J. Comp. Path. 1999 Vol. 120, 177–186 Phenotypic Characterization of Histiocytes Infiltrating a Leiomyofibrosarcoma A. Whyte, G. Rowlinson-Busza*, A. Dorling and R. White Department of Immunology, The Babraham Institute, Cambridge, CB2 4AT, *Imperial Cancer Research Fund Oncology Unit, Department of Clinical Oncology, Hammersmith Hospital, Du Cane Road, London W12 0HS and Department of Immunology, Imperial College of Science, Technology and Medicine, Hammersmith Hospital, Du Cane Road, London W12 0HS, UK Summary We described previously a unique cutaneous tumour in a young pig, which was characterized by several criteria as a histiocytic leiomyofibrosarcoma. The lipid-laden macrophages (histiocytes) which permeated the tumour were CD2 + /CD18 + /CD49d + but MAC387 (L1 antigen) and CD15 negative. The present study compared the phenotypes of histiocytes in tumour metastases in the liver with resident liver macrophages, revealing di erential expression of certain macrophage activation markers. After repeated subcutaneous pas- sage of the tumour in athymic (nu/nu) mice, flow cytometry demonstrated a rapid loss of porcine MHC Class II, but a more prolonged expression of porcine MHC Class I, consistent with our immunohistological observations. Mouse macrophages (CD2 + /F4·80 + ) infiltrated the later-passage tumours, suggesting that the histiocytes were not of neoplastic origin. 1999 W.B. Saunders Company Limited Introduction Tumours of smooth-muscle origin (leiomyomas and leiomyosarcomas) are rare in man (Fields and Helwig, 1981), and the report by Whyte et al. (1996b) appears to be the only one of such a tumour in the pig. This pig tumour was extensively infiltrated with lipid-laden macrophages (histiocytes), but it was not known whether these were of neoplastic origin or arose from the fibroblastic lineage that is also a feature of this tumour. In malignant fibrous histiocytoma (MFH), the most common soft-tissue sarcoma in human adults, histiocytes are intermingled with fibroblasts, but the origin of the histiocytes remains unresolved. The porcine histiocyte leiomyofibrosarcoma was predominantly subcutaneous in its distribution, but metastases occurred in various organs in lower numbers. The present study focused on liver metastases because these permitted direct comparison of the phenotype of the tumour-resident histio- cytes with that of the surrounding macrophages of the liver. Materials and Methods Tumour and Animals The origin and description of the tumour were reported previously by Whyte et al. (1996b). The mice used in the present studies were female athymic (nu/nu) animals 0021–9975/99/020177+10 $12.00/0 1999 W.B. Saunders Company Limited

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J. Comp. Path. 1999 Vol. 120, 177–186
Phenotypic Characterization of HistiocytesInfiltrating a Leiomyofibrosarcoma
A. Whyte, G. Rowlinson-Busza∗, A. Dorling† and R. WhiteDepartment of Immunology, The Babraham Institute, Cambridge, CB2 4AT,
∗Imperial Cancer Research Fund Oncology Unit, Department of Clinical Oncology,Hammersmith Hospital, Du Cane Road, London W12 0HS and
†Department of Immunology, Imperial College of Science, Technology and Medicine,Hammersmith Hospital, Du Cane Road, London W12 0HS, UK
Summary
We described previously a unique cutaneous tumour in a young pig, whichwas characterized by several criteria as a histiocytic leiomyofibrosarcoma.The lipid-laden macrophages (histiocytes) which permeated the tumour wereCD2+/CD18+/CD49d+ but MAC387 (L1 antigen) and CD15 negative. Thepresent study compared the phenotypes of histiocytes in tumour metastasesin the liver with resident liver macrophages, revealing differential expressionof certain macrophage activation markers. After repeated subcutaneous pas-sage of the tumour in athymic (nu/nu) mice, flow cytometry demonstrated arapid loss of porcine MHC Class II, but a more prolonged expression ofporcine MHC Class I, consistent with our immunohistological observations.Mouse macrophages (CD2+/F4·80+) infiltrated the later-passage tumours,suggesting that the histiocytes were not of neoplastic origin.
1999 W.B. Saunders Company Limited
Introduction
Tumours of smooth-muscle origin (leiomyomas and leiomyosarcomas) are rarein man (Fields and Helwig, 1981), and the report by Whyte et al. (1996b)appears to be the only one of such a tumour in the pig. This pig tumour wasextensively infiltrated with lipid-laden macrophages (histiocytes), but it wasnot known whether these were of neoplastic origin or arose from the fibroblasticlineage that is also a feature of this tumour. In malignant fibrous histiocytoma(MFH), the most common soft-tissue sarcoma in human adults, histiocytesare intermingled with fibroblasts, but the origin of the histiocytes remainsunresolved. The porcine histiocyte leiomyofibrosarcoma was predominantlysubcutaneous in its distribution, but metastases occurred in various organs inlower numbers. The present study focused on liver metastases because thesepermitted direct comparison of the phenotype of the tumour-resident histio-cytes with that of the surrounding macrophages of the liver.
Materials and Methods
Tumour and Animals
The origin and description of the tumour were reported previously by Whyte et al.(1996b). The mice used in the present studies were female athymic (nu/nu) animals
0021–9975/99/020177+10 $12.00/0 1999 W.B. Saunders Company Limited

178 A. Whyte et al.
of mixed genetic background. They were bred under specific pathogen-free conditionsat the Imperial Cancer Research Fund Animal Breeding Unit, South Mimms, Hert-fordshire, and subsequently housed in sterile filter-top cages with sterile bedding, andmaintained on an irradiated diet and autoclaved acidified water (pH 2·8).
The primary pig tumour and its subsequent mouse passages were chopped intopieces (c. 2 mm3) in sterile tissue-culture medium (RPMI 1640 containing fetal calfserum 10%, penicillin 100 U/ml and streptomycin 100 lm/ml; Flow Labs, Irvine,Scotland). Recipient mice were anaesthetized by methoxyfluorane inhalation (Meto-fane; C-Vet, Bury St Edmunds, Suffolk). Under aseptic conditions, a small (5 mm)incision was made in the skin on the flank of the mouse. Forceps were used to createa subcutaneous channel by blunt dissection into which the tumour fragment wasinserted, and the incision was closed with a 7·5-mm Michel clip. The clips wereremoved after one week and the tumour was allowed to grow to a diameter ofapproximately 1 cm before further passaging. This took about 2 months. At eachmouse to mouse passage, one sample of tumour was fixed in formol-saline and asecond sample was frozen in isopentane in liquid nitrogen and stored at −80°C. Thetumour was maintained for nine passages, showing no evidence of a reduced growthrate. The first passage was designated P0, and subsequent passages were designatedP1 to P8.
Immunohistochemistry and Antibodies
Immunohistochemistry was performed, as detailed elsewhere (Whyte et al., 1996a),on cryosections of pig liver tumour metastases, passaged nodules from nu/nu miceand pig cutaneous tumour lesions. The antibodies used are listed in Table 1; theirreactivities with skin and tumour metastases in the skin collected in this study arealso shown. Macrophage-specific antibodies were obtained from Dako (HighWycombe, Bucks) (M633, which reacts with macrophages in atherosclerotic lesions)and BMA Biomedicals (Augst, Switzerland). Those from BMA Biomedicals, whichwere a kind gift from Dr G. Burmeister, included 2·5F9 (late-stage inflammation andtumour marker) and PM2K (which reacts with an epitope present on macrophagesin various tissues but absent from monocytes and dendritic cells [Takeya et al., 1991];it may also recognize the macrophage scavenger receptor). Polyclonal anti-L1 antigen(MAC387) was supplied by Dako, and mouse mature macrophage marker (F4.80) bySerotec, UK. The anti-mouse CD2 antibodies were a kind gift from Dr S. Cobbold,University of Oxford. All other antibodies listed in Table 1 were produced in-houseand as detailed elsewhere (Whyte et al., 1996a,b). Monoclonal antibodies (mAbs) tothe macrophage L1 antigen (clones 8-5c2, S36·48 and S32·3), to lysozyme (cloneA099), to CD68 (clones EBM11 and PGM-1) and to CD16 (clones DJ130c, B88-9and Gran-1) were also tested but did not cross-react with pig determinants.
In-vitro Culture and Flow Cytometry (FACS) Analysis
Cells from the primary tumor or nu/nu passages were harvested into RPMI 1640medium containing fetal calf serum 10%, penicillin 50 U/ml, streptomycin 50 lg/mland 2 mM L-glutamine (Flow Labs, Irvine, Scotland). The tissue was chopped withcrossed scalpels into fragments (2 mm3), which were suspended in 75-ml2 flasks (Nunc)containing CMRL 1066 medium (Flow Labs), fetal calf serum 10%, penicillin 50 U/ml and streptomycin 50 lg/ml. From the fragments, single cells floated into suspensionand others (with a fibroblastic morphology) became adherent to the plastic andsurrounded by an expanding monolayer of cells. Only the cells in suspension showedcontinued growth, albeit slow, and only in complete medium (CMRL, not RPMI orDMEM). ‘‘Cytospin’’ preparations of the cells in suspension showed that they hadmorphological characteristics typical of histiocytes. When they first grew in suspension,1–5×105 cells were treated in 96-well plates (Sterilin, UK) with clone 74.11.10 (anti-pig Class I MHC) or clone MSA-3 (anti-class II MHC, SLA-DRw) supernates at
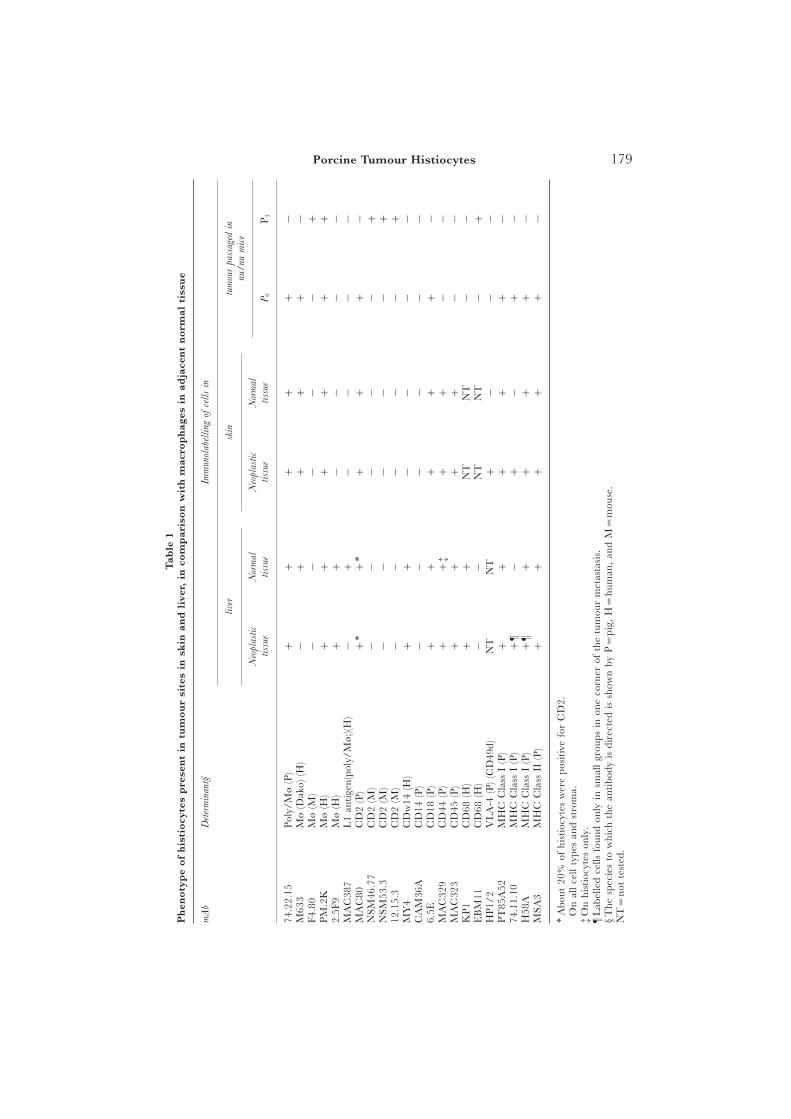
179Porcine Tumour HistiocytesT
able
1P
hen
oty
pe
of
his
tio
cyte
sp
rese
nt
intu
mo
ur
site
sin
skin
and
live
r,in
com
par
iso
nw
ith
mac
rop
hag
esin
adja
cen
tn
orm
alti
ssu
e
mA
bD
eter
min
ant§
Imm
unol
abel
ling
ofce
lls
in
live
rsk
intu
mou
rpa
ssag
edin
nu/
num
ice
Neo
plas
tic
Nor
mal
Neo
plas
tic
Nor
mal
tiss
ueti
ssue
tiss
ueti
ssue
P0
P3
74.2
2.15
Pol
y/M
ø(P
)+
++
++
−M
633
Mø
(Dak
o)(H
)−
++
++
−F4
.80
Mø
(M)
−−
−−
−+
PM
.2K
Mø
(H)
++
++
++
2.5F
9M
ø(H
)+
+−
−−
−M
AC
387
L1
anti
gen(
poly
/Mø;
)(H)
−+
−−
−−
MA
C80
CD
2(P
)+∗
+∗
++
+−
NSM
46.7
7C
D2
(M)
−−
−−
−+
NSM
53.3
CD
2(M
)−
−−
−−
+12
.15.
3C
D2
(M)
−−
−−
−+
MY
4C
Dw
14(H
)+
+−
−−
−C
AM
36A
CD
14(P
)−
−−
−−
−6.
5EC
D18
(P)
++
++
+−
MA
C32
9C
D44
(P)
+†
+‡
++
−−
MA
C32
3C
D45
(P)
++
++
−−
KP
1C
D68
(H)
++
NT
NT
−−
EB
M11
CD
68(H
)−
−N
TN
T−
+H
P1/
2V
LA
-4(P
)(C
D49
d)N
TN
T+
−−
−P
T85
A52
MH
CC
lass
I(P
)+
++
++
−74
.11.
10M
HC
Cla
ssI
(P)
+¶
−+
−+
−H
58A
MH
CC
lass
I(P
)+
¶+
++
+−
MSA
3M
HC
Cla
ssII
(P)
++
++
+−
∗A
bout
20%
ofhi
stio
cyte
sw
ere
posi
tive
for
CD
2.†
On
all
cell
type
san
dst
rom
a.‡
On
hist
iocy
tes
only
.¶
Lab
elle
dce
llsfo
und
only
insm
all
grou
psin
one
corn
erof
the
tum
our
met
asta
sis.
§T
hesp
ecie
sto
whi
chth
ean
tibo
dyis
dire
cted
issh
own
byP=
pig,
H=
hum
an,
and
M=
mou
se.
NT=
not
test
ed.

180 A. Whyte et al.
appropriate dilutions (1 in 50–1 in 500) for 15–20 min at 4°C. The cells were washedtwice in phosphate-buffered saline containing fetal calf serum 2·5% and incubatedfor 15–20 min at 4°C with sheep anti-mouse (1 in 300) (Sigma, UK) fluoresceinisothiocyanate (FITC)-conjugated second antibody. The cells were then washed, fixedin 1% paraformaldehyde and kept at 4°C in the dark until analysed (within oneweek). FACS analysis was performed on an EPICS XL Flow Cytometer (Coulter).
Results
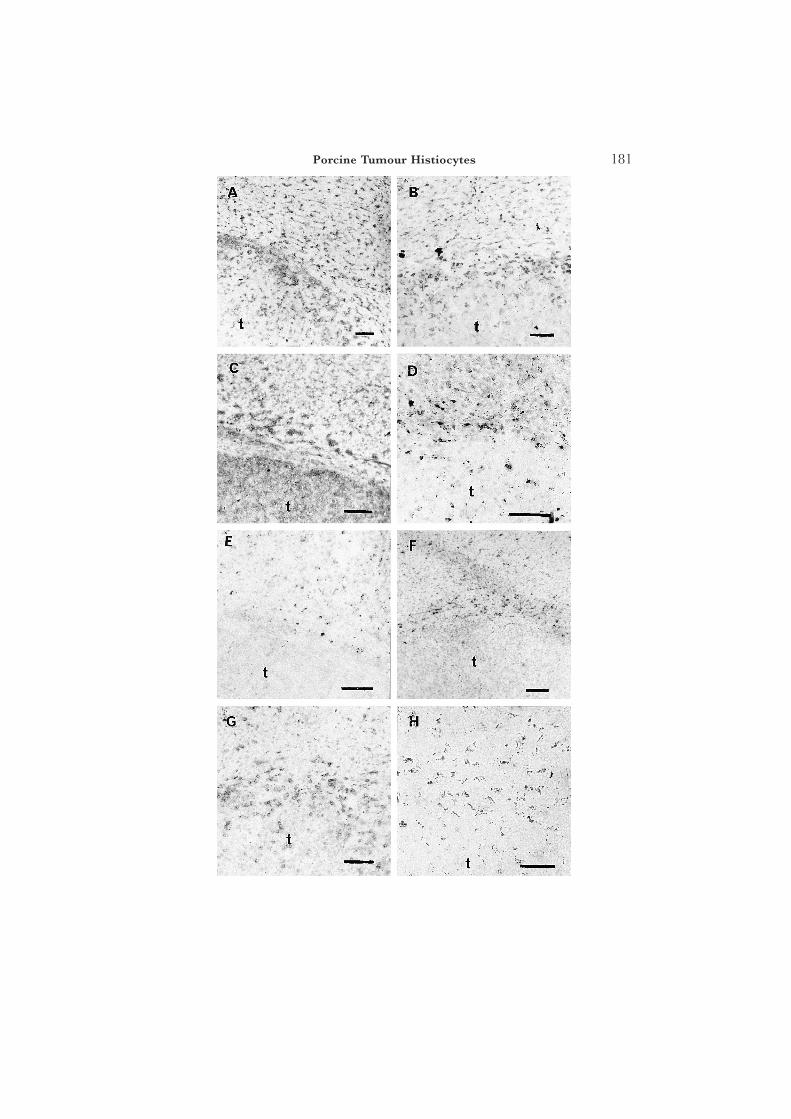
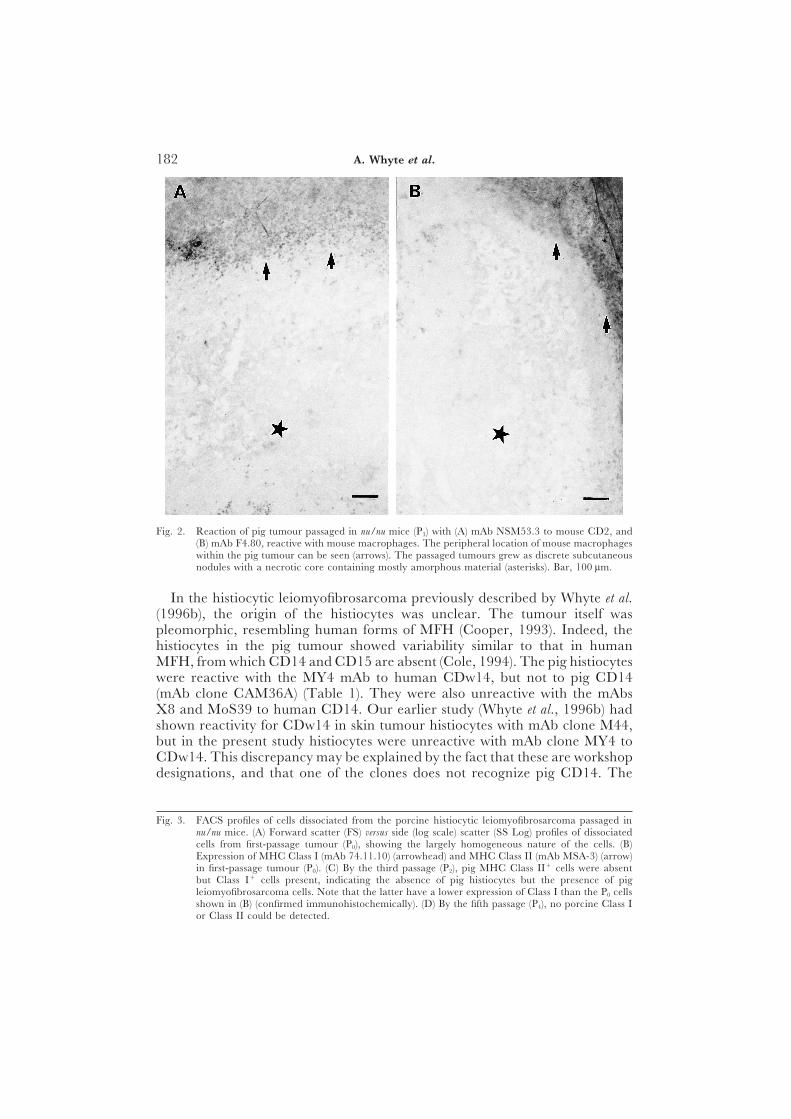
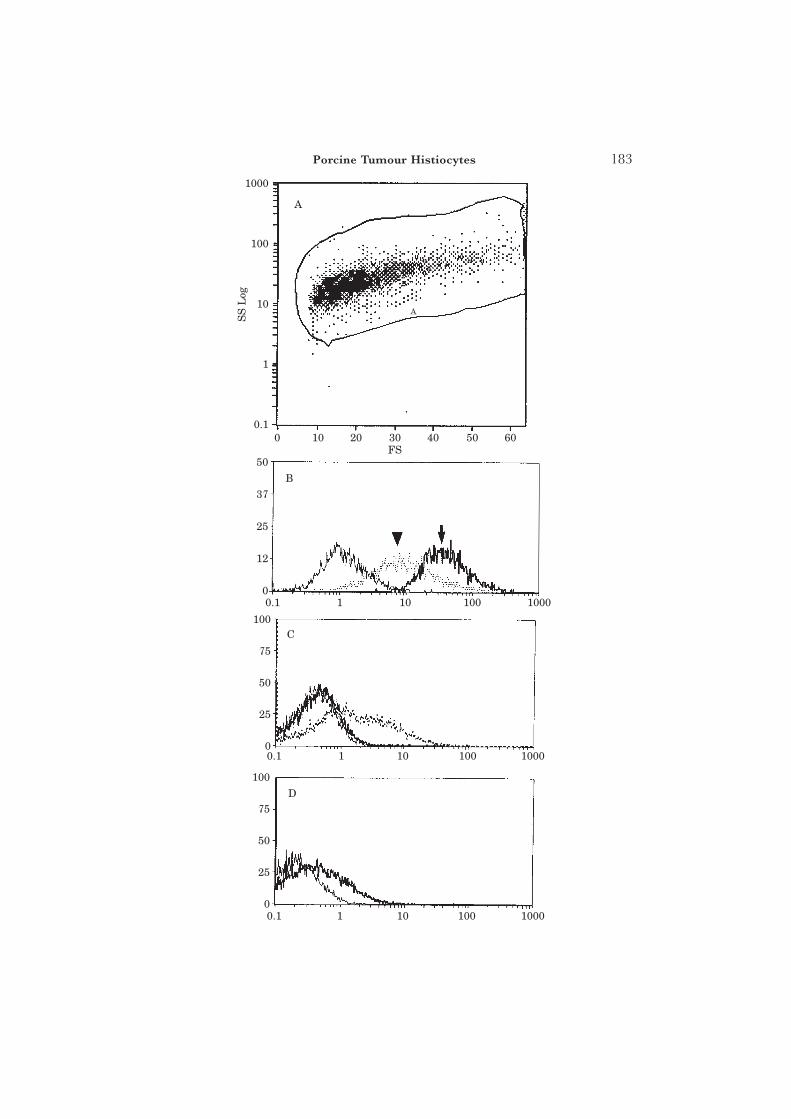
The immunohistological profile of the tumour, together with that of pig liverand skin, and passaged mouse tumour (first passage [P0] and fourth passage[P3]) with a wide range of antibodies is summarized in Table 1. The histiocytesin both skin and liver metastases were reactive for 74.22.15 (an accessory cellmarker), CD2 (about 20% of all macrophages were positive), CD44, CD45and pig (SLA) MHC Class I. Those in the liver metastases, but not in the skinmetastases, were additionally reactive for 2·5F9 and CDw14 (Fig. 1). Thetumour passaged in the nu/nu mice showed the presence of pig macrophages(74.22.15) at P0. Also, surprisingly, it showed the presence of mouse macro-phages [F4.80, EBM11 [CD68], and mouse CD2 at later passages (Fig. 2);the antibodies concerned did not recognize pig macrophages]. The mousemacrophages did not become apparent until later passages ([P3) of thetumour, by which time pig MHC Class II-reactive cells had disappeared andfew pig MHC Class I-positive cells remained (Fig. 3).
Discussion
The origins of the histiocytes that occur in various tumours remain a subjectof debate. Even in malignant fibrous histiocytoma (MFH), in which they area component of the tumour, they have been variously described as of histiocyticcell origin, of undifferentiated mesenchymal cell origin, or of fibroblast origin(Cooper, 1993). Both 9,10-dimethyl-1,2-benzanthracene (DMBA) (Kato et al.,1990) and methylcholanthrene (Katenkamp and Stiller, 1975) have been usedin rodents to induce neoplastic changes resembling MFH. A chemically-induced rat MFH, when transplanted into nu/nu mice, became progressivelypopulated with mouse macrophages, suggesting a ‘‘passenger’’ role for histio-cytes in MFH (Tsuchiya et al., 1993).
Fig. 1. Immunolabelling of histiocytes by a variety of antibodies within the pig tumour (t) in livermetastases. Adjacent (normal) liver tissue occupies the top portion of each picture. (A) CD45(clone MAC323) delineates all leucocytes. (B) The accessory cell marker, mAb clone 74.22.15,shows cells of granulocyte and macrophage lineage in the pig. (C) MHC Class I (mAb PT85A) inboth tumour and liver. The expression of MHC Class II (mAb MSA-3, SLA-DRw) was similar.(D) Antibody 2.5F9 recognized macrophages in both tumour and liver (but not the histiocytes inthe skin metastases; Table 1). (E) Conversely, liver but not tumour macrophages were reactivewith the M633 mAb. (F) The tumour, but not the liver, macrophages were negative for MAC387.Note here, as elsewhere, the accumulation of macrophages apparently within liver and at theborder of the well-circumscribed metastasis. (G) CD2 (MAC80) was present on both macrophagepopulations. (H) CDw14 (clone MY4) reacted less with tumour than liver macrophages (but notat all with the tumour histiocytes in skin metastases; Table 1). All scale bars are 100 lm except(H) where it represents 50 lm.
181Porcine Tumour Histiocytes
182 A. Whyte et al.
Fig. 2. Reaction of pig tumour passaged in nu/nu mice (P3) with (A) mAb NSM53.3 to mouse CD2, and(B) mAb F4.80, reactive with mouse macrophages. The peripheral location of mouse macrophageswithin the pig tumour can be seen (arrows). The passaged tumours grew as discrete subcutaneousnodules with a necrotic core containing mostly amorphous material (asterisks). Bar, 100 lm.
In the histiocytic leiomyofibrosarcoma previously described by Whyte et al.(1996b), the origin of the histiocytes was unclear. The tumour itself waspleomorphic, resembling human forms of MFH (Cooper, 1993). Indeed, thehistiocytes in the pig tumour showed variability similar to that in humanMFH, from which CD14 and CD15 are absent (Cole, 1994). The pig histiocyteswere reactive with the MY4 mAb to human CDw14, but not to pig CD14(mAb clone CAM36A) (Table 1). They were also unreactive with the mAbsX8 and MoS39 to human CD14. Our earlier study (Whyte et al., 1996b) hadshown reactivity for CDw14 in skin tumour histiocytes with mAb clone M44,but in the present study histiocytes were unreactive with mAb clone MY4 toCDw14. This discrepancy may be explained by the fact that these are workshopdesignations, and that one of the clones does not recognize pig CD14. The
Fig. 3. FACS profiles of cells dissociated from the porcine histiocytic leiomyofibrosarcoma passaged innu/nu mice. (A) Forward scatter (FS) versus side (log scale) scatter (SS Log) profiles of dissociatedcells from first-passage tumour (P0), showing the largely homogeneous nature of the cells. (B)Expression of MHC Class I (mAb 74.11.10) (arrowhead) and MHC Class II (mAb MSA-3) (arrow)in first-passage tumour (P0). (C) By the third passage (P2), pig MHC Class II+ cells were absentbut Class I+ cells present, indicating the absence of pig histiocytes but the presence of pigleiomyofibrosarcoma cells. Note that the latter have a lower expression of Class I than the P0 cellsshown in (B) (confirmed immunohistochemically). (D) By the fifth passage (P4), no porcine Class Ior Class II could be detected.
183Porcine Tumour Histiocytes
1000
100
10
1
0.1
50
37
25
12
0
100
75
50
25
0
D
C
B
A
10001001010.1
100
75
50
25
0
10001001010.1
10001001010.1
6050403020100
A
SS
Log
FS

184 A. Whyte et al.
pig tumour histiocytes also lacked expression of CD15 ([clone DU-HL60-3;Serotec], which recognizes pig CD15 [Whyte and Binns, 1994]). The pigtumour would therefore appear to possess histiocytes of a phenotype similarto those present in many cases of human MFH; even in the latter, however,a great deal of phenotypic diversity is present (Roholl et al., 1988). We areaware of only one report of MFH in the pig (Tanimoto et al., 1988), and it isnot clear why MFH is not observed with as great a frequency in pigs as inman; possible explanations include under-reporting and misdiagnosis. It isalso possible that human immune surveillance mechanisms differ from thosein domestic animals; indeed, unique phenotypes of lymphocytes are associatedwith neoplasms of human skin (Picker et al., 1990). MFH is rarely seenin domestic animals, including cats and dogs (Thomas, 1988). Cutaneoushistiocytoma, however, is a common benign neoplasm of dogs (Moore et al.,1996), but the origin of the histiocytes in such tumours would appear to beLangerhans cells rather than macrophages (Marchal et al., 1995).
Mouse macrophages were present in the tumour after three or more passagesin nu/nu mice (Fig. 2) and pig histiocytes (MHC Class II+) were absent afterthe first passage (Fig. 3). This pattern is similar to that observed in therepopulation of chemically-induced rat MFH by mouse histiocytes aftertransplantation into nu/nu recipients (Tsuchiya et al., 1993), indicating thatthe pig tumour has an ability to attract host macrophages. Once attracted,however, it would appear that these histiocytes differentiate along specificpathways, unlike macrophages in other sites, even in adjacent host tissue (e.g.,skin and liver; Table 1). Thus, within the various tumour sites in the pig, thehistiocytes expressed a variety of phenotypes. In the tumour metastases in theliver they were M633−/PM·2K+/2·5F9+/L1−, whereas in the skin tumoursites they were M633+/PM·2K+/2·5F9−/L1−. In man the PM·2K mAbrecognizes a tissue macrophage antigen (Takeya et al., 1991), and mAb 2·5F9recognizes a determinant (absent from monocytes) on late-stage inflammatorymacrophages. Liver macrophages expressed the M633 and L1 determinants,in contrast to the liver metastasis histiocytes. Skin and liver phenotypes weregenerally comparable, apart from the expression of the 2·5F9 determinant,and to a lesser extent the L1 antigen. Histiocytes in both tumour sites, togetherwith adjacent macrophages, did not express the S13-67 determinant, presenton the 8 kD subunit of the inflammatory L1 protein complex. However, thisdeterminant could be induced in pig skin macrophages by intradermal injectionof recombinant tumour necrosis factor (TNF)-a and interleukin (IL)-1a (datanot shown).
The phenotypic heterogeneity of both tumour and resident histiocyteswas apparent, indicating different degrees of activation dependent uponthe location. When transplanted into nu/nu mice, the porcine histiocyticleiomyofibrosarcoma initially retained its porcine characteristics (Table 1), butby the third passage the MHC Class II+ pig histiocytes had disappeared (Fig.3), and by the fourth passage mouse macrophages had infiltrated the tumourin large numbers (Fig. 2 and Table 1). There was also a progressive deteriorationof the tumour on repeated passage in nu/nu mice, with a loss of pig MHC

185Porcine Tumour Histiocytes
Class I (Fig. 3) and increasingly large necrotic central foci in the passagednodules (Fig. 2).
In transplanted rat MFH, the concentrations of the chemokine MCP-1(monocyte chemoattractant protein-1) showed a positive correlation with thedegree of infiltration of macrophages (Yamashiro et al., 1994), and it wasconcluded that MCP-1 induced infiltration of monocyte-derived macrophages,but not macrophages of the resident type (histiocytes). The immunophenotypeof the histiocytes in the pig leiomyofibrosarcoma was at variance with theresults for rat MFH, suggesting that MCP-1 was not the major agent responsiblefor recruitment of macrophages into the tumour. In human MFH cell lines,Takeya et al. (1995) concluded that histiocytes were not of neoplastic originbut rather represented a population of infiltrated macrophages. This conclusionis consistent with the results for the porcine leiomyofibrosarcoma; it is clear,however, that in the latter the infiltrating macrophages differentiated along adifferent pathway. Thus, although the pig histiocytes resembled human MFHhistiocytes in lacking CD15 and expressing CD68 (Takeya et al., 1995), theydiffered in that they were positive for CD45, MHC Class II and PM-2K.Definitive confirmation of the presence or absence of CD14 will requireimmunoprecipitation studies. It is also possible that MFH cell lines in cultureshow dedifferentiation of the macrophage population, losing markers expressedin the in-vivo tumour, due probably to the absence of the appropriatechemokine profile in vitro.
In conclusion, substantial phenotypic heterogeneity was demonstrated inhistiocytes within the porcine leiomyofibrosarcoma. These histiocytes appearednot to be a neoplastic component of the tumour, because after repeatedpassages in nu/nu mice they were replaced by mouse macrophages. However,although they were attracted into the tumour, they differentiated along uniquepathways, probably controlled by chemokines produced by the neoplasticcells. Their behaviour resembled that of histiocytes in many forms of MFH,in which histiocytes (1) appear to have a ‘‘passenger’’ role in a tumour offibroblastic origin, and (2) differentiate along variable pathways. It is possiblethat some tumours diagnosed in the past as MFH were in fact tumours offibroblastic origin with a non-neoplastic histiocytic component.
References
Cole, C. H. (1994). Malignant fibrous histiocytoma. Cancer Journal, 7, 63–66.Cooper, C. S. (1993). The molecular and genetic characterization of human soft tissue
tumors. Advances in Cancer Research, 60, 75–120.Fields, J. P. and Helwig, E. B. (1981). Leiomyosarcomas of the skin and subcutaneous
tissues. Cancer, 47, 156–169.Katenkamp, D. and Stiller, D. (1975). Structural patterns and histological behaviour
of experimental sarcomas: general considerations, histology and histochemistry.Experimental Pathology, 11, 182–189.
Kato, T., Takeya, M., Takagi, K. and Takahashi, K. (1990). Chemically inducedtransplantable malignant fibrous histiocytoma. Laboratory Investigation, 62, 635–645.
Marchal, T., Dezutter-Dambuyant, C., Fournel, C., Magnol, J. P. and Schmitt, D.(1995). Immunophenotypic and ultrastructural evidence of the Langerhans cellorigin of the canine cutaneous histiocytoma. Acta Anatomica (Basel), 153, 189–202.
Moore, P. F., Schrenzel, M. D., Affolter, V. K., Olivry, T. and Naydan, D. (1996).

186 A. Whyte et al.
Canine cutaneous histiocytoma is an epidermotropic Langerhans cell histiocytosisthat expresses CD1 and specific b2-integrin molecules. American Journal of Pathology,148, 1699–1708.
Picker, L. J., Michie, S. A., Rott, L. S. and Butcher, E. C. (1990). A unique phenotypeof skin-associated lymphocytes in humans. American Journal of Pathology, 136,1053–1068.
Roholl, P. J. M., Kleyne, J., Prins, M. E. F., Hooijkaas, H., Vroom, T. M. and vanUnnik, J. A. M. (1988). Immunologic marker analysis of normal and malignanthistiocytes. A comparative study of monoclonal antibodies for diagnostic purposes.American Journal of Clinical Pathology, 89, 187–194.
Takeya, M., Tsuchiya, T., Shimokawa, Y. and Takahashi, K. (1991). A new monoclonalantibody, PM-2K, specifically recognizes tissue macrophages but not bloodmonocytes. Journal of Pathology, 163, 315–321.
Takeya, M., Yamashiro, S., Yoshimura, T. and Takahashi, K. (1995). Immuno-phenotypic and immunoelectron microscopic characterization of major con-stituent cells in malignant fibrous histiocytoma using human cell lines andtheir transplanted tumours in immunodeficient mice. Laboratory Investigation, 72,679–688.
Tanimoto, T., Ohtsuki, Y., Sonobe, H., Takahashi, R. and Nomura, Y. (1988).Malignant fibrous histiocytoma in the spleen of a pig. Veterinary Pathology, 25,330–332.
Thomas, J. B. (1988). Malignant fibrous histiocytoma in a dog. Australian VeterinaryJournal, 65, 252–254.
Tsuchiya, T., Takahashi, K., Takeya, M., Hosokawa, Y., Hattori, T. and Takagi, K.(1993). Immunohistochemical, quantitative immunoelectron microscopic, andDNA characterization of chemically induced rat malignant fibrous histiocytoma.American Journal of Pathology, 143, 431–445.
Whyte, A. and Binns, R. M. (1994). Adhesion molecule expression and infiltratingmaternal leucocyte phenotypes during blastocyst implantation in the pig. CellBiology International, 18, 759–766.
Whyte, A., Licence, S. T., Robinson, M. K. and van der Lienden, K. (1996a).Lymphocyte subsets and adhesion molecules in cutaneous inflammation inducedby inflammatory agonists: correlation between E-selectin and cdTcR+ lym-phocytes. Laboratory Investigation, 75, 439–449.
Whyte, A., Ockleford, C. D., Byrne, S., Hubbard, A. and Wooley, S. T. (1996b).Leucocyte and endothelial cell adhesion molecule expression in porcine histiocyticleiomyofibrosarcoma. Journal of Comparative Pathology, 115, 429–440.
Yamashiro, S., Takeya, M., Nishi, T., Karatsu, J., Yoshimura, T., Ushio, Y. andTakahashi, K. (1994). Tumor-derived monocyte chemoattractant protein-1 in-duces intratumoral infiltration of monocyte-derived macrophage subpopulationin transplanted rat tumors. American Journal of Pathology, 145, 856–867.
C Received, June 26th, 1998Accepted, October 5th, 1998D
Related Documents











