Pharmacological Research 53 (2006) 501–507 Peroxisome proliferator-activated receptor (PPAR), a novel target site for drug discovery in metabolic syndrome Sadao Takahashi a , Toshiya Tanaka b , Tatsuhiko Kodama b , Juro Sakai b,c,∗ a Third Department of Internal Medicine, Faculty of Medical Sciences, University of Fukui, Fukui 910-0063, Japan b Laboratory for Systems Biology and Medicine, Research Center for Advanced Science and Technology, University of Tokyo, Komaba 153-8904, Japan c Exploratory Research for Advanced Technology of Japan Science and Technology Corporation, Aomi 153-0064, Japan Abstract The development of new treatments for metabolic syndrome is urgent project for decreasing the prevalence of coronary heart disease and diabetes mellitus in the advanced countries. Peroxisome proliferator-activated receptor (PPAR) and agonists have shed light on the treatment of hypertriglyceridemia and type 2 diabetes mellitus, respectively. Among PPARs, analysis of the PPAR functions is lagging behind because specific PPAR agonists have not been developed. The appearance of new PPAR agonists is brightening the prospects for elucidating the physiological role of PPAR. PPAR is a new target for the treatment of metabolic syndrome. In particular, the fact that fatty acid oxidation and energy dissipation in skeletal muscle and adipose tissue by PPAR agonists lead to improved lipid profile, reduced adiposity and insulin sensitivity is a breakthrough. It seems that treatment of PPAR agonists operate similarly to the caloric restriction and prolonged exercise. We suggest that the physiological role of PPAR may be an indicator for switching from glucose metabolism to fatty acid metabolism. To receive new benefits of PPAR agonists against metabolic syndrome by increasing fatty acid consumption in skeletal muscle and adipose tissue, we need to unveil more details on the functions of PPAR itself and its agonists in the future. © 2006 Elsevier Ltd. All rights reserved. Keywords: PPAR; Metabolic syndrome; Fatty acids oxidation; Adaptive thermogenesis The prevalence of obesity has risen dramatically in devel- oped countries. This phenomenon has led to an increase in the so-called metabolic syndrome. Because metabolic syndrome is associated with increased risk of coronary heart disease (CHD) and overt diabetes, it is important to prevent progress of serious diseases at an early stage. Although the precise reason for the global epidemic of obesity and metabolic syndrome has not been completely elucidated, it is accepted that environmental factors and changes in life style are major contributors. Metabolic syndrome is composed of abdominal obesity, hyperglycemia due to insulin resistance, atherogenic dyslipi- demia and hypertension [1]. Based on the Third Report of the National Cholesterol Education Program Expert Panel on Detection, Evaluation, and Treatment of High Blood Choles- terol in Adults (Adult Treatment Panel III), it has been reported that approximately 22% of adults in the USA have metabolic syndrome [2]. Furthermore based on data from the Third ∗ Corresponding author. Tel.: +81 3 5452 5472. E-mail address: [email protected] (J. Sakai). National Health and Nutrition Examination Survey (NHANES III), Ninomiya et al. [3] have evaluated the association between metabolic syndrome and history of myocardial infarction and/or stroke, and have given a definite opinion that the presence of metabolic syndrome identifies a cohort of the population that is at a substantial risk of cardiovascular disease. To address the major public health concerns faced by developed countries, numerous researches have emerged for elucidating the mecha- nisms of metabolic syndrome. Among them, peroxisome proliferator-activated receptors (PPARs) agonists have shed light on effective therapeutic approaches for correcting some aspects of metabolic syndrome. PPARs are ligand-activated transcription factors that control fatty acid metabolism, and members of the nuclear receptor superfamily. Together with retinoid X receptors (RXRs), the het- erodimers bind to the PPAR-responsive element (PPRE), which is a direct repeat of two similar hexanucleotide (5-AGGTCA- 3) half-sites separated by one or two nucleotides on its target genes. Three subtypes, PPAR, PPAR/ (refer to PPAR in this review), and PPAR have been identified with distinct tis- sue distribution and biological activities [4]. The roles of PPAR 1043-6618/$ – see front matter © 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.phrs.2006.03.019

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pharmacological Research 53 (2006) 501–507
Peroxisome proliferator-activated receptor � (PPAR�), a noveltarget site for drug discovery in metabolic syndrome
Sadao Takahashi a, Toshiya Tanaka b, Tatsuhiko Kodama b, Juro Sakai b,c,∗a Third Department of Internal Medicine, Faculty of Medical Sciences, University of Fukui, Fukui 910-0063, Japan
b Laboratory for Systems Biology and Medicine, Research Center for Advanced Science and Technology, University of Tokyo, Komaba 153-8904, Japanc Exploratory Research for Advanced Technology of Japan Science and Technology Corporation, Aomi 153-0064, Japan
Abstract
The development of new treatments for metabolic syndrome is urgent project for decreasing the prevalence of coronary heart disease anddiabetes mellitus in the advanced countries. Peroxisome proliferator-activated receptor (PPAR)� and � agonists have shed light on the treatment ofhypertriglyceridemia and type 2 diabetes mellitus, respectively. Among PPARs, analysis of the PPAR� functions is lagging behind because specificPPAR� agonists have not been developed. The appearance of new PPAR� agonists is brightening the prospects for elucidating the physiologicalrole of PPAR�. PPAR� is a new target for the treatment of metabolic syndrome. In particular, the fact that fatty acid oxidation and energy dissipationin skeletal muscle and adipose tissue by PPAR� agonists lead to improved lipid profile, reduced adiposity and insulin sensitivity is a breakthrough.Iomo©
K
osaadgca
hdtDtts
1d
t seems that treatment of PPAR� agonists operate similarly to the caloric restriction and prolonged exercise. We suggest that the physiological rolef PPAR� may be an indicator for switching from glucose metabolism to fatty acid metabolism. To receive new benefits of PPAR� agonists againstetabolic syndrome by increasing fatty acid consumption in skeletal muscle and adipose tissue, we need to unveil more details on the functions
f PPAR� itself and its agonists in the future.2006 Elsevier Ltd. All rights reserved.
eywords: PPAR�; Metabolic syndrome; Fatty acids oxidation; Adaptive thermogenesis
The prevalence of obesity has risen dramatically in devel-ped countries. This phenomenon has led to an increase in theo-called metabolic syndrome. Because metabolic syndrome isssociated with increased risk of coronary heart disease (CHD)nd overt diabetes, it is important to prevent progress of seriousiseases at an early stage. Although the precise reason for thelobal epidemic of obesity and metabolic syndrome has not beenompletely elucidated, it is accepted that environmental factorsnd changes in life style are major contributors.
Metabolic syndrome is composed of abdominal obesity,yperglycemia due to insulin resistance, atherogenic dyslipi-emia and hypertension [1]. Based on the Third Report ofhe National Cholesterol Education Program Expert Panel onetection, Evaluation, and Treatment of High Blood Choles-
erol in Adults (Adult Treatment Panel III), it has been reportedhat approximately 22% of adults in the USA have metabolicyndrome [2]. Furthermore based on data from the Third
∗ Corresponding author. Tel.: +81 3 5452 5472.E-mail address: [email protected] (J. Sakai).
National Health and Nutrition Examination Survey (NHANESIII), Ninomiya et al. [3] have evaluated the association betweenmetabolic syndrome and history of myocardial infarction and/orstroke, and have given a definite opinion that the presence ofmetabolic syndrome identifies a cohort of the population thatis at a substantial risk of cardiovascular disease. To addressthe major public health concerns faced by developed countries,numerous researches have emerged for elucidating the mecha-nisms of metabolic syndrome.
Among them, peroxisome proliferator-activated receptors(PPARs) agonists have shed light on effective therapeuticapproaches for correcting some aspects of metabolic syndrome.PPARs are ligand-activated transcription factors that controlfatty acid metabolism, and members of the nuclear receptorsuperfamily. Together with retinoid X receptors (RXRs), the het-erodimers bind to the PPAR-responsive element (PPRE), whichis a direct repeat of two similar hexanucleotide (5-AGGTCA-3) half-sites separated by one or two nucleotides on its targetgenes. Three subtypes, PPAR�, PPAR�/� (refer to PPAR� inthis review), and PPAR� have been identified with distinct tis-sue distribution and biological activities [4]. The roles of PPAR�
043-6618/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.oi:10.1016/j.phrs.2006.03.019

502 S. Takahashi et al. / Pharmacological Research 53 (2006) 501–507
and PPAR� in particular have been exclusively studied becausetheir agonists have been used in the clinical market. PPAR�that is primarily expressed in brown adipose tissue (BAT) andliver, and to a lesser extent in the kidney, skeletal muscle, andheart, binds to hypolipidemic fibrates and natural ligands includ-ing leukotriene B4 (LTB4) and 8-(S)-hydroxyeicosatetraenoicacid (8S-HETE). Fasted PPAR�−/− (knockout) mice showenhanced accumulation of lipid in the liver and heart due tosufferings from severe hypoglycemia and hypothermia. It sug-gests that PPAR� plays a crucial role in fatty acid uptake andoxidation in the liver and heart [5,6]. On the other hand, PPAR�is highly rich in adipocytes and macrophages, and is involved inadipocyte differentiation, lipid storage, and glucose homeosta-sis. The insulin-sensitizing thiazolidinediones (TZDs), whichare selective ligands of PPAR� consistently lower fasting andpostprandial glucose levels as well as free fatty acid levels. Sev-eral established mechanisms for TZDs have been reviewed indetail [4,7]. To the contrary, PPAR� is expressed ubiquitouslywith a less defined function because selective agonists have notbeen obtained. However, the development of selective PPAR�activators, has allowed investigations on the functions of PPAR�progress. In this review, we focus mainly on the recently eluci-dated functions of PPAR�.
1. Tissue distribution and ligands of PPAR�
sdPtmkPtt(PGGoleiemclsldi
2
l
lacking the ligand-binding domain of the murine PPAR� gene[11]. PPAR� −/− mice are smaller than normal from the fetalstage to the postnatal period and myelination of the corpus cal-losum in brain is altered. Gonadal adipose stores are smaller andCD 36 mRNA levels are higher. Enhanced hyperplasia of the epi-dermis has been detected in the skin of PPAR� −/− mice. Theysuggested that PPAR� is involved in development, myelinationin the brain, lipid metabolism, and epidermal cell proliferation.Meanwhile Barak et al. [12] also eliminated almost the entirePPAR� gene in mice. Over 90% of the embryos PPAR� −/−mice are prone to die due to placental defects and the micethat survive show an extremely lean phenotype. The differencebetween two knockout mice for lethality is likely due to the dif-ferent targeting strategies. The former truncated receptor micemight retain PPAR activity for survival. Michalik et al. [13] alsoindicated that only few PPAR� −/− mice could be obtained butno null mice line could be established for very high penetranceof a lethal phenotype.
3. Adipocyte differentiation
PPAR� is induced during adipocyte differentiation, and aforced expression of PPAR� in nonadipogenic cells effectivelyconverts them to mature adipocytes. Like PPAR�, it seems thatPPAR� might be involved in adipocyte differentiation. Grimaldiand co-workers [14] created the forced PPAR� expressing Swiss3foenTttactPtseipidPiaplaLtttam
Schmidt et al. [8] first discovered a novel member of theteroid hormone receptor superfamily cDNA as NUC1 (later onefined as PPAR�/�) from a human osteosarcoma cell library.PAR� is expressed ubiquitously and is the most abundant of
he three PPARs in most tissues except adipose tissue. PPAR�RNA levels are dramatically down-regulated in the liver and
idney by up to 80% after an overnight fast. On the other hand,PAR� expression is up-regulated in the liver and small intes-
ine, and PPAR� expression is decreased by 50% in adiposeissues in the fasted state in mature rat tissues [9]. ProstacyclinPGI2), carbaprostacyclin (cPGI2), and saturated fatty acids arePAR� ligands, but those ligands also activate PPAR� or �.laxoSmithKline first identified a high-affinity PPAR� ligand,W501516 that has more than 1000-fold selectivity for PPAR�ver other subtypes. Chawla et al. [10] reported that native veryow-density lipoprotein (VLDL) particles may be one of thendogenous sources of PPAR� ligands. Especially triglyceridesn VLDL that is hydrolyzed by lipoprotein lipase (LPL) are anfficient activator of PPAR� in macrophages. PPAR� deficientacrophages lost the transcriptional response by VLDL parti-
les. Those findings are interest because oxidized low-densityipoprotein (ox-LDL) activates PPAR� and induce the expres-ion of the scavenger receptor, CD36 and the oxysterol receptor,iver X receptor (LXR�). PPAR� activation induces adiposeifferentiation-related protein (ADRP) expression but does notnduce CD36 or LXR�.
. Homozygous PPAR� knockout mice
Two homozygous PPAR� −/− mice lines have been estab-ished. Gonzalez and colleagues established PPAR� −/− mice
T3C2 fibroblasts, which are normally refractory to adipose dif-erentiation. They found that a treatment of fatty acids aloner PPAR� agonist is not sufficient to trigger adipogenesis butxposure of cells to a combination of fatty acids and PPAR� ago-ist promotes the expression of a typical adipose differentiation.hey suggested that PPAR� gene expression is under the con-
rol of PPAR� activated by fatty acids. Furthermore they showedhat overexpression of PPAR� enhances fatty acid induction ofdipose-related genes in Ob1771 and 3T3-F442A preadipocyteells. A transactivation-deficient form of PPAR� mutated inhe AF2 domain severely reduced these effects indicating thatPAR� plays a central role in fatty acid-controlled differentia-
ion of preadipose cells [15]. Hansen et al. [16] concluded thatynergistic activation of endogenous PPAR� in preadipocytesnhances the expression of PPAR� but terminal differentiations only modestly promoted in 3T3-L1 and NIH-3T3 cells. PPAR�lays a role in the expansion of the pool of precursor cells. Thempact of PPAR� in adipocyte differentiation could be depen-ent on various types of preadipocyte cell lines, and it is true thatPAR� mainly regulates adipocyte differentiation. In contrast to
n vitro studies, adipose-specific PPAR� −/− mice do not affectdipose tissue mass [12]. Expression of activated PPAR� in adi-ose tissue (ligand independent active form of PPAR�) leads toeanness but features of irregular adipogenesis are not shown,nd no effect of PPAR� on adipocyte differentiation in 3T3-1 cells is re-confirmed [17]. Those in vivo data suggest that
he direct effect of PPAR� on adipose differentiation in adiposeissue in vitro is in the negative, and peripheral PPAR� func-ions in systemic lipid metabolism more than the direct effect ondipocyte differentiation that might be working in PPAR� −/−ice.
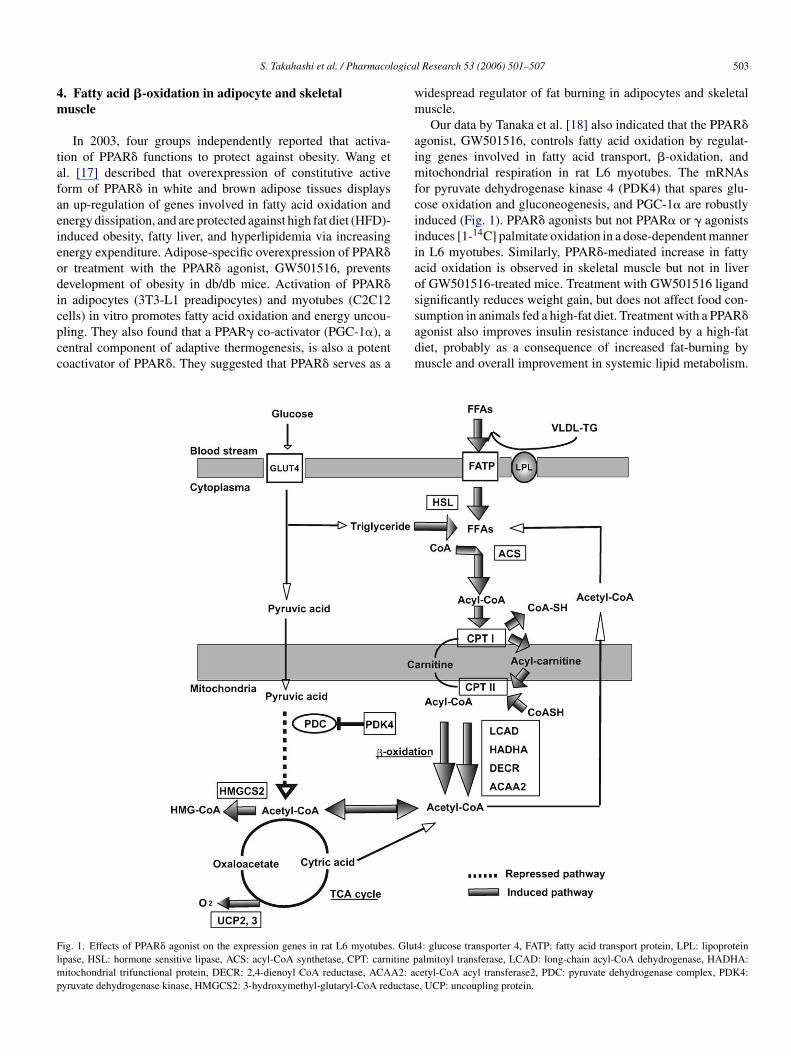
S. Takahashi et al. / Pharmacological Research 53 (2006) 501–507 503
4. Fatty acid �-oxidation in adipocyte and skeletalmuscle
In 2003, four groups independently reported that activa-tion of PPAR� functions to protect against obesity. Wang etal. [17] described that overexpression of constitutive activeform of PPAR� in white and brown adipose tissues displaysan up-regulation of genes involved in fatty acid oxidation andenergy dissipation, and are protected against high fat diet (HFD)-induced obesity, fatty liver, and hyperlipidemia via increasingenergy expenditure. Adipose-specific overexpression of PPAR�or treatment with the PPAR� agonist, GW501516, preventsdevelopment of obesity in db/db mice. Activation of PPAR�in adipocytes (3T3-L1 preadipocytes) and myotubes (C2C12cells) in vitro promotes fatty acid oxidation and energy uncou-pling. They also found that a PPAR� co-activator (PGC-1�), acentral component of adaptive thermogenesis, is also a potentcoactivator of PPAR�. They suggested that PPAR� serves as a
widespread regulator of fat burning in adipocytes and skeletalmuscle.
Our data by Tanaka et al. [18] also indicated that the PPAR�agonist, GW501516, controls fatty acid oxidation by regulat-ing genes involved in fatty acid transport, �-oxidation, andmitochondrial respiration in rat L6 myotubes. The mRNAsfor pyruvate dehydrogenase kinase 4 (PDK4) that spares glu-cose oxidation and gluconeogenesis, and PGC-1� are robustlyinduced (Fig. 1). PPAR� agonists but not PPAR� or � agonistsinduces [1-14C] palmitate oxidation in a dose-dependent mannerin L6 myotubes. Similarly, PPAR�-mediated increase in fattyacid oxidation is observed in skeletal muscle but not in liverof GW501516-treated mice. Treatment with GW501516 ligandsignificantly reduces weight gain, but does not affect food con-sumption in animals fed a high-fat diet. Treatment with a PPAR�agonist also improves insulin resistance induced by a high-fatdiet, probably as a consequence of increased fat-burning bymuscle and overall improvement in systemic lipid metabolism.
Flmp
ig. 1. Effects of PPAR� agonist on the expression genes in rat L6 myotubes. Glutipase, HSL: hormone sensitive lipase, ACS: acyl-CoA synthetase, CPT: carnitine p
itochondrial trifunctional protein, DECR: 2,4-dienoyl CoA reductase, ACAA2: ayruvate dehydrogenase kinase, HMGCS2: 3-hydroxymethyl-glutaryl-CoA reductase
4: glucose transporter 4, FATP: fatty acid transport protein, LPL: lipoproteinalmitoyl transferase, LCAD: long-chain acyl-CoA dehydrogenase, HADHA:
cetyl-CoA acyl transferase2, PDC: pyruvate dehydrogenase complex, PDK4:, UCP: uncoupling protein.

504 S. Takahashi et al. / Pharmacological Research 53 (2006) 501–507
Fig. 2. Model of PPAR� activation and gene expression in skeletal muscle induced by cold exposure, fasting, and prolonged exercise. PPAR: peroxisome proliferators-activated receptor, RXR: retinoid X receptor, PPRE: PPAR-responsive element, FATP: fatty acid transport protein, PGC-1�: PPAR� co-activator-1�.
GW501516 treatment prevents hepatic and intramuscular lipidaccumulation, and improves insulin sensitivity in high fat diet(HFD)-induced mice and ob/ob mice. It is interesting that thechanges in gene expression induced by GW501516 in culturedmyotubes and skeletal muscle in vivo are very similar to the geneexpression profile induced by cold exposure, fasting, and pro-longed exercise. We suggest that the physiological role of PPAR�may be a direct switch from glucose metabolism to fatty acidmetabolism. It is conceivable that free fatty acids released fromadipose tissues on fasting or exercise provide PPAR� ligandsto stimulate fatty acid oxidation and thermogenesis in skeletalmuscle (Fig. 2). This idea is consistent with the hypothesis thatPPAR� acts as a pivotal thermogenic transcription factor [17].Furthermore we observed that GW501516 treatment of the pan-creatic islets isolated from wild-type mice on normal chow dietsignificantly stimulated glucose-induced insulin release in vitro.It is important to investigate whether the effect of PPAR� ago-nists on the pancreas can prevent or delay �-cell loss in thedevelopment of type 2 diabetes.
Dressel et al. [19] reported a similar effect with GW501516in C2C12 myotubes, a line of mouse skeletal muscle cells, indi-cating that PPAR� agonists induces the expression of genesinvolved in lipid utilization, �-oxidation, cholesterol efflux, andenergy uncoupling. It is intriguing that PPAR� agonist increasesapolipoprotein-A1 specific efflux of intracellular cholesterol inskeletal muscle even though the effect has been reported inmuigCoirm
of oxidative myofibers is increased in muscles. Muscle remod-eling in muscle-specific PPAR� overexpression is reminiscentof that provoked by endurance training. They also showed thatmoderate exercise in wild-type mice increases PPAR� protein inmuscle compared to untrained mice in association with the effectof PPAR� in muscle for fatty acid �-oxidation. Wang et al. [23]showed that the targeted expression of constitutively active formof PPAR� in skeletal muscle or treatment with PPAR� agonistinduced oxidative type I fibers in muscle in 2004. Genes in thosemuscles for slow fiber contractile protein, mitochondrial bio-genesis, and �-oxidation are up-regulated. These fibers conferresistance to obesity with improved metabolic profiles. PPAR�transgenic mice remarkably increase and PPAR� −/− micedecrease the exercise endurance capacity. Most recently it hasbeen reported that PPAR� agonists (GW501516 and GW0472)increase glucose uptake independently of insulin in differenti-ated C2C12 and primary human skeletal myotubes. The effectwas coincident with increased expression and phosphoryla-tion of extracellular signal-related kinase (ERK)1/2 mitogen-activated kinase (MAPK), p38 MAPK, and AMP-activated pro-tein kinase (AMPK) [24]. PPAR� activation induces both fattyacid �-oxidation and glucose transport in skeletal muscle.
5. Lipoprotein metabolism
The PPAR� agonist, GW501516 dramatically increases high-dodica[l[
acrophages [20]. Holst et al. [21] indicated that 24 h of fastingp-regulates PPAR� expression in gastrocnemius muscle but notn the heart of mice, and PPAR� agonist GW1514 induces severalenes implicated in fatty acid uptake and metabolism in mouse2C12 myotubes. Furthermore, they generated an animal modelf mice overexpressing PPAR� specifically in skeletal musclen vivo [22]. Those mice promote a decrease in body fat contentelated to a reduced adipocyte size and increased oxidative enzy-atic activities. Histological examination shows that the number
ensity lipoprotein (HDL)-cholesterol level in insulin resistantbese rhesus monkeys, while lowering the levels of small-ense low-density lipoprotein, fasting triglycerides, and fastingnsulin [20]. Another PPAR� agonist, L-165041 increases HDL-holesterol levels in obese and diabetic db/db mice, but did notffect the plasma triglyceride (TG) levels [25]. Akiyama et al.26] investigated lipoprotein metabolism in PPAR� −/− miceacking the ligand-binding domain of the murine PPAR� gene11]. PPAR� −/− mice showed elevated plasma TG and free

S. Takahashi et al. / Pharmacological Research 53 (2006) 501–507 505
fatty acid (FFA) but similar total cholesterol (TC), free choles-terol (FC), and phospholipids (PL) on normal chow diet. Duringa high fat diet (HFD) challenge for 10 weeks, TG levels remainedsignificantly elevated in the PPAR� −/−mice. PPAR� −/− micehave an increased rate of hepatic VLDL production as well aslowered lipoprotein lipase (LPL) activity with increased hep-atic angiopoietin-like proteins (Angptl) 3 and 4 in PPAR� −/−mice compared to wild-mice. Those data are compatible withhypotriglyceridemia that was observed in targeted activation ofPPAR� [17]. However, in PPAR� −/− mice (almost completedeletion of the PPAR� gene) reported by Barak et al. [12], thereare no changes in TC, TG, HDL-C and FFA with a normal chowdiet; there is no data for lipoprotein profiles during a HFD chal-lenge. The reason why the two PPAR� −/− mice lines showeddifferent lipoprotein patterns in normal chow diet is unclear. Oneplausible reason is their method for the deletion of the PPAR�gene as mentioned above. The different results between effect ofPPAR� agonists or activation and PPAR� receptor deletion onlipoprotein metabolism are elusive. The physiological effects ofreceptor agonists may not necessarily be in direct opposition tothe effects of gene deletion of the receptor.
6. Atherosclerosis
In macrophages, GW501516 increased expression ofthe reverse cholesterol transporter ATP-binding cassette A1(ercamsIttPvmii(
imtaGidsipprtO
tor) increases MCP-1 levels in the primary macrophages bydecreasing the release of anti-inflammatory repressors. Theeffect is completely reversed by GW501516 (decreased unli-ganded PPAR� receptor induces anti-inflammatory repressorrelease). It seems that the amount of unliganded PPAR� recep-tor determines the inflammatory status. Those results indicatethat PPAR� agonists have anti-atherosclerotic effects in vivo,however, there is a contradictory paper reporting that GW0742,another PPAR� ligand, failed to decrease atherosclerosis in LDLreceptor −/− mice [32]. With regard to the lipoprotein profile ofHFD induced LDL receptor−/− mice, GW0742 had little effecton TC, HDL-C, and LDL-C but modestly decreased VLDL orTG [32,33].
7. Cardiomyocytes
The effect of PPAR� on skeletal muscles has expanded theresearch field to cardiac lipid metabolism. Gilde et al. [34]reported that PPAR� and PPAR�, but not PPAR� play a promi-nent role in the regulation of cardiac lipid metabolism. Moreoverthe PPAR� agonist, GW0742, partly restored the expressionof key genes of fatty acid oxidation in mouse adult cardiomy-ocytes isolated from PPAR� −/− mice [35]. Cheng et al. [36]examined myocardial fatty acid oxidation in cre-loxP-mediatedcardiomyocyte-restricted deletion of PPAR� mice. These micehmrsfdWotP
8
iors(TcUcBndeGpsm
ABCA1) and induced apolipoprotein A1-specific cholesterolfflux [20]. Opposite findings for atherosclerosis have beeneported. Overexpression of PPAR� in vascular smooth mus-le cells (VSMCs) increased postconfluent cell proliferationnd PPAR� activation promoted lipid accumulation in humanacrophages by increasing the expression of the class A and B
cavenger receptors (SR-A and CD36) and adipophilin [27,28].t has been accepted that inflammation couples dyslipidemiao atheroma formation [29]. Welch et al. [30] proposed thathe effect of a high concentration of PPAR� agonist is due toPAR� activation, and established the overlapping transacti-ation and transrepression functions of PPAR� and PPAR� inacrophages. It seems that PPAR� activation may induce anti-
nflammatory effect on macrophages because PPAR� activationnhibits the subset of lipopolysaccharide (LPS) or interferonIFN)-� responsive genes.
Lee et al. [31] finely demonstrated that PPAR� depletionn macrophages decreases atherosclerosis in LDL receptor−/−
ice. In contrast to in vitro data, there are no differences inhe expression of SR-A, CD36 and ABCA1 between wild-typend PPAR� −/− macrophages in the absence or presence ofW501516. Furthermore they found that PPAR� controls the
nflammation status by its association (pro-inflammatory) andissociation (anti-inflammatory) from transcriptional repres-ors. Deletion of PPAR� in macrophages leads to an anti-nflammatory status [low levels of the monocyte chemoattractantrotein (MCP-1), interleukin-1� (IL-1�), and matrix metallo-roteinase (MMP-9)] and showed less atherosclerosis in LDLeceptor−/− mice, suggesting that deletion of PPAR� enhanceshe released of anti-inflammatory transcriptional repressors.verexpression of PPAR� (increased unliganded PPAR� recep-
ave cardiac dysfunction, progressive myocardial lipid accu-ulation, cardiac hypertrophy, and congestive heart failure with
educed survival. PPAR� agonist, GW0742 induced the tran-cript levels of fatty acid oxidation in cultured cardiomyocytesrom PPAR� +/+ mice but the effect is not observed in car-iomyocytes from PPAR� −/− mice. The PPAR� agonist,Y14643 restored the decreased transcript levels of fatty acid
xidation in cells from PPAR� −/− mice. Those data indicatehat PPAR� is involved in cardiac fatty acid oxidation as doesPAR�.
. Oncogenesis
PPAR� expression is increased in colorectal cancer, andts mRNA level is markedly elevated in primary human col-rectal adenocarcinomas [37,38]. However, the expression wasepressed through the adenomatous polyposis coli (APC) tumoruppressor gene and non-steroidal anti-inflammatory drugsNSAIDs) inhibit tumorigenesis by inhibition of PPAR� [38].he up-regulation of PPAR� expression activity has also beenonfirmed in K-Ras transformed intestinal epithelial cells [39].sing PPAR�-deficient APCmin mutant mice (which lack a
opy of the adenomatous polyposis coli tumor suppressor gene),arak et al. [12] reported that PPAR� has no influence on intesti-al multiplicity and Harman et al. [40] showed that PPAR�epletion induces colon carcinogenesis implying a protectiveffect of PPAR� against colon carcinogenesis. PPAR� agonist,W501516 accelerates small-intestine polyp growth in cancer-rone Min mice, yet PPAR� itself is genetically dispensable formall intestine and colon polyp formation in the Min mouseodel [41].

506 S. Takahashi et al. / Pharmacological Research 53 (2006) 501–507
9. Therapeutic target of PPAR� agonists and issues ofconcern
The therapeutic potential of PPAR� agonists for obesity, dys-lipidemia, and type 2 diabetes have been tested in mice. PPAR�polymorphisms were associated with high LDL-cholesterollevel in a Caucasian or obesity in a Korean population in humanepidemiological studies [42,43]. In this context, it will be excit-ing to determine whether PPAR� agonists are clinically ben-eficial for obesity and its associated problems such as type 2diabetes and hyperlipidemia. However, potential unwanted con-sequences of PPAR� activation need to be considered. Furtherwork to elucidate the roles of the PPARs in oncogenesis andorgan toxicity will be crucial in determining the safety of chronicPPAR� ligand treatment in human.
Acknowledgments
This work was supported through supported in part byresearch grants from the Ministry of Education, Science andCulture of Japan, Special Coordination Funds for PromotingScience and Technology from the Ministry of Education, Cul-ture, Sports, Science and Technology of the Japanese Govern-ment, Astellas Foundation for Research on Metabolic Disorders,Research Fund of Mitsukoshi Health and Welfare Foundation,and Exploratory Research for Advanced Technology/Japan Sci-e
R
[
[
[12] Barak Y, Liao D, He W, Ong ES, Nelson MC, Olefsky JM, et al.Effects of peroxisome proliferator-activated receptor � on placentation,adiposity, and colorectal cancer. Proc Natl Acad Sci USA 2002;99:303–8.
[13] Michalik L, Desvergne B, Tan NS, Basu-Modak S, Escher P, Rieus-set J, et al. Impaired skin wound healing in peroxisome proliferator-activated receptor (PPAR)� and PPAR� mutant mice. J Cell Biol2001;154:799–814.
[14] Bastie C, Holst D, Gaillard D, Jehl-Pietri C, Grimaldi PA. Expression ofperoxisome proliferator-activated receptor PPAR� promotes induction ofPPAR� and adipocyte differentiation in 3T3C2 fibroblast. J Biol Chem1999;274:21920–5.
[15] Bastie C, Luquet S, Holst D, Jehl-Pietri C, Grimaldi PA. Alter-ations of peroxisome proliferator-activated receptor � activity affectfatty acid-controlled adipose differentiation. J Biol Chem 2000;275:38768–73.
[16] Hansen JB, Zhang H, Rasmussen TH, Petersen RK, Flindt EN,Kristiansen K. Peroxisome proliferator-activated receptor � (PPAR�)-mediated regulation of preadipocyte proliferation and gene expressionis dependent on cAMP signaling. J Biol Chem 2001;276:3175–82.
[17] Wang Y-X, Lee C-H, Tiep S, Yu RT, Ham J, Kang H, et al. Peroxisome-proliferator-activated receptor � activates fat metabolism to prevent obe-sity. Cell 2003;113:159–70.
[18] Tanaka T, Yamamoto J, Iwasaki S, Asaba H, Hamura H, Ikeda Y, et al.Activation of peroxisome proliferator-activated receptor � induces fattyacid �-oxidation in skeletal muscle and attenuates metabolic syndrome.Proc Natl Acad Sci USA 2003;100:15924–9.
[19] Dressel U, Allen TL, Pippal JB, Rohde PR, Lau P, Muscat GEO.The peroxisome proliferators-activated receptor �/� agonist, GW501516,regulates the expression of genes involved in lipid catabolism andenergy uncoupling in skeletal muscle cells. Mol Endocrinol 2003;17:
[
[
[
[
[
[
[
[
[
[[
nce and Technology Agency.
eferences
[1] Eckel RH, Grundy SM, Zimmet PZ. The metabolic syndrome. Lancet2005;365:1415–28.
[2] Ford ES, Giles WH, Dietz WH. Prevalence of the metabolic syndromeamong US adults: findings from the third National Health and NutritionExamination Survey. JAMA 2002;287:356–9.
[3] Ninomiya JK, L’Italien G, Criqui MH, Whyte JL, Gamst A, ChenRS. Association of the metabolic syndrome with history of myocardialinfarction and stroke in the third national health and nutrition examina-tion survey. Circulation 2004;109:42–6.
[4] Evans RM, Barish GD, Wang Y-X. PPARs and the complex journey toobesity. Nat Med 2004;10:1–7.
[5] Leone TC, Weinheimer CJ, Kelly DP. A critical role for the peroxi-some proliferators activated receptor � (PPAR�) in the cellular fastingresponse: The PPAR�-null mouse as a model of fatty acid oxidationdisorders. Proc Natl Acad Sci USA 1999;96:7473–8.
[6] Kersten S, Seydoux J, Peters JM, Gonzalez FJ, Desvergne B, WahliW. Peroxisome proliferator-activated receptor � mediates the adaptiveresponse to fasting. J Clin Invest 1999;103:1489–98.
[7] Yki-Jarvinen H. Thiazolidinedions. N Engl J Med 2004;351:1106–18.[8] Schmidt A, Endo N, Rutledge SJ, Vogel R, Shinar D, Rodan GA.
Identification of a new member of the steroid hormone receptor super-family that is activated by a peroxisome proliferator and fatty acids. MolEndocrinol 1992;6:1634–41.
[9] Escher P, Braissant O, Basu-Modak S, Michalik L, Wahli W, DesvergneB. Rat PPARs: quantitative analysis in adult rat tissues and regulationin fasting and refeeding. Endocrinology 2001;142:4195–202.
10] Chawla A, Lee C-H, Barak Y, He W, Rosenfeld J, Liao D, et al. PPAR�
is a very low-density lipoprotein sensor in macrophages. Proc Natl AcadSci USA 2003;100:1268–73.
11] Peters JM, Lee SST, Li W, Ward JM, Gavrilova O, Everett C, et al.Growth, adipose, brain, and skin alterations resulting from targeted dis-ruption of the mouse peroxisome proliferator-activated receptor � (�).Mol Cell Biol 2000;20:5119–28.
2477–93.20] Oliver Jr WR, Shenk JL, Snaith MR, Russell CS, Plunket KD, Bod-
kin NL, et al. A selective peroxisome proliferator-activated receptor� promotes reverse cholesterol transport. Proc Natl Acad Sci USA2001;98:5306–11.
21] Holst D, Luquet S, Nogueira V, Kristiansen K, Leverve X, GrimaldiPA. Nutritional regulation and role of peroxisome proliferator-activatedreceptor � in fatty acid catabolism in skeletal muscle. Biochim BiophysActa 2003;1633:43–50.
22] Luquet S, Lopez-Soriano J, Holst D, Fredenrich A, Melki J, Ras-soulzadegan M, et al. Peroxisome proliferator-activated receptor �
controls muscle development and oxidative capability. FASEB J2003;17:2299–301.
23] Wang Y-X, Zhang C-L, Yu RT, Cho HK, Nelson MC, Bayuga-OcampoCR, et al. Regulation of muscle fiber type and running endurance byPPAR�. PLoS Biol 2004;2:e294.
24] Kramer DK, Al-Khalili L, Perrini S, Skogsberg J, Wretenberg P, Kan-nisto K, et al. Direct activation of glucose transport in primary humanmyotubes after activation of peroxisome proliferator-activated receptor�. Diabetes 2005;54:1157–63.
25] Leibowitz MD, Fievet C, Hennuyer N, Peinado-Onsurb J, Duez H,Berger J, et al. Activation of PPAR� alters lipid metabolism in db/dbmice. FEBS Lett 2000;473:333–6.
26] Akiyama TE, Lambert G, Nicol CJ, Matsusue K, Peters JM, Brewer JrHB, et al. Peroxisome proliferator-activated receptor �/� regulates verylow density lipoprotein production and catabolism in mice on a westerndiet. J Biol Chem 2004;279:20874–81.
27] Zhang J, Fu M, Zhu X, Xiao Y, Mou Y, Zheng H, et al. Peroxi-some proliferator-activated receptor � is up-regulated during vascularlesion formation and promotes post-confluent cell proliferation in vas-cular smooth muscle cells. J Biol Chem 2002;277:11505–12.
28] Vosper H, Patel L, Graham TL, Khoudoli GA, Hill A, Macphee CH, etal. The peroxisome proliferator-activated receptor � promotes lipid accu-mulation in human macrophages. J Biol Chem 2001;276(47):44258–65.
29] Libby P. Inflammation in atherosclerosis. Nature 2002;420:868–74.30] Welch JS, Ricote M, Akiyama TE, Gonzalez FJ, Glass CK. PPAR�
and PPAR� negatively regulate specific subsets of lipopolysaccharide

S. Takahashi et al. / Pharmacological Research 53 (2006) 501–507 507
and IFN-� target genes in macrophages. Proc Natl Acad Sci USA2003;100:6712–7.
[31] Lee C-H, Chawla A, Urbiztondo N, Liao D, Boisvert WA, Evans RM.Transcriptional repression of atherogenic inflammation: modulation byPPAR�. Science 2003;302:453–7.
[32] Li AC, Binder CJ, Gutierrez A, Brown KK, Plotkin CR, PattisonJW, et al. Differential inhibition of macrophage foam-cell forma-tion and atherosclerosis in mice by PPAR�, �/�, �. J Clin Invest2004;114:1564–76.
[33] Graham TL, Mookherjee C, Suckling KE, Palmer CNA, Patel L. ThePPAR� agonist GW0742X reduces atherosclerosis in LDLR −/− mice.Atherosclerosis 2005;181:29–37.
[34] Gilde AJ, van der Lee KAJM, Willemsen PHM, Chinetti G, van der LeijFR, van der Vusse GJ, et al. Peroxisome proliferator-activated receptor(PPAR)� and PPAR�/�, but not PPAR�, modulate the expression ofgenes involved in cardiac lipid metabolism. Circ Res 2003;92:518–24.
[35] Cheng L, Ding G, Qin Q, Xiao Y, Woods D, Chen YE, et al. Per-oxisome proliferators-activated receptor � activates fatty acid oxidationin cultured neonatal and adult cardiomyocytes. Biochem Biophys ResCommun 2004;313:277–86.
[36] Cheng L, Ding G, Qin Q, Huang Y, Lewis W, He N, et al.Cardiomyocyte-restricted peroxisome proliferator-activated receptor-�deletion perturbs myocardial fatty acid oxidation and leads to cardiomy-opathy. Nat Med 2004;10:1245–50.
[37] He T-C, Chan TA, Vogelstein B, Kinzler KW. PPAR� is an APC-regulated target of nonsteroidal anti-inflammatory drugs. Cell 1999;99:335–45.
[38] Gupta RA, Tan J, Krause WF, Geraci MW, Willson TM, DeySK, et al. Prostacyclin-mediated activation of peroxisome proliferator-activated receptor � in colorectal cancer. Proc Natl Acad Sci USA2000;97:13275–80.
[39] Shao J, Sheng H, DuBois RN. Peroxisome proliferator-activated recep-tors modulate K-Ras-mediated transformation of intestinal epithelialcells. Cancer Res 2002;62:3282–8.
[40] Harman FS, Nicol CJ, Marin HE, Ward JM, Gonzalez FJ, Peters JM.Peroxisome proliferator-activated receptor-� attenuates colon carcinogen-esis. Nat Med 2004;10:481–3.
[41] Gupta RA, Wang D, Katkuri S, Wang H, Dey SK, DuBois RN.Activation of nuclear hormone receptor peroxisome proliferator-activated receptor-� accelerates intestinal adenoma growth. Nat Med2004;10:245–7.
[42] Skogsberg J, Kannisto K, Cassel TN, Hamsten A, Eriksson P, EhrenborgE. Evidence that peroxisome proliferator-activated receptor delta influ-ences cholesterol metabolism in men. Arterioscler Thromb Vasc Biol2003;23:637–43.
[43] Shin HD, Park BL, Kim LH, Jung HS, Cho YM, Moon MK, et al.Genetic polymorphisms in peroxisome proliferator-activated receptor �
associated with obesity. Diabetes 2004;53:847–51.
Related Documents











