Am. J. Respir. Cell Mol. Biol. Vol. 23, pp. 636–645, 2000 Internet address: www.atsjournals.org Phenotypically Different Cells with Heterogeneous Nuclear Ribonucleoprotein A2/B1 Overexpression Show Similar Genetic Alterations Yan-Gao Man, Alfredo Martinez, Ingalill M. Avis, Sung H. Hong, Frank Cuttitta, David J. Venzon, and James L. Mulshine Intervention Section, Department of Cell and Cancer Biology; and Biostatistics and Data Management Section, Medicine Branch, Division of Clinical Science, National Cancer Institute, Bethesda, Maryland Immunocytochemical studies have revealed that overexpres- sion of heterogeneous nuclear ribonucleoprotein (hnRNP) A2/ B1 in exfoliated epithelial cells is a potentially useful marker of early lung cancer. This study analyzed the correlation of hnRNP A2/B1 expression with molecular alterations in pheno- typically different epithelial cells of paraffin-embedded pul- monary tissues. Sections from 20 human subjects were ana- lyzed immunohistochemically for expression of hnRNP A2/B1. Normal-appearing, hyperplastic, and malignant epithelial cells with and without hnRNP A2/B1 expression (n 5 78) were mi- crodissected and assessed for microsatellite alterations (MA) and loss of heterozygosity (LOH) (n 5 14 markers) as well as for clonality. Results showed that (1) hnRNP A2/B1 immu- noreactive cells contained a significantly higher frequency of MA and LOH than did comparable cells that lacked detectable hnRNP A2/B1; (2) over 80% of MA and LOH seen in hnRNP A2/B1 immunoreactive normal-appearing and hyperplastic cells persisted in malignant cells; (3) preliminary analysis of methylation status of the androgen receptor gene in non-neo- plastic cells was suggestive of hnRNP A2/B1–expressing cells being of clonal origin; and (4) cells with cytoplasmic hnRNP A2/B1 immunoreactivity had a 3-fold higher frequency of MA and LOH than did cells with nuclear hnRNP A2/B1 immunore- activity. These findings suggest that phenotypically different respiratory epithelial cells with hnRNP A2/B1 overexpression might be clonally derived, and that the subcellular localization of hnRNP A2/B1 might be an important factor associated with tumor progression. Lung cancer is the leading cause of cancer death in both males and females, accounting for nearly one in three can- cer deaths in the United States (1). Previous efforts to re- duce lung cancer mortality through earlier cancer detec- tion by using a combined chest X-ray and sputum cytomorphology failed to show a significant reduction in lung cancer–related mortality (2). A pilot study of archival sputum specimens from a high-risk cohort identified a monoclonal antibody (mAb) that specifically reacted with “normal”-appearing bronchial epithelial cells from indi- viduals who subsequently developed lung cancer, provid- ing a more sensitive means for early detection of lung can- cer (3). This antibody was later found to specifically recognize human heterogeneous nuclear ribonucleopro- tein (hnRNP) A2/B1 (4). Preliminary results of prospec- tive studies in two new high-risk lung cancer cohorts showed that immunocytochemical assessments of hnRNP A2/B1 expression is sputum specimens with the same anti- body accurately predicted the outcome in 32 of 40 (80%) and in 69 of 94 (73.4%) subjects who subsequently devel- oped lung cancer, whereas cytologic changes suggestive of lung cancer were found in less than 10% of the subjects (5). In that report, many eventual cancers were found in cases with hnRNP A2/B1 overexpression that showed min- imal cytologic abnormalities during the initial examination (5). Subsequent studies in tissue samples with the same an- tibody, however, detected dynamic patterns of hnRNP A2/B1 expression during mammalian lung development (6), and also found hnRNP A2/B1 overexpression in both neoplastic and surrounding non-neoplastic lung tissues of patients with Stage I non–small cell lung cancer (7). In ad- dition, overexpression of hnRNP A2/B1 was detected in 41% of normal-appearing and in 37% of metaplastic respi- ratory epithelial cells from multiple biopsy specimens of chronic smokers with metaplasia obtained from a previ- ously published trial of 13-cis-retinoic acid chemopreven- tion for bronchial metaplasia (8). In that study hnRNP A2/ B1 expression status was not different between current smokers and recent former smokers. Inasmuch as only about 1% of smokers with metaplasia would be expected to develop lung cancer, the frequent expression of hnRNP A2/B1 in the central airway is hard to reconcile with the reported degree of diagnostic preci- sion of hnRNP A2/B1 expression in sputum specimens, raising an important question about the actual significance of hnRNP A2/B1 in the airway. We have previously suggested that part of the diagnos- tic utility of immunocytochemical evaluation of bronchial epithelial cells recovered in the sputum compared with im- munohistochemical evaluation of biopsies of intact bron- chial epithelium related to the distinct biology of the types of cells recovered in the different specimens (8). The bron- chial epithelial cells that comprise the “cells of interest” in the sputum are cells that exist as individual cells not con- nected to neighboring epithelial cells as is the case with the bronchial epithelial cells recovered from a bronchial bi- opsy. The solitary cells-of-interest from the sputum may represent a population of cells that are caught up in lung carcinogenesis related to the loss of contact with neighbor- ing cells and the basement membrane, accounting for why such cells may be more diagnostically informative than the interconnected bronchial epithelial cells recovered with a bronchial biopsy. In this study, our question was whether focusing on cells that overexpress hnRNP A2/B1 actually contributes to (Received in original form March 21, 2000 and in revised form July 10, 2000) Address correspondence to: James L. Mulshine, Head of Intervention Sec- tion, Dept. of Cell and Cancer Biology, Medicine Branch, Div. of Clinical Science, National Cancer Institute, Bldg. 10, Bethesda, MD 20892. E-mail: [email protected] Abbreviations: heterogeneous nuclear ribonucleoprotein, hnRNP; loss of heterozygosity, LOH; microsatellite alteration(s), MA; messenger RNA, mRNA; polymerase chain reaction, PCR.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Am. J. Respir. Cell Mol. Biol. Vol. 23, pp. 636–645, 2000Internet address: www.atsjournals.org
Phenotypically Different Cells with Heterogeneous Nuclear Ribonucleoprotein A2/B1 Overexpression Show Similar Genetic Alterations
Yan-Gao Man, Alfredo Martinez, Ingalill M. Avis, Sung H. Hong, Frank Cuttitta, David J. Venzon,and James L. Mulshine
Intervention Section, Department of Cell and Cancer Biology; and Biostatistics and Data Management Section, Medicine Branch,Division of Clinical Science, National Cancer Institute, Bethesda, Maryland
Immunocytochemical studies have revealed that overexpres-sion of heterogeneous nuclear ribonucleoprotein (hnRNP) A2/B1 in exfoliated epithelial cells is a potentially useful marker ofearly lung cancer. This study analyzed the correlation ofhnRNP A2/B1 expression with molecular alterations in pheno-typically different epithelial cells of paraffin-embedded pul-monary tissues. Sections from 20 human subjects were ana-lyzed immunohistochemically for expression of hnRNP A2/B1.Normal-appearing, hyperplastic, and malignant epithelial cells
with and without hnRNP A2/B1 expression (
n
5
78) were mi-crodissected and assessed for microsatellite alterations (MA)and loss of heterozygosity (LOH) (
n
5
14 markers) as well asfor clonality. Results showed that (
1
) hnRNP A2/B1 immu-noreactive cells contained a significantly higher frequency ofMA and LOH than did comparable cells that lacked detectablehnRNP A2/B1; (
2
) over 80% of MA and LOH seen in hnRNPA2/B1 immunoreactive normal-appearing and hyperplasticcells persisted in malignant cells; (
3
) preliminary analysis ofmethylation status of the androgen receptor gene in non-neo-plastic cells was suggestive of hnRNP A2/B1–expressing cellsbeing of clonal origin; and (
4
) cells with cytoplasmic hnRNPA2/B1 immunoreactivity had a 3-fold higher frequency of MAand LOH than did cells with nuclear hnRNP A2/B1 immunore-activity. These findings suggest that phenotypically differentrespiratory epithelial cells with hnRNP A2/B1 overexpressionmight be clonally derived, and that the subcellular localizationof hnRNP A2/B1 might be an important factor associated withtumor progression.
Lung cancer is the leading cause of cancer death in bothmales and females, accounting for nearly one in three can-cer deaths in the United States (1). Previous efforts to re-duce lung cancer mortality through earlier cancer detec-tion by using a combined chest X-ray and sputumcytomorphology failed to show a significant reduction inlung cancer–related mortality (2). A pilot study of archivalsputum specimens from a high-risk cohort identified amonoclonal antibody (mAb) that specifically reacted with“normal”-appearing bronchial epithelial cells from indi-viduals who subsequently developed lung cancer, provid-ing a more sensitive means for early detection of lung can-cer (3). This antibody was later found to specificallyrecognize human heterogeneous nuclear ribonucleopro-tein (hnRNP) A2/B1 (4). Preliminary results of prospec-
tive studies in two new high-risk lung cancer cohortsshowed that immunocytochemical assessments of hnRNPA2/B1 expression is sputum specimens with the same anti-body accurately predicted the outcome in 32 of 40 (80%)and in 69 of 94 (73.4%) subjects who subsequently devel-oped lung cancer, whereas cytologic changes suggestive oflung cancer were found in less than 10% of the subjects(5). In that report, many eventual cancers were found incases with hnRNP A2/B1 overexpression that showed min-imal cytologic abnormalities during the initial examination(5). Subsequent studies in tissue samples with the same an-tibody, however, detected dynamic patterns of hnRNPA2/B1 expression during mammalian lung development(6), and also found hnRNP A2/B1 overexpression in bothneoplastic and surrounding non-neoplastic lung tissues ofpatients with Stage I non–small cell lung cancer (7). In ad-dition, overexpression of hnRNP A2/B1 was detected in41% of normal-appearing and in 37% of metaplastic respi-ratory epithelial cells from multiple biopsy specimens ofchronic smokers with metaplasia obtained from a previ-ously published trial of 13-cis-retinoic acid chemopreven-tion for bronchial metaplasia (8). In that study hnRNP A2/B1 expression status was not different between currentsmokers and recent former smokers.
Inasmuch as only about 1% of smokers with metaplasiawould be expected to develop lung cancer, the frequentexpression of hnRNP A2/B1 in the central airway is hardto reconcile with the reported degree of diagnostic preci-sion of hnRNP A2/B1 expression in sputum specimens,raising an important question about the actual significanceof hnRNP A2/B1 in the airway.
We have previously suggested that part of the diagnos-tic utility of immunocytochemical evaluation of bronchialepithelial cells recovered in the sputum compared with im-munohistochemical evaluation of biopsies of intact bron-chial epithelium related to the distinct biology of the typesof cells recovered in the different specimens (8). The bron-chial epithelial cells that comprise the “cells of interest” inthe sputum are cells that exist as individual cells not con-nected to neighboring epithelial cells as is the case with thebronchial epithelial cells recovered from a bronchial bi-opsy. The solitary cells-of-interest from the sputum mayrepresent a population of cells that are caught up in lungcarcinogenesis related to the loss of contact with neighbor-ing cells and the basement membrane, accounting for whysuch cells may be more diagnostically informative than theinterconnected bronchial epithelial cells recovered with abronchial biopsy.
In this study, our question was whether focusing oncells that overexpress hnRNP A2/B1 actually contributes to
(
Received in original form March 21, 2000 and in revised form July 10, 2000
)
Address correspondence to:
James L. Mulshine, Head of Intervention Sec-tion, Dept. of Cell and Cancer Biology, Medicine Branch, Div. of ClinicalScience, National Cancer Institute, Bldg. 10, Bethesda, MD 20892. E-mail:[email protected]
Abbreviations:
heterogeneous nuclear ribonucleoprotein, hnRNP; loss ofheterozygosity, LOH; microsatellite alteration(s), MA; messenger RNA,mRNA; polymerase chain reaction, PCR.

Man, Martinez, Avis,
et al
.: hnRNP Expression and Genetic Alteration 637
the identification of individuals in the process of develop-ing a lung cancer. To evaluate the significance of hnRNPA2/B1 overexpression, we compared the frequency andpattern of several recognized markers of lung carcinogen-esis, including microsatellite alterations (MA) and loss ofheterozygosity (LOH), as well as clonality in phenotypi-cally different respiratory epithelial cells that did or didnot express high levels of hnRNP A2/B1. We conductedthis analysis for two reasons: (
1
) MA and LOH are earlysigns of defective DNA repair or replication, which pre-cede morphologic abnormalities and are changes sugges-tive of a tissue involved in carcinogenesis (9–14); (
2
)clonality analysis has been successfully applied to distin-guish the neoplastic or reactive nature of several types ofhuman tumors (15–19).
hnRNP A2/B1 appears not only to play important rolesin messenger RNA (mRNA) processing but also to partic-ipate actively in post-transcriptional events (20–23), sothat the overexpression of this protein could be causallyinvolved in the cancer process. If the overexpression ofhnRNP A2/B1 correlates with MA, LOH, and clonality,this association strengthens the possibility of hnRNP A2/B1 involvement in the cancer process. Conversely, if nocorrelation is found between hnRNP A2/B1 overexpres-sion and genetic alterations, it would then be unlikely thathnRNP A2/B1 expression was contributing to the processor identification of carcinogenesis.
Materials and Methods
Case Selection and Classification
In this initial study, the focus was to explore the correlation ofhnRNP A2/B1 expression status with genetic alterations in tis-sues with defined histologic differentiation. To do this, paraffin-embedded respiratory tissue samples from 20 human subjectswere used under an institutionally approved tissue procurementprotocol. This included lung tissue from six cancer patients. Toobtain a range of well-preserved non-neoplastic tissues, we usedlung tissue harvested from lung transplants and autoptic tissuesobtained from trauma victims. In the 12 cases from individualswhose lungs were harvested during the course of lung transplan-tation, and two cases from young victims who died of trauma-related causes, extensive clinical information was not available.In previous publications the expression of hnRNP A2/B1 was notrelated to either gender or type of lung cancer, so in this studyour analysis was directed at the coincidence of lung cancer–related molecular events with hnRNP A2/B1 expression (3, 5).
Serial sections of 5 to 7
m
m were placed on poly-
L
-lysine–coatedor positively charged microscopic slides. Sections were incubatedovernight at 37
8
C, deparaffinized with three changes of xylene,washed with descending concentrations of ethanol and tap water,and stained with hematoxylin and eosin for morphologic assess-ment. The morphologic designations were determined by inde-pendent reviews at the National Cancer Institute and the ArmedForces Institute of Pathology using published criteria (20). Ofthese, six were normal-appearing; eight were hyperplastic bron-chial and/or alveolar epithelium with no or mild atypia; two were
well-differentiated,
in situ
lung carcinomas; and four were well-differentiated invasive lung carcinomas with mixed (endocrine,spindle, and squamous) cell types. A total of 11 cases containedboth bronchial and alveolar regions, five contained only bron-chial epithelium, and four contained only alveolar tissues. Noneof the tissues from the six cancer patients contained clear-cut nor-mal epithelium, but all contained normal-appearing stromal ele-
ment. Only two cases, one with normal-appearing and the otherwith hyperplastic alveolar tissue, were nonsmokers.
Immunohistochemical Staining
A previously characterized mouse mAb that recognizes humanhnRNP A2/B1 was used (3). Detection kits with a biotinylatedsecondary antibody to mouse immunoglobulin G, streptavidin-conjugated alkaline phosphatase, and Fast Red chromogen, werepurchased from BioGenex (San Ramon, CA). Immunostainingwas performed using our published protocol (6–8). Briefly, de-paraffinized sections were incubated with the primary antibodysolution (4
m
m/ml or 1:100 dilution) or normal serum (4
m
g/ml)overnight at 4
8
C. After two 5-min washes in 1
3
phosphate-buff-ered saline (PBS) that followed each step, sections were sequen-tially incubated with the secondary antibody, streptavidin-conju-gated alkaline phosphatase, and chromogen. After chromogenreaction, sections were counterstained with hematoxylin, brieflywashed with tap and distilled water, and mounted in 3% gelatinmounting medium for microscopic evaluation. A staining distri-bution score (0: no immunoreactive cells; 1: 1 to 10% immunore-active cells; 2: 11 to 50% immunoreactive cells; 3: 51 to 100%immunoreactive cells) and staining intensity score (0: nonimmu-noreactive; 1:
1
; 2:
11
; 3:
111
) were obtained for each sample.Using the sum of these two scores, a staining index (SI) was es-tablished for each case. A given sample was considered immu-noreactive when the SI
.
2 (7, 8). The immunoreactive caseswere subclassified as cytoplasmic reactive if
>
85% of the immu-noreactive cells showed localization of hnRNP A2/B1 in their cy-toplasm, or nuclear immunoreactive when
>
85% of the reactivecells displayed hnRNP A2/B1 immunoreactivity in their nuclei.Positive controls included two hnRNP A2/B1 immunoreactivebronchial and alveolar samples identified in previous studies (6,7). Negative controls included substitution of the primary anti-body with nonimmune serum, or omission of the primary or sec-ondary antibody from the immunostaining sequence. Immuno-staining for each of the samples was repeated three times underthe same conditions.
Assessments for MA and LOH
The coverslips of immunostained sections were removed by soak-ing the slides in 50-to-60
8
C hot water. After a 3-to-5-min wash inwarm tap water, the sections were soaked in 1
3
PBS (pH 7.4)containing 10% glycerin. For each case, multiple foci of epithelialcell clusters with and without hnRNP A2/B1 expression, and stro-mal elements near or far away from the epithelium were microdis-sected under a standard microscope. The dissected sample fromeach focus was placed in a separate tube and subjected to protein-ase K digestion as previously described (16, 24). DNA extracts ofthese samples were assessed for MA and LOH by polymerasechain reaction (PCR) amplification with 14 polymorphic DNAmarkers, which were selected on the basis of the following criteria:(
1
) they were located on chromosomes that harbor proven or pu-tative tumor suppressor genes; and (
2
) they were found to havehigh frequencies of MA and LOH in a variety of malignant andpremalignant lung lesions (9–14). The primers were purchasedfrom Research Genetics (Huntsville, AL) and labeled with fluo-rescent dyes, and are listed in Table 1. The cases selected for as-sessments of MA and LOH and the types and foci of the epithelialcells microdissected are listed in Table 2. Gene Amp PCR kits,Taq gold DNA polymerase, and DNA size standard were ob-tained from Perkin–Elmer (Foster City, CA). PCR amplificationwas carried out in a programmable thermal cycler (Perkin–Elmer)at the following settings: after a denaturation at 94
8
C for 14 min,the samples were amplified for 35 to 40 cycles at 94
8
C, 55 to 60
8
C,and 72
8
C, each for 1 min, with a final extension at 72
8
C for 10 min.Amplified products were subjected to electrophoresis in 5 to 6%
638
AMERICAN JOURNAL OF RESPIRATORY CELL AND MOLECULAR BIOLOGY VOL. 23 2000
polyacrylamide gels (Bio-Rad, Foster City, CA) and the signalwas detected with an automated 377 DNA sequencer (Perkin–Elmer). The expected PCR products were located and examinedby comparing the mobility of the predominant DNA bands ineach case with that of the molecular weight size marker, and bycomparing the mobility and intensity of the amplified specificDNA bands from the tumor cells with those or normal controls(normal stromal or epithelial cells). MA was defined as an addi-tion or deletion of one or more repeat units that results in the shiftof microsatellite alleles; LOH was defined as the complete ab-sence or at least 95% reduction of one allele (16, 24).
Clonality Analysis
Specimens from all six female subjects, including three with inva-sive carcinoma, two with hyplastic epithelium, and one with nor-mal-appearing epithelium, were analyzed for clonality. Clonalityanalysis was carried out by assessing the DNA methylation pat-tern with a pair of fluorescent dye–labeled DNA markers (5
9
-GCTGTGAAGGTTGCTGTTCCTCAT-3
9
and 5
9
-TCCAGAA-TCTGTTCCAGAGCGTGC-3
9
) located on exon 1 of the humanandrogen receptor (HUMARA) gene (19). Rsa 1 and Hpa II en-zymes were purchased from GIBCO BRL/Life Technologies(Gaithersburg, MD). The primers were purchased from ResearchGenetics, and the same procedures described in our previous stud-ies (16, 24) were used with some modifications. Briefly, in eachcase, two or three different foci of epithelial cells with and withouthnRNP A2/B1 expression and stromal cells near or far away (atleast 15 mm) from the epithelium were separately collected into
different tubes. Duplicates of DNA extracts from each case weremixed with Rsa 1 or Hpa II enzymes and incubated overnight at42
8
C. After incubation, the mixtures were treated at 95
8
C for 10min to inactivate the enzymes. Mixtures were then subjected toPCR amplification and electrophoresis as described earlier. Mon-oclonality was defined as the presence of only one DNA band af-ter Hpa II digestion, compared with two bands in normal controlsand in samples treated with the control enzyme Rsa I.
Production of the Gel Images for LOH Assessment and Clonality Analysis
The production of gel images was carried out according to theUser’s Manual (Perkin–Elmer), and used protocol modificationsdeveloped in our laboratory (25).
Statistical Analysis
Tests of the equality of the probability of a genetic alteration(MA or LOH) across samples and across DNA markers wereconducted by exact computation of the Fisher–Freeman–Maltontest. Because significant heterogeneity was found, the data wereanalyzed using a generalized estimating equations technique asimplemented in the GENMOD procedures of SAS version 6.12(SAS Institute, Cary, NC). For each sample, the determinationsof genetic alterations in the 14 DNA markers were considered tobe repeated binary observations with an exchangeable correla-tion structure. Noninformative results were treated as missingvalues. A logistic regression model for the probability of geneticalterations as a function of sample and marker was used. Thismodel diverges for observed proportions of 0 or 1, and somemarkers and some samples had no alterations. Therefore, indica-tor variables for each sample and each marker could not be used;instead, the samples and the markers were classified by their ob-served marginal genetic alteration proportions
P
into categoriesof 0
,
P
,
0.2, 0.2
,
P
,
0.4, and so on. In one analysis, a markerproportion on the 0.2 boundary was assigned to the lower cate-gory to prevent model divergence. For each two-group compari-son, the relative risk parameters were estimated separately withineach group being compared, a linear contrast was formed byweighing each parameter by the frequency of its category withinthe group, and the contrast was tested by the likelihood ratiomethod. Analysis of the residuals showed consistency with themodeling assumptions.
Results
hnRNP A2/B1 Expression in Normal-Appearing and Hyperplastic Epithelial Cells
Distinct epithelial cells demonstrating immunoreactivityfor hnRNP A2/B1 were detected in five or six (83.3%) nor-mal-appearing, and in five of eight (62.5%) hyperplastic
TABLE 1
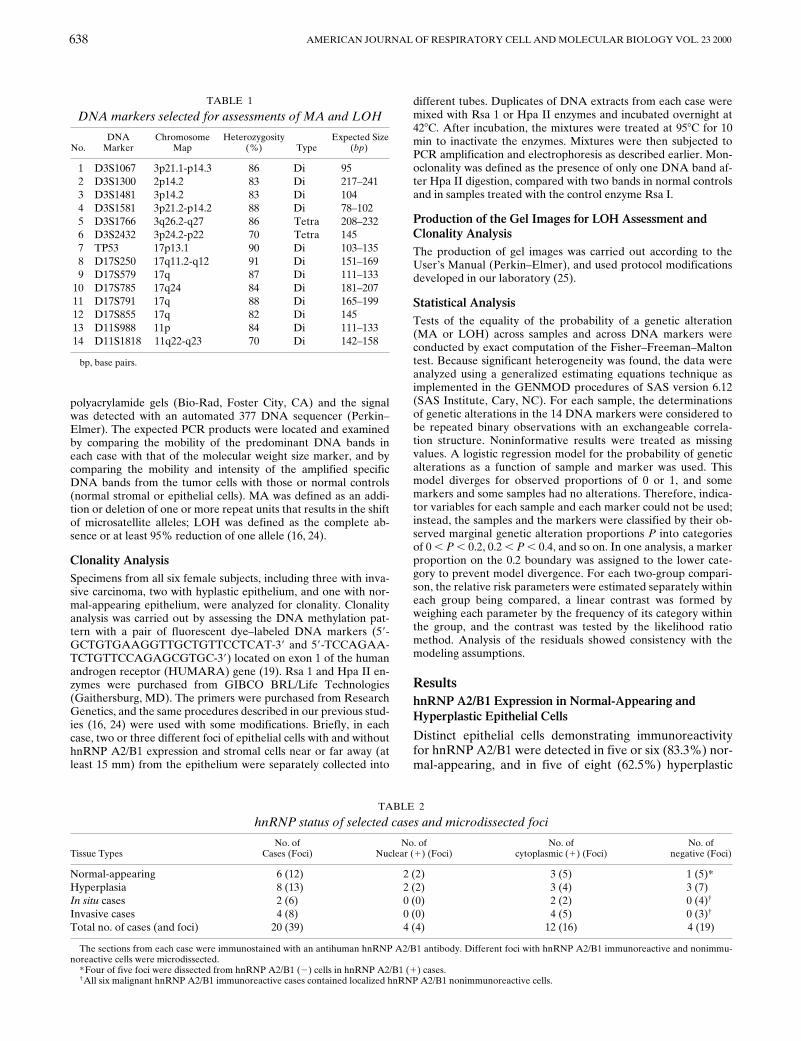
DNA markers selected for assessments of MA and LOH
No.DNA
MarkerChromosome
MapHeterozygosity
(
%
) Type Expected Size
(
bp
)
1 D3S1067 3p21.1-p14.3 86 Di 952 D3S1300 2p14.2 83 Di 217–2413 D3S1481 3p14.2 83 Di 1044 D3S1581 3p21.2-p14.2 88 Di 78–1025 D3S1766 3q26.2-q27 86 Tetra 208–2326 D3S2432 3p24.2-p22 70 Tetra 1457 TP53 17p13.1 90 Di 103–1358 D17S250 17q11.2-q12 91 Di 151–1699 D17S579 17q 87 Di 111–133
10 D17S785 17q24 84 Di 181–20711 D17S791 17q 88 Di 165–19912 D17S855 17q 82 Di 14513 D11S988 11p 84 Di 111–13314 D11S1818 11q22-q23 70 Di 142–158
bp, base pairs.
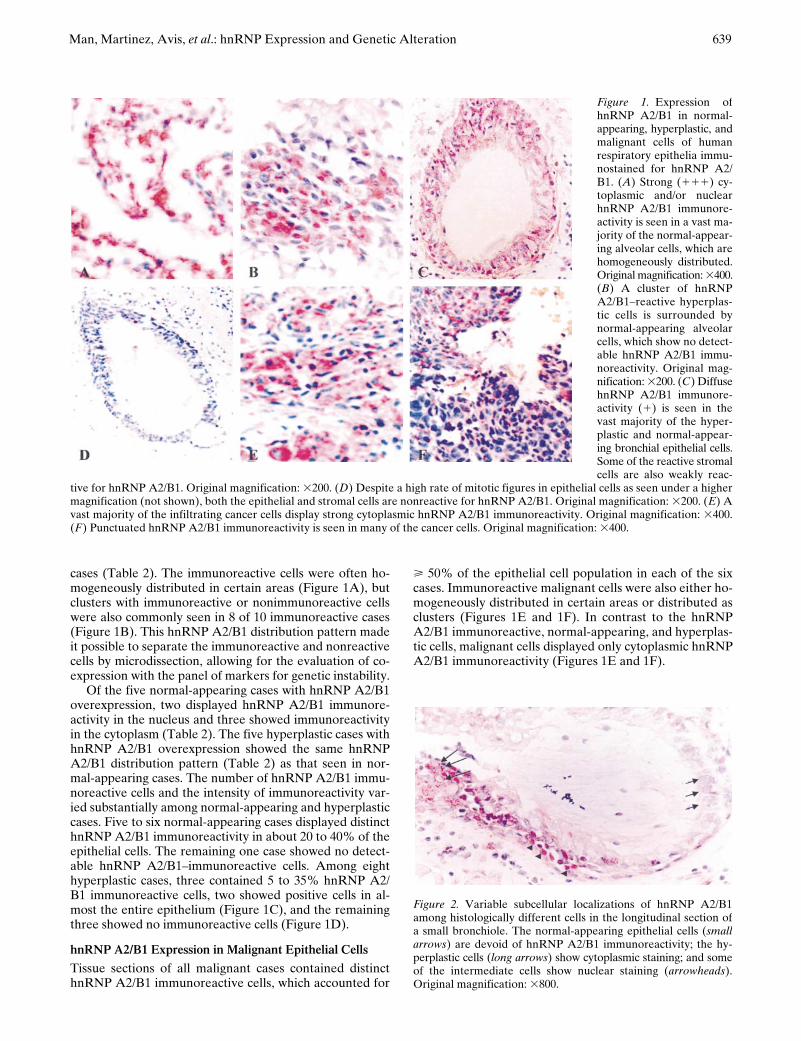
TABLE 2
hnRNP status of selected cases and microdissected foci
Tissue TypesNo. of
Cases (Foci)No. of
Nuclear (
1
) (Foci)No. of
cytoplasmic (
1
) (Foci)No. of
negative (Foci)
Normal-appearing 6 (12) 2 (2) 3 (5) 1 (5)*Hyperplasia 8 (13) 2 (2) 3 (4) 3 (7)
In situ
cases 2 (6) 0 (0) 2 (2)
0 (4)
†
Invasive cases 4 (8) 0 (0) 4 (5) 0 (3)
†
Total no. of cases (and foci) 20 (39) 4 (4) 12 (16) 4 (19)
The sections from each case were immunostained with an antihuman hnRNP A2/B1 antibody. Different foci with hnRNP A2/B1 immunoreactive and nonimmu-noreactive cells were microdissected.
*Four of five foci were dissected from hnRNP A2/B1 (
2
) cells in hnRNP A2/B1 (
1
) cases.
†
All six malignant hnRNP A2/B1 immunoreactive cases contained localized hnRNP A2/B1 nonimmunoreactive cells.
Man, Martinez, Avis,
et al
.: hnRNP Expression and Genetic Alteration 639
Figure 1. Expression ofhnRNP A2/B1 in normal-appearing, hyperplastic, andmalignant cells of humanrespiratory epithelia immu-nostained for hnRNP A2/B1. (A) Strong (111) cy-toplasmic and/or nuclearhnRNP A2/B1 immunore-activity is seen in a vast ma-jority of the normal-appear-ing alveolar cells, which arehomogeneously distributed.Original magnification: 3400.(B) A cluster of hnRNPA2/B1–reactive hyperplas-tic cells is surrounded bynormal-appearing alveolarcells, which show no detect-able hnRNP A2/B1 immu-noreactivity. Original mag-nification: 3200. (C) DiffusehnRNP A2/B1 immunore-activity (1) is seen in thevast majority of the hyper-plastic and normal-appear-ing bronchial epithelial cells.Some of the reactive stromalcells are also weakly reac-
tive for hnRNP A2/B1. Original magnification: 3200. (D) Despite a high rate of mitotic figures in epithelial cells as seen under a highermagnification (not shown), both the epithelial and stromal cells are nonreactive for hnRNP A2/B1. Original magnification: 3200. (E) Avast majority of the infiltrating cancer cells display strong cytoplasmic hnRNP A2/B1 immunoreactivity. Original magnification: 3400.(F) Punctuated hnRNP A2/B1 immunoreactivity is seen in many of the cancer cells. Original magnification: 3400.
cases (Table 2). The immunoreactive cells were often ho-mogeneously distributed in certain areas (Figure 1A), butclusters with immunoreactive or nonimmunoreactive cellswere also commonly seen in 8 of 10 immunoreactive cases(Figure 1B). This hnRNP A2/B1 distribution pattern madeit possible to separate the immunoreactive and nonreactivecells by microdissection, allowing for the evaluation of co-expression with the panel of markers for genetic instability.
Of the five normal-appearing cases with hnRNP A2/B1overexpression, two displayed hnRNP A2/B1 immunore-activity in the nucleus and three showed immunoreactivityin the cytoplasm (Table 2). The five hyperplastic cases withhnRNP A2/B1 overexpression showed the same hnRNPA2/B1 distribution pattern (Table 2) as that seen in nor-mal-appearing cases. The number of hnRNP A2/B1 immu-noreactive cells and the intensity of immunoreactivity var-ied substantially among normal-appearing and hyperplasticcases. Five to six normal-appearing cases displayed distincthnRNP A2/B1 immunoreactivity in about 20 to 40% of theepithelial cells. The remaining one case showed no detect-able hnRNP A2/B1–immunoreactive cells. Among eighthyperplastic cases, three contained 5 to 35% hnRNP A2/B1 immunoreactive cells, two showed positive cells in al-most the entire epithelium (Figure 1C), and the remainingthree showed no immunoreactive cells (Figure 1D).
hnRNP A2/B1 Expression in Malignant Epithelial Cells
Tissue sections of all malignant cases contained distincthnRNP A2/B1 immunoreactive cells, which accounted for
>
50% of the epithelial cell population in each of the sixcases. Immunoreactive malignant cells were also either ho-mogeneously distributed in certain areas or distributed asclusters (Figures 1E and 1F). In contrast to the hnRNPA2/B1 immunoreactive, normal-appearing, and hyperplas-tic cells, malignant cells displayed only cytoplasmic hnRNPA2/B1 immunoreactivity (Figures 1E and 1F).
Figure 2. Variable subcellular localizations of hnRNP A2/B1among histologically different cells in the longitudinal section ofa small bronchiole. The normal-appearing epithelial cells (smallarrows) are devoid of hnRNP A2/B1 immunoreactivity; the hy-perplastic cells (long arrows) show cytoplasmic staining; and someof the intermediate cells show nuclear staining (arrowheads).Original magnification: 3800.

640
AMERICAN JOURNAL OF RESPIRATORY CELL AND MOLECULAR BIOLOGY VOL. 23 2000
Cellular Localization of hnRNP A2/B1
The subcellular localizations of hnRNP A2/B1 often var-ied substantially in different cell clusters of the same case.Figure 2 shows a cross-section profile of a small bronchi-ole, in which the normal-appearing bronchial epithelialcells (
short arrows
) are devoid of hnRNP A2/B1 expres-sion, whereas the hyperplastic cells (
long arrows
) show cy-toplasmic hnRNP A2/B1 immunoreactivity. In addition,some of the cells located between the normal and hyper-plastic cell compartments (Figure 2,
arrowheads
) displaynuclear localization of hnRNP A2/B1.
Clonality Analysis
Samples for five of six female subjects were informativefor clonality analysis. Of these, two were malignant withheterogeneous expression of hnRNP A2/B1 (Figure 3A);one contained normal-appearing epithelium with uni-formly hnRNP A2/B1 immunoreactive cells (data notshown); one contained immunoreactive hyperplastic cellsthat constituted almost the entire epithelium (Figure 3D);and the remaining one contained hyperplastic epitheliumwith no detectable hnRNP A2/B1–expressing cells (Figure3G). hnRNP A2/B1 immunoreactive cells from the nor-mal-appearing and hyperplastic epithelia and cells fromthe two malignant cases showed the same monoclonal
Figure 3. Examples of micro-dissected cells and their clonalcomposition. (A–C) Invasivelung cancer with mixed celltypes. (A) Before microdissec-tion; (B) after microdissection;arrows indicate cells microdis-sected. hnRNP A2/B1 nonim-munoreactive cells were alsomicrodissected, but the loca-tion from which the cells wereharvested is not indicated inthe figure (c) Lanes 1 and 2:hnRNP A2/B1 immunoreac-tive cells; lane 3: hnRNP A2/B1 nonimmunoreactive cells;lanes 4–6: duplicates of lanes1–3. Lanes 1–3: treated withRsa 1 (control enzyme); lanes4–6: treated with Hpa II (a me-thylation-sensitive restriction en-donuclease). Both the hnRNPA2/B1–positive and –negativecells are monoclonal. (D–F)Homogeneous hnRNP A2/B1immunoreactive hyperplasticepithelium. (D) Before micro-dissection; (E) after microdis-section; arrows indicate cellsmicrodissected. (F) Lane 1:
hnRNP A2/B1 immunoreactive cells; lane 2: stromal cells far away from the epithelium; lane 3: stromal cells surrounding the epithelium;lanes 4–6: duplicates of lanes 1–3. Lanes 1–3: treated with Rsa 1 (control enzyme) lanes 4–6: treated with Hpa II. The hnRNP A2/B1 re-active epithelial cells (lane 4) and the adjacent stromal cells (lane 6) are monoclonal, whereas the normal stromal cells far away from theepithelium are polyclonal. (G–I) hnRNP A2/B1 nonimmunoreactive hyperplastic epithelium. (G) Before microdissection; (H) after mi-crodissection; arrows indicate cells microdissected. (I) Lane 1: hnRNP A2/B1 nonreactive epithelial cells; lane 2: stromal cells far awayfrom the epithelium; lane 3: stromal cells surrounding the epithelium; lanes 4–6: duplicates of lanes 1–3. Lanes 1–3: treated with Rsa 1(control enzyme); lanes 4–6: treated with Hpa II. All the cells are polyclonal.
Figure 4. Examples of co-occurrence of MA and LOH in normal-appearing (NA), hyperplastic (HP), and malignant (Malig) respi-ratory epithelial cells analyzed with different DNA markers. M:DNA size marker; NS: normal stromal cells; NE: normal epithe-lial cells; A2: hnRNP A2/B1 nonimmunoreactive alveolar cells;A1: hnRNP A2/B1 immunoreactive alveolar cells; B2: hnRNPnonimmunoreactive bronchial epithelial cells; B1: hnRNP im-munoreactive bronchial epithelial cells. Arrows indicate geneticalterations (either MA or LOH).

Man, Martinez, Avis, et al.: hnRNP Expression and Genetic Alteration 641
composition after Hpa II digestion (Figure 3C, lanes 4–6;Figure 3F, lane 4). The stromal cells near the hnRNP A2/B1 immunoreactive, normal-appearing, and hyperplasticepithelium showed the same monoclonality as did the epi-thelial cells (Figure 3F, lanes 4 and 6), whereas the stromalcells far away from the epithelium displayed a polyclonalcomposition (Figure 3F, lane 5). Cells from the hyperplas-tic lesion that lacked detectable hnRNP A2/B1 expression(Figure 3J, lanes 1–6), the normal controls, and the sam-ples treated with the control enzyme Rsa 1 all showed apolyclonal composition. Figure 3 shows the analysis of threerepresentative samples.
Frequenceis of MA and LOH in Normal-Appearing Epithelial Cells
The numbers of MA and LOH seen in each sample at eachchromosomal locus were summed and the combined fre-quencies of MA and LOH seen in hnRNP A2/B1 immu-noreactive or nonimmunoreactive cells are listed in Table3. Among a total of 85 informative foci in hnRNP A2/B1immunoreactive samples, a total of 34 MA and LOH weredetected (0.40 MA and LOH/focus) and each of the 14(100%) chromosomal loci displayed at least one MA andLOH; whereas among 65 informative loci in hnRNP A2/B1 nonreactive samples, a total of only 12 MA and LOHwere found (0.18 MA and LOH/focus), and 8 of the 14(57%) chromosomal loci showed genetic alterations. ThehnRNP A2/B1 immunoreactive cells had a significantlyhigher frequency (P , 0.01) of MA and LOH than didhnRNP A2/B1 nonreactive cells (Table 3).
Frequency of MA and LOH in Hyperplastic Epithelial Cells
The numbers of MA and LOH seen in each sample at eachchromosomal locus were summed and the combined fre-quencies of MA and LOH seen in hnRNP A2/B1 immu-noreactive or nonimmunoreactive cells are listed in Table3. Among a total of 75 informative foci in hnRNP A2/B1immunoreactive samples, a total of 22 MA and LOH weredetected (0.29 MA and LOH/focus), and MA and LOHwere distributed in 10 of the 14 (71.4%) chromosomal loci;whereas among 89 informative foci in hnRNP A2/B1 non-reactive samples, a total of only 13 MA and LOH werefound (0.15 MA and LOH/focus), and only 5 of 14 (36%)loci showed genetic alterations (Table 3). The hnRNP A2/
B1 immunoreactive cells had a significantly higher fre-quency of MA and LOH than did their hnRNP A2/B1nonreactive counterparts (P , 0.03).
Frequencies of MA and LOH in Malignant Epithelial Cells
The numbers of MA and LOH seen in each sample at eachchromosomal locus were summed and the combined fre-quencies of MA and LOH in hnRNP A2/B1 immunoreac-tive and nonimmunoreactive cells are listed in Table 3.Among 95 informative foci in hnRNP A2/B1 immunoreac-tive samples, a total of 47 MA and LOH were detected(0.49 MA or LOH/focus), and MA and LOH were seen in13 of the 14 (93%) chromosomal loci; whereas a total ofonly 28 MA and LOH were found in 91 informative foci(0.30 MA or LOH/focus) hnRNP nonreactive samples,and MA and LOH were seen in 11 of 14 (78%) chromo-somal loci (Table 3). The difference in the frequency ofMA and LOH between the reactive and nonreactive cellswas significant (P , 0.01).
The frequency and pattern of MA and LOH seenamong phenotypically different cells in replicated analyseswere consistent.
Concurrent MA or LOH in Normal-Appearing, Hyperplastic, and Malignant Cells
Concurrent MA or LOH was defined as the simultaneouspresence of either MA or LOH of either allele (of note forthe majority of LOH, the loss is from the same allele) at agiven chromosomal locus in both malignant and nonmalig-nant (including both normal-appearing and hyperplastic)tissues. MA and LOH detected in each cell phenotype ofdifferent cases were combined, and the combined fre-quency and pattern was used for comparison. For hnRNPA2/B1 immunoreactive nonmalignant cells compared withimmunoreactive malignant cells, concurrent MA and LOHwere found in 13 (92.9%) of the 14 chromosomal loci as-sessed (Table 4). In a parallel analysis, for the nonmalig-nant samples without hnRNP A2/B1 overexpression com-pared with the malignant samples with hnRNP A2/B1overexpression, concurrent genetic alterations were foundin 8 (57.1%) of the 14 loci (Table 4). For hnRNP A2/B1immunoreactive, nonmalignant samples compared withmalignant samples without hnRNP A2/B1 overexpression,concurrent MA and LOH were found in 11 (78.6%) of 14
TABLE 3
Combined frequencies of MA and LOH in normal-appearing, hyperplastic,and malignant epithelial cells in specimens from 20 patients
Cell TypeNo. of
Samples
MA and LOH at Each Chromosomal LocusNo. of
MA & LOHNo. of
IF FOCIAverage/
Focus P Value*1 2 3 4 5 6 7 8 9 10 11 12 13 14
NA (1) 7 3 2 3 2 3 2 3 2 2 1 4 2 2 3 34 85 0.40 , 0.01NA (2) 5 3 1 0 0 1 1 0 0 0 1 2 0 1 2 12 65 0.18
HP (1) 6 1 1 4 0 3 1 3 1 0 3 2 3 0 0 22 75 0.29 , 0.03HP (2) 7 1 3 0 0 3 3 0 0 0 0 3 0 0 0 13 89 0.15
MA (1) 7 3 2 3 2 6 4 4 2 0 3 6 4 4 4 47 95 0.49 , 0.01MA (2) 7 2 1 0 3 3 3 2 0 0 1 4 3 3 3 28 91 0.31
The numbers of MA and LOH seen in each sample at each chromosomal locus were summed. The total numbers of MA and LOH were divided by the total num-ber of informative foci to obtain the average of MA and LOH per focus. The average of MA and LOH per focus between the hnRNP immunoreactive and nonimmu-noreactive cells were compared by exact computation of the Fisher–Freeman–Malton test. NA, normal-appearing; HP, hyperplastic; MA, malignant; IF, informative.

642 AMERICAN JOURNAL OF RESPIRATORY CELL AND MOLECULAR BIOLOGY VOL. 23 2000
loci (Table 4). In the final comparison of nonmalignantand malignant cells without hnRNP A2/B1 overexpression,the frequency of shared genetic alterations was 57.1% (8of 14 loci).
Figure 4 shows examples of co-occurrence of MA andLOH at different chromosomal loci in hnRNP A2/B1 im-munoreactive, normal-appearing, and hyperplastic epithe-lial cells and their malignant counterparts.
Frequencies of MA and LOH in Cells with Cytoplasmic or Nuclear hnRNP A2/B1 Expression
Of the 13 hnRNP A2/B1 immunoreactive, normal-appear-ing, and hyperplastic samples, nine showed hnRNP A2/B1overexpression in the cytoplasm and four in the nucleus(Table 2). A total of 54 MA and LOH were detected in thenine samples with cytoplasmic hnRNP A2/B1 immunore-activity (6 MA and LOH/sample), whereas only eight MAand LOH were found in the four samples with nuclearhnRNP A2/B1 immunoreactivity (2 MA and LOH/sam-ple) (Table 5). The cells with cytoplasmic hnRNP A2/B1immunoreactivity had a significantly higher frequency ofMA and LOH than did cells with nuclear hnRNP A2/B1immunoreactivity (P , 0.01).
Of the seven hnRNP A2/B1 immunoreactive malignantsamples, all showed cytoplasmic immunoreactivity and nodistinct nuclear immunoreactivity was found in any ofthese samples. A total of 47 MA and LOH were detectedin these seven samples (6.7 MA and LOH/sample) (Ta-ble 5).
DiscussionAlthough hnRNP A2/B1 overexpression in sputum speci-mens has been consistently correlated with the eventualdevelopment of lung cancer in our previous studies in spu-tum specimens from over 8,000 patients (3, 5), the mecha-nism for hnRNP A2/B1 involvement in lung carcinogene-sis is unknown. To evaluate the correlation of hnRNP A2/B1 overexpression with molecular markers of lung car-cinogenesis, we compared the frequency and pattern ofMA and LOH among phenotypically different epithelialcells with and without hnRNP A2/B1 overexpression. Inthis study, we found that hnRNP A2/B1 immunoreactivecells had a significantly higher frequency of MA and LOH.
Over 80% of MA and LOH seen in hnRNP A2/B1 immu-noreactive, normal-appearing, and hyperplastic cells per-sisted in malignant cells. In addition, in the preliminaryanalysis, the hnRNP A2/B1 immunoreactive, normal-appearing, and hyperplastic cells tested were monoclonal,whereas comparable cells without hnRNP A2/B1 expres-sion were polyclonal. We also found that cytoplasmichnRNP A2/B1 immunoreactive cells had 3-fold higherrate of MA and LOH than did cells with nuclear immu-noreactivity. These data collectively suggest that (1) al-though normal-appearing or hyperplastic cells among dif-ferent cases or in the same case are morphologicallysimilar, they often display a substantially different geneticprofile, depending on their hnRNP A2/B1 expression sta-tus; and (2) cells with cytoplasmic hnRNP A2/B1 overex-pression appear to have accumulated more genetic alter-ations than other cells, which may account for their moreaggressive biologic behavior.
The DNA markers selected for this study were fromchromosomes 3, 11, and 17, which harbor known or puta-tive tumor suppressor genes, including fragile histidinetriad, Wilm’s tumor gene 1, and p53 (9–14). A number ofprevious studies with markers from these loci have shownhigh frequencies of MA and LOH, ranging from 15 to100%, in different malignant lung tissues (9–14). The fre-quencies of MA and LOH seen in this study ranged from14.3 to 57.1%, with an average of 48% in hnRNP A2/B1immunoreactive and 38.6% in nonimmunoreactive malig-nant cells. It is interesting to note that high frequencies ofMA and LOH have also been reported in about 50% ofhistologically normal specimens of smokers (11), in 31 to76% of specimens from smokers with metaplasia or dys-plasia (26), in 76 to 86% of nonmalignant lung tissues formlung cancer patients (27), and in 27% of patients with nocytologic or radiologic evidence of respiratory neoplasia(28). In this study, MA and LOH were seen in normal-appearing and hyperplastic cells without detectable hnRNPA2/B1 with an averaged frequency of 18 and 15%, respec-tively, and in cells with hnRNP A2/B1 overexpression withan averaged frequency of 40 and 29%, respectively (Table3). The consistent detection of high frequencies of MAand LOH in nonmalignant respiratory epithelial cells bydifferent studies on different populations suggests that
TABLE 4
Frequencies of co-occurrence of MA and LOH innon-neoplastic epithelial cells and their malignant
counterparts in specimens from 20 patients
Cell TypeNo. of
Chromosomal Loci
No. and % of Co-occurrence of
MA and LOH P Value*
NA & HP (1) : Malig (1) 14 13 (92.8%) , 0.02NA & HP (2) : Malig (1) 14 8 (57.1%)NA & HP (1) : Malig (2) 14 11 (78.5%) , 0.05NA & HP (2) : Malig (2) 14 8 (57.1%)
*The frequencies of co-occurrence of MA and LOH in non-neoplastic cellsand their malignant counterparts were obtained on the basis of the results listedin Table 3, and compared by exact computation of Fisher–Freeman–Maltontest. NA, normal-appearing epithelial cells; HP, hyperplastic epithelial cells;Malig, malignant epithelial cells.
TABLE 5
Frequencies of MA and LOH in epithelial cells with cytoplasmic versus nuclear overexpression ofhnRNP A2/B1 in specimens from 20 patients
Cell TypehnRNP
DistributionNo. of
SamplesNo. of
MA & LOHAverage MA
& LOH/Sample P Value*
Normal & Cytoplasm 9 54 6.0 , 0.01hyperplastic
Nucleus 4 8 2.0Malignant Cytoplasm 7 47 6.7
Nucleus 0 0 0
*The numbers of MA and LOH seen in each sample at all the chromosomalloci were summed. The total numbers of MA and LOH were divided by the to-tal number of the samples to obtain the average of MA and LOH per sampleThe average of MA and LOH in epithelial cells with cytoplasmic and nuclearhnRNP A2/B1 immunoreactivity was compared by exact computation of theFisher–Freeman–Malton test.

Man, Martinez, Avis, et al.: hnRNP Expression and Genetic Alteration 643
molecular biologic abnormalities precede morphologic ab-normalities. Therefore, assessment for MA and LOH maybe a more sensitive approach for the detection of earlylung cancer. MA and LOH assessment alone, however,can only suggest the general tendency but cannot preciselypredict when and which of the premalignant lesions mayprogress to lung cancer, for the following reasons: (1) thepresence of LOH at a given locus is not always a definitivesign of tumor suppressor gene inactivation because the re-maining allele may retain normal function; (2) there is cur-rently no consensus as to inactivation of which tumor sup-pressor gene defines the aggressive behavior of cells; and(3) LOH occurs at a low level (< 10%) randomly through-out the genome. Using a single molecular marker for lungcancer can be criticized because any individual marker suf-fers from low specificity for lung cancer, which is why weused a panel of markers frequently reported as being al-tered in lung cancer. Additional studies to better definethe relationship of molecular events to field carcinogenesisrelative to both synchronous and metachronous primarylung cancer would be of value.
Inasmuch as our previous immunocytochemical studiesof sputum specimens from high-risk cohorts have shownthat 73.4 to 80% of the individuals with exfoliated hnRNPA2/B1 immunoreactive epithelial cells eventually devel-oped lung cancer (6), it is likely that these cells may haveaccumulated additional abnormalities during the exfolia-tion process. Therefore, comparisons of the molecular bio-logic profile of hnRNP A2/B1 immunoreactive cells inrandom bronchial biopsies with that of comparable exfoli-ated cells recovered in sputum specimens may lead to theidentification of specific genetic markers that are associ-ated with the clinical progression of lung cancer. From thecurrent analysis, the expression of hnRNP A2/B1 in theairway should be considered not as a benign event but asan indication that at least some molecular events consis-tent with lung carcinogenesis have occurred. This state-ment reflects our current finding of a significant correlationbetween hnRNP A2/B1 immunoreactivity and molecularchanges associated with lung cancer even in tissue sec-tions. On the basis of our current findings, we speculatethat additional critical molecular events must occur whenthe bronchial epithelial cells are shed from the basementmembrane.
The further elucidation of a specific role of hnRNP A2/B1 and steps in the development of metastatic competencewill be of considerable interest, leading either to better un-derstanding of the mechanism of tumorigenesis and pro-gression or to more effective prevention and managementof lung cancer (28).
Clonality analysis based on the Lyon/Beutler’s hypoth-esis of random X chromosomal inactivation is thought tobe a reliable method of distinguishing between benign andneoplastic lesions (15–19). During the early stage of em-bryogenesis in the female, either the maternally or pater-nally derived X chromosome in each cell is randomly andpermanently inactivated, and this pattern of chromosomalinactivation is stable through subsequent cell cycles (15–19), which leads to somatic mosaicism of normal femaleswith half of the normal cells expressing the maternal andthe other half expressing the paternal allele. In contrast,
tumors arising from the clonal proliferation of a singleneoplastic cell will display only one, either the maternal orthe paternal phenotype. A highly polymorphic trinucle-otide CAG repeat in the X-linked HUMARA gene hasbeen used to distinguish between the inactivated and ac-tive chromosomes, and to determine the clonal composi-tion of a variety of lesions (15–19). In this study, two orthree foci of phenotypically different epithelial cells withand without hnRNP A2/B1 overexpression in each casewere microdissected for clonality analysis. This prelimi-nary result showed that hnRNP A2/B1 immunoreactivecells from non-neoplastic cases were monoclonal, whereascomparable cells without hnRNP A2/B1 were polyclonal.Although the small sample size has prevented us fromdrawing a general conclusion about the clonal compositionof non-neoplastic cells with hnRNP A2/B1 overexpres-sion, our results suggest that some of the hnRNP A2/B1immunoreactive, normal-appearing, and hyperplastic epi-thelial cells may be neoplastic and/or precursors of the ma-lignant lesions. Further, stromal cells near the hnRNP A2/B1 immunoreactive, normal-appearing, and hyperplasticepithelium shared the same monoclonal composition withthe epithelial cells, consistent with the assumption that aninteraction between stromal and epithelial cells may playan important role in carcinogenesis and tumor progression(24, 29, 30). Further investigation in this regard is ongoing.
The frequency and extent of hnRNP A2/B1 expressionwith molecular alterations appear to be independent of thecell differentiation status in non-neoplastic samples. Of sixnormal-appearing cases, five (83.3%) showed hnRNP A2/B1 overexpression, compared with five of eight (62.5%) caseswith hyperplastic cells. Also, in microdissected hnRNPA2/B1 immunoreactive, normal-appearing samples, a totalof 34 MA and LOH were detected in a total of 82 informa-tive foci (0.4 MA and LOH/focus), and all 14 (100%) chro-mosomal loci showed genetic abnormalities; whereas inhnRNP A2/B1 immunoreactive hyperplastic samples, a to-tal of 22 MA and LOH were found in a total of 75 infor-mative foci (0.29 MA and LOH/focus), and only 10 of the14 (71%) chromosomal loci displayed molecular alter-ations. The slightly higher frequency of hnRNP A2/B1 co-expression with genetic alterations in normal-appearingcompared with hyperplastic cells is consistent with ourpreviously reported finding with immunohistochemicalanalysis of 1,078 foci of phenotypically different respira-tory cells in biopsy specimens from smokers with metapla-sia, where overexpression occurred in 41% of the normal-appearing and in 37% of squamous metaplastic samples(8). Our early detection report also showed that a majorityof the individuals with hnRNP A2/B1 overexpression intheir sputum cells who subsequently developed lung can-cer had minimal cytomorphologic abnormalities in theirsputum cells (5). Other studies also showed that some his-tologically normal biopsy specimens from bronchial epi-thelium of current and former smokers showed allelic lossequal to or greater than that seen in in situ carcinoma le-sions (11), and that the cancerous and adjacent normal-appearing mammary epithelial cells share a similar fre-quency and pattern of genetic alterations (29).
The subcellular localization of hnRNP A2/B1 expres-sion correlates with both the cell differentiation status

644 AMERICAN JOURNAL OF RESPIRATORY CELL AND MOLECULAR BIOLOGY VOL. 23 2000
and molecular alterations. Among 14 cases with normal-appearing and hyperplastic epithelium, four (28.5%) con-tained no detectable hnRNP A2/B1 immunoreactive cells,four (28.5%) displayed nuclear immunoreactivity, and six(43%) showed cytoplasmic immunoreactivity. In contrast,all six malignant cases showed exclusive cytoplasmic local-ization of hnRNP A2/B1. The cytoplasmic localization ofhnRNP A2/B1 may have useful diagnostic implications.Assessments for MA and LOH revealed a 3-fold higherfrequency of MA and LOH in cells with cytoplasmichnRNP A2/B1 overexpression than in comparable cellswith nuclear hnRNP A2/B1 overexpression. Also, clonal-ity analysis of normal-appearing and hyperplastic cellswith cytoplasmic hnRNP A2/B1 overexpression showed amonoclonal composition, whereas the comparable cellsthat lacked detectable hnRNP A2/B1 expression displayeda polyclonal composition. These findings suggest that cyto-plasmic overexpression of hnRNP A2/B1 may be an im-portant factor that is closely associated with or signifiesthe progression of lung neoplasia.
This study was initiated because of the complexity inunderstanding the significance of hnRNP A2/B1 overex-pression. In previous studies, the expression of hnRNPA2/B1 in the exfoliated bronchial cells recovered in thesputum was highly associated with the eventual develop-ment of cancer (3, 5), but hnRNP A2/B1 overexpression inthe intact bronchial epithelial tissues harvested by bron-chial biopsy showed frequent expression in smokers withbronchial metaplasia. In the original study of the mAb tohnRNP A2/B1, we did not find significant immunoexpres-sion in the airways of young, healthy nonsmokers whodied of acute trauma. Relatively few of the smokers withmetaplasia would be expected to progress to lung cancer(8). In light of this disparate result, we wanted to betterunderstand how hnRNP A2/B1 expression is permittingeffective early lung cancer identification. To do this it isimportant to consider why there may be serious molecularchanges suggestive of lung cancer at a higher frequencythan the disease frequency actually observed in the gen-eral population. The first potentially confounding factor isthat the most important risk for mortality in individualwith tobacco consumption is cardiovascular diseases. Insmokers, death due to heart disease is a dominant compet-ing risk which would decrease the number of people withhnRNP A2/B1–associated abnormalities that would ulti-mately manifest as lung cancer. Similarly, because devel-oping a cancer is a multistep process, initiated or geneti-cally altered cells may be present for many years insmokers; only with protracted follow-up will the true rateof development of clinically significant lung cancer be es-tablished.
Another possible explanation as to why the sputum testis informative but the analysis of cells from intact bron-chial mucosa obtained by biopsy is not related to the biol-ogy of cancer progression. Evolving cancer cells lose con-tact with neighboring cells and the basement membrane asthey acquire the ability to invade or metastasize. Conse-quently, these evolving cancer cells are also prone to ex-foliate into the sputum. Solitary bronchial epithelial cellsrecovered in the sputum may preferentially indicate in-volvement with field carcinogenesis (3, 5, 8). The shed
cells-of-interest in the sputum have presumably escapednormal cellular clearance mechanisms because they are re-covered intact. Although this type of bronchial epithelialcell is present in low frequency, when these cells displaydetectable levels of hnRNP A2/B1 a high correlation ex-ists with the eventual development of an evident clinicalcancer. This utility presumably reflects not only the biol-ogy of hnRNP A2/B1 but also the biology of exfoliation(31). Comparisons of the genetic profiles of hnRNP A2/B1–immunoreactive cells in random bronchial biopsieswith those of comparable cells in sputum specimens maylead to the identification of more specific genetic markersthat are associated with true commitment to clinically sig-nificant cancer.
The mechanism underlying a correlation betweenhnRNP A2/B1 overexpression with genetic alterations isnot evident, inasmuch as a definite function for hnRNPA2/B1 in the progression of lung cancer has not been elu-cidated. It has been reported that telomere and telomeraseare associated with the regulation of diverse biologicevents such as cell growth, immortalization, and chromo-somal stability, and there is a growing literature about thepotential for interactions between hnRNP A2/B1 andclosely homologous family members with telomeric se-quences (21, 32, 33). Inasmuch as hnRNP A2/B1 hasbinding domains for pre-mRNAs and shuttles between thenucleus and cytoplasm, it is also possible that hnRNP A2/B1 may also function through the mechanism that regu-lates the biogenesis, localization, and metabolism of cer-tain mRNAs (20–23). A recent study demonstrated thathnRNP A2 not only selectively binds but also remains as-sociated with the cytoplasmic transport sequence of mye-lin basic protein mRNA (22). In oogenesis and embryonicdevelopment, hnRNP family members, including hnRNPA2/B1, selectively target mRNAs encoding many essentialmorphogenic proteins to different subcellular domainsand other proteins, and mislocalization of these mRNAsresults in aberrant embryogenesis (23). For all these rea-sons, further evaluation of the biology of hnRNP A2/B1related to carcinogenesis is warranted.
Acknowledgments: The authors are very grateful to Dr. Jill Siegfried, Univer-sity of Pittsburgh Cancer Center (Pittsburgh, PA), for providing the non-neo-plastic tissue samples. The authors appreciate the assistance of Drs. Zhang-PingZhuang of the National Cancer Institute and Farid Moifar of the Armed ForcesInstitute of Pathology in the morphologic review of the tissues selected for usein this study.
References
1. Greenlee, R. T., T. Murray, S. Bolden, and P. A. Wing. 2000. Cancer statis-tics, 2000. CA Cancer J. Clin. 50:7–33.
2. Szabo, E., and J. L. Mulshine. 1996. Lung cancer prevention: historical andfuture perspectives. In Comprehensive Textbook of Thoracic Oncology. J.Aisner, R. Arrigada, and M. R. Green, editors. Williams & Wilkins, Balti-more. 90–102.
3. Tockman, M. S., P. K. Gupta, J. D. Myers, J. K. Frost, S. B. Baylin, E. B.Gold, A. M. Chase, P. H. Wilkinson, and J. L. Mulshine. 1988. Sensitiveand specific monoclonal antibody recognition of human lung cancer anti-gen on preserved sputum cells: a new approach to early lung cancer detec-tion. J. Clin. Oncol. 6:1685–1693.
4. Zhou, J., J. L. Mulshine, E. J. Unsworth, I. Avis, F. Cuttitta, and A. Treston.1996. Purification and characterization of a protein that permits early de-tection of lung cancer. Identification of the heterogeneous nuclear ribonu-cleoprotein-A2/B1 as the antigen for monoclonal antibody 703D4. J. Biol.Chem. 271:10760–10766.
5. Tockman, M. S., J. L. Mulshine, S. Piantadosi, Y. S. Erozan, P. K. Gupta,

Man, Martinez, Avis, et al.: hnRNP Expression and Genetic Alteration 645
J. C. Ruckdeschel, P. R. Taylor, T. Zhikov, W. H. Zhou, Y. L. Qiao, andS. X. Yao, Lung cancer early detection working group investigators, andYunan Tin company investigators. 1997. Prospective detection of preclini-cal lung cancer: results from two studies of hnRNP overexpression. Clin.Cancer Res. 3:2237–2246.
6. Montuenga, L. M., J. Zhou, I. Avis, M. Vos, A. Martinez, F. Cuttitta, A. M.Treston, M. Sunday, and J. L. Mulshine. 1998. Expression of heteroge-neous nuclear ribonucleoprotein A2/B1 changes with critical stages ofmammalian lung development. Am. J. Respir. Cell Mol. Biol. 19:554–562.
7. Zhou, J., S. M. Jensen, S. M. Steinberg, J. L. Mulshine, and R. I. Linnoila.1996. Expression of early lung cancer detection marker p31 in neoplasticand non-neoplastic respiratory epithelium. Lung Cancer 14:85–97.
8. Zhou, J., J. L. Mulshine, J. Y. Ro, I. Avis, R. Yu, J. J. Lee, R. Morice, S. M.Lippman, and J. S. Lee. 1998. Expression of heterogeneous nuclear ribo-nucleoprotein A2/B1 in bronchial epithelium of chronic smokers. Clin.Cancer Res. 4:1631–1640.
9. Nelson, H. H., J. K. Wiencke, L. Gunn, J. C. Wain, and D. C. Christiani.1998. Chromosome 3p14 alterations in lung cancer: evidence that FHITexon deletion is a target of tobacco carcinogens and asbestos. Cancer Res.58:1804–1807.
10. Field, J. K., E. M. Neville, M. P. Stewart, A. Swift, T. Liloglou, J. M. Risk,H. Ross, J. R. Gosney, and R. J. Donelly. 1996. Fractional allele loss dataindicate distinct genetic populations in the development of non-small-celllung cancer. Br. J. Cancer 74:1968–1974.
11. Wistuba, I. I., S. Lam, C. Behrens, A. K. Virmani, K. M. Fong, J. LeRiche,J. M. Samet, S. Srivastava, J. D. Minna, and A. F. Gazdar. 1997. Moleculardamage in the bronchial epithelium of current and former smokers. J.Natl. Cancer Inst. 89:1366–1373.
12. Hittelman, W. N., H. J. Kim, J. S. Lee, D. M. Shin, and S. M. Lippman. 1996.Detection of chromosome instability of tissue fields at risk: in situ hybrid-ization. J. Cell. Biochem. (Suppl) 25:57–62.
13. Froudarakis, M. E., G. Sourvinos, P. Fournel, D. Bouros, J. M. Vergnon, D. A.Spandidos, and N. M. Siafakas. 1998. Microsatellite instability and loss ofheterozygosity at chromosomes 9 and 17 in non-small lung cancer. Chest113:1091–1094.
14. Bepler, G., K. M. Fong, B. E. Johnson, K. C. O’Brian, A. Daly, P. V. Zim-merman, M. A. Garcia-Blanco, and B. Peterson. 1998. Association ofchromosome 11 locus D11S12 with histology, stage, and metastases in lungcancer. Cancer Detect. Prev. 22:14–19.
15. Chung, G. T., V. Sundaresan, P. Hasleton, R. Rudd, R. Taylor, and P. H.Rabbitts. 1996. Clonal evolution of lung tumors. Cancer Res. 56:1609–1614.
16. Mooney, E. E., Y. G. Man, G. L. Bratthauer, and F. A. Tavassoli. 1999. Ev-idence that Leydig cells in Sertoli-Leydig cell tumors have a reactiverather than a neoplastic molecular profile. Cancer 86:2312–2319.
17. Lyon, M. F. 1988. The William Allan Memorial Award address: X-chromo-some inactivation and the location and expression of X-linked genes. Am.J. Hum. Genet. 42:8–16.
18. Vogelstein, B., E. R. Fearon, and S. R. Hamilton. 1987. Clonal analysis us-
ing recombinant DNA probes form the X-chromosome. Cancer Res. 47:4806–4813.
19. Allen, R. C., H. Y. Zoghbi, A. B. Moseley, H. M. Rosenblatt, and J. W. Bel-mont. 1992. Methylation of Hpa II and Hha 1 sites near the polymorphicCAG repeat in the human androgen receptor gene correlates with X-chro-mosome inactivation. Am. J. Hum. Genet. 51:1229–1239.
20. Weighardt, F., G. Biamonti, and S. Riva. 1996. The roles of heterogeneousribonucleoproteins (hnRNP) in RNA metabolism. Bioassays 18:747–756.
21. McKay, S. J., and H. Cooke. 1992. hnRNP A2/B1 binds specifically to singlestranded vertebrate telomeric repeat TTAGGGn. Nucleic Acids Res. 20:6461–6464.
22. Hoek, K. S., G. L. Kidd, J. H. Carson, and R. Smith. 1998. hnRNP A2 selec-tively binds the cytoplasmic transport sequence of myelin basic proteinmRNA. Biochemistry 37:7021–7029.
23. Pokrywka, N. J. 1995. RNA localization and cytoskeleton in Drosophila oo-cytes. Curr. Top. Dev. Biol. 31:864–876.
24. Moinfar, F., Y. G. Man, L. Arnould, G. L. Bratthauer, M. Ratschek, and F. A.Tavassoli. 2000. Current and independent genetic alterations in the stro-mal and epithelial cells of mammary carcinoma: implication for carcino-genesis. Cancer Res. 60:2562–2566.
25. Man, Y. G., F. Moinfar, G. L. Bratthauer, and F. A. Tavassoli. 2000. Fiveuseful approaches for generating more valid gel image of LOH and clonal-ity analysis with an automated 377 DNA sequencer. Mol. Diagn. Pathol.9:84–90.
26. Mao, L., J. S. Lee, J. M. Kurie, Y. H. Fan, S. M. Lippman, W. Hittelman,and W. K. Hong. 1997. Clonal genetic alterations in the lungs of currentand former smokers. J. Natl. Cancer Inst. 89:857–862.
27. Hung, J., Y. Kishimoto, K. Sugio, A. Virmani, D. D. McIntire, J. D. Minna,and A. F. Gazdar. 1995. Allele specific chromosome 3p deletions occur atan early stage in the pathogenesis of lung carcinoma. JAMA 273:558–563.
28. Field J. K., T. Liloglou, G. Xinarianos, W. Prime, and P. Fielding. 1999. Ge-netic alterations in bronchial lavage as a potential marker for individualswith a high risk of developing lung cancer. Cancer Res. 59:2690–2695.
29. Moinfar, F., Y. G. Man, G. L. Brathauer, and F. A. Tavassoli. 2000. Geneticabnormality in ductal intraepithelial neoplasia of the flat type (clingingcarcinoma in situ) of the breast. Cancer 88:2072–2081.
30. Hanahan, D., and R. A. Weinberg. 2000. The hallmarks of cancer. Cell 100:57–70.
31. Churchill, L., R. H. Gundel, L. G. Letts, and C. D. Wagner. 1993. Contribu-tion of specific cell-adhesive glycoproteins to airway and alveolar inflam-mation and dysfunction. Am. Rev. Respir. Dis. 148:S83–S88.
32. LaBranche, H., S. Dupuis, Y. Ben-David, M. R. Bani, R. J. Wellinger, andB. Chabot. 1998. Telomere elongation by hnRNP A1 and a derivative thatinteracts with telomeric repeats and telomerase. Nat. Genet. 19:199–202.
33. Pontius, B. W., and P. Berg. 1990. Renaturation of complementary DNAstrands mediated by purified mammalian heterogeneous nuclear ribonu-cleoprotein A1 protein: implications for a mechanism for rapid molecularassembly. Proc. Natl. Acad. Sci. USA 87:8403–8407.
Related Documents











