Peripheral Glucagon-like Peptide-1 (GLP-1) and Satiation Mukesh Punjabi, Myrtha Arnold, Nori Geary, Wolfgang Langhans, Gustavo Pacheco-López ⁎ Physiology and Behaviour Laboratory, Institute of Food, Nutrition and Health, ETH Zurich, 8603 Schwerzenbach, Switzerland abstract article info Article history: Received 21 February 2011 Accepted 23 February 2011 Available online xxxx Keywords: Exendin-4 Food intake Eating Vagus Hindbrain Hepatic portal vein Incretin Meal size Peripheral GLP-1 is produced by post-translational processing of pro-glucagon in enteroendocrine L-cells and is released in response to luminal nutrient (primarily carbohydrate and fat) stimulation. GLP-1 is well known for its potent insulinotropic and gluco-regulatory effects. GLP-1 receptors (GLP-1R) are expressed in the periphery and in several brain areas that are implicated in the control of eating. Both central and peripheral administration of GLP-1 have been shown to reduce food intake. Unresolved, however, is whether these effects reflect functions of endogenous GLP-1. Data collected in our laboratory indicate that in chow-fed rats: 1) Remotely controlled, intra-meal intravenous (IV) or intraperitoneal (IP) GLP-1 infusions selectively reduce meal size; 2) hindbrain GLP-1R activation is involved in the eating-inhibitory effect of IV infused GLP-1, whereas intact abdominal vagal afferents are necessary for the eating-inhibitory effect of IP, but not IV, infused GLP-1; 3) GLP-1 degradation in the liver prevents a systemic increase in endogenous GLP-1 during normal chow meals in rats; and 4) peripheral or hindbrain GLP-1R antagonism by exendin-9 does not affect spontaneous eating. Also, although our data indicate that peripheral GLP-1 can act in two different sites to inhibit eating, they argue against a role of systemic increases in endogenous GLP-1 in satiation in chow-fed rats. Therefore, further studies should examine whether a local paracrine action of GLP-1 in the intestine or and endocrine action in the hepatic-portal area is physiologically relevant for satiation. © 2011 Elsevier Inc. All rights reserved. 1. Introduction Glucagon-like-peptide (7–36) amide (GLP-1) is the product of post- translational processing of pro-glucagon in the gut and the brain [1]. In the brain, GLP-1 is expressed in some neurons of the nucleus tractus solittarii (NTS) [2,3]. In the gut, GLP-1 is released primarily from enteroendocrine L-cells located in the distal jejunum and ileum [1]. Luminal carbohydrates and fats are potent stimuli for GLP-1 secretion, and these effects appear to be mediated in part by taste receptors expressed on enteroendocrine cells [4] (Fig. 1). Recently GLP-1 has also been identified in sweet- and umami-taste receptor cells in the oral cavity [5], suggesting that GLP-1 signaling is involved in taste. GLP-1 receptors (GLP-1R) are widely distributed in the brain and in peripheral organs such as the pancreatic islets and the whole gastrointestinal (GI) tract [6,7]. In the brain, GLP-1R are found in several hindbrain and forebrain areas [8], including areas that are implicated in the control of food intake and energy balance, such as the area postrema (AP), NTS, hypothalamus and amygdala [3]. GLP-1 has several physiological effects: it acts as a strong incretin, i.e., stimulates glucose-induced insulin release [9], it inhibits glucagon release [1], and it inhibits gastric emptying, i.e., contributes to the ileal brake mechanism [10]. Further potential physiological effects of GLP-1 include an influence on learning and memory [11], neuroprotection [12,13], and the inhibition of eating and drinking [14,15]. Administration of GLP-1 or of potent, long-acting natural or synthetic GLP-1 analogs, such as exendin-4 (Ex-4) or liraglutide, inhibits eating in many species, including humans [16–22]. Ex-4 is a naturally occurring peptide resistant to rapid enzymatic degradation by dipeptidyl peptidase 4 (DPP-4) [23]. The synthetic GLP-1 analog liraglutide also resists enzymatic degradation and, because it binds to albumin in the plasma, delays renal excretion [23]. Chronic administrations of GLP-1 or its analogs have effectively reduced weight and improved glucose metabolism in overweight individuals [18,24], suggesting GLP-1 may offer an effective therapeutic option for overweight and type II diabetes [25]. Endogenous intestinal GLP-1 has been implicated in meal termination (satiation) because: 1) exogenous GLP-1 inhibited eating primarily by reducing meal size [26,27] and 2) intraperitoneal (IP) injection of the specific GLP-1R antagonist exendin (9–39)(Ex-9) stimulated eating under some conditions [26], indicating that satiation can be delayed when endogenous GLP-1 signaling is blocked. In other studies, however, Ex-9 administration failed to stimulate eating, suggesting that endogenous GLP-1 is not required for the control of meal size under all conditions. Thus, further research is warranted to identify the situations under which endogenous GLP-1 is physiologically relevant for satiation. Furthermore, the site and mechanism of GLP-1's eating-inhibitory action, including the intra- cellular signaling cascades involved, are still largely unknown. Physiology & Behavior xxx (2011) xxx–xxx ⁎ Corresponding author at: Physiology and Behaviour Laboratory, ETH Zurich, Schorenstr. 16, 8603 Schwerzenbach, Switzerland. Tel.: + 41 44 655 7420; fax: +41 44 655 7206. E-mail address: [email protected] (G. Pacheco-López). PHB-09381; No of Pages 6 0031-9384/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.physbeh.2011.02.038 Contents lists available at ScienceDirect Physiology & Behavior journal homepage: www.elsevier.com/locate/phb Please cite this article as: Punjabi M, et al, Peripheral Glucagon-like Peptide-1 (GLP-1) and Satiation, Physiol Behav (2011), doi:10.1016/j. physbeh.2011.02.038

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Physiology & Behavior xxx (2011) xxx–xxx
PHB-09381; No of Pages 6
Contents lists available at ScienceDirect
Physiology & Behavior
j ourna l homepage: www.e lsev ie r.com/ locate /phb
Peripheral Glucagon-like Peptide-1 (GLP-1) and Satiation
Mukesh Punjabi, Myrtha Arnold, Nori Geary, Wolfgang Langhans, Gustavo Pacheco-López ⁎Physiology and Behaviour Laboratory, Institute of Food, Nutrition and Health, ETH Zurich, 8603 Schwerzenbach, Switzerland
⁎ Corresponding author at: PhysiologyandBehaviour Lab16, 8603 Schwerzenbach, Switzerland. Tel.: +41 44 655 7
E-mail address: [email protected] (G. Pache
0031-9384/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.physbeh.2011.02.038
Please cite this article as: Punjabi M, et al,physbeh.2011.02.038
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 February 2011Accepted 23 February 2011Available online xxxx
Keywords:Exendin-4Food intakeEatingVagusHindbrainHepatic portal veinIncretinMeal size
Peripheral GLP-1 is produced by post-translational processing of pro-glucagon in enteroendocrine L-cellsand is released in response to luminal nutrient (primarily carbohydrate and fat) stimulation. GLP-1 is wellknown for its potent insulinotropic and gluco-regulatory effects. GLP-1 receptors (GLP-1R) are expressed inthe periphery and in several brain areas that are implicated in the control of eating. Both central andperipheral administration of GLP-1 have been shown to reduce food intake. Unresolved, however, iswhether these effects reflect functions of endogenous GLP-1. Data collected in our laboratory indicate thatin chow-fed rats: 1) Remotely controlled, intra-meal intravenous (IV) or intraperitoneal (IP) GLP-1infusions selectively reduce meal size; 2) hindbrain GLP-1R activation is involved in the eating-inhibitoryeffect of IV infused GLP-1, whereas intact abdominal vagal afferents are necessary for the eating-inhibitoryeffect of IP, but not IV, infused GLP-1; 3) GLP-1 degradation in the liver prevents a systemic increase inendogenous GLP-1 during normal chowmeals in rats; and 4) peripheral or hindbrain GLP-1R antagonism byexendin-9 does not affect spontaneous eating. Also, although our data indicate that peripheral GLP-1 can actin two different sites to inhibit eating, they argue against a role of systemic increases in endogenous GLP-1in satiation in chow-fed rats. Therefore, further studies should examine whether a local paracrine action ofGLP-1 in the intestine or and endocrine action in the hepatic-portal area is physiologically relevant forsatiation.
oratory, ETHZurich, Schorenstr.420; fax: +41 44 655 7206.co-López).
l rights reserved.
Peripheral Glucagon-like Peptide-1 (GLP-1)
© 2011 Elsevier Inc. All rights reserved.
1. Introduction
Glucagon-like-peptide (7–36) amide (GLP-1) is the product of post-translational processing of pro-glucagon in the gut and the brain [1]. Inthe brain, GLP-1 is expressed in some neurons of the nucleus tractussolittarii (NTS) [2,3]. In the gut, GLP-1 is released primarily fromenteroendocrine L-cells located in the distal jejunum and ileum [1].Luminal carbohydrates and fats are potent stimuli for GLP-1 secretion,and these effects appear to be mediated in part by taste receptorsexpressed on enteroendocrine cells [4] (Fig. 1). Recently GLP-1 has alsobeen identified in sweet- and umami-taste receptor cells in the oralcavity [5], suggesting that GLP-1 signaling is involved in taste. GLP-1receptors (GLP-1R) arewidely distributed in the brain and in peripheralorgans such as the pancreatic islets and the whole gastrointestinal (GI)tract [6,7]. In the brain, GLP-1R are found in several hindbrain andforebrain areas [8], including areas that are implicated in the control offood intake and energy balance, such as the area postrema (AP), NTS,hypothalamus and amygdala [3]. GLP-1 has several physiologicaleffects: it acts as a strong incretin, i.e., stimulates glucose-inducedinsulin release [9], it inhibits glucagon release [1], and it inhibits gastricemptying, i.e., contributes to the ileal brake mechanism [10]. Further
potential physiological effects of GLP-1 include an influence on learningand memory [11], neuroprotection [12,13], and the inhibition of eatingand drinking [14,15]. Administration of GLP-1 or of potent, long-actingnatural or synthetic GLP-1 analogs, such as exendin-4 (Ex-4) orliraglutide, inhibits eating in many species, including humans [16–22].Ex-4 is a naturally occurring peptide resistant to rapid enzymaticdegradationbydipeptidyl peptidase 4 (DPP-4) [23]. The syntheticGLP-1analog liraglutide also resists enzymatic degradation and, because itbinds to albumin in the plasma, delays renal excretion [23]. Chronicadministrations of GLP-1 or its analogs have effectively reduced weightand improved glucose metabolism in overweight individuals [18,24],suggesting GLP-1 may offer an effective therapeutic option foroverweight and type II diabetes [25].
Endogenous intestinal GLP-1 has been implicated in mealtermination (satiation) because: 1) exogenous GLP-1 inhibited eatingprimarily by reducing meal size [26,27] and 2) intraperitoneal (IP)injection of the specific GLP-1R antagonist exendin (9–39)(Ex-9)stimulated eating under some conditions [26], indicating thatsatiation can be delayedwhen endogenous GLP-1 signaling is blocked.In other studies, however, Ex-9 administration failed to stimulateeating, suggesting that endogenous GLP-1 is not required for thecontrol of meal size under all conditions. Thus, further research iswarranted to identify the situations under which endogenous GLP-1 isphysiologically relevant for satiation. Furthermore, the site andmechanism of GLP-1's eating-inhibitory action, including the intra-cellular signaling cascades involved, are still largely unknown.
and Satiation, Physiol Behav (2011), doi:10.1016/j.

2 M. Punjabi et al. / Physiology & Behavior xxx (2011) xxx–xxx
After secretion, GLP-1 is rapidly degraded into GLP-1 (9–36) byDPP-IV [28]. As a result, the biological half-life of GLP-1 is less than5 min [29]. If prandial secretion is maintained and exceeds degrada-tion, endogenous GLP-1 may still have an endocrine satiating effect bydirect central nervous system action. In addition, such a route ofaction may be particularly relevant for pharmacological interventionsthat lead to sustained, and substantial increases in circulating levels ofnative GLP-1 or GLP-1R agonists. It is also relevant to mention thatparacrine effects of GLP-1 in rats may be endocrine effects in humans,as is the case for the satiating effect of cholecystokinin (CCK). Finally,gut hormones can also signal the brain through an endocrine orparacrine effect on afferent nerves, with the vagus being theprominent neural route of gut-brain signaling [30,31]. Whether ornot endogenous peripheral GLP-1 induces satiation through such anendocrine or paracrine action is still unresolved. This review brieflysummarizes what is known about these open questions, capitalizingon some experiments that we recently did to address them.
2. Endocrine effects of GLP-1
The insulinotropic effect of GLP-1 is mediated in part by GLP-1R onhepatic branch vagal afferents terminating in the wall of the hepaticportal vein close to the liver [32]. Activation of these receptors triggersa vago-vagal reflex that increases pancreatic vagal-efferent activityand stimulates insulin release [33]. Therefore, it appeared logical totest whether a similar mechanism is involved in the eating-inhibitoryeffect of peripheral GLP-1. In an attempt to mimic the meal-inducedrelease of endogenous GLP-1 from the small intestinal L-cells into thehepatic portal vein as closely as possible, we equipped rats withchronic hepatic portal vein catheters and infused various doses ofGLP-1 during the first spontaneous nocturnal meal. The animals werecontinuously monitored via infra-red video cameras, and theinfusions were triggered by remote control from an adjoining roomto prevent disturbing them. Under these conditions, hepatic portalvein infusions of GLP-1 reduced the size and duration of the ongoingmeal but did not affect the subsequent inter-meal interval or thesize or duration of the second meal [27]. Furthermore, intra-mealhepatic portal vein and vena cava (i.e., systemic) infusions of GLP-1(1 nmol/kg body weight [BW]) produced similar effects on eating,suggesting that the satiating effect of circulating GLP-1 is notmediated by GLP-1R located in the hepatic portal system or liver.Interestingly, although hepatic portal vein infusion of Ex-4 under thesame conditions reduced the sizes of both the first and the secondmeals, it did not affect the duration of the intervening inter-mealinterval, suggesting that GLP-1R activation has a selective effect onsatiation. Whether the briefer and smaller increases in plasma GLP-1levels produced by GLP-1 infusions and the more prolonged andgreater increases in GLP-1R activation produced by Ex-4 reduce foodintake by acting at the same site(s) or whether Ex-4 recruitsadditional receptors for its extended effect remains to be investigated.In addition, while all these findings are consistent with a possiblephysiological endocrine satiating effect of endogenous GLP-1, our datato date do not support a role of GLP-1 in the graded satiating effect offood because we have not found a clearly dose-related satiating effectof exogenous GLP-1 [27].
We also examinedwhether the eating-inhibitory effect of circulatingGLP-1 is mediated by abdominal vagal afferents. Rats underwent sub-diaphragmatic vagal deafferentation (SDA) [34] or sham surgery andwere equipped with hepatic portal vein catheters. SDA produces themost specific and complete lesionof abdominal vagal afferents. That is, itlesions both vagal A- and C-fibers, whereas capsaicin lesions onlyunmyelinatedvagalC-fibers and lesionsnon-vagal aswell as vagalfibers[35]. In addition, SDA leaves about 50% of the vagal efferents intact[34,36], which reduces the adverse side effects related to impaired GImotility and secretion that are produced by complete subdiaphragmaticvagotomy. Intra-meal hepatic portal vein infusions of GLP-1 reduced
Please cite this article as: Punjabi M, et al, Peripheral Glucagon-like Pephysbeh.2011.02.038
meal size and duration similarly in SDA and sham-operated rats [27]. Inaddition, Ex-4 reduced first (Fig. 2) and second meal size (data notshown)without a significant effect on othermeal parameters. Ex-4 alsoproduced similar 2 and 4 h decreases in cumulative food intake in SDAand sham-operated rats (Fig. 3). Overall, these data indicate thatabdominal vagal afferents are not necessary for the eating-inhibitoryeffect of circulating GLP-1 or Ex-4.
In order to identify the neural substrates that may be involved inmediating the satiating effect of circulating GLP-1, we assessed c-Fosexpression in some hindbrain (area postrema [AP], NTS) and forebrainstructures (central nucleus of the amygdala [CeA], hypothalamicparaventricular and arcuate nuclei [PVN and Arc]) implicated in thecontrol of eating. We found that hepatic portal vein GLP-1 infusion, at adose (1 nmol/kg BW) that reliably inhibited eating, activated the NTS,AP, and CeA, suggesting that these brain areas participate in theendocrine eating-inhibitory effect of GLP-1 [37]. One previous studyusing femoral vein infusion of a lower dose of native GLP-1(1 μg=0.24 nmol/kg BW) failed to detect a significant increase in c-Fos expression in the brainstem [38], whereas another study usingfemoral vein infusions of Ex-4 [39] found awider-spread activation thanwe did. Because of themuch longer biological half-life of Ex-4, however,these results cannot directly be compared to ours.
In addition to causing satiation, GLP-1 may inhibit eating non-specifically by inducing malaise. GLP-1 can trigger a conditioned tasteaversion and pica (kaolin intake) in rodents [40,41]. On the otherhand, behavioral observations in macaques did not reveal any overtsigns of malaise, such as decreased alertness, drooling, or vomiting,in response to GLP-1 [16]. Under which conditions GLP-1 producessatiation or malaise remains unclear. GLP-1R in the CeA appear tomediate some of the response to peripheral illness [42]. This isbecause 1) intra-amygdala administration of GLP-1, but not of theinactive analog, GLP-1 (9–39), produced a strong conditioned tasteaversion, similar to those produced by visceral malaise or foodpoisoning, and 2) intra-amygdala administration of Ex-9 preventedtaste aversion learning in response to IP injections of the toxin lithiumchloride [42]. Importantly, intra-amygdala injection of GLP-1 did notitself reduce food intake, suggesting that this mechanism is not anecessary component of the eating-inhibitory effect of endocrineGLP-1. Presumably, GLP-1 projections from the NTS are involved inthis receptor activation. At the same time, however, our c-Fos data areconsistent with the possibility that the CeA may also be involved inmediating the eating-inhibitory effect of circulating GLP-1. Whetheror not this increase in CeA c-Fos occurred in GLP-1R-expressingneurons or was related to an aversive component of GLP-1's eating-inhibitory effect under our conditions requires further research.
We recently found that infusion of Ex-9 (10 μg/rat) into the 4thcerebral ventricle blocked the eating-inhibitory effect of hepatic portalvein GLP-1 infusion in rats, indicating that GLP-1R that are accessiblefrom the 4th ventricle (e.g. NTS and AP) are involved in mediating thiseffect [43]. Also, preliminary findings of ours indicate that AP lesionsblock the eating-inhibitory effect of hepatic portal vein GLP-1infusions, suggesting that the AP is involved as well (Punjabi et al., inpreparation). IP injection of (2.0 μg/kg BW)Ex-4was recently reportedto reduce food intake in AP-lesioned rats after IP injection [44], whichat first glance seems to contradict the idea that the AP mediates theeating-inhibitory effect of circulating GLP-1. It is possible, however,that the greater and more prolonged GLP-1R activation produced byEx-4 may have recruited redundant neural circuits to inhibit eating.It may be relevant in this connection that it was not determinedwhether Ex-4 elicited a conditioned taste aversion after AP lesions.
3. GLP-1R mechanisms
GLP-1R are G-protein-coupled and activate diverse intracellularsignaling pathways involving cyclic adenosine monophosphate,protein kinase A (PKA), phospholipase C, phosphatidylinositol-3
ptide-1 (GLP-1) and Satiation, Physiol Behav (2011), doi:10.1016/j.

3M. Punjabi et al. / Physiology & Behavior xxx (2011) xxx–xxx
kinase, protein kinase C, p44/42 mitogen-activated protein kinase(p44/42 MAPK; also known as extracellular signal-regulated kinase[ERK]-1/2), and intracellular Ca2+ [45,46]. The neuronal intracellularsignaling pathways mediating the eating-inhibitory effect of GLP-1are still poorly characterized. We recently started to address thisquestion using strategies based on the mediation of the satiatingeffect of CCK and on the intracellular signaling pathway activated byGLP-1 in pancreatic β-cells and in neurons. CCK increases activity ofthe ERK signaling cascade in NTS neurons, and inhibiting thispathwaywith an ERK kinase (MEK) inhibitor significantly attenuatedthe eating-inhibitory effect of IP CCK in rats [47,48]. GLP-1Ractivation also leads to ERK phosphorylation through a MEK-dependent pathway in pancreatic β-cells [49]. Furthermore, ERKphosphorylation was significantly increased in the hippocampus20 min after intranasal administration of the short GLP-1 analog, [Ser(2)] exendin(1–9), and the enhancement of associative learning bythis GLP-1 analogwas completely blocked by intracerebroventricularinfusion of aMEK inhibitor [11]. In linewith this, it has been proposedthat hindbrain GLP-1R activation leads to an increase in ERK activity[14]. Preliminary data from our laboratory in fact suggest that hepaticportal vein GLP-1 infusion increases phosphorylation of ERK 2, butnot ERK 1, in the NTS (Punjabi et al., unpublished). Whetheractivation of the ERK 1/2 signaling pathway is involved in thesatiating effect of circulating GLP-1, however, remains to beinvestigated.
4. Prandial increases in circulating GLP-1 levels
Increases in circulating endogenous GLP-1 levels in laboratoryanimals have been reported only in response to intragastric [50,51]or intraintestinal [52] administration of liquid diets or nutrientsolutions in anesthetized animals, i.e., in situations that do notnecessarily mimic ingestion of a solid, mixed-nutrient meal inundisturbed, awake animals. To determine whether meals doincrease circulating GLP-1 levels, we equipped rats with hepaticportal vein and vena cava catheters, took blood samples in parallelfrom both blood vessels during and after spontaneous nocturnalchowmeals (fixed in size to 3 g), and assayed active GLP-1.We foundthat GLP-1 increased during the meal in the hepatic portal vein, butnot in the vena cava [53]. Hepatic portal vein plasma concentration ofGLP-1 increased rapidly, reached a peak (∼17 pM) 6 min after mealonset, and returned to baseline values (5–7 pM) by 20 min aftermealonset. Vena cava GLP-1 levels did not increase at any time. When1 nmol/kg BW GLP-1 was infused into the hepatic portal vein, activeGLP-1 in the vena cava increased to∼360 pM immediately after theend of infusion and returned to baseline levels 20 min later. Theprandial increase in endogenous GLP-1 that we observed was muchsmaller than in previous tests of intragastric or intraintestinalnutrient administration [50,51]. We presume that this discrepancyis due to the different test conditions for the reasons described above.Also, we measured active GLP-1 [i.e. GLP-1 (7–36 amide) and GLP-1(7–37)], whereas D'Alessio et al. [50], for instance, determined totalGLP-1 levels using an antiserum that reacts with an epitope includingthe COOH-terminal amide and therefore also recognizes the inactiveprecursor GLP-1 (1–36) and metabolite GLP-1 (9–36). Our failure tofind a significant prandial increase of active GLP-1 in the vena cava isconsistent with other reports. For instance, a 5 ml meal of Ensure(Abbott Laboratories) failed to increase systemic active GLP-1 levelsin rats [54], and intragastric intubation of glucose and corn oil causeda much smaller increase in GLP-1 concentration in the inferior venacava than in the hepatic portal vein [51]. Meal-related changes inGLP-1 have been reported in humans (e.g. [55,56]), but these studiesdid not include GLP-1 measurements in the HPV. Some of thesestudies also measured total instead of active GLP-1. Although totalGLP-1 is a reliable index of secreted hormone, it is not a useful
Please cite this article as: Punjabi M, et al, Peripheral Glucagon-like Pephysbeh.2011.02.038
measure of the amount of active hormone present because GLP-1degradation rates exceed clearance rates [57].
All in all, our findings suggest that during a nocturnal chow mealin rats, GLP-1 released from intestinal L-cells may have local,paracrine effects or endocrine effects in the hepatic portal vein orliver, but not systemic endocrine effects (Fig. 1). Furthermore,1 nmol/kg BW GLP-1 is only about twice the threshold dose for asatiating effect under these conditions [27], but increased systemicGLP-1 levels almost 25-fold the prandial physiological level in thehepatic portal vein. Our findings therefore also suggest that theinhibition of eating after intravenous infusion of exogenous GLP-1under these conditions is a pharmacological effect. Thus, our dataargue against a role of systemic increases in endogenous GLP-1 insatiation, at least under our conditions.
5. Paracrine effects of GLP-1
Because meals do not appear to increase systemic GLP-1 levels, wehave also tested the effects of IP infusionsof GLP-1 on eating. IP infusionsmay be better suited than hepatic portal vein infusions to target thevagal afferents terminating in thewall of the small intestine and, hence,address the possible role of a paracrine effect of endogenous GLP-1 insatiation. Remotely controlled, intra-meal IP GLP-1 infusions weretested under the same conditions as described above for hepatic portalvein infusions [58]. Similar to its effect after hepatic portal vein infusion,IP GLP-1 selectively reduced meal size and duration, although thethreshold dose (10 nmol/kg BW) for a reliable reduction of meal sizeand duration was about ten times higher than for hepatic portal veininfusion [58]. In contrast to its failure to block the eating-inhibitoryeffect of GLP-1 after hepatic portal vein infusion, however, SDA blockedthe satiating effect of IP infusion of 10 nmol/kg BW GLP-1 [58]. Thus,vagal afferent signaling is necessary for the full eating-inhibitory effectof IP, but not hepatic portal vein, GLP-1 [53,58]. Whether or not theeating-inhibitory effect of IP Ex-4 also requires intact abdominal vagalafferents remains to be examined. The acute hyperglycemia induced byIP Ex-4 administrationwas recently reported to be independent of vagalafferent signaling [59], suggesting that IP administered Ex-4 can reachnon-vagal GLP-1R.
The interstitial fluid in the lamina propria of the intestinal mucosaenters the lymphatic ducts by bulk flow, so that the intestinal lymphreflects the composition of the interstitial fluid in the lamina propriaof the mucosa fairly well [60,61]. Vagal afferents also terminate in thelamina propria (Fig. 1)[62,63]. Furthermore, IP injected peptides alsoreach the lamina propria and, thus, the intestinal lymph [50,64,65]. Inaddition, intragastric nutrient infusions increase the GLP-1 concen-tration in intestinal lymph (i.e. chyle) more than in the hepatic portalblood [50]. Finally, GLP-1 has been shown to activate gastric vagalafferents in whole nerve recordings [66], suggesting a similar effect onintestinal vagal afferents. These facts together with our SDA findingsdescribed above are consistent with the idea that IP GLP-1 may act onintestinal vagal afferents to inhibit eating, i.e., that IP GLP-1administration mimics a paracrine satiating effect of endogenousGLP-1 during meals.
To more directly examine the possible physiological relevance ofendogenous intestinal GLP-1, we IP infused Ex-9 [58]. Although Ex-9clearly antagonizes the incretin effect of endogenous GLP-1 inhumans [67] and rats [68,69], currently available reports on theeffects of peripheral Ex-9 on eating are inconsistent. IP Ex-9increased food intake only under some of the conditions tested(i.e. a moderate dose [30 μg/kg=8.9 nmol/kg BW] given in themiddle of the light phase or a higher dose [100 μg/kg=29.7 nmol/kgBW] given at 1 h into the dark phase [26]). Furthermore, othersfound that hepatic portal vein or jugular vein infusion of anotherGLP-1 receptor antagonist (desHis1,Glu8exendin-4) did not increasefood intake in rats [70]. Under our conditions, intra-meal infusion of30 nmol/kg BW Ex-9 did not increase the size of the first or second
ptide-1 (GLP-1) and Satiation, Physiol Behav (2011), doi:10.1016/j.
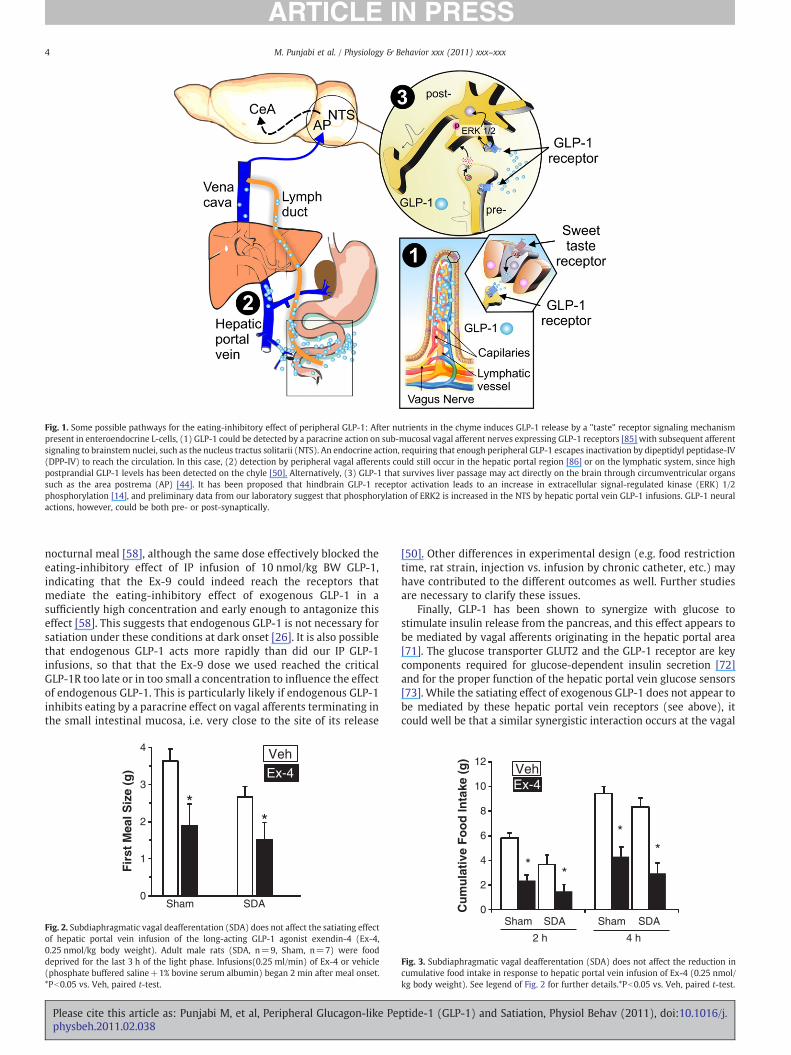
Fig. 1. Some possible pathways for the eating-inhibitory effect of peripheral GLP-1: After nutrients in the chyme induces GLP-1 release by a "taste" receptor signaling mechanismpresent in enteroendocrine L-cells, (1) GLP-1 could be detected by a paracrine action on sub-mucosal vagal afferent nerves expressing GLP-1 receptors [85] with subsequent afferentsignaling to brainstem nuclei, such as the nucleus tractus solitarii (NTS). An endocrine action, requiring that enough peripheral GLP-1 escapes inactivation by dipeptidyl peptidase-IV(DPP-IV) to reach the circulation. In this case, (2) detection by peripheral vagal afferents could still occur in the hepatic portal region [86] or on the lymphatic system, since highpostprandial GLP-1 levels has been detected on the chyle [50]. Alternatively, (3) GLP-1 that survives liver passage may act directly on the brain through circumventricular organssuch as the area postrema (AP) [44]. It has been proposed that hindbrain GLP-1 receptor activation leads to an increase in extracellular signal-regulated kinase (ERK) 1/2phosphorylation [14], and preliminary data from our laboratory suggest that phosphorylation of ERK2 is increased in the NTS by hepatic portal vein GLP-1 infusions. GLP-1 neuralactions, however, could be both pre- or post-synaptically.
4 M. Punjabi et al. / Physiology & Behavior xxx (2011) xxx–xxx
nocturnal meal [58], although the same dose effectively blocked theeating-inhibitory effect of IP infusion of 10 nmol/kg BW GLP-1,indicating that the Ex-9 could indeed reach the receptors thatmediate the eating-inhibitory effect of exogenous GLP-1 in asufficiently high concentration and early enough to antagonize thiseffect [58]. This suggests that endogenous GLP-1 is not necessary forsatiation under these conditions at dark onset [26]. It is also possiblethat endogenous GLP-1 acts more rapidly than did our IP GLP-1infusions, so that that the Ex-9 dose we used reached the criticalGLP-1R too late or in too small a concentration to influence the effectof endogenous GLP-1. This is particularly likely if endogenous GLP-1inhibits eating by a paracrine effect on vagal afferents terminating inthe small intestinal mucosa, i.e. very close to the site of its release
0
1
2
3
4
Fir
st M
eal S
ize
(g) Ex-4
Veh
Sham SDA
**
Fig. 2. Subdiaphragmatic vagal deafferentation (SDA) does not affect the satiating effectof hepatic portal vein infusion of the long-acting GLP-1 agonist exendin-4 (Ex-4,0.25 nmol/kg body weight). Adult male rats (SDA, n=9, Sham, n=7) were fooddeprived for the last 3 h of the light phase. Infusions(0.25 ml/min) of Ex-4 or vehicle(phosphate buffered saline+1% bovine serum albumin) began 2 min after meal onset.*Pb0.05 vs. Veh, paired t-test.
Please cite this article as: Punjabi M, et al, Peripheral Glucagon-like Pephysbeh.2011.02.038
[50]. Other differences in experimental design (e.g. food restrictiontime, rat strain, injection vs. infusion by chronic catheter, etc.) mayhave contributed to the different outcomes as well. Further studiesare necessary to clarify these issues.
Finally, GLP-1 has been shown to synergize with glucose tostimulate insulin release from the pancreas, and this effect appears tobe mediated by vagal afferents originating in the hepatic portal area[71]. The glucose transporter GLUT2 and the GLP-1 receptor are keycomponents required for glucose-dependent insulin secretion [72]and for the proper function of the hepatic portal vein glucose sensors[73]. While the satiating effect of exogenous GLP-1 does not appear tobe mediated by these hepatic portal vein receptors (see above), itcould well be that a similar synergistic interaction occurs at the vagal
0
2
4
6
8
10
12
2 h 4 h
Cu
mu
lati
ve F
oo
d In
take
(g
)
Sham
**
**
Ex-4Veh
SDA Sham SDA
Fig. 3. Subdiaphragmatic vagal deafferentation (SDA) does not affect the reduction incumulative food intake in response to hepatic portal vein infusion of Ex-4 (0.25 nmol/kg body weight). See legend of Fig. 2 for further details.*Pb0.05 vs. Veh, paired t-test.
ptide-1 (GLP-1) and Satiation, Physiol Behav (2011), doi:10.1016/j.

5M. Punjabi et al. / Physiology & Behavior xxx (2011) xxx–xxx
afferents GLP-1R in the intestinal wall. We therefore started toexamine whether IP GLP-1 may also interact with glucose to inhibiteating. Preliminary data from these experiments suggest that sub-threshold doses of GLP-1 and glucose can combine to reduce mealsize after intra-meal, IP infusion (Rüttimann et al., unpublished). Thisraises the possibility that GLP-1 may generally require glucose toactivate vagal afferents, which would extend the concept of the gluco-incretin functional unit model [72–74].
6. Perspectives
Spurred by the limited effectiveness of the few currently availabletreatment options for obesity and by the multiple side effects andproblems frequently encountered with drugs acting directly in thebrain, basic and clinical obesity researchers have begun to shift theirattention from the brain to the GI tract and in particular to thepotential of GI peptides in the control of energy homeostasis. GIpeptides link intestinal nutrient sensing to systemic effects mediatedby neuroendocrine or neural reflexes, and many GI peptides havebeen found to reduce food intake. As mentioned above, GLP-1 and itsanalogs are particularly promising for the pharmacotherapy of obesityand type II diabetes because GLP-1 is still effective in obese individuals[16–22,75] and has potent beneficial effects on metabolism [1]. Also,recent evidence from intestinal nutrient infusion studies in laboratoryanimal models as well as from pharmacological antagonist studies inhumans indicate that the dramatic obesity-curbing effect of bariatricsurgery is at least in part related to the enhanced release of GLP-1 andother, primarily distal small intestinal, eating-inhibitory GI peptides[54,76]. Moreover, previous concerns about tolerance to the eating-inhibitory effects of GI peptides with chronic administration inanimal models and, hence, about their therapeutic potential, may beunwarranted. Thus, CCK administered just prior to each meal in obeseZucker and dietary-induced obese rats adapted to three scheduledmeals/day (a feeding paradigm that is presumably more relevant forhuman eating than continuous ad libitum access to food) persistentlyreduced meal size and body weight over several weeks of treatment[77,78]. In addition, chronic administration of GLP-1 analogs resultedin significant weight loss in obese humans [79], implying that GLP-1receptor activation preserves its effectiveness with chronic use andthat this is an effective and promising strategy that deserves furtherexploration [1]. The apparent synergistic effects of GLP-1 and itssynthetic analogs with other peptides on food intake, insulin releaseand glucose utilization support the idea of an increased therapeuticpotential for "cocktails" of GLP-1 and other agents. Such peptidecocktails might combine GLP-1 or its analogs with leptin or amylin,combinations which have been shown to produce synergistic effectson eating and body weight [80,81]. Combination of GLP-1 withanother peptide in a chimeric molecule represents also a promisingapproach. Such a molecule combining GLP-1 and pancreatic glucagon(which was identified as the first physiologically relevant peripheralsatiating peptide almost 30 years ago [82]), produced dramatic effectson body weight in mice [83]. Another exciting area of new research isthe discovery of GLP-1 preparations that are rapidly absorbed andprotected from intestinal enzymatic breakdown and can therefore beadministered orally [22]. Last but not least, the fact that L-cells –
similar to other enteroendocrine cells - express some of the same"taste" receptors as the taste cells in the oral cavity [84] and releasephysiologically relevant amounts of GLP-1 in response to stimulationby tastants raises further new opportunities. For example, combina-tions of in-vitro models of the GI epithelium, in-vivo animal modelsand food technology may lead to successful targeting of the intestinalendocrine cells to exploit novel therapeutic approaches to themodulation of GI peptide release to influence eating and metabolism.
All these discoveries suggest that the sensing and secretoryfunctions of the wall of the GI tract represent a major, heretoforeunderappreciated regulatory interface in the control of eating and
Please cite this article as: Punjabi M, et al, Peripheral Glucagon-like Pephysbeh.2011.02.038
energy balance, and a viable target for pharmacological approaches tocurb obesity and metabolic disease. Although a number of mecha-nisms have already been implicated in these actions, their precisemediating mechanisms in the gut and brain as well as their truephysiological relevance remain unclear. To eventually identify themost promising strategy – or combination of approaches – to fightobesity and type II diabetes, it is critically important to thoroughlycharacterize the site(s) and mechanism(s) of action of the endoge-nous peptides or their analogs involved. We hope that this reviewsucceeded to briefly summarize some key features of currentknowledge about intestinal GLP-1 and to identify gaps in knowledgewere further research is required.
References
[1] Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev 2007;87:1409–39.
[2] Goke R, Larsen PJ, Mikkelsen JD, Sheikh SP. Distribution of GLP-1 binding sites inthe rat brain: evidence that exendin-4 is a ligand of brain GLP-1 binding sites. Eur JNeurosci 1995;7:2294–300.
[3] Merchenthaler I, Lane M, Shughrue P. Distribution of pre-pro-glucagon andglucagon-like peptide-1 receptor messenger RNAs in the rat central nervoussystem. J Comp Neurol 1999;403:261–80.
[4] Kokrashvili Z, Mosinger B, Margolskee RF. T1r3 and alpha-gustducin in gutregulate secretion of glucagon-like peptide-1. Ann NY Acad Sci 2009;1170:91–4.
[5] Shin YK, Martin B, Golden E, Dotson CD, Maudsley S, Kim W, et al. Modulation oftaste sensitivity by GLP-1 signaling. J Neurochem 2008;106:455–63.
[6] Wei Y, Mojsov S. Distribution of GLP-1 and PACAP receptors in human tissues. ActaPhysiol Scand 1996;157:355–7.
[7] Dunphy JL, Taylor RG, Fuller PJ. Tissue distribution of rat glucagon receptorand GLP-1 receptor gene expression. Mol Cell Endocrinol 1998;141:179–86.
[8] Shimizu I, Hirota M, Ohboshi C, Shima K. Identification and localization ofglucagon-like peptide-1 and its receptor in rat brain. Endocrinology 1987;121:1076–82.
[9] Burcelin R, Da Costa A, Drucker D, Thorens B. Glucose competence of thehepatoportal vein sensor requires the presence of an activated glucagon-likepeptide-1 receptor. Diabetes 2001;50:1720–8.
[10] Holst JJ. Enteroglucagon. Annu Rev Physiol 1997;59:257–71.[11] During MJ, Cao L, Zuzga DS, Francis JS, Fitzsimons HL, Jiao X, et al. Glucagon-like
peptide-1 receptor is involved in learning and neuroprotection. Nat Med 2003;9:1173–9.
[12] Harkavyi A, Whitton PS. Glucagon-like peptide 1 receptor stimulation as a meansof neuroprotection. Br J Pharmacol 2010;159:495–501.
[13] Holst JJ, Burcelin R, Nathanson E. Neuroprotective properties of GLP-1: theoreticaland practical applications. Curr Med Res Opin.; 2011.
[14] Hayes MR, De Jonghe BC, Kanoski SE. Role of the glucagon-like-peptide-1 receptorin the control of energy balance. Physiol Behav 2010;100:503–10.
[15] van Dijk G, Thiele TE, Seeley RJ, Woods SC, Bernstein IL. Glucagon-like peptide-1and satiety. Nature 1997;385:214.
[16] Scott KA, Moran TH. The GLP-1 agonist exendin-4 reduces food intake innonhuman primates through changes in meal size. Am J Physiol Regul IntegrComp Physiol 2007;293:R983–7.
[17] Reidelberger RD, Haver AC, Apenteng BA, Anders KL, Steenson SM. Effects ofexendin-4 alone and with peptide YY(3–36) on food intake and body weight indiet-induced obese rats. Obes Silver Spring 2011;19:121–7.
[18] Bradley DP, Kulstad R, Schoeller DA. Exenatide andweight loss. Nutrition 2010;26:243–9.
[19] Larsen PJ, Fledelius C, Knudsen LB, Tang-Christensen M. Systemic administrationof the long-acting GLP-1 derivative NN2211 induces lasting and reversible weightloss in both normal and obese rats. Diabetes 2001;50:2530–9.
[20] Gutzwiller JP, Goke B, Drewe J, Hildebrand P, Ketterer S, Handschin D, et al.Glucagon-like peptide-1: a potent regulator of food intake in humans. Gut1999;44:81–6.
[21] Gutzwiller JP, Degen L, Heuss L, Beglinger C. Glucagon-like peptide 1 (GLP-1) andeating. Physiol Behav 2004;82:17–9.
[22] Steinert RE, Poller B, Castelli MC, Drewe J, Beglinger C. Oral administration ofglucagon-like peptide 1 or peptide YY 3–36 affects food intake in healthy malesubjects. Am J Clin Nutr 2010;92:810–7.
[23] Choukem SP, Gautier JF. How do different GLP-1 mimetics differ in their actions?Curr Diab Rep 2006;6:365–72.
[24] Young AA, Gedulin BR, Bhavsar S, Bodkin N, Jodka C, Hansen B, et al. Glucose-lowering and insulin-sensitizing actions of exendin-4: studies in obese diabetic(ob/ob, db/db) mice, diabetic fatty Zucker rats, and diabetic rhesus monkeys(Macaca mulatta). Diabetes 1999;48:1026–34.
[25] Joy SV, Rodgers PT, Scates AC. Incretin mimetics as emerging treatments for type 2diabetes. Ann Pharmacother 2005;39:110–8.
[26] Williams DL, Baskin DG, Schwartz MW. Evidence that intestinal glucagon-likepeptide-1 plays a physiological role in satiety. Endocrinology 2009;150:1680–7.
ptide-1 (GLP-1) and Satiation, Physiol Behav (2011), doi:10.1016/j.

6 M. Punjabi et al. / Physiology & Behavior xxx (2011) xxx–xxx
[27] Rüttimann EB, Arnold M, Hillebrand JJ, Geary N, Langhans W. Intrameal hepaticportal and intraperitoneal infusions of glucagon-like peptide-1 reduce spontane-ous meal size in the rat via different mechanisms. Endocrinology 2009;150:1174–81.
[28] Orskov C, Rabenhoj L, Wettergren A, Kofod H, Holst JJ. Tissue and plasmaconcentrations of amidated and glycine-extended glucagon-like peptide I inhumans. Diabetes 1994;43:535–9.
[29] Hassan M, Eskilsson A, Nilsson C, Jonsson C, Jacobsson H, Refai E, et al. In vivodynamic distribution of 131I-glucagon-like peptide-1 (7–36) amide in the ratstudied by gamma camera. Nucl Med Biol 1999;26:413–20.
[30] Berthoud HR. The vagus nerve, food intake and obesity. Regul Pept 2008;149:15–25.
[31] Berthoud HR, Neuhuber WL. Functional and chemical anatomy of the afferentvagal system. Auton Neurosci 2000;85:1–17.
[32] Nishizawa M, Nakabayashi H, Kawai K, Ito T, Kawakami S, Nakagawa A, et al. Thehepatic vagal reception of intraportal GLP-1 is via receptor different from thepancreatic GLP-1 receptor. J Auton Nerv Syst 2000;80:14–21.
[33] Nakabayashi H, Nishizawa M, Nakagawa A, Takeda R, Niijima A. Vagalhepatopancreatic reflex effect evoked by intraportal appearance of tGLP-1. Am JPhysiol 1996;271:E808–13.
[34] Norgren R, Smith GP. A method for selective section of vagal afferent or efferentaxons in the rat. Am J Physiol 1994;267:R1136–41.
[35] Holzer P. Capsaicin: cellular targets, mechanisms of action, and selectivity for thinsensory neurons. Pharmacol Rev 1991;43:143–201.
[36] Smith GP, Jerome C, Norgren R. Afferent axons in abdominal vagus mediate satietyeffect of cholecystokinin in rats. Am J Physiol 1985;249:R638–41.
[37] Baumgartner I, Pacheco-López G, Rüttimann EB, Arnold M, Asarian L, LanghansW, et al. Hepatic-portal vein infusions of glucagon-like peptide-1 reduce mealsize and increase c-Fos expression in the nucleus tractus solitarii, areapostrema and central nucleus of the amygdala in rats. J Neuroendocrinol2010;22:557–63.
[38] Rowland NE, Crews EC, Gentry RM. Comparison of Fos induced in rat brain byGLP-1 and amylin. Regul Pept 1997;71:171–4.
[39] Yamamoto H, Kishi T, Lee CE, Choi BJ, Fang H, Hollenberg AN, et al. Glucagon-likepeptide-1-responsive catecholamine neurons in the area postrema link peripheralglucagon-like peptide-1 with central autonomic control sites. J Neurosci 2003;23:2939–46.
[40] Thiele TE, Van Dijk G, Campfield LA, Smith FJ, Burn P, Woods SC, et al. Centralinfusion of GLP-1, but not leptin, produces conditioned taste aversions in rats. Am JPhysiol 1997;272:R726–30.
[41] Seeley RJ, Blake K, Rushing PA, Benoit S, Eng J, Woods SC, et al. The role of CNSglucagon-like peptide-1 (7–36) amide receptors in mediating the visceral illnesseffects of lithium chloride. J Neurosci 2000;20:1616–21.
[42] Kinzig KP, D'Alessio DA, Seeley RJ. The diverse roles of specific GLP-1 receptors inthe control of food intake and the response to visceral illness. J Neurosci 2002;22:10470–6.
[43] Pacheco-López G, Punjabi M, Graber M, Geary N, Arnold M, Langhans W.Hindbrain glucagon-like peptide-1 receptors (GLP-1R) mediate the eating-inhibitory effect of hepatic portal vein (HPV) GLP-1 infusions. Society for theStudy of Ingestive Behavior, Annual meeting; 2010. Pittsburg, Appetite 54, 688.
[44] Baraboi ED, Smith P, Ferguson AV, Richard D. Lesions of area postrema andsubfornical organ alter exendin-4-induced brain activation without preventingthe hypophagic effect of the GLP-1 receptor agonist. Am J Physiol Regul IntegrComp Physiol 2010;298:R1098–110.
[45] Montrose-Rafizadeh C, Avdonin P, Garant MJ, Rodgers BD, Kole S, Yang H, et al.Pancreatic glucagon-like peptide-1 receptor couples to multiple G proteins andactivates mitogen-activated protein kinase pathways in Chinese hamster ovarycells. Endocrinology 1999;140:1132–40.
[46] Holz GGT, Leech CA, Habener JF. Activation of a cAMP-regulated Ca(2+)-signalingpathway in pancreatic beta-cells by the insulinotropic hormone glucagon-likepeptide-1. J Biol Chem 1995;270:17749–57.
[47] Sutton GM, Patterson LM, Berthoud HR. Extracellular signal-regulated kinase 1/2signaling pathway in solitary nucleus mediates cholecystokinin-induced suppres-sion of food intake in rats. J Neurosci 2004;24:10240–7.
[48] Sutton GM, Duos B, Patterson LM, Berthoud HR. Melanocortinergic modulation ofcholecystokinin-induced suppression of feeding through extracellular signal-regu-lated kinase signaling in rat solitary nucleus. Endocrinology 2005;146:3739–47.
[49] Gomez E, Pritchard C, Herbert TP. cAMP-dependent protein kinase and Ca2+influx through L-type voltage-gated calcium channels mediate Raf-independentactivation of extracellular regulated kinase in response to glucagon-like peptide-1in pancreatic beta-cells. J Biol Chem 2002;277:48146–51.
[50] D'Alessio D, LuW, SunW, Zheng S, Yang Q, Seeley R, et al. Fasting and postprandialconcentrations of GLP-1 in intestinal lymph and portal plasma: evidence forselective release of GLP-1 in the lymph system. Am J Physiol Regul Integr CompPhysiol 2007;293:R2163–9.
[51] Iritani N, Sugimoto T, Fukuda H, Komiya M, Ikeda H. Oral triacylglycerols regulateplasma glucagon-like peptide-1(7–36) and insulin levels in normal and especiallyin obese rats. J Nutr 1999;129:46–50.
[52] Anini Y, Fu-Cheng X, Cuber JC, Kervran A, Chariot J, Roz C. Comparison of thepostprandial release of peptide YY and proglucagon-derived peptides in the rat.Pflugers Arch 1999;438:299–306.
[53] Rüttimann EB. Glucagon-like peptide-1 (GLP-1) and satiation [Diss, EidgenössischeTechnische Hochschule ETH Zürich, Nr 18686, 2010]. Zürich: ETH; 2010.
[54] Shin AC, Zheng H, Townsend RL, Sigalet DL, Berthoud HR. Meal-induced hormoneresponses in a rat model of Roux-en-Y gastric bypass surgery. Endocrinology2010;151:1588–97.
Please cite this article as: Punjabi M, et al, Peripheral Glucagon-like Pephysbeh.2011.02.038
[55] Beglinger C, Degen L. Gastrointestinal satiety signals in humans–physiologic rolesfor GLP-1 and PYY? Physiol Behav 2006;89:460–4.
[56] Elliott RM, Morgan LM, Tredger JA, Deacon S, Wright J, Marks V. Glucagon-likepeptide-1 (7–36)amide andglucose-dependent insulinotropic polypeptide secretionin response to nutrient ingestion in man: acute post-prandial and 24-h secretionpatterns. J Endocrinol 1993;138:159–66.
[57] Kieffer TJ, McIntosh CH, Pederson RA. Degradation of glucose-dependentinsulinotropic polypeptide and truncated glucagon-like peptide 1 in vitro and invivo by dipeptidyl peptidase IV. Endocrinology 1995;136:3585–96.
[58] Rüttimann EB, Arnold M, Geary N, Langhans W. GLP-1 antagonism with exendin(9–39) fails to increase spontaneousmeal size in rats. Physiol Behav 2010;100:291–6.
[59] Perez-Tilve D, Gonzalez-Matias L, Aulinger BA, Alvarez-Crespo M, Gil-Lozano M,Alvarez E, et al. Exendin-4 increases blood glucose levels acutely in rats byactivation of the sympathetic nervous system. Am J Physiol Endocrinol Metab2010;298:E1088–96.
[60] Berthoud HR, Kressel M, Raybould HE, Neuhuber WL. Vagal sensors in the ratduodenal mucosa: distribution and structure as revealed by in vivo DiI-tracing.Anat Embryol Berl 1995;191:203–12.
[61] Williams RM, Berthoud HR, Stead RH. Vagal afferent nerve fibres contact mast cells inrat small intestinal mucosa. Neuroimmunomodulation 1997;4:266–70.
[62] Miller MJ, McDole JR, Newberry RD. Microanatomy of the intestinal lymphaticsystem. Ann NY Acad Sci 2010;1207(Suppl 1):E21–8.
[63] Ohtani O. Three-dimensional organization of lymphatics and its relationship toblood vessels in rat small intestine. Cell Tissue Res 1987;248:365–74.
[64] Manolas KJ, Adrian TE, Dunlop HM, Bacarese-Hamilton AJ, Bloom SR, WelbournRB. Lymph, pancreatic and gastrointestinal hormones in response to feeding in theconscious pig. Eur Surg Res 1985;17:324–32.
[65] Tong J, TschopMH, Aulinger BA, Davis HW, Yang Q, Liu J, et al. The intestinal lymphfistula model–a novel approach to study ghrelin secretion. Am J PhysiolGastrointest Liver Physiol 2010;298:G474–80.
[66] Bucinskaite V, Tolessa T, Pedersen J, Rydqvist B, Zerihun L, Holst JJ, et al. Receptor-mediated activation of gastric vagal afferents by glucagon-like peptide-1 in the rat.Neurogastroenterol Motil 2009;21:978-e78.
[67] Edwards CM, Todd JF, Mahmoudi M, Wang Z, Wang RM, Ghatei MA, et al.Glucagon-like peptide 1 has a physiological role in the control of postprandialglucose in humans: studies with the antagonist exendin 9–39. Diabetes 1999;48:86–93.
[68] Kolligs F, Fehmann HC, Goke R, Goke B. Reduction of the incretin effect in rats bythe glucagon-like peptide 1 receptor antagonist exendin (9–39) amide. Diabetes1995;44:16–9.
[69] Wang Z, Wang RM, Owji AA, Smith DM, Ghatei MA, Bloom SR. Glucagon-likepeptide-1 is a physiological incretin in rat. J Clin Invest 1995;95:417–21.
[70] Kim DH, D'Alessio DA, Woods SC, Seeley RJ. The effects of GLP-1 infusion in thehepatic portal region on food intake. Regul Pept 2009;155:110–4.
[71] Balkan B, Li X. Portal GLP-1 administration in rats augments the insulin responseto glucose via neuronal mechanisms. Am J Physiol Regul Integr Comp Physiol2000;279:R1449–54.
[72] Guillam MT, Hummler E, Schaerer E, Yeh JI, Birnbaum MJ, Beermann F, et al. Earlydiabetes and abnormal postnatal pancreatic islet development in mice lackingGlut-2. Nat Genet 1997;17:327–30.
[73] Burcelin R, DolciW, Thorens B. Glucose sensing by the hepatoportal sensor is GLUT2-dependent: in vivo analysis in GLUT2-null mice. Diabetes 2000;49:1643–8.
[74] Cani PD, Holst JJ, Drucker DJ, Delzenne NM, Thorens B, Burcelin R, et al. GLUT2 andthe incretin receptors are involved in glucose-induced incretin secretion. Mol CellEndocrinol 2007;276:18–23.
[75] Naslund E, Barkeling B, King N, Gutniak M, Blundell JE, Holst JJ, et al. Energy intakeand appetite are suppressed by glucagon-like peptide-1 (GLP-1) in obese men. IntJ Obes Relat Metab Disord 1999;23:304–11.
[76] Vincent RP, Ashrafian H, le Roux CW. Mechanisms of disease: the role ofgastrointestinal hormones in appetite and obesity. Nat Clin Pract GastroenterolHepatol 2008;5:268–77.
[77] Campbell R, Smith G. CCK-8 decreases body weight in Zucker rats. Annual meetingof Society for Neuroscience; 1983. Boston, MA., USA.
[78] Smith GP, Gibbs J. The Satiety Effect of Cholecystokinin - Recent Progress andCurrent Problems. Ann NY Acad Sci 1985;448:417–23.
[79] Schmitz O. The GLP-1 concept in the treatment of type 2 diabetes–still standing atthe gate of dawn? J Clin Endocrinol Metab 2008;93:375–7.
[80] Roth JD, Coffey T, Jodka CM, Maier H, Athanacio JR, Mack CM, et al. Combinationtherapy with amylin and peptide YY[3–36] in obese rodents: anorexigenicsynergy and weight loss additivity. Endocrinology 2007;148:6054–61.
[81] Williams DL, BaskinDG, SchwartzMW. Leptin regulation of the anorexic response toglucagon-like peptide-1 receptor stimulation. Diabetes 2006;55:3387–93.
[82] Langhans W, Zieger U, Scharrer E, Geary N. Stimulation of feeding in rats byintraperitoneal injection of antibodies to glucagon. Science 1982;218:894–6.
[83] Day JW, Ottaway N, Patterson JT, Gelfanov V, Smiley D, Gidda J, et al. A new glucagonand GLP-1 co-agonist eliminates obesity in rodents. Nat Chem Biol 2009;5:749–57.
[84] Margolskee RF, Dyer J, Kokrashvili Z, Salmon KS, Ilegems E, Daly K, et al. T1R3 andgustducin in gut sense sugars to regulate expression of Na+−glucosecotransporter 1. Proc Natl Acad Sci USA 2007;104:15075–80.
[85] Imeryuz N, Yegen BC, Bozkurt A, Coskun T, Villanueva-Penacarrillo ML, Ulusoy NB.Glucagon-like peptide-1 inhibits gastric emptying via vagal afferent-mediatedcentral mechanisms. Am J Physiol Gastrointest Liver Physiol 1997;273:G920–7.
[86] Vahl TP, Tauchi M, Durler TS, Elfers EE, Fernandes TM, Bitner RD, et al. Glucagon-like peptide-1 (GLP-1) receptors expressed on nerve terminals in the portal veinmediate the effects of endogenous GLP-1 on glucose tolerance in rats.Endocrinology 2007;148:4965–73.
ptide-1 (GLP-1) and Satiation, Physiol Behav (2011), doi:10.1016/j.
Related Documents











