-deae, Mesembryanthemum aitonis and M. hyper-trophicum and one of Tetragonioideae, Tetragoniatetragonoides, were used as outgroups.
DNA extraction, PCR amplification and DNA
sequencing of ITS regions. Total genomic DNAwas extracted from herbarium and fresh materialswith DNeasy� Plant MiniKit (QIAGEN, made inGermany), following the protocol of the manufac-turer. The whole ITS region including 5.8 S wasamplified using the two primers P17S (5¢ CTACCG ATT GAA TGG TCC GGT GAA 3¢) andP26S (5¢ TCC CGG TTC GCT CGC CGT TACTA 3¢) (Pop and Oxelman 2001) according to thefollowing protocol: to 1.5 lL template DNA, 1 lL10 pmol of each primer, 0.2 lL Taq DNA poly-merase, 2.5 lL l0 x puffer (included in the poly-merase kit), 2 lL MgCl2, 1.5 lL 2 mM/L dNTPS,were added with 15.3 lL H2O to a total volume of25 lL. Cycling included heating to 97�C for90 sec., followed by 36 thermal cycles, each with
124 N. S. Hassan et al.: Sesuvioideae
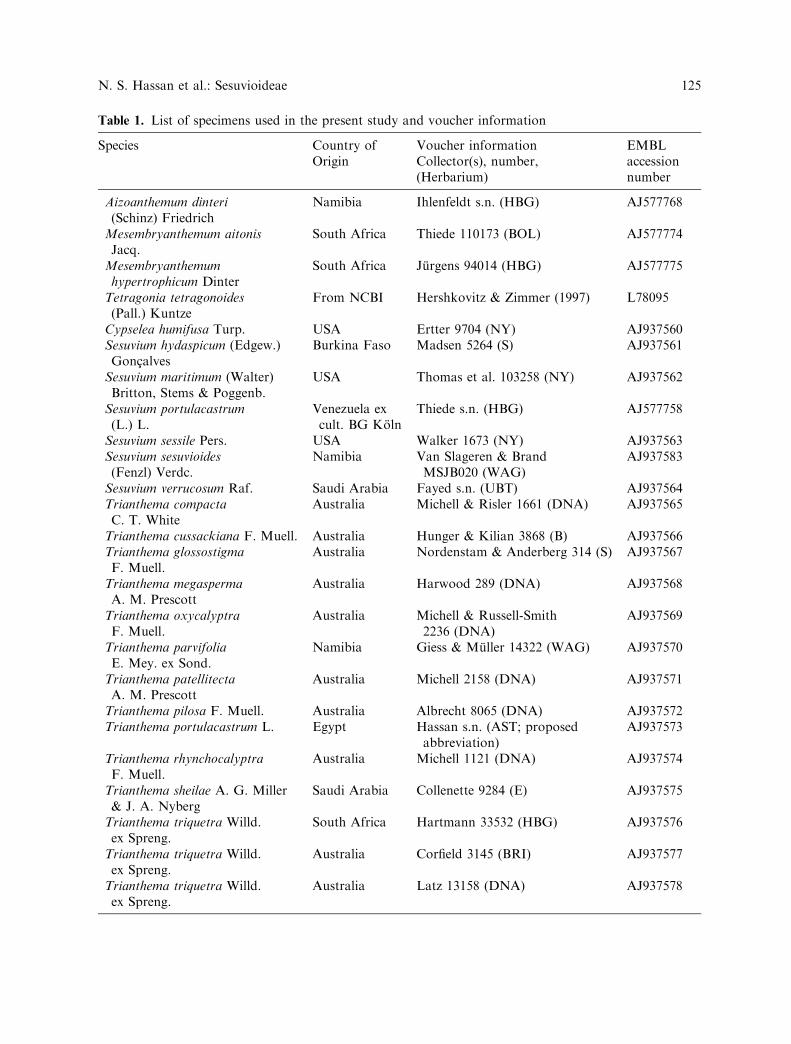
Table 1. List of specimens used in the present study and voucher information
Species Country ofOrigin
Voucher informationCollector(s), number,(Herbarium)
EMBLaccessionnumber
Aizoanthemum dinteri(Schinz) Friedrich
Namibia Ihlenfeldt s.n. (HBG) AJ577768
Mesembryanthemum aitonisJacq.
South Africa Thiede 110173 (BOL) AJ577774
Mesembryanthemumhypertrophicum Dinter
South Africa Jurgens 94014 (HBG) AJ577775
Tetragonia tetragonoides(Pall.) Kuntze
From NCBI Hershkovitz & Zimmer (1997) L78095
Cypselea humifusa Turp. USA Ertter 9704 (NY) AJ937560Sesuvium hydaspicum (Edgew.)Goncalves
Burkina Faso Madsen 5264 (S) AJ937561
Sesuvium maritimum (Walter)Britton, Stems & Poggenb.
USA Thomas et al. 103258 (NY) AJ937562
Sesuvium portulacastrum(L.) L.
Venezuela excult. BG Koln
Thiede s.n. (HBG) AJ577758
Sesuvium sessile Pers. USA Walker 1673 (NY) AJ937563Sesuvium sesuvioides(Fenzl) Verdc.
Namibia Van Slageren & BrandMSJB020 (WAG)
AJ937583
Sesuvium verrucosum Raf. Saudi Arabia Fayed s.n. (UBT) AJ937564Trianthema compactaC. T. White
Australia Michell & Risler 1661 (DNA) AJ937565
Trianthema cussackiana F. Muell. Australia Hunger & Kilian 3868 (B) AJ937566Trianthema glossostigmaF. Muell.
Australia Nordenstam & Anderberg 314 (S) AJ937567
Trianthema megaspermaA. M. Prescott
Australia Harwood 289 (DNA) AJ937568
Trianthema oxycalyptraF. Muell.
Australia Michell & Russell-Smith2236 (DNA)
AJ937569
Trianthema parvifoliaE. Mey. ex Sond.
Namibia Giess & Muller 14322 (WAG) AJ937570
Trianthema patellitectaA. M. Prescott
Australia Michell 2158 (DNA) AJ937571
Trianthema pilosa F. Muell. Australia Albrecht 8065 (DNA) AJ937572Trianthema portulacastrum L. Egypt Hassan s.n. (AST; proposed
abbreviation)AJ937573
Trianthema rhynchocalyptraF. Muell.
Australia Michell 1121 (DNA) AJ937574
Trianthema sheilae A. G. Miller& J. A. Nyberg
Saudi Arabia Collenette 9284 (E) AJ937575
Trianthema triquetra Willd.ex Spreng.
South Africa Hartmann 33532 (HBG) AJ937576
Trianthema triquetra Willd.ex Spreng.
Australia Corfield 3145 (BRI) AJ937577
Trianthema triquetra Willd.ex Spreng.
Australia Latz 13158 (DNA) AJ937578
N. S. Hassan et al.: Sesuvioideae 125

denaturation step at 97�C for 20 sec., annealingstep at 69�C for 90 sec., extention step at 72�C for90 sec. A final extention step at 72�C for 7 min.followed.
The PCR products were cleaned with the QIAquick� PCR Purification Kit (QIAGEN, made inGermany) following the manufacturer’s protocol.Sequencing reactions were done in a total volumeof 10 lL containing of 4 lL dye terminator pre-mix and 0.7 lL 5 pmol primer; the volume ofpurified DNA ranged from 1 to 4 lL depending onconcentration, on the expense of water volume. Thesequencing reaction was run with 26 thermal cycles;each cycle starting at 96�C for 30 sec., 50�C for15 sec. and 60�C for 4 min. Afterwards the DNAproducts were precipitated by addition of 250 lL95 % ethanol and 10 lL 3 M/L (PH 4.6) Na-acetate, then centrifuged for 10 min. (at14000 rpm) at room temperature. The ethanolwas discarded and the precipitated pellet waswashed twice with 80% ethanol, then the pelletwas dried in the oven at 30�C for 5 min. Finallyboth strands were sequenced using an ABI PrismModel 310 version 3.0 sequencer.
Morphological data. Morphological charactersand character states used in this analysis are basedon study of herbarium material (Table 1) whichincluded about 200 specimens from differentherbaria (AQ, B, DNA, E, K, NY, S, andWAG; complete list available from the corre-sponding author). These specimens were studiedwith a stereo-microscope (Wild M3) and 23variable characters (Table 2) were coded, compris-ing both vegetative and reproductive structures.Autapomorphic characters were excluded in thisanalysis and multistate characters coded as poly-morphisms. Seed characters were obtained fromSEM studies (for details, see Hassan et al., 2005).
Seed terminology follows Barthlott (1981, 1984)and Cutler (1979) with some modifications by theauthors.
Data analysis. Sequences were imported inMacClade version 4.0 (Maddison and Maddison2000), pre-aligned and trimmed both in the 26Sand 18S region to equal length. The boundariesof ITS1, 5.8S and ITS2 were defined throughcomparison with a sequence of Tetragonia tetrag-onoides (Hershkovitz and Zimmer 1997). Thealignment was adapted manually and a numberof gaps was required to align the studied taxa.Aligned sequences (available from the correspond-ing author and from TreeBASE (study accessionnumber = S1292, matrix accession numbers =M2261, M2262; Sanderson et al. 1994) included29 taxa and 653 characters. DNA sequences forall taxa studied have been submitted to EMBLand accession numbers are indicated in Table 1.The morphological matrix comprising 23 charac-ters (Table 3) was added to the molecular matrixafter trimming all species with more than oneaccession. From the results of molecular analysis,the polyphyly of Trianthema triquetra was obvi-ous, so that one African and one Australianrepresentative were coded for morphologicalcharacters (Table 3). In addition, the strict con-sensus tree obtained in the molecular analysis wasimported into MacClade version 4.0 (Maddisonand Maddison 2000) and the morphologicalcharacters were plotted on this tree.
The molecular matrix and the combined matrixwere analyzed using PAUP version 4.0b8a (Swof-ford 1998) on a power MacIntosh G3 computer.Sequence divergences (Table 4) were calculatedusing pairwise distances for ingroup and ingroup/outgroup taxa. Both data matrices were analyzedwith the heuristic search option with tree-bisection-
Table 1. (Continued)
Species Country ofOrigin
Voucher informationCollector(s), number,(Herbarium)
EMBLaccessionnumber
Trianthema triquetra Willd.ex Spreng.
Australia Michell & Russell-Smith2223 (DNA)
AJ937579
Trianthema turgidifolia F. Muell. Australia Latz 12861 (DNA) AJ937580Zaleya galericulata (Melville)H. Eichler
Australia Michell & Risler 1717(DNA)
AJ937581
Zaleya pentandra (L.) C. Jeffrey Saudi Arabia Fayed s.n. (UBT) AJ937582
126 N. S. Hassan et al.: Sesuvioideae

reconnection (TBR) branch swapping with MUL-TREES options in effect. The robustness of theresults was estimated by bootstrap (BS) analysis(Felsenstein 1985) with 10,000 bootstrap replicatesand 100 random sequences addition replicates(options as above). We used the following catego-ries of bootstrap support: <50% unsupported,50%–74% weakly supported, 75%–84% moder-ately supported, 85%–100% highly supported.Second, decay values (DI; Bremer 1988, Donoghueet al. 1992) were computed using AutoDecayversion 4.0 (Eriksson 1998) in combination withthe reverse constraint option of PAUP*. Consis-tency index (CI), retention index (RI) and rescaledconsistency index (RC) were calculated underexclusion of uninformative characters. In the mainanalyses, all characters were given equal weight andgaps were treated as missing data. A furtheranalysis was conducted using the combined datasetand applying successive weighing of all charactersaccording to the rescaled consistency index. Afterconstancy was reached, the resulting tree wascompared to the consensus trees retrieved fromthe unweighted analyses. Then, the relative weightof the morphological characters was kept while theweight of the molecular characters was reset to 1and the resulting tree was again compared to theconsensus trees retrieved from the unweightedanalyses.
Results
ITS data. In angiosperm taxa, it has beenfound that ITS1 and ITS2 are less than 300base pair (bp) long, where ITS1 varies inlength from 187–298, while ITS2 ranges from187–252 bp (e.g. Baldwin et al. 1995, Downieand Katz-Downie 1996). The length range ofboth ITS1 (208–232) and ITS2 (199–214) lieswithin that range and match closely those
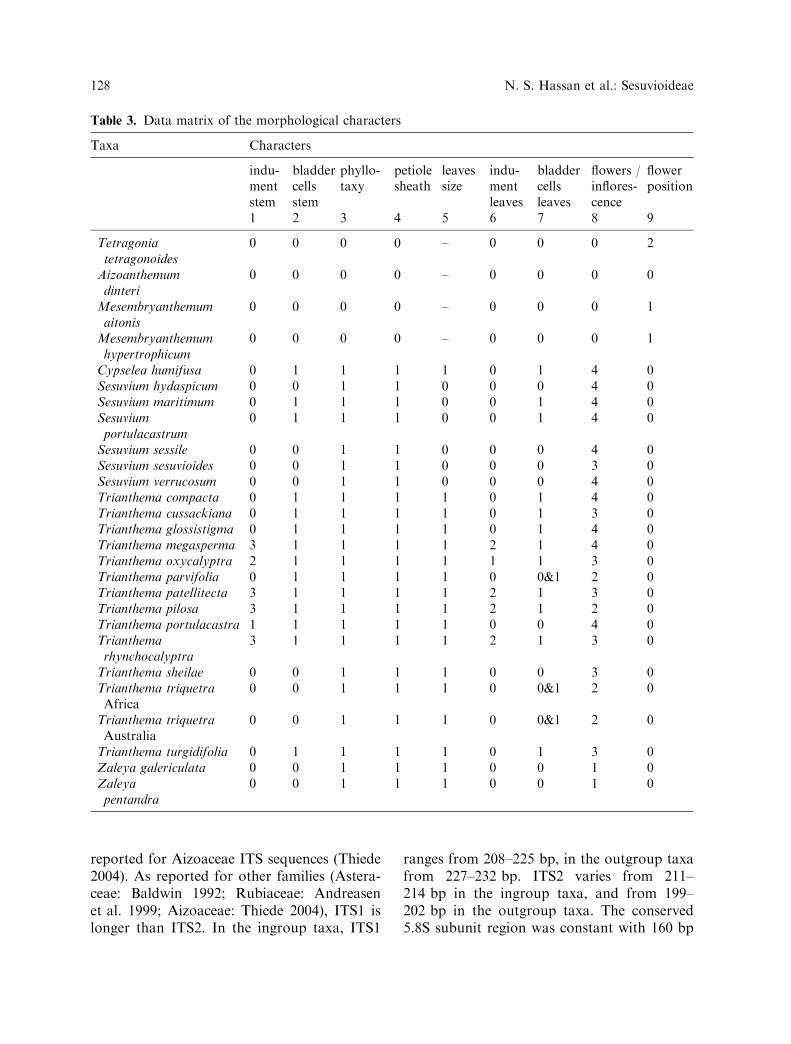
Table 2. Characters and their states used in themorphological analysis
1. Indumentum on the stem surface: 0– ab-sent 1– pubescent on young twigs only 2–sparsely pubescent all over 3– denselypubescent all over
2. Bladder cell idioblasts on the stem surface:0– present 1– absent
3. Phyllotaxy: 0– alternate 1– opposite4. Petiole at the base: 0– without membra-
nous sheath 1– with membranous sheath5. Leaves of pair (for opposite leaves only):
0– ±equal 1– unequal6. Indumentum on the leaf surface: 0– ab-
sent 1– pubescent on the midrib belowand at the apex 2– pubescent all over
7. Bladder cell idioblasts on the leaf surface:0– present 1– absent
8. Flowers per inflorescence: 0– plant is in-florescence 1– more than ten 2– four tonine 3– one to three 4– always one
9. Flowers position: 0– perigynous 1– semi-epigynous 2– epigynous
10. Pedicel: 0– present 1– absent11. Indumentum on the outside surface of
perianth: 0– always absent 1– usuallysparsely pubescent on the tube only 2–ciliate on the apex only 3– densely pub-escent all over
12. Number of stamens: 0– ten or more 1– five- nine 2– less than five
13. Number of stigmas: 0– five 1– four 2–three 3– two 4– one
14. Placentation type: 0– axillary 1– basal 2–apical
15. Fruit: 0– hygrochastic 1– nut 2–circumscissile
16. Top of gynoecium in fruiting stage: 0–without projections 1– with 3-5 projec-tions around the flat top 2– with un-dulating rim around the top
17. Operculum: 0– not splitting 1– splitting18. Seed number: 0– more than twelve 1– four
– twelve 2– one- three19. Seed shape: 0– orbicular 1– suborbicular
2– ± ovoid 3– reniform 4– D- shaped 5–± pear-shaped
20. Seed color: 0– shiny black 1– black 2–dark brown 3– light brown
21. Seed coat surface: 0– ± strongly ridged allover 1– slightly ridged along the dorsalside 2–slightly ridged only towards thehilum 3– not ridged
22. Papillae: 0– absent 1– present23. Aril: 0– with aril 1– without aril
N. S. Hassan et al.: Sesuvioideae 127
reported for Aizoaceae ITS sequences (Thiede2004). As reported for other families (Astera-ceae: Baldwin 1992; Rubiaceae: Andreasenet al. 1999; Aizoaceae: Thiede 2004), ITS1 islonger than ITS2. In the ingroup taxa, ITS1
ranges from 208–225 bp, in the outgroup taxafrom 227–232 bp. ITS2 varies from 211–214 bp in the ingroup taxa, and from 199–202 bp in the outgroup taxa. The conserved5.8S subunit region was constant with 160 bp
Table 3. Data matrix of the morphological characters
Taxa Characters
indu-mentstem
bladdercellsstem
phyllo-taxy
petiolesheath
leavessize
indu-mentleaves
bladdercellsleaves
flowers /inflores-cence
flowerposition
1 2 3 4 5 6 7 8 9
Tetragoniatetragonoides
0 0 0 0 – 0 0 0 2
Aizoanthemumdinteri
0 0 0 0 – 0 0 0 0
Mesembryanthemumaitonis
0 0 0 0 – 0 0 0 1
Mesembryanthemumhypertrophicum
0 0 0 0 – 0 0 0 1
Cypselea humifusa 0 1 1 1 1 0 1 4 0Sesuvium hydaspicum 0 0 1 1 0 0 0 4 0Sesuvium maritimum 0 1 1 1 0 0 1 4 0Sesuviumportulacastrum
0 1 1 1 0 0 1 4 0
Sesuvium sessile 0 0 1 1 0 0 0 4 0Sesuvium sesuvioides 0 0 1 1 0 0 0 3 0Sesuvium verrucosum 0 0 1 1 0 0 0 4 0Trianthema compacta 0 1 1 1 1 0 1 4 0Trianthema cussackiana 0 1 1 1 1 0 1 3 0Trianthema glossistigma 0 1 1 1 1 0 1 4 0Trianthema megasperma 3 1 1 1 1 2 1 4 0Trianthema oxycalyptra 2 1 1 1 1 1 1 3 0Trianthema parvifolia 0 1 1 1 1 0 0&1 2 0Trianthema patellitecta 3 1 1 1 1 2 1 3 0Trianthema pilosa 3 1 1 1 1 2 1 2 0Trianthema portulacastra 1 1 1 1 1 0 0 4 0Trianthemarhynchocalyptra
3 1 1 1 1 2 1 3 0
Trianthema sheilae 0 0 1 1 1 0 0 3 0Trianthema triquetraAfrica
0 0 1 1 1 0 0&1 2 0
Trianthema triquetraAustralia
0 0 1 1 1 0 0&1 2 0
Trianthema turgidifolia 0 1 1 1 1 0 1 3 0Zaleya galericulata 0 0 1 1 1 0 0 1 0Zaleyapentandra
0 0 1 1 1 0 0 1 0
128 N. S. Hassan et al.: Sesuvioideae
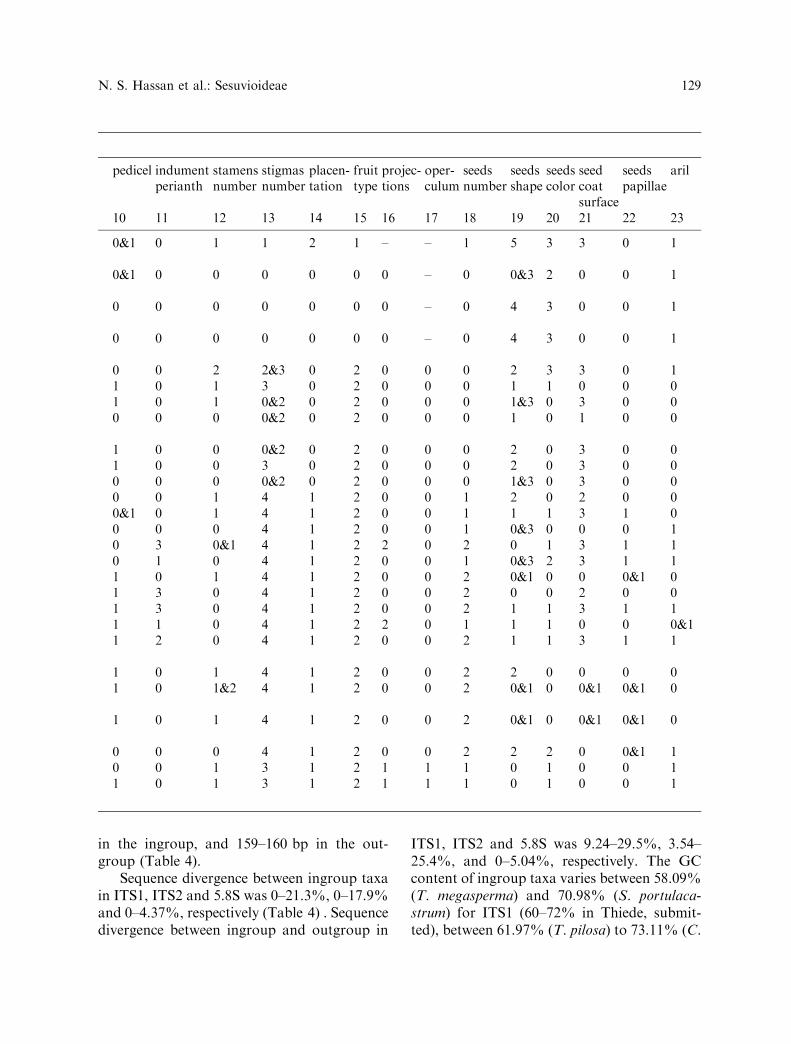
in the ingroup, and 159–160 bp in the out-group (Table 4).
Sequence divergence between ingroup taxain ITS1, ITS2 and 5.8S was 0–21.3%, 0–17.9%and 0–4.37%, respectively (Table 4) . Sequencedivergence between ingroup and outgroup in
ITS1, ITS2 and 5.8S was 9.24–29.5%, 3.54–25.4%, and 0–5.04%, respectively. The GCcontent of ingroup taxa varies between 58.09%(T. megasperma) and 70.98% (S. portulaca-strum) for ITS1 (60–72% in Thiede, submit-ted), between 61.97% (T. pilosa) to 73.11% (C.
pedicel indumentperianth
stamensnumber
stigmasnumber
placen-tation
fruittype
projec-tions
oper-culum
seedsnumber
seedsshape
seedscolor
seedcoatsurface
seedspapillae
aril
10 11 12 13 14 15 16 17 18 19 20 21 22 23
0&1 0 1 1 2 1 – – 1 5 3 3 0 1
0&1 0 0 0 0 0 0 – 0 0&3 2 0 0 1
0 0 0 0 0 0 0 – 0 4 3 0 0 1
0 0 0 0 0 0 0 – 0 4 3 0 0 1
0 0 2 2&3 0 2 0 0 0 2 3 3 0 11 0 1 3 0 2 0 0 0 1 1 0 0 01 0 1 0&2 0 2 0 0 0 1&3 0 3 0 00 0 0 0&2 0 2 0 0 0 1 0 1 0 0
1 0 0 0&2 0 2 0 0 0 2 0 3 0 01 0 0 3 0 2 0 0 0 2 0 3 0 00 0 0 0&2 0 2 0 0 0 1&3 0 3 0 00 0 1 4 1 2 0 0 1 2 0 2 0 00&1 0 1 4 1 2 0 0 1 1 1 3 1 00 0 0 4 1 2 0 0 1 0&3 0 0 0 10 3 0&1 4 1 2 2 0 2 0 1 3 1 10 1 0 4 1 2 0 0 1 0&3 2 3 1 11 0 1 4 1 2 0 0 2 0&1 0 0 0&1 01 3 0 4 1 2 0 0 2 0 0 2 0 01 3 0 4 1 2 0 0 2 1 1 3 1 11 1 0 4 1 2 2 0 1 1 1 0 0 0&11 2 0 4 1 2 0 0 2 1 1 3 1 1
1 0 1 4 1 2 0 0 2 2 0 0 0 01 0 1&2 4 1 2 0 0 2 0&1 0 0&1 0&1 0
1 0 1 4 1 2 0 0 2 0&1 0 0&1 0&1 0
0 0 0 4 1 2 0 0 2 2 2 0 0&1 10 0 1 3 1 2 1 1 1 0 1 0 0 11 0 1 3 1 2 1 1 1 0 1 0 0 1
N. S. Hassan et al.: Sesuvioideae 129

humifusa) for ITS2 (65–72 % in Thiede,unpublished data), and between 55%(T. cussackiana and T. oxycalyptra) to56.87% (Cypselea humifusa, S. maritimum,S. portulacastrum, S. sessile, S. sesuvioides,S. verrucosum, T. parvifolia, T. sheilae andT. triquetra) for 5.8S (55.1% in Thiede 2004).The mean GC content of the whole sequencedarea was 63.76% (65.3% in Thiede 2004).
The final molecular data matrix of sub-family Sesuvioideae included 29 OTUs, 653positions and 168 parsimony informative char-acters. Heuristic search for the whole ITSregion yielded 12 shortest trees with 546 steps,CI = 0.66 (excluding uninformative charac-ters), RI = 0.73 and RC = 0.48. The strictconsensus tree of these 12 MPT’s is shown inFig. 1.
ITS results strongly support the mono-phyly of subfamily Sesuvioideae (BS 100%and DI 34). The ITS tree topology suggeststhat Sesuvium L. and Trianthema L. areparaphyletic while Zaleya Burm.f. is mono-phyletic. The ITS cladogram (Fig. 1) can bedivided into two major clades: Sesuvium andTrianthema. The Sesuvium clade is weaklysupported (BS 71%, DI 1) and sister to theTrianthema clade which is unsupported (BS <50%, DI 1). Cypselea (C. humifusa) is nestedwithin Sesuvium (Fig. 1) and therefore is moreclosely related to Sesuvium than to Trianthemaor Zaleya. The Sesuvium clade comprises threesubclades. The single Cypselea specimen formsa subclade of its own (Fig. 1: II), while bothSesuvium subclades are strongly supported.The first Sesuvium subclade comprisesS. hydaspicum and S. sesuvioides (Fig. 1: I,BS 97%, DI 4). The second Sesuvium subclade,
which is sister to Cypselea, includes S. mariti-mum, S. portulacastrum, S. sessile and S. ver-rucosum and does not show any furtherresolution (Fig. 1: III, BS 97%, DI 4).
The Trianthema clade (Figs. 1,2) can bedivided into three subclades. The first subclade(Fig. 1: IV) comprises T. sheilae, T. parvifoliaand T. triquetra and receives moderate support(BS 84 %, DI 4). The second subclade (Fig. 1:V) comprising the two Zaleya species (Z. gale-riculata and Z. pentandra) is stronglysupported (BS 100%, DI 9). The third sub-clade, comprising all remaining species (Fig. I:VI) is weakly supported (BS 71%, DI 2).Within this subclade, only the terminalT. patellitecta, T. rhynchocalyptra, T. mega-sperma, and T. pilosa subclade shows somesupport (BS 71%, DI 3); all other internalbranches are without or with very weak (51–52%) bootstrap support.
Combined data. The combined matrix thusresulted in 27 OTUs, 676 positions and 189parsimony informative characters. Heuristicsearch yielded 40 shortest trees with 646 steps,CI = 0.56 (excluding uninformative charac-ters), RI = 0.72 and RC = 0.45. The strictconsensus tree of these 40 MPT’s is shown inFig. 2. The topology of the combined tree issimilar to that of the molecular tree, but withsome differences in support and resolution.The combined analysis (Fig. 2) supportsmonophyly of the ingroup even more stronglythan the molecular analysis (BS 100 %, DI 38).The Sesuvium clade, however, falls apart, withthe main Sesuvium subclade (Fig. 2: III) stillpresent, but with less support (BS 84%, DI 3)than in the purely molecular subclade. Sesuvi-um hydaspicum and S. sesuvioides form an
Table 4. Sequence characteristics
ITS1 ITS2 5.8S
Aligned total length (bp) 249 232 162Length range (bp) 208–232 199–214 159–160Length mean (bp) 220.03 210.45 159.97Sequence divergence (ingroup) (%) 0–21.3 0–17.9 0–4.37Sequence divergence (ingroup/outgroup) (%) 9.24–29.5 3.54–25.4 0–5.04
130 N. S. Hassan et al.: Sesuvioideae
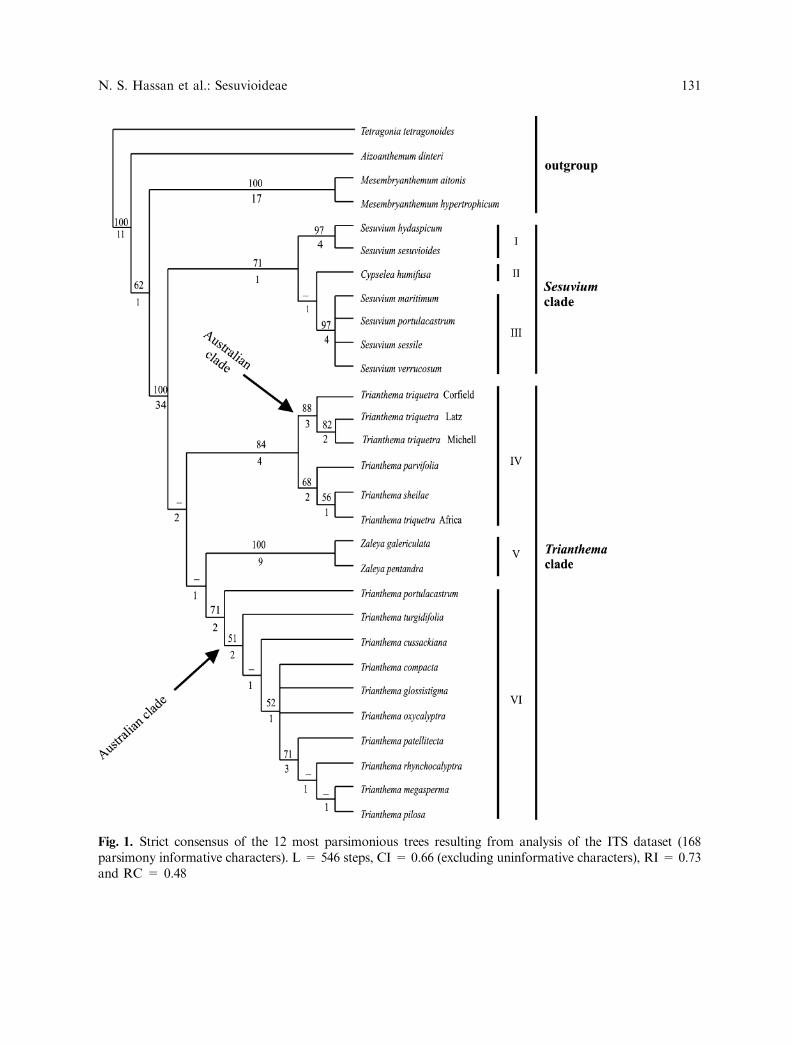
Fig. 1. Strict consensus of the 12 most parsimonious trees resulting from analysis of the ITS dataset (168parsimony informative characters). L = 546 steps, CI = 0.66 (excluding uninformative characters), RI = 0.73and RC = 0.48
N. S. Hassan et al.: Sesuvioideae 131
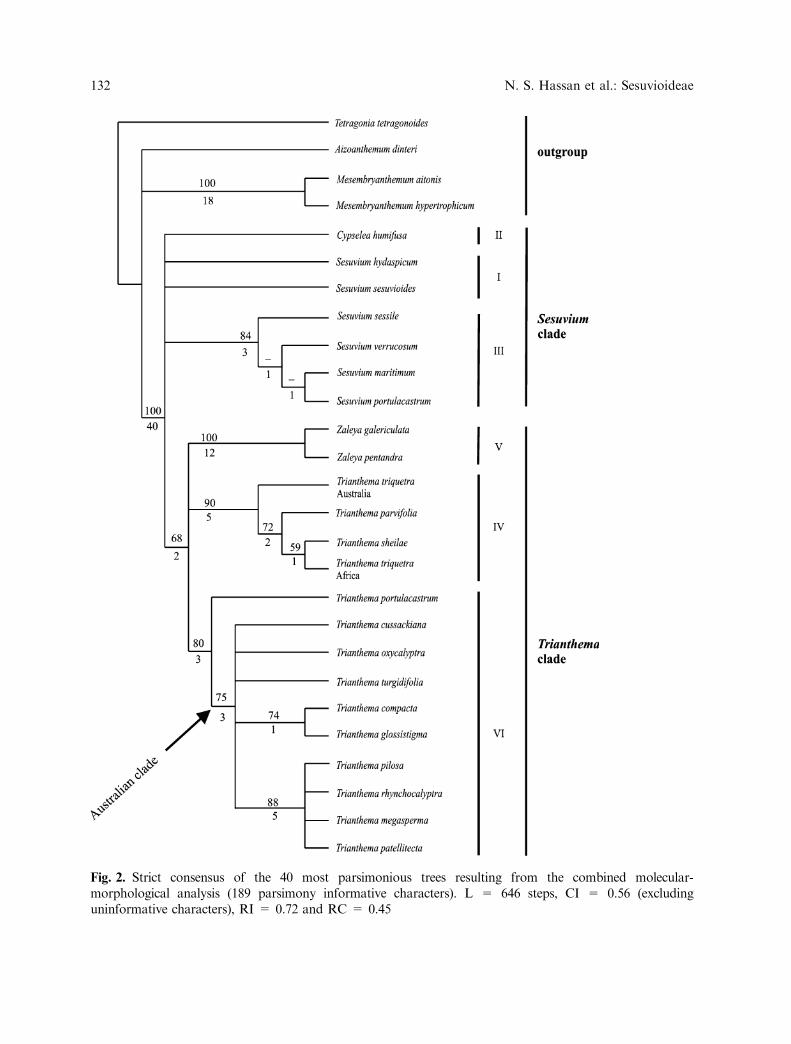
Fig. 2. Strict consensus of the 40 most parsimonious trees resulting from the combined molecular-morphological analysis (189 parsimony informative characters). L = 646 steps, CI = 0.56 (excludinguninformative characters), RI = 0.72 and RC = 0.45
132 N. S. Hassan et al.: Sesuvioideae

unresolved polytomy with subclade III, Cyps-elea humifusa and the Trianthema clade. Sup-port for the Trianthema clade is slightly betterthan in the molecular analysis (BS 68%, DI 2).Zaleya is placed in an unresolved polytomywith the two Trianthema subclades, and itssupport raises (DI 12 versus DI 9, Fig. 2).Support for subclade IV, the Papularia sub-clade, raises (from BS 84% DI 4 to BS 90% DI5) and a similar increase in support can beobserved for subclade VI (from BS 71% DI 2to BS 80% DI 3, Fig. 2). Most impressive isthe increase in support for the terminal groupT. megasperma, T. patellitecta, T. pilosa, andT. rhynchocalyptra, which raises from BS 71%DI 3 to BS 88% DI 5.
Constancy is reached after two rounds ofsuccessive reweighting and a single tree ofL=307.71 steps results (CI=0.76, RI=0.87,RC=0.73). In this tree, the same subclades asin the purely molecular analysis are retrieved,but Cypselea is sister to the two Sesuviumsubclades and Zaleya to the two Trianthemasubclades. The Cypselea/Sesuvium clade ismoderately supported with BS 79% DI 1.4,the Zaleya/Trianthema clade with BS 81% DI1.9. Of the 23 morphological characters, only 9remain with a weight of 1 (1, 3–6, 9, 14–15, 17).If the weight of all molecular characters is resetto 1, two shortest trees result, and the consen-sus is identical to the ITS tree shown in Fig. 1.
Morphological characters were plotted onthe ITS tree to evaluate the morphologicaltraits with respect to phylogenetical signal.Some of the characters coded, such as oppositeleaves, presence of a membranous sheathtowards the petiole base and circumscissilecapsules support the monophyly of Sesuvioi-deae (Fig. 3a). Possession of one stigma(Fig. 4a) characterizes Trianthema, basal pla-centation supports the clade of Zaleya andTrianthema, while a splitting operculum isunique for Zaleya. Some characters supportclades also retrieved in the molecular analysis,for instance, presence of indumentum on theplant surface characterizes the T. megasperma,T. patellitecta, T. pilosa, and T. rhynchocalyp-tra group (Fig. 5b). However, most other
characters, in particular the seed characters,are variable throughout Sesuvioideae and thusnot useful for phylogenetic conclusions(Figs. 3b, 4b, 5a). Figure 4b also reveals thatarillate seeds do not represent a synapomorphyfor Sesuvioideae as stated by Bittrich andHartmann (1988), but are instead restricted tohalf of the species only and of multiple origin.By successive weighting, characters 10 (pres-ence of pedicel), 22 (presence of papillae ontesta), 19 (seed shape), 12 (number of sta-mens), and 23 (presence of aril, Fig. 4b) wereidentified as the characters with the worst fit(weight assigned < 0.1; listed in sequence).
Discussion
Our study of both a molecular and a morpho-logical dataset confirms the monophyly ofSesuvioideae, as suggested by Bittrich (1990).Undoubtedly, the congruence between ITSand combined data displays that the resultsare stable to a large extent.
The monophyly of Sesuvioideae at least intheir old circumscription (without Tribulocar-pus) is impressively demonstrated (BS 100% inboth analyses and DI 34 and 40). Morpholog-ically, stipule-like appendages and circumscis-sile capsules are probably the most reliablemorphological synapomorphies (Fig. 3a: char-acter 4).
The ingroup is split into two groups,Sesuvium-Cypselea and Trianthema-Zaleya.The close relationship of Trianthema andZaleya is reflected in the taxonomical historyduring which Zaleya was split from andreunited with Trianthema several times (seeintroduction). The most surprising result ofthis study, however, is the position of Cypselea(subclade II) within the Sesuvium clade in theITS tree (Fig. 1) and in both trees resultingfrom reweighting either all or the morpholog-ical characters. In the unweighted combinedtree, Cypselea and Sesuvium are no moremonophyletic, reflecting the absence of mor-phological synapomorphies for the group.Cypselea is represented in our study by onlyone of the three species (C. humifusa) and can
N. S. Hassan et al.: Sesuvioideae 133

134 N. S. Hassan et al.: Sesuvioideae

be circumscribed by a combination of charac-ters: very slender and low-growing herbs;leaves opposite, very variable in size, petioleat least as long as the blade, basally withlaciniate stipule-like appendages; perianth veryshort (up to 1.5 mm long), lobes withoutdorsal appendages, stamens 2–3, and septaincomplete in mature fruit (Hartmann 1993,2001a). The seed coat of Cypselea is unique inSesuvioideae with its raised reticulate surfaceassociated with a raised cuticle layer above theanticlinal boundaries (Hassan et al., 2005).Morphologically, the relationship betweenCypselea and Sesuvium is best supported byaxillary placentation and many-seeded cap-sules (Fig. 3b: character 18). However, axillaryplacentation most likely constitutes a plesio-morphic character because it is the commoncondition in all other subfamilies of Aizoaceaeexcept for the Ruschioideae; and the sameholds true for many-seeded capsules, which arefound in all subfamilies. Reduction of flowernumber per inflorescence to one or very few isprobably apomorphic for Sesuvium-Cypselea,but occurs within Trianthema as well. InTrianthema, the number of flowers – besidethe number of seeds – was considered byJeffrey (1960) to form a base for the distinctionof two subgenera, but the present study showsthat reduction o flowers per inflorescence isprobably subject to relatively rapid evolution.
Hofmann (1973: 289) notes that onlySesuvium and Cypselea possess simple opercu-late capsules, while Trianthema und Zaleyashow some additional features of indehiscence:in Trianthema portulacastrum, a secondaryhorizontal ring is formed in the ovary, dividingthe fruit in two unequal storeys (Payer 1957after Hofmann 1973). Ovules in the upperstorey remain enclosed in the operculum upondehiscence of the capsules. In other species ofTrianthema, however, the seeds lie loosely inthe carpels and fall out once the lid is lifted
(Hartmann, pers. comm.). In Zaleya, a verticaldivision results in more or less completesecondary walls, and the operculum is splitalong these (Melville 1952); each chambercontains one seed.
Muller (1909: p. 91) states that the twogroups differ in the possession of a sclerenchy-matic ring being present in the stems ofSesuvium and Cypselea, but absent in Trian-thema (incl. Zaleya). As such a ring is presentin a number of genera now all considered tobelong to Molluginaceae, while it is absent inthe other genera now considered Aizoaceae aswell (Mesembryanthemum and Tetragonia;Muller 1909), the presence of the ring has tobe considered synapomorphic in the context ofAizoaceae and might thus constitute a validsynapomorphy for the Sesuvium-Cypseleaclade. However, other anatomical characters,such as the type of the anomalous secondarygrowth in stems and the deposition of calciumoxalate, are not supportive of these two clades(Muller 1909). Trianthema (incl. Zaleya),together with Mesembryanthemum (sensu Mul-ler 1909), and Acrosanthes (Aizooideae) showsanomalous secondary growth of the Mesem-bryanthemum-type with single strands of vas-cular tissue, while Cypselea, together withAizoon and Tetragonia, possesses the Tetra-gonia-type with complete rings of xylem andphloem. Sesuvium, finally, shows a mixed type.However, as the genera of Molluginaceaestudied (Muller 1909) are equally ambiguousas to the type of secondary growth, thischaracter is probably not of systematic valueat higher ranks than species. Calcium oxalatecrystals are deposited as druses in both Sesu-vium and Trianthema (incl. Zaleya), as well asin Aizoon and Tetragonia, whereas only singlecrystals are present in Cypselea and raphids areformed in Mesembryanthemum, i.e. Mesem-bryanthemoideae and Ruschioideae (Muller1909).
Fig. 3. Distribution patterns of morphological characters on the strict consensus tree derived from the ITSdataset (Fig. 1). 3a. Petiole base (synapomorphy for Sesuvioideae). 3b. Seed number (synapomorphy forTrianthema/Zaleya)
b
N. S. Hassan et al.: Sesuvioideae 135

Fig. 4a. Stigma number [synapomorphy for Trianthema (1) and for subclade I within Sesuvium (2)]. 4b.
Presence of an aril (synapomorphy for subclade III within Sesuvium and for subclade IV within Trianthema)
136 N. S. Hassan et al.: Sesuvioideae
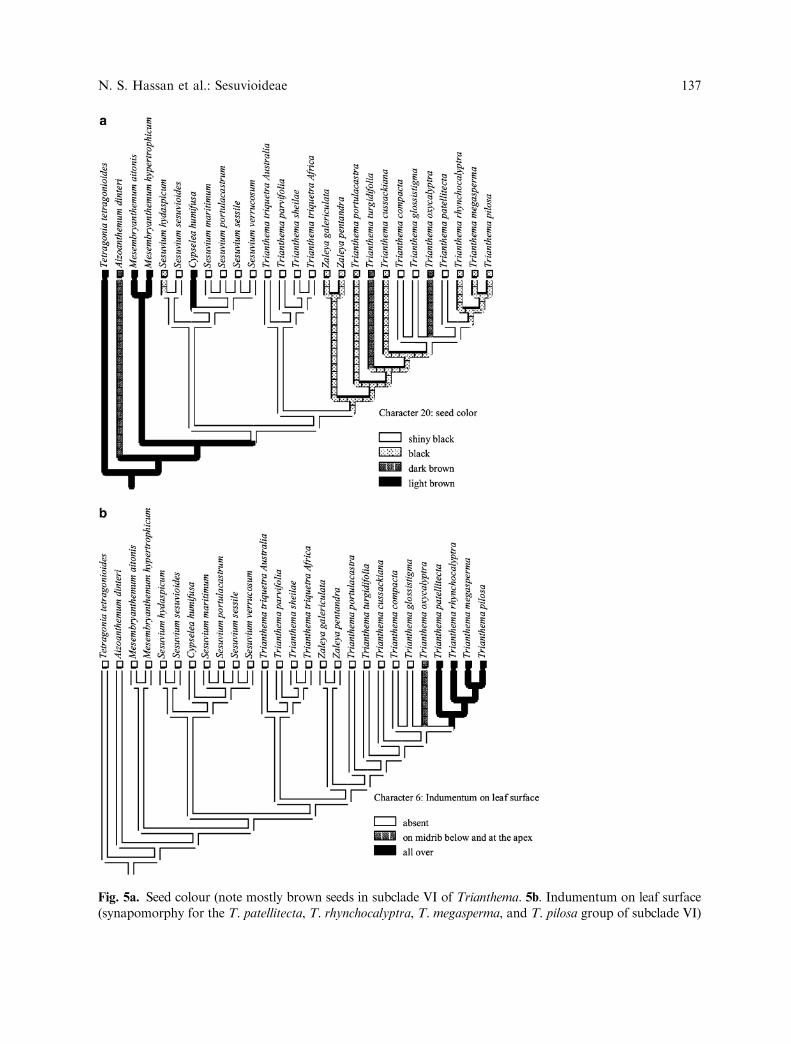
Fig. 5a. Seed colour (note mostly brown seeds in subclade VI of Trianthema. 5b. Indumentum on leaf surface(synapomorphy for the T. patellitecta, T. rhynchocalyptra, T. megasperma, and T. pilosa group of subclade VI)
N. S. Hassan et al.: Sesuvioideae 137

Sesuvium includes ca. 12 species (Gilbertet al. 2000) with a world wide tropical distri-bution (cf. Hartmann 2001b). They often growin saline soils in coastal or inland habitats(Hartmann 2001b). Numerous seeds, 2–5 stig-mas, a short taproot (Hartmann 2001b),sometimes stems with adventitious roots atthe nodes and usually uniflorous inflorescencesare considered the main character states per-mitting the identification of the genus. Sesuvi-um in its present circumscription falls into twosubclades. Subclade I , comprising S. hydaspi-cum and S. sesuvioides, is highly supported bythe ITS analysis, but not retrieved by theunweighted combined analysis (Figs. 1, 2).Reduction of stigma number to two (Fig. 4a)can be considered a morphological synapo-morphy for subclade I within Sesuvium, inwhich the common number of stigmas is 3–5.Furthermore, the two species are characterizedby a perianth tube with stout appendages,rough internodes (after drying) and a ± ellip-soid fruit. However, the two species differ inthe number of stamens (ranging from 5–7 inS. hydaspicum, but numerous in S. sesuvioides),and seed character states (arillate smooth seedsin S. sesuvioides versus non-arillate ridgedseeds in S. hydaspicum). Sesuvium hydaspicumwas frequently placed in synonymy ofS. sesuvioides (e.g. Gilbert et al. 2000, Hart-mann 2001b), but Goncalves (1965) restrictedS. sesuvioides to Angolan and Namibian spec-imens with smooth seeds. Our two specimens,S. sesuvioides from Namibia (Table 1) andS. hydaspicum from Burkina Faso, whichindeed possess ridged seeds and seven stamens,support the concept of Goncalves (1965) toseparate the two taxa as species. Sequencedivergence amounts to ca. 0.8% betweenS. sesuvioides and S. hydaspicum, and is thusin the same order of magnitude as between theundisputed species of subclade II and, there-fore, not indicative for conspecifity (sequencedivergence between species in different sub-clades amounts to ca. 2–3%) . However, astudy comprising numerous accessions in thecomplex over its whole distribution rangewould be desirable.
Subclade III of Sesuvium (S. maritimum,S. portulacastrum, S. sessile and S. verrucosum)is also better supported by ITS data analysisthan by the combined data (Fig. 2), indicatingvariability in morphological features. The fourabove species share the features of numerous,smooth, shiny black arillate seeds, but suchseeds occur also in S. sesuvioides and in someTrianthema species (Fig. 5a). In floral charac-ters, the members of subclade III are ratherdiverse. The occurrence of three to five stigmasis shared only with Cypselea (Fig. 4a), but fivelocules are considered a plesiomorphic featurefor the entire family. The number of stamensvaries from five in S. maritimum to numerousin the remaining taxa, but again, five stamensconstitute the plesiomorphic condition alsofound in the other Aizoaceae subfamiliesexcept for Tetragonia and Gunniopsis. Almostsessile flowers are found in S. sessile,S. maritimum and S. verrucosum, but those ofS. portulacastrum sensu Hartmann (2001b)possess a distinct pedicel of 5–10 mm length.All four taxa grow in similar habitats on thesea shore (Hartmann 2001b), except forS. verrucosum, which is also reported as aninvader and a pioneer of reclaimed land(Phillips 1988).
Trianthema and Zaleya share the synapo-morphic character of a reduced number ofseeds (Fig. 3b), and perhaps an incompletehorizontal division of the locules. Trianthemais widely distributed in dry areas of tropicaland subtropical regions and can best becharacterized by the synapomorphic characterstate of a single stigma (Fig. 4a) and a singlelocule (e.g. Hartmann 1993). Of the approx. 20species worldwide, ten are endemic to Austra-lia (e.g. Prescott and Venning 1984, Gilbert2000). The circumscription and subdivision ofTrianthema has changed in the course ofhistory. Candolle (1828) split Trianthema intotwo sections, Trianthema section Zaleya withten stamens and Trianthema section Rocamawith five stamens. Jeffrey (1960) recognizedZaleya, based on fruit construction, anddivided Trianthema into two subgenera, sub-gen. Trianthema with four ovules and usually
138 N. S. Hassan et al.: Sesuvioideae

uniflorous inflorescences, and subgen. Papu-laria (Forssk.) C. Jeffrey ex Adamson with twoovules, one above the other, and cymoseinflorescences. The type species of Trianthema,T. portulacastrum, is placed at the base ofsubclade VI, so that this subclade is the typicalone.
Hofmann (1973: p. 260) states that subgen.Papularia shares with Zaleya dichotomouslybranched inflorescences. Unfortunately, nei-ther Jeffrey (1960) nor any of the subsequentworkers stated explicitly which species belongto subgen. Papularia apart from the typespecies, T. crystallina Forssk. Of T. crystallina,however, no sequencable material was avail-able, so the question whether the namePapularia should be applied to subclade IV(Figs. 1, 2) or not, cannot be answered withcertainty at present. The name Papularia isavailable, because Papularia E. M. Fries(1825), a member of the Dematiaceae(Hyphoymycetes), is a younger homonym.Whereas the species of subclade IV(T. parvifolia, T. sheilae and T. triquetra) showtwo ovules (Fig. 3b) and cymose inflorescencesdefining Papularia, these character states arealso found in subclade VI. Trianthemacrystallina, the type of Papularia, possessesprominent bladder cell idioblasts on bothstems and leaves, but the name has been usedfor plants of T. triquetra with sparse and smallbladder cells as well. On the other hand,T. parvifolia and T. sheilae develop distinctbladder cells on the leaf blades, whereas otherspecies show bladder cells restricted tothe margins, e.g. T. salsoloides. Originally,T. triquetra had been described as a veriety ofof T. crystallina by Willdenow (1803) empha-sizing the close relationship between the twospecies. Thus, subclade IV almost certainlycan be considered as subgen. PapulariaC. Jeffrey. The situation is further confusedby the fact that Willdenow (1803) in annotat-ing Rottler’s report of a trip in India, onwhich the material named provisionallyT. triquetra was collected, considered T. triqu-etra as ‘‘Spielart’’ (= variety) of T. crystallina.Later, Sonder (1862) used the name T. crystal-
lina for two varieties from southern Africawhich lack bladder cells at first inspection; atthe same time, Sonder (1862) establishedT. parvifolia with papillate leaves.
Subclade IV (T. parvifolia, T. sheilae andT. triquetra) is well supported by ITS andcombined results (Fig. 2) and also morpholog-ically well defined by (almost) sessile flowersarranged in three-flowered clusters and thepossession of five stamens. However, the lastcharacter shows some variation in T. triquetra(5 stamens in both Asian and Australianmaterial to less than 5 [(2) )3 ()4)] in Africanexcept for Sudanese, Angolan, and Namibianmaterial; Hassan, unpubl. data). Members ofsubclade IV also share arillate, shiny blackseeds (Figs. 4b, 5a) with a ridged surface andpenta-hexagonal epidermal cells (Hassan et al.,2005), but again, these character states areneither apomorphic nor unique in Sesuvioi-deae. The relationships of T. parvifolia andT. triquetra are still confused, and the formerwas often considered a subspecies of the latter(e.g. Jeffrey 1960, 1961; Friedrich 1970).Recently, Hartmann (2001b) recognized T. par-vifolia as a distinct species, a conclusionconfirmed by our results. The material inves-tigated by her as T. triquetra from Africa ismore closely related to Trianthema sheilaefrom Ethiopia, Saudi Arabia and Yemen(acc. to Chaudhary 1999) than to T. triquetrafrom Australia according to both trees(Figs. 1, 2). The two specimens of T. triquetraof the same geographical origin, NorthernAustralia, form a monophyletic clade which issister to the specimen from Queensland(Fig. 1). Bittrich (1990) mentions the occur-rence of hygrochastic tepals in T. triquetra andrelated species (as well as in some Aizooideae;e.g. Aizoon giessii Friedrich and related spe-cies); these may be of systematic importance inthis subclade, but our material was insufficientfor an assessment. Subclade VI is weaklysupported by ITS data (Fig. 1) but supportincreases in the combined analysis (Fig. 2).The only reasonably supported group withinthis clade consists of T. megasperma,T. patellitecta,T. pilosa, andT. rhynchocalyptra.
N. S. Hassan et al.: Sesuvioideae 139

These species share the apomorphic characterof a dense indumentum on internodes, leaves(Fig. 5b) and perianth. Seed surface is papil-lose in T. megaspema, T. pilosa andT. rhynchocalyptra, but smooth to very slightlyridged towards the hilum in T. patellitecta(Hassan et al., 2005). All four taxa occur inAustralia, and both molecular and morpho-logical data support their monophyly. Thisgroup of taxa shares with subclade IV thereduced number of seeds (Fig. 3b) indicatingthat a reduction of seed number must haveoccurred at least twice in Trianthema. Allspecies of subclade VI come from Australia,except for T. portulacastrum, which is sister tothe remainder of the clade and is foundworldwide. Whether the widespread T. portul-acastrum originated in Australia, or whetherthe Australian species have radiated followingcolonization of the continent by a T. portula-castrum-like ancestor cannot be decided on thebase of the material studied here.
Our molecular results (Fig. 1) support theclose relationship between Zaleya and Trian-thema. The placement of Zaleya within Tri-anthema (Fig. 1: V) is supported by threecharacter states: unequal opposite leaves,basal placentation and few seeds. Zaleyawas included in Trianthema by some authorsof floristic treatments (e.g. Hutchinson andDalziel 1954, Tackholm 1956, Zohary 1966).However, in the combined analysis Zaleya isplaced in an unresolved polytomy togetherwith the two Trianthema clades (Fig. 2).Prescott and Venning (1984) mentioned thatZaleya probably comprises three closelyrelated species in tropical and subtropicalregions, and they expect that closer investi-gation will reveal that these three taxa con-stitute only one variable species. Recently,Gilbert et al. (2000) regards Zaleya to consistof six closely related species. Based on ITSand combined results, the monophyly of Z.galericulata and Z. pentandra is stronglysupported. Sequence divergence between thetwo species is less than 5%, and a number ofmorphological synapomorphies corroboratesthis clade in addition to the generic synapo-
morphies discussed above, namely, turret-shaped fruits with orbicular seeds with aclosely ridged surface. However, Z. pentandrahas short fruit projections (to 0.4 mm) andoccurs in Africa and Asia, while Z. galeric-ulata has longer fruit projections (0.7–1 mm),and a more easily splitting operculum, and itis restricted to Australia. A detailed popula-tion-level study over the whole distributionrange of Zaleya would therefore be necessarybefore species delimitation in Zaleya can beput on a sufficient data base.
The present study sheds some light on themain taxonomic problems in the traditionalSesuvioideae, the position of Cypselea andZaleya. Our results can be used to supportan inclusion of the two genera in a morebroadly conceived Sesuvium or Trianthema,respectively; but they can just as well beinterpreted to support a split of Sesuvium andTrianthema into (at least ?) two genera each,leaving Cypselea and Zaleya independent intheir present circumscriptions. As both pos-sibilities would require nomenclaturalchanges, and the present data are not favor-ing one of the two, no nomenclatural changesare proposed here.
The present work is part of the Ph.D. thesis ofthe first author. He acknowledges financial sup-port through the Egypt Channel System and theUniversity of Bayreuth. Prof. Fayed, Assiut Uni-versity, provided plant material. We are particu-larly grateful to the directors and correspondentsof all herbaria lending material for the morpho-logical study and / or granting permission to usetheir specimens for the molecular study. Wefurther thank Heidrun E. K. Hartmann, Univer-sity of Hamburg, for her support and criticalremarks.
References
Andreasen K., Baldwin B. G., Bremer B. (1999)Phylogenetic utility of the nuclear rDNA ITSregion in the subfamily Ixoroideae (Rubiaceae):comparison with cpDNA rbcL sequence data.Pl. Syst. Evol. 217: 119–135.
140 N. S. Hassan et al.: Sesuvioideae

Backer C. A. (1954) Aizoaceae. In: Van Steenis C.G. C. (ed.) Flora Malesiana, ser. 1, vol. 4.Leiden, pp. 266–274.
Baldwin B. G. (1992) Phylogenetic utility of theinternal transcribed spacer of nuclear ribosomalDNA in plants: An example from the Compos-itae. Molec. Phylogenet. Evol. 1: 3–16.
Baldwin B. G., Sanderson M. J., Porter J. M.,Wojciechowski M. F., Campbell C. S., Don-oghue M. J. (1995) The ITS region of nuclearribosomal DNA: a valuable source of evidenceon Angiosperm phylogeny. Ann. Missouri Bot.Gard. 82: 247–277.
Barthlott W. (1981) Epidermal and seed surfaceapplicability and some evolutionary aspects.Nord. J. Bot. 1: 345–355.
Barthlott W. (1984) Microstructural features andseed surface. In: Heywood V., Moore D.M.(eds.) Current concepts in plant taxonomy.Academic Press, London, pp. 95–105.
Bhambie S., Joshi M. C., Gupta M. L. (1977)Anatomical studies on certain members of Aizo-aceae. Proc. Indian Acad. Sci. 85B: 399–406.
Bittrich V. (1990) Systematic studies in Aizoaceae.Mitt. Inst. Allg. Bot. Hamburg 23b: 491–507.
Bittrich V., Hartmann H. E. K. (1988) TheAizoaceae – a new approach. Bot. J. Linn. Soc.97: 239–254.
Bittrich V., Struck M. (1989) What is primitive inthe Mesembryanthemaceae? An analysis of evo-lutionary polarity of characters states. S. Afri-can. J. Bot. 55: 321–331.
Bremer K. (1988) The limits of amino acidsequence data in angiosperm phylogenetic recon-struction. Evolution 42: 795–803.
Candolle A. P. de (1828) Prodromus SystematisNaturalis Regni Vegetabilis. Treuttel and Wurtz,Paris, 3: 415–455.
Chaudhary S. A. (1999) Flora of the Kingdom ofSaudi Arabia., vol. 1. Riyadh, pp.141–155.
Corner E. J. H. (1976) The seeds of Dicotyledonsvol.1. Cambridge University Press, Cambridge,pp. 66–67.
Cronquist A. (1981) An integrated system ofclassification of flowering plants. ColumbiaUniversity Press, New York.
CuenoudP. (2003) Introduction to expandedCaryo-phyllales. Conspectus of families of expandedCaryophyllales. In: Kubitzki K., Bayer C. (eds.)The families and genera of vascular plants.V. Flowering plants. Dicotyledons. Malvales,
Capparales, and non-betalain Caryophyllales.Springer, Berlin, pp. 1–6.
Cuenoud P., Savolainen V., Chatrou L. W., PowellM., Grayer R. J., Chase, M. W. (2002) Molecularphylogenetics of Caryophyllales based on nuclear18S rDNA and plastid rbcL, atpB, and matKDNA sequences. Amer. J. Bot. 89: 132–144.
Cutler D. F. (1979) Leaf surface studies in Aloe andHaworthia species (Liliaceae), taxonomicimplications. Trop. Subtrop. Pflanzenwelt(Akad.Wiss. Lit. Mainz) 28: 8–29.
Donoghue M. J., Olmstead R. G., Smith J. F.,Palmer J. D. (1992) Phylogenetic relationships ofDipsacales based on rbcL sequences. Ann. Mis-souri Bot. Gard. 79:333–345.
Downie S. R., Katz-Downie D. S. (1996) Amolecular phylogeny of Apiaceae subfamilyApioideae: evidence from nuclear ribosomalDNA internal transcribed spacer sequences.Amer. J. Bot. 83: 234–251.
Downie S. R., Palmer J. D. (1994) A chloroplastDNA phylogeny of the Caryophyllales based onstructural and inverted repeat restriction sitevariation. Syst. Bot. 19: 236–252.
Downie S. R., Olmstead R. G., Zurawski G., SoltisD. E., Soltis P. S., Watson J., Palmer J. D. (1991)Six independent losses of the chloroplast DNArpL2 intron in dicotyledons: molecular andphylogenetic implications. Evolution 45: 1245–1259.
Eckardt T. (1964) Reihe Centrospermae. In: EnglerA. (ed.) Syllabus der Pflanzenfamilien, ed. 12,2.Borntraeger, Berlin, pp. 79–102.
Ehrendorfer F. (1976) Closing remark: systematicsand evolution of Centrospermae families. Pl.Syst. Evol. 126: 99–105.
Endress M. E., Bittrich V. (1993) Molluginaceae.In: Kubitzki K., Rohwer J. G., Bittrich V. (eds.)Volume II. Flowering plants. Dicotyledons.Magnoliid, Hamamelid and Caryophyllid fami-lies. Springer, Berlin, pp. 419–425.
Eriksson T. (1998) AutoDecay, ver. 4.0 ed. Dis-tributed by the authors, Bergius Foundation,Royal Swedish Acadamy of Sciences. Stock-holm, Sweden.
Felsenstein J. (1985) Confidence limits on phylog-enies: an approach using bootstrap. Evolution39: 783–791.
Friedrich H.-Ch. (1970) 27. Aizoaceae. In: Merx-muller H. (ed.) Prodromus einer Flora vonSudwestafrika. Cramer, Lehre, pp. 1–135.
N. S. Hassan et al.: Sesuvioideae 141

GilbertM.G.,HartmannH.E.K., EdwardsS. (2000)Aizoaceae. In: Edwards S., TadesseM., DemissowS., Hedberg I. (eds.) Flora of Ethiopia and Eritrea,vol. 2. Addis Ababa, Uppsala, pp. 240–248.
Goncalves M. L. (1965) Sobsidios para o conheci-mento da flora de Angola – I. Garcia de Orta 13:381–382
Hartmann H. E. K. (1993) Aizoaceae. In: KubitzkiK., Rohwer J.G., Bittrich V. (eds.) The familiesand genera of vascular plants ed.2. Springer,Berlin, pp. 37–69.
Hartmann H. E. K. (2001a) Aizoaceae A-E. In:Illustrated handbook of succulent plants.Springer, Berlin.
Hartmann H. E. K. (2001b) Aizoaceae F-Z. In:Illustrated Handbook of Succulent Plants.Springer, Berlin.
Hassan N. M. S., Meve U., Liede-Schumann S.(2005) Seed coat morphology of Aizoaceae-Sesuvioideae, Gisekiaceae and Molluginaceae,and its systematic significance. Bot. J. Linn. Soc.148:189–206.
Hershkovitz M. A., Zimmer E. A. (1997) On theevolutionary origins of the cacti. Taxon 46: 217–232.
Hofmann U. (1973) Morphologische Untersuch-ungen zur Umgrenzung und Gliederung derAizoaceen. Bot. Jahrb. Syst. 93: 247–324.
Hofmann U. (1977) Die Stellung von Stegnospermainnerhalb der Centrospermen. Centrospermen-studien 9. Ber. Deutsch. Bot. Ges. 90: 39–52.
Horaninow P. F. (1834) Prima lineae SystemisNature. St. Petersburg.
Hutchinson J., Dalziel J. M. (1954) Flora of WestTropical Africa, vol. 1. Milbank, London, pp.135–136.
Jeffrey C. (1960) Notes on tropical African Aizo-aceae. Kew Bull. 14: 235–238.
Jeffrey C. (1961) Aizoaceae. In: Hubbard C.,Milne-Redhead E. (eds.) Flora of tropical EastAfrica. Crown Agents for Oversea Govern-ments, London.
Klak C., Khunou A., Reeves A., Hedderson T. A.(2003) A phylogenetic hypothesis for the Aizo-aceae (Caryophyllales) based on four plastidDNA regions. Amer. J. Bot. 90: 1433–1445.
Landrum J. V. (2001) Wide-band tracheides inleaves of genera in Aizoaceae: the systematicoccurrence of a novel cell type and its implica-tions for the monophyly of the subfamily Ru-schioideae. Pl. Syst. Evol. 227: 49–61.
Lindley J. (1846) Vegetable Kingdom, ed. 1.Longmans, London.
Maddison W. P., Maddison D. R. (2000) Mac-Clade: analysis of phylogeny and characterevolution. Version 4. Sinauer Asscociates, Sun-derland, Massachusetts.
Manhart J. R., Rettig J. H. (1994) Gene sequencedata. In: Behnke H.-D., Mabry T. J. (eds.)Caryophyllales: evolution and systematics.Springer, Berlin, pp. 235–246.
Melville R. (1952) Trianthema pentandra L. andsome related species. Kew Bull. 7: 266–278.
Muller K. (1909) Beitrage zur Systematik derAizoaceen. Bot. Jahrb. Syst. 42. Beibl. 97: 54–94.
Muschler R. (1912) A manual Flora of Egypt 1.Friedlander, Berlin, pp. 319–326.
Oliver D. (1871) Flora of Tropical Africa, vol. 2.Reeve, Kent, pp. 581–596.
Phillips D. C. (1988) Wild flowers of Bahrain. D. C.Phillips, Manama (Bahrain), pp. 59–63.
Pop M., Oxelman B. (2001) Inferring the history ofthe polyploid Silene aegaea (Caryophyllaceae)using plastid and homoeologous nuclear DNAsequences. Molec. Phylogenet. Evol. 20: 474–481.
Prescott A., Venning J. (1984) Aizoaceae. In: Alex-ander S. (ed.) Flora of Australia, vol. 4. AustralianGovernment Publ. Service, Canberra, pp. 19–62.
Raghavan T. S., Srinivasan V. K. (1940) Studies inIndian Aizoaceae. Ann. Bot. (new series) 4: 651–661.
Sage R. F. (2001) Environmental and evolutionarypreconditions for the origin and diversificationof the C4 photosynthetic syndrome. Plant Biol.3: 202–213.
Sanderson M. J., Donoghue M. J., Piel W.,Eriksson T. (1994) TreeBASE: a prototypedatabase of phylogenetic analyses and an inter-active tool for browsing the phylogeny of life.Amer. J. Bot. 81: 183.
Schmid W. (1925) Morphologische, anatomischeund entwicklungsgeschichtliche Untersuchungenan Mesembryanthemum pseudotruncatellum Ber-ger. Beibl. Vierteljahrsschr. Naturforsch. Ges.Zur. 8: 1–80.
Singh B. P. (1992) Chemotaxonomy of Aizoaceaeand Molluginaceae — a study of free aminoand organic acids. J. Econ. Tax. Bot. 16(1):103–106.
Sonder O. W. (1862) Trianthema. In: Harvey W.H., Sonder O. W. (eds.) Flora Capensis, vol. 2.Hodges, Smith & Co., Dublin.
142 N. S. Hassan et al.: Sesuvioideae

Swofford D. L. (1998) PAUP*. Phylogenetic anal-ysis using parsimony (*and other methods).Version 4. Sinauer Associates, Sunderland, Mas-sachusetts.
Tackholm V. (1956) Student’s Flora of Egypt, ed.1. Univ. Press, Cairo, pp. 406–410.
Thiede J. (2004) Phylogeny, systematics and clas-sification of the Aizoaceae: a reconsiderationbased on molecular data. In: Thiede J., MetzingD., Veste M., Jurgens N. (eds.) Succulent plantresearch in Africa. Festschrift Hans-Dieter Ih-lenfeldt. Schumannia 4.
Wallace R. S. (1998) Systematic implications ofevolutionary relationships within the Aizoaceaes.l. IOS Bull. 6: 25.
Willdenow C. L. in Rottler, J. B. (1803) BotanischeBemerkungen auf der Hin- und Ruckreise vonTrankenbar nach Madras von Herrn Missionar
Rottler mit Anmerkungen von Herrn Prof. D. L.Willdenow. Ges. Naturf. Freunde Berlin NeueSchriften 4: 181.
Zohary M. (1966) Flora Palaestina 1. Israel Acad-emy of Sciences and Humanities, Jerusalem, pp.74–77.
Addresses of the authors: N. S. Hassan, BotanyDepartment, Faculty of Science, Assiut University,Assiut 71516, Egypt; J. Thiede, Biozentrum KleinFlottbek, Systematik der Pflanzen, Hamburg Uni-versity, Ohnhorststrasse 18, 22609 Hamburg, Ger-many; Sigrid Liede-Schumann (e-mail:[email protected]), Department ofplant Systematics, University of Bayreuth, 95440Bayreuth, Germany.
N. S. Hassan et al.: Sesuvioideae 143
Related Documents











