REGULAR PAPER Phosphatidylglycerol depletion affects photosystem II activity in Synechococcus sp. PCC 7942 cells Bala ´zs Bogos Bettina Ughy Ildiko ´ Domonkos Hajnalka Laczko ´-Dobos Josef Komenda Leyla Abasova Krisztia ´n Cser Imre Vass Anna Sallai Hajime Wada Zolta ´n Gombos Received: 19 March 2009 / Accepted: 3 September 2009 / Published online: 18 September 2009 Ó Springer Science+Business Media B.V. 2009 Abstract The role of phosphatidylglycerol (PG) in pho- tosynthetic membranes of cyanobacteria was analyzed in a Synechococcus sp. PCC 7942 mutant produced by inacti- vating its cdsA gene presumably encoding cytidine 5 0 -diphosphate-diacylglycerol synthase, a key enzyme in PG synthesis. In a medium supplemented with PG the Synechococcus sp. PCC 7942/DcdsA cells grew photoau- totrophically. Depletion of PG in the medium resulted (a) in an arrest of cell growth and division, (b) in a suppression of O 2 evolving activity, and (c) in a modification of Chl fluorescence induction curves. Two-dimensional PAGE showed that in the absence of PG (a) the amount of the PSI monomers increased at the expense of the PSI trimers and (b) PSII dimers were decomposed into monomers. [ 35 S]methionine labeling confirmed that PG depletion did not block the de novo synthesis of PSII proteins but slowed down the assembly of the newly synthesized D1 protein into PSII core complexes. Retailoring of PG was observed during PG depletion: the exogenously added artificial dioleoyl PG was transformed into photosynthetically more essential PG derivatives. Concomitantly with a decrease in PG content, SQDG content increased, but it could not restore photosynthetic activity. Keywords Oxygen-evolving activity Phosphatidylglycerol PS II acceptor side Synechococcus Abbreviations 2D-BN Two-dimensional blue native gel electrophoresis Chl Chlorophyll CP43 43 kDa chlorophyll-binding protein DCMU 3-(3, 4-dichlorophenyl)-1, 1 0 -dimethylurea DGDG Digalactosyldiacylglycerol OD Optical density pBQ 1, 4-p-benzoquinone PG Phosphatidylglycerol PQ Plastoquinone Q A Primary quinone electron acceptor of PS II Q B Secondary quinone electron acceptor of PS II RC Reaction center SQDG Sulfoquinovosyldiacylglycerol WT Wild type Introduction The lipid composition of photosynthetic membranes is highly conserved among cyanobacterial strains and higher plant chloroplasts (Wada and Murata 1998). One of the lipid classes is phosphatidylglycerol (PG), which is an indis- pensable component of photosynthetic membranes. In cyanobacterial cells PG is the only representative of the B. Bogos B. Ughy I. Domonkos H. Laczko ´-Dobos L. Abasova K. Cser I. Vass A. Sallai Z. Gombos (&) Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, P. O. Box 521, 6701 Szeged, Hungary e-mail: [email protected] J. Komenda Institute of Microbiology, Academy of Sciences, Opatovicky ´ mly ´n, 37981 Tr ˇebon ˇ, Czech Republic J. Komenda Institute of Physical Biology, University of South Bohemia, Za ´mek 136, 37333 Nove ´ Hrady, Czech Republic H. Wada Department of Life Sciences, Graduate School of Arts and Sciences, University of Tokyo, Komaba, Tokyo 153-8902, Japan 123 Photosynth Res (2010) 103:19–30 DOI 10.1007/s11120-009-9497-0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REGULAR PAPER
Phosphatidylglycerol depletion affects photosystem II activityin Synechococcus sp. PCC 7942 cells
Balazs Bogos Æ Bettina Ughy Æ Ildiko Domonkos Æ Hajnalka Laczko-Dobos ÆJosef Komenda Æ Leyla Abasova Æ Krisztian Cser Æ Imre Vass ÆAnna Sallai Æ Hajime Wada Æ Zoltan Gombos
Received: 19 March 2009 / Accepted: 3 September 2009 / Published online: 18 September 2009
� Springer Science+Business Media B.V. 2009
Abstract The role of phosphatidylglycerol (PG) in pho-
tosynthetic membranes of cyanobacteria was analyzed in a
Synechococcus sp. PCC 7942 mutant produced by inacti-
vating its cdsA gene presumably encoding cytidine
50-diphosphate-diacylglycerol synthase, a key enzyme in
PG synthesis. In a medium supplemented with PG the
Synechococcus sp. PCC 7942/DcdsA cells grew photoau-
totrophically. Depletion of PG in the medium resulted (a)
in an arrest of cell growth and division, (b) in a suppression
of O2 evolving activity, and (c) in a modification of Chl
fluorescence induction curves. Two-dimensional PAGE
showed that in the absence of PG (a) the amount of the PSI
monomers increased at the expense of the PSI trimers and
(b) PSII dimers were decomposed into monomers.
[35S]methionine labeling confirmed that PG depletion did
not block the de novo synthesis of PSII proteins but slowed
down the assembly of the newly synthesized D1 protein
into PSII core complexes. Retailoring of PG was observed
during PG depletion: the exogenously added artificial
dioleoyl PG was transformed into photosynthetically more
essential PG derivatives. Concomitantly with a decrease in
PG content, SQDG content increased, but it could not
restore photosynthetic activity.
Keywords Oxygen-evolving activity �Phosphatidylglycerol � PS II acceptor side � Synechococcus
Abbreviations
2D-BN Two-dimensional blue native gel electrophoresis
Chl Chlorophyll
CP43 43 kDa chlorophyll-binding protein
DCMU 3-(3, 4-dichlorophenyl)-1, 10-dimethylurea
DGDG Digalactosyldiacylglycerol
OD Optical density
pBQ 1, 4-p-benzoquinone
PG Phosphatidylglycerol
PQ Plastoquinone
QA Primary quinone electron acceptor of PS II
QB Secondary quinone electron acceptor of PS II
RC Reaction center
SQDG Sulfoquinovosyldiacylglycerol
WT Wild type
Introduction
The lipid composition of photosynthetic membranes is
highly conserved among cyanobacterial strains and higher
plant chloroplasts (Wada and Murata 1998). One of the lipid
classes is phosphatidylglycerol (PG), which is an indis-
pensable component of photosynthetic membranes. In
cyanobacterial cells PG is the only representative of the
B. Bogos � B. Ughy � I. Domonkos � H. Laczko-Dobos �L. Abasova � K. Cser � I. Vass � A. Sallai � Z. Gombos (&)
Institute of Plant Biology, Biological Research Center,
Hungarian Academy of Sciences, P. O. Box 521, 6701 Szeged,
Hungary
e-mail: [email protected]
J. Komenda
Institute of Microbiology, Academy of Sciences, Opatovicky
mlyn, 37981 Trebon, Czech Republic
J. Komenda
Institute of Physical Biology, University of South Bohemia,
Zamek 136, 37333 Nove Hrady, Czech Republic
H. Wada
Department of Life Sciences, Graduate School of Arts
and Sciences, University of Tokyo, Komaba, Tokyo 153-8902,
Japan
123
Photosynth Res (2010) 103:19–30
DOI 10.1007/s11120-009-9497-0

glycerophospholipids (Wada and Murata 1998). PG is an
anionic lipid with a negatively charged phosphoglyceryl
headgroup attached to the diacyl-glycerol backbone. This
negative charge plays an important role in establishing pro-
tein–lipid interactions in photosynthetic membranes,
resulting in the formation of protein oligomers (Domonkos
et al. 2008). Although PG is a minor component of photo-
synthetic membranes, it plays important roles, in addition to
the formation of the photosynthetic apparatus, in a number of
various photosynthetic functions as presented below.
The importance of PG was studied in chloroplasts of
eukaryotic photosynthetic organisms, such as Chlamydo-
monas reinhardtii (Sato et al. 1995), and in higher plants
(Ariizumi et al. 2002; Hagio et al. 2002; Xu et al. 2002). It
has been demonstrated by using isolated PSII particles
treated with phospholipase A2 that PG molecules partici-
pate in PSII dimer formation (Kruse et al. 2000). The
availability of the complete genomic sequence of Syn-
echocystis sp. PCC 6803 (Kaneko et al. 1996) and the
unpublished, draft genomic sequence of Synechococcus sp.
PCC 7942 opened the way for using molecular genetic
approaches to study the structural and functional roles of
PG in vivo. Studies on Synechocystis sp. PCC 6803 iden-
tified PG-phosphate synthase (PgsA) as a key enzyme
involved in PG biosynthesis (Hagio et al. 2000). Experi-
ments with the Synechocystis sp. PCC 6803/DpgsA mutant
revealed the important role of PG in the structural orga-
nization and functioning of the photosynthetic apparatus.
Maintenance of this mutant, defective in PG synthesis
required exogenously added PG to the medium. When
cultured in the absence of PG the PG content of the isolated
thylakoid membranes decreased gradually leading to lethal
impairments in a number of photosynthetic processes.
The effect of PG on individual steps of the electron
transport chain was also studied in this mutant, and a
perturbation in the vicinity of the secondary quinone
acceptor (QB) was observed (Gombos et al. 2002). We
found that depletion of PG resulted in decreased photo-
synthetic oxygen-evolving activity. Similar results were
obtained with a cytidine 50-diphosphate (CDP)-diacyl-
glycerol synthase (cdsA) mutant. This mutant harbors an
upstream mutation in PG biosynthesis in the same strain as
described by Sato et al. (2000).
X-ray crystallographic data showed the presence of PG
molecules in PSI and its absence in PSII reaction centers
(RCs) (Jordan et al. 2001; Zouni et al. 2001). Even 3.8 A
resolution measurements did not allow the detection of lipid
molecules in PSII (Zouni et al. 2001). The structure of PSII
was published with better resolution, suggesting the presence
of a PG molecule in the vicinity of D1 and the 43 kDa
chlorophyll-binding protein (CP43) (Loll et al. 2005).
However, recent X-ray crystallographic data with 2.9 A
resolution identified the presence of two PG molecules per
monomer of PSII. The previously known (Loll et al. 2005)
and the newly identified PG molecule are located on the
cytoplasmic side, close to the QB site (Guskov et al. 2009).
Both of the PG molecules are situated on the cytoplasmic
face of a bilayer domain formed by 3 MGDG, 2 DGDG, an
SQDG and the mentioned two PG molecules. This bilayer
island is encircled by protein subunits D1, CP43, PsbE, PsbF,
PsbJ, and PsbK that form the PQ-PQH2 exchange cavity. In
the case of the PSI RCs X-ray crystallographic measure-
ments also indicated the presence of three PG molecules
(Jordan et al. 2001), but PSI activity was not significantly
affected by short-term PG depletion (Hagio et al. 2000).
Nevertheless, PG depletion affected PSII activity earlier than
it did PSI processes. This finding suggested that the strength
of PG binding by PSII might be much weaker than that by the
PSI RCs. Recent reports confirmed the deleterious effect of
PG deprivation not only on photosynthetic function but also
on the structure of photosynthetic RCs in cyanobacteria
(Domonkos et al. 2004; Sakurai et al. 2003). Since PG is
required for functional binding of extrinsic proteins to the
PSII core (Sakurai et al. 2007) we concluded that PG affects
the functionally important proteins involved in photosyn-
thetic electron transport.
More detailed investigation of the effects of PG deple-
tion revealed the importance of PG in the maintenance of
primary quinone acceptor (QA) to QB electron transfer
within the PSII RC (Gombos et al. 2002). The character-
ization of a phycobilisomeless mutant of Synechocystis sp.
PCC 6803, PAL/DcdsA, that was defective also in PG
synthesis has strengthened the view that active PSII RC
needs PG (Laczko-Dobos et al. 2008). The results
emphasized that PG is active on the acceptor side of PSII
rather than on its donor side.
In addition to Synechocystis sp. PCC 6803, Synecho-
coccus sp. PCC 7942 would be another experimental object
to study the effect of PG depletion on photosynthetic and
other cellular processes. The unicellular Synechococcus sp.
PCC 7942 is a non-diazotrophic, obligatory photoautotro-
phic cyanobacterial strain.
In Synechocystis sp. PCC 6803 both PG and sulfoquino-
vosyldiacylglycerol (SQDG) are essential lipids (Aoki et al.
2004; Hagio et al. 2000) for the organism to maintain growth
and photosynthetic activity. However, in Synechococcus sp.
PCC 7942 SQDG is not essential, as it has been demonstrated
by Guler and co-workers (1996). Loss of the sulfolipid in
SQDG-deficient cells was compensated for by an increase in
cellular content of PG. To investigate the role of PG
in photosynthetic processes of Synechococcus sp. PCC 7942,
we inactivated the cdsA gene of this organism.
In the present article we demonstrate, by using fluores-
cence decay kinetics and two-dimensional blue native
SDS-PAGE (2D-BN/SDS-PAGE), that in the mutant Syn-
echococcus sp. PCC 7942/DcdsA PG depletion suppresses
20 Photosynth Res (2010) 103:19–30
123

electron transport around QB and affects the oligomeric
state of PSI and PSII, respectively. These results demon-
strate that PG is an indispensable lipid in both Synecho-
cystis sp. PCC 6803 (Hagio et al. 2000) and Synechococcus
sp. PCC 7942 cells unlike SQDG which is dispensable for
the maintenance of cellular functions in Synechococcus sp.
PCC 7942 (Sato 2004).
Materials and methods
Organisms and growth conditions
Synechococcus sp. PCC 7942/DcdsA cells were selected
and grown on BG11 agar plates supplemented with
500 lM sodium acetate. Synechococcus sp. PCC 7942 WT
and mutant cells were cultured in unbuffered BG11 liquid
medium supplemented with 500 lM sodium acetate. The
cultures were irradiated at 30�C with 30–35 lmol pho-
tons m-2 s-1 of continuous white light. Aeration was
performed without additional CO2, using a gyratory shaker
at 100 rpm. 50 ml liquid cultures were maintained in
150 ml Erlenmeyer flasks in the presence of 20 lM diol-
eoyl-PG (18:1/18:1 PG; P-9664; Sigma, St. Louis),
50 lg ml-1 kanamycin or 8 lg ml-1 chloramphenicol. PG
depletion was carried out by washing the cells twice with
PG-free medium, and culturing them thereafter in PG-free
medium.
Synechocystis sp. PCC 6803/DpgsA mutant cells (Hagio
et al. 2000) were grown photoautotrophically in BG11
medium supplemented with 5 mM HEPES–NaOH (pH
7.5), and 20 lg ml-1 kanamycin. Growth conditions, PG
supplementation and PG depletion were similar to those
used for Synechococcus sp. PCC 7942 cells.
Mutant generation, transformation of cells
Standard molecular biological methods were used during
the targeted insertional mutagenesis of Synechococcus sp.
PCC 7942. Escherichia coli DH5a and XL1-Blue cells
(grown in Luria broth at 37�C) were transformed using
standard methods (Sambrook et al. 1989). RCU1 50-CTC
GAGCAACGCTTGCTTAT-30 and RCD1: 50-AATTCGC
ATTGCCGCTGAGG-30 primer pair was used to amplify
the cdsA locus of Synechococcus sp. PCC 7942/DcdsA.
Primers were designed on the basis of the draft genome
sequence of Synechococcus sp. PCC 7942 available at
http://genome.ornl.gov/microbial/syn_PCC7942/. The cdsA
gene was identified by homology search using the published
sequence of the Synechocystis sp. PCC 6803 cdsA gene (http://
bacteria.kazusa.or.jp/cyanobase/). The amplified 1,596 bp
cdsA PCR fragments were cloned into pMPM-A2 recipient
vector (Mayer 1995) and verified by automatic sequencing.
The 209 bp MunI/HincII fragment of the cdsA gene was
replaced by a kanamycin resistance cassette or a chlor-
amphenicol resistance cassette from pZE21 and pZA3
vectors (Lutz and Bujard 1997), respectively. The resulting
plasmids carrying chloramphenicol resistance, pRC5C1,
and kanamycin resistance, pRC5K1 were used for targeted
insertional mutagenesis of the cdsA gene. Transformation
of WT Synechococcus sp. PCC 7942 cells was done
according to an optimized method (Golden and Sherman
1984). The segregation state of the transformants was
checked by PCR amplification of the cdsA locus from the
mutant chromosome.
Lipid analysis
Lipids were extracted from intact cells according to
standard lipid analytic methods (Bligh and Dyer 1959).
Lipid classes were separated by thin layer chromatogra-
phy, and fatty acids were analyzed on Supelco SP2330
capillary columns in a Hewlett Packard HP6890 gas
chromatography equipment as described earlier (Wada
and Murata 1989).
Spectroscopic measurements of cell density, proteins,
and pigments
Aliquots of cyanobacterial cells were collected and OD
was measured at 750 nm using a Shimadzu UV-3000
Spectrophotometer (Columbia, MD). Cells were pelleted
and extracted with acetone:methanol (7:2) (Ihlenfeldt and
Gibson 1975). The concentration of Chl was calculated
from the absorbance at 663 nm of the extracted samples
using an extinction coefficient of 82 mM-1 cm-1. The
protein content of extracted and pelleted cells was mea-
sured by the Lowry method (Yeang et al. 1998). For
measurements of protein concentration Bovine Serum
Albumin solution was used as a standard.
Absorption spectra were taken using protein normalized
cultures in 3 ml quartz cuvettes using a Shimadzu UV-
1601 spectrophotometer. The cultures were adjusted to
200 lg ml-1 protein content. The baseline was corrected
using two blank cuvettes filled with BG11 medium turned
with the opaque side toward the light path. The initial OD
at 750 nm was adjusted to zero and spectra were taken
using normal scanning speed against a blank sample. Each
of the spectra contains average data of three independent
PG-supplemented and PG-depleted samples in a given time
point. The resulting spectra were further normalized to
each other using the absorption maximum of the phyco-
biliproteins (625 nm) (Shibata 1958).
Photosynth Res (2010) 103:19–30 21
123

Measurements of photosynthetic oxygen-evolving
activity
Photosynthetic oxygen evolution in intact cells was mea-
sured with a Clark-type oxygen electrode (Hansatech
Instruments, Kings Lynn, UK) (Gombos et al. 1991).
Whole electron transport chain was measured from H2O to
CO2 and PSII oxygen-evolving activity from H2O to
exogenously added artificial quinone molecules (500 lM
pBQ). Light from an incandescent lamp at a saturating light
intensity of 500 lmol photons m-2 s-1, equipped with a
red optical filter was used in all oxygen evolution mea-
surements. The Chl concentration of the samples was
adjusted to 5 lg ml-1.
Flash-induced fluorescence relaxation kinetics
The kinetics of the decay of flash-induced variable Chl
fluorescence were measured in the 150 ls to 100 s time
range by a double modulation fluorometer (PS Instruments,
Brno) (Trtilek et al. 1997). Samples containing 200 lg
protein were used for the measurements. In order to show
the changes induced by PG depletion in the shape of the
flash fluorescence transients, the following normalization
was used: the basic level of fluorescence intensities (F0),
measured before the saturating flash, were shifted to the
same level, which was set as 0. Then the initial fluores-
cence amplitudes measured from the 0 level were nor-
malized to the same value, which was set to 1. After this
normalization the initial fluorescence rise in each samples
occurred from the 0 level to 1, and the shape of the decay
kinetics could be compared.
Protein analysis
Cells containing 75 lg of Chl were radioactively labeled at
29�C and at 60 lmol photons m-2 s-1 with [35S]Met (at
[1,000 Ci mmol-1, Isotope Institute Ltd, Hungary) for
20 min and then used for the isolation of membranes
(Komenda et al. 2004). Cyanobacterial membranes were
prepared by breaking the cells with glass beads (150–
200 lm diameter) at 4�C followed by differential centri-
fugation (Komenda and Barber 1995).
The isolated membranes were solubilized with dodecyl-
b-D-maltoside (dodecylmaltoside/Chl ratio of 40:1, w/w)
and analyzed by BN/SDS-PAGE at 4�C in a 5–14%
polyacrylamide gel (Schagger and von Jagow 1991). The
whole lane from the gel was excised, incubated for 30 min
in 25 mM Tris–HCl buffer (pH 7.5) containing 1% SDS
and then layered onto the top of a denaturing gel. The
protein composition of the complexes was then assayed by
the second dimension SDS-PAGE in a denaturing 12–20%
linear gradient polyacrylamide gel containing 7 M urea
(Komenda et al. 2004). The protein bands were stained
with Coomassie blue.
Results
Inactivation of the cdsA gene of Synechococcus sp.
PCC 7942
The cdsA gene encoding CDP-diacylglycerol synthase,
which is a crucial enzyme in PG synthesis, was identified in
the draft genome sequence of Synechococcus sp. PCC 7942
by homology search from the genomic data of Synecho-
cystis sp. PCC 6803. Following a targeted disruption of the
chromosomal cdsA gene by insertion of a chloramphenicol
(CmR) or a kanamycin (KmR) resistance cassette (Fig. 1a),
the transformants were segregated and verified by PCR
analysis (Fig. 1b). The kanamycin and chloramphenicol
resistance cassettes-containing Synechococcus sp. PCC
7942/DcdsA mutants did not show any phenotypical dif-
ferences compared to each other.
Effect of PG depletion on the growth and pigment
composition of Synechococcus sp. PCC 7942/DcdsA
cells
The growing of Synechococcus sp. PCC 7942/DcdsA
mutant cultures on solid or in liquid media requires exog-
enously added PG. The segregated transformants were able
to grow on PG-supplemented agar plates (Fig. 2a) and
were unable to grow on agar without added PG (Fig. 2b).
We also investigated the growth of Synechococcus sp.
PCC 7942/DcdsA in liquid medium under photoautotrophic
conditions in the presence and in the absence of PG. In the
medium containing PG the growth of Synechococcus sp.
PCC 7942/DcdsA cells was continuous even after 13 days
Fig. 1 Inactivation of chromosomal cdsA. a Physical map of the
Synechococcus sp. PCC 7942 genomic region containing the cdsAgene. In the inactivated strain the KmR or CmR gene cassette replaces
a 209 bp-long MunI/HincII gene fragment. Small black arrowsindicate positions of PCR primers used to amplify the region and to
verify the replacement. b PCR analysis of the WT (lane 1) and of
DcdsA kanamycin- (lane 2) and chloramphenicol- (lane 3) resistant
transformant genomic fragments
22 Photosynth Res (2010) 103:19–30
123
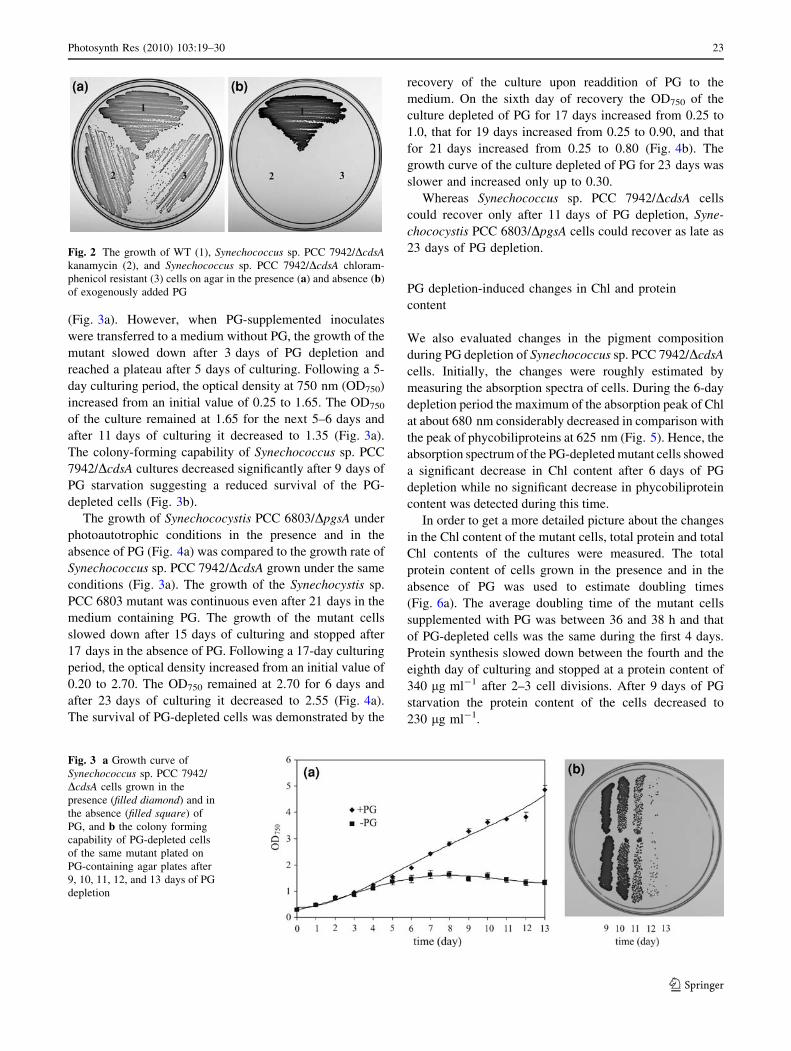
(Fig. 3a). However, when PG-supplemented inoculates
were transferred to a medium without PG, the growth of the
mutant slowed down after 3 days of PG depletion and
reached a plateau after 5 days of culturing. Following a 5-
day culturing period, the optical density at 750 nm (OD750)
increased from an initial value of 0.25 to 1.65. The OD750
of the culture remained at 1.65 for the next 5–6 days and
after 11 days of culturing it decreased to 1.35 (Fig. 3a).
The colony-forming capability of Synechococcus sp. PCC
7942/DcdsA cultures decreased significantly after 9 days of
PG starvation suggesting a reduced survival of the PG-
depleted cells (Fig. 3b).
The growth of Synechococystis PCC 6803/DpgsA under
photoautotrophic conditions in the presence and in the
absence of PG (Fig. 4a) was compared to the growth rate of
Synechococcus sp. PCC 7942/DcdsA grown under the same
conditions (Fig. 3a). The growth of the Synechocystis sp.
PCC 6803 mutant was continuous even after 21 days in the
medium containing PG. The growth of the mutant cells
slowed down after 15 days of culturing and stopped after
17 days in the absence of PG. Following a 17-day culturing
period, the optical density increased from an initial value of
0.20 to 2.70. The OD750 remained at 2.70 for 6 days and
after 23 days of culturing it decreased to 2.55 (Fig. 4a).
The survival of PG-depleted cells was demonstrated by the
recovery of the culture upon readdition of PG to the
medium. On the sixth day of recovery the OD750 of the
culture depleted of PG for 17 days increased from 0.25 to
1.0, that for 19 days increased from 0.25 to 0.90, and that
for 21 days increased from 0.25 to 0.80 (Fig. 4b). The
growth curve of the culture depleted of PG for 23 days was
slower and increased only up to 0.30.
Whereas Synechococcus sp. PCC 7942/DcdsA cells
could recover only after 11 days of PG depletion, Syne-
chococystis PCC 6803/DpgsA cells could recover as late as
23 days of PG depletion.
PG depletion-induced changes in Chl and protein
content
We also evaluated changes in the pigment composition
during PG depletion of Synechococcus sp. PCC 7942/DcdsA
cells. Initially, the changes were roughly estimated by
measuring the absorption spectra of cells. During the 6-day
depletion period the maximum of the absorption peak of Chl
at about 680 nm considerably decreased in comparison with
the peak of phycobiliproteins at 625 nm (Fig. 5). Hence, the
absorption spectrum of the PG-depleted mutant cells showed
a significant decrease in Chl content after 6 days of PG
depletion while no significant decrease in phycobiliprotein
content was detected during this time.
In order to get a more detailed picture about the changes
in the Chl content of the mutant cells, total protein and total
Chl contents of the cultures were measured. The total
protein content of cells grown in the presence and in the
absence of PG was used to estimate doubling times
(Fig. 6a). The average doubling time of the mutant cells
supplemented with PG was between 36 and 38 h and that
of PG-depleted cells was the same during the first 4 days.
Protein synthesis slowed down between the fourth and the
eighth day of culturing and stopped at a protein content of
340 lg ml-1 after 2–3 cell divisions. After 9 days of PG
starvation the protein content of the cells decreased to
230 lg ml-1.
Fig. 2 The growth of WT (1), Synechococcus sp. PCC 7942/DcdsAkanamycin (2), and Synechococcus sp. PCC 7942/DcdsA chloram-
phenicol resistant (3) cells on agar in the presence (a) and absence (b)
of exogenously added PG
Fig. 3 a Growth curve of
Synechococcus sp. PCC 7942/
DcdsA cells grown in the
presence (filled diamond) and in
the absence (filled square) of
PG, and b the colony forming
capability of PG-depleted cells
of the same mutant plated on
PG-containing agar plates after
9, 10, 11, 12, and 13 days of PG
depletion
Photosynth Res (2010) 103:19–30 23
123
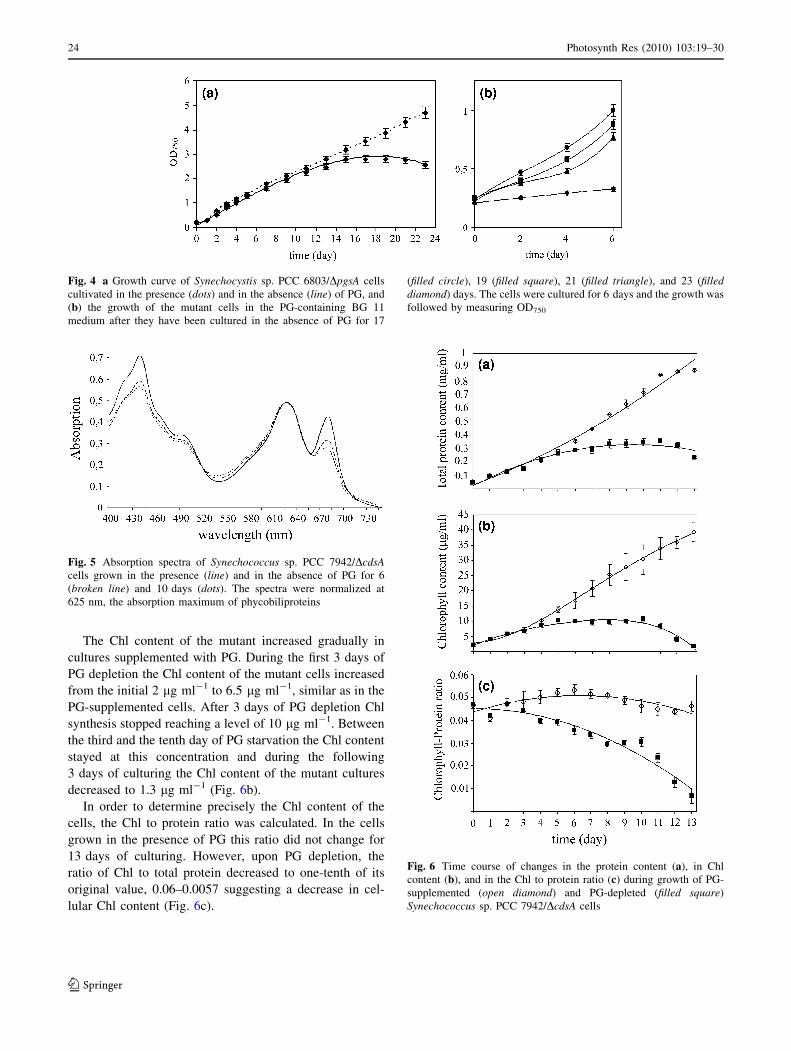
The Chl content of the mutant increased gradually in
cultures supplemented with PG. During the first 3 days of
PG depletion the Chl content of the mutant cells increased
from the initial 2 lg ml-1 to 6.5 lg ml-1, similar as in the
PG-supplemented cells. After 3 days of PG depletion Chl
synthesis stopped reaching a level of 10 lg ml-1. Between
the third and the tenth day of PG starvation the Chl content
stayed at this concentration and during the following
3 days of culturing the Chl content of the mutant cultures
decreased to 1.3 lg ml-1 (Fig. 6b).
In order to determine precisely the Chl content of the
cells, the Chl to protein ratio was calculated. In the cells
grown in the presence of PG this ratio did not change for
13 days of culturing. However, upon PG depletion, the
ratio of Chl to total protein decreased to one-tenth of its
original value, 0.06–0.0057 suggesting a decrease in cel-
lular Chl content (Fig. 6c).
Fig. 4 a Growth curve of Synechocystis sp. PCC 6803/DpgsA cells
cultivated in the presence (dots) and in the absence (line) of PG, and
(b) the growth of the mutant cells in the PG-containing BG 11
medium after they have been cultured in the absence of PG for 17
(filled circle), 19 (filled square), 21 (filled triangle), and 23 (filleddiamond) days. The cells were cultured for 6 days and the growth was
followed by measuring OD750
Fig. 5 Absorption spectra of Synechococcus sp. PCC 7942/DcdsAcells grown in the presence (line) and in the absence of PG for 6
(broken line) and 10 days (dots). The spectra were normalized at
625 nm, the absorption maximum of phycobiliproteins
Fig. 6 Time course of changes in the protein content (a), in Chl
content (b), and in the Chl to protein ratio (c) during growth of PG-
supplemented (open diamond) and PG-depleted (filled square)
Synechococcus sp. PCC 7942/DcdsA cells
24 Photosynth Res (2010) 103:19–30
123
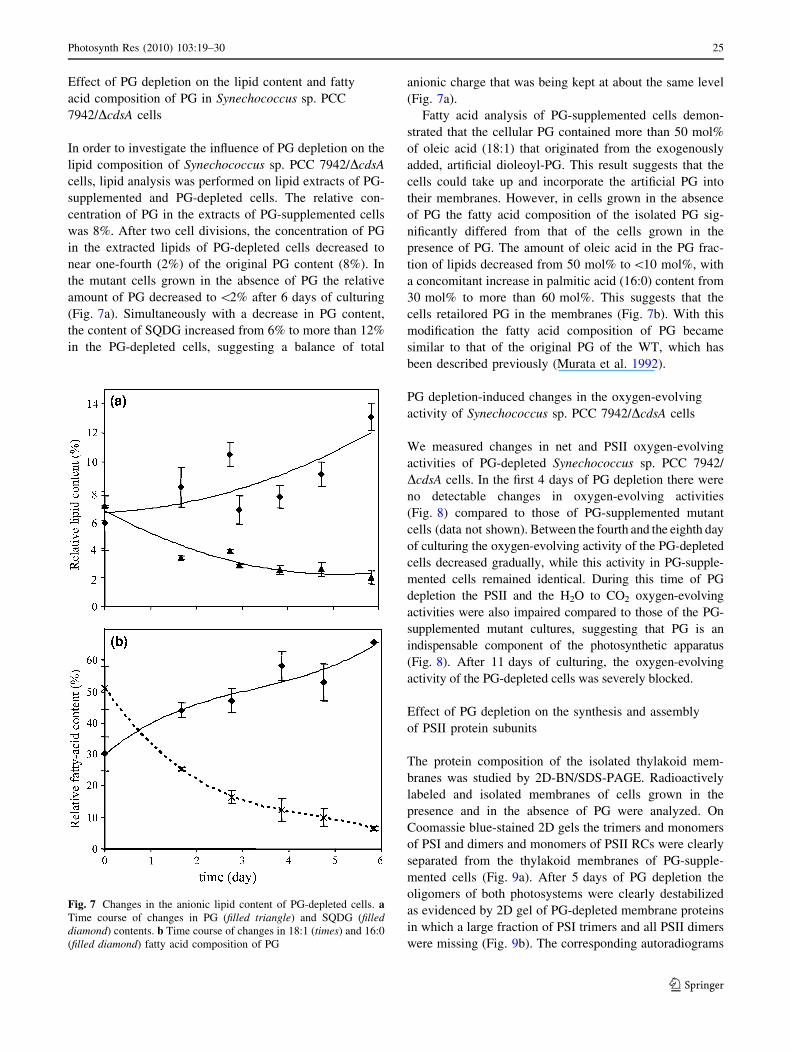
Effect of PG depletion on the lipid content and fatty
acid composition of PG in Synechococcus sp. PCC
7942/DcdsA cells
In order to investigate the influence of PG depletion on the
lipid composition of Synechococcus sp. PCC 7942/DcdsA
cells, lipid analysis was performed on lipid extracts of PG-
supplemented and PG-depleted cells. The relative con-
centration of PG in the extracts of PG-supplemented cells
was 8%. After two cell divisions, the concentration of PG
in the extracted lipids of PG-depleted cells decreased to
near one-fourth (2%) of the original PG content (8%). In
the mutant cells grown in the absence of PG the relative
amount of PG decreased to \2% after 6 days of culturing
(Fig. 7a). Simultaneously with a decrease in PG content,
the content of SQDG increased from 6% to more than 12%
in the PG-depleted cells, suggesting a balance of total
anionic charge that was being kept at about the same level
(Fig. 7a).
Fatty acid analysis of PG-supplemented cells demon-
strated that the cellular PG contained more than 50 mol%
of oleic acid (18:1) that originated from the exogenously
added, artificial dioleoyl-PG. This result suggests that the
cells could take up and incorporate the artificial PG into
their membranes. However, in cells grown in the absence
of PG the fatty acid composition of the isolated PG sig-
nificantly differed from that of the cells grown in the
presence of PG. The amount of oleic acid in the PG frac-
tion of lipids decreased from 50 mol% to\10 mol%, with
a concomitant increase in palmitic acid (16:0) content from
30 mol% to more than 60 mol%. This suggests that the
cells retailored PG in the membranes (Fig. 7b). With this
modification the fatty acid composition of PG became
similar to that of the original PG of the WT, which has
been described previously (Murata et al. 1992).
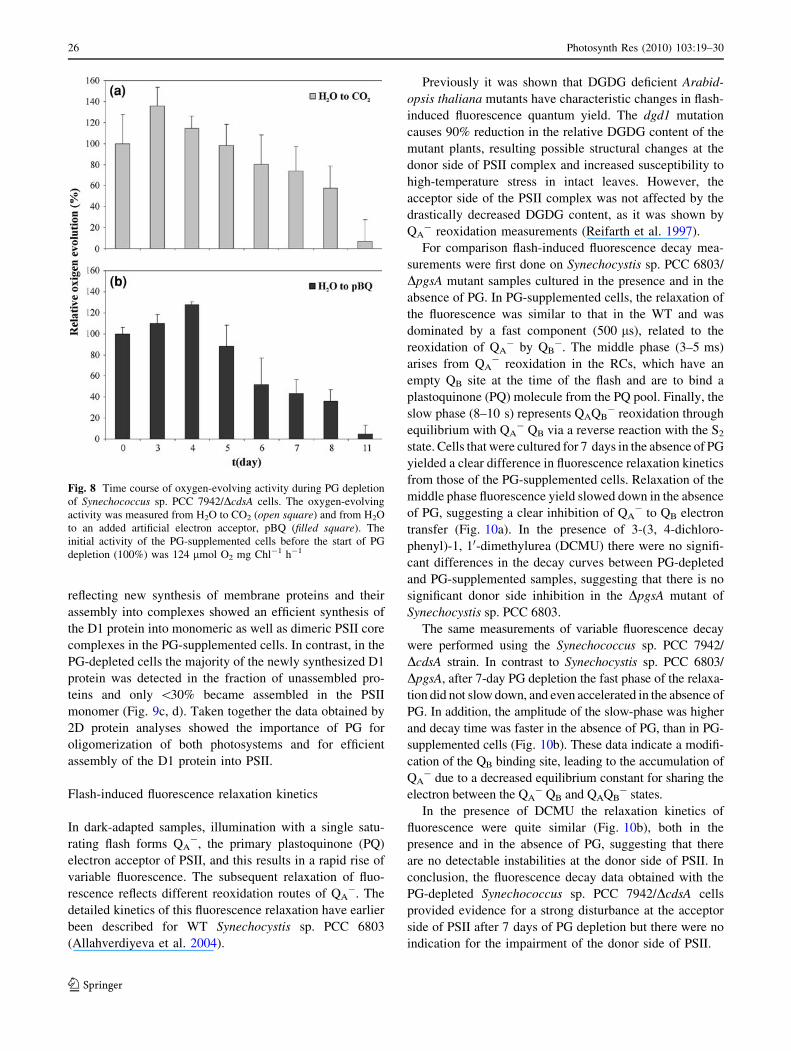
PG depletion-induced changes in the oxygen-evolving
activity of Synechococcus sp. PCC 7942/DcdsA cells
We measured changes in net and PSII oxygen-evolving
activities of PG-depleted Synechococcus sp. PCC 7942/
DcdsA cells. In the first 4 days of PG depletion there were
no detectable changes in oxygen-evolving activities
(Fig. 8) compared to those of PG-supplemented mutant
cells (data not shown). Between the fourth and the eighth day
of culturing the oxygen-evolving activity of the PG-depleted
cells decreased gradually, while this activity in PG-supple-
mented cells remained identical. During this time of PG
depletion the PSII and the H2O to CO2 oxygen-evolving
activities were also impaired compared to those of the PG-
supplemented mutant cultures, suggesting that PG is an
indispensable component of the photosynthetic apparatus
(Fig. 8). After 11 days of culturing, the oxygen-evolving
activity of the PG-depleted cells was severely blocked.
Effect of PG depletion on the synthesis and assembly
of PSII protein subunits
The protein composition of the isolated thylakoid mem-
branes was studied by 2D-BN/SDS-PAGE. Radioactively
labeled and isolated membranes of cells grown in the
presence and in the absence of PG were analyzed. On
Coomassie blue-stained 2D gels the trimers and monomers
of PSI and dimers and monomers of PSII RCs were clearly
separated from the thylakoid membranes of PG-supple-
mented cells (Fig. 9a). After 5 days of PG depletion the
oligomers of both photosystems were clearly destabilized
as evidenced by 2D gel of PG-depleted membrane proteins
in which a large fraction of PSI trimers and all PSII dimers
were missing (Fig. 9b). The corresponding autoradiograms
Fig. 7 Changes in the anionic lipid content of PG-depleted cells. aTime course of changes in PG (filled triangle) and SQDG (filleddiamond) contents. b Time course of changes in 18:1 (times) and 16:0
(filled diamond) fatty acid composition of PG
Photosynth Res (2010) 103:19–30 25
123
reflecting new synthesis of membrane proteins and their
assembly into complexes showed an efficient synthesis of
the D1 protein into monomeric as well as dimeric PSII core
complexes in the PG-supplemented cells. In contrast, in the
PG-depleted cells the majority of the newly synthesized D1
protein was detected in the fraction of unassembled pro-
teins and only \30% became assembled in the PSII
monomer (Fig. 9c, d). Taken together the data obtained by
2D protein analyses showed the importance of PG for
oligomerization of both photosystems and for efficient
assembly of the D1 protein into PSII.
Flash-induced fluorescence relaxation kinetics
In dark-adapted samples, illumination with a single satu-
rating flash forms QA-, the primary plastoquinone (PQ)
electron acceptor of PSII, and this results in a rapid rise of
variable fluorescence. The subsequent relaxation of fluo-
rescence reflects different reoxidation routes of QA-. The
detailed kinetics of this fluorescence relaxation have earlier
been described for WT Synechocystis sp. PCC 6803
(Allahverdiyeva et al. 2004).
Previously it was shown that DGDG deficient Arabid-
opsis thaliana mutants have characteristic changes in flash-
induced fluorescence quantum yield. The dgd1 mutation
causes 90% reduction in the relative DGDG content of the
mutant plants, resulting possible structural changes at the
donor side of PSII complex and increased susceptibility to
high-temperature stress in intact leaves. However, the
acceptor side of the PSII complex was not affected by the
drastically decreased DGDG content, as it was shown by
QA- reoxidation measurements (Reifarth et al. 1997).
For comparison flash-induced fluorescence decay mea-
surements were first done on Synechocystis sp. PCC 6803/
DpgsA mutant samples cultured in the presence and in the
absence of PG. In PG-supplemented cells, the relaxation of
the fluorescence was similar to that in the WT and was
dominated by a fast component (500 ls), related to the
reoxidation of QA- by QB
-. The middle phase (3–5 ms)
arises from QA- reoxidation in the RCs, which have an
empty QB site at the time of the flash and are to bind a
plastoquinone (PQ) molecule from the PQ pool. Finally, the
slow phase (8–10 s) represents QAQB- reoxidation through
equilibrium with QA- QB via a reverse reaction with the S2
state. Cells that were cultured for 7 days in the absence of PG
yielded a clear difference in fluorescence relaxation kinetics
from those of the PG-supplemented cells. Relaxation of the
middle phase fluorescence yield slowed down in the absence
of PG, suggesting a clear inhibition of QA- to QB electron
transfer (Fig. 10a). In the presence of 3-(3, 4-dichloro-
phenyl)-1, 10-dimethylurea (DCMU) there were no signifi-
cant differences in the decay curves between PG-depleted
and PG-supplemented samples, suggesting that there is no
significant donor side inhibition in the DpgsA mutant of
Synechocystis sp. PCC 6803.
The same measurements of variable fluorescence decay
were performed using the Synechococcus sp. PCC 7942/
DcdsA strain. In contrast to Synechocystis sp. PCC 6803/
DpgsA, after 7-day PG depletion the fast phase of the relaxa-
tion did not slow down, and even accelerated in the absence of
PG. In addition, the amplitude of the slow-phase was higher
and decay time was faster in the absence of PG, than in PG-
supplemented cells (Fig. 10b). These data indicate a modifi-
cation of the QB binding site, leading to the accumulation of
QA- due to a decreased equilibrium constant for sharing the
electron between the QA- QB and QAQB
- states.
In the presence of DCMU the relaxation kinetics of
fluorescence were quite similar (Fig. 10b), both in the
presence and in the absence of PG, suggesting that there
are no detectable instabilities at the donor side of PSII. In
conclusion, the fluorescence decay data obtained with the
PG-depleted Synechococcus sp. PCC 7942/DcdsA cells
provided evidence for a strong disturbance at the acceptor
side of PSII after 7 days of PG depletion but there were no
indication for the impairment of the donor side of PSII.
Fig. 8 Time course of oxygen-evolving activity during PG depletion
of Synechococcus sp. PCC 7942/DcdsA cells. The oxygen-evolving
activity was measured from H2O to CO2 (open square) and from H2O
to an added artificial electron acceptor, pBQ (filled square). The
initial activity of the PG-supplemented cells before the start of PG
depletion (100%) was 124 lmol O2 mg Chl-1 h-1
26 Photosynth Res (2010) 103:19–30
123
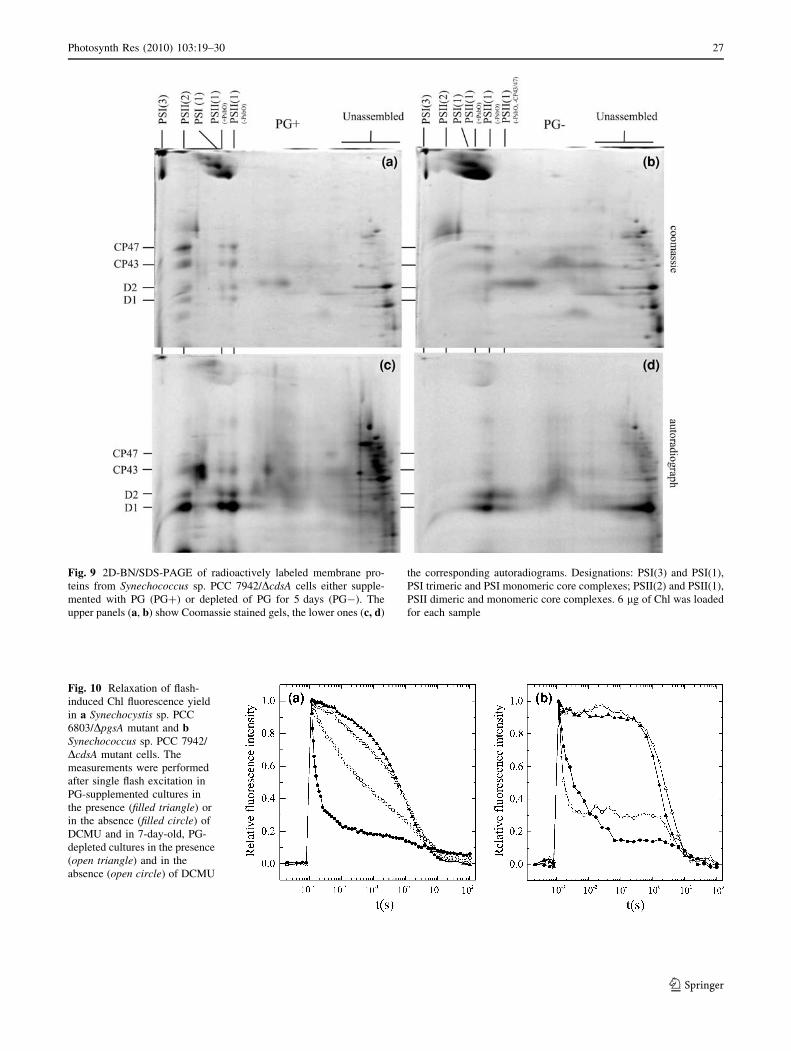
Fig. 10 Relaxation of flash-
induced Chl fluorescence yield
in a Synechocystis sp. PCC
6803/DpgsA mutant and bSynechococcus sp. PCC 7942/
DcdsA mutant cells. The
measurements were performed
after single flash excitation in
PG-supplemented cultures in
the presence (filled triangle) or
in the absence (filled circle) of
DCMU and in 7-day-old, PG-
depleted cultures in the presence
(open triangle) and in the
absence (open circle) of DCMU
Fig. 9 2D-BN/SDS-PAGE of radioactively labeled membrane pro-
teins from Synechococcus sp. PCC 7942/DcdsA cells either supple-
mented with PG (PG?) or depleted of PG for 5 days (PG-). The
upper panels (a, b) show Coomassie stained gels, the lower ones (c, d)
the corresponding autoradiograms. Designations: PSI(3) and PSI(1),
PSI trimeric and PSI monomeric core complexes; PSII(2) and PSII(1),
PSII dimeric and monomeric core complexes. 6 lg of Chl was loaded
for each sample
Photosynth Res (2010) 103:19–30 27
123

Discussion
To get a generalized view of the effects of PG on the
functions and structure of photosystems in prokaryotic
photosynthetic organisms, a particular mutant strain, Syn-
echococcus sp. PCC 7942/DcdsA, was constructed, and
subjected to studies as outlined below.
Effect of PG depletion on the growth and pigment
composition of Synechococcus sp. PCC 7942/DcdsA
cells
Inactivation of the cdsA gene gave rise to a Synechococcus
sp. PCC 7942/DcdsA mutant defective in PG synthesis. The
growth of mutant cells was suppressed upon depletion of
PG from the cells. However, similar to other mutants
defective in PG synthesis (Hagio et al. 2000; Laczko-
Dobos et al. 2008), addition of PG to the medium made the
growth rate of mutants indistinguishable from that of the
WT, suggesting that the cells require a continuous supply
of PG for photoautotrophic growth. The pigment compo-
sition of cells was altered by PG depletion. Similar to
Synechocystis sp. PCC 6803/DpgsA, the Chl content
gradually decreased simultaneously with a decrease in PG
content of the cells, although the phycobiliprotein content
did not change significantly. The growth of Synechococcus
sp. PCC 7942/DcdsA slowed down and stopped earlier than
that of Synechocystis sp. PCC 6803/DpgsA. This demon-
strates that PG depletion has a more severe effect on cel-
lular functions in Synechococcus sp. PCC 7942 than in
Synechocystis sp. PCC 6803 cells. While the Synechocystis
sp. PCC 6803 mutant defective in PG synthesis stopped
growing after 17 days of PG depletion, this occurred earlier
(7 days after PG depletion) with Synechococcus sp. PCC
7942/DcdsA cells. The recovery of PG-depleted Synecho-
cystis sp. PCC 6803/DpgsA was superior to that of Syn-
echococcus sp. PCC 7942/DcdsA. Following 23 days of PG
depletion the readdition of PG led to an incomplete
recovery.
While complete recovery occurred even after 21 days of
PG depletion corresponding to four cell divisions of Syn-
echocystis sp. PCC 6803 mutant cells, the Synechococcus
sp. PCC 7942 mutant cells could not recover after 12 days
of PG depletion which corresponds to two cell divisions.
Effect of PG depletion on the lipid content and fatty
acid composition of PG in DcdsA cells
Decrease in PG content of Synechococcus sp. PCC 7942/
DcdsA cells was followed by gas chromatography. Fol-
lowing two cell divisions, the PG content of the cells
decreased to one-fourth of the original value. The decrease
of PG was compensated for by an increase in SQDG
content of Synechococcus sp. PCC 7942/DcdsA cells.
Earlier it has been postulated that in photosynthetic
organisms loss in PG following severe phosphate limitation
can be balanced by an increase in SQDG content (Yu and
Benning 2003). PG can be substituted by SQDG ensuring a
constant sum of anionic charged lipids in the cells. Sulfur-
starvation concomitantly upregulated PG synthesis and this
could compensate for a decrease in SQDG content
(Sugimoto et al. 2008). Our analysis of fatty acid content
showed that the exogenously added dioleoyl-phosphati-
dylglycerol was converted to more essential PG which
contains palmitic acid at position sn-2 (Murata et al. 1992).
The oleic acid esterified to the sn-1 position of the glycerol
backbone was replaced by a stearyl residue that normally
binds to this position (Murata et al. 1992). This retailoring
process was earlier detected in Synechococcus sp. PCC
6301 as a result of nitrate starvation (Gombos et al. 1987).
Effect of PG depletion on the synthesis and assembly
of PSII protein subunits
Protein analysis of Synechococcus sp. PCC 7942/DcdsA by
2D-BN/SDS-PAGE supported our earlier results on the
effect of PG depletion on the protein content of PAL/
DcdsA (Laczko-Dobos et al. 2008). PG depletion leads to
conversion of both the dimeric form of PSII and the tri-
meric form of PSI into their respective monomer forms.
This is a sign of a general destabilization of RC oligomers.
However, Takahashi and coworkers suggested that in intact
cells PSII exists in monomeric form (Takahashi et al.
2009). Formation of PSII RCs was suppressed and it
occurred much earlier in PG-depleted Synechococcus sp.
PCC 7942/DcdsA than in PG-depleted Synechocystis sp.
PCC 6803/DpgsA cells. This suggests that PG is a more
important partner of protein subunits for RC oligomer
formation in Synechococcus sp. PCC 7942 than in Syn-
echocystis sp. PCC 6803. In addition, the finding of the
majority of newly synthesized D1 among the non-assem-
bled thylakoid proteins supports the view that PG is also
necessary for a correct incorporation of membrane proteins
into the complexes.
Effect of PG depletion on photosynthetic activities
PG depletion affects photosynthetic processes in both
cyanobacterial species studied so far. In Synechocystis sp.
PCC 6803/DpgsA defective in PG synthesis the oxygen-
evolving activity may decrease to 60% of its original value
following 9 days of PG depletion (Hagio et al. 2000).
Suppression of photosynthetic activity was even more
extensive in Synechococcus sp. PCC 7942/DcdsA in which
an 11-day-long PG depletion was enough to decrease
photosynthetic activity to about 10% of its original value.
28 Photosynth Res (2010) 103:19–30
123

Whereas the effects of a 21-day-long PG depletion were
reversible in Synechocystis sp. PCC 6803/DpgsA cells, in
Synechococcus sp. PCC 7942/DcdsA cells 14 days of PG
depletion were sufficient to inhibit photosynthetic pro-
cesses irreversibly and readdition of PG to the cells did not
restore photosynthesis. Apparently, SQDG, another anionic
lipid constituent of photosynthetic membranes is not an
obligatory component of the photosynthetic machinery.
The SQDG-deficient mutant of Synechococcus sp. PCC
7942 grew without exogenously added lipid and had active
PSII without SQDG (Aoki et al. 2004; Guler et al. 2000).
Since the increase in SQDG content on the expense of PG
content could not restore photosynthetic activities in Syn-
echococcus sp. PCC 7942/DcdsA cells, we conclude that
PG is indispensably needed for this photosynthetic organ-
ism. The result suggests that maintaining the constant sum
of negatively charged lipids is not enough for the mainte-
nance of active electron transport processes.
We concluded as follows:
i. Synechococcus sp. PCC 7942/DcdsA is a mutant
defective in PG synthesis and is more susceptible to
PG depletion than Synechocystis sp. PCC 6803/DpgsA
or PAL/DcdsA.
ii. PG depletion affects the lipid composition of Syn-
echococcus sp. PCC 7942/DcdsA. The decrease in PG
content is counterbalanced by an increase in SQDG
content in the cells. Added artificial dioleoyl-PG was
transformed to essential PG species originally present
in the WT cells. However, neither the increased level
of SQDG nor the retailoring of fatty acid composition
can compensate for the negative effects of the reduced
PG content.
iii. The oligomerization of PSII and PSI was severely
perturbed by PG depletion in Synechococcus sp. PCC
7942/DcdsA following a short-term PG depletion. The
newly synthesized D1 protein was observed in the
non-assembled protein region of 2D gels showing the
importance of the lipid for the efficient assembly of
proteins into complexes.
iv. PG depletion resulted in a relatively rapid decrease in
PSII oxygen-evolving activity related to the pertur-
bation on the acceptor side of PSII while its donor side
was not primarily inactivated. In contrast to Synecho-
cystis sp. PCC 6803/DpgsA, perturbation of the PSII
acceptor site in Synechococcus sp. PCC 7942/DcdsA
is not caused by the PG-deficiency-induced inhibition
of electron transport between QA and QB, but seems to
be due to a decreased equilibrium constant for sharing
the electron between the QA- QB and QAQB
- states.
v. Synechococcus sp. PCC 7942/DcdsA provided a new
experimental system to support the importance of PG
in the correct formation of photosynthetic RCs and in
the functions of electron transport in an obligatory
photoautotrophic cyanobacterium.
Acknowledgments The authors thank Dr. Ghada Ajlani, CEA Sa-
clay, Gif-sur-Yvette, France, for her advice in preparing the cdsAplasmid construction, as well as for reading and correcting the man-
uscript. We are grateful to Prof. Ferenc Solymosy for reading and
correcting the manuscript. This work was supported by grants from
the Hungarian Science Foundation (OTKA; grant nos. T 60109 and T
68692), by the Ministry of Education, Youth and Sports of the Czech
Republic (project no. MSM6007665808) and by the Czech Academy
of Sciences (Institutional Research Concept no. AV0Z50200510,
Czech–Hungarian bilateral research priority project and project
IAA400200801).
References
Allahverdiyeva Y, Deak Z, Szilard A, Diner BA, Nixon PJ, Vass I
(2004) The function of D1-H332 in photosystem II electron
transport studied by thermoluminescence and chlorophyll fluo-
rescence in site-directed mutants of Synechocystis 6803. Eur J
Biochem 271:3523–3532
Aoki M, Sato N, Meguro A, Tsuzuki M (2004) Differing involvement
of sulfoquinovosyl diacylglycerol in photosystem II in two
species of unicellular cyanobacteria. Eur J Biochem 271:685–
693
Ariizumi T, Kishitani S, Inatsugi R, Nishida I, Murata N, Toriyama K
(2002) An increase in unsaturation of fatty acids in phosphat-
idylglycerol from leaves improves the rates of photosynthesis
and growth at low temperatures in transgenic rice seedlings.
Plant Cell Physiol 43:751–758
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction
and purification. Can J Biochem Physiol 37:911–917
Domonkos I, Malec P, Sallai A, Kovacs L, Itoh K, Shen GZ, Ughy B,
Bogos B, Sakurai I, Kis M, Strzalka K, Wada H, Itoh S, Farkas
T, Gombos Z (2004) Phosphatidylglycerol is essential for
oligomerization of photosystem I reaction center. Plant Physiol
134:1471–1478
Domonkos I, Laczko-Dobos H, Gombos Z (2008) Lipid-assisted
protein–protein interactions that support photosynthetic and
other cellular activities. Prog Lipid Res 47:422–435
Golden SS, Sherman LA (1984) Optimal conditions for genetic
transformation of the cyanobacterium Anacystis nidulans R2.
J Bacteriol 158:36–42
Gombos Z, Kis M, Pali T, Vigh L (1987) Nitrate starvation induces
homeoviscous regulation of lipids in the cell envelope of the
blue-green-alga, Anacystis nidulans. Eur J Biochem 165:461–
465
Gombos Z, Wada H, Murata N (1991) Direct evaluation of effects of
fatty-acid unsaturation on the thermal properties of photosyn-
thetic activities, as studied by mutation and transformation of
Synechocystis PCC6803. Plant Cell Physiol 32:205–211
Gombos Z, Varkonyi Z, Hagio M, Iwaki M, Kovacs L, Masamoto K,
Itoh S, Wada H (2002) Phosphatidylglycerol requirement for the
function of electron acceptor plastoquinone Q(B) in the photo-
system II reaction center. Biochemistry 41:3796–3802
Guler S, Seeliger A, Hartel H, Renger G, Benning C (1996) A null
mutant of Synechococcus sp PCC7942 deficient in the sulfolipid
sulfoquinovosyl diacylglycerol. J Biol Chem 271:7501–7507
Guler S, Essigmann B, Benning C (2000) A cyanobacterial gene,
sqdX, required for biosynthesis of the sulfolipid sulfoquinovo-
syldiacylglyerol. J Bacteriol 182:543–545
Photosynth Res (2010) 103:19–30 29
123

Guskov A, Kern J, Gabdulkhakov A, Broser M, Zouni A, Saenger W
(2009) Cyanobacterial photosystem II at 2.9-angstrom resolution
and the role of quinones, lipids, channels and chloride. Nat Struct
Mol Biol 16:334–342
Hagio M, Gombos Z, Varkonyi Z, Masamoto K, Sato N, Tsuzuki M,
Wada H (2000) Direct evidence for requirement of phosphati-
dylglycerol in photosystem II of photosynthesis. Plant Physiol
124:795–804
Hagio M, Sakurai I, Sato S, Kato T, Tabata S, Wada H (2002)
Phosphatidylglycerol is essential for the development of thyla-
koid membranes in Arabidopsis thaliana. Plant Cell Physiol 43:
1456–1464
Ihlenfeldt MJA, Gibson J (1975) Phosphate utilization and alkaline-
phosphatase activity in Anacystis nidulans (Synechococcus).
Arch Microbiol 102:23–28
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, Krauss N
(2001) Three-dimensional structure of cyanobacterial photosys-
tem I at 2.5 angstrom resolution. Nature 411:909–917
Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y,
Miyajima N, Hirosawa M, Sugiura M, Sasamoto S, Kimura T,
Hosouchi T, Matsuno A, Muraki A, Nakazaki N, Naruo K,
Okumura S, Shimpo S, Takeuchi C, Wada T, Watanabe A,
Yamada M, Yasuda M, Tabata S (1996) Sequence analysis of the
genome of the unicellular cyanobacterium Synechocystis sp.
strain PCC6803. II. Sequence determination of the entire
genome and assignment of potential protein-coding regions.
DNA Res 3:109–136
Komenda J, Barber J (1995) Comparison of psbO and psbH deletion
mutants of Synechocystis PCC6803 indicates that degradation of
D1 protein is regulated by the Q(B) site and dependent on
protein synthesis. Biochemistry 34:9625–9631
Komenda J, Reisinger V, Muller BC, Dobakova M, Granvogl B,
Eichacker LA (2004) Accumulation of the D2 protein is a key
regulatory step for assembly of the photosystem II reaction
center complex in Synechocystis PCC 6803. J Biol Chem
279:48620–48629
Kruse O, Hankamer B, Konczak C, Gerle C, Morris E, Radunz A,
Schmid GH, Barber J (2000) Phosphatidylglycerol is involved in
the dimerization of photosystem II. J Biol Chem 275:6509–6514
Laczko-Dobos H, Ughy B, Toth SZ, Komenda J, Zsiros O, Domonkos
I, Parducz A, Bogos B, Komura M, Itoh S, Gombos Z (2008)
Role of phosphatidylglycerol in the function and assembly of
photosystem II reaction center, studied in a cdsA-inactivated
PAL mutant strain of Synechocystis sp. PCC6803 that lacks
phycobilisomes. Biochim Biophys Acta 1777:1184–1194
Loll B, Kern J, Saenger W, Zouni A, Biesiadka J (2005) Towards
complete cofactor arrangement in the 3.0 angstrom resolution
structure of photosystem II. Nature 438:1040–1044
Lutz R, Bujard H (1997) Independent and tight regulation of
transcriptional units in Escherichia coli via the LacR/O, the
TetR/O and AraC/I-1-I-2 regulatory elements. Nucleic Acids Res
25:1203–1210
Mayer MP (1995) A new set of useful cloning and expression vectors
derived from pBlueScript. Gene 163:41–46
Murata N, Wada H, Gombos Z (1992) Modes of fatty-acid
desaturation in cyanobacteria. Plant Cell Physiol 33:933–941
Reifarth F, Christen G, Seeliger AG, Dormann P, Benning C, Renger G
(1997) Modification of the water oxidizing complex in leaves of the
dgd1 mutant of Arabidopsis thaliana deficient in the galactolipid
digalactosyldiacylglycerol. Biochemistry 36:11769–11776
Sakurai I, Hagio M, Gombos Z, Tyystjarvi T, Paakkarinen V, Aro EM,
Wada H (2003) Requirement of phosphatidylglycerol for mainte-
nance of photosynthetic machinery. Plant Physiol 133:1376–1384
Sakurai I, Mizusawa N, Ohashi S, Kobayashi M, Wada H (2007)
Effects of the lack of phosphatidylglycerol on the donor side of
photosystem II. Plant Physiol 144:1336–1346
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a
laboratory manual. Cold Spring Harbor Laboratory Press, New
York
Sato N (2004) Roles of the acidic lipids sulfoquinovosyl diacylgly-
cerol and phosphatidylglycerol in photosynthesis: their specific-
ity and evolution. J Plant Res 117:495–505
Sato N, Tsuzuki M, Matsuda Y, Ehara T, Osafune T, Kawaguchi A
(1995) Isolation and characterization of mutants affected in
lipid-metabolism of Chlamydomonas reinhardtii. Eur J Biochem
230:987–993
Sato N, Hagio M, Wada H, Tsuzuki M (2000) Requirement of
phosphatidylglycerol for photosynthetic function in thylakoid
membranes. Proc Natl Acad Sci USA 97:10655–10660
Schagger H, von Jagow G (1991) Blue native electrophoresis for
isolation of membrane-protein complexes in enzymatically active
form. Anal Biochem 199:223–231
Shibata K (1958) Spectrophotometry of intact biological materials.
Absolute and relative measurements of their transmission, reflec-
tion and absorption spectra. J Biochem (Tokyo) 45:599–623
Sugimoto K, Midorikawa T, Tsuzuki M, Sato N (2008) Upregulation
of PG synthesis on sulfur-starvation for PS I in Chlamydomonas.
Biochem Biophys Res Commun 369:660–665
Takahashi T, Inoue-Kashino N, Ozawa S, Takahashi Y, Kashino Y,
Satoh K (2009) Photosystem II complex in vivo is a monomer.
J Biol Chem 284:15598–15606
Trtilek M, Kramer DM, Koblizek M, Nedbal L (1997) Dual-
modulation LED kinetic fluorometer. J Lumin 72(4):597–599
Wada H, Murata N (1989) Synechocystis PCC6803 mutants defective
in desaturation of fatty-acids. Plant Cell Physiol 30:971–978
Wada H, Murata N (1998) Membrane lipids in cyanobacteria. In:
Siegenthaler P-A, Murata N (eds) Lipids in photosynthesis.
Kluwer, Dordrecht, pp 65–81
Xu CC, Hartel H, Wada H, Hagio M, Yu B, Eakin C, Benning C (2002)
The pgp1 mutant locus of Arabidopsis encodes a phosphatidyl-
glycerolphosphate synthase with impaired activity. Plant Physiol
129:594–604
Yeang HY, Yusof F, Abdullah L (1998) Protein purification for the
Lowry assay: acid precipitation of proteins in the presence of
sodium dodecyl sulfate and other biological detergents. Anal
Biochem 265:381–384
Yu B, Benning C (2003) Anionic lipids are required for chloroplast
structure and function in Arabidopsis. Plant J 36:762–770
Zouni A, Witt HT, Kern J, Fromme P, Krauss N, Saenger W, Orth P
(2001) Crystal structure of photosystem II from Synechococcuselongatus at 3.8 angstrom resolution. Nature 409:739–743
30 Photosynth Res (2010) 103:19–30
123
Related Documents











