Phagocytosis of bacteria is enhanced in macrophages undergoing nutrient deprivation Wim Martinet 1, *, Dorien M. Schrijvers 1, *, Jean-Pierre Timmermans 2 , Arnold G. Herman 1 and Guido R. Y. De Meyer 1 1 Division of Pharmacology, University of Antwerp, Belgium 2 Laboratory of Cell Biology and Histology, University of Antwerp, Belgium Nutrient deprivation triggers a variety of signaling events that enable energy conservation by cells. Among the different nutrient-sensing pathways, it is worth not- ing: (a) AMP-activated protein kinase, a metabolic stress sensor that is stimulated by elevated AMP ⁄ ATP ratios; (b) Per-Arnt-Sim kinase, which acts as a molec- ular sensor of oxygen, redox status, ATP and other indicators of the cellular metabolism; and (c) the hexosamine biosynthetic pathway that produces uri- dine 5¢-diphospho-N-acetylglucosamine as a substrate for O-N-acetylglucosamine transferase [1]. One energy conservation strategy that has attracted much attention is enhanced autophagocytosis, also known as macro- autophagy or simply autophagy [2]. This process is a catabolic pathway involving the engulfment and degra- dation of a cell’s own components through the lyso- somal machinery [3,4]. Autophagic signaling in response to nutrients is mainly relayed through the serine-threonine kinase mammalian target of rapa- mycin (mTOR). Indeed, mTOR is activated by nutri- ent-rich conditions, especially high levels of amino acids and insulin. Blocking mTOR function using rapamycin or its analogs mimics nutrient deprivation and triggers autophagy [1,4]. Keywords autophagy; heterophagy; p38 MAP kinase; scavenger receptor A; starvation Correspondence W. Martinet, Division of Pharmacology, University of Antwerp, Universiteitsplein 1, B-2610 Antwerp, Wilrijk, Belgium Fax: +32 3 820 25 67 Tel: +32 3 820 26 79 E-mail: [email protected] *These authors contributed equally to this work (Received 28 November 2008, revised 28 January 2009, accepted 5 February 2009) doi:10.1111/j.1742-4658.2009.06951.x Phagocytosis represents a mechanism used by macrophages to remove pathogens and cellular debris. Recent evidence suggests that phagocytosis is stimulated under specific conditions of stress, such as extracellular pres- sure and hypoxia. In the present study, we show that amino acid or glucose deprivation caused an increase in the phagocytosis of heat-inactivated Escherichia coli and Staphylococcus aureus by macrophages, but not the uptake of platelets, apoptotic cells or beads. Increased phagocytosis of bac- teria could be blocked by phagocytosis inhibitors and was found to be dependent on p38 mitogen-activated protein kinase activity and scavenger receptor A. Although nutrient deprivation is a strong stimulus of auto- phagy, autophagosome formation was not critical for the uptake of bacte- ria because phagocytic clearance was not inhibited after down-regulation of the autophagy essential gene Atg7. Moreover, enhanced uptake of bacteria should not be considered as a general stress response because phagocytosis of bacteria was not stimulated after exposure of macrophages to the genotoxic agent camptothecin, heat (40 °C) or thapsigargin-induced endo- plasmic reticulum stress. Overall, the results obtained in the present study indicate that nutrient deprivation can stimulate macrophages to fight bacterial infections. Abbreviations AC, apoptotic cell; CHOP, C ⁄ EBP homologous protein; EBSS, Earle’s balanced salt solution; eGFP, enhanced green fluorescent protein; ER, endoplasmic reticulum; LPS, lipopolysaccharide; MAP, mitogen-activated protein; MARCO, macrophage receptor with collagenous structure; mTOR, mammalian target of rapamycin; NDRG1, N-myc downstream-regulated gene 1; PI, propidium iodide; PI3-kinase, phosphoinositide 3-kinase; PLT, platelet; RORa, RAR-related orphan receptor alpha gene; siRNA, small interfering RNA; SR-A, scavenger receptor A; TLR, Toll-like receptor. FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2227

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Phagocytosis of bacteria is enhanced in macrophagesundergoing nutrient deprivationWim Martinet1,*, Dorien M. Schrijvers1,*, Jean-Pierre Timmermans2, Arnold G. Herman1 andGuido R. Y. De Meyer1
1 Division of Pharmacology, University of Antwerp, Belgium
2 Laboratory of Cell Biology and Histology, University of Antwerp, Belgium
Nutrient deprivation triggers a variety of signaling
events that enable energy conservation by cells. Among
the different nutrient-sensing pathways, it is worth not-
ing: (a) AMP-activated protein kinase, a metabolic
stress sensor that is stimulated by elevated AMP ⁄ATP
ratios; (b) Per-Arnt-Sim kinase, which acts as a molec-
ular sensor of oxygen, redox status, ATP and other
indicators of the cellular metabolism; and (c) the
hexosamine biosynthetic pathway that produces uri-
dine 5¢-diphospho-N-acetylglucosamine as a substrate
for O-N-acetylglucosamine transferase [1]. One energy
conservation strategy that has attracted much attention
is enhanced autophagocytosis, also known as macro-
autophagy or simply autophagy [2]. This process is a
catabolic pathway involving the engulfment and degra-
dation of a cell’s own components through the lyso-
somal machinery [3,4]. Autophagic signaling in
response to nutrients is mainly relayed through the
serine-threonine kinase mammalian target of rapa-
mycin (mTOR). Indeed, mTOR is activated by nutri-
ent-rich conditions, especially high levels of amino
acids and insulin. Blocking mTOR function using
rapamycin or its analogs mimics nutrient deprivation
and triggers autophagy [1,4].
Keywords
autophagy; heterophagy; p38 MAP kinase;
scavenger receptor A; starvation
Correspondence
W. Martinet, Division of Pharmacology,
University of Antwerp, Universiteitsplein 1,
B-2610 Antwerp, Wilrijk, Belgium
Fax: +32 3 820 25 67
Tel: +32 3 820 26 79
E-mail: [email protected]
*These authors contributed equally to this
work
(Received 28 November 2008, revised 28
January 2009, accepted 5 February 2009)
doi:10.1111/j.1742-4658.2009.06951.x
Phagocytosis represents a mechanism used by macrophages to remove
pathogens and cellular debris. Recent evidence suggests that phagocytosis
is stimulated under specific conditions of stress, such as extracellular pres-
sure and hypoxia. In the present study, we show that amino acid or glucose
deprivation caused an increase in the phagocytosis of heat-inactivated
Escherichia coli and Staphylococcus aureus by macrophages, but not the
uptake of platelets, apoptotic cells or beads. Increased phagocytosis of bac-
teria could be blocked by phagocytosis inhibitors and was found to be
dependent on p38 mitogen-activated protein kinase activity and scavenger
receptor A. Although nutrient deprivation is a strong stimulus of auto-
phagy, autophagosome formation was not critical for the uptake of bacte-
ria because phagocytic clearance was not inhibited after down-regulation of
the autophagy essential gene Atg7. Moreover, enhanced uptake of bacteria
should not be considered as a general stress response because phagocytosis
of bacteria was not stimulated after exposure of macrophages to the
genotoxic agent camptothecin, heat (40 �C) or thapsigargin-induced endo-
plasmic reticulum stress. Overall, the results obtained in the present
study indicate that nutrient deprivation can stimulate macrophages to fight
bacterial infections.
Abbreviations
AC, apoptotic cell; CHOP, C ⁄ EBP homologous protein; EBSS, Earle’s balanced salt solution; eGFP, enhanced green fluorescent protein; ER,
endoplasmic reticulum; LPS, lipopolysaccharide; MAP, mitogen-activated protein; MARCO, macrophage receptor with collagenous structure;
mTOR, mammalian target of rapamycin; NDRG1, N-myc downstream-regulated gene 1; PI, propidium iodide; PI3-kinase, phosphoinositide
3-kinase; PLT, platelet; RORa, RAR-related orphan receptor alpha gene; siRNA, small interfering RNA; SR-A, scavenger receptor A; TLR,
Toll-like receptor.
FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2227

Beside autophagocytosis, cells have the ability to
internalize and degrade extracellular particles via het-
erophagocytosis (hereafter referred to as phagocytosis)
[5,6]. Lower organisms use phagocytosis for the acqui-
sition of nutrients, whereas phagocytosis in metazoa
primarily occurs in specialized cells, such as macro-
phages and neutrophils, where it has evolved into an
extraordinarily complex process. Phagocytosis by mac-
rophages is critical for the uptake and removal of
infectious agents and senescent or dying cells and is
stimulated under specific conditions of stress, such as
extracellular pressure [7] and hypoxia [8]. Several lines
of evidence indicate that many nutrients regulate phag-
ocytic activity in macrophages. For example, a choles-
terol-rich diet inhibits macrophage phagocytosis by
down-regulating plasma membrane fluidity and inter-
ference with receptor movement [9]. By contrast, ascor-
bate, as well as some trace elements, such as zinc, lead
and cadmium, stimulate the phagocytic capacity of
macrophages [10–12]. Furthermore, a diminished avail-
ability of iron may impair the ability of phagocytic
cells to kill ingested bacteria and fungi by down-regu-
lating myeloperoxidase activity [13].
Recent evidence was provided showing that auto-
phagy is essential for the phagocytosis of apoptotic
corpses during embryonic development because the
autophagic process contributes to the generation of
engulfment signals in apoptotic cells (ACs) by main-
taining cellular ATP production [14]. Moreover, engag-
ing the autophagic pathway via Toll-like receptor
(TLR) signaling also enhances phagosome maturation
and the destruction of engulfed material [15]. It should
also be noted that members of the phosphoinositide
3-kinase (PI3-kinase) family participate in autophago-
some formation [16,17] and in phagocytosis through
the delivery of membranes into extending pseudopodia
[18]. These findings clearly indicate that the autophagic
pathway is tightly linked to phagocytosis. In the pres-
ent study, we examined whether nutrient deprivation,
one of the main triggers of autophagy, stimulates the
phagocytosis capacity of macrophages.
Results
Nutrient deprivation leads to an increase in the
phagocytosis of heat-inactivated bacteria by
macrophages in vitro
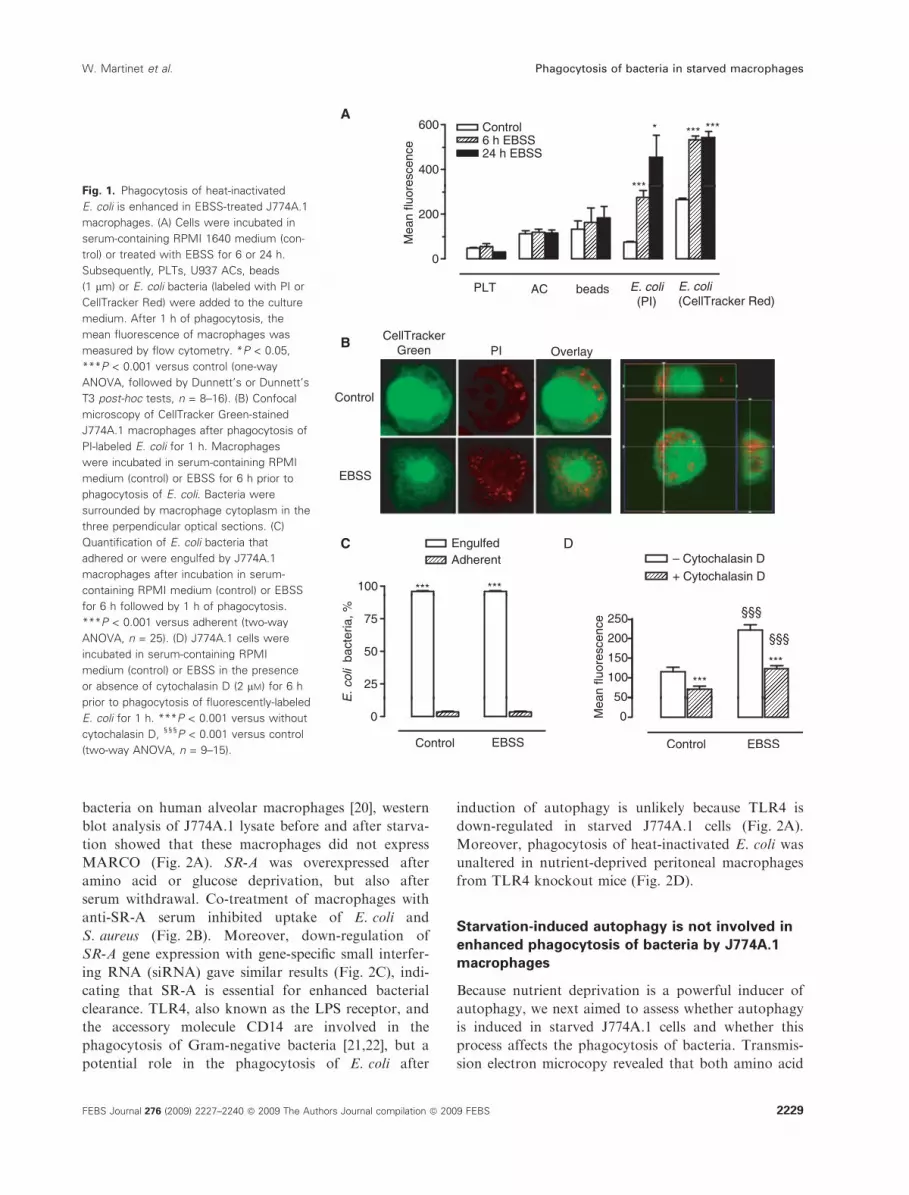
Mouse J774A.1 macrophages were incubated in amino
acid-free Earle’s balanced salt solution (EBSS) or con-
trol medium supplemented with 10% fetal bovine
serum for 6 or 24 h, followed by 1 h of incubation
with fluorescently-labeled platelets (PLT), U937 ACs,
carboxylated beads (0.1 or 1 lm in diameter) or heat-
inactivated Escherichia coli bacteria. Flow cytometric
analysis demonstrated that the uptake of PLTs, U937
AC or beads was not changed in EBSS-starved macro-
phages versus control cells (Fig. 1A). By contrast, the
uptake of E. coli bacteria was clearly increased after 6
and 24 h (Fig. 1A). To examine the potential role of
TLR signaling in the enhanced clearance of bacteria,
phagocytosis of beads was measured in the presence of
10 lgÆmL)1 lipopolysaccharide (LPS), which is a TLR
ligand and important surface constituent of E. coli.
Despite activation of macrophages by LPS, as deter-
mined by nitrite measurements in the culture medium
(12 ± 2 lm nitrite versus < 0.1 lm after 24 h of incu-
bation with or without LPS, respectively), phagocytosis
of beads was not stimulated. Increased phagocytosis of
E. coli in macrophages undergoing EBSS-induced
nutrient deprivation was confirmed by confocal
microscopy (Fig. 1B,C). Optical slides in the three
perpendicular axes showed that most bacteria were
surrounded by macrophage cytoplasm in all dimen-
sions (Fig. 1B). Moreover, enhanced uptake of bacteria
was blocked by the phagocytosis inhibitor cytochala-
sin D (Fig. 1D). We may therefore assume that the
flow cytometry data truly reflect phagocytosis and not
merely adherence of the bacteria to the macrophage
surface. Apart from amino acid deprivation, glucose
deprivation significantly increased the internalization
of heat-inactivated E. coli, whereas incubation in
serum-free medium had no effect (Fig. 2B). Similar
findings were obtained after phagocytosis of the
Gram-positive bacterium Staphylococcus aureus
(Fig. 2B). Titration experiments demonstrated that glu-
cose in the medium must be below 1 mgÆdL)1 to trig-
ger enhanced bacterial uptake (not shown).
Nutrient deprivation was also found to cause an
increase in the extent of phagocytosis by peritoneal
macrophages in vitro, as reflected by an increase in
mean fluorescence of macrophages after phagocytosis
of propidium iodide (PI)-labeled E. coli in amino
acid deprivation versus control conditions (mean fluo-
rescence: 116 ± 14 versus 48 ± 5, respectively;
P < 0.001; unpaired Student’s t-test, n = 5).
Enhanced phagocytosis of bacteria by starved
J774A.1 macrophages is scavenger receptor A
(SR-A) dependent
Both SR-A and macrophage receptor with collagenous
structure (MARCO) mediate the binding of unopson-
ized bacteria in vitro and in vivo, and are suggested to
play a pivotal role in bacterial clearance [19]. Although
MARCO is the dominant receptor for unopsonized
Phagocytosis of bacteria in starved macrophages W. Martinet et al.
2228 FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS
bacteria on human alveolar macrophages [20], western
blot analysis of J774A.1 lysate before and after starva-
tion showed that these macrophages did not express
MARCO (Fig. 2A). SR-A was overexpressed after
amino acid or glucose deprivation, but also after
serum withdrawal. Co-treatment of macrophages with
anti-SR-A serum inhibited uptake of E. coli and
S. aureus (Fig. 2B). Moreover, down-regulation of
SR-A gene expression with gene-specific small interfer-
ing RNA (siRNA) gave similar results (Fig. 2C), indi-
cating that SR-A is essential for enhanced bacterial
clearance. TLR4, also known as the LPS receptor, and
the accessory molecule CD14 are involved in the
phagocytosis of Gram-negative bacteria [21,22], but a
potential role in the phagocytosis of E. coli after
induction of autophagy is unlikely because TLR4 is
down-regulated in starved J774A.1 cells (Fig. 2A).
Moreover, phagocytosis of heat-inactivated E. coli was
unaltered in nutrient-deprived peritoneal macrophages
from TLR4 knockout mice (Fig. 2D).
Starvation-induced autophagy is not involved in
enhanced phagocytosis of bacteria by J774A.1
macrophages
Because nutrient deprivation is a powerful inducer of
autophagy, we next aimed to assess whether autophagy
is induced in starved J774A.1 cells and whether this
process affects the phagocytosis of bacteria. Transmis-
sion electron microcopy revealed that both amino acid
Fig. 1. Phagocytosis of heat-inactivated
E. coli is enhanced in EBSS-treated J774A.1
macrophages. (A) Cells were incubated in
serum-containing RPMI 1640 medium (con-
trol) or treated with EBSS for 6 or 24 h.
Subsequently, PLTs, U937 ACs, beads
(1 lm) or E. coli bacteria (labeled with PI or
CellTracker Red) were added to the culture
medium. After 1 h of phagocytosis, the
mean fluorescence of macrophages was
measured by flow cytometry. *P < 0.05,
***P < 0.001 versus control (one-way
ANOVA, followed by Dunnett’s or Dunnett’s
T3 post-hoc tests, n = 8–16). (B) Confocal
microscopy of CellTracker Green-stained
J774A.1 macrophages after phagocytosis of
PI-labeled E. coli for 1 h. Macrophages
were incubated in serum-containing RPMI
medium (control) or EBSS for 6 h prior to
phagocytosis of E. coli. Bacteria were
surrounded by macrophage cytoplasm in the
three perpendicular optical sections. (C)
Quantification of E. coli bacteria that
adhered or were engulfed by J774A.1
macrophages after incubation in serum-
containing RPMI medium (control) or EBSS
for 6 h followed by 1 h of phagocytosis.
***P < 0.001 versus adherent (two-way
ANOVA, n = 25). (D) J774A.1 cells were
incubated in serum-containing RPMI
medium (control) or EBSS in the presence
or absence of cytochalasin D (2 lM) for 6 h
prior to phagocytosis of fluorescently-labeled
E. coli for 1 h. ***P < 0.001 versus without
cytochalasin D, §§§P < 0.001 versus control
(two-way ANOVA, n = 9–15).
W. Martinet et al. Phagocytosis of bacteria in starved macrophages
FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2229
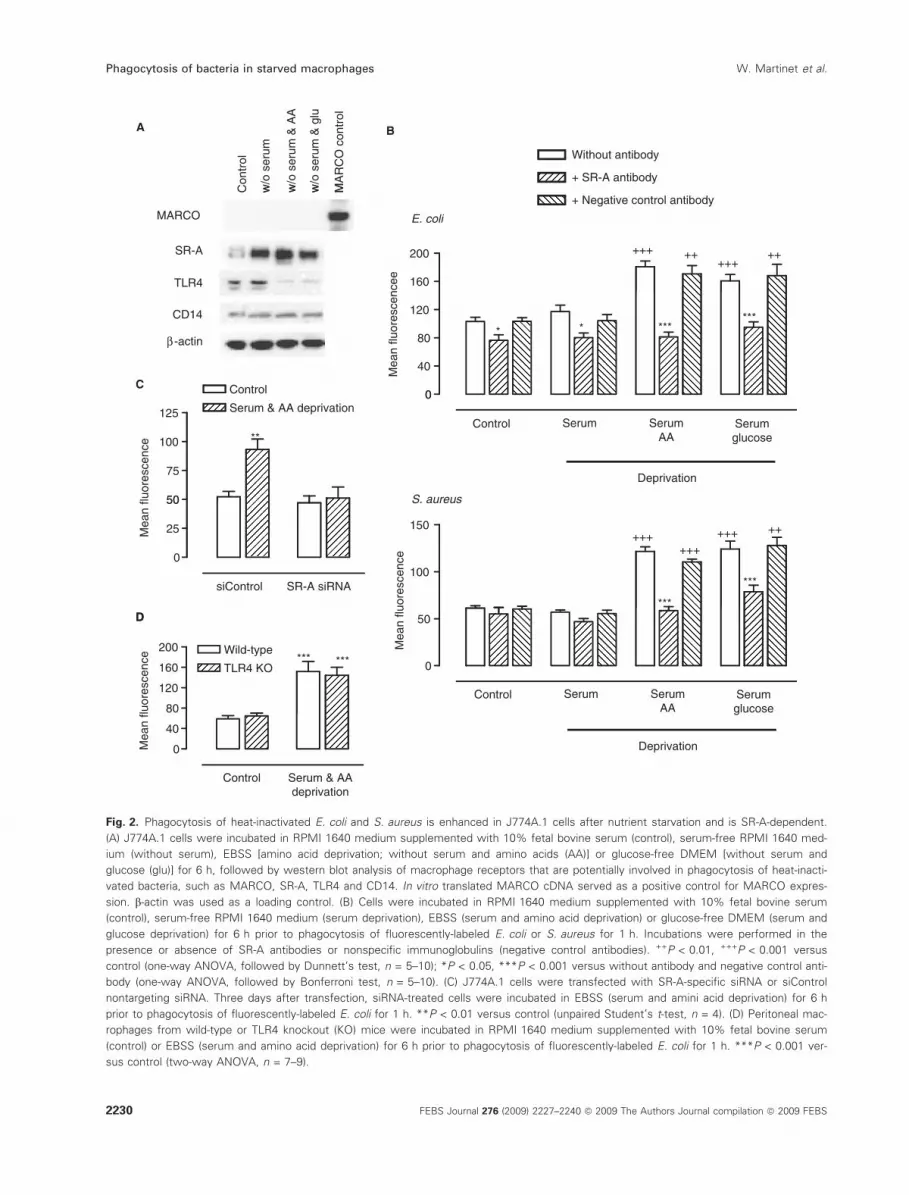
Fig. 2. Phagocytosis of heat-inactivated E. coli and S. aureus is enhanced in J774A.1 cells after nutrient starvation and is SR-A-dependent.
(A) J774A.1 cells were incubated in RPMI 1640 medium supplemented with 10% fetal bovine serum (control), serum-free RPMI 1640 med-
ium (without serum), EBSS [amino acid deprivation; without serum and amino acids (AA)] or glucose-free DMEM [without serum and
glucose (glu)] for 6 h, followed by western blot analysis of macrophage receptors that are potentially involved in phagocytosis of heat-inacti-
vated bacteria, such as MARCO, SR-A, TLR4 and CD14. In vitro translated MARCO cDNA served as a positive control for MARCO expres-
sion. b-actin was used as a loading control. (B) Cells were incubated in RPMI 1640 medium supplemented with 10% fetal bovine serum
(control), serum-free RPMI 1640 medium (serum deprivation), EBSS (serum and amino acid deprivation) or glucose-free DMEM (serum and
glucose deprivation) for 6 h prior to phagocytosis of fluorescently-labeled E. coli or S. aureus for 1 h. Incubations were performed in the
presence or absence of SR-A antibodies or nonspecific immunoglobulins (negative control antibodies). ++P < 0.01, +++P < 0.001 versus
control (one-way ANOVA, followed by Dunnett’s test, n = 5–10); *P < 0.05, ***P < 0.001 versus without antibody and negative control anti-
body (one-way ANOVA, followed by Bonferroni test, n = 5–10). (C) J774A.1 cells were transfected with SR-A-specific siRNA or siControl
nontargeting siRNA. Three days after transfection, siRNA-treated cells were incubated in EBSS (serum and amini acid deprivation) for 6 h
prior to phagocytosis of fluorescently-labeled E. coli for 1 h. **P < 0.01 versus control (unpaired Student’s t-test, n = 4). (D) Peritoneal mac-
rophages from wild-type or TLR4 knockout (KO) mice were incubated in RPMI 1640 medium supplemented with 10% fetal bovine serum
(control) or EBSS (serum and amino acid deprivation) for 6 h prior to phagocytosis of fluorescently-labeled E. coli for 1 h. ***P < 0.001 ver-
sus control (two-way ANOVA, n = 7–9).
Phagocytosis of bacteria in starved macrophages W. Martinet et al.
2230 FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS
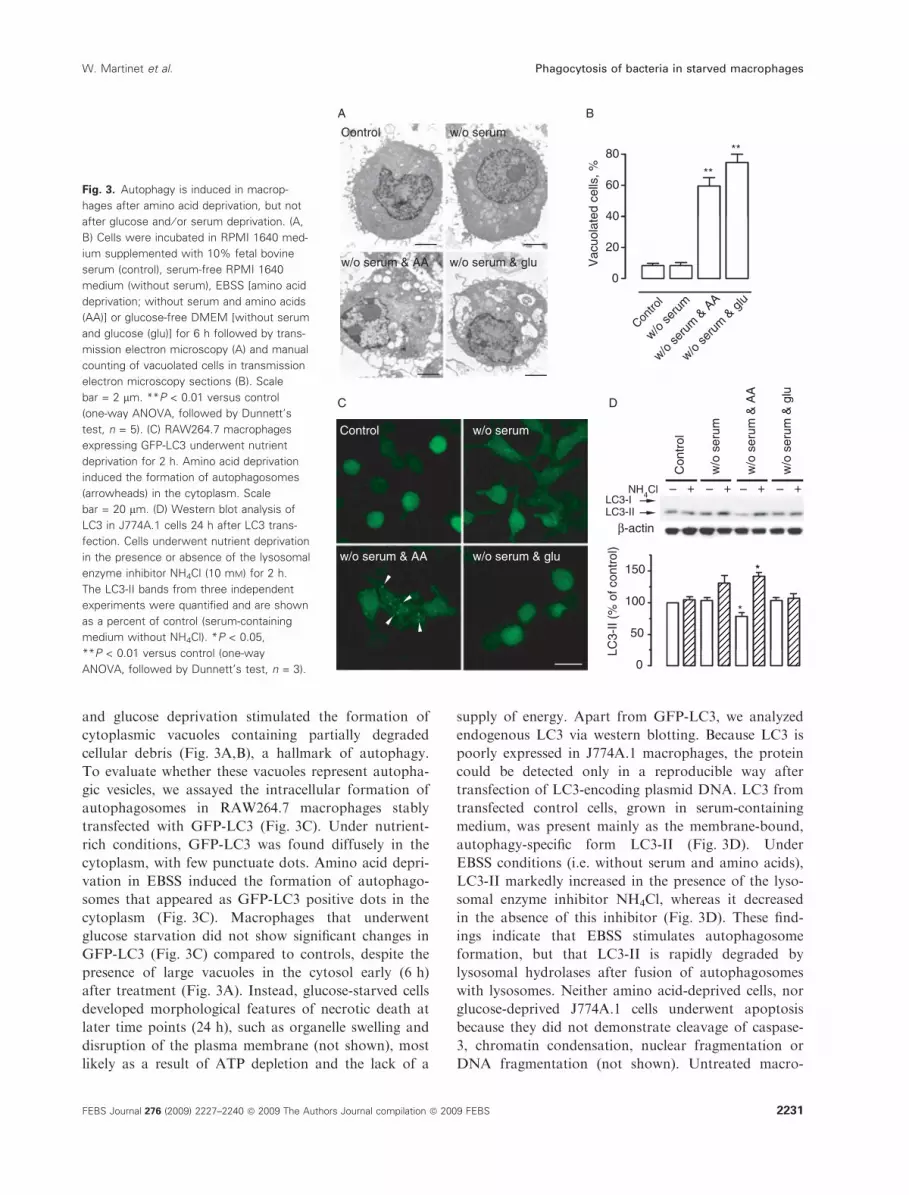
and glucose deprivation stimulated the formation of
cytoplasmic vacuoles containing partially degraded
cellular debris (Fig. 3A,B), a hallmark of autophagy.
To evaluate whether these vacuoles represent autopha-
gic vesicles, we assayed the intracellular formation of
autophagosomes in RAW264.7 macrophages stably
transfected with GFP-LC3 (Fig. 3C). Under nutrient-
rich conditions, GFP-LC3 was found diffusely in the
cytoplasm, with few punctuate dots. Amino acid depri-
vation in EBSS induced the formation of autophago-
somes that appeared as GFP-LC3 positive dots in the
cytoplasm (Fig. 3C). Macrophages that underwent
glucose starvation did not show significant changes in
GFP-LC3 (Fig. 3C) compared to controls, despite the
presence of large vacuoles in the cytosol early (6 h)
after treatment (Fig. 3A). Instead, glucose-starved cells
developed morphological features of necrotic death at
later time points (24 h), such as organelle swelling and
disruption of the plasma membrane (not shown), most
likely as a result of ATP depletion and the lack of a
supply of energy. Apart from GFP-LC3, we analyzed
endogenous LC3 via western blotting. Because LC3 is
poorly expressed in J774A.1 macrophages, the protein
could be detected only in a reproducible way after
transfection of LC3-encoding plasmid DNA. LC3 from
transfected control cells, grown in serum-containing
medium, was present mainly as the membrane-bound,
autophagy-specific form LC3-II (Fig. 3D). Under
EBSS conditions (i.e. without serum and amino acids),
LC3-II markedly increased in the presence of the lyso-
somal enzyme inhibitor NH4Cl, whereas it decreased
in the absence of this inhibitor (Fig. 3D). These find-
ings indicate that EBSS stimulates autophagosome
formation, but that LC3-II is rapidly degraded by
lysosomal hydrolases after fusion of autophagosomes
with lysosomes. Neither amino acid-deprived cells, nor
glucose-deprived J774A.1 cells underwent apoptosis
because they did not demonstrate cleavage of caspase-
3, chromatin condensation, nuclear fragmentation or
DNA fragmentation (not shown). Untreated macro-
Fig. 3. Autophagy is induced in macrop-
hages after amino acid deprivation, but not
after glucose and ⁄ or serum deprivation. (A,
B) Cells were incubated in RPMI 1640 med-
ium supplemented with 10% fetal bovine
serum (control), serum-free RPMI 1640
medium (without serum), EBSS [amino acid
deprivation; without serum and amino acids
(AA)] or glucose-free DMEM [without serum
and glucose (glu)] for 6 h followed by trans-
mission electron microscopy (A) and manual
counting of vacuolated cells in transmission
electron microscopy sections (B). Scale
bar = 2 lm. **P < 0.01 versus control
(one-way ANOVA, followed by Dunnett’s
test, n = 5). (C) RAW264.7 macrophages
expressing GFP-LC3 underwent nutrient
deprivation for 2 h. Amino acid deprivation
induced the formation of autophagosomes
(arrowheads) in the cytoplasm. Scale
bar = 20 lm. (D) Western blot analysis of
LC3 in J774A.1 cells 24 h after LC3 trans-
fection. Cells underwent nutrient deprivation
in the presence or absence of the lysosomal
enzyme inhibitor NH4Cl (10 mM) for 2 h.
The LC3-II bands from three independent
experiments were quantified and are shown
as a percent of control (serum-containing
medium without NH4Cl). *P < 0.05,
**P < 0.01 versus control (one-way
ANOVA, followed by Dunnett’s test, n = 3).
W. Martinet et al. Phagocytosis of bacteria in starved macrophages
FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2231
phages or cells incubated in medium without serum
showed a normal cell morphology and did not undergo
autophagy, apoptosis or necrotic death (Fig. 3A–D).
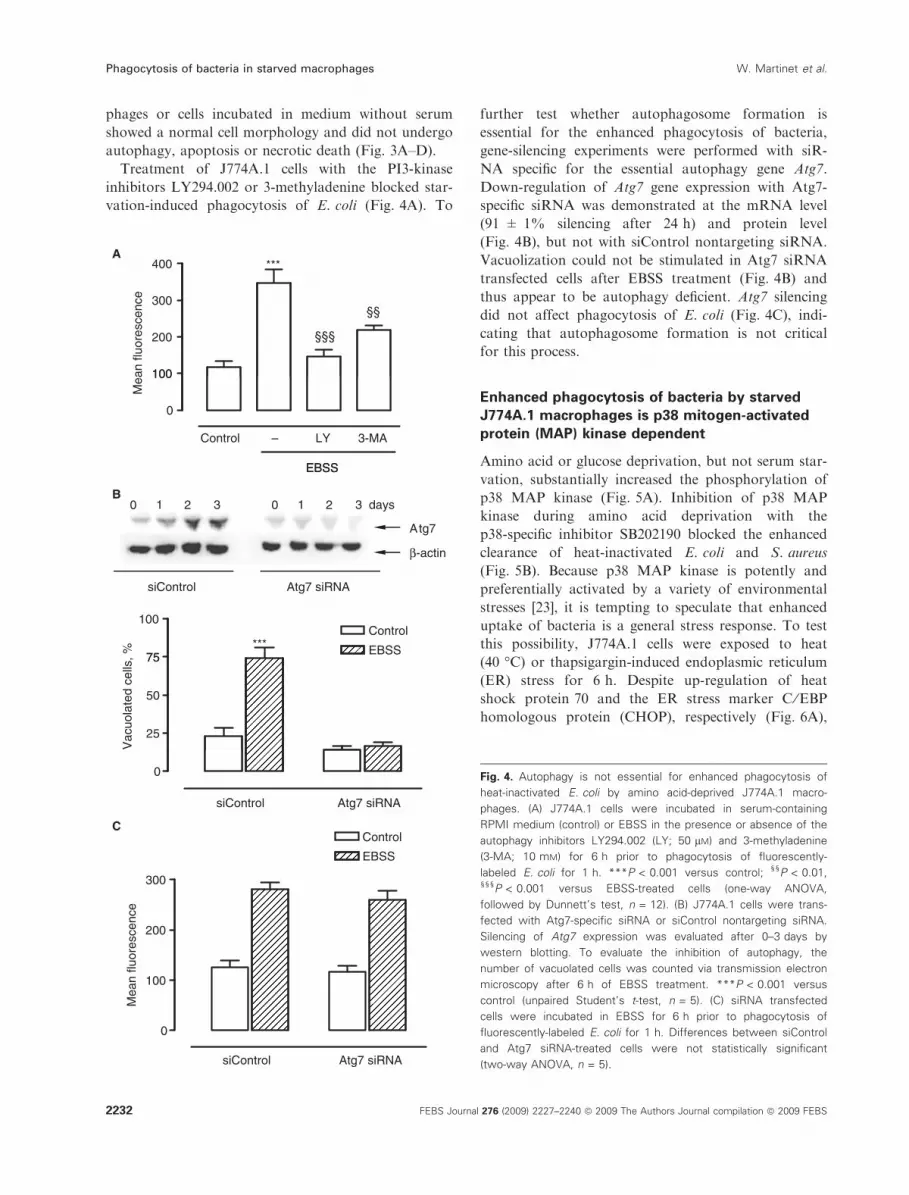
Treatment of J774A.1 cells with the PI3-kinase
inhibitors LY294.002 or 3-methyladenine blocked star-
vation-induced phagocytosis of E. coli (Fig. 4A). To
further test whether autophagosome formation is
essential for the enhanced phagocytosis of bacteria,
gene-silencing experiments were performed with siR-
NA specific for the essential autophagy gene Atg7.
Down-regulation of Atg7 gene expression with Atg7-
specific siRNA was demonstrated at the mRNA level
(91 ± 1% silencing after 24 h) and protein level
(Fig. 4B), but not with siControl nontargeting siRNA.
Vacuolization could not be stimulated in Atg7 siRNA
transfected cells after EBSS treatment (Fig. 4B) and
thus appear to be autophagy deficient. Atg7 silencing
did not affect phagocytosis of E. coli (Fig. 4C), indi-
cating that autophagosome formation is not critical
for this process.
Enhanced phagocytosis of bacteria by starved
J774A.1 macrophages is p38 mitogen-activated
protein (MAP) kinase dependent
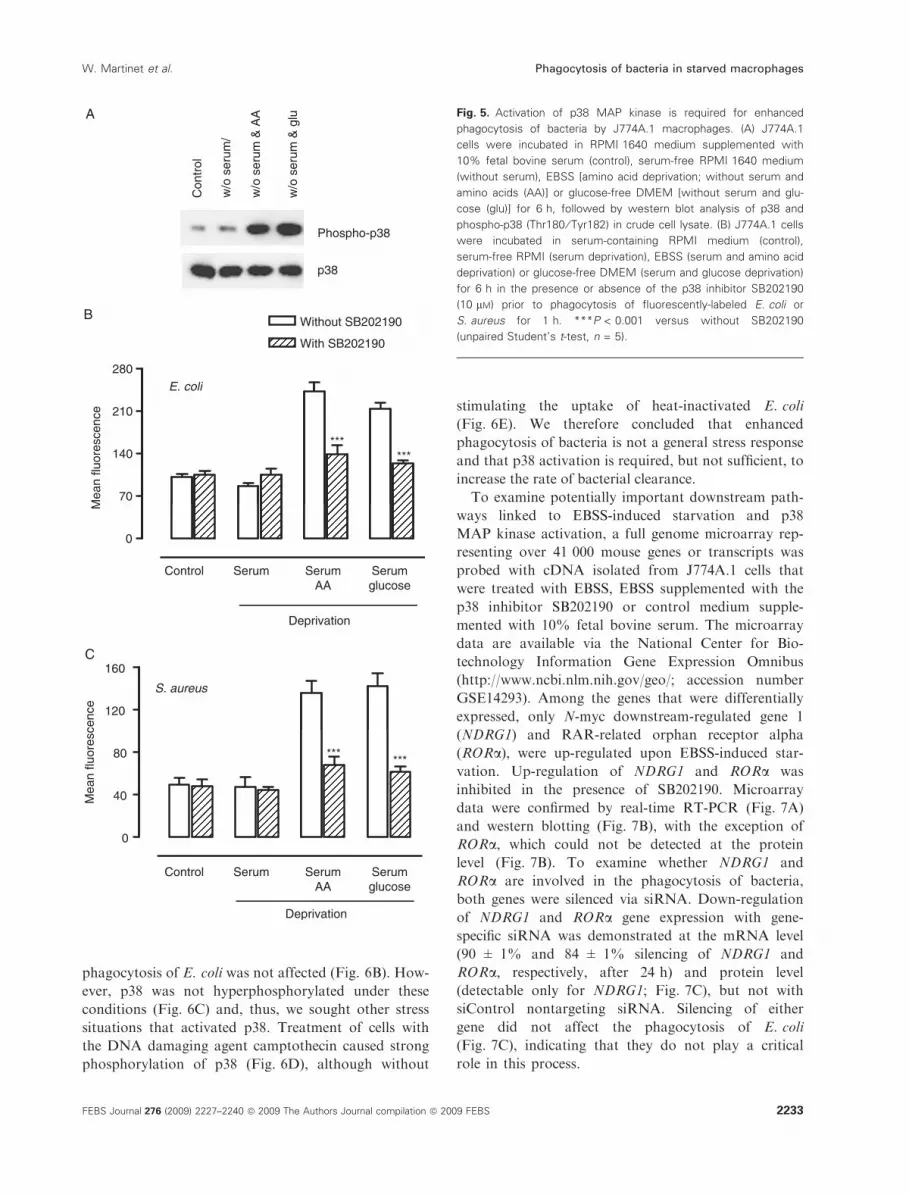
Amino acid or glucose deprivation, but not serum star-
vation, substantially increased the phosphorylation of
p38 MAP kinase (Fig. 5A). Inhibition of p38 MAP
kinase during amino acid deprivation with the
p38-specific inhibitor SB202190 blocked the enhanced
clearance of heat-inactivated E. coli and S. aureus
(Fig. 5B). Because p38 MAP kinase is potently and
preferentially activated by a variety of environmental
stresses [23], it is tempting to speculate that enhanced
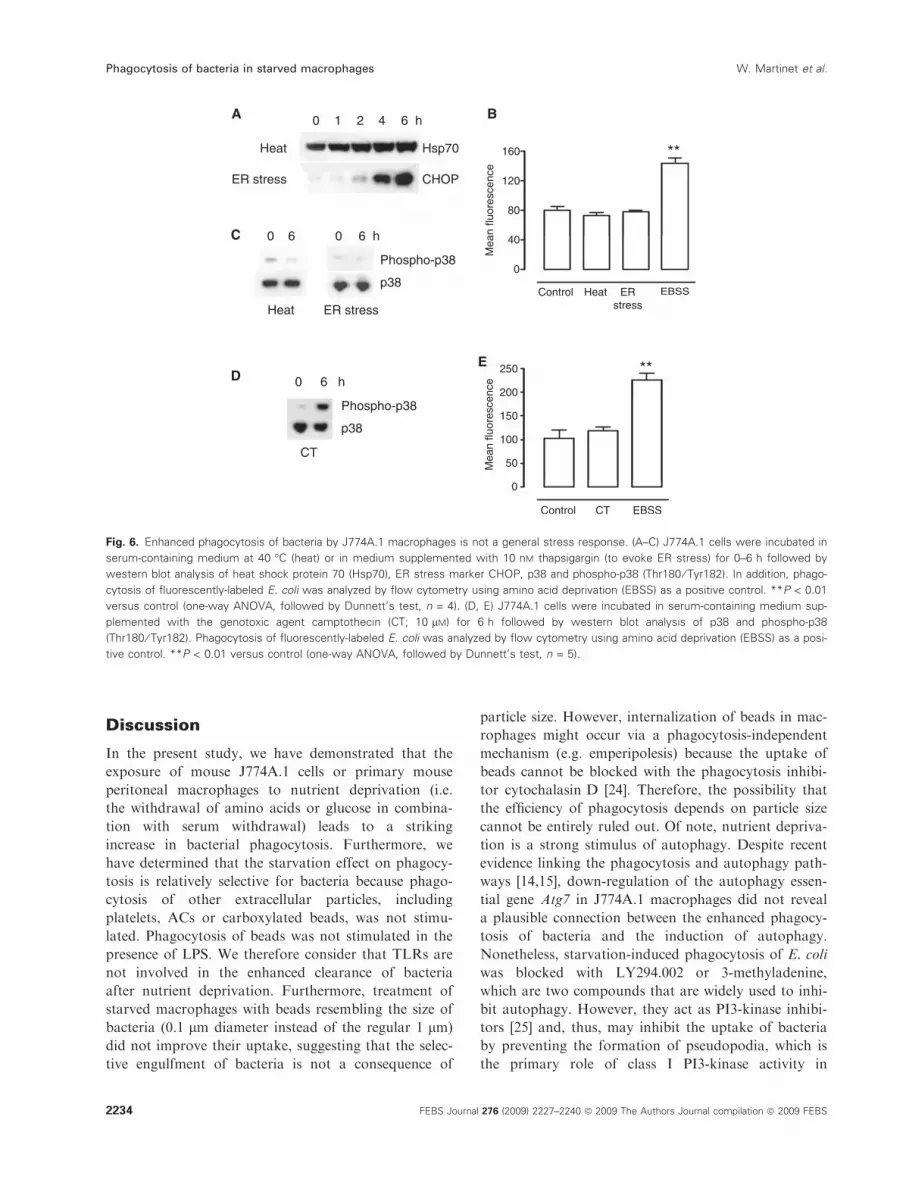
uptake of bacteria is a general stress response. To test
this possibility, J774A.1 cells were exposed to heat
(40 �C) or thapsigargin-induced endoplasmic reticulum
(ER) stress for 6 h. Despite up-regulation of heat
shock protein 70 and the ER stress marker C ⁄EBPhomologous protein (CHOP), respectively (Fig. 6A),
Fig. 4. Autophagy is not essential for enhanced phagocytosis of
heat-inactivated E. coli by amino acid-deprived J774A.1 macro-
phages. (A) J774A.1 cells were incubated in serum-containing
RPMI medium (control) or EBSS in the presence or absence of the
autophagy inhibitors LY294.002 (LY; 50 lM) and 3-methyladenine
(3-MA; 10 mM) for 6 h prior to phagocytosis of fluorescently-
labeled E. coli for 1 h. ***P < 0.001 versus control; §§P < 0.01,§§§P < 0.001 versus EBSS-treated cells (one-way ANOVA,
followed by Dunnett’s test, n = 12). (B) J774A.1 cells were trans-
fected with Atg7-specific siRNA or siControl nontargeting siRNA.
Silencing of Atg7 expression was evaluated after 0–3 days by
western blotting. To evaluate the inhibition of autophagy, the
number of vacuolated cells was counted via transmission electron
microscopy after 6 h of EBSS treatment. ***P < 0.001 versus
control (unpaired Student’s t-test, n = 5). (C) siRNA transfected
cells were incubated in EBSS for 6 h prior to phagocytosis of
fluorescently-labeled E. coli for 1 h. Differences between siControl
and Atg7 siRNA-treated cells were not statistically significant
(two-way ANOVA, n = 5).
Phagocytosis of bacteria in starved macrophages W. Martinet et al.
2232 FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS
phagocytosis of E. coli was not affected (Fig. 6B). How-
ever, p38 was not hyperphosphorylated under these
conditions (Fig. 6C) and, thus, we sought other stress
situations that activated p38. Treatment of cells with
the DNA damaging agent camptothecin caused strong
phosphorylation of p38 (Fig. 6D), although without
stimulating the uptake of heat-inactivated E. coli
(Fig. 6E). We therefore concluded that enhanced
phagocytosis of bacteria is not a general stress response
and that p38 activation is required, but not sufficient, to
increase the rate of bacterial clearance.
To examine potentially important downstream path-
ways linked to EBSS-induced starvation and p38
MAP kinase activation, a full genome microarray rep-
resenting over 41 000 mouse genes or transcripts was
probed with cDNA isolated from J774A.1 cells that
were treated with EBSS, EBSS supplemented with the
p38 inhibitor SB202190 or control medium supple-
mented with 10% fetal bovine serum. The microarray
data are available via the National Center for Bio-
technology Information Gene Expression Omnibus
(http://www.ncbi.nlm.nih.gov/geo/; accession number
GSE14293). Among the genes that were differentially
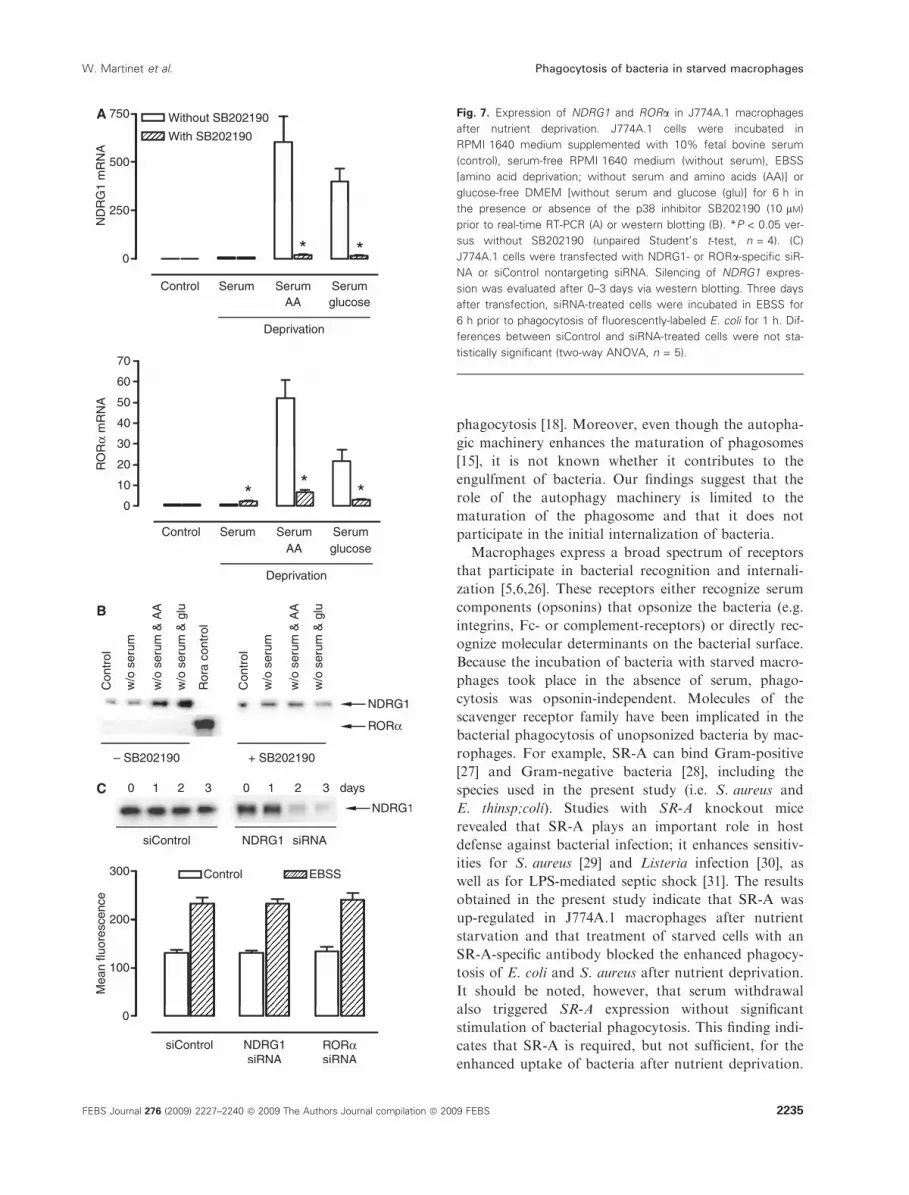
expressed, only N-myc downstream-regulated gene 1
(NDRG1) and RAR-related orphan receptor alpha
(RORa), were up-regulated upon EBSS-induced star-
vation. Up-regulation of NDRG1 and RORa was
inhibited in the presence of SB202190. Microarray
data were confirmed by real-time RT-PCR (Fig. 7A)
and western blotting (Fig. 7B), with the exception of
RORa, which could not be detected at the protein
level (Fig. 7B). To examine whether NDRG1 and
RORa are involved in the phagocytosis of bacteria,
both genes were silenced via siRNA. Down-regulation
of NDRG1 and RORa gene expression with gene-
specific siRNA was demonstrated at the mRNA level
(90 ± 1% and 84 ± 1% silencing of NDRG1 and
RORa, respectively, after 24 h) and protein level
(detectable only for NDRG1; Fig. 7C), but not with
siControl nontargeting siRNA. Silencing of either
gene did not affect the phagocytosis of E. coli
(Fig. 7C), indicating that they do not play a critical
role in this process.
Fig. 5. Activation of p38 MAP kinase is required for enhanced
phagocytosis of bacteria by J774A.1 macrophages. (A) J774A.1
cells were incubated in RPMI 1640 medium supplemented with
10% fetal bovine serum (control), serum-free RPMI 1640 medium
(without serum), EBSS [amino acid deprivation; without serum and
amino acids (AA)] or glucose-free DMEM [without serum and glu-
cose (glu)] for 6 h, followed by western blot analysis of p38 and
phospho-p38 (Thr180 ⁄ Tyr182) in crude cell lysate. (B) J774A.1 cells
were incubated in serum-containing RPMI medium (control),
serum-free RPMI (serum deprivation), EBSS (serum and amino acid
deprivation) or glucose-free DMEM (serum and glucose deprivation)
for 6 h in the presence or absence of the p38 inhibitor SB202190
(10 lM) prior to phagocytosis of fluorescently-labeled E. coli or
S. aureus for 1 h. ***P < 0.001 versus without SB202190
(unpaired Student’s t-test, n = 5).
W. Martinet et al. Phagocytosis of bacteria in starved macrophages
FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2233
Discussion
In the present study, we have demonstrated that the
exposure of mouse J774A.1 cells or primary mouse
peritoneal macrophages to nutrient deprivation (i.e.
the withdrawal of amino acids or glucose in combina-
tion with serum withdrawal) leads to a striking
increase in bacterial phagocytosis. Furthermore, we
have determined that the starvation effect on phagocy-
tosis is relatively selective for bacteria because phago-
cytosis of other extracellular particles, including
platelets, ACs or carboxylated beads, was not stimu-
lated. Phagocytosis of beads was not stimulated in the
presence of LPS. We therefore consider that TLRs are
not involved in the enhanced clearance of bacteria
after nutrient deprivation. Furthermore, treatment of
starved macrophages with beads resembling the size of
bacteria (0.1 lm diameter instead of the regular 1 lm)
did not improve their uptake, suggesting that the selec-
tive engulfment of bacteria is not a consequence of
particle size. However, internalization of beads in mac-
rophages might occur via a phagocytosis-independent
mechanism (e.g. emperipolesis) because the uptake of
beads cannot be blocked with the phagocytosis inhibi-
tor cytochalasin D [24]. Therefore, the possibility that
the efficiency of phagocytosis depends on particle size
cannot be entirely ruled out. Of note, nutrient depriva-
tion is a strong stimulus of autophagy. Despite recent
evidence linking the phagocytosis and autophagy path-
ways [14,15], down-regulation of the autophagy essen-
tial gene Atg7 in J774A.1 macrophages did not reveal
a plausible connection between the enhanced phagocy-
tosis of bacteria and the induction of autophagy.
Nonetheless, starvation-induced phagocytosis of E. coli
was blocked with LY294.002 or 3-methyladenine,
which are two compounds that are widely used to inhi-
bit autophagy. However, they act as PI3-kinase inhibi-
tors [25] and, thus, may inhibit the uptake of bacteria
by preventing the formation of pseudopodia, which is
the primary role of class I PI3-kinase activity in
Fig. 6. Enhanced phagocytosis of bacteria by J774A.1 macrophages is not a general stress response. (A–C) J774A.1 cells were incubated in
serum-containing medium at 40 �C (heat) or in medium supplemented with 10 nM thapsigargin (to evoke ER stress) for 0–6 h followed by
western blot analysis of heat shock protein 70 (Hsp70), ER stress marker CHOP, p38 and phospho-p38 (Thr180 ⁄ Tyr182). In addition, phago-
cytosis of fluorescently-labeled E. coli was analyzed by flow cytometry using amino acid deprivation (EBSS) as a positive control. **P < 0.01
versus control (one-way ANOVA, followed by Dunnett’s test, n = 4). (D, E) J774A.1 cells were incubated in serum-containing medium sup-
plemented with the genotoxic agent camptothecin (CT; 10 lM) for 6 h followed by western blot analysis of p38 and phospho-p38
(Thr180 ⁄ Tyr182). Phagocytosis of fluorescently-labeled E. coli was analyzed by flow cytometry using amino acid deprivation (EBSS) as a posi-
tive control. **P < 0.01 versus control (one-way ANOVA, followed by Dunnett’s test, n = 5).
Phagocytosis of bacteria in starved macrophages W. Martinet et al.
2234 FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS
phagocytosis [18]. Moreover, even though the autopha-
gic machinery enhances the maturation of phagosomes
[15], it is not known whether it contributes to the
engulfment of bacteria. Our findings suggest that the
role of the autophagy machinery is limited to the
maturation of the phagosome and that it does not
participate in the initial internalization of bacteria.
Macrophages express a broad spectrum of receptors
that participate in bacterial recognition and internali-
zation [5,6,26]. These receptors either recognize serum
components (opsonins) that opsonize the bacteria (e.g.
integrins, Fc- or complement-receptors) or directly rec-
ognize molecular determinants on the bacterial surface.
Because the incubation of bacteria with starved macro-
phages took place in the absence of serum, phago-
cytosis was opsonin-independent. Molecules of the
scavenger receptor family have been implicated in the
bacterial phagocytosis of unopsonized bacteria by mac-
rophages. For example, SR-A can bind Gram-positive
[27] and Gram-negative bacteria [28], including the
species used in the present study (i.e. S. aureus and
E. thinsp;coli). Studies with SR-A knockout mice
revealed that SR-A plays an important role in host
defense against bacterial infection; it enhances sensitiv-
ities for S. aureus [29] and Listeria infection [30], as
well as for LPS-mediated septic shock [31]. The results
obtained in the present study indicate that SR-A was
up-regulated in J774A.1 macrophages after nutrient
starvation and that treatment of starved cells with an
SR-A-specific antibody blocked the enhanced phagocy-
tosis of E. coli and S. aureus after nutrient deprivation.
It should be noted, however, that serum withdrawal
also triggered SR-A expression without significant
stimulation of bacterial phagocytosis. This finding indi-
cates that SR-A is required, but not sufficient, for the
enhanced uptake of bacteria after nutrient deprivation.
Fig. 7. Expression of NDRG1 and RORa in J774A.1 macrophages
after nutrient deprivation. J774A.1 cells were incubated in
RPMI 1640 medium supplemented with 10% fetal bovine serum
(control), serum-free RPMI 1640 medium (without serum), EBSS
[amino acid deprivation; without serum and amino acids (AA)] or
glucose-free DMEM [without serum and glucose (glu)] for 6 h in
the presence or absence of the p38 inhibitor SB202190 (10 lM)
prior to real-time RT-PCR (A) or western blotting (B). *P < 0.05 ver-
sus without SB202190 (unpaired Student’s t-test, n = 4). (C)
J774A.1 cells were transfected with NDRG1- or RORa-specific siR-
NA or siControl nontargeting siRNA. Silencing of NDRG1 expres-
sion was evaluated after 0–3 days via western blotting. Three days
after transfection, siRNA-treated cells were incubated in EBSS for
6 h prior to phagocytosis of fluorescently-labeled E. coli for 1 h. Dif-
ferences between siControl and siRNA-treated cells were not sta-
tistically significant (two-way ANOVA, n = 5).
W. Martinet et al. Phagocytosis of bacteria in starved macrophages
FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2235

Analogous to SR-A, the scavenger receptor MARCO
can adhere to Gram-positive and Gram-negative bacte-
ria. MARCO is considered to be the major binding
receptor for unopsonized bacteria on human alveolar
macrophages [20], but was not detectable in J774A.1
macrophages, suggesting that its involvement in
enhanced bacterial clearance, as reported in the present
study, is unlikely. Similarly, the changes in TLR4 and
CD14 cannot fully explain the enhanced uptake of
bacteria by starved J774A.1 cells because these mole-
cules only bind Gram-negative bacteria [21,22]. More-
over, these receptors were not up-regulated after
nutrient deprivation. Possibly, other receptors on the
surface of J774A.1 cells are involved in binding and ⁄orphagocytosis of bacteria [19]. The macrophage
mannose receptor and lectin-like oxidized low-density
lipoprotein receptor 1, for example, support the adhe-
sion of Gram-positive and Gram-negative bacteria, but
these receptors are not professional phagocytic recep-
tors, and only bind receptors that require a partner to
trigger efficient phagocytosis [32,33].
Although macrophage receptors are key proteins
involved in the phagocytosis of bacteria [19], several
lines of evidence suggest that other proteins may be
equally important. For example, Anand et al. [8] dem-
onstrated that hypoxia triggers phagocytosis of bacte-
ria by macrophages in a p38 MAP kinase-dependent
manner. Analogous with hypoxia, nutrient deprivation
(both amino acids and glucose, but not serum) sub-
stantially increased the phosphorylation of p38 MAP
kinase. Inhibition of p38 MAP kinase activation by
the p38-specific inhibitor SB202190 attenuated bacte-
rial phagocytosis induced by nutrient deprivation. p38
MAP kinase regulates cell growth, cell differentiation,
cell activation and cell death, and responses to inflam-
mation and stress stimuli at the transcriptional and
translational levels [34]. Many downstream substrates
of p38 MAP kinase have been described, both in the
cytoplasm and in the nucleus, that mediate these
effects. Doyle et al. [35] reported that numerous TLR
ligands specifically enhance the phagocytosis of bacte-
ria through myeloid differentiation factor 88, interleu-
kin-1 receptor-associated kinase-4 and p38, and that
activation of this pathway is essential for the up-regu-
lation of several scavenger receptors, including
MARCO and SR-A, as well as lectin-like oxidized
low-density lipoprotein receptor 1 and interstitial cell
adhesion molecule-1. Hypoxia-induced phagocytosis of
bacteria by macrophages occurs under the control of
hypoxia-inducible factor-1a, whose expression is
reversed after p38 inhibition [8]. In the present study, a
microarray screening of the whole mouse genome did
not reveal up-regulation of macrophage receptors or
hypoxia-inducible factor-1a in starved J774A.1 cells,
but led to the identification of two hypoxia-regulated
genes (i.e. NDRG1 and RORa) that were strongly
up-regulated at the mRNA level after amino acid or
glucose deprivation, but not after serum withdrawal.
The expression of both genes was controlled by p38
because up-regulation of NDRG1 and RORa mRNA
was inhibited in the presence of SB202190. The overex-
pression of NDRG1 was confirmed via western blot-
ting; however, expressed protein from RORa was
undetectable in J774A.1 cells even after starvation.
Given that the uptake of bacteria is enhanced after
hypoxia in macrophages [8] and that NDRG1 is
up-regulated after hypoxia [36], it is tempting to specu-
late that NDRG1 overexpression is associated with the
induction of bacterial clearance. However, gene-silenc-
ing experiments showed that neither NDRG1 nor
RORa are essential for the phagocytosis of bacteria.
Most likely, other factors may be expressed or released
during starvation, which are undetectable via micro-
array technology, and serve to augment phagocytosis
independent of NDRG1 and RORa. Indeed, macro-
phages are specialized immune cells that, once acti-
vated, may release large amounts of different cytokines
and ⁄or reactive oxygen species. Secretion of interleu-
kin-10 is a well-known stimulus for phagocytosis in
human monocytes [37,38]. We previously demonstrated
that J774A.1 macrophages secrete large amounts of
the pro-inflammatory cytokines interleukin-6 and
tumor necrosis factor a upon amino acid deprivation
[39]; however, Anand et al. [8] demonstrated that treat-
ment of macrophages with interleukin-1, tumor necro-
sis factor a or reactive oxygen species (H2O2 or NO)
does not affect phagocytosis significantly.
In summary, exposure of macrophages to amino
acid or glucose deprivation selectively stimulates the
phagocytosis of heat-inactivated Gram-positive and
Gram-negative bacteria via an SR-A- and p38-depen-
dent mechanism. From a clinical perspective, these
results are promising with respect to the development
of nutritional strategies for the treatment of patients
who are infected with multi-resistant bacteria.
Experimental procedures
Cell culture
Mouse J774A.1 macrophages and U937 monocytic cells
were grown in RPMI 1640 medium (Invitrogen, Carlsbad,
CA, USA) supplemented with antibiotics and 10% fetal
bovine serum. RAW264.7 macrophages stably expressing
GFP-LC3 [15] (a gift from M. Sanjuan, St Jude Children’s
Research Institute, Memphis, TN, USA) were grown in
Phagocytosis of bacteria in starved macrophages W. Martinet et al.
2236 FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS

serum-containing DMEM medium. Peritoneal macrophages
were isolated 5 days after injection of 2 mL of Brewer’s
thioglycolate medium (Sigma, St Louis, MO, USA) into the
peritoneal cavity of fasting C57Bl ⁄ 6 mice. To generate
ACs, U937 cells were incubated with 50 lm etoposide for
4 h. Macrophages, U937 ACs and human blood platelet
concentrates (kindly provided by the blood transfusion cen-
ter of the University Hospital of Antwerp, Belgium) were
fluorescently labeled with 10 lm CellTracker Green (Molec-
ular Probes, Eugene, OR, USA) for 30 min at 37 �C. Forphagocytosis experiments, macrophages were washed in
NaCl ⁄Pi and incubated in RPMI 1640 medium with or
without serum, DMEM without glucose or EBSS in the
presence or absence of the autophagy ⁄phagocytosis inhibi-
tors LY294.002 (50 lm; Sigma) or 3-methyladenine
(10 mm; Sigma), the phagocytosis inhibitor cytochalasin D
(2 lm; Sigma) or the p38 MAP kinase inhibitor SB202190
(10 lm; Sigma). These inhibitors did not affect macrophage
viability within the studied time frame. In some phagocyto-
sis experiments, the medium was supplemented with
10 lgÆmL)1 LPS from E. coli O111:B4 (Sigma) to assess the
potential role of TLR signaling. Activation of macrophages
by LPS was assessed by measuring nitrite, a stable NO
metabolite and indicator of NO synthase activity, using the
Griess reaction [40]. In other experiments, SR-A was
blocked prior to phagocytosis of bacteria by addition of rat
anti-SR-A (2F8; AbD Serotec, Oxford, UK) to the culture
medium (10 lgÆmL)1). Immunoglobulin G from rat serum
(Sigma) was used as a negative control.
Bacteria
E. coli and S. aureus bacteria were grown in LB medium at
37 �C. Bacteria were harvested by centrifugation and
washed three times with NaCl ⁄Pi to remove LB. Subse-
quently, bacteria were heat inactivated (1 h at 70 �C) and
fluorescently labeled with 10 lgÆmL)1 PI or 10 lm Cell-
Tracker Red (Molecular Probes) for 1 h on a shaker at
room temperature. Finally, bacteria were washed two times
in NaCl ⁄Pi and diluted to a concentration of 108 bacte-
riaÆmL)1.
Phagocytosis assay
Macrophages (106) were washed with NaCl ⁄Pi and incu-
bated for 1 h with PLTs (2 · 108), U937 ACs (3 · 106),
bacteria (5 · 107) or carboxylate-modified fluospheres
(5 · 107, 0.1 or 1.0 lm; Molecular Probes) at 37 �C. To
remove adherent and unphagocytosed particles, macrophag-
es were washed with cold NaCl ⁄Pi and briefly treated with
a solution of 0.05% trypsin and 0.02% EDTA. Finally,
macrophages were scraped from the plate and 20 000 cells
per sample were analyzed on a FACSort cytometer (BD
Biosciences, Franklin Lakes, NJ, USA). The acquisition
threshold was set to include cells but to exclude debris
and remaining unbound particles from further analysis.
Data were analyzed using cell quest pro Software (BD
Biosciences).
Plasmid construction, transfection and in vitro
translation
The enhanced green fluorescent protein (eGFP) coding
sequence was excised from plasmid pEGFP-N3 (BD Bio-
sciences Clontech, Mountain View, CA, USA) by digestion
with EcoRI and NotI. Vector ends were blunted with
Klenow polymerase and self ligated to produce plasmid pN3.
Full-length cDNA encoding mouse LC3 protein was ampli-
fied by PCR from C2C12 cells using Pfu DNA polymerase
(Stratagene, La Jolla, CA, USA) and primers 5¢-CAAGA
TCTCGCGCGATGCCCTCMGACCGG-3¢ and 5¢-CAAA
GCTTTCAGAAGCCGAAGGTTTCYTGGGAG-3¢. The
resultant PCR product was BglII ⁄HindIII digested and
cloned in the similarly opened pN3 to produce pMuLC3.
J774A.1 macrophages were transiently transfected using
Fugene HD transfection reagent (Roche, Mannheim,
Germany) at a ratio of 5 lL of reagent to 2 lg of plasmid
DNA.
In vitro translation of mouse MARCO was performed
using XbaI linearized pcDNA3MARCO plasmid (a gift
from T. Pikkarainen, Karolinska Institute, Stockholm, Swe-
den) and the Rabbit Reticulocyte Lysate System (Promega,
Madison, WI, USA). To obtain a positive control for
mouse RORa in western blot experiments, mouse RORacDNA was amplified by PCR using the oligonucleotides
5¢-TCAAAGCTTGCGATGAAAGCTCAAATTGAAATTA
TTCC-3¢ and 5¢-TCAGGATCCTTACCCATCGATTTG
CATGGCTGGCTC-3¢. The PCR product was HindIII ⁄BamHI digested and cloned in the similarly opened plasmid
pUC19 to yield pUC19RORa. Plasmid pGEM4ZGFP-A64
(gift from V. Van Tendeloo, University Hospital of
Antwerp, Edegem, Belgium) containing the eGFP coding
sequence downstream of a T7 promoter was BamHI
digested to remove eGFP and self ligated to produce
pGEM4Z-A64. RORa was then excised from pUC19RORaand cloned into the HindIII ⁄BamHI sites of pGEM4Z-A64
to yield pGEM4Z-RORa. In vitro translation of RORa was
finally performed using nonlinearized pGEM4Z-RORa and
the TNT Quick Coupled Transcription ⁄ translation System
(Promega). All constructs were confirmed by sequencing.
siRNA-mediated gene silencing
J774A.1 macrophages were transfected with 50 nm
ON-TARGETplus SMARTpool siRNA specific to mouse
Atg7, SR-A, NDRG1 or RORa (Dharmacon, Lafayette,
CO, USA) using HiPerfect transfection reagent (Qiagen,
Valencia, CA, USA) according to the manufacturer’s
instructions. siControl RISC-free siRNA (Dharmacon) was
used as a negative control.
W. Martinet et al. Phagocytosis of bacteria in starved macrophages
FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2237

Microarray analysis
Total RNA was prepared from cultured cells using the
Absolutely RNA Miniprep Kit (Stratagene). All RNA
samples were treated with RNase-free DNase I. RNA
quality was verified on an Agilent 2100 Bioanalyser using
the RNA 6000 Nano LabChip kit (Agilent Technologies,
Palo Alto, CA, USA). Samples were then analyzed by the
Microarray Facility of the Flanders Interuniversity Insti-
tute for Biotechnology (VIB, Leuven, Belgium) using the
Whole Mouse Genome Oligo Microarray Kit (Agilent
Technologies) representing over 41 000 mouse genes and
transcripts.
Real-time quantitative RT-PCR
cDNA was prepared from cultured cells using the Fastlane
Cell cDNA kit (Qiagen, Venlo, The Netherlands). TaqMan
gene expression assays for Atg7 (assay Id:
Mm01183587_m1), SR-A (assay Id: Mm01197876_m1),
NDRG1 (assay Id: Mm00440447_m1) and RORa (assay Id:
Mm00443103_m1) (Applied Biosystems, Foster City, CA,
USA) were then performed in duplicate on an ABIPrism
7300 sequence detector system (Applied Biosystems) in
25 lL reaction volumes containing 1· Universal PCR
Master Mix (Applied Biosystems). The parameters for
PCR amplification were 95 �C for 10 min followed by 40
cycles of 95 �C for 15 s and 60 �C for 1 min. The relative
expression of mRNA species was calculated using the
comparative threshold cycle method. All data were con-
trolled for quantity of cDNA input by performing measure-
ments on the endogenous reference gene b-actin (assay Id:
Mm00607939_s1; Applied Biosystems).
Immunoblot assays
Cultured cells were lysed in an appropriate volume of
Laemmli sample buffer (Bio-Rad, Richmond, CA, USA).
Cell lysates were then heat-denatured for 4 min in boiling
water and loaded onto SDS ⁄PAGE. After electrophoresis,
proteins were transferred to an Immobilon-P Transfer
Membrane (Millipore, Bedford, MA, USA) according to
standard procedures. Membranes were blocked in NaCl ⁄Tris containing 0.05% Tween-20 (NaCl ⁄Tris-T) and 5%
nonfat dry milk (Bio-Rad) for 1 h. After blocking, mem-
branes were probed overnight at 4 �C with primary anti-
bodies in antibody dilution buffer (NaCl ⁄Tris-Tcontaining 1% nonfat dry milk), followed by 1 h of incu-
bation with secondary antibody at room temperature.
Antibody detection was accomplished with SuperSignal
West Pico or SuperSignal West Femto Maximum Sensi-
tivity Substrate (Pierce, Rockford, IL, USA) using a
Lumi-Imager (Roche).
The primary antibodies used were: goat anti-MARCO
and rat anti-CD14 (R&D Systems, Abingdon, UK); rat
anti-SR-A (clone 2F8) (AbD Serotec); mouse anti-caspase-3
(clone 19) and anti-HSP70 (clone 7) (BD Transduction
Laboratories, Lexington, KY, USA); rabbit anti-Atg7, anti-
p38 and anti-phospho-p38 (Thr180 ⁄Tyr182) (Cell SignalingTechnology, Beverly, MA, USA); rabbit anti-LC3 (Novus
Biologicals, Littleton, CO, USA); chicken anti-NDRG1
(Abcam, Cambridge, UK); rabbit anti-RORa (clone H-65)
and mouse anti-CHOP (clone B-3) (Santa Cruz Biotechnol-
ogy, Santa Cruz, CA, USA); mouse anti-b-actin (clone
AC-15) (Sigma); and anti-Toll-like receptor 4 (eBioscience,
San Diego, CA, USA). Peroxidase-conjugated secondary
antibodies were purchased from DakoCytomation (Glost-
rup, Denmark).
Confocal microscopy
Macrophages were grown on coverslips, fluorescently
labeled with 10 lm CellTracker Green or 1 lm LysoTracker
(Molecular Probes) and incubated with PI-labeled bacteria
for 1 h. Finally, coverslips were formalin fixed and
mounted on microscope slides using Slowfade Gold
Reagent (Molecular Probes). Dual-channel images were
taken with a LSM510 confocal microscope (Carl Zeiss,
Jena, Germany), using an Argon laser (488 nm line) and a
HeNe laser (543 nm line) with a bandpass emission filter
(BP500-530) and a longpass emission filter (LP560), respec-
tively. Regions of interest containing individual macro-
phages were selected and Z-stacks were further analyzed
using the ortho and gallery displays of the LSM510 imag-
ing software. After confocal imaging, adherent and
engulfed bacteria were quantified via manual counting of
PI-labeled particles.
Transmission electron microscopy
J774A.1 macrophages were fixed in 0.1 m sodium cacody-
late-buffered (pH 7.4), 2.5% glutaraldehyde solution for
2 h and then rinsed (3 · 10 min) in 0.1 m sodium cacody-
late-buffered (pH 7.4), 7.5% saccharose and postfixed in
1% OsO4 solution for 1 h. After dehydration in an ethanol
gradient (70% ethanol for 20 min; 96% ethanol for 20 min;
100% ethanol for 2 · 20 min), samples were embedded in
Durcupan ACM. Ultrathin sections were stained with
uranyl acetate and lead citrate. Sections were examined in
a Philips CM 10 microscope (Philips, Eindhoven, The
Netherlands) at 80 kV.
Statistical analysis
Results are expressed as the mean ± SEM. All analyses
were performed using spss software, version 14.0 (SPSS
Inc., Chicago, IL, USA). The statistical tests used in the
present study are noted in the appropriate figure legends.
P < 0.05 was considered statistically significant.
Phagocytosis of bacteria in starved macrophages W. Martinet et al.
2238 FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS

Acknowledgements
Research was supported by the Fund for Scientific
Research (FWO)-flanders (Belgium) (projects No.
G.0308.04, G.0113.06 and G.0112.08), the University
of Antwerp (NOI-BOF and TOP-BOF) and the
Bekales Foundation. The authors thank Jan Van
Daele, Lieve Svensson, Francis Terloo and Dominique
De Rijck for their excellent technical assistance.
Wim Martinet and Dorien M Schrijvers are post-
doctoral fellows of the FWO-Flanders.
References
1 Lindsley JE & Rutter J (2004) Nutrient sensing and
metabolic decisions. Comp Biochem Physiol B Biochem
Mol Biol 139, 543–559.
2 Mizushima N, Levine B, Cuervo AM & Klionsky DJ
(2008) Autophagy fights disease through cellular self-
digestion. Nature 451, 1069–1075.
3 Yoshimori T (2004) Autophagy: a regulated bulk degra-
dation process inside cells. Biochem Biophys Res Com-
mun 313, 453–458.
4 Yorimitsu T & Klionsky DJ (2005) Autophagy: molecu-
lar machinery for self-eating. Cell Death Differ 12,
1542–1552.
5 Aderem A & Underhill DM (1999) Mechanisms of
phagocytosis in macrophages. Annu Rev Immunol 17,
593–623.
6 Stuart LM & Ezekowitz RA (2005) Phagocytosis: ele-
gant complexity. Immunity 22, 539–550.
7 Shiratsuchi H & Basson MD (2004) Extracellular pres-
sure stimulates macrophage phagocytosis by inhibiting
a pathway involving FAK and ERK. Am J Physiol Cell
Physiol 286, C1358–C1366.
8 Anand RJ, Gribar SC, Li J, Kohler JW, Branca MF,
Dubowski T, Sodhi CP & Hackam DJ (2007) Hypoxia
causes an increase in phagocytosis by macrophages in a
HIF-1alpha-dependent manner. J Leukoc Biol 82, 1257–
1265.
9 Dianzani MU, Torrielli MV, Canuto RA, Garcea R &
Feo F (1976) The influence of enrichment with choles-
terol on the phagocytic activity of rat macrophages.
J Pathol 118, 193–199.
10 Thomas WR & Holt PG (1978) Vitamin C and immu-
nity: an assessment of the evidence. Clin Exp Immunol
32, 370–379.
11 Chvapil M (1976) Effect of zinc on cells and biomem-
branes. Med Clin North Am 60, 799–812.
12 Koller LD & Roan JG (1977) Effects of lead and cad-
mium on mouse peritoneal macrophages. J Reticuloend-
othel Soc 21, 7–12.
13 Chandra RK (1973) Reduced bactericidal capacity of
polymorphs in iron deficiency. Arch Dis Child 48, 864–
866.
14 Qu X, Zou Z, Sun Q, Luby-Phelps K, Cheng P, Hogan
RN, Gilpin C & Levine B (2007) Autophagy gene-
dependent clearance of apoptotic cells during embryonic
development. Cell 128, 931–946.
15 Sanjuan MA, Dillon CP, Tait SW, Moshiach S, Dorsey
F, Connell S, Komatsu M, Tanaka K, Cleveland JL,
Withoff S et al. (2007) Toll-like receptor signalling in
macrophages links the autophagy pathway to phagocy-
tosis. Nature 450, 1253–1257.
16 Pattingre S, Espert L, Biard-Piechaczyk M & Codogno
P (2008) Regulation of macroautophagy by mTOR and
Beclin 1 complexes. Biochimie 90, 313–323.
17 Backer JM (2008) The regulation and function of Class
III PI3Ks: novel roles for Vps34. Biochem J 410, 1–17.
18 Stephens L, Ellson C & Hawkins P (2002) Roles of
PI3Ks in leukocyte chemotaxis and phagocytosis. Curr
Opin Cell Biol 14, 203–213.
19 Palecanda A & Kobzik L (2001) Receptors for unops-
onized particles: the role of alveolar macrophage scav-
enger receptors. Curr Mol Med 1, 589–595.
20 Arredouani MS, Palecanda A, Koziel H, Huang YC,
Imrich A, Sulahian TH, Ning YY, Yang Z, Pikkarainen
T, Sankala M et al. (2005) MARCO is the major bind-
ing receptor for unopsonized particles and bacteria on
human alveolar macrophages. J Immunol 175, 6058–
6064.
21 Neal MD, Leaphart C, Levy R, Prince J, Billiar TR,
Watkins S, Li J, Cetin S, Ford H, Schreiber A et al.
(2006) Enterocyte TLR4 mediates phagocytosis and
translocation of bacteria across the intestinal barrier.
J Immunol 176, 3070–3079.
22 Schiff DE, Kline L, Soldau K, Lee JD, Pugin J, Tobias
PS & Ulevitch RJ (1997) Phagocytosis of gram-negative
bacteria by a unique CD14-dependent mechanism.
J Leukoc Biol 62, 786–794.
23 Obata T, Brown GE & Yaffe MB (2000) MAP kinase
pathways activated by stress: the p38 MAPK pathway.
Crit Care Med 28, N67–N77.
24 Schrijvers DM, Martinet W, De Meyer GRY, Andries
L, Herman AG & Kockx MM (2004) Flow cytomet-
ric evaluation of a model for phagocytosis of cells
undergoing apoptosis. J Immunol Methods 287, 101–
108.
25 Klionsky DJ, Abeliovich H, Agostinis P, Agrawal DK,
Aliev G, Askew DS, Baba M, Baehrecke EH, Bahr BA,
Ballabio A et al. (2008) Guidelines for the use and
interpretation of assays for monitoring autophagy in
higher eukaryotes. Autophagy 4, 151–175.
26 Underhill DM & Ozinsky A (2002) Phagocytosis of
microbes: complexity in action. Annu Rev Immunol 20,
825–852.
27 Dunne DW, Resnick D, Greenberg J, Krieger M &
Joiner KA (1994) The type I macrophage scavenger
receptor binds to gram-positive bacteria and recognizes
W. Martinet et al. Phagocytosis of bacteria in starved macrophages
FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS 2239

lipoteichoic acid. Proc Natl Acad Sci USA 91, 1863–
1867.
28 Peiser L, Gough PJ, Kodama T & Gordon S (2000)
Macrophage class A scavenger receptor-mediated
phagocytosis of Escherichia coli: role of cell heterogene-
ity, microbial strain, and culture conditions in vitro.
Infect Immun 68, 1953–1963.
29 Thomas CA, Li Y, Kodama T, Suzuki H, Silverstein
SC & El KJ (2000) Protection from lethal gram-positive
infection by macrophage scavenger receptor-dependent
phagocytosis. J Exp Med 191, 147–156.
30 Suzuki H, Kurihara Y, Takeya M, Kamada N, Kat-
aoka M, Jishage K, Ueda O, Sakaguchi H, Higashi T,
Suzuki T et al. (1997) A role for macrophage scavenger
receptors in atherosclerosis and susceptibility to infec-
tion. Nature 386, 292–296.
31 Haworth R, Platt N, Keshav S, Hughes D, Darley E,
Suzuki H, Kurihara Y, Kodama T & Gordon S
(1997) The macrophage scavenger receptor type A is
expressed by activated macrophages and protects the
host against lethal endotoxic shock. J Exp Med 186,
1431–1439.
32 Le Cabec V, Emorine LJ, Toesca I, Cougoule C &
Maridonneau-Parini I (2005) The human macrophage
mannose receptor is not a professional phagocytic
receptor. J Leukoc Biol 77, 934–943.
33 Shimaoka T, Kume N, Minami M, Hayashida K,
Sawamura T, Kita T & Yonehara S (2001) LOX-1
supports adhesion of Gram-positive and Gram-negative
bacteria. J Immunol 166, 5108–5114.
34 Cuenda A & Rousseau S (2007) p38 MAP-kinases path-
way regulation, function and role in human diseases.
Biochim Biophys Acta 1773, 1358–1375.
35 Doyle SE, O’Connell RM, Miranda GA, Vaidya SA,
Chow EK, Liu PT, Suzuki S, Suzuki N, Modlin RL, Yeh
WC et al. (2004) Toll-like receptors induce a phagocytic
gene program through p38. J Exp Med 199, 81–90.
36 Ellen TP, Ke Q, Zhang P & Costa M (2008) NDRG1,
a growth and cancer related gene: regulation of gene
expression and function in normal and disease states.
Carcinogenesis 29, 2–8.
37 Capsoni F, Minonzio F, Ongari AM, Carbonelli V,
Galli A & Zanussi C (1995) IL-10 up-regulates human
monocyte phagocytosis in the presence of IL-4 and
IFN-gamma. J Leukoc Biol 58, 351–358.
38 Lingnau M, Hoflich C, Volk HD, Sabat R & Docke
WD (2007) Interleukin-10 enhances the CD14-depen-
dent phagocytosis of bacteria and apoptotic cells by
human monocytes. Hum Immunol 68, 730–738.
39 Martinet W, Schrijvers DM, Herman AG & De Meyer
GRY (2006) z-VAD-fmk-induced non-apoptotic cell
death of macrophages: possibilities and limitations for
atherosclerotic plaque stabilization. Autophagy 2, 312–
314.
40 Jans DM, Martinet W, Fillet M, Kockx MM, Merville
MP, Bult H, Herman AG & De Meyer GRY (2004)
Effect of non-steroidal anti-inflammatory drugs on
amyloid-beta formation and macrophage activation
after platelet phagocytosis. J Cardiovasc Pharmacol 43,
462–470.
Phagocytosis of bacteria in starved macrophages W. Martinet et al.
2240 FEBS Journal 276 (2009) 2227–2240 ª 2009 The Authors Journal compilation ª 2009 FEBS
Related Documents











