Persistent Production of Neurons from Adult Brain Stem Cells During Recovery after Stroke PA ¨ R THORED, a,c ANDREAS ARVIDSSON, a,c EMANUELE CACCI, b,c HENRIK AHLENIUS, b,c THERE ´ SE KALLUR, b,c VLADIMER DARSALIA, b,c CHRISTINE T. EKDAHL, a,c ZAAL KOKAIA, b,c OLLE LINDVALL a,c a Laboratory of Neurogenesis and Cell Therapy, Section of Restorative Neurology, Wallenberg Neuroscience Center, University Hospital, Lund, Sweden; b Laboratory of Neural Stem Cell Biology, Section of Restorative Neurology, Stem Cell Institute, University Hospital, Lund, Sweden; c Lund Strategic Research Center for Stem Cell Biology and Cell Therapy, Lund, Sweden Key Words. Stem cells • Neurogenesis • Striatum • Stroke ABSTRACT Neural stem cells in the subventricular zone of adult rodents produce new striatal neurons that may replace those that have died after stroke; however, the neurogenic response has been considered acute and transient, yielding only small numbers of neurons. In contrast, we show herein that stri- atal neuroblasts are generated without decline at least for 4 months after stroke in adult rats. Neuroblasts formed early or late after stroke either differentiate into mature neurons, which survive for several months, or die through caspase- mediated apoptosis. The directed migration of the new neu- rons toward the ischemic damage is regulated by stromal cell-derived factor-1 and its receptor CXCR4. These re- sults show that endogenous neural stem cells continuously supply the injured adult brain with new neurons, which suggests novel self-repair strategies to improve recovery after stroke. STEM CELLS 2006;24:739 –747 INTRODUCTION Stroke is a leading cause of chronic disability in humans for which effective treatment is lacking. Recent experimental find- ings raise the possibility that functional improvement after stroke may be induced through neuronal replacement by endog- enous neural stem cells (NSCs) in the subventricular zone (SVZ). Ischemic stroke caused by middle cerebral artery occlu- sion (MCAO) triggers increased cell proliferation in the rat SVZ [1–3]. The newly formed neuroblasts migrate into the damaged striatum [4 –7], a region where neurogenesis does not occur in the intact brain. After maturation, a substantial portion of the new neurons express markers characteristic of those mature neurons that have died, that is, striatal projection neurons [4, 5]. However, this potential self-repair mechanism is thought to operate only acutely after stroke, with the number of generated neurons being small and their existence transitory [8]. We show herein that stroke-induced neurogenesis is ex- tensive and long-lasting, with continuous production of ma- ture striatal neurons for several months after the insult. We also find that stromal cell– derived factor-1 (SDF-1)/ CXCR4 signaling regulates the directed migration of new neurons to the injured area. MATERIALS AND METHODS Induction of Stroke and 5-Bromo-2-Deoxyuridine Labeling Under halothane anesthesia, the middle cerebral artery of arti- ficially ventilated male Wistar rats was occluded with a filament inserted through the common carotid artery [9, 10]. After 30 minutes or 2 hours, the filament was withdrawn. For sham surgery, the filament was advanced a few millimeters inside the internal carotid artery. Physiological parameters were kept within a predetermined range. Intraperitoneal injections of 5-bromo-2-deoxyuridine (BrdU; 50 mg/kg, Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com) were given twice daily during weeks 1–2, 5– 6, or 7– 8 after surgery. In one experiment, three BrdU injections with 2-hour intervals were given 4 days or 6 weeks after MCAO, and the animals were killed 2 hours thereafter. Caspase Inhibitor and CXCR4 Antagonist Treatments Daily intraventricular infusions of a caspase inhibitor cocktail, comprising equal portions of a multicaspase, a caspase 3, and a caspase 9 inhibitor, were given over 12 days, starting on day 3 Correspondence: Olle Lindvall, M.D., Ph.D., Laboratory of Neurogenesis and Cell Therapy, Section of Restorative Neurology, Wallenberg Neuroscience Center, University Hospital, SE-221 84 Lund, Sweden. Telephone: 46-46-222 0543; Fax: 46-46-222 0560; e-mail: [email protected] Received June 23, 2005; accepted for publication September 27, 2005; first published in STEM CELLS EXPRESS October 6, 2006. ©AlphaMed Press 1066-5099/2006/$20.00/0 doi: 10.1634/stemcells.2005-0281 TISSUE-SPECIFIC STEM CELLS S TEM CELLS 2006;24:739 –747 www.StemCells.com

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Persistent Production of Neurons from Adult Brain Stem CellsDuring Recovery after Stroke
PAR THORED,a,c ANDREAS ARVIDSSON,a,c EMANUELE CACCI,b,c HENRIK AHLENIUS,b,c THERESE KALLUR,b,c
VLADIMER DARSALIA,b,c CHRISTINE T. EKDAHL,a,c ZAAL KOKAIA,b,c OLLE LINDVALLa,c
aLaboratory of Neurogenesis and Cell Therapy, Section of Restorative Neurology, Wallenberg Neuroscience Center,
University Hospital, Lund, Sweden; bLaboratory of Neural Stem Cell Biology, Section of Restorative Neurology,
Stem Cell Institute, University Hospital, Lund, Sweden; cLund Strategic Research Center for Stem Cell Biology and
Cell Therapy, Lund, Sweden
Key Words. Stem cells • Neurogenesis • Striatum • Stroke
ABSTRACT
Neural stem cells in the subventricular zone of adult rodentsproduce new striatal neurons that may replace those thathave died after stroke; however, the neurogenic responsehas been considered acute and transient, yielding only smallnumbers of neurons. In contrast, we show herein that stri-atal neuroblasts are generated without decline at least for 4months after stroke in adult rats. Neuroblasts formed earlyor late after stroke either differentiate into mature neurons,
which survive for several months, or die through caspase-mediated apoptosis. The directed migration of the new neu-rons toward the ischemic damage is regulated by stromalcell-derived factor-1� and its receptor CXCR4. These re-sults show that endogenous neural stem cells continuouslysupply the injured adult brain with new neurons, whichsuggests novel self-repair strategies to improve recoveryafter stroke. STEM CELLS 2006;24:739–747
INTRODUCTIONStroke is a leading cause of chronic disability in humans forwhich effective treatment is lacking. Recent experimental find-ings raise the possibility that functional improvement afterstroke may be induced through neuronal replacement by endog-enous neural stem cells (NSCs) in the subventricular zone(SVZ). Ischemic stroke caused by middle cerebral artery occlu-sion (MCAO) triggers increased cell proliferation in the rat SVZ[1–3]. The newly formed neuroblasts migrate into the damagedstriatum [4–7], a region where neurogenesis does not occur inthe intact brain. After maturation, a substantial portion of thenew neurons express markers characteristic of those matureneurons that have died, that is, striatal projection neurons [4, 5].However, this potential self-repair mechanism is thought tooperate only acutely after stroke, with the number of generatedneurons being small and their existence transitory [8].
We show herein that stroke-induced neurogenesis is ex-tensive and long-lasting, with continuous production of ma-ture striatal neurons for several months after the insult. Wealso find that stromal cell– derived factor-1� (SDF-1�)/CXCR4 signaling regulates the directed migration of newneurons to the injured area.
MATERIALS AND METHODS
Induction of Stroke and 5-Bromo-2�-DeoxyuridineLabelingUnder halothane anesthesia, the middle cerebral artery of arti-ficially ventilated male Wistar rats was occluded with a filamentinserted through the common carotid artery [9, 10]. After 30minutes or 2 hours, the filament was withdrawn. For shamsurgery, the filament was advanced a few millimeters inside theinternal carotid artery. Physiological parameters were keptwithin a predetermined range. Intraperitoneal injections of5-bromo-2�-deoxyuridine (BrdU; 50 mg/kg, Sigma-Aldrich, St.Louis, http://www.sigmaaldrich.com) were given twice dailyduring weeks 1–2, 5–6, or 7–8 after surgery. In one experiment,three BrdU injections with 2-hour intervals were given 4 days or6 weeks after MCAO, and the animals were killed 2 hoursthereafter.
Caspase Inhibitor and CXCR4 AntagonistTreatmentsDaily intraventricular infusions of a caspase inhibitor cocktail,comprising equal portions of a multicaspase, a caspase 3, and acaspase 9 inhibitor, were given over 12 days, starting on day 3
Correspondence: Olle Lindvall, M.D., Ph.D., Laboratory of Neurogenesis and Cell Therapy, Section of Restorative Neurology,Wallenberg Neuroscience Center, University Hospital, SE-221 84 Lund, Sweden. Telephone: 46-46-222 0543; Fax: 46-46-222 0560;e-mail: [email protected] Received June 23, 2005; accepted for publication September 27, 2005; first published in STEMCELLS EXPRESS October 6, 2006. ©AlphaMed Press 1066-5099/2006/$20.00/0 doi: 10.1634/stemcells.2005-0281
TISSUE-SPECIFIC STEM CELLS
STEM CELLS 2006;24:739–747 www.StemCells.com

after MCAO [11]. Mini-osmotic pumps (model 2002, flow rate,0.5 �l/hour; Alzet, Palo Alto, CA, http://www.alzet.com) wereimplanted 4 weeks after 2-hour MCAO, infusing either theCXCR4 antagonist AMD3100 (Sigma-Aldrich, 5 mg/ml inphosphate-buffered saline [PBS]) or PBS for 2 weeks into thelateral ventricle.
In Vitro AssaysNeurospheres were grown in Dulbecco’s modified Eagle’s me-dium(DMEM)/F12(Gibco-BRL,Gaithersburg,MD,http://www.gibcobrl.com) supplemented with B27 containing 20 ng/ml epi-dermal growth factor and 10 ng/ml basic fibroblast growthfactor (R&D Systems Inc., Minneapolis, http://www.rndsystems.com) [12]. After trituration of SVZ tissue or primary neuro-spheres, cells were plated in 24-well plates for 7 days, and thenumber and diameter of spheres were quantified.
For studies of phosphorylation, adult rat SVZ cells wereplated in 96-well plates. After 5 days in vitro, serum and growthfactors were withdrawn for 18 hours. Cells were treated with thebasal medium either alone or containing 500 ng/ml SDF-1�(R&D Systems) for 10 minutes. Phosphorylation of extracellularsignal-related kinase (ERK)-1/2 was assessed using FACEenzyme-linked immunosorbent assay kits (Active Motif, Rixen-sart, Belgium, http://www.activemotif.com).
For the migration assay, neurospheres from the SVZ weredistributed on a poly-L-lysine coated polyvinylpyrrolidine-freepolycarbonate filter in the upper section of a 96-well microche-motaxis chamber (Neuro Probe, Gaithersburg, MD, http://www.neuroprobe.com). SDF-1� (100 or 500 ng/ml) was added toeither the lower wells, upper wells, or both. Basal mediumwithout SDF-1� served as control. Neurospheres were allowedto migrate for 12 hours. The filter was fixed in 4% phosphate-buffered paraformaldehyde (PFA) and stained with Hoechst33342 (Molecular Probes Inc., Eugene, OR, http://probes.invitrogen.com), and migrated cells were counted using anepifluorescence microscope.
ImmunohistochemistryAfter transcardial perfusion with 4% ice-cold PFA, brains werepostfixed overnight in PFA and sectioned coronally at 30 �m ondry ice. Prior to staining using diaminobenzidine (DAB), free-floating sections were quenched for 20 minutes in 3% hydrogenperoxide and 10% methanol, and prior to staining for BrdU,sections were incubated in 1 M HCl at 65°C for 10 minutes andat 37°C for 20 minutes. Following preincubation with the ap-propriate normal sera, sections were incubated for 36 hours at4°C with either two of the following antibodies—rat anti-BrdU(1:100; Harlan Sera-Lab, Loughborough, U.K., http://www.har-laneurope.com), goat anti-doublecortin (Dcx) antibody (1:400;Santa Cruz Biotechnology Inc., Santa Cruz, CA, http://www.scbt.com), guinea-pig anti-Dcx (1:3,000; Abcam, Cambridge,U.K., http://www.abcam.com), mouse anti-neuronal nuclei(NeuN; 1:100; Chemicon, Temecula, CA, http://www.chemicon.com), goat anti-CXCR4 (recognizes SDF-1� andSDF-1�, 1:100; Santa Cruz Biotechnology), goat anti-SDF-1(recognizes SDF-1� and SDF-1�, 1:100; Santa Cruz Biotech-nology), mouse anti-ED1 (1:200; SeroTec, Ltd., Oslo, Norway,http://www.serotec.com), or mouse glial fibrillary acidic protein(GFAP; 1:1,000; Sigma-Aldrich)—or overnight with mouseanti-NeuN only, in normal sera. Stainings were visualized by
incubation for 2 hours with Cy3-conjugated (1:200; JacksonImmunoResearch Laboratories, West Grove, PA, http://www.jacksonimmuno.com) and biotinylated secondary antibodies(1:200; Vector Laboratories, Burlingame, CA, http://www.vectorlabs.com) followed by Alexa 488-conjugated streptavidin(1:200; Molecular Probes) for 2 hours for double stainings, oravidin-biotin-peroxidase complex for 1 hour followed by treat-ment with DAB (0.5 mg/ml) and hydrogen peroxide for singlestaining. Sections were then mounted on glass slides and cov-erslipped. For caspase-cleaved poly(ADP-ribose) polymerase(cPARP)/Dcx double staining, sections were stained for Dcx asabove and mounted on glass slides before incubation overnightwith rabbit anti-rat cPARP antibody (1:20; Cell Signaling Tech-nology, Beverly, MA, http://www.cellsignal.com) in 3% bovineserum albumin at 4°C. For in vitro stainings, neurospheres wereplated on poly-L-lysine-coated four-well chamber slides anddifferentiated for 7 days in basal medium containing 1% fetalbovine serum (differentiation medium). Cells were fixed in 4%PFA at room temperature and stained using mouse anti-�III-tubulin antibody (1:350; Sigma-Aldrich). For CXCR4 staining,SVZ-derived neurospheres or primary cells were plated over-night in expansion medium, fixed, and stained with goat anti-CXCR4 antibody (1:100). Both stainings were counterstainedwith Hoechst 33342.
Microscopical AnalysisCells double-labeled with BrdU and Dcx or NeuN were countedusing an epifluorescence microscope with 40� objective(Olympus A/S, Glostrup, Denmark, http://www2.olympus.dk).The accuracy of double labeling was validated using a confocallaser-scanning microscope (Leica, Heerbrugg, Switzerland,http://www.leica.com). Dcx�, Dcx�/BrdU�, and NeuN�/BrdU� cells were counted, using a 0.25-mm-wide quadraticgrid, in the entire striatum in four sections, at �1.60, �1.00,�0.48, and �0.26 mm from bregma, the most medial columnlining the SVZ. Striatal volumes were measured using stereo-logical equipment in the same sections. The size of the SVZ wasmeasured in sections from the same rostrocaudal levels stainedwith NeuN and cresyl violet. The SVZ was delineated by cellsstained with cresyl violet only. BrdU� cells in the SVZ werecounted in three sections (�1.60, �0.70, and �0.26 mm frombregma) with the optical fractionator method on stereologicalequipment.
Behavioral TestsBehavioral tests were performed 24 hours before (control) and48 hours, 4 weeks, and 16 weeks after 30-minute MCAO. In themodified horizontal ladder walking test [13], rats were allowedto walk on the ladder for 5 minutes. The degree of behavioraldeficit was calculated for each animal using a formula that takesinto account the total number of steps and falls, as well as thetime and speed of active walking. The stepping test, for evalu-ation of forelimb akinesia [14], and the cylinder test of forelimbasymmetry [15] were performed as described previously [16].
Animal Groups and Statistical AnalysisQuantification of the number of Dcx�, Dcx�/BrdU�, andNeuN�/BrdU� cells 2, 6, and 16 weeks after stroke, and cellmigration and SVZ volume, was performed on the same groupsof animals, whereas cell numbers at 8 and 12 weeks were taken
740 Neurogenesis after Stroke
from the vehicle-treated animals in the AMD 3100 experiment.Data in the caspase inhibitor and AMD3100 experiments, fol-lowing BrdU pulses 4 days and 6 weeks after stroke as well asin all behavioral tests, were obtained from separate animalgroups. Values are reported as mean � standard error of themean. Group differences were assessed with unpaired t tests andside differences were assessed with paired t tests. A one-wayanalysis of variance with Fischer’s post hoc test was used toassess differences in the number of migrated cells and level ofphosphorylation in vitro. For studies of correlation, Pearson’scorrelation coefficients were generated and analyzed for signif-icance using Fisher’s r to z transformation.
RESULTS
Striatal Neuroblasts Are Generated from SVZPrecursors for Several Months after StrokeWe first studied the occurrence of Dcx� neuroblasts in the adultrat striatum at various time points after 2-hour MCAO. Con-
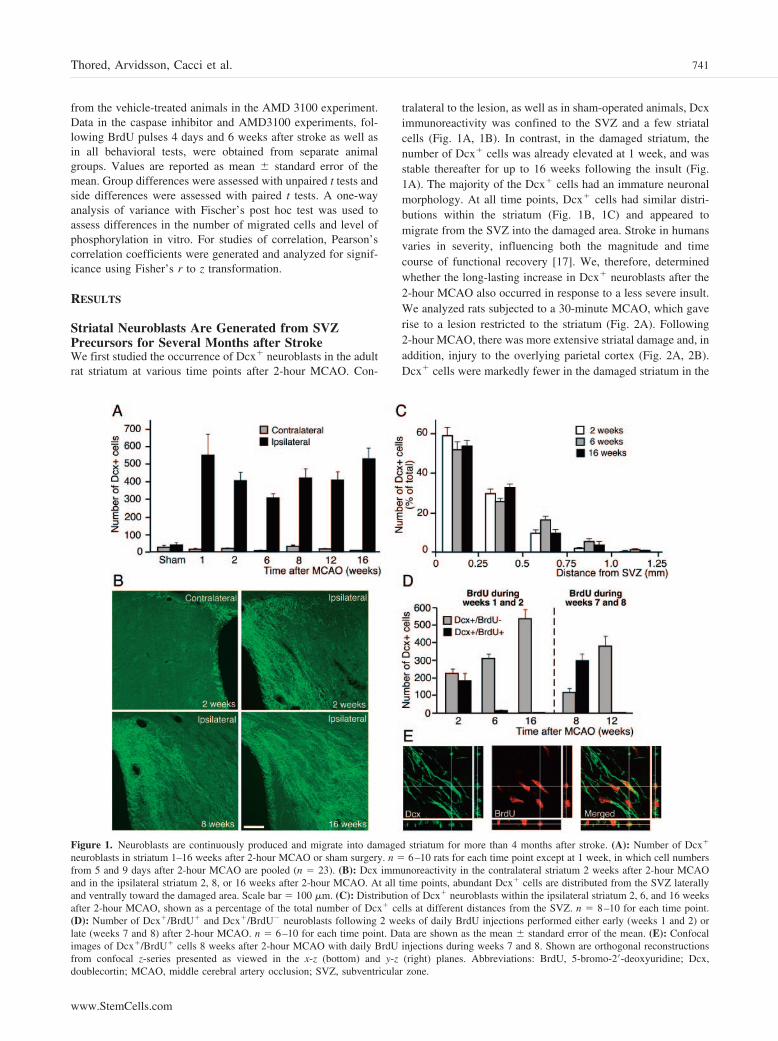
tralateral to the lesion, as well as in sham-operated animals, Dcximmunoreactivity was confined to the SVZ and a few striatalcells (Fig. 1A, 1B). In contrast, in the damaged striatum, thenumber of Dcx� cells was already elevated at 1 week, and wasstable thereafter for up to 16 weeks following the insult (Fig.1A). The majority of the Dcx� cells had an immature neuronalmorphology. At all time points, Dcx� cells had similar distri-butions within the striatum (Fig. 1B, 1C) and appeared tomigrate from the SVZ into the damaged area. Stroke in humansvaries in severity, influencing both the magnitude and timecourse of functional recovery [17]. We, therefore, determinedwhether the long-lasting increase in Dcx� neuroblasts after the2-hour MCAO also occurred in response to a less severe insult.We analyzed rats subjected to a 30-minute MCAO, which gaverise to a lesion restricted to the striatum (Fig. 2A). Following2-hour MCAO, there was more extensive striatal damage and, inaddition, injury to the overlying parietal cortex (Fig. 2A, 2B).Dcx� cells were markedly fewer in the damaged striatum in the
Figure 1. Neuroblasts are continuously produced and migrate into damaged striatum for more than 4 months after stroke. (A): Number of Dcx�
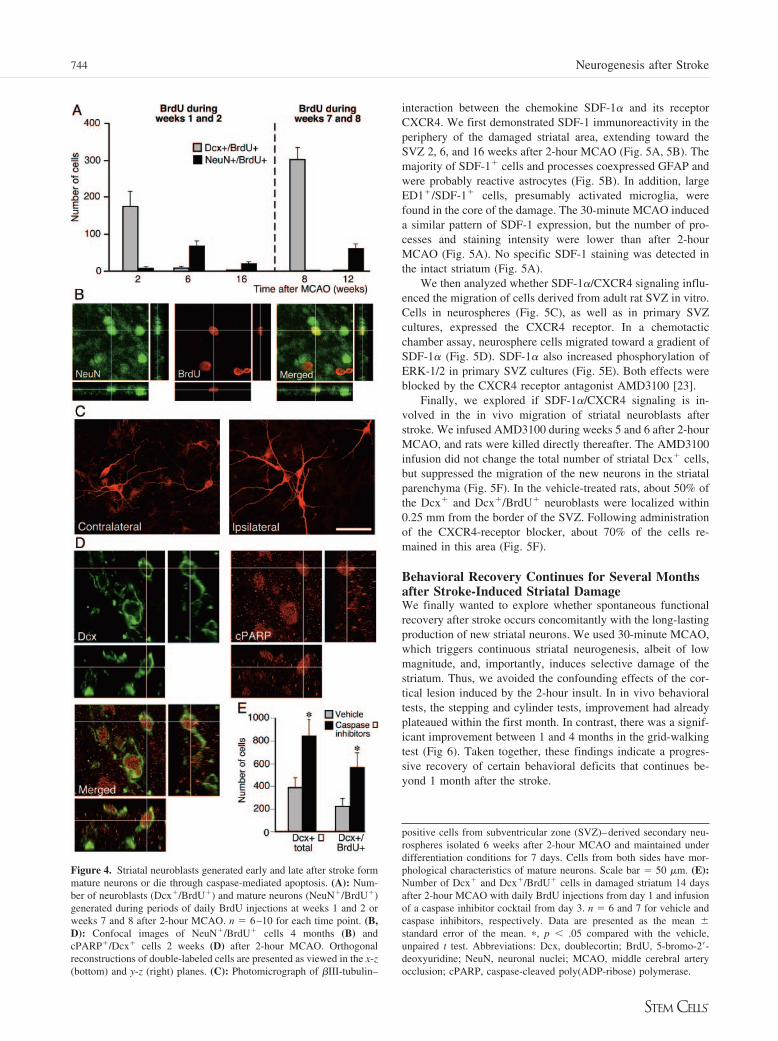
neuroblasts in striatum 1–16 weeks after 2-hour MCAO or sham surgery. n � 6–10 rats for each time point except at 1 week, in which cell numbersfrom 5 and 9 days after 2-hour MCAO are pooled (n � 23). (B): Dcx immunoreactivity in the contralateral striatum 2 weeks after 2-hour MCAOand in the ipsilateral striatum 2, 8, or 16 weeks after 2-hour MCAO. At all time points, abundant Dcx� cells are distributed from the SVZ laterallyand ventrally toward the damaged area. Scale bar � 100 �m. (C): Distribution of Dcx� neuroblasts within the ipsilateral striatum 2, 6, and 16 weeksafter 2-hour MCAO, shown as a percentage of the total number of Dcx� cells at different distances from the SVZ. n � 8–10 for each time point.(D): Number of Dcx�/BrdU� and Dcx�/BrdU� neuroblasts following 2 weeks of daily BrdU injections performed either early (weeks 1 and 2) orlate (weeks 7 and 8) after 2-hour MCAO. n � 6–10 for each time point. Data are shown as the mean � standard error of the mean. (E): Confocalimages of Dcx�/BrdU� cells 8 weeks after 2-hour MCAO with daily BrdU injections during weeks 7 and 8. Shown are orthogonal reconstructionsfrom confocal z-series presented as viewed in the x-z (bottom) and y-z (right) planes. Abbreviations: BrdU, 5-bromo-2�-deoxyuridine; Dcx,doublecortin; MCAO, middle cerebral artery occlusion; SVZ, subventricular zone.
741Thored, Arvidsson, Cacci et al.
www.StemCells.com
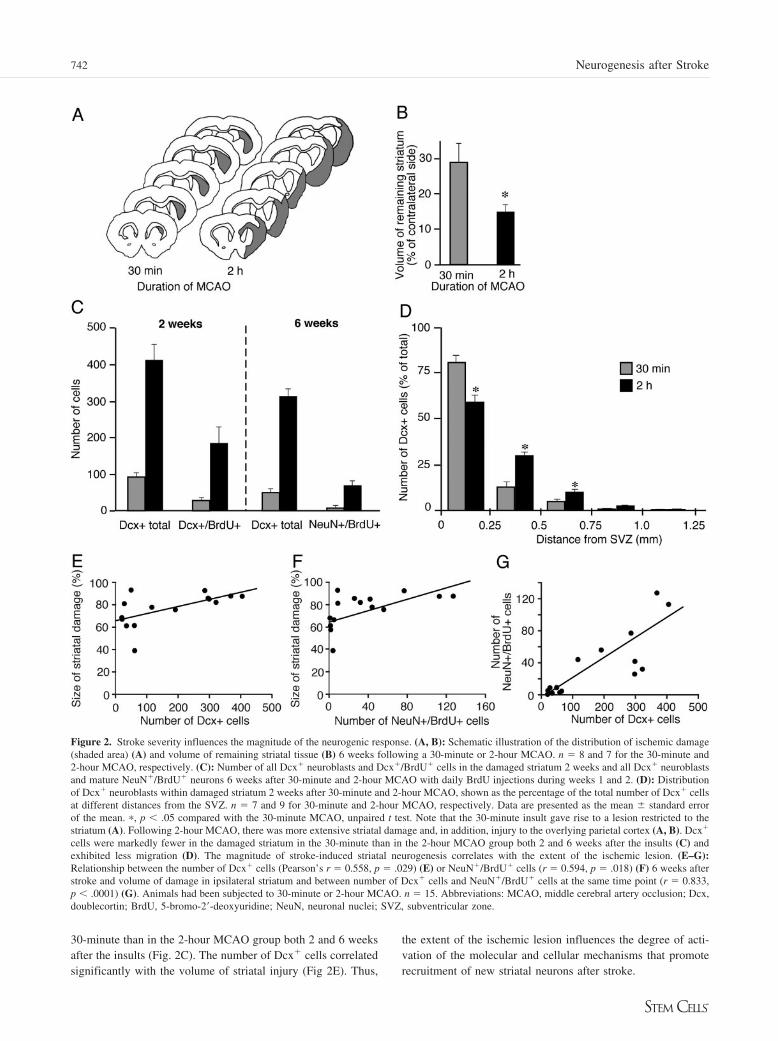
30-minute than in the 2-hour MCAO group both 2 and 6 weeksafter the insults (Fig. 2C). The number of Dcx� cells correlatedsignificantly with the volume of striatal injury (Fig 2E). Thus,
the extent of the ischemic lesion influences the degree of acti-vation of the molecular and cellular mechanisms that promoterecruitment of new striatal neurons after stroke.
Figure 2. Stroke severity influences the magnitude of the neurogenic response. (A, B): Schematic illustration of the distribution of ischemic damage(shaded area) (A) and volume of remaining striatal tissue (B) 6 weeks following a 30-minute or 2-hour MCAO. n � 8 and 7 for the 30-minute and2-hour MCAO, respectively. (C): Number of all Dcx� neuroblasts and Dcx�/BrdU� cells in the damaged striatum 2 weeks and all Dcx� neuroblastsand mature NeuN�/BrdU� neurons 6 weeks after 30-minute and 2-hour MCAO with daily BrdU injections during weeks 1 and 2. (D): Distributionof Dcx� neuroblasts within damaged striatum 2 weeks after 30-minute and 2-hour MCAO, shown as the percentage of the total number of Dcx� cellsat different distances from the SVZ. n � 7 and 9 for 30-minute and 2-hour MCAO, respectively. Data are presented as the mean � standard errorof the mean. �, p � .05 compared with the 30-minute MCAO, unpaired t test. Note that the 30-minute insult gave rise to a lesion restricted to thestriatum (A). Following 2-hour MCAO, there was more extensive striatal damage and, in addition, injury to the overlying parietal cortex (A, B). Dcx�
cells were markedly fewer in the damaged striatum in the 30-minute than in the 2-hour MCAO group both 2 and 6 weeks after the insults (C) andexhibited less migration (D). The magnitude of stroke-induced striatal neurogenesis correlates with the extent of the ischemic lesion. (E–G):Relationship between the number of Dcx� cells (Pearson’s r � 0.558, p � .029) (E) or NeuN�/BrdU� cells (r � 0.594, p � .018) (F) 6 weeks afterstroke and volume of damage in ipsilateral striatum and between number of Dcx� cells and NeuN�/BrdU� cells at the same time point (r � 0.833,p � .0001) (G). Animals had been subjected to 30-minute or 2-hour MCAO. n � 15. Abbreviations: MCAO, middle cerebral artery occlusion; Dcx,doublecortin; BrdU, 5-bromo-2�-deoxyuridine; NeuN, neuronal nuclei; SVZ, subventricular zone.
742 Neurogenesis after Stroke
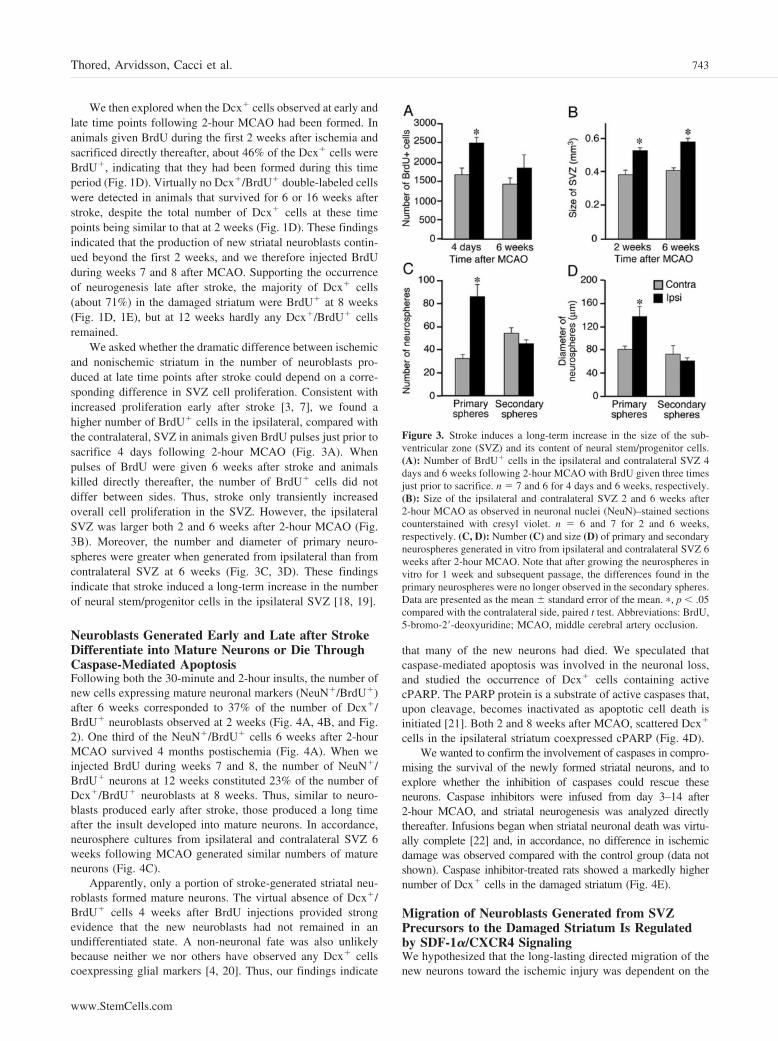
We then explored when the Dcx� cells observed at early andlate time points following 2-hour MCAO had been formed. Inanimals given BrdU during the first 2 weeks after ischemia andsacrificed directly thereafter, about 46% of the Dcx� cells wereBrdU�, indicating that they had been formed during this timeperiod (Fig. 1D). Virtually no Dcx�/BrdU� double-labeled cellswere detected in animals that survived for 6 or 16 weeks afterstroke, despite the total number of Dcx� cells at these timepoints being similar to that at 2 weeks (Fig. 1D). These findingsindicated that the production of new striatal neuroblasts contin-ued beyond the first 2 weeks, and we therefore injected BrdUduring weeks 7 and 8 after MCAO. Supporting the occurrenceof neurogenesis late after stroke, the majority of Dcx� cells(about 71%) in the damaged striatum were BrdU� at 8 weeks(Fig. 1D, 1E), but at 12 weeks hardly any Dcx�/BrdU� cellsremained.
We asked whether the dramatic difference between ischemicand nonischemic striatum in the number of neuroblasts pro-duced at late time points after stroke could depend on a corre-sponding difference in SVZ cell proliferation. Consistent withincreased proliferation early after stroke [3, 7], we found ahigher number of BrdU� cells in the ipsilateral, compared withthe contralateral, SVZ in animals given BrdU pulses just prior tosacrifice 4 days following 2-hour MCAO (Fig. 3A). Whenpulses of BrdU were given 6 weeks after stroke and animalskilled directly thereafter, the number of BrdU� cells did notdiffer between sides. Thus, stroke only transiently increasedoverall cell proliferation in the SVZ. However, the ipsilateralSVZ was larger both 2 and 6 weeks after 2-hour MCAO (Fig.3B). Moreover, the number and diameter of primary neuro-spheres were greater when generated from ipsilateral than fromcontralateral SVZ at 6 weeks (Fig. 3C, 3D). These findingsindicate that stroke induced a long-term increase in the numberof neural stem/progenitor cells in the ipsilateral SVZ [18, 19].
Neuroblasts Generated Early and Late after StrokeDifferentiate into Mature Neurons or Die ThroughCaspase-Mediated ApoptosisFollowing both the 30-minute and 2-hour insults, the number ofnew cells expressing mature neuronal markers (NeuN�/BrdU�)after 6 weeks corresponded to 37% of the number of Dcx�/BrdU� neuroblasts observed at 2 weeks (Fig. 4A, 4B, and Fig.2). One third of the NeuN�/BrdU� cells 6 weeks after 2-hourMCAO survived 4 months postischemia (Fig. 4A). When weinjected BrdU during weeks 7 and 8, the number of NeuN�/BrdU� neurons at 12 weeks constituted 23% of the number ofDcx�/BrdU� neuroblasts at 8 weeks. Thus, similar to neuro-blasts produced early after stroke, those produced a long timeafter the insult developed into mature neurons. In accordance,neurosphere cultures from ipsilateral and contralateral SVZ 6weeks following MCAO generated similar numbers of matureneurons (Fig. 4C).
Apparently, only a portion of stroke-generated striatal neu-roblasts formed mature neurons. The virtual absence of Dcx�/BrdU� cells 4 weeks after BrdU injections provided strongevidence that the new neuroblasts had not remained in anundifferentiated state. A non-neuronal fate was also unlikelybecause neither we nor others have observed any Dcx� cellscoexpressing glial markers [4, 20]. Thus, our findings indicate
that many of the new neurons had died. We speculated thatcaspase-mediated apoptosis was involved in the neuronal loss,and studied the occurrence of Dcx� cells containing activecPARP. The PARP protein is a substrate of active caspases that,upon cleavage, becomes inactivated as apoptotic cell death isinitiated [21]. Both 2 and 8 weeks after MCAO, scattered Dcx�
cells in the ipsilateral striatum coexpressed cPARP (Fig. 4D).We wanted to confirm the involvement of caspases in compro-
mising the survival of the newly formed striatal neurons, and toexplore whether the inhibition of caspases could rescue theseneurons. Caspase inhibitors were infused from day 3–14 after2-hour MCAO, and striatal neurogenesis was analyzed directlythereafter. Infusions began when striatal neuronal death was virtu-ally complete [22] and, in accordance, no difference in ischemicdamage was observed compared with the control group (data notshown). Caspase inhibitor-treated rats showed a markedly highernumber of Dcx� cells in the damaged striatum (Fig. 4E).
Migration of Neuroblasts Generated from SVZPrecursors to the Damaged Striatum Is Regulatedby SDF-1�/CXCR4 SignalingWe hypothesized that the long-lasting directed migration of thenew neurons toward the ischemic injury was dependent on the
Figure 3. Stroke induces a long-term increase in the size of the sub-ventricular zone (SVZ) and its content of neural stem/progenitor cells.(A): Number of BrdU� cells in the ipsilateral and contralateral SVZ 4days and 6 weeks following 2-hour MCAO with BrdU given three timesjust prior to sacrifice. n � 7 and 6 for 4 days and 6 weeks, respectively.(B): Size of the ipsilateral and contralateral SVZ 2 and 6 weeks after2-hour MCAO as observed in neuronal nuclei (NeuN)–stained sectionscounterstained with cresyl violet. n � 6 and 7 for 2 and 6 weeks,respectively. (C, D): Number (C) and size (D) of primary and secondaryneurospheres generated in vitro from ipsilateral and contralateral SVZ 6weeks after 2-hour MCAO. Note that after growing the neurospheres invitro for 1 week and subsequent passage, the differences found in theprimary neurospheres were no longer observed in the secondary spheres.Data are presented as the mean � standard error of the mean. �, p � .05compared with the contralateral side, paired t test. Abbreviations: BrdU,5-bromo-2�-deoxyuridine; MCAO, middle cerebral artery occlusion.
743Thored, Arvidsson, Cacci et al.
www.StemCells.com
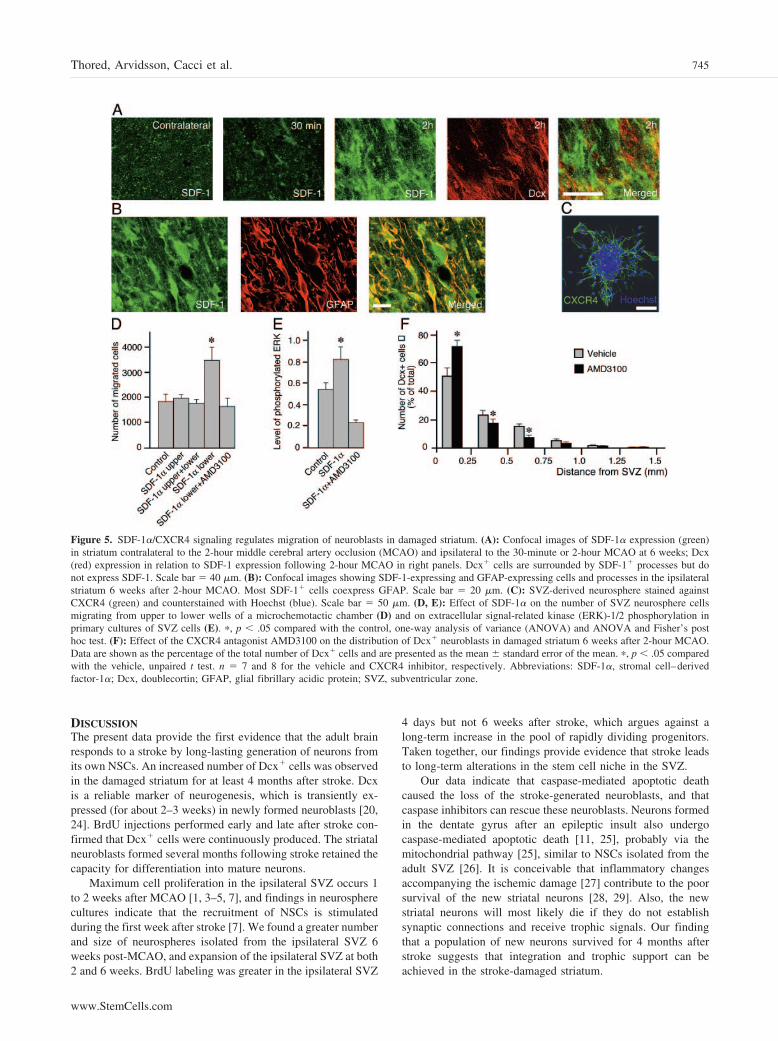
interaction between the chemokine SDF-1� and its receptorCXCR4. We first demonstrated SDF-1 immunoreactivity in theperiphery of the damaged striatal area, extending toward theSVZ 2, 6, and 16 weeks after 2-hour MCAO (Fig. 5A, 5B). Themajority of SDF-1� cells and processes coexpressed GFAP andwere probably reactive astrocytes (Fig. 5B). In addition, largeED1�/SDF-1� cells, presumably activated microglia, werefound in the core of the damage. The 30-minute MCAO induceda similar pattern of SDF-1 expression, but the number of pro-cesses and staining intensity were lower than after 2-hourMCAO (Fig. 5A). No specific SDF-1 staining was detected inthe intact striatum (Fig. 5A).
We then analyzed whether SDF-1�/CXCR4 signaling influ-enced the migration of cells derived from adult rat SVZ in vitro.Cells in neurospheres (Fig. 5C), as well as in primary SVZcultures, expressed the CXCR4 receptor. In a chemotacticchamber assay, neurosphere cells migrated toward a gradient ofSDF-1� (Fig. 5D). SDF-1� also increased phosphorylation ofERK-1/2 in primary SVZ cultures (Fig. 5E). Both effects wereblocked by the CXCR4 receptor antagonist AMD3100 [23].
Finally, we explored if SDF-1�/CXCR4 signaling is in-volved in the in vivo migration of striatal neuroblasts afterstroke. We infused AMD3100 during weeks 5 and 6 after 2-hourMCAO, and rats were killed directly thereafter. The AMD3100infusion did not change the total number of striatal Dcx� cells,but suppressed the migration of the new neurons in the striatalparenchyma (Fig. 5F). In the vehicle-treated rats, about 50% ofthe Dcx� and Dcx�/BrdU� neuroblasts were localized within0.25 mm from the border of the SVZ. Following administrationof the CXCR4-receptor blocker, about 70% of the cells re-mained in this area (Fig. 5F).
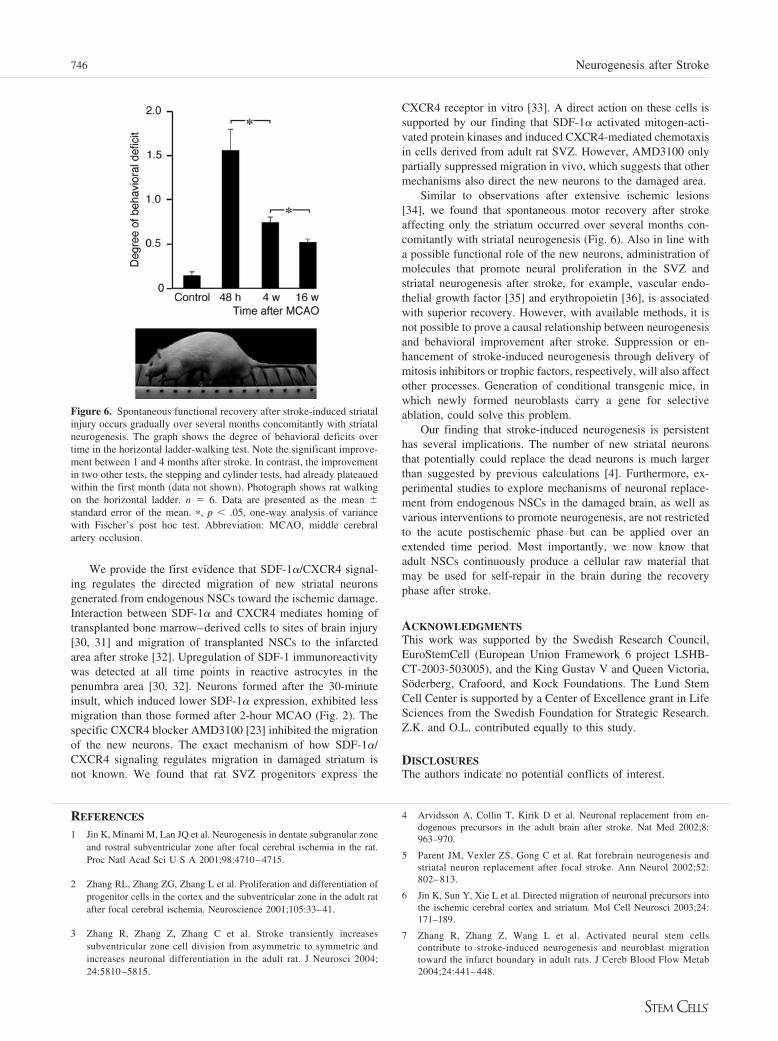
Behavioral Recovery Continues for Several Monthsafter Stroke-Induced Striatal DamageWe finally wanted to explore whether spontaneous functionalrecovery after stroke occurs concomitantly with the long-lastingproduction of new striatal neurons. We used 30-minute MCAO,which triggers continuous striatal neurogenesis, albeit of lowmagnitude, and, importantly, induces selective damage of thestriatum. Thus, we avoided the confounding effects of the cor-tical lesion induced by the 2-hour insult. In in vivo behavioraltests, the stepping and cylinder tests, improvement had alreadyplateaued within the first month. In contrast, there was a signif-icant improvement between 1 and 4 months in the grid-walkingtest (Fig 6). Taken together, these findings indicate a progres-sive recovery of certain behavioral deficits that continues be-yond 1 month after the stroke.
positive cells from subventricular zone (SVZ)–derived secondary neu-rospheres isolated 6 weeks after 2-hour MCAO and maintained underdifferentiation conditions for 7 days. Cells from both sides have mor-phological characteristics of mature neurons. Scale bar � 50 �m. (E):Number of Dcx� and Dcx�/BrdU� cells in damaged striatum 14 daysafter 2-hour MCAO with daily BrdU injections from day 1 and infusionof a caspase inhibitor cocktail from day 3. n � 6 and 7 for vehicle andcaspase inhibitors, respectively. Data are presented as the mean �standard error of the mean. �, p � .05 compared with the vehicle,unpaired t test. Abbreviations: Dcx, doublecortin; BrdU, 5-bromo-2�-deoxyuridine; NeuN, neuronal nuclei; MCAO, middle cerebral arteryocclusion; cPARP, caspase-cleaved poly(ADP-ribose) polymerase.
Figure 4. Striatal neuroblasts generated early and late after stroke formmature neurons or die through caspase-mediated apoptosis. (A): Num-ber of neuroblasts (Dcx�/BrdU�) and mature neurons (NeuN�/BrdU�)generated during periods of daily BrdU injections at weeks 1 and 2 orweeks 7 and 8 after 2-hour MCAO. n � 6–10 for each time point. (B,D): Confocal images of NeuN�/BrdU� cells 4 months (B) andcPARP�/Dcx� cells 2 weeks (D) after 2-hour MCAO. Orthogonalreconstructions of double-labeled cells are presented as viewed in the x-z(bottom) and y-z (right) planes. (C): Photomicrograph of �III-tubulin–
744 Neurogenesis after Stroke
DISCUSSIONThe present data provide the first evidence that the adult brainresponds to a stroke by long-lasting generation of neurons fromits own NSCs. An increased number of Dcx� cells was observedin the damaged striatum for at least 4 months after stroke. Dcxis a reliable marker of neurogenesis, which is transiently ex-pressed (for about 2–3 weeks) in newly formed neuroblasts [20,24]. BrdU injections performed early and late after stroke con-firmed that Dcx� cells were continuously produced. The striatalneuroblasts formed several months following stroke retained thecapacity for differentiation into mature neurons.
Maximum cell proliferation in the ipsilateral SVZ occurs 1to 2 weeks after MCAO [1, 3–5, 7], and findings in neurospherecultures indicate that the recruitment of NSCs is stimulatedduring the first week after stroke [7]. We found a greater numberand size of neurospheres isolated from the ipsilateral SVZ 6weeks post-MCAO, and expansion of the ipsilateral SVZ at both2 and 6 weeks. BrdU labeling was greater in the ipsilateral SVZ
4 days but not 6 weeks after stroke, which argues against along-term increase in the pool of rapidly dividing progenitors.Taken together, our findings provide evidence that stroke leadsto long-term alterations in the stem cell niche in the SVZ.
Our data indicate that caspase-mediated apoptotic deathcaused the loss of the stroke-generated neuroblasts, and thatcaspase inhibitors can rescue these neuroblasts. Neurons formedin the dentate gyrus after an epileptic insult also undergocaspase-mediated apoptotic death [11, 25], probably via themitochondrial pathway [25], similar to NSCs isolated from theadult SVZ [26]. It is conceivable that inflammatory changesaccompanying the ischemic damage [27] contribute to the poorsurvival of the new striatal neurons [28, 29]. Also, the newstriatal neurons will most likely die if they do not establishsynaptic connections and receive trophic signals. Our findingthat a population of new neurons survived for 4 months afterstroke suggests that integration and trophic support can beachieved in the stroke-damaged striatum.
Figure 5. SDF-1�/CXCR4 signaling regulates migration of neuroblasts in damaged striatum. (A): Confocal images of SDF-1� expression (green)in striatum contralateral to the 2-hour middle cerebral artery occlusion (MCAO) and ipsilateral to the 30-minute or 2-hour MCAO at 6 weeks; Dcx(red) expression in relation to SDF-1 expression following 2-hour MCAO in right panels. Dcx� cells are surrounded by SDF-1� processes but donot express SDF-1. Scale bar � 40 �m. (B): Confocal images showing SDF-1-expressing and GFAP-expressing cells and processes in the ipsilateralstriatum 6 weeks after 2-hour MCAO. Most SDF-1� cells coexpress GFAP. Scale bar � 20 �m. (C): SVZ-derived neurosphere stained againstCXCR4 (green) and counterstained with Hoechst (blue). Scale bar � 50 �m. (D, E): Effect of SDF-1� on the number of SVZ neurosphere cellsmigrating from upper to lower wells of a microchemotactic chamber (D) and on extracellular signal-related kinase (ERK)-1/2 phosphorylation inprimary cultures of SVZ cells (E). �, p � .05 compared with the control, one-way analysis of variance (ANOVA) and ANOVA and Fisher’s posthoc test. (F): Effect of the CXCR4 antagonist AMD3100 on the distribution of Dcx� neuroblasts in damaged striatum 6 weeks after 2-hour MCAO.Data are shown as the percentage of the total number of Dcx� cells and are presented as the mean � standard error of the mean. �, p � .05 comparedwith the vehicle, unpaired t test. n � 7 and 8 for the vehicle and CXCR4 inhibitor, respectively. Abbreviations: SDF-1�, stromal cell–derivedfactor-1�; Dcx, doublecortin; GFAP, glial fibrillary acidic protein; SVZ, subventricular zone.
745Thored, Arvidsson, Cacci et al.
www.StemCells.com
We provide the first evidence that SDF-1�/CXCR4 signal-ing regulates the directed migration of new striatal neuronsgenerated from endogenous NSCs toward the ischemic damage.Interaction between SDF-1� and CXCR4 mediates homing oftransplanted bone marrow–derived cells to sites of brain injury[30, 31] and migration of transplanted NSCs to the infarctedarea after stroke [32]. Upregulation of SDF-1 immunoreactivitywas detected at all time points in reactive astrocytes in thepenumbra area [30, 32]. Neurons formed after the 30-minuteinsult, which induced lower SDF-1� expression, exhibited lessmigration than those formed after 2-hour MCAO (Fig. 2). Thespecific CXCR4 blocker AMD3100 [23] inhibited the migrationof the new neurons. The exact mechanism of how SDF-1�/CXCR4 signaling regulates migration in damaged striatum isnot known. We found that rat SVZ progenitors express the
CXCR4 receptor in vitro [33]. A direct action on these cells issupported by our finding that SDF-1� activated mitogen-acti-vated protein kinases and induced CXCR4-mediated chemotaxisin cells derived from adult rat SVZ. However, AMD3100 onlypartially suppressed migration in vivo, which suggests that othermechanisms also direct the new neurons to the damaged area.
Similar to observations after extensive ischemic lesions[34], we found that spontaneous motor recovery after strokeaffecting only the striatum occurred over several months con-comitantly with striatal neurogenesis (Fig. 6). Also in line witha possible functional role of the new neurons, administration ofmolecules that promote neural proliferation in the SVZ andstriatal neurogenesis after stroke, for example, vascular endo-thelial growth factor [35] and erythropoietin [36], is associatedwith superior recovery. However, with available methods, it isnot possible to prove a causal relationship between neurogenesisand behavioral improvement after stroke. Suppression or en-hancement of stroke-induced neurogenesis through delivery ofmitosis inhibitors or trophic factors, respectively, will also affectother processes. Generation of conditional transgenic mice, inwhich newly formed neuroblasts carry a gene for selectiveablation, could solve this problem.
Our finding that stroke-induced neurogenesis is persistenthas several implications. The number of new striatal neuronsthat potentially could replace the dead neurons is much largerthan suggested by previous calculations [4]. Furthermore, ex-perimental studies to explore mechanisms of neuronal replace-ment from endogenous NSCs in the damaged brain, as well asvarious interventions to promote neurogenesis, are not restrictedto the acute postischemic phase but can be applied over anextended time period. Most importantly, we now know thatadult NSCs continuously produce a cellular raw material thatmay be used for self-repair in the brain during the recoveryphase after stroke.
ACKNOWLEDGMENTSThis work was supported by the Swedish Research Council,EuroStemCell (European Union Framework 6 project LSHB-CT-2003-503005), and the King Gustav V and Queen Victoria,Soderberg, Crafoord, and Kock Foundations. The Lund StemCell Center is supported by a Center of Excellence grant in LifeSciences from the Swedish Foundation for Strategic Research.Z.K. and O.L. contributed equally to this study.
DISCLOSURESThe authors indicate no potential conflicts of interest.
REFERENCES
1 Jin K, Minami M, Lan JQ et al. Neurogenesis in dentate subgranular zoneand rostral subventricular zone after focal cerebral ischemia in the rat.Proc Natl Acad Sci U S A 2001;98:4710–4715.
2 Zhang RL, Zhang ZG, Zhang L et al. Proliferation and differentiation ofprogenitor cells in the cortex and the subventricular zone in the adult ratafter focal cerebral ischemia. Neuroscience 2001;105:33–41.
3 Zhang R, Zhang Z, Zhang C et al. Stroke transiently increasessubventricular zone cell division from asymmetric to symmetric andincreases neuronal differentiation in the adult rat. J Neurosci 2004;24:5810 –5815.
4 Arvidsson A, Collin T, Kirik D et al. Neuronal replacement from en-dogenous precursors in the adult brain after stroke. Nat Med 2002;8:963–970.
5 Parent JM, Vexler ZS, Gong C et al. Rat forebrain neurogenesis andstriatal neuron replacement after focal stroke. Ann Neurol 2002;52:802– 813.
6 Jin K, Sun Y, Xie L et al. Directed migration of neuronal precursors intothe ischemic cerebral cortex and striatum. Mol Cell Neurosci 2003;24:171–189.
7 Zhang R, Zhang Z, Wang L et al. Activated neural stem cellscontribute to stroke-induced neurogenesis and neuroblast migrationtoward the infarct boundary in adult rats. J Cereb Blood Flow Metab2004;24:441– 448.
Figure 6. Spontaneous functional recovery after stroke-induced striatalinjury occurs gradually over several months concomitantly with striatalneurogenesis. The graph shows the degree of behavioral deficits overtime in the horizontal ladder-walking test. Note the significant improve-ment between 1 and 4 months after stroke. In contrast, the improvementin two other tests, the stepping and cylinder tests, had already plateauedwithin the first month (data not shown). Photograph shows rat walkingon the horizontal ladder. n � 6. Data are presented as the mean �standard error of the mean. �, p � .05, one-way analysis of variancewith Fischer’s post hoc test. Abbreviation: MCAO, middle cerebralartery occlusion.
746 Neurogenesis after Stroke

8 Bernal GM, Peterson DA. Neural stem cells as therapeutic agents forage-related brain repair. Aging Cell 2004;3:345–351.
9 Koizumi J, Yoshida Y, Nakazawa T et al. Experimental studies ofischemic brain edema. 1. A new experimental model of cerebral embo-lism in rats in which recirculation can be introduced in the ischemic area.Jpn J Stroke 1986;8:1–8.
10 Zhao Q, Memezawa H, Smith ML et al. Hyperthermia complicatesmiddle cerebral artery occlusion induced by an intraluminal filament.Brain Res 1994;649:253–259.
11 Ekdahl CT, Mohapel P, Elmer E et al. Caspase inhibitors increaseshort-term survival of progenitor-cell progeny in the adult rat dentategyrus following status epilepticus. Eur J Neurosci 2001;14:937–945.
12 Parmar M, Sjoberg A, Bjorklund A et al. Phenotypic and molecularidentity of cells in the adult subventricular zone. in vivo and afterexpansion in vitro. Mol Cell Neurosci 2003;24:741–752.
13 Metz GA, Schwab ME, Welzl H. The effects of acute and chronic stresson motor and sensory performance in male Lewis rats. Physiol Behav2001;72:29–35.
14 Schallert T, Norton D, Jones T. A clinically relevant unilateral rat modelof Parkinsonian akinesia. J Neural Transplant Plast 1992;3:332–333.
15 Schallert T, Lindner MD. Rescuing neurons from trans-synaptic degen-eration after brain damage: Helpful, harmful, or neutral in recovery offunction? Can J Psychol 1990;44:276–292.
16 Andsberg G, Kokaia Z, Klein RL et al. Neuropathological and behavioralconsequences of adeno-associated viral vector-mediated continuous in-trastriatal neurotrophin delivery in a focal ischemia model in rats. Neu-robiol Dis 2002;9:187–204.
17 Jorgensen HS, Nakayama H, Raaschou HO et al. Outcome and timecourse of recovery in stroke. Part II: Time course of recovery. TheCopenhagen Stroke Study. Arch Phys Med Rehabil 1995;76:406–412.
18 Doetsch F, Petreanu L, Caille I et al. EGF converts transit-amplifyingneurogenic precursors in the adult brain into multipotent stem cells.Neuron 2002;36:1021–1034.
19 Morshead CM, van der Kooy D. Disguising adult neural stem cells. CurrOpin Neurobiol 2004;14:125–131.
20 Couillard-Despres S, Winner B, Schaubeck S et al. Doublecortinexpression levels in adult brain reflect neurogenesis. Eur J Neurosci2005;21:1–14.
21 Herceg Z, Wang ZQ. Functions of poly(ADP-ribose) polymerase(PARP) in DNA repair, genomic integrity and cell death. Mutat Res2001;477:97–110.
22 Kokaia Z, Andsberg G, Martinez-Serrano A et al. Focal cerebral isch-emia in rats induces expression of P75 neurotrophin receptor in resistantstriatal cholinergic neurons. Neuroscience 1998;84:1113–1125.
23 Hatse S, Princen K, Bridger G et al. Chemokine receptor inhibition byAMD3100 is strictly confined to CXCR4. FEBS Lett 2002;527:255–262.
24 Brown JP, Couillard-Despres S, Cooper-Kuhn CM et al. Transient ex-pression of doublecortin during adult neurogenesis. J Comp Neurol2003;467:1–10.
25 Ekdahl CT, Zhu C, Bonde S et al. Death mechanisms in status epilepti-cus-generated neurons and effects of additional seizures on their survival.Neurobiol Dis 2003;14:513–523.
26 Ceccatelli S, Tamm C, Sleeper E et al. Neural stem cells and cell death.Toxicol Lett 2004;149:59–66.
27 Stoll G, Jander S, Schroeter M. Inflammation and glial responses inischemic brain lesions. Prog Neurobiol 1998;56:149–171.
28 Ekdahl CT, Claasen JH, Bonde S et al. Inflammation is detrimental forneurogenesis in adult brain. Proc Natl Acad Sci U S A 2003;100:13632–13637.
29 Monje ML, Toda H, Palmer TD. Inflammatory blockade restores adulthippocampal neurogenesis. Science 2003;302:1760–1765.
30 Hill WD, Hess DC, Martin-Studdard A et al. SDF-1 (CXCL12) isupregulated in the ischemic penumbra following stroke: Association withbone marrow cell homing to injury. J Neuropathol Exp Neurol 2004;63:84–96.
31 Ji JF, He BP, Dheen ST et al. Interactions of chemokines and chemokinereceptors mediate the migration of mesenchymal stem cells to the im-paired site in the brain after hypoglossal nerve injury. STEM CELLS2004;22:415–427.
32 Imitola J, Raddassi K, Park KI et al. Directed migration of neural stemcells to sites of CNS injury by the stromal cell-derived factor 1alpha/CXC chemokine receptor 4 pathway. Proc Natl Acad Sci U S A 2004;101:18117–18122.
33 Tran PB, Ren D, Veldhouse TJ et al. Chemokine receptors are expressedwidely by embryonic and adult neural progenitor cells. J Neurosci Res2004;76:20–34.
34 Modo M, Stroemer RP, Tang E et al. Neurological sequelae and long-term behavioural assessment of rats with transient middle cerebral arteryocclusion. J Neurosci Methods 2000;104:99–109.
35 Sun Y, Jin K, Xie L et al. VEGF-induced neuroprotection, neurogenesis,and angiogenesis after focal cerebral ischemia. J Clin Invest Jun 2003;111:1843–1851.
36 Wang L, Zhang Z, Wang Y et al. Treatment of stroke with erythropoietinenhances neurogenesis and angiogenesis and improves neurologicalfunction in rats. Stroke 35:1732–1737.
747Thored, Arvidsson, Cacci et al.
www.StemCells.com
Related Documents