Vol. 138, No. 3, 1986 August 14, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Pages 1395-1404 PHOSPIIATIDYLINOSITOL 4 ,5-BISPHOSPHATE FORi IATION RABBIT SI',ELETAL AND HEART MUSCLE ~EbiBRANES IN biagdolna Varsanyi, blichael Messer, Neil R. Brandt, Ludwig M.G. Heilmeyer, Jr. Institut f~r Physiologische Chemie, Abteilung f~r Biochemie Supralnolekularer Systeme, Ruhr-Universitgt Bochum, Universit~tsstraBe 150, 4630 Bochum I, West-Germany Received June 25, 1986 Incubation of rabbit skeletal muscle microsomes or isolated triads with 7 32P-ATP/Mg2+ in the absence and in the presence of added phosphatidylinositol resulted in the formation of phosphatidylinositol 4-phosphate catalyzed by phosphatidylinosi- tol kinase. When phosphatidylinositol 4-phosphate was added as exogenous substrate, phosphatidylinositol 4,5-bisphosphate was also formed demonstrating the presence of a membrane bound phos' phatidylinositol 4-phosphate kinase. Triads were broken mechani- cally in a French press and separated on a continuous sucrose gradient. Incubation of these fractions with ~ 32P- ATP/t2g2+ resulted in a rapid labeling of phospholipid in a membrane fraction banding between transverse tubules and the terminal cysternae. Partial triad breakage and triad reformation experiments indicated that this phosphatidylinositol kinase was associated with T-tubules. When exogenous phosphatidylinositol 4- phosphate was employed as substrate phosphatidylinositol 4,5- bisphosphate and phosphatidic acid were formed, indicating the presence of all the enzymes of the polyphosphoinositide signaling system in this special membrane fraction. In contrast, heart muscle microsomes or plasma membranes can catalyze this reaction sequence from endogenous formed phosphatidylinositol 4- phosphate. © 1986~ademiePress. Ine. In fast skeletal muscle until now only the first step in the PI- phosphorylation sequence has been demonstrated (I); the consecutive phosphorylation of PIP to PIP 2 has not yet been observed. A membrane bound PI kinase catalyzes the PIP formation in SR isolated from rabbit skeletal muscle (I). The PIP formation rate and the steady state level is enhanced by phosphorylase ABBREVIATIONS: DG: diacylglycerol; PA: phosphatidic acid; PI: phosphatidylinositol; PIP: phosphatidylinositol 4-phosphate; PIP2: phosphatidylinositol 4,5-bisphosphate; PPI: polyphospho- inositides; IP3: inositol 1,4,5-trisphosphate; SR: sarcoplasmic reticulum; QnB: quinuclidinyl-benzilate; DHP: dihydropyridine. 1395 0006-291X/86 $1.50 Copyright © 1986 by Academic Press, Inc. All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vol. 138, No. 3, 1986
August 14, 1986
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Pages 1395-1404
PHOSPIIATIDYLINOSITOL 4 , 5-BISPHOSPHATE FORi IATION RABBIT SI',ELETAL AND HEART MUSCLE ~EbiBRANES
IN
biagdolna Varsanyi, blichael Messer, Neil R. Brandt, Ludwig M.G. Heilmeyer, Jr.
Institut f~r Physiologische Chemie, Abteilung f~r Biochemie Supralnolekularer Systeme, Ruhr-Universitgt Bochum,
Universit~tsstraBe 150, 4630 Bochum I, West-Germany
Received June 25, 1986
Incubation of rabbit skeletal muscle microsomes or isolated triads with 7 32P-ATP/Mg2+ in the absence and in the presence of added phosphatidylinositol resulted in the formation of phosphatidylinositol 4-phosphate catalyzed by phosphatidylinosi- tol kinase. When phosphatidylinositol 4-phosphate was added as exogenous substrate, phosphatidylinositol 4,5-bisphosphate was also formed demonstrating the presence of a membrane bound phos' phatidylinositol 4-phosphate kinase. Triads were broken mechani- cally in a French press and separated on a continuous sucrose gradient. Incubation of these fractions with ~ 32P- ATP/t2g2+ resulted in a rapid labeling of phospholipid in a membrane fraction banding between transverse tubules and the terminal cysternae. Partial triad breakage and triad reformation experiments indicated that this phosphatidylinositol kinase was associated with T-tubules. When exogenous phosphatidylinositol 4- phosphate was employed as substrate phosphatidylinositol 4,5- bisphosphate and phosphatidic acid were formed, indicating the presence of all the enzymes of the polyphosphoinositide signaling system in this special membrane fraction. In contrast, heart muscle microsomes or plasma membranes can catalyze this reaction sequence from endogenous formed phosphatidylinositol 4- phosphate. © 1986 ~ademiePress. Ine.
In fast skeletal muscle until now only the first step in the PI-
phosphorylation sequence has been demonstrated (I); the
consecutive phosphorylation of PIP to PIP 2 has not yet been
observed. A membrane bound PI kinase catalyzes the PIP formation
in SR isolated from rabbit skeletal muscle (I). The PIP formation
rate and the steady state level is enhanced by phosphorylase
ABBREVIATIONS: DG: diacylglycerol; PA: phosphatidic acid; PI: phosphatidylinositol; PIP: phosphatidylinositol 4-phosphate; PIP2: phosphatidylinositol 4,5-bisphosphate; PPI: polyphospho- inositides; IP3: inositol 1,4,5-trisphosphate; SR: sarcoplasmic reticulum; QnB: quinuclidinyl-benzilate; DHP: dihydropyridine.
1395
0006-291X/86 $1.50 Copyright © 1986 by Academic Press, Inc.
All rights of reproduction in any form reserved.

Vol. 138, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
kinase showing that this protein kinase expresses, as a side
activity, also lipid kinase activity (2,3).
Recently, IP 3 has been shown to enhance reversibly the isometric
submaximum steady state force of skinned skeletal muscle fibres
(4). Additionally, IP 3 has been discussed Controversially as a
possible chemical link in excitation/contraction coupling
(5,6,7,8). Whatever the predominant role of IP 3 is in skeletal
muscle, if it has a physiological role, the whole cascade from PI
to IP 3 must take place in this tissue. Therefore, a search has
been undertaken for lipid kinase activities in membrane fractions
from rabbit skeletal muscle. In heart preliminary studies
indicated PPI formation in ventricular muscle (9).
Here we will show that T-tubules isolated from skeletal muscle
triads contain PI kinase, PIP kinase and DG kinase activities.
However, the formation of the intermediate product PIP 2 is
observed only when PIP is added as exogenous substrate. In
contrast, heart muscle plasma membranes catalyze the whole
reaction sequence from endogenous PI.
MATERIALS AND METHODS
Rabbit skeletal muscle microsomes, triads, T-tubules and SR subfractions were prepared from back muscle as described by Lau et al., (10). Rabbit heart muscle microsomes were isolated from ventricles according to (11) and further separated into dyads and plasma membranes by centrifugation on a linear continuous sucrose gradient from 15 to 45 %. Protein was determined by the methods of Lowry (12) or Bradford (13) using bovine serum albumin as standard. DHP and muscarine binding sites were assayed with (3H)PN 200/110 (0.5 nM for skeletal and 0.25 nM for cardiac muscle) and (3H) QnB, 0.5 nM, respectively (14). Preparation of lipid substrates and activity assay of the lipid kinases were carried out according to (3). Both lipid substrates, PI and PIP, were used at final concentrations of ca. I mM in the phosphorylation mixture. Assay of lipid phosphorylation and lipid identification: membrane fractions were incubated in a total volume of 150 ~i at pH 7.5 and 30 ° C containing 100 mM Hepes, 10 mM EDTA, I mM EGTA, 10 mM
7-32P-ATP, 25 mM Mg2+ in absence or presence of exogenously added phospholipid substrates. During the incubation aliquots of 30 ~i were removed , suspended in 300 ~i chloroform: methanol, I : 2 (v/v) and stored on ice. Determination of radioactivity incorporated into lipids was carried out as described by Schacht (15). For identification of the labeled lipids duplicate aliquots
1396

V o l . 1 3 8 , N o . 3 , 1 9 8 6 B I O C H E M I C A L A N D B I O P H Y S I C A L R E S E A R C H C O M M U N I C A T I O N S
of the chloroform phase were evaporated to dryness under a stream of nitrogen. The dried samples were taken up in a small volume of chloroform and were spotted on silica gel plates (Merck Darmstadt, Art.-No. 5715). The mobil phase was chloroform/ aceton/methanol/acetic acid/water, 40:15:13:12:8, v/v) as described in (16). Comigration of labeled products with authentic PIP and PIP~ (Sigma) were visualized following autoradiography at -70 ° C withZKODAK X Omat film.
RESULTS
i:~abbit skeletal muscle microsomes can be separated by isopycnic
centrifugation on a linear sucrose gradient into a heavy fraction
banding at ca. 40 % sucrose containing triads and a light
fraction banding at ca. 26 % sucrose. The heavy band contains
intact triads as indicated by the DHP marker for T-tubules (Fig.
I). The light band has been identified as derived mainly from the
longitudinal reticulum of the SR (I0). A low DHP binding
capacity is observed in this light fraction too, which could
indicate an approximately 10 % contamination by free tubules or
non-tubular plasma membranes. The microsomes and both fractions,
triads and the longitudinal SR, exhibited PI kinase activity.
20 40 60
froction number
.3oL x
20"~=
- E o"
o o 10- z 0_
sucrose(%w/w}
44.5 40 34 32.5 30.5 27 22 14 I I I I I I I
Figure
7 c_
E x 7
250 x o
200 o
15o -~
I00 o
o 50 k2
I." Separation of rabbit skeletal muscle microsomes into triads and longitudinal sarcoplasmic reticulum Microsomes from rabbit skeletal muscle were prepared as described in Methods. 3 ml (ca. 30 mg protein) were layered on top of 15 ml sucrose gradient from 10 - 45 % (w/w~. Centrifugation was carried out at 100,000 xg for 12 hours at 4 C. Protein (O) and ligand binding (O) were determined as described in Methods. PI-kinase activity (A) was tested as described in (3) with the modification that PI associated with a heat denatured Ca2+ transport ATPase was used as substrate.
1397

Vol. 138, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Table 1
PI and PIP kinase activities in rabbit skeletal muscle membrane fractions
fraction kinase
microsomes triads longitudinal SR
-I -I pmole x min x mg
PI kinase
endogenous substrate 173
exogenous substrate 154
PIP kinase
exogenous substrate 11
149 55
159 40
32 3
Lipid kinase activities were determined as described in Methods. The PIP kinase activities were calculated from the initial velocity of lipid phosphorylation and the ratio of PIP 2 formation to the total amount of phosphorylated lipids unoer assay condition.
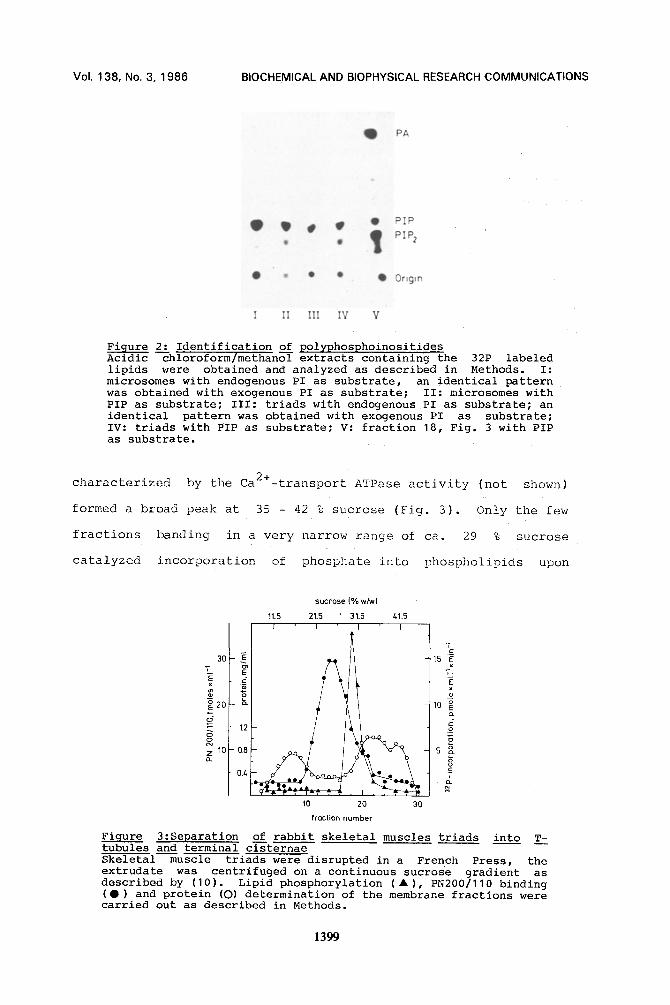
Table 1 summarizes the 32~ incorporation rates into the PI
either associated endogenously with tile membranes or into the PI
added exogenously as an ultrasonicated Triton mixture. Product
identification showed that only PIP but no£ PIP 2 was formed under
these conditions (Fig. 2, lane I and III). An identical result
was obtained from the longitudinal Si< fraction (not shown).
IIowever, when PIP was added as exogenous substrate a new
product, PIP2, appeared (Fig. 2, lane II and IV). The isolated
triad fraction accumulates PIP 2 to a higher percentage in
relation to the total amount of PPI than the microsomal fraction
(Fig. 2; compare lane II and IV). Consistent with this
observation, laost of the PI kinase and PIP kinase activities
present in the microsomes are found in the triad fraction (Tablel).
Triads were mechanically disrupted and centrifuged on isopycnic
gradients. The peak of the DHP binding sites which are present in
T-tubules appeared at 24 % sucrose while the terminal cysternae
1398
Vol. 138, No. 3, 1 9 8 6 B IOCHEMICAL A N D BIOPHYSICAL RESEARCH C O M M U N I C A T I O N S
PA
i PIP iI~ ~ lip f PiP2 u
• • I Q Origin
I I] III IV V
Figure 2: Identification of polyphosphoinositides Acidic --chloroform/methan~ extracts containing the 32P labeled lipids were obtained and analyzed as described in Methods. I: microsomes with endogenous PI as substrate, an identical pattern was obtained with exogenous PI as substrate; II: microsomes with PIP as substrate; III: triads with endogenous PI as substrate; an identical pattern was obtained with exogenous PI as substrate; IV: triads with PIP as substrate; V: fraction 18, Fig. 3 with PIP as substrate.
characterized by the Ca2+-transport ATPase activity (not shown)
formed a broad peak at 35 - 42 % sucrose (Fig. 3). Only the few
fractions banding in a very narrow range of ca. 29 % sucrose
catalyzed incorporation of phosphate into phospholiPids upon
sucrose (% w/w)
11,5 21,5 ' 31.5 41.5 [ I ' I I ' ]
'E "l 1o
d o~ 12
~ l o - o 8 5
10 20 30 frQction number
Figure 3:Separation of rabbit skeletal muscles triads into T- tubules and terminal cisternae Skeletal muscle triads were disrupted in a French Press, the extrudate was centrifuged on a continuous sucrose gradient as described by (10). Lipid phosphorylation ( • ), PN200/110 binding (•) and protein (O) determination of the membrane fractions were carried out as described in Methods.
1399

Voi. 138, No. 3 ,1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
incubation of each fraction with ~2p-ATP/Mg2+ This band is
completely contained within a shoulder of the DHP binding site
distribution and does not appear to be with the terminal
cysternae. PI kinase activity was not found at 29 % sucrose when
the components of disrupted triads were rejoined in presence of
potassium-cacodylate following a second centrifugation in an
identical isopycnic gradient (not shown). In contrast, when
partially disrupted triads were centrifuged, the content of PI
kinase in the 29 % sucrose band correlated with the amount of
free T-tubules. Dombradl et al. (17) have employed similar
protocols to demonstrate the association of phosphorylase kinase
with the T-tubule membranes.
The initial phosphate incorporation rate of the 29 % band was
enhanced nearly 2-fold by addition of exogenous PI and a further
2-fold by addition of exogenous PIP as substrates. Product
identification for the PIP reaction showed that a high amount of
PIP 2 and a considerably lower amount of PIP were formed (Fig. 2,
lane V). Most surprisingly, PA was also produced as a further
major product. The data suggest that part of the formed PIP 2 has
already been split into DG and IP 3 . DG was apparently
phosphorylated to PA, implying the presence of phospholipase C
and DG kinase activities in the 29 % sucrose fraction, blinor
amounts of different lysoforms can also be observed which
probably were formed by degradation during phospho!ipid
isolation.
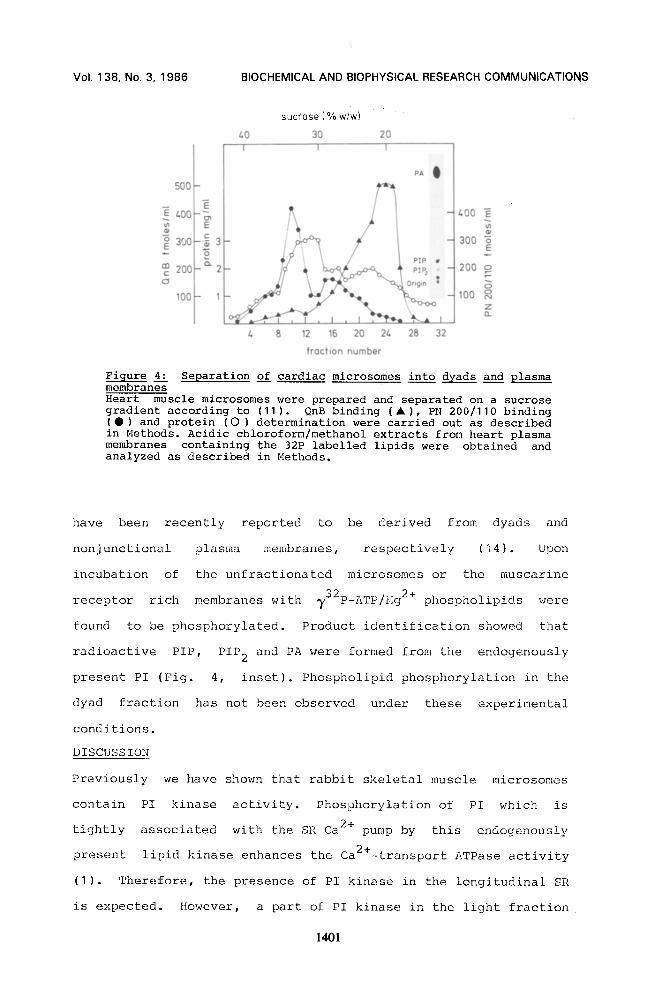
Heart muscle microsomes can be separated on a continuous sucrose
gradient into several fractions (Fig. 4). One band at 33 %
sucrose which is more dense than the bulk of the SR is
characterized by a high DHP binding activity - relative to
muscarine receptors - while another band at ca. 20 % sucrose
has a low ratio of DHP to muscarine sites. 'Phese microsomal bands
1400
VoI. 138 , No. 3, 1 9 8 6 BIOCHEMICAL A N D BIOPHYSICAL RESEARCH C O M M U N I C A T I O N S
500
400
300 E
m 200 c
o
I00
sucrose (% Wiw)
40 30 20
c~ 400 -~
c 3 300
o_ 2 200 o
S 1 100 o
Z 0_
8 12 16 20 24 28 32
f ract ion number
Figure 4." Separation of cardiac microsomes into dyads and plasma membranes Heart muscle microsomes were prepared and separated on a sucrose gradient according to (11). QnB binding (A), PN 200/110 binding (Q) and protein (O) determination were carried out as described in Methods. Acidic chloroform/methanol extracts from heart plasma membranes containing the 32P labelled lipids were obtained and analyzed as described in Methods.
have been recently reported to be derived from dyads and
nonjunctional plasma membranes, respectively (14). Upon
incubation of the unfractionated microsomes or the muscarine
receptor rich membranes with 732p-ATP/Hg 2+ phospholipids were
found to be phosphorylated. Product identification showed that
radioactive PIP, PIP 2 and PA were formed from the endogenously
present PI (Fig. 4, inset). Phospholipid phosphorylation in the
dyad fraction has not been observed under these experimental
conditions.
DISCUSSION
Previously we have shown that rabbit skeletal muscle microsomes
contain PI kinase activity. Phosphorylation of PI which is
tightly associated with the SR Ca 2+ pump by this endogenously
present lipid kinase enhances the Ca2+-transport ATPase activity
(I). Therefore, the presence of PI kinase in the longitudinal SR
is expected. However, a part of PI kinase in the light fraction
140]

Vol. 138, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
might originate from free tubules or non tubular plasma
membranes.
~ioreover, we have observed that lipid kinase activity is also
present in the triad fraction. Using either endogenous or
exogenous PI as substrate, only PIP has been detected as product;
neither PIP 2 nor PA has been found. Under the same conditions,
cardiac microsomes and cardiac plasma membranes rich in muscarine
receptor binding sites can form PIP 2 as a result of the
consecutive phosphorylation of the endogenously produced PIP
(Eig. 4, inset). Based on the values reported in Table I it can
be calculated that during an incubation of 40 min PIP accumulates
in the skeletal muscle microsomes or triads up to ca. 50 p H. At
this substrate concentration PIP kinase from other sources
(18,19) can reach 5 to 30 % of its maximal activity found at
saturating substrate concentrations. Therefore, a PIP kinase
could be expected to be active under these incubation conditions.
The lack of PIP 2 formation from the endogenous PI and PIP can
also not be due to an absence of PIP kinase. The skeletal muscle
microsomes and triads converted exogenous PIP to the
bisphosphorylated product. Thus, at the moment it is unclear why
in skeletal muscle PIP kinase cannot act on the endogenously
formed PIP whereas it is active in heart muscle plasma membrane
preparation.
It ai~pears that in the skeletal muscle triads both of these lipid
hinases are predominantly associated with a subfraction of the T-
tubules which appears as a shoulder in the distribution pattern
for DHP binding sites (Fig. 3). The densitiy of these membrane
fractions is, among other factorsja function of the amount of
associated proteins. Thus, the presence of lipid kinase might
enhance the density. Furthermore, other membrane proteins e.g.
spanning protein or ~Jfeet '~ mig]lt also be distributed unequally
1402

VoI. 138, No .3 ,1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
in these tubular fractions with no causal relationship to the
lipid kinase activity distribution. The determined activity is
also a function of the substrate concentration. Therefore, an
activity peak as measured by endogenous lipid phosphorylation may
be a result of an enhanced PI availability rather than an
increased amount of lipid kinase protein. Employing exogenous PI
as substrate the PI kinase activity yields a pea k which is super-
imposed on the lipid phosphorylation band shown in Fig. 3.
Additionally, low PI kinase activity is detectable in fractions
of terminal cisternae (not shown).
The highest activity of the two lipid kinases in cardiac
microsomes is associated with the plasma membranes enriched in
muscarine receptors. This subpopulation is derived from non
junctional domaines of the surface membrane (14). These
observations may indicate different locations of the PPI
signaling system in these two types of striated muscle.
Alternatively, the cardiac dyad fraction may contain more active
lipid phosphatases and phospholipases than the skeletal muscle
triads. The cardiac and skeletal muscle membrane also show DG
kinase activity which has been reported previously to exist in
soluble form (2). The formation of PA in our experiments can be
explained by the action of a phosphodiesterase which in part
splits the formed PIP 2 into IP 3 and DG; the latter is then
converted to PA.
The pattern of phosphorylated phospholipids observed in the
skeletal as well as the heart muscle membrane fractions indicates
that all of the enzymes of the PPI signaling system are located
in these membranes. Since IP 3 can modulate contraction at a
remarkable low concentration (4) this IP 3 probably originates
from T-tubules or plasma membranes and thus, in principle, can be
formed upon extracellular stimulation.
1403

Vol. 138, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ACKNOWLEDGMENTS
N. R. B. is supported by grants from NIH (HL28804), the American Heart Association (Florida Affiliate) and ~iiles Institute for Preclinical Studies which is greatfully acknowledged. This work was supported by the Deutsche Forschungsgemeinschaft, SFB 168, and Fonds der Chemie. We thank Mrs. M. Cochu, Mrs. M. PreiS and ~ir. U. Siemen for their excellent technical assistance.
I~EFERENCES
I ) Varsanyi, M., TSlle, H.G., Dawson, R.:I.C., Irvine, R.F. and Heilmeyer, L.~I.G., Jr. (1983), E~i[30-J. 2, 1543-1548.
2) Georgoussi, Z. and IIeilmeyer, L.~,I. G. , Jr. (1986), Biochemistry, in press.
3) Varslnyi, M., Behle, G., Sch[fer, M. (1986), Z. Naturforsch. 41 c, 310-314.
4) Thieleczek, R. and Heilmeyer, L.M.G. Jr. (1986), Biochem. Biophys. Res. Commun. 135, 662-669.
5) Volpe, P. Salviati, G., Di Virgilio, F. and Pozzan, T. (1985), Nature 316, 347-349.
6) Vergara, J., Tsien, R.Y. and Delay, M. (1985), Proc. Nat. Acad. Sci. USA 82, 6352-6356.
(7) Scherer, N.I<I. and Ferguson, J.E. (1985), Biochem. Biophys. Res. Commun. 128, 1064-1070.
( 8) Ashley, C.C., Griffiths, P.J., Lea, T.J. and Tregear, R.T. (1985), J. Physiol. 369, 185.
~P (9) Enyedi, A., Farag6, A., Sarkadi, 13. and Gardos, G. (1984),
FEiBS-Lett. 176, 235-238. (10) Lau, Y.H., Caswell, A.H. and i~runschwig, J.P. (1977), J.
Biol. Chem. 252, 5565-5574. (11) Brandt, N. (1985), Arch. Biochem. Biophys. 242, 306-319. (12) Lowry, O.I{., P, osebrough, N.J., Farr, A.L. and Randall, R.J.
(1951), J. Biol. Chem. 193, 265-275. (13) Bradford, M.M. (1976), Anal. Biochem. 72, 248-254. (14) Brandt, N.R. and Susseff, A.L. (1986), Arch. ~iochem.
Biophys. 244, 872-875. (15) Schacht, J. (1981), ~eth. Snzymol. 72, 626-631. (16) Shaigk, N.A. and Palmer, F.B. St.C. (1977), J. Neurochem.
28, 395-402. (17) Dombr~di, V.K., Silbermann, S.R., Lee, E.Y.C., Caswell,
A.H. and Brandt, N.R. (1984), Arch. Diochem. Biophys. 230, 615-630.
(18) Cooper , P.Ii. and Iiawthorne, J.}]. (1976), Liochem. J. 160, 97-105.
(19) Kai, i.]., Salway, J.G. and Hawthorne, J.N. (1968), Biochem. J. 106, 791 -801.
]404
Related Documents











