Phenylbenzopyrones structure-activity studies identify betuletol derivatives as potential antitumoral agents Sara Rubio a , José Quintana a , Mariana López b , José Luis Eiroa b , Jorge Triana b , Francisco Estévez a, ⁎ a Department of Biochemistry, I.C.I.C., University of Las Palmas de Gran Canaria, Plaza Dr. Pasteur s/n, 35016 Las Palmas de Gran Canaria, Spain b Department of Chemistry, I.C.I.C., University of Las Palmas de Gran Canaria, Campus Universitario de Tafira, 35017 Las Palmas de Gran Canaria, Spain Received 10 March 2006; received in revised form 20 June 2006; accepted 13 July 2006 Available online 22 July 2006 Abstract We have analyzed the cytotoxicity of 22 compounds with a phenylbenzo-γ-pirone core structure, most of them obtained from natural sources, in five human tumor cell lines (HL-60, A431, SK-OV-3, HeLa and HOS). Betuletol 3-methyl ether and its diacetate were the most cytotoxic compounds. The HL-60 cell line was especially sensitive to these compounds, with IC 50 values of approximately 1 μM. Treatment of HL-60 cells with betuletol 3-methyl ether was associated with apoptosis induction which was prevented by a non-specific caspase inhibitor (z-VAD-fmk) and also by a specific inhibitor of caspase-8 (z-IETD-fmk) indicating activation of the extrinsic apoptotic pathway. The results suggest that betuletol 3- methyl ether has potential as new anticancer agent. © 2006 Elsevier B.V. All rights reserved. Keywords: Apoptosis; Flavonoids; Cytotoxic activity; Caspases; DNA fragmentation; poly(ADP-ribose) polymerase 1. Introduction Many anticancer drugs have been shown to cause the death of sensitive cells through the induction of apoptosis. This cellular suicide program is initiated by ligation of death receptors, growth factor deprivation or various environmental stresses which are mediated by molecular pathways that culminate in the activation of a conserved family of aspartate- specific cysteine proteases, known as the caspases, which orchestrate the dismantling and clearance of the dying cell. Caspases themselves are present as proenzymes that are readily cleaved and activated during apoptosis, providing the cell with a means to rapidly amplify its apoptotic response (Cohen, 1997; Thornberry and Lazebnik, 1998). Two major pathways have been identified for the induction of apoptosis. The intrinsic pathway is activated by release of mitochondrial cytochrome c (Kluck et al., 1997; Liu et al., 1996; Yang et al., 1997) which forms a complex with Apaf-1 and activates caspase-9 leading to activation of caspase-3 (Li et al., 1997; Srinivasula et al., 1998). The extrinsic pathway is initiated by activation of death receptors and thereby cleavage of procaspase-8 (Boldin et al., 1996; Muzio et al., 1996). Caspase-8 directly activates caspase- 3(Stennicke et al., 1998). Caspase-8 also cleaves Bid-a Bcl-2 family member— and thereby plays a role in the release of cytochrome c. However, it is becoming increasingly clear that apoptosis can also occur independently of caspase activation (Chipuk and Green, 2005). Flavonoids comprise a vast array of biologically active compounds ubiquitous in plants, many of which have been used in traditional Eastern medicine for thousands of years. These polyphenolic compounds exert several biological functions such as apoptosis-inducing activity, free radical scavenging activity, and anti-tumorigenic activity (Ko et al., 2002; Lee et al., 2002). Here we have studied the effect of twenty two flavonoids, most of them isolated from endemic plants to the Canary Islands, on cell viability of five human tumor cell lines. The betuletol derivatives, betuletol 3-methyl ether and betuletol 3-methyl ether diacetate were the most potent cytotoxic compounds in all cell lines studied, but also the most potent apoptotic inducers on European Journal of Pharmacology 548 (2006) 9 – 20 www.elsevier.com/locate/ejphar ⁎ Corresponding author. Tel.: +34 928 451443; fax: +34 928 451441. E-mail address: [email protected] (F. Estévez). 0014-2999/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2006.07.020

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

logy 548 (2006) 9–20www.elsevier.com/locate/ejphar
European Journal of Pharmaco
Phenylbenzopyrones structure-activity studies identify betuletol derivativesas potential antitumoral agents
Sara Rubio a, José Quintana a, Mariana López b, José Luis Eiroa b, Jorge Triana b,Francisco Estévez a,⁎
a Department of Biochemistry, I.C.I.C., University of Las Palmas de Gran Canaria, Plaza Dr. Pasteur s/n, 35016 Las Palmas de Gran Canaria, Spainb Department of Chemistry, I.C.I.C., University of Las Palmas de Gran Canaria, Campus Universitario de Tafira, 35017 Las Palmas de Gran Canaria, Spain
Received 10 March 2006; received in revised form 20 June 2006; accepted 13 July 2006Available online 22 July 2006
Abstract
We have analyzed the cytotoxicity of 22 compounds with a phenylbenzo-γ-pirone core structure, most of them obtained from natural sources,in five human tumor cell lines (HL-60, A431, SK-OV-3, HeLa and HOS). Betuletol 3-methyl ether and its diacetate were the most cytotoxiccompounds. The HL-60 cell line was especially sensitive to these compounds, with IC50 values of approximately 1 μM. Treatment of HL-60 cellswith betuletol 3-methyl ether was associated with apoptosis induction which was prevented by a non-specific caspase inhibitor (z-VAD-fmk) andalso by a specific inhibitor of caspase-8 (z-IETD-fmk) indicating activation of the extrinsic apoptotic pathway. The results suggest that betuletol 3-methyl ether has potential as new anticancer agent.© 2006 Elsevier B.V. All rights reserved.
Keywords: Apoptosis; Flavonoids; Cytotoxic activity; Caspases; DNA fragmentation; poly(ADP-ribose) polymerase
1. Introduction
Many anticancer drugs have been shown to cause the deathof sensitive cells through the induction of apoptosis. Thiscellular suicide program is initiated by ligation of deathreceptors, growth factor deprivation or various environmentalstresses which are mediated by molecular pathways thatculminate in the activation of a conserved family of aspartate-specific cysteine proteases, known as the caspases, whichorchestrate the dismantling and clearance of the dying cell.Caspases themselves are present as proenzymes that are readilycleaved and activated during apoptosis, providing the cell with ameans to rapidly amplify its apoptotic response (Cohen, 1997;Thornberry and Lazebnik, 1998). Two major pathways havebeen identified for the induction of apoptosis. The intrinsicpathway is activated by release of mitochondrial cytochrome c(Kluck et al., 1997; Liu et al., 1996; Yang et al., 1997) whichforms a complex with Apaf-1 and activates caspase-9 leading to
⁎ Corresponding author. Tel.: +34 928 451443; fax: +34 928 451441.E-mail address: [email protected] (F. Estévez).
0014-2999/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2006.07.020
activation of caspase-3 (Li et al., 1997; Srinivasula et al., 1998).The extrinsic pathway is initiated by activation of deathreceptors and thereby cleavage of procaspase-8 (Boldin et al.,1996; Muzio et al., 1996). Caspase-8 directly activates caspase-3 (Stennicke et al., 1998). Caspase-8 also cleaves Bid-a Bcl-2family member— and thereby plays a role in the release ofcytochrome c. However, it is becoming increasingly clear thatapoptosis can also occur independently of caspase activation(Chipuk and Green, 2005).
Flavonoids comprise a vast array of biologically activecompounds ubiquitous in plants, many of which have been usedin traditional Eastern medicine for thousands of years. Thesepolyphenolic compounds exert several biological functions suchas apoptosis-inducing activity, free radical scavenging activity,and anti-tumorigenic activity (Ko et al., 2002; Lee et al., 2002).Here we have studied the effect of twenty two flavonoids, mostof them isolated from endemic plants to the Canary Islands, oncell viability of five human tumor cell lines. The betuletolderivatives, betuletol 3-methyl ether and betuletol 3-methylether diacetate were the most potent cytotoxic compounds in allcell lines studied, but also the most potent apoptotic inducers on

10 S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
human myeloid leukemia HL-60 cells. Increased levels of anti-apoptotic or decreased levels of pro-apoptotic proteins can causeacquired or intrinsic resistance against apoptosis induction byantineoplastic drugs thereby contributing to treatment failureand poor clinical prognosis. Thus, novel agents targetingaberrant apoptosis pathways or inducing alternative deathpathways may be suited to overcome treatment resistance(Igney and Krammer, 2002). The present studies demonstratethat betuletol 3-methyl ether induces apoptosis, at least in largepart, by activation of the extrinsic caspase-8 pathway of death.
2. Materials and methods
2.1. Materials
Most of the flavonoids used in this study were obtained fromthe Canary Islands' endemic plants and were isolated accordingto published methods with minor modifications. Structuralidentities of flavonoids were determined spectroscopically(proton nuclear magnetic resonance and 13C nuclear magneticresonance, infrared and UV/Visible spectroscopy and massspectrometry) as described previously: Betuletol 3-methyl ether1, quercetin 3,3′-dimethyl ether 7, kaempferol 3-methyl ether 9,5,7-dihydroxy-3,3′,4′-trimethoxyflavone 10, 4′, 5, 7-trihy-droxy-3,6-dimethoxyflavone 12 and eriodictyol 19 wereisolated from the aerial parts of Allagopappus dichotomus asdescribed (González et al., 1995). Quercetin 3-methyl ether 5,naringenin 21, eriodictyol 19 and kaempferol 3-methyl ether 9were isolated from Allagopappus viscosissimus as described(González et al., 1992a). Axillarin 11 and apigenin 22 wereisolated from the aerial parts of Tanacetum ferulaceum and Ta-nacetum ptarmiciflorumas described (González et al., 1990,1992b). Acetyl derivatives of betuletol 3-methyl ether (com-pound 2), quercetin 3-methyl ether (compound 6), quercetin3,3′-dimethyl ether (compound 8) and eriodictyol (compound20) were obtained by treatment of the corresponding alcoholswith acetic anhydride (Ac2O) in pyridine for 12 h at roomtemperature. Betuletol 3, 5, 7-trimethyl ether 3 was obtained bymethylation of 4′,5,7-trihydroxy-3, 6-dimethoxyflavone 12.Mono-di- and triacetyl derivatives (compounds 13, 14 and 15)of 4′,5,7-trihydroxy-3, 6-dimethoxyflavone 12were obtained byacetylation of this compound with acetic anhydride in pyridineas above. The acetyl derivatives were purified by chromatog-raphy on a silica gel column, eluted with hexane, and hexane-ethyl acetate mixtures (8:2). The mono and diacetyl derivatives,7-Acetoxy-4′,5-dihydroxy-3,6-dimethoxyflavone 13 and 5,7-diacetoxy-4′-hydroxy-3,6-dimethoxyflavone 14, are describedfor the first time.
7-Acetoxy-4′,5-dihydroxy-3,6-dimethoxyflavone 13: UV/Vis λmax nm, (MeOH): 289, 316,330; (+NaOMe): 286, 325,398; (+AlCl3): 294, 350, 396; (+AlCl3+ HCl): same as AlCl3;(+NaOAc): 290, 376; (+NaOAc+H3BO3): same as NaOAc.PMR (300 MHz, CDCl3): δ 8.11 (d, J=8.8 Hz, H-2′, H-6′),7.23 (d, J=8.8 Hz, H-3′, H-5′), 6.76 (s, H-8), 4.04 (s, OMe),3.87 (s, OMe), 2.35 (s, OAc). EIMS m/z (rel. int.): 372 [M]+
(100), 357 (15.6), 330 (41.1), 329 (45.8), 287 (39.1), 269 (17.8),121 (22.9), 69 (66.0).
5,7-diacetoxy-4′-hydroxy-3,6-dimethoxyflavone 14: UV/Vis λmax nm, (MeOH): 287, 323,329; (+NaOMe): 284, 325,393; (+AlCl3): same as MeOH; (+AlCl3+ HCl): same as AlCl3;(+NaOAc): 287, 360; (+NaOAc+H3BO3): 287,325. PMR(300 MHz, CDCl3): δ 8.06 (d, J=8.8, H-2′, H-6′), 7.21 (d,J=8.5, H-3′, H-5′), 6.89 (s, H-8), 3.94 (s, OMe), 3.76 (s, OMe),2.50 (s, OAc), 2.33 (s, OAc). EIMS m/z (rel. int.): 413 [M]+
(36.6), 372 (100), 330 (49.2), 315 (46.8), 312 (42.8), 287 (22.5),269 (10.6), 121 (14.9), 69 (15.6).
Quercetin 4 (3,3′,4′,5,7-pentahydroxyflavone) was obtainedfrom Sigma. Cryptostrobin 18 was isolated from Rumohraadiantiformis. The structure of this compound was determinedby a combination of spectroscopic analysis and comparisonwith reported data (Byrne et al., 1982).
Dihydroquercetin 7,3′-dimethyl ether 16 and dihydrokaemp-ferol 7,4′-dimethyl ether 17 were isolated from the aerial partsof Pulicaria canariensis as recently described (Triana et al.,2005). All other reagents were of analytical grade. Purity of allcompounds was 99.0% as judged by high-performance liquidchromatography. Stock solutions of 10 mM flavonoids weremade in dimethyl sulfoxide (DMSO), and aliquots were frozenat −20 °C. Polyvinylidene difluoride (PVDF) membranes werepurchased from Millipore. Antibodies for poly(ADP-ribose)polymerase (PARP), caspase-3, caspase-8 and caspase-9 werepurchased from Stressgen. Antibody for cytochrome c waspurchased from BD PharMingen. Secondary antibodies werefrom Amersham Pharmacia Biotech. The caspase inhibitorswere from Sigma. The caspase-3 and caspase-8 substrates werefrom Sigma. The caspase-9 substrate was from AnaSpec.
2.2. Cell culture
HL-60 cells were cultured in RPMI 1640 medium containing10% (v/v) heat-inactivated fetal bovine serum and 100 units/mlpenicillin and 100 μg/ml streptomycin at 37 °C in a humidifiedatmosphere containing 5% CO2. The cultures were passed twiceweekly exhibiting characteristic doubling times of∼24 h. The cellnumbers were counted by a hematocytometer, and the viability wasalways greater that 95% in all experiments as assayed by the0.025% trypan blue exclusion method. Further dilutions of stocksolutions of flavonoids were made in culture media just before use.In all experiments, the final concentration of DMSOdid not exceed0.3% (v/v), a concentration which is non-toxic to the cells. Thesame concentration was present in control experiments. The A431cell linewas grown asmonolayers in plastic tissue flasks containingDulbecco's modified Eagle's medium (DMEM) supplementedwith 10% fetal bovine serum, 100 units/ml penicillin and 100μg/mlstreptomycin. SK-OV-3 was grown as monolayers in DMEM. Thehuman cervix carcinoma HeLa and human osteosarcoma HOScells were grown in Eagle's minimum essential medium (EMEM)supplementedwith 2mMglutamine, 1%Non essential amino acidsand 10% fetal bovine serum. Cells at logarithmic growth phasewere harvested using 0.25% trypsin-EDTA solution for 5 min,pelleted by centrifugation at 500 ×g for 5 min, washed once withmedium and resuspended at a concentration of 1×104 cells/ml.These cell lineswere obtained from theEuropeanCollection ofCellCultures (Salisbury, UK).

11S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
Human peripheral blood mononuclear cells (PBMC) wereisolated from heparin-anticoagulated blood of healthy volunteersby centrifugation with Ficoll–Paque Plus (Amersham Bios-ciences). PBMCs were also stimulated with phytohemagglutinine(PHA, 2 μg/ml) for 48 h before experimental treatment.
2.3. Cytotoxicity of flavonoids on HL-60 cells, A431 cells,HeLa cells, SK-OV-3 cells and HOS cells
The cytotoxicity of flavonoids in HL-60 cells was analyzedby colorimetric 3-(4,5-dimethyl-2-thiazolyl-)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay. Briefly, 1×104 exponential-ly growing cells were seeded in 96-well microculture plateswith various flavonoids concentrations. After the addition ofMTT (0.5 mg/ml) cells were incubated at 37 °C for 4 h. Sodiumdodecyl sulfate (SDS) (10% w/v) in 0.05 M HCl was added tothe wells and then incubated at room temperature overnightunder dark conditions. The absorbance was measured at570 nm. Concentrations inducing a 50% inhibition of cellgrowth (IC50) were determined graphically for each experimentas described (Rivero et al., 2003). A431, SK-OV-3, HeLa andHOS cells (5×103−1×104) were inoculated in 96-well platesand were then incubated at 37 °C for 24 h to allow theattachment to plates after which the culture media were changedand flavonoids were added and provided different finalconcentrations. Cells were then incubated at 37 °C in a 5%CO2–95% air atmosphere for 72 h. Cell viability wasdetermined as above using a colorimetric assay with thereduction of MTT.
2.4. Quantitative fluorescent microscopy
Cells were harvested and fixed in 3% paraformaldehyde andincubated at room temperature for 10 min. The fixative wasremoved and the cells werewashedwith PBS, resuspended in 30–50 μl of PBS containing 20 μg/ml bis-benzimide trihydrochloride(Hoechst 33258), and incubated at room temperature for 15 min.Ten-microliter aliquots of the cells were placed on glass slides,and triplicate samples of 500 cells each were counted and scoredfor the incidence of apoptotic chromatin condensation using aZeiss fluorescent microscopy. Stained nuclei with condensedchromatin (supercondensed chromatin at the nuclear periphery),or nuclei that were fragmented into multiple smaller dense bodieswere considered as apoptotic. Nuclei with uncondensed anddispersed chromatin were considered as not apoptotic.
2.5. Quantification of apoptosis by flow cytometry
To study changes in the cell DNA content, histogrammeasurements of hypodiploid DNA formation was performedby flow cytometry using a Coulter EPICS™ cytometer (Beck-man Coulter). Histograms were analyzed with the Expo 32 ADCSoftware™. Cells were collected and centrifuged at 500 ×g,washed with PBS and resuspended in 50 μl of PBS. Followingdropwise addition of 1 ml of ice-cold 75% ethanol, fixed cellswere stored at −20 °C for 1 h. Samples were then centrifuged at500 ×g andwashedwith PBS before resuspension in 1ml of PBS
containing 50 μg/ml propidium iodide and 100 μg/ml RNase Aand incubation for 1 h at 37 °C in the dark. The percentage ofcells with decreased DNA staining, composed of apoptotic cellsresulting from either fragmentation or decreased chromatin, of aminimum of 10,000 cells per experimental condition wascounted. Cell debris was excluded from analysis by selectivegating based on anterior and right angle scattering.
2.6. Analysis of DNA fragmentation
HL-60 cells were washed with PBS and incubated in 20 μl of50 mM Tris-HCl (pH 8.0), 10 mM EDTA, 0.5% SDS and 1 μg/μl RNase A (Sigma) at 37 °C for 1 h. Then, 10 μg/μl proteinaseK (Sigma) (2 μl) was added, and the mixture was incubated at50 °C for 2 h more. DNAwas extracted with 100 μl of phenol-chloroform-isoamyl alcohol (24:24:1) and mixed with 5 μl ofloading solution (10 mM EDTA, pH 8.0, containing 1% (w/v)low melting-point agarose, 0.25% bromophenol blue and 40%sucrose). Samples were separated by electrophoresis in 2%agarose gels and visualized by UV illumination after ethidiumbromide staining.
2.7. Immunoblotting of caspase-3
HL-60 cells (1×106) were treated with flavonoids at theindicated concentrations in RPMI 1640 medium. Cell pelletswere lysed in lysis buffer containing 125 mM Tris-HCl pH 6.8,2% SDS, 5% glycerol, and 1% β-mercaptoethanol, and boiledfor 5 min. The samples were separated on 15% SDS-polyacrylamide gel, electrotransferred to a PVDF membrane,immunoblotted with anti-caspase-3 antibody [Stressgen; 1:1000dilution in Tris-buffered saline containing 0.1% Tween-20(TBST) supplemented with 3% nonfat milk] overnight. Afterwashing and incubation with horseradish peroxidase-conjugat-ed anti-rabbit (Amersham Pharmacia Biotech), the antigen–antibody complexes were visualized by enhanced chemilumi-nescence (ECL, Amersham Pharmacia Biotech) using themanufacturer's protocol.
2.8. Immunoblot analysis of poly(ADP-ribose) polymerasedegradation
Induction of apoptosis was also examined by proteolyticcleavage of poly(ADP-ribose) polymerase. After treatments, cellswere pelleted by centrifugation, and resuspended in lysis buffercontaining 25 mM PBS, 0.1 mM phenylmethylsufonylfluorideand protease inhibitors leupeptin, aprotinin and pepstatin A (5 μg/ml each). After centrifugation, the pellet was resuspended in theloading buffer containing 125 mM Tris-HCl, pH 6.8, 2% SDS,5% glycerol, and 1% β-mercaptoethanol. The mixture wassonicated for 30 s at 4 °C and then boiled to 100 °C for 3 min. Thecell lysates were fractionated on a 7.5% polyacrylamide gelcontaining 0.1% SDS and proteins were electrophoreticallytransferred onto a PVDFmembrane.Membranewas blockedwith5%nonfat milk in Tris-buffered saline containing 0.1%Tween-20for 1 h, followed by incubation with anti-poly(ADP-ribose)polymerase polyclonal antibody (Stressgen; 1:3000 dilution in

12 S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
TBST supplemented with 3% nonfat milk) overnight. Afterwashing and incubation with anti-rabbit antibody conjugated tohorseradish peroxidase (Amersham Pharmacia Biotech), theantigen–antibody complexes were visualized by enhancedchemiluminiscence (ECL, Amersham Pharmacia Biotech) usingthe manufacturer's protocol. The appearance of an 85 kDacleavage product was used as a measure of apoptosis.
2.9. Detection of cytochrome c release from mitochondria
Release of cytochrome c from mitochondria was detected byWestern blot analysis. After treatments, HL-60 cells werewashed twice with PBS and then suspended in ice-cold buffer[20 mM HEPES (pH 7.5), 1.5 mM MgCl2, 10 mM KCl, 1 mMEDTA, 1 mM EGTA, 1 mM dithiothreitol, 0.1 mM phenyl-methylsulfonylfluoride, and 5 μg/ml leupeptin, aprotinin, andpepstatin A] containing 250 mM sucrose. After 15 minincubation on ice, cells were lysed by pushing them severaltimes through a 22-gauge needle and the lysate spun down at1000 ×g for 5 min at 4 °C. The supernatant fraction wascentrifuged at 105,000×g for 45 min at 4 °C and the resultingsupernatant was used as the soluble cytosolic fraction. Cytosolicproteins (50 μg) were resolved on an SDS/15% polyacrylamidegel and electrotransferred onto a PVDF membrane. Themembrane was probed with monoclonal anti-cytochrome cantibody (BD Transduction Laboratories) (1:250 dilution) andthen with secondary antibody conjugated to horseradishperoxidase. Proteins bands were detected by chemiluminescence(ECL, Amersham Pharmacia Biotech) as described above.
2.10. Assay of caspase activity
After treatments, cells were harvested by centrifugation at1000 ×g for 5 min at 4 °C and washed with PBS, and the cellpellets were kept on ice. The cells were resuspended in cell lysisbuffer (50 mM HEPES, pH 7.4, 1 mM dithiothreitol, 0.1 mMEDTA, 0.1% Chaps) and held on ice for 5 min. Aftercentrifugation for 10 min at 17,000 ×g at 4 °C, the supernatantswere analyzed for protein concentration by the Bradford dye-binding assay and stored at −20 °C until used to study caspasecolorimetric enzymatic activity. Equal amounts of protein(∼20 μg) from different treatments were used, and the assayswere set up on ice. The net increase of absorbance at 405 nmafter incubation at 37 °C was indicative of enzyme activity.Specific labelled substrates for caspase-3, -8 and 9 activitieswere N-acetyl-Asp-Glu-Val-Asp-p-nitroaniline (DEVD-pNA), N-acetyl-Ile-Glu-Thr-Asp-p-nitroaniline (IETD-pNA)and N-acetyl-Leu-Glu-His-Asp-p-nitroaniline (LEHD-pNA)respectively.
3. Results
3.1. Betuletol derivatives inhibit the growth and cell viability ofhuman tumor cell lines
In the present study, we examined the effects of 22 flavonoids(Fig. 1), on the growth of five human tumor cell lines.
Antiproliferative studies on betuletol 3-methyl ether 1 indicatethat this compound displays strong cytotoxic properties in allcell lines assayed, although HL-60 cells was especially sensitiveto this substance with an IC50 as low as 1.8±0.9 μM (Table 1).Since betuletol 3-methyl ether 1 has two hydroxyl groups on C5and C7, respectively, we next obtained the diacetate 2- anddimethyl ether 3-derivatives of betuletol 3-methyl ether 1 inorder to know more about the structural requirement for theantiproliferative action of this compound. Therefore, thecytotoxic studies performed with the di-acetylated compoundindicate that theses changes have no impact on the IC50 in HL-60cells (1.8 ± 0.9 μM vs 1.3 ± 0.4 μM). However, in thedimethylated derivative a dramatic consequence on the growthinhibitory activity is observed. Contrary to betuletol 3-methylether 1 and betuletol 3-methyl ether diacetate 2, betuletol 3, 5, 7-trimethyl ether 3 is ineffective as an antiproliferative agent in allcell lines tested. Since the methoxy group is smaller than theacetyl group, it seems clear that this is not a consequence of sterichindrance. Moreover, if the hydroxyl groups in the betuletol 3-methyl ether 1 interact by hydrogen bridges with vicinity groupson the biological targets, then the presence of the keto groups onthe di-acetyl derivative could be important. One group thatseems to play a key role in determining the potency of thesecompounds on cell viability is the methyl group on position 4′ofthe B ring (2-phenyl group) since betuletol 3-methyl ether 1 issignificantly more potent than 4′,5,7-trihydroxy-3,6-dimethox-yflavone 12 in all cell lines studied.
The critical relationship of fruit and vegetable intake andcancer prevention has been thoroughly documented (Block et al.,1992). Since quercetin 4 is the most abundant flavonoid in thehuman diet we have investigated the effect of a series ofstructurally related flavonoids. The results indicate that methyl-ation of hydroxyl group at position C3 of quercetin 4 yieldsquercetin 3-methyl ether 5, a compound with a higherantiproliferative activity. Cytotoxic effects were observed in allcancer cell lines tested except in HOS cells, which indicates thatthe above chemical change in the quercetin molecule 4 affects cellproliferation in a cell-type specific manner. However, thehydroxyl group located at the 3′ position seems to have littleimportance for the growth inhibitory activity of the abovequercetin derivative since the additional methylation at this levelto yield quercetin 3,3′-dimethyl ether 7 does not significantlychange the IC50 value. The presence of a hydroxyl group in the 3′position of ring B has different consequences depending on theconfiguration of molecule. For example, kaempferol 3-methylether 9 lacks the 3′-hydroxyl group and displays similar IC50 to its3′-hydroxylated counterpart, quercetin 3-methyl ether 5 in all celllines assayed. Similarly, quercetin 3-methyl ether tetracetate 6 andquercetin 3,3′-dimethyl ether triacetate 8 differ only by thepresence of an acetyl- ormethylether-group, respectively, at the 3′position, but they showed similar potency. On the other hand, thepresence of the 3′ hydroxyl group improves the antiproliferativeactivity of axillarin 11 as compared with the 3′-dehydroxylatedcounterpart 12 in all cell types tested except HL-60 cells. When amethoxy group is placed at C6 instead of hydrogen in quercetin 3-methyl ether 5, the impact on the IC50 is dependent on the cell typeunder study. The resulting compound (axillarin 11) is significantly

Fig. 1. Structures of the investigated flavonoids.
13S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
less cytotoxic on HL-60 (IC50=14.3±4.6 μM versus 32.6±8.8 μM) but significantly more cytotoxic on HOS cells(IC50=90.4±27.6 μM versus 25.6±9.6 μM). In contrast, thepresence of this methyl ether group does not change its cytotoxicpotency on A431, HeLa and SK-OV-3 cells.
The influence of the hydroxyl group at position 4′ of the B ring(2-phenyl group) on cell viability was also tested. A dramaticdecrease in the growth inhibitory activity was observed whenquercetin 3,3′-dimethyl ether 7was methylated in that position toproduce 5,7-dihydroxy-3,3′,4′-trimethoxyflavone 10. The IC50
increased from 12.2±0.4 μM and 30.6±7.3 μM to N100 μM, inHL-60 and HeLa cells respectively. As occurs with otherssubstitutions, this chemical change affected the cell lines understudy to different degrees. The HOS cell line was resistant to bothquercetin 3,3′-dimethyl ether 7 and 5,7-dihydroxy-3,3′,4′-trimethoxyflavone 10. Thus, acetylation of all free hydroxylgroups locating at positions 3′, 4′, 5 and 7 in the quercetin 3-methyl ether 5 molecule to produce quercetin 3-methyl ethertetracetate 6 greatly improved the cytotoxic activity onHOS cells.In this cell line the IC50 decreased about one order of magnitude
(90.4±27.6 μM versus 9.4±3.0). Unlike the HOS cells, theremaining cell lines were resistant to these chemical changes.Also, an increase in the growth inhibitory activity was clearlyobserved when quercetin 3,3′-dimethyl ether 7 was acetylated atpositions 4′, 5 and 7 to produce quercetin 3,3′-dimethyl ethertriacetate 8. HL-60 cells, A431 cells and particularly HOS cellswere susceptible to these chemicals substitutions.
Selected acetylations were also performed on the freehydroxyl groups of the 4′, 5, 7-trihydroxy-3, 6-dimethoxy-flavone molecule 12. Acetylation of the hydroxyl group atposition 7 yielded a monoacetylated derivative, 7-acetoxy-4′,5-dihydroxy-3,6-dimethoxyflavone 13, that was significantlymore active on HL-60 cells. This effect is abolished when asecond and a third acetyl group are sequentially introduced atposition 5 and at position 4′ to produce the derivativecompounds 5,7-diacetoxy-4′-hydroxy-3,6-dimethoxyflavone14 and 4′,5,7-triacetoxy-3,6-dimethoxyflavone 15 respectively.
Another set of flavonoids are those related to the structure ofthe low cytotoxic compound naringenin 21. This flavanonedecreased the proliferation rate of the cell lines studied only at

Fig. 2. Effects of betuletol 3-methyl ether and betuletol 3-methyl ether diacetateon apoptosis on HL-60 cells. (A) Cells were stained with bisbenzimidetrihydrochloride after treatment with 10 μM of betuletol 3-methyl ether for 12 h,and apoptotic cells were determined by fluorescence microscopy. Valuesrepresent means±S.E. This histogram is representative of two independentexperiments each performed in duplicate. ⁎Pb0.05, significantly different fromthe untreated control. (B) Photomicrographs of representative fields of HL-60cells treated as above and stained with bisbenzimide trihydrochloride to evaluatenuclear chromatin condensation (i.e. apoptosis). (C) Analysis for the cleavage ofpoly(ADP-ribose) polymerase (PARP) by immunoblotting. (D) Cells wereincubated as above and DNA extracted. Laddered electrophoretic patterns ofoligonucleosomal DNA fragments were resolved by conventional agarose gelelectrophoresis, stained with ethidium bromide and visualized under UV light.Control lane refers to untreated cells.
Table 1Effects of phenylbenzo-γ-pirone derivatives on the growth of human tumor celllines
Compound IC50 (μM)
HL-60 A431 HeLa SK-OV-3 HOS
1 1.8±0.9 3.0±1.0 9.1±4.0 21.0±7.0 6.8±0.62 1.3±0.4 8.0±2.9 8.1±4.7 30.0±3.0 7.5±0.83 N100 N100 N100 N100 N1004 27.8±4.1 71.1±7.1 N100 N100 93.3±1.15 14.3±4.6 26.8±7.9 35.0±11.7 44.6±11.9 90.4±27.66 9.2±3.4 19.3±8.2 24.7±3.6 43.6±15.3 9.4±3.07 12.2±0.4 33.9±6.0 30.6±7.3 30.3±12.3 N1008 3.8±0.2 10.0±5.2 29.3±5.7 29.6±6.6 18.3±3.39 7.2±2.9 40.6±7.9 32.8±11.3 30.0±11.3 77.5±4.510 N100 54.2±3.7 N100 68.0±41.0 N10011 32.6±8.8 33.0±9.2 47.7±8.8 29.5±11.5 25.6±9.612 27.5±1.6 59.6±3.6 N100 88.2±1.8 60.5±14.613 13.4±3.1 58.6±2.5 N100 N100 60.0±1.014 33.5±0.3 55.9±1.0 N100 N100 80.3±28.015 20.6±3.1 56.1±7.6 N100 69.8±5.2 49.8±25.016 N100 90.3±0.1 N100 N100 75.6±13.917 N100 N100 N100 N100 N10018 73.5±10.1 71.2±10.1 N100 N100 91.7±12.319 35.0±3.0 28.8±5.8 N100 N100 N10020 4.5 ±1.3 10.8±1.3 20.5±6.1 22.5±2.9 N10021 N100 N100 N100 N100 N10022 13.1±1.0 76.3±18.9 32.7±12.4 71.3±6.1 82.6±15.3
Cells were cultured for 72 h and the IC50 values were calculated as described inthe Experimental Section. The data shown represent the mean±S.E.M. of 3–5independent experiments with three determinations in each.
14 S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
very high concentrations (IC50N100 μM) but the introductionof a hydroxyl group at position 3′ (eriodictyol 19) greatlyincreases its antiproliferative activity although only on HL-60and A431 cells. The 3′, 4′, 5 and 7 tetracetylated derivative,eriodictyol tetracetate 20, is even more active with IC50 valuesas low as 4.5±1.3 μM and 10.8±1.3 μM on HL-60 and A431,respectively. A moderate increase in the cytotoxicity ofnaringenin 21 is also observed on HL-60 and A431 cellswhen the hydroxyl group placed at the 4′ position on the B ring(2-phenyl group) is absent and the molecule has a methyl groupon C8 (cryptostrobin 18). Another structural requirement toimprove the cytotoxic activity of naringenin 21 is theintroduction of a double bond between C2 and C3. Theresulting flavone (apigenin 22) showed antiproliferative activityin all cell lines tested although HL-60 cell line was the mostsusceptible to this chemical change (IC50=13.1±1.0 μM).
A detailed inspection of Table 1 reveals that among the fivetumor cell lines used in this study the growth of promyelocyticHL-60 cells were, in general, most susceptible to the cytotoxicityinduced by flavonoids. Interestingly, quercetin 4, a flavonoidwidely studied due to abundance in the fruit and vegetableusually consumed by humans, displayed significantly cytotoxicproperties for HL-60 cells alone (IC50=27.8±4.1 μM). Theremaining lines were insensitive to quercetin 4 effects on cellproliferation.
Since HL-60 cell line was highly sensitive to the anti-proliferative effect of betuletol 3-methyl ether 1 we decided todetermine whether this natural flavonoid displays its cytotoxicaction through activation of the apoptotic pathway.
3.2. Betuletol derivatives induce apoptosis on human myeloidleukemia cells
In order to evaluate whether betuletol derivatives decrease cellviability through apoptosis activation, morphological changescharacteristic of apoptotic cells (condensed and fragmentedchromatin) were analyzed and quantified by fluorescent micros-copy (Fig. 2A and B). Evaluation of nuclear morphology (Fig.2B), indicates that the percentage of apoptotic cells increasedfrom 6±1% to 24±2% (4 times increase) in betuletol 3-methylether treated cells, after 12 h exposure at a concentration as low as10 μM (Fig. 2A). Next, we examined whether these flavonoidsinduce poly(ADP-ribose) polymerase cleavage, a hallmark ofapoptosis that indicates activation of caspase. Hydrolysis of the116 kDa poly(ADP-ribose) polymerase protein to the 85 kDafragment was detected in betuletol 3-methyl ether and betuletol 3-methyl ether diacetate treated cells after 12 h exposure at 10 μM(Fig. 2C), and increased in a dose-dependent manner (data notshown). These results indicate that poly(ADP-ribose) polymerasecleavage was involved in apoptosis induced by these compoundsand, as expected (due to cytotoxic ineffectiveness), betuletol3,5,7-trimethyl ether did not induce poly(ADP-ribose) polymer-ase cleavage (Fig. 2C). When cells were incubated with these

15S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
flavonoids, the DNA showed the typical fragmentation patternsformed by internucleosomal hydrolysis of chromatin thusconfirming the apoptosis-inducing effects. As expected, betuletol3,5,7-trimethyl ether was unable to induce DNA fragmentation(Fig. 2D). Taken together, these results indicate that both betuletolderivatives, betuletol 3-methyl ether and betuletol 3-methyl etherdiacetate, are potent inducers of human myeloid leukemia HL-60cells apoptosis and that both compounds are equally active.
3.3. A general caspase inhibitor abolished betuletolderivatives-induced poly(ADP-ribose) polymerase cleavageand DNA fragmentation
To confirm that betuletol derivatives-triggered apoptosisrequires the activation of caspases, HL-60 cells were pretreatedwith the broad-spectrum caspase inhibitor z-VAD-fmk (benzy-loxycarbonyl-Val-Ala-Asp(OMe) fluoromethyl ketone). Theresults demonstrate that this inhibitor completely blockedbetuletol derivatives-induced DNA fragmentation (Fig. 3A).The effect of z-VAD-fmk on poly(ADP-ribose) polymerasecleavage by betuletol 3-methyl ether was also analyzed. Asobserved (Fig. 3B), pretreatment of HL-60 cells with thisinhibitor completely abolished the generation of the 85 kDafragment. Taken together, these results indicate that these eventsare caspase dependent.
Fig. 3. Involvement of activation of caspases in the induction of apoptosis. HL-60 cells were incubated with the indicated doses of compounds for 12 h inabsence or presence of broad-spectrum caspase inhibitor z-VAD-fmk (100 μM).DNA fragmentation (A) and proteolysis of poly(ADP-ribose) polymerase (B)were then analyzed. The migration positions of full-length poly(ADP-ribose)polymerase and the 85 kDa cleavage product are indicated. The results arerepresentative of three independent experiments.
3.4. Betuletol derivatives-induced apoptosis involves caspase-3activation
To determine whether caspase-3 is activated in HL-60 cells bythe betuletol 3-methyl ether, dose-response experiments wereperformed and cell lysates assayed for cleavage of the tetrapeptidesubstrate N-acetyl-Asp-Glu-Val-Asp-p-nitroaniline (DEVD-pNA). The results (Fig. 4A), demonstrate that caspase-3 activityincreased 1.5- and 2.3-fold in betuletol 3-methyl ether treatedcells with 3- or 10-μM of compound, respectively, for 12 h.Hydrolysis of procaspase-3 to the active enzyme, caspase-3, bybetuletol 3-methyl ether and betuletol 3-methyl ether diacetatewas also determined by immunobloting. The results indicate thatthese compounds at concentrations as low as 3 μM promotecleavage of pro-caspase-3 into active caspase-3 (Fig. 4B). At highconcentrations (100 μM) of a specific caspase-3 inhibitor,benzyloxycarbonyl-Asp-Glu-Val-Asp-(OMe) fluoromethyl ke-tone (z-DEVD-fmk), the percentage of apoptotic cells and DNAladdering induced by betuletol 3-methyl ether was significantlydecreased. In contrast, there was not any effect when theconcentration of the inhibitor used was lowered to a minimumof 10 μM (results not shown).
3.5. Effects of betuletol derivatives on mitochondrial cytochromec release
To determine whether betuletol 3-methyl ether and betuletol3-methyl ether diacetate-induced apoptosis on HL-60 cellsinvolves the release of cytochrome c from mitochondria tocytosol, dose-response experiments were performed and cyto-solic preparations were analyzed by immunoblotting. Asdemonstrated (Fig. 5A), a low concentration of both compounds(3 μM) induce cytochrome c release at the same proportion andno difference was observed with higher concentrations ofcompounds. The effect of these compounds on caspase-9formation was also analyzed. The results indicate that bothbetuletol derivatives stimulate cleavage of inactive pro-caspase-9 to the active 37 kDa fragment (Fig. 5B). To determine whetherthis fragment was associated with caspase-9 activity, lysatesfrom treated cells were assayed for cleavage of the specificsubstrate N-acetyl-Leu-Glu-His-Asp-p-nitroaniline (LEHD-pNA). The results indicated that compound 1 (10 μM, 12 h)also increases caspase-9 activity (Fig. 5C). In order to determinethe contribution of caspase-9 to the betuletol derivatives-inducedapoptosis we examined the impact of the irreversible caspase-9inhibitor benzyloxycarbonyl-Leu-Glu-His-Asp(OMe)-fluoro-methyl ketone (z-LEHD-fmk). As shown (Fig. 5D) the inhibitorwas completely ineffective at blocking betuletol 3-methyl ether-induced DNA nucleosomal fragmentation typical of apoptoticcell death.
3.6. Caspase-8 is activated by betuletol derivatives
Since the intrinsic pathway does not seem to contributesignificantly to betuletol derivatives-induced apoptosis, wedecided to determine the contribution of the extrinsic pathway.Therefore, HL-60 cells were treated with increasing doses of
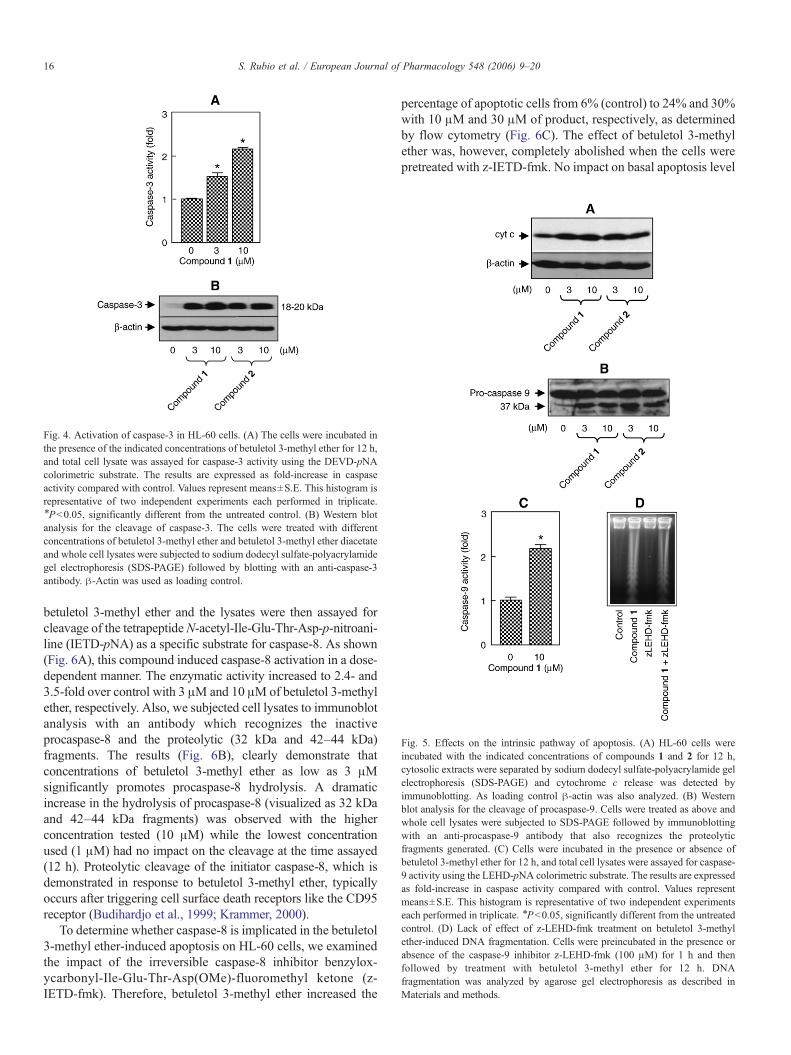
Fig. 5. Effects on the intrinsic pathway of apoptosis. (A) HL-60 cells wereincubated with the indicated concentrations of compounds 1 and 2 for 12 h,cytosolic extracts were separated by sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) and cytochrome c release was detected byimmunoblotting. As loading control β-actin was also analyzed. (B) Westernblot analysis for the cleavage of procaspase-9. Cells were treated as above andwhole cell lysates were subjected to SDS-PAGE followed by immunoblottingwith an anti-procaspase-9 antibody that also recognizes the proteolyticfragments generated. (C) Cells were incubated in the presence or absence ofbetuletol 3-methyl ether for 12 h, and total cell lysates were assayed for caspase-9 activity using the LEHD-pNA colorimetric substrate. The results are expressedas fold-increase in caspase activity compared with control. Values representmeans±S.E. This histogram is representative of two independent experimentseach performed in triplicate. ⁎Pb0.05, significantly different from the untreatedcontrol. (D) Lack of effect of z-LEHD-fmk treatment on betuletol 3-methylether-induced DNA fragmentation. Cells were preincubated in the presence orabsence of the caspase-9 inhibitor z-LEHD-fmk (100 μM) for 1 h and thenfollowed by treatment with betuletol 3-methyl ether for 12 h. DNAfragmentation was analyzed by agarose gel electrophoresis as described inMaterials and methods.
Fig. 4. Activation of caspase-3 in HL-60 cells. (A) The cells were incubated inthe presence of the indicated concentrations of betuletol 3-methyl ether for 12 h,and total cell lysate was assayed for caspase-3 activity using the DEVD-pNAcolorimetric substrate. The results are expressed as fold-increase in caspaseactivity compared with control. Values represent means±S.E. This histogram isrepresentative of two independent experiments each performed in triplicate.⁎Pb0.05, significantly different from the untreated control. (B) Western blotanalysis for the cleavage of caspase-3. The cells were treated with differentconcentrations of betuletol 3-methyl ether and betuletol 3-methyl ether diacetateand whole cell lysates were subjected to sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) followed by blotting with an anti-caspase-3antibody. β-Actin was used as loading control.
16 S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
betuletol 3-methyl ether and the lysates were then assayed forcleavage of the tetrapeptideN-acetyl-Ile-Glu-Thr-Asp-p-nitroani-line (IETD-pNA) as a specific substrate for caspase-8. As shown(Fig. 6A), this compound induced caspase-8 activation in a dose-dependent manner. The enzymatic activity increased to 2.4- and3.5-fold over control with 3 μM and 10 μM of betuletol 3-methylether, respectively. Also, we subjected cell lysates to immunoblotanalysis with an antibody which recognizes the inactiveprocaspase-8 and the proteolytic (32 kDa and 42–44 kDa)fragments. The results (Fig. 6B), clearly demonstrate thatconcentrations of betuletol 3-methyl ether as low as 3 μMsignificantly promotes procaspase-8 hydrolysis. A dramaticincrease in the hydrolysis of procaspase-8 (visualized as 32 kDaand 42–44 kDa fragments) was observed with the higherconcentration tested (10 μM) while the lowest concentrationused (1 μM) had no impact on the cleavage at the time assayed(12 h). Proteolytic cleavage of the initiator caspase-8, which isdemonstrated in response to betuletol 3-methyl ether, typicallyoccurs after triggering cell surface death receptors like the CD95receptor (Budihardjo et al., 1999; Krammer, 2000).
To determine whether caspase-8 is implicated in the betuletol3-methyl ether-induced apoptosis on HL-60 cells, we examinedthe impact of the irreversible caspase-8 inhibitor benzylox-ycarbonyl-Ile-Glu-Thr-Asp(OMe)-fluoromethyl ketone (z-IETD-fmk). Therefore, betuletol 3-methyl ether increased the
percentage of apoptotic cells from 6% (control) to 24% and 30%with 10 μM and 30 μM of product, respectively, as determinedby flow cytometry (Fig. 6C). The effect of betuletol 3-methylether was, however, completely abolished when the cells werepretreated with z-IETD-fmk. No impact on basal apoptosis level
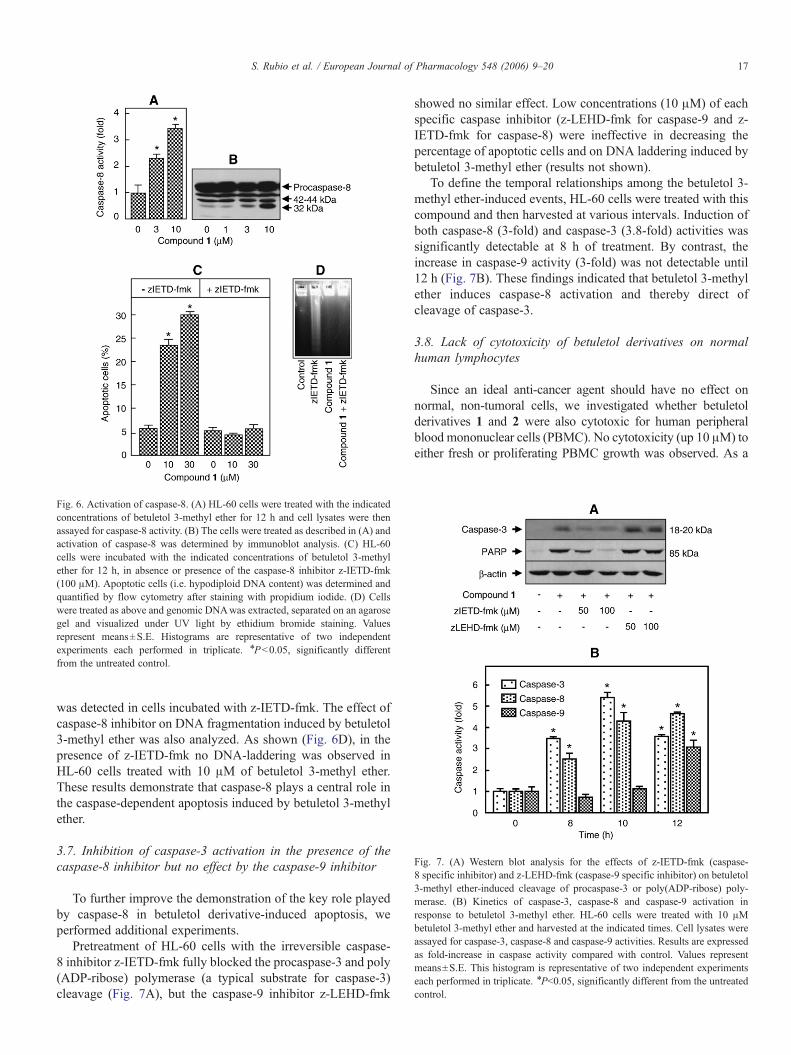
Fig. 7. (A) Western blot analysis for the effects of z-IETD-fmk (caspase-8 specific inhibitor) and z-LEHD-fmk (caspase-9 specific inhibitor) on betuletol3-methyl ether-induced cleavage of procaspase-3 or poly(ADP-ribose) poly-merase. (B) Kinetics of caspase-3, caspase-8 and caspase-9 activation inresponse to betuletol 3-methyl ether. HL-60 cells were treated with 10 μMbetuletol 3-methyl ether and harvested at the indicated times. Cell lysates wereassayed for caspase-3, caspase-8 and caspase-9 activities. Results are expressedas fold-increase in caspase activity compared with control. Values representmeans±S.E. This histogram is representative of two independent experimentseach performed in triplicate. ⁎Pb0.05, significantly different from the untreatedcontrol.
Fig. 6. Activation of caspase-8. (A) HL-60 cells were treated with the indicatedconcentrations of betuletol 3-methyl ether for 12 h and cell lysates were thenassayed for caspase-8 activity. (B) The cells were treated as described in (A) andactivation of caspase-8 was determined by immunoblot analysis. (C) HL-60cells were incubated with the indicated concentrations of betuletol 3-methylether for 12 h, in absence or presence of the caspase-8 inhibitor z-IETD-fmk(100 μM). Apoptotic cells (i.e. hypodiploid DNA content) was determined andquantified by flow cytometry after staining with propidium iodide. (D) Cellswere treated as above and genomic DNAwas extracted, separated on an agarosegel and visualized under UV light by ethidium bromide staining. Valuesrepresent means±S.E. Histograms are representative of two independentexperiments each performed in triplicate. ⁎Pb0.05, significantly differentfrom the untreated control.
17S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
was detected in cells incubated with z-IETD-fmk. The effect ofcaspase-8 inhibitor on DNA fragmentation induced by betuletol3-methyl ether was also analyzed. As shown (Fig. 6D), in thepresence of z-IETD-fmk no DNA-laddering was observed inHL-60 cells treated with 10 μM of betuletol 3-methyl ether.These results demonstrate that caspase-8 plays a central role inthe caspase-dependent apoptosis induced by betuletol 3-methylether.
3.7. Inhibition of caspase-3 activation in the presence of thecaspase-8 inhibitor but no effect by the caspase-9 inhibitor
To further improve the demonstration of the key role playedby caspase-8 in betuletol derivative-induced apoptosis, weperformed additional experiments.
Pretreatment of HL-60 cells with the irreversible caspase-8 inhibitor z-IETD-fmk fully blocked the procaspase-3 and poly(ADP-ribose) polymerase (a typical substrate for caspase-3)cleavage (Fig. 7A), but the caspase-9 inhibitor z-LEHD-fmk
showed no similar effect. Low concentrations (10 μM) of eachspecific caspase inhibitor (z-LEHD-fmk for caspase-9 and z-IETD-fmk for caspase-8) were ineffective in decreasing thepercentage of apoptotic cells and on DNA laddering induced bybetuletol 3-methyl ether (results not shown).
To define the temporal relationships among the betuletol 3-methyl ether-induced events, HL-60 cells were treated with thiscompound and then harvested at various intervals. Induction ofboth caspase-8 (3-fold) and caspase-3 (3.8-fold) activities wassignificantly detectable at 8 h of treatment. By contrast, theincrease in caspase-9 activity (3-fold) was not detectable until12 h (Fig. 7B). These findings indicated that betuletol 3-methylether induces caspase-8 activation and thereby direct ofcleavage of caspase-3.
3.8. Lack of cytotoxicity of betuletol derivatives on normalhuman lymphocytes
Since an ideal anti-cancer agent should have no effect onnormal, non-tumoral cells, we investigated whether betuletolderivatives 1 and 2 were also cytotoxic for human peripheralblood mononuclear cells (PBMC). No cytotoxicity (up 10 μM) toeither fresh or proliferating PBMC growth was observed. As a
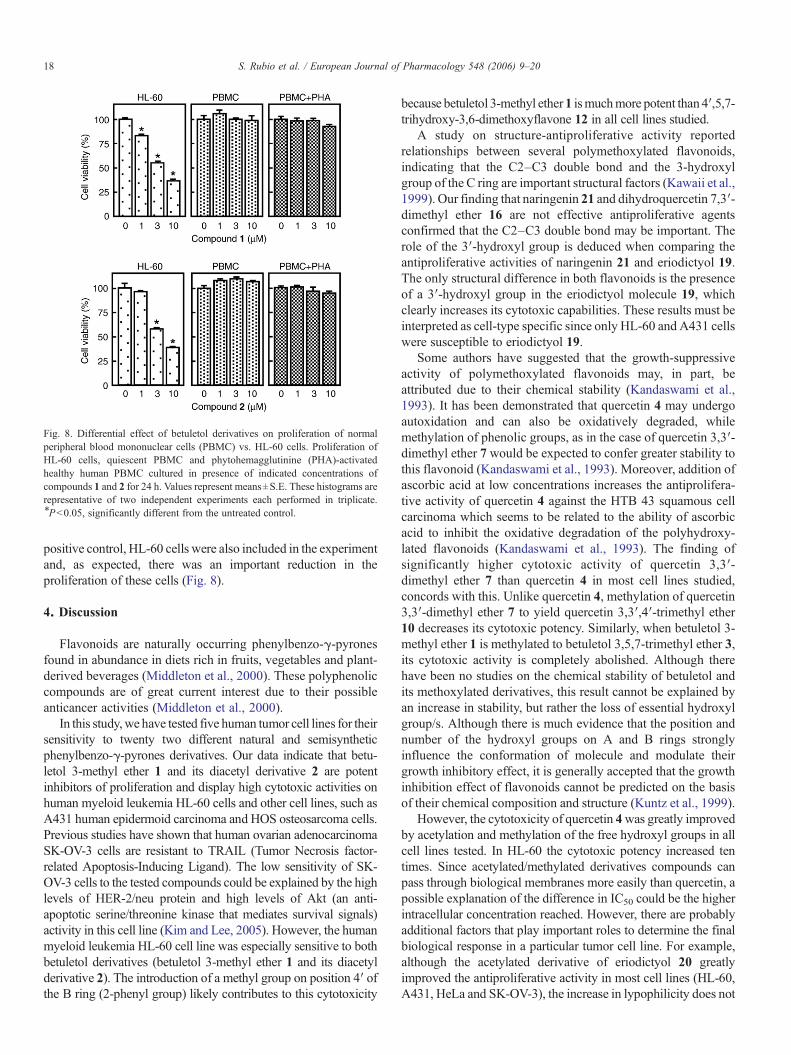
Fig. 8. Differential effect of betuletol derivatives on proliferation of normalperipheral blood mononuclear cells (PBMC) vs. HL-60 cells. Proliferation ofHL-60 cells, quiescent PBMC and phytohemagglutinine (PHA)-activatedhealthy human PBMC cultured in presence of indicated concentrations ofcompounds 1 and 2 for 24 h. Values represent means±S.E. These histograms arerepresentative of two independent experiments each performed in triplicate.⁎Pb0.05, significantly different from the untreated control.
18 S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
positive control, HL-60 cells were also included in the experimentand, as expected, there was an important reduction in theproliferation of these cells (Fig. 8).
4. Discussion
Flavonoids are naturally occurring phenylbenzo-γ-pyronesfound in abundance in diets rich in fruits, vegetables and plant-derived beverages (Middleton et al., 2000). These polyphenoliccompounds are of great current interest due to their possibleanticancer activities (Middleton et al., 2000).
In this study,we have tested five human tumor cell lines for theirsensitivity to twenty two different natural and semisyntheticphenylbenzo-γ-pyrones derivatives. Our data indicate that betu-letol 3-methyl ether 1 and its diacetyl derivative 2 are potentinhibitors of proliferation and display high cytotoxic activities onhuman myeloid leukemia HL-60 cells and other cell lines, such asA431 human epidermoid carcinoma and HOS osteosarcoma cells.Previous studies have shown that human ovarian adenocarcinomaSK-OV-3 cells are resistant to TRAIL (Tumor Necrosis factor-related Apoptosis-Inducing Ligand). The low sensitivity of SK-OV-3 cells to the tested compounds could be explained by the highlevels of HER-2/neu protein and high levels of Akt (an anti-apoptotic serine/threonine kinase that mediates survival signals)activity in this cell line (Kim and Lee, 2005). However, the humanmyeloid leukemia HL-60 cell line was especially sensitive to bothbetuletol derivatives (betuletol 3-methyl ether 1 and its diacetylderivative 2). The introduction of a methyl group on position 4′ ofthe B ring (2-phenyl group) likely contributes to this cytotoxicity
because betuletol 3-methyl ether1 ismuchmore potent than 4′,5,7-trihydroxy-3,6-dimethoxyflavone 12 in all cell lines studied.
A study on structure-antiproliferative activity reportedrelationships between several polymethoxylated flavonoids,indicating that the C2–C3 double bond and the 3-hydroxylgroup of the C ring are important structural factors (Kawaii et al.,1999). Our finding that naringenin 21 and dihydroquercetin 7,3′-dimethyl ether 16 are not effective antiproliferative agentsconfirmed that the C2–C3 double bond may be important. Therole of the 3′-hydroxyl group is deduced when comparing theantiproliferative activities of naringenin 21 and eriodictyol 19.The only structural difference in both flavonoids is the presenceof a 3′-hydroxyl group in the eriodictyol molecule 19, whichclearly increases its cytotoxic capabilities. These results must beinterpreted as cell-type specific since only HL-60 and A431 cellswere susceptible to eriodictyol 19.
Some authors have suggested that the growth-suppressiveactivity of polymethoxylated flavonoids may, in part, beattributed due to their chemical stability (Kandaswami et al.,1993). It has been demonstrated that quercetin 4 may undergoautoxidation and can also be oxidatively degraded, whilemethylation of phenolic groups, as in the case of quercetin 3,3′-dimethyl ether 7 would be expected to confer greater stability tothis flavonoid (Kandaswami et al., 1993). Moreover, addition ofascorbic acid at low concentrations increases the antiprolifera-tive activity of quercetin 4 against the HTB 43 squamous cellcarcinoma which seems to be related to the ability of ascorbicacid to inhibit the oxidative degradation of the polyhydroxy-lated flavonoids (Kandaswami et al., 1993). The finding ofsignificantly higher cytotoxic activity of quercetin 3,3′-dimethyl ether 7 than quercetin 4 in most cell lines studied,concords with this. Unlike quercetin 4, methylation of quercetin3,3′-dimethyl ether 7 to yield quercetin 3,3′,4′-trimethyl ether10 decreases its cytotoxic potency. Similarly, when betuletol 3-methyl ether 1 is methylated to betuletol 3,5,7-trimethyl ether 3,its cytotoxic activity is completely abolished. Although therehave been no studies on the chemical stability of betuletol andits methoxylated derivatives, this result cannot be explained byan increase in stability, but rather the loss of essential hydroxylgroup/s. Although there is much evidence that the position andnumber of the hydroxyl groups on A and B rings stronglyinfluence the conformation of molecule and modulate theirgrowth inhibitory effect, it is generally accepted that the growthinhibition effect of flavonoids cannot be predicted on the basisof their chemical composition and structure (Kuntz et al., 1999).
However, the cytotoxicity of quercetin 4was greatly improvedby acetylation and methylation of the free hydroxyl groups in allcell lines tested. In HL-60 the cytotoxic potency increased tentimes. Since acetylated/methylated derivatives compounds canpass through biological membranes more easily than quercetin, apossible explanation of the difference in IC50 could be the higherintracellular concentration reached. However, there are probablyadditional factors that play important roles to determine the finalbiological response in a particular tumor cell line. For example,although the acetylated derivative of eriodictyol 20 greatlyimproved the antiproliferative activity in most cell lines (HL-60,A431, HeLa and SK-OV-3), the increase in lypophilicity does not

19S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
explain its ineffectiveness in HOS cells. Also, among the 22compounds presented in Table 1, betuletol 3-methyl ether 1 andits diacetate-derivative 2 exhibited the highest cytotoxic activitiesagainst all human cancer cell lines evaluated. Inmost cell lines theIC50 values for these betuletol derivatives were lower than 10μM,although in HL-60 cells these values were about 1 μM.
As for all of the agents used or developed for cancertreatment, selectivity toward cancer cells is an importantcriterion. We therefore compared the effects of compounds 1and 2 between HL-60 cells and human peripheral bloodmononuclear cells (PBMC). Interestingly, dose-response stud-ies revealed that quiescent PBMC and proliferating PBMCwereresistant toward both betuletol derivatives 1 and 2.
In this work, we demonstrate that the exposure of HL-60cells to betuletol 3-methyl ether and its diacetyl derivativeexerts a strong antiproliferative effect and both compoundsdisplayed similar IC50 values. Among the number of pathwaysthat can be exploited for cancer prevention and treatment,apoptosis is a physiological process by which cells are removedwhen an agent damages their DNA, therefore compounds thatcan induce apoptosis may be useful in management and therapyof cancer. Although previous studies have documented theinduction of apoptotic cell death and DNA fragmentation byphenylbenzo-γ-pirones derivatives in the human myeloidleukemia HL-60 cells (Lee et al., 2002) betuletol derivativeshave yet to be assessed. Our results show that betuletol 3-methylether and its diacetyl derivative induced apoptosis in thehematopoietic cell line HL-60.
Both betuletol derivatives promote the formation of apoptoticbodies and the internucleosomal degradation of DNA, resulting inthe formation and eventual release of oligonucleosomal DNAfragments. The apoptosis induction was accompanied by poly(ADP-ribose) polymerase cleavage which is considered to be oneof the hallmarks of apoptosis (Nicholson and Thornberry, 1997).
We report in the present study that z-VAD-fmk, a broad-spectrum caspase inhibitor, completely abrogated the morpho-logical changes associated with apoptosis, DNA fragmentationand the cleavage of poly(ADP-ribose) polymerase, whichindicated that caspases are essential components in betuletol3-methyl ether-induced apoptosis.
Caspase-3 is the most active effector caspase, in both theintrinsic and extrinsic pathways where it is processed andactivated by caspase-9 and caspase-8 respectively. Caspase-3has been found to be involved in leukemia-cell apoptosis inducedby cytotoxic agents such as 1-β-D-arabinofuranosylcytosine,mitoxantrone, etoposide and CPT-11 (Motwani et al., 1999). Ahigh level of caspase-3 activation and processing was found aftertreatment of HL-60 cells with betuletol derivatives 1 and 2.
Previous studies have already demonstrated that most ofcytotoxic agents induce the release of mitochondrial cyto-chrome c (Szewczyk and Wojtczak, 2002), and this proteintriggers a caspase-dependent assembly of the apoptosome (VanGurp et al., 2003). Although we demonstrate that betuletolderivatives initiated redistribution of cytochrome c into thecytosol and procaspase-9 processing, our results indicate thatthe role of caspase-9 in the betuletol induced-apoptosis overallis, at most, minimal. Moreover, we analyzed the kinetic patterns
of cytochrome c release in time course experiments afterbetuletol derivatives treatments. The cytochrome c release wasnot an early event in the apoptotic cell death induced by bothbetuletol derivatives (results not shown). Although the exposureof HL-60 cells to both betuletol derivatives resulted in increasedcytosolic cytochrome c, this response was not detected untilafter 12 h of treatment. Consistently with this finding, theactivation of caspase-9 (as determined by a colorimetric assayand western blot), was not observed until the same time period.Furthermore, there have been ample examples of cross-talkbetween cell death receptor-mediated signaling through thecaspase-8 and the apoptosome cascade. Caspase-8 directlyactivates downstream effector caspases and also cleaves Bid andtriggers mitochondrial damage that in turn leads to cytochromec release. Studies have demonstrated that the intrinsic pathwayfor apoptosis initiation is controlled by members of the Bcl-2family (Marsden and Strasser, 2003; Borner, 2003), which arecrucial regulators of apoptosis in mammalian cells. However,we were unable to demonstrate alterations in the expression orin the mobility of Bcl-2 on sodium dodecyl sulfate-polyacryl-amide gel electrophoresis, a phenomenon that generally(although not invariably) accompanies perturbations in phos-phorylation state (Haldar et al., 1995) (data not shown).
Previous studies have shown that apoptosis caused by anti-cancer drugsmay bemediated via theCD95 system (Friesen et al.,1996), although many drugs seem to initiate the mitochondrialpathway directly. An ample variety of mechanisms of resistanceto apoptosis that interfere at different levels of apoptosissignalling have been described (Igney and Krammer, 2002).Thus a search for novel agents that target aberrant apoptosispathways or induce alternative death pathways may be suited toovercome treatment resistance.
Using irreversible inhibitors for selected caspases, weattempted to further delineate the pathways of caspase activationand their relationship to poly(ADP-ribose) polymerase cleavageand DNA fragmentation induced by betuletol derivatives. Theeffective blockage of apoptosis induction by a cell-permeableinhibitor of caspase-8, z-IETD-fmk, but not by a cell-permeableinhibitor of caspase-9, z-LEHD-fmk (also confirmed by kineticstudies of caspases activation) supports a model in whichbetuletol 3-methyl ether induces apoptosis by a caspase-8 mediated mechanism.
5. Conclusion
Our results indicate that betuletol 3-methyl ether inhibitshuman HL-60 cell growth and induces apoptotic cell deathinvolving the extrinsic pathway and could be useful in thedevelopment of novel anticancer agents.
Acknowledgments
This work was supported by Grants from the Ministerio deEducación y Ciencia of Spain and FEDER (SAF2004-07928),from Consejería de Educación, Cultura y Deportes (CanaryIslands Government) and FEDER (GRUP2004/44) and Insti-tuto Canario de Investigación del Cáncer (08/2004). We thank J.

20 S. Rubio et al. / European Journal of Pharmacology 548 (2006) 9–20
Estévez (Hospital Universitario Insular de Gran Canaria) for hiscollaboration in the Western blot assays, J.C. Hernández fortechnical assistance as well as the encouragement and support ofLennart Loven and Dr. Jaime Bermejo.
References
Block, G., Patterson, B., Subar, A., 1992. Fruit, vegetables and cancerprevention: a review of the epidemiological evidence. Nutr. Cancer 18,1–29.
Boldin, M., Goncharov, T., Goltsev, Y., Wallach, D., 1996. Involvement ofMACH, a novel MORT1/FADD-interacting protease, in Fas/APO-1- andTNF receptor-induced cell death. Cell 85, 803–815.
Borner, C., 2003. The Bcl-2 protein family: sensors and checkpoints for life-or-death decisions. Mol. Immunol. 39, 615–647.
Budihardjo, I., Oliver, H., Lutter, M., Luo, X., Wang, X., 1999. Biochemicalpathways of caspase activation during apoptosis. Annu. Rev. Cell Dev. Biol.15, 269–290.
Byrne, L.T., Cannon, J.R., Gawad, D.H., Joshi, B.S., Skelton, B.W., Toia, R.F.,White, A.H., 1982. The crystal structure of (S)-(−)-6-Bromo-5,7-dihydroxy-8-methyl-2-phenyl-2,3-dihydro-4H-1-benzopyran-4-one [(−)-6-bromocryp-tostrobin] and a 13C N.M.R. Study of (±)-cryptostrobin and relatedsubstances. Revision of the structures of the natural products (±)-lawinal,unonal, 7-O-methylunonal and isounonal. Aust. J. Chem. 35, 1851–1858.
Chipuk, J.E., Green, D.R., 2005. Do inducers of apoptosis trigger caspase-independent cell death? Nat. Rev., Mol. Cell Biol. 6, 268–275.
Cohen, G.M., 1997. Caspases: the executioners of apoptosis. Biochem. J. 326,1–16.
Friesen, C., Herr, I., Krammer, P.H., Debatin, K.M., 1996. Involvement of theCD95 (APO-1/FAS) receptor/ligand system in drug-induced apoptosis inleukemia cells. Nat. Med. 2, 574–577.
González, A.G., Bermejo, J., Triana, J., López, M., Eiroa, J.L., 1990.Sesquiterpene lactones from Tanacetum ferulaceum. Phytochemistry 29,2339–2341.
González, A.G., Bermejo, J., Triana, J., Eiroa, J.L., López, M., 1992a.Germacranolides from Allagopappus viscosissimus. Phytochemistry 31,330–331.
González, A.G., Bermejo, J., Triana, J., López, M., Eiroa, J.L., 1992b.Sesquiterpene lactones and other constituents of Tanacetum species.Phytochemistry 31, 1821–1822.
González, A.G., Bermejo, J., Triana, J., Eiroa, J.L., López,M., 1995. Sesquiterpenelactones and other constituents of Allagopappus species. J. Nat. Prod. 58,432–437.
Haldar, S., Jena, N., Croce, C.M., 1995. Inactivation of Bcl-2 by phosphory-lation. Proc. Natl. Acad. Sci. U. S. A. 92, 4507–4511.
Igney, F.H., Krammer, P.H., 2002. Death and anti-death: tumour resistance toapoptosis. Nat. Rev., Cancer 2, 277–288.
Kandaswami, C., Perkins, E., Soloniuk, D.S., Drzewiecki, G., Middleton, E.,1993. Ascorbic acid-enhanced antiproliferative effect of flavonoids onsquamous cell carcinoma in vitro. Anticancer Drugs 4, 91–96.
Kawaii, S., Tomono, Y., Katase, E., Ogawa, K., Yano, M., 1999. Antiprolifera-tive activity of flavonoids on several cancer cell lines. Biosci. Biotechnol.Biochem. 63, 896–899.
Kim, K.M., Lee, Y.J., 2005. Role of HER-2/neu signaling in sensitivity to tumornecrosis factor-related apoptosis-inducing ligand: enhancement of TRAIL-mediated apoptosis by amiloride. J. Cell. Biochem. 96, 376–389.
Kluck, R.M., Bozy-Wetzel, E., Green, D.R., Newmeyer, D.D., 1997. The releaseof cytochrome c from mitochondria: a primary site for Bcl-2 regulation ofapoptosis. Science 275, 1132–1136.
Ko, C.H., Shen, S.C., Lin, H.Y., Hou, W.C., Lee, W.R., Yang, L.L., Chen, Y.C.,2002. Flavanones structure related inhibition on TPA-induced tumorpromotion through suppression of extracellular signal-regulated proteinkinases: involvement of prostaglandin E2 in anti-promotive process. J. Cell.Physiol. 193, 93–102.
Krammer, P.H., 2000. CD95's deadly mission in the immune system. Nature407, 789–795.
Kuntz, S., Wenzel, U., Daniel, H., 1999. Comparative analysis of the effects offlavonoids on proliferation, cytotoxicity, and apoptosis in human coloncancer cell lines. Eur. J. Nutr. 38, 133–142.
Lee, W.R., Shen, S.C., Lin, H.Y., Hou, W.C., Yang, L.L., Chen, Y.C., 2002.Wogonin and fisetin induce apoptosis in human promyeloleukemic cells,accompanied by a decrease of reactive oxygen species, and activation ofcaspase 3 and Ca2+-dependent endonuclease. Biochem. Pharmacol. 63,225–236.
Li, P., Nijhawan, D., Budihardjo, I., Srinivasula, S.M., Ahmad, M., Alnemri, E.S.,Wang, X., 1997. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 91, 479–489.
Liu, X., Kim, C., Yang, J., Jemmerson, R., Wang, X., 1996. Induction ofapoptotic program in cell-free extracts: requirement for dATP andcytochrome c. Cell 86, 147–157.
Marsden, V.S., Strasser, A., 2003. Control of apoptosis in the immune system:Bcl-2, BH3-only proteins and more. Annu. Rev. Immunol. 21, 71–105.
Middleton, E., Kandaswami, C., Theoharides, T.C., 2000. The effects of plantflavonoids on mammalian cells: implications for inflammation, heartdisease, and cancer. Pharmacol. Rev. 52, 673–751.
Motwani, M., Delohery, T.M., Schwartz, G.K., 1999. Sequential dependentenhancement of caspase activation and apoptosis by flavopiridol onpaclitaxel-treated human gastric and breast cancer cells. Clin. Cancer Res.5, 1876–1883.
Muzio,M., Chinnaiyan, A.M., Kischkel, F.C., O'Rourke,K., Schevchenko,A., Ni,J., Scaffidi, C., Bretz, J.D., Zhang, M., Gentz, R., Mann, M., Krammer, P.H.,Peter, M.E., Dixit, V.M., 1996. FLICE, a novel FADD-homologous ICE/CED-3-like protease, is recruited to theCD95 (Fas/APO-1) death-inducing signallingcomplex. Cell 85, 817–827.
Nicholson, D.W., Thornberry, N.A., 1997. Caspases: killer proteases. TrendsBiochem. Sci. 22, 299–306.
Rivero, A., Quintana, J., Eiroa, J.L., López, M., Triana, J., Bermejo, J., Estévez,F., 2003. Potent induction of apoptosis by germacranolide sesquiterpenelactones on human myeloid leukemia cells. Eur. J. Pharmacol. 482, 77–84.
Srinivasula, S., Ahmad, M., Alnemri, T., Alnemri, E., 1998. Autoactivation ofprocaspase-9 by Apaf-1 mediated oligomerization. Mol. Cell 1, 949–957.
Stennicke, H.R., Jurgensmeier, J.M., Shin, H., Deveraux, Q., Wolf, B.B., Yang,X., Zhou, Q., Ellerby, H.M., Ellerby, L.M., Bredesen, D., Green, D.R., Reed,J.C., Froelich, C.J., Salvesen, G.S., 1998. Procaspase-3 is a majorphysiologic target of caspase-8. J. Biol. Chem. 273, 27084–27090.
Szewczyk, A., Wojtczak, L., 2002. Mitochondria as a pharmacological target.Pharmacol. Rev. 54, 101–127.
Thornberry, N.A., Lazebnik, Y., 1998. Caspases: enemies within. Science 281,1312–1316.
Triana, J., López, M., Pérez, F.J., González-Platas, J., Quintana, J., Estévez, F.,León, F., Bermejo, J., 2005. Sesquiterpenoids from Pulicaria canariensisand their cytotoxic activities. J. Nat. Prod. 68, 523–531.
Van Gurp, M., Festjens, N., van Loo, G., Saelens, X., Vandenabeele, P., 2003.Mitochondrial intermembrane proteins in cell death. Biochem. Biophys.Res. Commun. 304, 487–497.
Yang, J., Liu, X., Bhalla, K., Kim, C.N., Ibrado, A.M., Cai, J., Peng, T.I., Jones,D.P., Wang, X., 1997. Prevention of apoptosis by Bcl-2: release ofcytochrome c from mitochondria blocked. Science 275, 1129–1132.
Related Documents











