of June 16, 2015. This information is current as Peroxynitrite-Driven Apoptotic Death Impairment of Tyrosine Phosphorylation and Activation and Proliferation by Promoting Peroxynitrite Inhibits T Lymphocyte Rafael Radi and Alfonso M. Cayota Francoise Vuillier, Gabriela Gualco, Guillaume Dighiero, Cecilia Brito, Mercedes Naviliat, Adriana C. Tiscornia, http://www.jimmunol.org/content/162/6/3356 1999; 162:3356-3366; ; J Immunol References http://www.jimmunol.org/content/162/6/3356.full#ref-list-1 , 26 of which you can access for free at: cites 75 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 1999 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on June 16, 2015 http://www.jimmunol.org/ Downloaded from by guest on June 16, 2015 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of June 16, 2015.This information is current as
Peroxynitrite-Driven Apoptotic DeathImpairment of Tyrosine Phosphorylation andActivation and Proliferation by Promoting Peroxynitrite Inhibits T Lymphocyte
Rafael Radi and Alfonso M. CayotaFrancoise Vuillier, Gabriela Gualco, Guillaume Dighiero, Cecilia Brito, Mercedes Naviliat, Adriana C. Tiscornia,
http://www.jimmunol.org/content/162/6/33561999; 162:3356-3366; ;J Immunol
Referenceshttp://www.jimmunol.org/content/162/6/3356.full#ref-list-1
, 26 of which you can access for free at: cites 75 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 16, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Peroxynitrite Inhibits T Lymphocyte Activation andProliferation by Promoting Impairment of TyrosinePhosphorylation and Peroxynitrite-Driven Apoptotic Death1
Cecilia Brito,* Mercedes Naviliat,*¶ Adriana C. Tiscornia,* Francoise Vuillier, §
Gabriela Gualco,† Guillaume Dighiero,§ Rafael Radi,* and Alfonso M. Cayota2‡
Peroxynitrite (ONOO2) is a potent oxidizing and nitrating agent produced by the reaction of nitric oxide with superoxide. Itreadily nitrates phenolic compounds such as tyrosine residues in proteins, and it has been demonstrated that nitration of tyrosineresidues in proteins inhibits their phosphorylation. During immune responses, tyrosine phosphorylation of key substrates byprotein tyrosine kinases is the earliest of the intracellular signaling pathways following activation through the TCR complex. Thiswork was aimed to evaluate the effects of ONOO2 on lymphocyte tyrosine phosphorylation, proliferation, and survival. Addi-tionally, we studied the generation of nitrating species in vivo and in vitro during immune activation. Our results demonstrate thatONOO2, through nitration of tyrosine residues, is able to inhibit activation-induced protein tyrosine phosphorylation in purifiedlymphocytes and prime them to undergo apoptotic cell death after PHA- or CD3-mediated activation but not upon phorbolester-mediated stimulation. We also provide evidence indicating that peroxynitrite is produced during in vitro immune activation,mainly by cells of the monocyte/macrophage lineage. Furthermore, immunohistochemical studies demonstrate the in vivo gener-ation of nitrating species in human lymph nodes undergoing mild to strong immune activation. Our results point to a physiologicalrole for ONOO2 as a down-modulator of immune responses and also as key mediator in cellular and tissue injury associated withchronic activation of the immune system. The Journal of Immunology,1999, 162: 3356–3366.
N itric oxide (zNO)3 is a diffusible messenger known todisplay a variety of physiological functions, includingvasorelaxation, bronchodilation, inhibition of platelet
aggregation, and neurotransmission (1–5). Additionally it appearsto be involved in the macrophage-dependent killing of intracellularparasites and possibly cancer cells, indicating the potential of thisfree radical to mediate cytotoxic and pathological effects (6, 7).When produced in excess,zNO can have a multitude of potentiallytoxic effects, which are highly dependent on its concentration andthe particular microenvironment in which it is produced (7). Thus,zNO has been reported to inhibit mitochondrial respiration (8, 9)and ribonucleotide reductase (10) and to damage DNA (11). Inaddition, it has been implicated in the pathogenesis of glutamate-mediated neuronal toxicity, inflammation, graft-vs-host disease,and reoxygenation injury following ischemia (12–16). A signifi-cant portion of these toxic effects ofzNO are due to the generationof peroxynitrite (ONOO2), the reaction product ofzNO with
superoxide (O2.) (17–19). The second order rate constant for the
reaction betweenzNO and O2. anion to yield ONOO2 is 6.73 109
M/s (20), which is 3 times faster than the reaction of O2. with
superoxide dismutase (SOD). Therefore,zNO is one of the fewbiological molecules capable of outcompeting SOD for O2
., mak-ing ONOO2 formation a favored reaction under in vivo conditionswhere cellular production ofzNO and O2
. is increased. Peroxyni-trite is a powerful oxidant (6, 17–19, 21, 22) that nitrates free andprotein-associated tyrosines and other phenolic moieties (7, 23,24). Stimulated macrophages, neutrophils, motoneurons, andendothelial cells have been demonstrated to generate peroxyni-trite (23, 25–27), and as assessed by nitrotyrosine detection,recent data have provided evidence for the in vivo formation ofperoxynitrite in human atherosclerosis, sepsis, human acutelung injury, and chronic inflammation (28 –31). It has beendemonstrated that peroxynitrite-mediated nitration of theorthoposition of tyrosine residues in proteins inhibits tyrosine phos-phorylation and targets proteins for degradation (32–34). Inlymphocytes, the earliest of the intracellular signaling eventsinitiated by Ag binding to the TCR/CD3 complex involves theactivation of several tyrosine kinases and phosphatases that, inturn, results in the phosphorylation of several key cellular sub-strates (35, 36). Thus, it can be speculated that nitration oftyrosine residues on proteins could prevent T lymphocyte-de-pendent immune responses. It has been demonstrated that thesetransduction signals leading to the activation of several tran-scriptional factors are also strongly influenced by cellular redoxstatus (reviewed in Refs. 37 and 38). This work was aimed toexamine the effects of peroxynitrite in normal lymphocyte ac-tivation, proliferation, and survival. We have also analyzed thegeneration of endogenous nitrating species during normal im-mune responses and their consequences on activation-inducedtyrosine phosphorylation and apoptotic cell death.
Departments of *Biochemistry,¶Rheumatology,†Pathology, and‡Medicine, Facultadde Medicina, Universidad de la Republica, Montevideo, Uruguay; and‡Unit of Im-munohematology and Immunopathology, Pasteur Institute, Paris, France
Received for publication July 29, 1998. Accepted for publication December 2, 1998.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by research grants from Consejo Nacional de Investiga-ciones Cientıficas y Tecnicas (Ministerio de Educacion y Cultura, Uruguay) and inpart by grants from Comite Evaluation-Orientation Cooperation Scientifique, Univer-site Paris VII (France), and Universidad de la Republica, Montevideo Uruguay.2 Address correspondence and reprint requests to Dr. A. Cayota, Departmento deBioquımica-Facultad de Medicina, Av. Gral Flores 2125 CP 11800, Montevideo,Uruguay.3 Abbreviations used in this paper:zNO, nitric oxide; ONOO2, peroxynitrite; O2
.,superoxide; SOD, superoxide dismutase; Mn-TBAP, manganese [III] tetrakis [4-ben-zoic acid] porphyrin; MC-540, merocyanin 540; KLH, keyhole limpet hemocyanin;sCD3, soluble CD3; iCD3, immobilized CD3; PARS, poly(A)DP ribosyl synthetase.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsMaterials
A stock solution of aminoguanidine was prepared from a bicarbonate salt(Sigma, St. Louis, MO) by suspending it in a minimum quantity of distilledwater. Concentrated phosphoric acid was then added until no more CO2 evo-lution was visualized. The pH was adjusted to 7.4 with saturated NaOH, andthe final volume was adjusted with PBS to the desired stock concentration.Mn-TBAP (manganese [III] tetrakis [4-benzoic acid] porphyrin) was pur-chased from Alexis (San Diego, CA). PHA, PMA,L-tyrosine, 3-amino-L-tyrosine, o-phospho-L-tyrosine, 3-nitro-L-tyrosine, and monoclonal anti-phosphotyrosine (mouse IgG1, clone PT-66) were purchased from Sigma.
Cell preparation and culture conditions
PBMC were isolated from peripheral venous blood obtained from healthyvolunteers by centrifugation over a Ficoll gradient. T lymphocytes werepurified from nonadherent cells obtained after adherence of PBMC to plas-tic for 2 h at37°C. Nonadherent cells were then incubated with saturatingconcentrations of anti-CD14, anti-CD19, anti-CD16, and anti-CD56 mu-rine mAbs (Dakopatts, Glostrup, Denmark) for 1 h at 4°C tocompletelyeliminate contaminating monocytes, B lymphocytes, and NK cells. Afterwashing, cells were incubated in the presence of magnetic beads precoatedwith sheep anti-mouse Abs (Dynabeads, Dynal, Oslo, Norway) to achievea 5:1 bead to cell ratio. After incubation (1 h at 4°C) cell fractions wereseparated using a magnet applied to the outside of the tube. Unbound Tlymphocytes were carefully aspirated, and a second round of immunomag-netic separation was performed with a bead to cell ratio of 10:1; this frac-tion was referred to as purified lymphocytes. Purified monocytes were iso-lated by adherence to plastic for 2 h at37°C from a fraction of untreatedwhole PBMC that were not exposed to the Abs used in lymphocyte prep-aration. The purity of the subsets was evaluated by either flow cytometryor immunofluorescence, and the purity of the cell preparations used was.95%. The possibility of minor contaminant cells includes plasma cells,which, in contrast to B cells, do not react with CD19. Cells were culturedin RPMI 1640 supplemented with 10% heat-inactivated FCS (LPS free;Life Technologies, Grand Island, NY), 4 mM glutamine, 1 mM HEPES, 1mM sodium pyruvate, and penicillin (200 IU/ml)-streptomycin (50mg/ml;Life Technologies).
Synthesis of peroxynitrite
Peroxynitrite was synthesized in a quenched flow reactor as previouslyreported (17, 18). Briefly, solutions of 1) 0.6 M NaNO2 and 2) 0.6 M H2O2
were pumped into a T junction and mixed in a glass tube. The reaction wasquenched by pumping 1.5 M NaOH at the same rate into a second Tjunction at the end of glass tube. Excess H2O2 was removed by addition ofMnO2 powder. The solution was then frozen at280°C, at which temper-ature peroxynitrite forms a yellow top layer due to freeze fractionation. Theconcentration of this layer was usually in the range of 250–400 mM asdetermined spectrophotometrically at 302 nm (e302 5 1670 M21zcm21).Working dilutions were made in 1 N NaOH.
Agarose gel analysis of DNA fragmentation
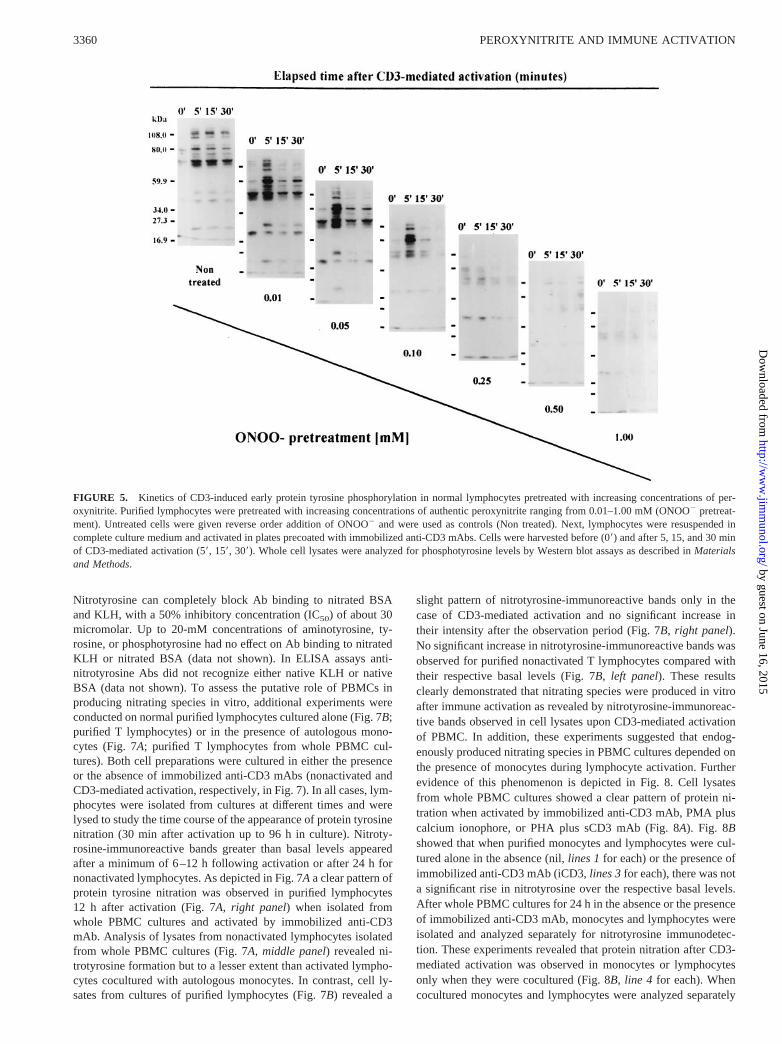
The presence of apoptotic cells was assessed by visualization of a charac-teristic banding pattern of internucleosomal DNA fragmentation. For this,8 3 106 cells were pelleted for 10 min at 2003 g and resuspended in 0.5ml of TTE buffer (10 mM Tris-HCl (pH 7.6) and 1 mM EDTA containing0.2% Triton X-100). Fragmented DNA was separated from intact chroma-tin by microcentrifugation for 10 min at 13,0003 g at 4°C. Supernatants(0.5 ml) were mixed with 0.1 ml of ice-cold 5 M NaCl and vortexedvigorously. After addition of 0.7 ml of ice-cold isopropanol, DNA wasprecipitated overnight at220°C. Samples were centrifuged for 10 min at13,0003 g at 4°C and washed in ice-cold 70% ethanol. Dried pellets weresolubilized in 20ml of TE buffer (10 mM Tris-HCl (pH 7.6) and 1 mMEDTA). Samples were mixed with loading buffer, heated for 10 min at65°C, and electrophoresed on 1% agarose gels with TBE buffer until bro-mophenol blue dye migrated 3–4 cm. Fragmented DNA was visualized bystaining with ethidium bromide.
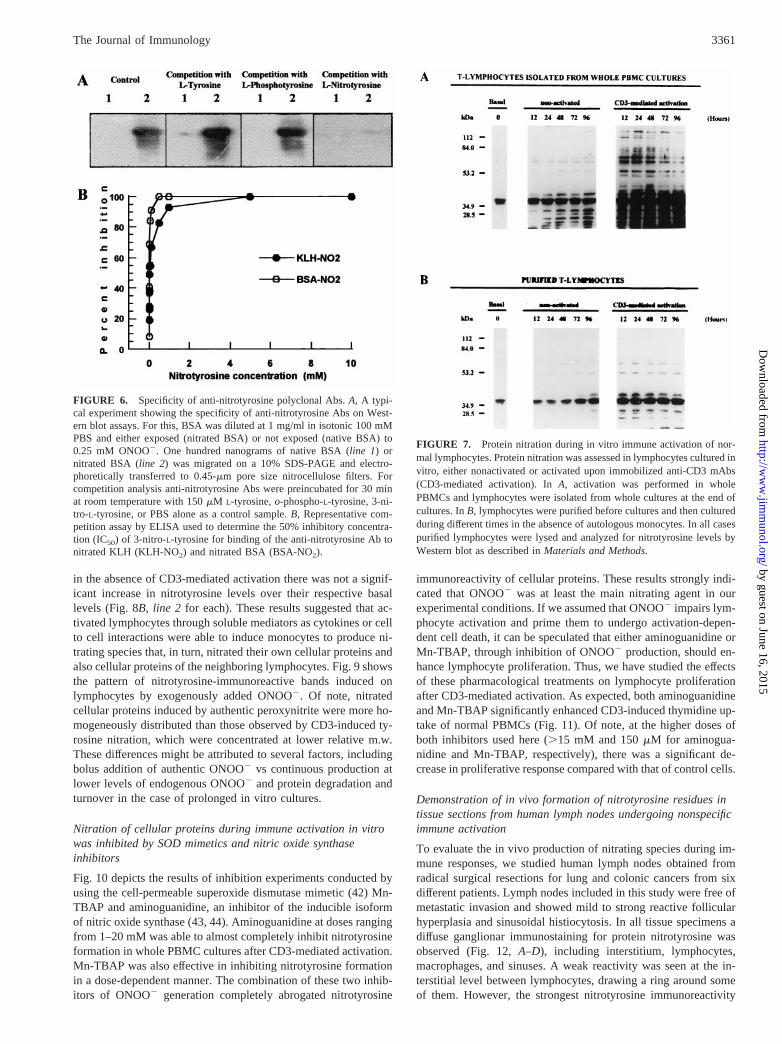
Flow cytometric analysis of apoptotic cells
Flow cytometric analysis was performed on an EPICS 752 FACS (Coulter,Hialeah, FL). The forward and right angle scatter signals were used to gateout cellular debris, damaged cells, and aggregates. The fluorescence emis-sion was displayed on a log scale to yield histograms of log fluorescenceintensity (x-axis) vs cell number (y-axis). The fluorescent dye merocyanin540 (MC-540, Fluka, Ronkonkona, NY) has been successfully used as afluorescent probe in cytometric analysis by its ability to bind symmetric
membranes with exposed phosphatidylserine on apoptotic cells (39). Stain-ing of cells with MC-540 was performed according to the method de-scribed by McEvoy et al. (40) with minor modifications. In brief, a stocksolution of MC-540 was prepared in 50% ethanol at 1 mg/ml and stored inthe dark at220°C. A working solution was freshly prepared by dilutingmerocyanine in HEPES-buffered salt solution with 0.1% BSA to achieve afinal concentration of 2mg/ml. Cells (106) were resuspended in 100ml ofworking solution and incubated for 5 min in the dark at room temperature,then washed once with HEPES-buffered salt solution without BSA andimmediately analyzed by flow cytometry.
Production of rabbit anti-nitrotyrosine polyclonal Abs
New Zealand rabbits were immunized with 500mg of peroxynitrite-modified keyhole limpet hemocyanin (NO2-KLH) emulsioned with anequal volume of CFA according to the procedure of Ye et al. (41). KLH(Sigma) diluted in 100 mM PBS was nitrated in the presence of 0.5 mMperoxynitrite. Boosters were performed every 3 mo after the first injectionwith NO2-KLH emulsioned in IFA. Rabbits were bled 15 days after the lastbooster according to serum anti-nitrotyrosine titration as assessed byELISAs against native KLH and NO2-KLH. Pooled sera were fractionatedin ammonium sulfate, and after dialysis overnight unwanted Abs againstnative KLH were eliminated using a Sepharose-4B column coupled tonative KLH. Specific anti-nitrotyrosine Abs were affinity purified using aSepharose-4B column coupled to 3-nitro-L-tyrosine.
ELISAs for specificity and competition analysis of anti-nitrotyrosine Abs
Polyclonal anti-nitrotyrosine Abs were screened for specificity and un-wanted cross-reactions by Western blot and ELISA. Briefly, 96-well poly-styrene ELISA plates were coated with either native or peroxynitrite-nitrated proteins diluted at 30mg/ml in carbonate buffer (50 mM; pH 9.5)and incubated overnight at 4°C. After washing, plates were blocked with5% gelatin in PBS plus 0.05% Tween-20 (v/v) for 1 h at 37°C. Afterwashing, the anti-nitrotyrosine polyclonal Ab was added at a 1/2000 di-lution (0.12mg/ml) in PBS-0.05% Tween for 2 h at 37°C. The plates werewashed and then incubated with goat anti-rabbit IgG conjugated withhorseradish peroxidase (Dakopatts) for 2 h at 37°C. The plates weredeveloped with o-phenylenediamine dihydrochloride, and absorbancewas read at 492 using a microplate reader. SDS-PAGE and Western blot-ting were performed as described below. When competition analysis wereperformed, the anti-nitrotyrosine Ab was preincubated for 30 min atroom temperature with several concentrations ofL-tyrosine, 3-amino-L-tyrosine,o-phospho-L-tyrosine, and 3-nitro-L-tyrosine as competitors forAb binding.
Nitrotyrosine and phosphotyrosine immunodetection by Westernblot assays
For Western blot analysis, cell suspensions were washed twice in PBS andresuspended in hot lysis buffer (10 mM Tris-HCl (pH 6.8), 1% SDS, and5% glycerol) followed by immediate boiling for 5 min, sonication, anddetermination of protein concentration by the bicinchoninic acid method(Pierce, Rockford, IL). Samples were stored at270°C until analysis on10% SDS-PAGE (30mg of protein/lane). Next, proteins were electro-phoretically transferred to 0.45-mm pore size nitrocellulose membranes,and nonspecific binding sites were blocked overnight in blocking in TBS-Tblocking buffer (5% BSA and 0.3% Tween in Tris-buffered saline, pH 7.5).For nitrotyrosine immunodetection, nitrocellulose filters were probed withanti-nitrotyrosine polyclonal Abs diluted 1/1000 in blocking buffer for 1 hat room temperature. After extensive washings in TBS-Tween 0.3%, blotswere further incubated for 45 min with a horseradish peroxidase-linkedanti-rabbit IgG polyclonal Ab raised in donkeys and developed using theenhanced chemiluminescence detection system (Amersham, ArlingtonHeights, IL). For phosphotyrosine immunodetection, nitrocellulose filterswere blocked as described above and then probed with an anti-phosphotyrosine mAb diluted in TBS-T with 0.1% BSA. After washing inTBS-T, blots were incubated for 45 min with a horseradish peroxidase-linked anti-mouse IgG polyclonal Ab and developed as described above.
Proliferation assays
For [3H]thymidine uptake, cells were cultured in 96-well culture plates at5 3 104 cells/well in a total volume of 0.2 ml. Stimulation was conductedin the presence of immobilized anti-CD3 mAbs, PHA, and soluble (sCD3)anti-CD3 mAbs (at 5 and 10mg/ml, respectively), or PMA (100 ng/ml)plus calcium ionophore A-23187 (0.2mg/ml). After 3 days, cultures werepulsed with 1mCi/well of [3H]thymidine and further cultured for 18 h.
3357The Journal of Immunology
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Cells were then harvested, and radioactivity was measured in a beta scin-tillation counter. All samples were incubated in quadruplicate, and for eachexperimental condition a control with culture medium alone was included.
Immunohistochemical studies for nitrotyrosine detection
Immunohistochemistry to detect anti-nitrotyrosine was performed on hu-man lymph nodes obtained from routine surgical resections for lung andcolonic cancers obtained at the Hospital de Clınicas (Facultad de Medicina,Universidad de la Republica, Montevideo, Uruguay). Lymph nodes werefixed in 10% formalin and paraffin embedded, and 5-mm sections weremounted in silanized microscope slides. Tissue sections were probed withanti-nitrotyrosine polyclonal Abs (working dilution, 1/50) and developedwith a secondary Ab coupled to biotin using a streptavidin-peroxidase kit(Sigma) and either aminoethylcarbazol or diaminobenzidine as chromo-gens. Histological sections were counterstained with hemalum. Controlswere performed with an irrelevant polyclonal rabbit serum as the first Ab.The technical control was omission of the primary Ab. The specificity ofimmunostaining was always assessed by competition with soluble3-nitrotyrosine at 5 mM.
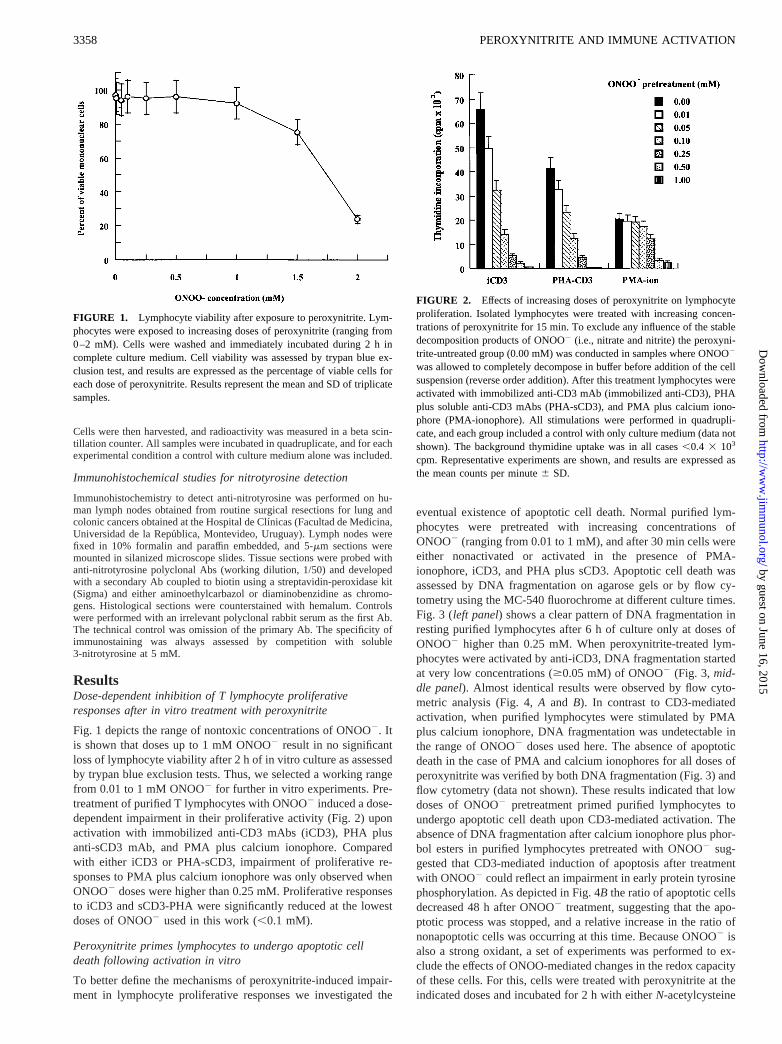
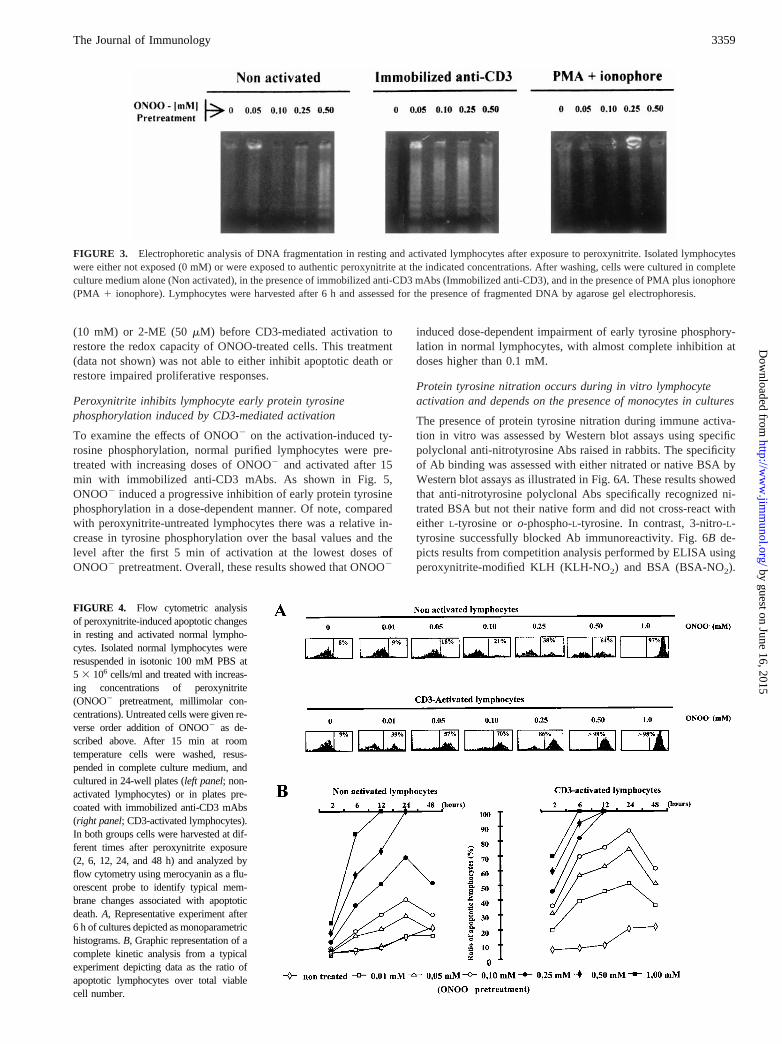
ResultsDose-dependent inhibition of T lymphocyte proliferativeresponses after in vitro treatment with peroxynitrite
Fig. 1 depicts the range of nontoxic concentrations of ONOO2. Itis shown that doses up to 1 mM ONOO2 result in no significantloss of lymphocyte viability after 2 h of invitro culture as assessedby trypan blue exclusion tests. Thus, we selected a working rangefrom 0.01 to 1 mM ONOO2 for further in vitro experiments. Pre-treatment of purified T lymphocytes with ONOO2 induced a dose-dependent impairment in their proliferative activity (Fig. 2) uponactivation with immobilized anti-CD3 mAbs (iCD3), PHA plusanti-sCD3 mAb, and PMA plus calcium ionophore. Comparedwith either iCD3 or PHA-sCD3, impairment of proliferative re-sponses to PMA plus calcium ionophore was only observed whenONOO2 doses were higher than 0.25 mM. Proliferative responsesto iCD3 and sCD3-PHA were significantly reduced at the lowestdoses of ONOO2 used in this work (,0.1 mM).
Peroxynitrite primes lymphocytes to undergo apoptotic celldeath following activation in vitro
To better define the mechanisms of peroxynitrite-induced impair-ment in lymphocyte proliferative responses we investigated the
eventual existence of apoptotic cell death. Normal purified lym-phocytes were pretreated with increasing concentrations ofONOO2 (ranging from 0.01 to 1 mM), and after 30 min cells wereeither nonactivated or activated in the presence of PMA-ionophore, iCD3, and PHA plus sCD3. Apoptotic cell death wasassessed by DNA fragmentation on agarose gels or by flow cy-tometry using the MC-540 fluorochrome at different culture times.Fig. 3 (left panel) shows a clear pattern of DNA fragmentation inresting purified lymphocytes after 6 h ofculture only at doses ofONOO2 higher than 0.25 mM. When peroxynitrite-treated lym-phocytes were activated by anti-iCD3, DNA fragmentation startedat very low concentrations ($0.05 mM) of ONOO2 (Fig. 3,mid-dle panel). Almost identical results were observed by flow cyto-metric analysis (Fig. 4,A and B). In contrast to CD3-mediatedactivation, when purified lymphocytes were stimulated by PMAplus calcium ionophore, DNA fragmentation was undetectable inthe range of ONOO2 doses used here. The absence of apoptoticdeath in the case of PMA and calcium ionophores for all doses ofperoxynitrite was verified by both DNA fragmentation (Fig. 3) andflow cytometry (data not shown). These results indicated that lowdoses of ONOO2 pretreatment primed purified lymphocytes toundergo apoptotic cell death upon CD3-mediated activation. Theabsence of DNA fragmentation after calcium ionophore plus phor-bol esters in purified lymphocytes pretreated with ONOO2 sug-gested that CD3-mediated induction of apoptosis after treatmentwith ONOO2 could reflect an impairment in early protein tyrosinephosphorylation. As depicted in Fig. 4B the ratio of apoptotic cellsdecreased 48 h after ONOO2 treatment, suggesting that the apo-ptotic process was stopped, and a relative increase in the ratio ofnonapoptotic cells was occurring at this time. Because ONOO2 isalso a strong oxidant, a set of experiments was performed to ex-clude the effects of ONOO-mediated changes in the redox capacityof these cells. For this, cells were treated with peroxynitrite at theindicated doses and incubated for 2 h with eitherN-acetylcysteine
FIGURE 1. Lymphocyte viability after exposure to peroxynitrite. Lym-phocytes were exposed to increasing doses of peroxynitrite (ranging from0–2 mM). Cells were washed and immediately incubated during 2 h incomplete culture medium. Cell viability was assessed by trypan blue ex-clusion test, and results are expressed as the percentage of viable cells foreach dose of peroxynitrite. Results represent the mean and SD of triplicatesamples.
FIGURE 2. Effects of increasing doses of peroxynitrite on lymphocyteproliferation. Isolated lymphocytes were treated with increasing concen-trations of peroxynitrite for 15 min. To exclude any influence of the stabledecomposition products of ONOO2 (i.e., nitrate and nitrite) the peroxyni-trite-untreated group (0.00 mM) was conducted in samples where ONOO2
was allowed to completely decompose in buffer before addition of the cellsuspension (reverse order addition). After this treatment lymphocytes wereactivated with immobilized anti-CD3 mAb (immobilized anti-CD3), PHAplus soluble anti-CD3 mAbs (PHA-sCD3), and PMA plus calcium iono-phore (PMA-ionophore). All stimulations were performed in quadrupli-cate, and each group included a control with only culture medium (data notshown). The background thymidine uptake was in all cases,0.4 3 103
cpm. Representative experiments are shown, and results are expressed asthe mean counts per minute6 SD.
3358 PEROXYNITRITE AND IMMUNE ACTIVATION
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
(10 mM) or 2-ME (50mM) before CD3-mediated activation torestore the redox capacity of ONOO-treated cells. This treatment(data not shown) was not able to either inhibit apoptotic death orrestore impaired proliferative responses.
Peroxynitrite inhibits lymphocyte early protein tyrosinephosphorylation induced by CD3-mediated activation
To examine the effects of ONOO2 on the activation-induced ty-rosine phosphorylation, normal purified lymphocytes were pre-treated with increasing doses of ONOO2 and activated after 15min with immobilized anti-CD3 mAbs. As shown in Fig. 5,ONOO2 induced a progressive inhibition of early protein tyrosinephosphorylation in a dose-dependent manner. Of note, comparedwith peroxynitrite-untreated lymphocytes there was a relative in-crease in tyrosine phosphorylation over the basal values and thelevel after the first 5 min of activation at the lowest doses ofONOO2 pretreatment. Overall, these results showed that ONOO2
induced dose-dependent impairment of early tyrosine phosphory-lation in normal lymphocytes, with almost complete inhibition atdoses higher than 0.1 mM.
Protein tyrosine nitration occurs during in vitro lymphocyteactivation and depends on the presence of monocytes in cultures
The presence of protein tyrosine nitration during immune activa-tion in vitro was assessed by Western blot assays using specificpolyclonal anti-nitrotyrosine Abs raised in rabbits. The specificityof Ab binding was assessed with either nitrated or native BSA byWestern blot assays as illustrated in Fig. 6A. These results showedthat anti-nitrotyrosine polyclonal Abs specifically recognized ni-trated BSA but not their native form and did not cross-react witheither L-tyrosine oro-phospho-L-tyrosine. In contrast, 3-nitro-L-tyrosine successfully blocked Ab immunoreactivity. Fig. 6B de-picts results from competition analysis performed by ELISA usingperoxynitrite-modified KLH (KLH-NO2) and BSA (BSA-NO2).
FIGURE 3. Electrophoretic analysis of DNA fragmentation in resting and activated lymphocytes after exposure to peroxynitrite. Isolated lymphocyteswere either not exposed (0 mM) or were exposed to authentic peroxynitrite at the indicated concentrations. After washing, cells were cultured in completeculture medium alone (Non activated), in the presence of immobilized anti-CD3 mAbs (Immobilized anti-CD3), and in the presence of PMA plus ionophore(PMA 1 ionophore). Lymphocytes were harvested after 6 h and assessed for the presence of fragmented DNA by agarose gel electrophoresis.
FIGURE 4. Flow cytometric analysisof peroxynitrite-induced apoptotic changesin resting and activated normal lympho-cytes. Isolated normal lymphocytes wereresuspended in isotonic 100 mM PBS at5 3 106 cells/ml and treated with increas-ing concentrations of peroxynitrite(ONOO2 pretreatment, millimolar con-centrations). Untreated cells were given re-verse order addition of ONOO2 as de-scribed above. After 15 min at roomtemperature cells were washed, resus-pended in complete culture medium, andcultured in 24-well plates (left panel; non-activated lymphocytes) or in plates pre-coated with immobilized anti-CD3 mAbs(right panel; CD3-activated lymphocytes).In both groups cells were harvested at dif-ferent times after peroxynitrite exposure(2, 6, 12, 24, and 48 h) and analyzed byflow cytometry using merocyanin as a flu-orescent probe to identify typical mem-brane changes associated with apoptoticdeath.A, Representative experiment after6 h of cultures depicted as monoparametrichistograms.B, Graphic representation of acomplete kinetic analysis from a typicalexperiment depicting data as the ratio ofapoptotic lymphocytes over total viablecell number.
3359The Journal of Immunology
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Nitrotyrosine can completely block Ab binding to nitrated BSAand KLH, with a 50% inhibitory concentration (IC50) of about 30micromolar. Up to 20-mM concentrations of aminotyrosine, ty-rosine, or phosphotyrosine had no effect on Ab binding to nitratedKLH or nitrated BSA (data not shown). In ELISA assays anti-nitrotyrosine Abs did not recognize either native KLH or nativeBSA (data not shown). To assess the putative role of PBMCs inproducing nitrating species in vitro, additional experiments wereconducted on normal purified lymphocytes cultured alone (Fig. 7B;purified T lymphocytes) or in the presence of autologous mono-cytes (Fig. 7A; purified T lymphocytes from whole PBMC cul-tures). Both cell preparations were cultured in either the presenceor the absence of immobilized anti-CD3 mAbs (nonactivated andCD3-mediated activation, respectively, in Fig. 7). In all cases, lym-phocytes were isolated from cultures at different times and werelysed to study the time course of the appearance of protein tyrosinenitration (30 min after activation up to 96 h in culture). Nitroty-rosine-immunoreactive bands greater than basal levels appearedafter a minimum of 6–12 h following activation or after 24 h fornonactivated lymphocytes. As depicted in Fig. 7Aa clear pattern ofprotein tyrosine nitration was observed in purified lymphocytes12 h after activation (Fig. 7A, right panel) when isolated fromwhole PBMC cultures and activated by immobilized anti-CD3mAb. Analysis of lysates from nonactivated lymphocytes isolatedfrom whole PBMC cultures (Fig. 7A, middle panel) revealed ni-trotyrosine formation but to a lesser extent than activated lympho-cytes cocultured with autologous monocytes. In contrast, cell ly-sates from cultures of purified lymphocytes (Fig. 7B) revealed a
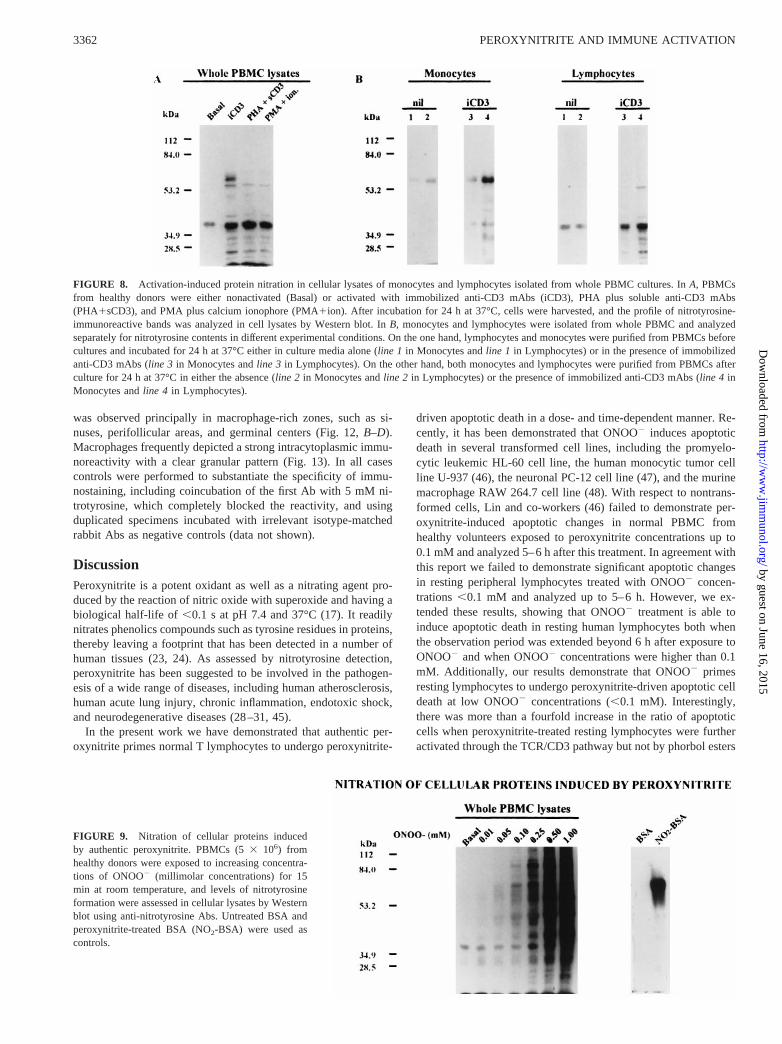
slight pattern of nitrotyrosine-immunoreactive bands only in thecase of CD3-mediated activation and no significant increase intheir intensity after the observation period (Fig. 7B, right panel).No significant increase in nitrotyrosine-immunoreactive bands wasobserved for purified nonactivated T lymphocytes compared withtheir respective basal levels (Fig. 7B, left panel). These resultsclearly demonstrated that nitrating species were produced in vitroafter immune activation as revealed by nitrotyrosine-immunoreac-tive bands observed in cell lysates upon CD3-mediated activationof PBMC. In addition, these experiments suggested that endog-enously produced nitrating species in PBMC cultures depended onthe presence of monocytes during lymphocyte activation. Furtherevidence of this phenomenon is depicted in Fig. 8. Cell lysatesfrom whole PBMC cultures showed a clear pattern of protein ni-tration when activated by immobilized anti-CD3 mAb, PMA pluscalcium ionophore, or PHA plus sCD3 mAb (Fig. 8A). Fig. 8Bshowed that when purified monocytes and lymphocytes were cul-tured alone in the absence (nil,lines 1for each) or the presence ofimmobilized anti-CD3 mAb (iCD3,lines 3for each), there was nota significant rise in nitrotyrosine over the respective basal levels.After whole PBMC cultures for 24 h in the absence or the presenceof immobilized anti-CD3 mAb, monocytes and lymphocytes wereisolated and analyzed separately for nitrotyrosine immunodetec-tion. These experiments revealed that protein nitration after CD3-mediated activation was observed in monocytes or lymphocytesonly when they were cocultured (Fig. 8B, line 4 for each). Whencocultured monocytes and lymphocytes were analyzed separately
FIGURE 5. Kinetics of CD3-induced early protein tyrosine phosphorylation in normal lymphocytes pretreated with increasing concentrations of per-oxynitrite. Purified lymphocytes were pretreated with increasing concentrations of authentic peroxynitrite ranging from 0.01–1.00 mM (ONOO2 pretreat-ment). Untreated cells were given reverse order addition of ONOO2 and were used as controls (Non treated). Next, lymphocytes were resuspended incomplete culture medium and activated in plates precoated with immobilized anti-CD3 mAbs. Cells were harvested before (09) and after 5, 15, and 30 minof CD3-mediated activation (59, 159, 309). Whole cell lysates were analyzed for phosphotyrosine levels by Western blot assays as described inMaterialsand Methods.
3360 PEROXYNITRITE AND IMMUNE ACTIVATION
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
in the absence of CD3-mediated activation there was not a signif-icant increase in nitrotyrosine levels over their respective basallevels (Fig. 8B,line 2 for each). These results suggested that ac-tivated lymphocytes through soluble mediators as cytokines or cellto cell interactions were able to induce monocytes to produce ni-trating species that, in turn, nitrated their own cellular proteins andalso cellular proteins of the neighboring lymphocytes. Fig. 9 showsthe pattern of nitrotyrosine-immunoreactive bands induced onlymphocytes by exogenously added ONOO2. Of note, nitratedcellular proteins induced by authentic peroxynitrite were more ho-mogeneously distributed than those observed by CD3-induced ty-rosine nitration, which were concentrated at lower relative m.w.These differences might be attributed to several factors, includingbolus addition of authentic ONOO2 vs continuous production atlower levels of endogenous ONOO2 and protein degradation andturnover in the case of prolonged in vitro cultures.
Nitration of cellular proteins during immune activation in vitrowas inhibited by SOD mimetics and nitric oxide synthaseinhibitors
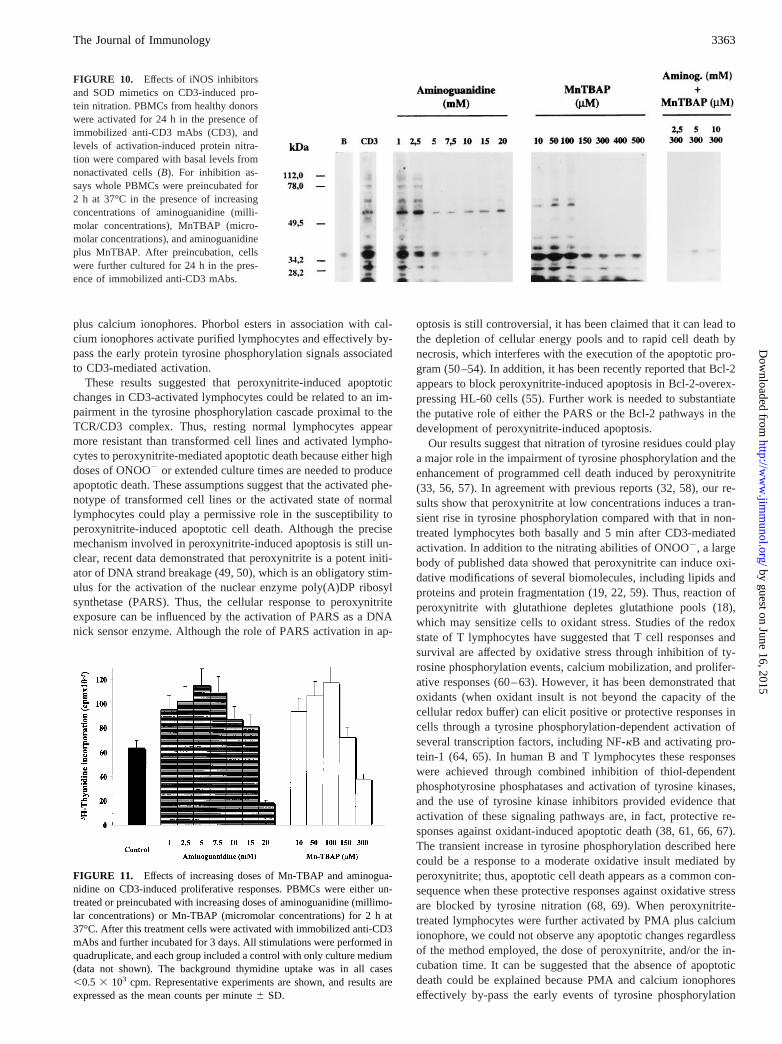
Fig. 10 depicts the results of inhibition experiments conducted byusing the cell-permeable superoxide dismutase mimetic (42) Mn-TBAP and aminoguanidine, an inhibitor of the inducible isoformof nitric oxide synthase (43, 44). Aminoguanidine at doses rangingfrom 1–20 mM was able to almost completely inhibit nitrotyrosineformation in whole PBMC cultures after CD3-mediated activation.Mn-TBAP was also effective in inhibiting nitrotyrosine formationin a dose-dependent manner. The combination of these two inhib-itors of ONOO2 generation completely abrogated nitrotyrosine
immunoreactivity of cellular proteins. These results strongly indi-cated that ONOO2 was at least the main nitrating agent in ourexperimental conditions. If we assumed that ONOO2 impairs lym-phocyte activation and prime them to undergo activation-depen-dent cell death, it can be speculated that either aminoguanidine orMn-TBAP, through inhibition of ONOO2 production, should en-hance lymphocyte proliferation. Thus, we have studied the effectsof these pharmacological treatments on lymphocyte proliferationafter CD3-mediated activation. As expected, both aminoguanidineand Mn-TBAP significantly enhanced CD3-induced thymidine up-take of normal PBMCs (Fig. 11). Of note, at the higher doses ofboth inhibitors used here (.15 mM and 150mM for aminogua-nidine and Mn-TBAP, respectively), there was a significant de-crease in proliferative response compared with that of control cells.
Demonstration of in vivo formation of nitrotyrosine residues intissue sections from human lymph nodes undergoing nonspecificimmune activation
To evaluate the in vivo production of nitrating species during im-mune responses, we studied human lymph nodes obtained fromradical surgical resections for lung and colonic cancers from sixdifferent patients. Lymph nodes included in this study were free ofmetastatic invasion and showed mild to strong reactive follicularhyperplasia and sinusoidal histiocytosis. In all tissue specimens adiffuse ganglionar immunostaining for protein nitrotyrosine wasobserved (Fig. 12,A–D), including interstitium, lymphocytes,macrophages, and sinuses. A weak reactivity was seen at the in-terstitial level between lymphocytes, drawing a ring around someof them. However, the strongest nitrotyrosine immunoreactivity
FIGURE 6. Specificity of anti-nitrotyrosine polyclonal Abs.A, A typi-cal experiment showing the specificity of anti-nitrotyrosine Abs on West-ern blot assays. For this, BSA was diluted at 1 mg/ml in isotonic 100 mMPBS and either exposed (nitrated BSA) or not exposed (native BSA) to0.25 mM ONOO2. One hundred nanograms of native BSA (line 1) ornitrated BSA (line 2) was migrated on a 10% SDS-PAGE and electro-phoretically transferred to 0.45-mm pore size nitrocellulose filters. Forcompetition analysis anti-nitrotyrosine Abs were preincubated for 30 minat room temperature with 150mM L-tyrosine,o-phospho-L-tyrosine, 3-ni-tro-L-tyrosine, or PBS alone as a control sample.B, Representative com-petition assay by ELISA used to determine the 50% inhibitory concentra-tion (IC50) of 3-nitro-L-tyrosine for binding of the anti-nitrotyrosine Ab tonitrated KLH (KLH-NO2) and nitrated BSA (BSA-NO2).
FIGURE 7. Protein nitration during in vitro immune activation of nor-mal lymphocytes. Protein nitration was assessed in lymphocytes cultured invitro, either nonactivated or activated upon immobilized anti-CD3 mAbs(CD3-mediated activation). InA, activation was performed in wholePBMCs and lymphocytes were isolated from whole cultures at the end ofcultures. InB, lymphocytes were purified before cultures and then culturedduring different times in the absence of autologous monocytes. In all casespurified lymphocytes were lysed and analyzed for nitrotyrosine levels byWestern blot as described inMaterials and Methods.
3361The Journal of Immunology
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
was observed principally in macrophage-rich zones, such as si-nuses, perifollicular areas, and germinal centers (Fig. 12,B–D).Macrophages frequently depicted a strong intracytoplasmic immu-noreactivity with a clear granular pattern (Fig. 13). In all casescontrols were performed to substantiate the specificity of immu-nostaining, including coincubation of the first Ab with 5 mM ni-trotyrosine, which completely blocked the reactivity, and usingduplicated specimens incubated with irrelevant isotype-matchedrabbit Abs as negative controls (data not shown).
DiscussionPeroxynitrite is a potent oxidant as well as a nitrating agent pro-duced by the reaction of nitric oxide with superoxide and having abiological half-life of ,0.1 s at pH 7.4 and 37°C (17). It readilynitrates phenolics compounds such as tyrosine residues in proteins,thereby leaving a footprint that has been detected in a number ofhuman tissues (23, 24). As assessed by nitrotyrosine detection,peroxynitrite has been suggested to be involved in the pathogen-esis of a wide range of diseases, including human atherosclerosis,human acute lung injury, chronic inflammation, endotoxic shock,and neurodegenerative diseases (28–31, 45).
In the present work we have demonstrated that authentic per-oxynitrite primes normal T lymphocytes to undergo peroxynitrite-
driven apoptotic death in a dose- and time-dependent manner. Re-cently, it has been demonstrated that ONOO2 induces apoptoticdeath in several transformed cell lines, including the promyelo-cytic leukemic HL-60 cell line, the human monocytic tumor cellline U-937 (46), the neuronal PC-12 cell line (47), and the murinemacrophage RAW 264.7 cell line (48). With respect to nontrans-formed cells, Lin and co-workers (46) failed to demonstrate per-oxynitrite-induced apoptotic changes in normal PBMC fromhealthy volunteers exposed to peroxynitrite concentrations up to0.1 mM and analyzed 5–6 h after this treatment. In agreement withthis report we failed to demonstrate significant apoptotic changesin resting peripheral lymphocytes treated with ONOO2 concen-trations,0.1 mM and analyzed up to 5–6 h. However, we ex-tended these results, showing that ONOO2 treatment is able toinduce apoptotic death in resting human lymphocytes both whenthe observation period was extended beyond 6 h after exposure toONOO2 and when ONOO2 concentrations were higher than 0.1mM. Additionally, our results demonstrate that ONOO2 primesresting lymphocytes to undergo peroxynitrite-driven apoptotic celldeath at low ONOO2 concentrations (,0.1 mM). Interestingly,there was more than a fourfold increase in the ratio of apoptoticcells when peroxynitrite-treated resting lymphocytes were furtheractivated through the TCR/CD3 pathway but not by phorbol esters
FIGURE 8. Activation-induced protein nitration in cellular lysates of monocytes and lymphocytes isolated from whole PBMC cultures. InA, PBMCsfrom healthy donors were either nonactivated (Basal) or activated with immobilized anti-CD3 mAbs (iCD3), PHA plus soluble anti-CD3 mAbs(PHA1sCD3), and PMA plus calcium ionophore (PMA1ion). After incubation for 24 h at 37°C, cells were harvested, and the profile of nitrotyrosine-immunoreactive bands was analyzed in cell lysates by Western blot. InB, monocytes and lymphocytes were isolated from whole PBMC and analyzedseparately for nitrotyrosine contents in different experimental conditions. On the one hand, lymphocytes and monocytes were purified from PBMCs beforecultures and incubated for 24 h at 37°C either in culture media alone (line 1 in Monocytes andline 1 in Lymphocytes) or in the presence of immobilizedanti-CD3 mAbs (line 3 in Monocytes andline 3 in Lymphocytes). On the other hand, both monocytes and lymphocytes were purified from PBMCs afterculture for 24 h at 37°C in either the absence (line 2 in Monocytes andline 2 in Lymphocytes) or the presence of immobilized anti-CD3 mAbs (line 4 inMonocytes andline 4 in Lymphocytes).
FIGURE 9. Nitration of cellular proteins inducedby authentic peroxynitrite. PBMCs (53 106) fromhealthy donors were exposed to increasing concentra-tions of ONOO2 (millimolar concentrations) for 15min at room temperature, and levels of nitrotyrosineformation were assessed in cellular lysates by Westernblot using anti-nitrotyrosine Abs. Untreated BSA andperoxynitrite-treated BSA (NO2-BSA) were used ascontrols.
3362 PEROXYNITRITE AND IMMUNE ACTIVATION
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
plus calcium ionophores. Phorbol esters in association with cal-cium ionophores activate purified lymphocytes and effectively by-pass the early protein tyrosine phosphorylation signals associatedto CD3-mediated activation.
These results suggested that peroxynitrite-induced apoptoticchanges in CD3-activated lymphocytes could be related to an im-pairment in the tyrosine phosphorylation cascade proximal to theTCR/CD3 complex. Thus, resting normal lymphocytes appearmore resistant than transformed cell lines and activated lympho-cytes to peroxynitrite-mediated apoptotic death because either highdoses of ONOO2 or extended culture times are needed to produceapoptotic death. These assumptions suggest that the activated phe-notype of transformed cell lines or the activated state of normallymphocytes could play a permissive role in the susceptibility toperoxynitrite-induced apoptotic cell death. Although the precisemechanism involved in peroxynitrite-induced apoptosis is still un-clear, recent data demonstrated that peroxynitrite is a potent initi-ator of DNA strand breakage (49, 50), which is an obligatory stim-ulus for the activation of the nuclear enzyme poly(A)DP ribosylsynthetase (PARS). Thus, the cellular response to peroxynitriteexposure can be influenced by the activation of PARS as a DNAnick sensor enzyme. Although the role of PARS activation in ap-
optosis is still controversial, it has been claimed that it can lead tothe depletion of cellular energy pools and to rapid cell death bynecrosis, which interferes with the execution of the apoptotic pro-gram (50–54). In addition, it has been recently reported that Bcl-2appears to block peroxynitrite-induced apoptosis in Bcl-2-overex-pressing HL-60 cells (55). Further work is needed to substantiatethe putative role of either the PARS or the Bcl-2 pathways in thedevelopment of peroxynitrite-induced apoptosis.
Our results suggest that nitration of tyrosine residues could playa major role in the impairment of tyrosine phosphorylation and theenhancement of programmed cell death induced by peroxynitrite(33, 56, 57). In agreement with previous reports (32, 58), our re-sults show that peroxynitrite at low concentrations induces a tran-sient rise in tyrosine phosphorylation compared with that in non-treated lymphocytes both basally and 5 min after CD3-mediatedactivation. In addition to the nitrating abilities of ONOO2, a largebody of published data showed that peroxynitrite can induce oxi-dative modifications of several biomolecules, including lipids andproteins and protein fragmentation (19, 22, 59). Thus, reaction ofperoxynitrite with glutathione depletes glutathione pools (18),which may sensitize cells to oxidant stress. Studies of the redoxstate of T lymphocytes have suggested that T cell responses andsurvival are affected by oxidative stress through inhibition of ty-rosine phosphorylation events, calcium mobilization, and prolifer-ative responses (60–63). However, it has been demonstrated thatoxidants (when oxidant insult is not beyond the capacity of thecellular redox buffer) can elicit positive or protective responses incells through a tyrosine phosphorylation-dependent activation ofseveral transcription factors, including NF-kB and activating pro-tein-1 (64, 65). In human B and T lymphocytes these responseswere achieved through combined inhibition of thiol-dependentphosphotyrosine phosphatases and activation of tyrosine kinases,and the use of tyrosine kinase inhibitors provided evidence thatactivation of these signaling pathways are, in fact, protective re-sponses against oxidant-induced apoptotic death (38, 61, 66, 67).The transient increase in tyrosine phosphorylation described herecould be a response to a moderate oxidative insult mediated byperoxynitrite; thus, apoptotic cell death appears as a common con-sequence when these protective responses against oxidative stressare blocked by tyrosine nitration (68, 69). When peroxynitrite-treated lymphocytes were further activated by PMA plus calciumionophore, we could not observe any apoptotic changes regardlessof the method employed, the dose of peroxynitrite, and/or the in-cubation time. It can be suggested that the absence of apoptoticdeath could be explained because PMA and calcium ionophoreseffectively by-pass the early events of tyrosine phosphorylation
FIGURE 10. Effects of iNOS inhibitorsand SOD mimetics on CD3-induced pro-tein nitration. PBMCs from healthy donorswere activated for 24 h in the presence ofimmobilized anti-CD3 mAbs (CD3), andlevels of activation-induced protein nitra-tion were compared with basal levels fromnonactivated cells (B). For inhibition as-says whole PBMCs were preincubated for2 h at 37°C in the presence of increasingconcentrations of aminoguanidine (milli-molar concentrations), MnTBAP (micro-molar concentrations), and aminoguanidineplus MnTBAP. After preincubation, cellswere further cultured for 24 h in the pres-ence of immobilized anti-CD3 mAbs.
FIGURE 11. Effects of increasing doses of Mn-TBAP and aminogua-nidine on CD3-induced proliferative responses. PBMCs were either un-treated or preincubated with increasing doses of aminoguanidine (millimo-lar concentrations) or Mn-TBAP (micromolar concentrations) for 2 h at37°C. After this treatment cells were activated with immobilized anti-CD3mAbs and further incubated for 3 days. All stimulations were performed inquadruplicate, and each group included a control with only culture medium(data not shown). The background thymidine uptake was in all cases,0.5 3 103 cpm. Representative experiments are shown, and results areexpressed as the mean counts per minute6 SD.
3363The Journal of Immunology
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

and, in turn, trigger protective responses against oxidative stress,which could able to inhibit apoptotic death. In contrast, apoptoticchanges observed in resting lymphocytes exposed to high doses ofONOO2 could be explained by the inability to trigger these ty-rosine phosphorylation-dependent protective responses as a con-
sequence of tyrosine nitration. Decreased proliferative responses toPMA and calcium ionophores in lymphocytes pretreated with highdoses of peroxynitrite were not associated with either apoptoticchanges or cellular loss due to a necrotic process, which couldsuggest a peroxynitrite-induced anergic state.
Given the reactive nature of ONOO2 in biological systems (70),one could speculate that peroxynitrite-mediated oxidative modifi-cations could explain the impairment in lymphocyte activation andsurvival described here. Despite the strong oxidizing potential ofONOO2, kinetic factors determine that a limited number of reac-tions are relevant in vivo. In particular, the reactions with thiols,metal centers, and CO2 account for the most of peroxynitrite con-sumption. Since nitration is strongly favored by metals and CO2,nitration constitutes a key cellular event after peroxynitrite forma-tion in vivo. It is unlikely that depletion of endogenous antioxi-dants by ONOO2 could have a relevant place in explaining ourresults because we cannot prevent apoptotic cell death after resto-ration of cellular redox capacity with eitherN-acetylcysteine or2-ME. In addition, the absence of apoptotic cell death after PMA-mediated activation supports the idea that oxidizing ability ofONOO2 is not the main factor explaining our results. Neverthe-less, further work is necessary to definitively confirm this idea.
Although other factors could play a role in peroxynitrite-mediated impairment of lymphocyte signaling proliferation andsurvival, our results suggest that nitration-mediated impairment ofprotein tyrosine phosphorylation plays a major role in this process
FIGURE 12. Immunohistochemical staining for protein nitrotyrosine residues of human lymph nodes.A, Topographic view showing subcapsular andmedular sinuses and scanty cortical immunoreactivity with anti-nitrotyrosine Abs (diaminobenzidene; original magnification,340). B, Subcapsular sinusplenty of positive macrophages (diaminobenzidene; original magnification,3400).C, Dilated sinuses containing large numbers of sinus histiocytes withstrong cytoplasmic immunolabeling (diaminobenzidene; original magnification,3100).D, Tissue section showing multiple parenchymal macrophages withstrong granular reactivity and a poorly defined perilymphocyte and lymphocyte labeling with random distribution (diaminobenzidene; original magnifi-cation,3400). Stars show sinuses. Thin arrows indicate lymphoid follicles with germinal center and a well-defined mantle zone. The thick arrow indicatesa subcapsular sinus. Asterisks show parenchymal macrophages, and the arrowhead shows perilymphocyte immunostaining.
FIGURE 13. Cytoplasmic immunostaining of nitrotyrosine in macro-phages from human mediastinal lymph node specimens. Weak immuno-staining in lymphocytic and perilymphocytic areas showing several mac-rophages with a strong specific immunoreactivity (aminoethylcarbazol;original magnification,3100).Inset, Cytoplasmic granular pattern of anti-nitrotyrosine reactivity in macrophages (aminoethylcarbazol; original mag-nification,31000).
3364 PEROXYNITRITE AND IMMUNE ACTIVATION
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

through a defective coupling of the TCR-CD3 complex to proxi-mal intracellular events such as tyrosine phosphorylation (67, 70–73). In addition, we provide strong experimental evidence for theendogenous production of nitrating species by cells of the mono-cyte-macrophage lineage in cultures of PBMC activated either bythe CD3 pathway or in the presence of PMA plus ionophores.Generation of reactive nitrogen intermediates by human mono-cytes/macrophages requires the production of nitric oxide as oneessential precursor for the generation of nitrating species either invivo or in vitro. Indeed, evidence does exist for the identificationof the endothelial constitutive and inducible isoforms of nitric ox-ide synthase mRNA, nitric oxide synthase protein, and nitric oxideproduction in human monocytes and macrophages (71–73). On theother hand, recent work demonstrated that nitrotyrosine is not theunique biomarker of peroxynitrite production and other NO-derivednitrating species could be formed in vivo (45). In this respect it hasbeen recently reported that nitrite (NO2
2), a major end productderived from nitric oxide metabolism, readily promotes tyrosinenitration through formation of nitryl chloride (NO2Cl) and nitro-gen dioxide (NO2) through reaction with the inflammatory medi-ator hypochlorous acid (HOCl) or myeloperoxidase (74–76).
Thus, it has been proposed that the nitrite oxidation mechanismsare probably relevant during inflammatory processes, where neu-trophils are recruited and activated, providing an additional path-way for the generation of reactive nitrating intermediates. Al-though with the present evidence, it cannot be definitively statedthat the formation of nitrotyrosine in vitro is due to peroxynitrite,the absence of neutrophils in our culture conditions and the resultsof experiments performed with Mn-TBAP associated or not asso-ciated with aminoguanidine favor the view that under our exper-imental conditions formation of nitrotyrosine was dependent onthe presence of superoxide and NOS activity. These argumentssupport the idea that peroxynitrite is the main nitrating species.Our results also showed that monocyte-derived nitrating specieswere able to induce long-lasting protein tyrosine nitration of theirown cellular proteins and neighboring lymphocytes present inwhole cultures. The persistence of nitrated proteins over time sug-gests that lymphocytes lack the enzymatic ability to remove nitrogroups from tyrosine residues as has been demonstrated fortyrosine kinases/phosphatases.
It is presently unclear whether activated lymphocytes are able toinduce the production of nitrating species by monocytes eitherthrough membrane-associated interactions or through soluble media-tors as cytokines, including TNF-a (77). The pattern of nitrated pro-teins after in vitro activation showed that the proteins more heavilynitrated were principally concentrated,50 kDa. However, the patternof tyrosine nitration induced by authentic peroxynitrite (Fig. 9)evolves over time to acquire a profile indistinguishable from that in-duced by in vitro activation (data not shown). In this respect it hasbeen demonstrated that peroxynitrite-induced protein modificationsare associated with increased degradation, which could explain ourresults (33). Since our results suggested that the pattern of nitratedproteins induced during immune activation might be the consequenceof endogenously produced ONOO2, we have showed in vivo thatlymph nodes from surgical specimens with histological evidence ofhyperplasia and nonspecific immune activation displayed a strong pat-tern of reactivity with anti-nitrotyrosine polyclonal Abs. Although wecannot elucidate which species nitrated tyrosines, the results show thegeneration of nitrating species in vivo.
Overall, our results suggest a role for nitrating species such asperoxynitrite as major modulators of immune responses. This as-sumption is reinforced by the enhancement of lymphocyte prolif-
erative responses in conditions of low to moderate inhibition ofeither zNO or O2
. (Fig. 11), which provides experimental evidencefor the physiological relevance of peroxynitrite-mediated tyrosinenitration. Our results also lead to the idea that this effect is at leastin part mediated through nitration of tyrosine residues, which re-sult in inhibition of tyrosine phosphorylation. In this respect, onecan speculate that upon activation only heavily tyrosine-phospho-rylated lymphocytes will proceed to acquire adequate effector andregulator functions. Besides the potential of peroxynitrite as adown-modulator of normal immune responses, nitration of cellularproteins may be injurious via multiple mechanisms, including al-tered protein function, increased turnover due to enhanced prote-olysis, and formation of neoantigens with eventual autoimmunereactions. Thus, peroxynitrite generation in vivo may represent amajor mediator in inflammation-mediated protein modification andtissue injury at sites of chronic or intense immune activation.
AcknowledgmentsWe thank Beatriz Alvarez and Gerardo Ferrer (Facultad de Medicina, Uni-versidad de la Republica) for technical assistance. We are also grateful forhelpful discussions with Otto Pritsch and Gerard Dumas (Facultad de Me-dicina, Universidad de la Republica, and Immunohematology Unit, PasteurInstitut-Paris, respectively).
References1. Skaper, S. D., L. Facci, and A. Leon. 1995. Inflammatory mediator stimulation of
astrocytes and meningeal fibroblasts induces neuronal degeneration via the nitri-dergic pathway.J. Neurochem. 64:266.
2. Ignarro, L. J. 1993. Nitric oxide-mediated vasorelaxation.Thromb. Haemost. 70:148.
3. Hibbs, J. J., R. R. Taintor, Z. Vavrin, and E. M. Rachlin. 1988. Nitric oxide: acytotoxic activated macrophage effector molecule.Biochem. Biophys. Res. Com-mun. 157:87.
4. Hibbs, J. R. J. 1991. Synthesis of nitric oxide fromL-arginine: a recently dis-covered pathway induced by cytokines with antitumour and antimicrobial activ-ity. Res. Immunol. 142:565.
5. Moncada, S., R. M. Palmer, and E. A. Higgs. 1991. Nitric oxide: physiology,pathophysiology, and pharmacology.Pharmacol. Rev. 43:109.
6. Crow, J. P., and J. S. Beckman. 1995. The role of peroxynitrite in nitric oxide-mediated toxicity.Curr. Top. Microbiol. Immunol. 196:57.
7. Beckman, J. S., and W. H. Koppenol. 1996. Nitric oxide, superoxide, and per-oxynitrite: the good, the bad, and the ugly.Am. J. Physiol. 271:C1424.
8. Cassina, A., and R. Radi. 1996. Differential inhibitory action of nitric oxide andperoxynitrite on mitochondrial electron transport.Arch. Biochem. Biophys. 328:309.
9. Brown, G. C., A. G. McBride, E. J. Fox, K. S. McNaught, and V. Borutaite. 1997.Nitric oxide and oxygen metabolism.Biochem. Soc. Trans. 25:901.
10. Lepoivre, M., F. Fieschi, J. Coves, L. Thelander, and M. Fontecave. 1991. In-activation of ribonucleotide reductase by nitric oxide.Biochem. Biophys. Res.Commun. 179:442.
11. Nguyen, T., D. Brunson, C. L. Crespi, B. W. Penman, J. S. Wishnok, andS. R. Tannenbaum. 1992. DNA damage and mutation in human cells exposed tonitric oxide in vitro.Proc. Natl. Acad. Sci. USA 89:3030.
12. Dawson, V. L., T. M. Dawson, D. A. Bartley, G. R. Uhl, and S. H. Snyder. 1993.Mechanisms of nitric oxide-mediated neurotoxicity in primary brain cultures.J. Neurosci. 13:2651.
13. Jaeschke, H., V. B. Schini, and A. Farhood. 1992. Role of nitric oxide in theoxidant stress during ischemia/reperfusion injury of the liver.Life Sci. 50:1797.
14. Kroncke, K. D., V. Kolb-Bachofen, B. Berschick, V. Burkart, and H. Kolb. 1991.Activated macrophages kill pancreatic syngeneic islet cells via arginine-depen-dent nitric oxide generation.Biochem. Biophys. Res. Commun. 175:752.
15. Hoffman, R. A., J. M. Langrehr, and R. L. Simmons. 1992. The role of induciblenitric oxide synthetase during graft-versus-host disease.Transplant. Proc. 24:2856.
16. Green, S. J., S. Mellouk, S. L. Hoffman, M. S. Meltzer, and C. A. Nacy. 1990.Cellular mechanism of nonspecific immunity to intracellular infection: cytokine-induced synthesis of toxic nitrogen oxides fromL-arginine by macrophages andhepatocytes.Immunol. Lett. 330:181.
17. Beckman, J. S., T. W. Beckman, J. Chen, P. A. Marshall, and B. A. Freeman.1990. Apparent hydroxyl radical production by peroxynitrite: implications forendothelial injury from nitric oxide and superoxide.Proc. Natl. Acad. Sci. USA87:1620.
18. Radi, R., J. S. Beckman, K. M. Bush, and B. A. Freeman. 1991a. Peroxynitriteoxidation of sulfhydryls: the cytotoxic potential of superoxide and nitric oxide.J. Biol. Chem. 266:4244.
19. Radi, R., J. S. Beckman, K. M. Bush, and B. A. Freeman. 1991b. Peroxynitrite-induced membrane lipid peroxidation: the cytotoxic potential of superoxide andnitric oxide.Arch. Biochem. Biophys. 288:481.
3365The Journal of Immunology
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

20. Huie, R. E., and S. Padmaja. 1993. The reaction of NO with superoxide.FreeRadical Res. Commun. 18:195.
21. Inoue, S., and S. Kawanishi. 1995. Oxidative damage induced by simultaneousgeneration of nitric oxide and superoxide.FEBS Lett. 371:86.
22. Ischiropoulos, H., and A. B. Al-Mehdi. 1995. Peroxynitrite-mediated oxidativeprotein modifications.FEBS Lett. 364:279.
23. Ischiropoulos, H., L. Zhu, J. Chen, M. Tsai, J. C. Martin, C. D. Smith, andJ. S. Beckman. 1992. Peroxynitrite-mediated tyrosine nitration catalyzed by su-peroxide dismutase.Arch. Biochem. Biophys. 298:431.
24. Crow, J. P., and H. Ischiropoulos. 1996. Detection and quantitation of nitroty-rosine residues in proteins: in vivo marker of peroxynitrite.Methods Enzymol.269:185.
25. Carreras, M. C., G. A. Pargament, S. D. Catz, J. J. Poderoso, and A. Boveris.1994. Kinetics of nitric oxide and hydrogen peroxide production and formation ofperoxynitrite during the respiratory burst of human neutrophils.FEBS Lett. 341:65.
26. Kooy, N. W., and J. A. Royall. 1994. Agonist-induced peroxynitrite productionfrom endothelial cells.Arch. Biochem. Biophys. 310:352.
27. Estevez, A. G., N. Spear, S. M. Manuel, R. Radi, C. E. Henderson, L. Barbeito,and J. S. Beckman. 1998. Nitric oxide and superoxide contribute to motor neuronapoptosis induced by trophic factor deprivation.J. Neurosci. 18:923.
28. Beckman, J. S., Y. Zu Ye, P. G. Anderson, J. Chen, M. A. Accavitti,M. M. Tarpey, and C. R. White. 1994. Extensive nitration of protein tyrosines inhuman atherosclerosis detected by immunohistochemistry.Biol. Chem. Hoppe-Seyler 375:81.
29. Haddad, I. Y., G. Pataky, P. Hu, C. Galliani, J. S. Beckman, and S. Matalon.1994. Quantitation of nitrotyrosine levels in lung sections of patients and animalswith acute lung injury.J. Clin. Invest. 94:2407.
30. Kooy, N. W., J. A. Royall, Y. Z. Ye, D. R. Kelly, and J. S. Beckman. 1995.Evidence for in vivo peroxynitrite production in human acute lung injury.Am J. Respir. Crit. Care Med. 151:1250.
31. Kaur, H., and B. Halliwell. 1994. Evidence for nitric oxide-mediated oxidativedamage in chronic inflammation: nitrotyrosine in serum and synovial fluid fromrheumatoid patients.FEBS Lett. 350:9.
32. Mandoro, T. H., B. C. Shafer, and J. G. Vostal. 1997. Peroxynitrite-inducedtyrosine nitration and phosphorylation in human platelets.Free Radical Biol.Med. 22:1055.
33. Gow, A. J., D. Duran, S. Malcolm, and H. Ischiropoulos. 1996. Effects of per-oxynitrite-induced protein modifications on tyrosine phosphorylation and degra-dation.FEBS Lett. 385:63.
34. Kong, S. K., M. B. Yim, E. R. Stadtman, and P. B. Chock. 1996. Peroxynitritedisables the tyrosine phosphorylation regulatory mechanism: lymphocyte-specifictyrosine kinase fails to phosphorylate nitrated cdc2(6–20)NH2 peptide.Proc.Natl. Acad. Sci. USA 93:3377.
35. June, C. H., M. C. Fletcher, J. A. Ledbetter, and L. E. Samelson. 1990. Increasesin tyrosine phosphorylation are detectable before phospholipase C activation afterT cell receptor stimulation.J. Immunol. 144:1591.
36. Isakov, N. 1993. Tyrosine phosphorylation and dephosphorylation in T lympho-cyte activation.Mol. Immunol. 30:197.
37. Baeuerle, P. A., and T. Henkel. 1994. Function and activation of NF-kappa B inthe immune system.Annu. Rev. Immunol. 12:141.
38. Sen, C. K., and L. Packer. 1996. Antioxidant and redox regulation of gene tran-scription.FASEB J. 10:709.
39. Fadok, V. A., D. R. Voelker, P. A. Campbell, J. J. Cohen, D. L. Bratton, andP. M. Henson. 1992. Exposure of phosphatidylserine on the surface of apoptoticlymphocytes triggers specific recognition and removal by macrophages.J. Im-munol. 148:2207.
40. McEvoy, L., R. A. Schlegel, P. Williamson, and B. J. Del Buono. 1988. Mero-cyanine 540 as a flow cytometric probe of membrane lipid organization in leu-kocytes.J. Leukocyte Biol. 44:337.
41. Ye, Y. Z., M. Strong, Z. Q. Huang, and J. S. Beckman. 1996. Antibodies thatrecognize nitrotyrosine.Methods Enzymol. 269:201.
42. Faulkner, K. M., S. I. Liochev, and I. Fridovich. 1994. Stable Mn(III) porphyrinsmimic superoxide dismutase in vitro and substitute for it in vivo.J. Biol. Chem.268:23471.
43. Southan, G. J., and C. Szabo. 1996. Selective pharmacological inhibition of dis-tinct nitric oxide synthase isoforms.Biochem. Pharmacol. 51:383.
44. Wolff, D. J., and A. Lubeskie. 1995. Aminoguanidine is an isoform-selective,mechanism-based inactivator of nitric oxide synthase.Arch. Biochem. Biophys.316:290.
45. Halliwell, B. 1997. What nitrates tyrosine? Is nitrotyrosine specific as a biomar-ker of peroxynitrite formation in vivo?FEBS Lett. 411:157.
46. Lin, T. K., J. Y. Xue, M. Nomen, B. Spur, and P. Y. K. Wong. 1995. Peroxyni-trite-induced apoptosis in HL-60 cells.J. Biol. Chem. 270:16487.
47. Estevez, A. G., R. Radi, L. Barbeito, J. T. Shin, J. A. Thompson, andJ. S. Beckman. 1995. Peroxynitrite-induced cytotoxicity in PC-12 cells: evidencefor an apoptotic mechanism differentially modulated by neurotrophic factors.J. Neurochem. 65:1543.
48. Sandoval, M., X. J. Zhang, X. Liu, E. E. Mannick, D. A. Clark, and M. J. Miller.1997. Peroxynitrite-induced apoptosis in T84 and RAW 264.7 cells: attenuationby L-ascorbic acid.Free Radical Biol. Med. 22:489.
49. Salgo, M. G., K. Stone, G. L. Squadrito, J. R. Battista, and W. A. Pryor. 1995.Peroxynitrite causes DNA nicks in plasmid pBR322.Biochem. Biophys. Res.Commun. 210:1025.
50. Szabo, C. 1996. DNA strand breakage and activation of poly-ADP ribosyltrans-ferase: a cytotoxic pathway triggered by peroxynitrite.Free Radical Biol. Med.21:855.
51. Rice, W. G., S. D. Hillyer, B. Harten, C. A. Schaeffer, M. Dorminy,D. A. Lackey, E. Kirstein, J. Mendeleyev, K. Buki, A. Hakam, et al. 1992.Induction of endonuclease-mediated apoptosis in tumor cells by C-nitroso-sub-stituted ligands of poly(ADP-ribose) polymerase.Proc. Natl. Acad. Sci. USA89:7703.
52. Ueta, K., and O. Hayashi. 1985. ADP-ribosylation.Annu. Rev. Biochem. 54:73.53. Virag, L., G. S. Scott, S. Cuzzocrea, D. Marmer, A. L. Salzman, and C. Szabo´.
1998. Peroxynitrite-induced thymocyte apoptosis: the role of caspases and poly(ADP-ribose) synthase (PARS) activation.Immunology 94:345.
54. Nicholson, D. W., A. All, N. A. Thornberry, J. P. Vaillantcourt, J. P. Ding,C. K. Gallant, Y. Gareau, P. R. Griffin, M. Labelle, Y. A. Lazebnik, et al. 1995.Identification and inhibition of the ICE/CED-3 protease necessary for mammalianapoptosis.Nature 376:37.
55. Lin, T. K., J. Y. Xue, and P. Y. K. Wong. 1997. Bcl-2 blocks peroxynitrite-induced apoptosis in HL-60 cells, an association with reactive oxygen species.Inflamm. Res. 46:S157.
56. Martin, B. L., D. Wu, S. Jakes, and D. J. Graves. 1990. Chemical influences onthe specificity of tyrosine phosphorylation.J. Biol. Chem. 265:7108.
57. van der Vliet, A., J. P. Eiserich, C. A. O’Neill, B. Halliwell, and E. C. Cross.1995. Tyrosine modification by reactive nitrogen species: a closer look.Arch.Biochem. Biophys. 319:341.
58. Mallozzi, C., A. M. Di Stasi, and M. Minetti. 1997. Peroxynitrite modulatestyrosine-dependent signal transduction pathway of human erythrocyte band 3.FASEB J. 11:1281.
59. Rubbo, H., R. Radi, M. Trujillo, R. Telleri, B. Kalyanaraman, S. Barnes, S. Kirk,and B. A. Freeman. 1994. Nitric oxide regulation of superoxide and peroxynitrite-dependent lipid peroxidation: formation of novel nitrogen-containing oxidizedlipid derivatives.J. Biol. Chem. 269:26066.
60. Kanner, S. B., T. J. Kavanagh, A. Grossmann, S. L. Hu, J. B. Bolen,P. S. Rabinovitch, and J. A. Ledbetter. 1992. Sulfhydryl oxidation down-regu-lates T-cell signalling and inhibits tyrosine phosphorylation of phospholipaseCg1. Proc. Natl. Acad. Sci. USA 89:300.
61. Staal, F. J., M. T. Anderson, G. E. Staal, L. A. Herzenberg, C. Gitler, andL. A. Herzenberg. 1994. Redox regulation of signal transduction: tyrosine phos-phorylation and calcium influx.Proc. Natl. Acad. Sci. USA 91:3619.
62. Smyth, M. J. 1991. Glutathione modulates activation-dependent proliferation ofhuman peripheral blood lymphocyte populations without regulating their acti-vated function.J. Immunol. 146:1921.
63. Kavanagh, T. J., A. Grossmann, E. P. Jaecks, J. C. Jinneman, D. L. Eaton,G. M. Martin, and P. S. Rabinovitch. 1990. Proliferative capacity of humanperipheral blood lymphocytes sorted on the basis of glutathione content.J. Cell.Physiol. 145:472.
64. Nakamura, H., K. Nakamura, and J. Yodoi. 1997. Redox regulation of cellularactivation.Annu. Rev. Immunol. 15:351.
65. Los, M., W. Droge, K. Stricker, P. A. Baeuerle, and K. Schulze-Osthoff. 1995.Hydrogen peroxide as a potent activator of T lymphocyte functions.Eur. J. Im-munol. 125:159.
66. Schieven, G. L., J. M. Kirihara, D. E. Myers, J. A. Ledbetter, and F. M. Uckun.1993. Reactive oxygen intermediates activate NF-kB in a tyrosine kinase-depen-dent mechanism and in combination with vanadate activate the p56lck and p59fyn
tyrosine kinases in human lymphocytes.Blood 82:1212.67. Cayota, A., F. Vuillier, G. Gonzalez, and G. Dighiero. 1997. In vitro antioxidant
treatment recovers proliferative responses of anergic CD41 lymphocytes fromHIV-infected individuals.Blood 87:4746.
68. Devary, Y., R. A. Gottlieb, T. Smeal, and M. Karin. 1992. The mammalianultraviolet response is triggered by activation of Src kinases.Cell 71:1081.
69. Guyton, K. Z., Y. Liu, M. Gorospe, Q. Xu, and N. J. Holbrook. 1996. Activationof mitogen activated protein kinase: role in cell survival following oxidant injury.J. Biol. Chem. 271:7138.
70. Radi, R. 1996. Kinetic analysis of reactivity of peroxynitrite with biomolecules.Methods Enzymol. 269:354.
71. Dugas, B., D. Mossalayi, C. Damais, and J. P. Kolb. 1995. Nitric oxide produc-tion by human monocytes: evidence for a role of CD23.Immunol. Today 16:574.
72. Paul-Eugene, N., D. Mossalayi, M. Sarfati, K. Yamaoka, J. P. Aubry,J. Y. Bonnefoy, B. Dugas, and J. P. Kolb. 1995. Evidence for a role of FceRII/CD23 in the IL-4 induced nitric oxide production by normal human mononuclearphagocytes.Cell. Immunol. 163:314.
73. Weinberg, J. B., M. A. Misukonis, P. J. Shami, S. N. Mason, D. L. Sauls,W. A. Dittman, E. R. Wood, G. K. Smith, B. McDonald, K. E. Bachus, et al.1995. Human mononuclear phagocyte inducible nitric oxide synthase (iNOS):analysis of iNOS mRNA, iNOS protein, biopterin, and nitric oxide production byblood monocytes and peritoneal macrophages.Blood 86:1184.
74. van der Vliet, A., J. P. Eiserich, B. Halliwell, and C. A. Cross. 1997. Formationof reactive nitrogen species during peroxidase-catalyzed oxidation of nitrite.J. Biol. Chem. 272:7617.
75. Eiserich, J. P., M. Hristova, C. E. Cross, A. D. Jones, B. A. Freeman,B. Halliwell, and A. van der Vliet. 1998. Formation of nitric oxide-derived in-flammatory oxidants by myeloperoxidase in neutrophils.Nature 391:393.
76. Eiserich, J. P., C. E. Cross, A. D. Jones, B. Halliwell, and A. van der Vliet. 1996.Formation of nitrating and chlorinating species by reaction of nitrite with hypo-chlorous acid: a novel mechanism for nitric oxide-mediated protein modification.J. Biol. Chem. 271:19199.
77. Phelps, D. T., T. J. Ferro, P. J. Higgins, R. Shankar, D. M. Parker, andA. Johnson. 1995. TNF-a induces peroxynitrite-mediated depletion of lung en-dothelial glutathione via protein kinase C.Am. J. Physiol. 269:L551.
3366 PEROXYNITRITE AND IMMUNE ACTIVATION
by guest on June 16, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Related Documents











