doi:10.1152/jn.00538.2013 113:156-171, 2015. First published 8 October 2014; J Neurophysiol Kwee-Yum Lee, Nicholas O'Dwyer, Mark Halaki and Richard Smith stick-balancing skill Perceptual and motor learning underlies human You might find this additional info useful... 69 articles, 17 of which can be accessed free at: This article cites /content/113/1/156.full.html#ref-list-1 including high resolution figures, can be found at: Updated information and services /content/113/1/156.full.html can be found at: Journal of Neurophysiology about Additional material and information http://www.the-aps.org/publications/jn This information is current as of January 14, 2015. American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at http://www.the-aps.org/. (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2015 by the publishes original articles on the function of the nervous system. It is published 12 times a year Journal of Neurophysiology on January 14, 2015 Downloaded from on January 14, 2015 Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1152/jn.00538.2013 113:156-171, 2015. First published 8 October 2014;J NeurophysiolKwee-Yum Lee, Nicholas O'Dwyer, Mark Halaki and Richard Smithstick-balancing skillPerceptual and motor learning underlies human
You might find this additional info useful...
69 articles, 17 of which can be accessed free at:This article cites /content/113/1/156.full.html#ref-list-1
including high resolution figures, can be found at:Updated information and services /content/113/1/156.full.html
can be found at:Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of January 14, 2015.
American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at http://www.the-aps.org/.(monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2015 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on January 14, 2015D
ownloaded from
on January 14, 2015D
ownloaded from

Perceptual and motor learning underlies human stick-balancing skill
Kwee-Yum Lee,1,3 Nicholas O’Dwyer,2,3 Mark Halaki,3 and Richard Smith3
1School of Exercise Science, Australian Catholic University, Strathfield, New South Wales, Australia; 2School of HumanMovement Studies, Charles Sturt University, Bathurst, New South Wales, Australia; and 3Discipline of Exercise and SportScience, The University of Sydney, New South Wales, Australia
Submitted 26 July 2013; accepted in final form 5 October 2014
Lee KY, O’Dwyer N, Halaki M, Smith R. Perceptual andmotor learning underlies human stick-balancing skill. J Neuro-physiol 113: 156 –171, 2015. First published October 8, 2014;doi:10.1152/jn.00538.2013.—We investigated the acquisition ofskill in balancing a stick (52 cm, 34 g) on the fingertip in nineparticipants using three-dimensional motion analysis. After 3.5 h ofpractice over 6 wk, the participants could more consistently balancethe stick for longer durations with greatly reduced magnitude andspeed of stick and finger movements. Irrespective of level of skill, thebalanced stick behaved like a normal noninverted pendulum oscillat-ing under greater-than-gravity torque with simple harmonic motionabout a virtual pivot located at the radius of gyration above the centerof mass. The control input parameter was the magnitude ratio betweenthe torque applied on the stick by the participant and the torque due togravity. The participants utilized only a narrow range of this param-eter, which did not change with practice, to rotate the stick like alinear mass-spring system. With increased skill, the stick thereforemaintained the same period of oscillation but showed marked reduc-tions in magnitude of both oscillation and horizontal translation.Better balancing was associated with 1) more accurate visual local-ization of the stick and proprioceptive localization of the finger and 2)reduced cross-coupling errors between finger and stick movements inorthogonal directions; i.e., finger movements in the anteroposteriorplane became less coupled with stick tip movements in the mediolat-eral plane, and vice versa. Development of this fine motor skilltherefore depended on perceptual and motor learning to provideimproved estimation of sensorimotor state and precision of motorcommands to an unchanging internal model of the rotational dynam-ics.
pole balancing; perception-action coupling; depth perception; kines-thesis; gravitational dynamics
BALANCING AN OBJECT can be categorized as a continuous skillwhereby an inherently unstable object has to be kept uprightwithin the physical limitations and boundaries of the controller.When an object with a relatively small base of support has tobe kept upright or transported, fine control is required tobalance the object to prevent it from falling under the force ofgravity. In one aspect at least, human adults are expert balanc-ers since they can balance their own body well by achievingupright posture over a small base of support and can walk andrun without falling. These feats seem natural for adults, butmastering the motor skills needed to perform these taskscertainly requires a significant amount of practice for infants(Payne and Isaacs 2008). Hence, studying the mechanisms oflearning to balance through the behavior of infants requireslong periods of observation. In this respect, the skill of balanc-ing a stick on the fingertip is a challenging task that provides
a good opportunity to investigate the mechanisms of skilllearning, since it is a skill that can be acquired in several hoursof practice by young adults (Cabrera and Milton 2004a, 2012).To control an unstable object such as this requires constantvigilance and sustained visual attention. The subjective expe-rience of the participants indicates that although it is possibleto think about something else or to speak while balancing, ifvisual or proprioceptive attention is removed from the stick formore than a fleeting moment, stability is lost. Hence, even askilled performer can maintain balancing for only a few min-utes (Cabrera and Milton 2004b). Another interesting featureof the task is that augmented feedback from an expert trainerapparently cannot be used to guide the learner, and no obvious“rules” can be expressed to aid skill acquisition, which insteadappears to depend on implicit learning (Stevens et al. 2012).
Since the work of Barto et al. (1983), object balancing, alsoknown as the inverted pendulum problem, has been a benchmarkproblem in the field of control systems engineering. The object tobe kept upright is usually a pole which pivots on a motorized cartthat moves on a one-dimensional track. Investigators in numerousstudies have successfully designed various controllers that can betrained to balance and stabilize such an object rapidly with highefficiency (Connell and Utgoff 1987; Guez and Selinsky 1988;Koza and Keane 1990; Lee and Berenji 1989; Widrow 1987).
While the mechanism(s) underlying the learning behavior ofthese machine controllers can in principle be specified, itremains unclear how relevant they are to learning in humans.Lakie et al. (2003) have shown that a large inverted pendulumwith low intrinsic stiffness can be controlled via anticipatoryhand movements, with stability being provided by a neurallygenerated, intermittent pattern of impulses despite irregular andaperiodic pendulum sway. In a previous study of balancing a stickon the fingertip by skilled performers (Lee et al. 2012), weobserved that the movement of the stick tip was only about halfthat of the finger. This fundamental kinematic finding showed thatthe balanced stick pivoted more like a normal noninverted pen-dulum than the conventional inverted pendulum model fromcontrol systems engineering, a point also noted in previous studies(Cabrera and Milton 2002; Treffner and Kelso 1999). Indeed, thebehavior of the balanced stick resembled angular simple harmonicmotion of a normal pendulum, but operating under greater-than-gravity torque (applied by the balancer) and with a translatingpivot point. Oscillatory movements between the finger and sticktip also have been emphasized in other studies of stick balancing,using time-delayed inverted pendulum models (Milton et al.2009a, 2009b), but the noninverted pendulum model is purely amechanical description of the behavior of the balanced stick andtherefore is independent of the issue of time delay (see Lee et al.2012).
Address for reprint requests and other correspondence: N. O’Dwyer, Schoolof Human Movement Studies, Charles Sturt Univ., Bathurst, NSW 2795,Australia (e-mail: [email protected]).
J Neurophysiol 113: 156–171, 2015.First published October 8, 2014; doi:10.1152/jn.00538.2013.
156 0022-3077/15 Copyright © 2015 the American Physiological Society www.jn.org
on January 14, 2015D
ownloaded from

In this descriptive model of a pendulum, the relevant inputcontrol parameter was the magnitude ratio between the activetorque applied by the participant and the torque due to gravity.This parameter determined the distinctive location of the vir-tual pivot point of the pendulum, which was approximatelyat the radius of gyration (measured about the center of mass,CoM) above the CoM of the stick. This location pointed to theimportance of knowledge of the gravitational dynamics andmass distribution of the stick for the interaction of the partic-ipants with the movement of the stick. A subsequent control-theoretical interpretation of this noninverted pendulum model(Gawthrop et al. 2013) showed that the virtual pivot is the keyto successful balancing, enabling humans to simplify the con-trol by approximately splitting the fourth-order dynamics of thesystem into two noninteracting second-order systems: controlof the angle (stick rotations) using finger acceleration at anappropriate frequency to create the virtual pivot, and control ofthe horizontal position of the virtual pivot (stick translations)using the pendulum angle controller set point. The structure ofthe two subsystems is such that they are amenable to cascadecontrol, and it is known that humans can successfully controltwo noninteracting second-order systems simultaneously (Oy-tam et al. 2005).
Although there have been a number of previous studies ofstick balancing (Cabrera and Milton 2002, 2004a, 2004b, 2012;Cluff and Balasubramaniam 2009; Cluff et al. 2009, 2011,2012; Foo et al. 2000; Mehta and Schaal 2002; Milton 2011;Milton et al. 2008, 2009a, 2009b; Reeves et al. 2013; Treffnerand Kelso 1999), there has not been extensive study of thechanges in stick-balancing behavior with practice. Cabrera andMilton (2004a) had participants practice stick balancing over anumber of days. The change in the speed of hand movementswas used to estimate the “corrective” movements of the hand,and faster changes were observed after practice. Cluff andBalasubramaniam (2009) also reported faster changes in fin-gertip speed after practice and concluded, in agreement withCabrera and Milton, that learning resulted in an increasedtolerance for larger speed step sizes of the finger. Cluff et al.(2009) analyzed the time series dynamics of the finger dis-placement data of Cluff and Balasubramaniam (2009) andfound that learning was accompanied by increased dynamicalstability of movement trajectories.
Whereas these studies provided insights into skill-relatedchanges in the statistical properties of hand movements, twofurther studies by Cluff and colleagues examined concomitantchanges in postural and limb control. Cluff et al. (2011)showed that the trajectories of both finger and center ofpressure displacements became more variable and discontinu-ous with practice and that the coupling of finger displacementswith postural sway increased. They suggested that this cou-pling resulted from the development of a hierarchical controlmechanism that switches intermittently between coupled andindependent control of the finger and posture. Cluff et al.(2012) analyzed the stick angle and six upper and lower limbjoint angles in the sagittal plane as subjects learned to balance.They found a reduction in individual joint angle variationconcomitant with an increase in multijoint covariation, indi-cating that subjects relied less on the control of individualjoints and more on distributed error compensation, a transitionthat was correlated with improved balancing performanceacross subjects.
In our previous study of skilled stick balancing (Lee et al.2012), we presented detailed three-dimensional (3-D) kine-matic and kinetic measures that are not available from otherstudies to date. In the present study we apply these measures toidentify hitherto unreported basic behavioral changes withpractice, with the aim of extending understanding of thelearning mechanisms for skilled balancing. In addition, thependulum model proposed previously (Lee et al. 2012) wasinvestigated further to determine whether changes in the be-havior of the model occurred as a result of learning.
MATERIALS AND METHODS
Participants
Fourteen right-handed young adult participants (6 women, 8 men,ages 18–35 yr) with no history of motor disorders volunteered toparticipate in the study. All were naive to the stick-balancing task. Theexperimental procedures were approved by the Human ResearchEthics Committee of The University of Sydney, and written informedconsent was obtained. Each participant received a pair of cinemavouchers at the beginning of the study and another pair on completion.
Task and practice
The task goal was for the participant to balance a stick on the padof the middle finger of the right hand while standing upright for atleast 20 s without taking a step. The participants were given 10 minof familiarization immediately before data collection was begun. Atthe beginning of each trial the participants supported the stick withtheir left hand, which they released once they were given the “go”signal for data collection. Trials were stopped once 20 s had beenreached. Twenty trials were performed, and the whole experimenttook about 1 h to complete. An identical procedure was repeated forthe postpractice trials.
To acquire the skill of balancing, each participant was providedwith a stick and asked to practice at home for a total of 3.5 hdistributed over 6 wk. Participants were instructed to practice for atleast 5 min daily and were given a log book to encourage compliancewith this schedule. An extra pair of cinema vouchers was promised ifthey were able to 1) double the number of successful trials they hadachieved in the prepractice session or 2) succeed at all trials in thepostpractice session. The log books indicated that all participantsachieved the target hours of practice.
Apparatus and Data Acquisition
The experimental methods used in the current study are the same asthose reported in Lee et al. (2012) and are presented only in summaryform in this report. The stick was a telescopic television antenna(length, 0.52 m; diameter, 5–7 mm; mass, 0.0338 kg; moment ofinertia, 0.0029 kg·m2; distance from base to CoM, 0.27 m). Asemi-cone-shaped base (8-mm diameter), originally the tip of theantenna, was in full contact with the finger pad, preventing slippagebetween the finger and the base of the stick during balancing. Tworetroreflective markers (0.56-g Styrofoam spheres) were attached tothe stick, one at the tip and another 9 cm above the base, to track the3-D movement of the stick. To track the 3-D movement of eachparticipant’s left and right arm and head, five reflective markers(25-mm diameter) were attached on the left and right radial styloidprocess, the forehead, and the left and right zygomatic bone. Tencameras (Eagle cameras; Motion Analysis, Santa Rosa, CA) wereused to track the movements of the stick and participants with 100-Hzcapture frequency. The laboratory coordinate system was defined asmediolateral (ML), anteroposterior (AP), and vertical (V) axes paral-leling the orientation of the participants at the start of the task.
157PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

Captured trial data were imported for analysis into Matlab 7.0 soft-ware (The MathWorks).
Trimming of the raw data was necessary to ensure that only datafrom true balancing of the stick were analyzed. The starting point ofeach trial was chosen when the supporting left hand released the stick,identified from the onset of movement of the marker on the left radialstyloid process. For trials less than 20 s in duration, the end point wasselected by examining the sway cycles between the upper and lowermarkers on the stick. A failure of balancing was easily identified bythe desynchronization of the trajectories of the two markers on thestick in the ML and/or AP axes. The end point of such trials waschosen two sway cycles prior to the initiation of such a desynchroni-zation. Only trials comprising more than 5 s of data were analyzed.
Data Analysis
The variables measured are described briefly below. All variablesexcept the head-centered coordinate system were described in detailby Lee et al. (2012). In addition, frequency domain and linear systemsanalyses were used to investigate the relations between kinematic andkinetic variables to garner further insights into the effects of practiceon stick-balancing performance.
Assessment of balancing performance. The number and duration ofsuccessful stick-balancing trials were the most basic performancemeasurements. The duration of balancing is perhaps the most tellingmeasure of level of skill, because balancing can be maintained foronly a limited duration even by a skilled stick balancer (Cabrera andMilton 2004b). The range of movement in each 3-D direction (themaximum–minimum position) of the stick and finger was measured toassess the stability of the stick and the amount of finger movement.The total distance traveled in the laboratory space by the stick tip andfinger was calculated for each trial by summing the 3-D distancetraveled across sample intervals. The normalized length of path (LoP)was then defined as the total distance traveled by the stick tip or fingerdivided by the duration of each trial in seconds.
Kinematic variables. The position, velocity, and acceleration of thefinger and stick tip were measured in each axis. To reveal how thefinger and stick tip moved vertically in the laboratory space, the meanvalue of the stick tip position in the V axis was calculated. The anglesof deviation of the stick from the vertical in the V � ML plane (MLangle) and V � AP plane (AP angle) were calculated. The axial angleof the stick, which represents its rotation about the longitudinal axis,was estimated as described previously (Lee et al. 2012). The velocityand acceleration of these angles were calculated. Root-mean-square(RMS) values of these kinematic variables of the stick tip and fingerwere calculated. To provide a measure of the most common angulardisplacement of the stick from the vertical, the absolute magnitude ofthe angle of deviation of the stick from the vertical was calculated foreach data point (10-ms interval) from each trial, and a histogramnormalized to the length of the trial was constructed.
Half-cycles of stick oscillation. To identify the periodic nature ofthe stick movement, the average duration of the interval betweenzero-crossings of the positive and negative angular velocity signalswas obtained separately for the ML and AP directions. This durationis equivalent to one-half the period of the sway cycle of the stick.These cycles indicate the following directions of movement of thestick: toppling to the left for M¡L toppling cycle, toppling to theright for M¢L toppling cycle, toppling backward for A¡P topplingcycle, and toppling forward for A¢P toppling cycle.
Relative coordinates. A relative coordinate system that measuredthe stick angle with reference to the participant’s head was employedto gain insight into how the stick moved relative to the visual field.The origin of this relative coordinate system (MLrel, APrel, and Vrel)was the forehead marker. The MLrel axis was aligned with the linebetween the markers on the left and right zygomatic bone. Theorientation of the Vrel axis was parallel to that of the V axis of thelaboratory coordinate system (and the gravitational field). The APrel
axis was orthogonal to the MLrel and Vrel axes. Hence, both the MLrel
and APrel axes moved with rotations of the head in the horizontalplane. Although the head-referenced motion of the stick in the MLrel
axis was not a precise measure of its motion relative to the angle ofgaze, substantial coupling of head with eye movements could beassumed to occur during stick balancing. Coupling of head and eyemovements occurs in saccades of less than 3° in reading text (Lee 1999),and the contribution of head movements increases as the size of gazeshift increases (Freedman 2008; Guitton and Volle 1987). Further-more, head movements contribute significantly to predictive gazeshifts (Bizzi et al. 1972), which could be assumed during stickbalancing because the trajectory of motion of the stick when it fallsunder gravity is familiar and predictable (McIntyre et al. 2001; Senotet al. 2005; Zago and Lacquaniti 2005; Zago et al. 2004, 2008) fromthe moment the fall begins (only the precise moment and direction offalling are unpredictable). Even for small gaze shifts where the headdoes not move, the MLrel position of the head would still be closelyaligned with that of the stick tip.
In this relative coordinate system we defined the angle of the stickprojected onto the transverse MLrel � APrel plane as the “toppling”angle. This angle is equivalent to the relative direction of toppling ofthe stick from the vertical (0–360°) when viewed from above. Weexamined the amount of time spent at each toppling direction, inde-pendent of its magnitude, by constructing a frequency histogram ofthe toppling angle at each data point (10-ms interval). The frequencycount of the toppling direction in each 5° bin was normalized to thetrial duration and expressed as a percentage. The pattern of these anglehistograms was fitted with an ellipse to quantify their shape (see Fig.4A) by using a nonlinear fitting method that minimizes the geometricerror (Gander et al. 1994). The center coordinate of the ellipse (ameasure of whether the toppling was more frequent in one direction orthe other), the length of the major and minor axes (a measure of theratio of the toppling frequency in one axis vs. the other), and the tiltangle of the major axis (a measure of the deviation from the APrel
axis) were calculated.Kinetic variables. The derivations of these variables are described
in detail by Lee et al. (2012). The MoI with respect to the pivot pointat the base of the stick was derived empirically using the mass of thestick and its oscillation period under gravity (Tipler 1999) andchecked theoretically using the equations for composite MoI ofhollow cylinders (Halliday et al. 2001). The MoI of axial rotation(MoIaxial) of the stick was obtained by the equation for a hollowcylinder rotating about a longitudinal axis (Halliday et al. 2001). Thenet torque (�net) acting on the stick was computed from the angular accel-
eration (�̈) and the MoI in both the V � ML and V � AP planes.Similarly, the axial torque (�axial) was calculated from the axialangular acceleration and the MoIaxial. The net torque is the sum ofgravitational torque (�grav) and active torque (�act), where gravitationaltorque is the torque produced by gravity that rotates the stick about itsbase and active torque is caused by finger acceleration. The fingerreaction force (the translational stick force acting on the fingertip) wascalculated in ML, AP, and V axes using the acceleration data of thestick CoM. RMS values of the kinetic variables (3-D net torque, 2-Dactive torque, 2-D gravitational torque, and 3-D stick force) of thestick and finger were also calculated.
Frequency domain analyses. The frequency content of all thekinematic and kinetic variables of the stick and finger were analyzedusing fast Fourier transformation with a 512-point Hamming window.The frequency resolution was 0.1 Hz, and frequencies up to 5 Hz wereexamined. The spectrum describes the total power (� mean squarevalue) of the signals and its distribution across frequency. It was thedistribution across frequency that was of primary interest here, ratherthan the absolute power, which was already adequately assessed by themagnitude measures of the variables described above. Therefore, thepower spectral density of each signal was computed. This is the “relativepower spectrum,” that is, the power at each frequency expressed as a
158 PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

proportion of the total power in the spectrum. Thus the total area undera spectral density curve is unity. The median power frequency (MPF) foreach spectrum was also computed across the trials.
Linear systems analyses. To investigate the dynamic physical prop-erties of the control of stick translation and rotation, linear systemstechniques of cross-correlational and spectrographic analysis (Ada et al.1993; Bendat and Piersol 1986; Neilson 1972; Winter and Patla 1997)were employed. Coherence, gain, and phase frequency-response func-tions were calculated in the ML and AP axes between finger position andstick tip position, finger position and stick angle, finger reaction force andstick tip position, and net torque and stick angle.
Crossed-axes analysis. The relation between the finger position andthe stick tip position across the ML and AP axes was analyzed toidentify cross talk between these orthogonal axes (i.e., finger MLposition vs. stick tip AP position and finger AP position vs. stick tipML position). To quantify the degree of cross talk between thesevariables, the “overall coherence” (Lay et al. 2002; Oytam et al. 2005;Porges et al. 1980; Smith et al. 2001; Wang et al. 2013) was computedfrom a cross-correlational and spectrographic analysis. The overallcoherence provides a measure across all frequencies of the proportionof the total variance of the output that is linearly related to the input.Thus the overall coherence provides a single measure of the totalvariance accounted for by the linear transfer function and in this wayis analogous to the square of the Pearson product moment correlation(r2, the coefficient of determination). Hence, this measure revealed theamount of coupling between mediolateral and anteroposterior move-ments and how this coupling changed with practice.
Characteristic dynamics of the stick. The movements of the sticktip and finger included both oscillatory movement and translations. Toreveal the typical oscillatory cycle of the stick unaffected by cycle-to-cycle variations in translation, the stick angle, angular velocity, andactive torque in the ML and AP directions were time normalized foreach half-cycle of oscillation (M¡L, M¢L, A¡P, and A¢P top-pling cycle) and averaged across all trials and participants. For eachcycle of oscillation, the mean relation between active torque and angleand between angular velocity and angle was calculated for eachparticipant. The maximum positive and negative value of the averageangle from each cycle was defined as the “correction” angle, whichrepresents the angle at which the stick reversed its direction ofmovement to prevent it from falling.
Frequency histograms of the pendulum length. In the descriptivekinematic model of the balanced stick as angular simple harmonicmotion of a normal pendulum developed by Lee et al. (2012), anequation was developed using the ratio (k) between the magnitudes ofthe active torque and the gravitational torque to calculate where thepivot point was located. The mean value of this ratio for each topplingdirection in each participant was derived from the ratio of the slope oftheir mean relation between active torque and angle and the slope ofthe gravitational line. This ratio sets both the pivot point of the pendulumand the sway cycle period of the stick. The length from the pivot point tothe base of the stick contacting the finger was termed the pendulumlength (PL). Different values of PL indicated different behaviors ofthe stick. Three main stick behaviors were operationally defined (seeFig. 4 of Lee et al. 2012): 1) pure falling of the stick, when PL � 0m (the pivot point is at or below the base of the stick, which showsfalling on the finger due to gravitational torque); 2) rotation of thestick, when the PL ranged from 0 m to twice the most frequent PL (thepivot point is located at a point along the stick or up to 30 cm abovethe tip of the stick, which shows oscillatory movement); or 3) puretranslation of the stick, when the PL is equal to or greater than twicethe most frequent PL (the pivot point is beyond the tip of the stick,and the base and tip travel in parallel). To assess changes with skill,frequency histograms of the PL were calculated at 10-ms intervals forcollated trials for each participant before and after practice. Thefrequency values of the histogram are expressed as a percentage oftotal trial duration and averaged across participants.
Statistical analyses. The effect of practice on the dependent vari-ables was examined using 2 � 2 (Practice [Pre, Post] � Axes [ML,AP]) repeated-measures analyses of variance (ANOVA) for stickangle and net torque variables; 2 � 3 (Practice [Pre, Post] � Axes[ML, AP, V]) repeated-measures ANOVAs for finger and stick tiprange, position, and finger force variables; and t-tests for dependentsamples for trial duration, LoP, and axial angle variables (� level �0.05). The influence of the axes and the sequence of toppling wasmeasured for some variables by 2 � 2 � 2 (Practice [Pre, Post] �Axes [ML, AP] � Toppling Sequence [M¡L, M¢L or A¡P,A¢P]) repeated-measures ANOVAs. Tukey post hoc tests were usedwhen significant results were found. All data are expressed asmeans � SD. Statistical analyses were performed in Statistica 7software (StatSoft, version 7.1).
RESULTS
Assessment of Balance Performance
All participants attempted 20 trials of balancing the stick for20 s in both pre- and postpractice sessions. Five participantscould not balance for more than 5 s during prepractice trials.They showed a similar pattern whereby once the stick wasreleased from the left hand, it fell with no more than oneoscillatory hand movement. The data from these participantswere excluded from further analysis, and the number of par-ticipants in the statistical analyses was thus reduced to nine.For these nine participants, the data from all trials longer than5 s were averaged and the mean values were used to obtaingroup mean and SD values.
After practicing for 6 wk (� 5 min per day, 3.5 h in total),the participants were more consistently able to balance thestick for a longer duration (Table 1). The average number ofsuccessful trials increased with practice from 3.7 � 5.5 to 15.9 �4.0 (single-sided t-test, P � 0.05), while the average duration
Table 1. Number of analyzable trials and mean duration pertrial pre- and postpractice for each participant
Participant
Prepractice Postpractice
No. oftrials
Meanduration, s
No. oftrials
Meanduration, s
1 0 20 17.62 4 8.9 20 19.13 14 9.3 20 14.94 0 14 16.45 3 6.2 18 15.86 0 18 16.17 8 7.9 19 15.68 20 17.2 20 17.89 6 8.7 20 19.5
10 15 13.4 20 18.711 20 16.2 15 16.612 0 16 10.213 3 5.6 17 14.914 0 16 16.0
Data are no. of analyzable trials (�5-s duration) and mean duration per trial(in seconds) pre- and postpractice for each participant. The duration ofpostpractice trials was �20 s in most cases due to the data-trimming process(mean duration of data trimmed � 3.4 s). Only the 9 participants who had morethan 3 analyzable trials in the prepractice session were included in furtheranalyses. Participants 8 and 11 were already successful in many prepracticetrials and so showed no improvement on these measures, but they wereincluded in further analyses because they showed improvement on othermeasures of balancing.
159PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

of trials increased from 10.4 � 4.2 s to 17.0 � 1.8 s (single-sided t-test, P � 0.05).
Sample raw kinematic traces in the ML axis from oneparticipant before and after practice are shown in Fig. 1. It canbe seen that the finger and stick tip moved in the same directionfor most of the time, with the finger covering a wider rangethan the tip (Fig. 1, A and C). Most of the movements shownby these traces are slow changes in position that representtranslations of varying amplitude of the finger and stick tip inspace. Rotation of the stick is not readily apparent in theposition traces because rotation occurs only when there is achange in the relative positions of the finger and stick tip. The
difference between the positions of the finger and stick tip ateach time interval (Fig. 1, A and C) is directly related to theangular deviation from the vertical (Fig. 1, B and D). This ismost obvious when the finger and tip move in opposite direc-tions and can be seen more clearly in Fig. 1, C and D, wherethe translations are smaller than in Fig. 1, A and B. A period-icity at �1.7 Hz (�0.6 s per cycle) apparent in the rotationaldisplacements coincided with the MPF of stick angular veloc-ity shown by the spectral analyses.
The linear systems analyses indicated that 0.75 Hz was thefrequency boundary between translational and rotational mo-tion. Below 0.75 Hz, the finger and stick tip movements were
A
B
C
D
ML
Pos
ition
(m) -
PR
EM
L A
ngle
(deg
) - P
RE
Time (s)
ML
Ang
le (d
eg) -
PO
ST
ML
Pos
ition
(m) -
PO
ST
−0.1
0
0.1
0.2
0.3
0.4
0.5
−0.2
−8
−6
−4
−2
0
2
4
6
8
10
0 1 2 3 4 5 6 7 8 9 10−8
−6
−4
−2
0
2
4
6
8
10
−0.1
0
0.1
0.2
0.3
0.4
0.5
−0.2
Fig. 1. A 10-s sample of raw kinematic datain the mediolateral (ML) axis from 1 partic-ipant. A and C: finger (black line) and sticktip (gray line) position in laboratory coordi-nate system pre- and postpractice, respec-tively. B and D: stick angle pre- (Pre) andpostpractice (Post), respectively.
160 PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

highly coherent and the motion mostly comprised translationswhere the finger and stick tip moved approximately in parallel.The finger movements and the stick angle were less coherent atlow frequency because low-speed finger movement was noteffective in producing rotation of the stick. Above 0.75 Hz, thefinger movements and the stick angle were highly coherent andthe motion mostly comprised rotational movements where thefinger and stick tip movements diverged. Hence, the finger andstick tip movements were less coherent because almost allhigh-speed finger movements were coupled to stick rotation. Itcan be seen that the trajectory of the stick tip resembled asmoothed version of the trajectory of the finger (Fig. 1, A andC). This was confirmed by filtering the finger trajectory usinga low-pass, zero-phase filter (Butterworth 4th-order, dual-pass)with a cutoff frequency of 0.75 Hz. The filtered finger trajec-tory then very closely matched the trajectory of the stick tip.This result reflects the fact that it was translational motion ofthe finger that mostly determined both the translational motionof the stick tip and the change in angle of the stick.
Kinematics of Finger and Stick Movement
All of the kinematic measures showed that the magnitude ofmovement of both the stick and the finger decreased with
practice. Reductions in magnitude of one-third to one-halfwere observed: the normalized 3-D length of path of the stickand finger (Fig. 2), the RMS values and range of finger andstick tip movements (Fig. 3A), the RMS values of the angle ofdeviation of the stick from the vertical in ML and AP directions(Fig. 3B), the frequency distribution of the absolute angle ofdeviation of the stick from the vertical (Fig. 3C), and thecorrection angles at which the stick reversed its direction ofmovement to prevent it from falling (see Fig. 8). A diagram-matic representation of the 3-D path traveled by the stick tipand finger during pre- and postpractice trials is depicted for oneparticipant in Fig. 2A. The LoP of both the stick tip and finger(Fig. 2B) decreased significantly with practice [F(1, 8) �18.74, P � 0.05]. Irrespective of practice, however, the LoP forthe stick tip was substantially less than that of the finger [F(1,8) � 49.79, P � 0.05], and the ratio between the two did notchange significantly with practice (stick tip LoP/finger LoP:prepractice � 0.60 � 0.06 m/s, postpractice � 0.57 � 0.06m/s, single-sided t-test, P � 0.11), confirming that the stickpivoted more like a normal pendulum than an inverted pendu-lum. Note that the LoP is normalized by the total duration ofthe trial, and hence the reduction in LoP corresponds to areduction almost by half in the average velocity (m/s) of the
Stick Tip
Finger
PRE POST
00.1
0.20.3
0.4
0.20.1
1.0
1.1
1.2
1.3
1.4
1.5
1.6
00.1
0.20.3
0.4
0.20.1
1.0
1.1
1.2
1.3
1.4
1.5
1.6
Infe
rior
Sup
erio
r (m
)
Posterior Anterior (m)Medial
Lateral (m
)
Nor
m. L
oP (m
/s) *
*
0
0.1
0.2
0.3
0.4
0.5
0.6
Fing S-Tip
†PRE POST
A
B
Fig. 2. A: 3-dimensional (3-D) movement tracesof the stick and fingertip for 10 s from 1 partic-ipant. The total distances traveled by each of thestick and the finger markers represent the lengthof path (LoP) values. B: group mean and SD ofthe normalized (Norm.) LoP values for the fin-gertip (Fing) and stick tip (S-Tip) pre- and post-practice. Note that the normalized LoP also cor-responds to the average velocity of the stick tipand finger. *P � 0.05, significant difference withpractice. †P � 0.05, significant difference be-tween the finger and stick tip.
161PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

stick tip and finger with practice. This velocity change wassupported by a decrease with practice in the MPF for the fingerand stick tip position [F(1, 8) � 17.66, P � 0.05].
RMS and range of finger and stick tip movements. Changesin movement amplitude expressed as RMS displacement in thelaboratory space are shown in Fig. 3A. The mean amplitude ofthe fingertip and the stick tip movement in all directionsdecreased significantly with practice [F(1, 8) � 7.24, P �0.05]. For both finger and stick tip, the RMS amplitude in theV direction was on average only 24% of that in the ML and APdirections, and this ratio did not change with practice (single-sided t-tests, P � 0.20). The ranges (maximum–minimum) ofboth the finger and stick tip movement also decreased signif-icantly with practice in the ML (finger: 0.58 � 0.16 to 0.42 �0.17 m; stick tip: 0.47 � 0.12 m to 0.35 � 0.15 m), AP (finger:0.62 � 0.13 m to 0.49 � 0.15 m; stick tip: 0.49 � 0.09 m to0.41 � 0.13 m) and V (finger: 0.17 � 0.04 to 0.11 � 0.03 m;stick tip: 0.17 � 0.05 to 0.12 � 0.03 m) directions [F(1, 8) �6.58, P � 0.05]. The decreases in displacement and range of
the finger were greater than for the stick tip [F(1, 8) � 6.68,P � 0.05]. These differences reflect the fact that there was areduction in the magnitude of the stick angle with practice, asdescribed in the next section.
RMS values of stick angles. The RMS value of the stickangle in all directions decreased significantly with practice(Fig. 3B) [F(1, 8) � 5.77, P � 0.05]. It can be seen that theamplitude of the axial angle was substantially greater thanboth the ML and AP angles. Since the stick base contactedfirmly with the finger pad, the axial angle reflects thecomposite rotational movements of the wrist, shoulder,trunk, and hips (see Lee et al. 2012), but it contributesminimally to balance of the stick and did not changesignificantly with practice (single-sided t-tests, P � 0.60). Acomparison of the ML and AP angles shows that theamplitude of the stick angle was greater in the AP comparedwith the ML direction irrespective of practice [F(1, 8) �11.30, P � 0.01]. Both angles decreased with practice, butthe angle in the AP direction decreased more [F(1, 8) �
0
0.05
0.10
0.15
0.20
S-Tip Fing0
0.05
0.10
0.15
0.20
S-Tip Fing
Dis
plac
emen
t RM
S (m
)
PRE
POST
ML AP V
0
0.05
0.10
0.15
0.20
S-Tip Fing
* **
* *
A
B
0
2
4
6
8
ML AP
* *
05
1015202530
Axial
†
Stic
k A
ngle
RM
S (d
eg)
Stic
k A
ngle
RM
S (d
eg)
Deviation from the Vertical (deg)
% o
f Tot
al D
urat
ion
0
5
10
15
20
25
0 2 4 6 8 10 12 14 16 18 20
4.6°2.6° POST
PRE*
C
Fig. 3. A: group mean and SD of the fingertip andstick tip root-mean-square (RMS) displacement inthe ML, anteroposterior (AP), and vertical (V) axespre- and postpractice. B: group mean and SD of theRMS stick angle values pre- and postpractice: left,ML and AP angle; right, axial angle. Note that theaxial angle could be related to rotations of the wrist,shoulder, trunk, and/or hips. *P � 0.05, significantdifference with practice. †P � 0.05, significant dif-ference between ML and AP angles. C: group mean(thick lines) � SD (thin lines) of the time-normalizedhistogram of the angle of deviation of the stick fromthe vertical in the laboratory space before and afterpractice. Pre- and postpractice median angles areindicated in boxes next to the vertical lines. *P �0.05, significant difference with practice.
162 PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

6.30, P � 0.05] so that the ML and AP angles became closerin amplitude.
The angle of deviation of the stick from the vertical indicatesthe absolute magnitude of deviation without considerationof the direction of toppling. Changes with practice in thetime-normalized histograms of the deviation angle are shownin Fig. 3C. The median angle decreased significantly withpractice from 4.6 � 1.8° to 2.6 � 1.2° (single-sided t-test, P �0.05). The distribution pattern narrowed toward 0°, and thefrequency of occurrence of angles greater than �5° was re-duced almost by half.
Duration of half-cycle of stick oscillation. Perhaps the mostimportant finding with regard to the characteristic dynamics ofthe balanced stick was the duration of the stick sway (half)cycles. The zero-crossings of the positive and negative angularvelocity signals can be seen as the peaks and troughs of theangle signals in Fig. 1 (B and D). The mean intervals betweenzero-crossings for each toppling cycle are presented in Table 2.This simplest empirical measure showed no evidence ofchange with practice despite marked reductions in the magni-tude of sway (Figs. 1, 3B, and 3C) [F(1, 8) � 0.21, P � 0.66].Moreover, despite the clear differences in amplitude of swaybetween the AP and ML axes, especially before practice (Fig.3B), the sway durations were very similar for all directions oftoppling [F(1, 8) � 0.00, P � 0.96]. The mean duration of0.30 � 0.05 s across all conditions also coincided with thetheoretical half-cycle time derived from the pendulum modelof Lee et al. (2012). As shown by Lee et al. (2012), if this stickwere suspended as a physical pendulum at the radius ofgyration (rg) from its CoM and oscillated under the 4.53 (meank ratio) times greater-than-gravity torque exerted by the par-ticipants, then the general expression for the period of angularsimple harmonic motion of a pendulum predicts a period of0.60 s. The corresponding theoretical half-cycle duration of0.30 s therefore coincides with the measured values. Since aconstant period despite varying amplitude of oscillation is oneof the fundamental characteristics of a pendulum followingsimple harmonic motion, the cycle duration employed by theparticipants was consistent with angular simple harmonic mo-
tion of this stick as a physical pendulum operating undergreater-than-gravity torque.
Relative Coordinates
An example of the time-normalized histogram of the top-pling angle in the MLrel-APrel plane from one participant pre-and postpractice is depicted in Fig. 4A. It can be seen from thelength of the major and minor axes of the ellipse fitted to eachhistogram that irrespective of practice, the stick toppled morefrequently in the APrel direction than in the MLrel direction.With practice, a decrease in the frequency of toppling in theAPrel direction can be observed. It can also be seen that thecentroid of the ellipse was positioned more posteriorly preprac-tice than postpractice, indicating that the stick fell posteriorlymore often than anteriorly before practice. After practice,however, a more even distribution in the APrel direction can beidentified from a shift in the centroid (prepractice: 0.01%medial, 0.78% posterior; postpractice: 0.02% medial, 0.22%posterior). The group mean and SD of the features of theellipses are illustrated in Fig. 4B. There was a significantreduction with practice only in the major axis [F(1, 8) � 5.71,P � 0.05], whereas the length of the minor axis did not showany change (Tukey post hoc test: P � 0.99). The frequency oftoppling in the APrel direction was over three times greater thanin the MLrel direction before practice, but after practice it haddecreased to about twice the frequency in the MLrel direction.The tilt angle of the major axis of the ellipse and its centroidlocation in the MLrel direction did not change significantly withpractice (single-sided t-tests, P � 0.29). However, the relativefrequency of anterior-to-posterior compared with posterior-to-anterior toppling was reduced considerably, as indicated by ashift in the centroid location anteriorly with practice (single-sided t-test, P � 0.05).
Kinetics of Finger and Stick Movement
In parallel with the kinematic changes, the kinetic measuresshowed corresponding reductions in the forces and torquesapplied between the finger and stick (Fig. 5).
Net torque and finger reaction force. The average amplitudeof net torque in the three planes of motion (Fig. 5) reducedsignificantly with practice [F(1, 8) � 17.53, P � 0.05]. Similarto the results for the amplitude of the stick angle, the net torquewas greater in the AP compared with the ML direction irre-spective of practice [F(1, 8) � 6.24, P � 0.05], and the torquein the AP direction decreased more with practice [F(1, 8) �5.52, P � 0.05]. Although the amplitude of the axial torquewas extremely small compared with the ML and AP torques, italso decreased significantly with practice (single-sided t-test,P � 0.05). The average amplitude of the finger reaction forcealso decreased with practice in all three directions [F(1, 8) �16.27, P � 0.05]. Post hoc Tukey tests of a significantdifference across the three axes [F(1, 8) � 6.77, P � 0.05]revealed a significantly greater force in the AP compared withthe ML direction in prepractice but not postpractice trials.
Frequency Domain Analyses
Crossed-axes analysis: finger position (input) and stick tip(output). The overall coherence between crossed axes de-creased with practice (Fig. 6) [F(1, 8) � 13.74, P � 0.05].There was no difference between the two directions of the
Table 2. Sway half-cycle durations and k ratios with theoreticalsway half-cycle duration
Toppling Cycles
Sway duration, s k Ratio
Prepractice Postpractice Prepractice Postpractice
M¡Ltopl 0.31 � 0.06 0.29 � 0.04 4.4 � 0.3 4.7 � 0.6M¢Ltopl 0.29 � 0.05 0.32 � 0.05 4.6 � 0.3 4.8 � 0.5A¡Ptopl 0.31 � 0.03 0.30 � 0.07 4.3 � 0.5 4.6 � 0.5A¢Ptopl 0.29 � 0.06 0.31 � 0.03 4.3 � 0.4 4.6 � 0.4Theoretical duration
from mean k ratio 0.30
Data are group mean (�SD) sway half-cycle durations and k ratios of themagnitudes of the active torque applied by the participants and the torque onthe stick due to the gravity for M¡L, M¢L, A¡P, and A¢P topplingdirections pre- and postpractice. The theoretical sway half-cycle durationderived from the pendulum model of Lee et al. (2012) is also shown. The ratio(k) between the magnitudes of the active torque and the gravitational torquedetermines both the sway cycle period and the pivot point of the pendulum.The ratio for each participant was calculated from the slope of the meantorque-angle line for each toppling direction relative to the slope of additiveinverse of the gravitational torque-angle line.
163PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

cross talk between axes (finger ML vs. stick tip AP and fingerAP vs. stick tip ML values) [F(1, 8) � 0.02, P � 0.88]. Thechange in coupling between axes with practice represented aone-third reduction in movements of the finger which producederroneous off-axis movements of the stick rather than correct-ing the stick angle.
Characteristic Dynamics of the Stick
The performance improvement with practice did not resultfrom changes in the dynamics between the finger and stickmovement, because there was no change in the characteristicdynamics of the stick oscillations, as shown by the mean
torque-angle relations (Fig. 7) and the phase portrait of therotational dynamics (Fig. 8A). These findings again support themodel of the observed balancing behavior of the stick asconsistent with angular simple harmonic motion of a physicalpendulum (Lee et al. 2012).
Torque-angle relationship. The characteristic oscillatory cy-cle of the stick, unaffected by cycle-to-cycle variations intranslation, is shown by the mean relationship between active
1%
2%
3%
4%
30°
210°
60°
240°
(A)90°
270°(P)
120°
300°
150°
330°
180° (L)
0°(M)
%%111%%1%%%%1%1 1%
2%
3%
4%
30°
210°
60°
240°
(A)90°
270°(P)
120°
300°
150°
330°
180° (L)
0°(M)
%%11%11%%%%%1%1
α: 2.4°, [M: 0.01 %, P: 0.78 %]Major: 2.4 %, Minor: 0.9 %
α: 14.9°, [M: 0.02 %, P: 0.22 %]Major: 1.7 %, Minor: 1.0 %
-1.5
-1
-0.5
0
0
1
2
3
4
Mj Mn
5
10
15
*
Cen
troid
Loc
atio
n of
Elli
pse
(%)
Leng
th o
f Sem
i-axe
s (%
)
Tilt
Ang
le o
f Elli
pse
(deg
)
(M) (A)
(L) (P)
0
PRE POSTA
B
*
PRE
POST
Fig. 4. A: time-normalized histograms of thetoppling angle in the relative coordinate spacewith the ellipse fitted to the pre- and post-practice histograms from 1 participant. Major,length of major axis of the ellipse; minor, lengthof minor axis of the ellipse; and �, tilt angle ofthe major axis of the ellipse in reference to theAPrel axis (positive values indicate clockwise di-rection). The coordinates and direction of thecentroid of the ellipse are shown in square brack-ets. B: group mean and SD of the features of theellipses fitted to the time-normalized histo-grams of the toppling angle: left, length ofsemi-axes (Mj, major; Mn, minor); middle: tiltangle of the major axis in relation to the APrel
axis; right, location of the centroid. M, medial;L, lateral; A, anterior; P, posterior. *P � 0.05,significant difference with practice.
PRE POST
Net
Tor
que
RM
S (N
m)
Fing
er R
x. F
orce
RM
S (N
)
0
0.005
0.010
0.015
0.020
0.025
ML AP0
2.0
4.0
6.0
Axial0
0.02
0.04
0.06
ML AP V
x 10-6
**
*
*
**
†
Fig. 5. Group mean and SD of the RMS net and axial torque (left) and fingerreaction force (right) in all directions. Note the difference in scale for axialtorque. *P � 0.05, significant difference with practice. †P � 0.05, significantdifference between ML and AP torque.
PRE
POST
* *
00.050.100.150.200.250.300.350.400.45
Fing MLS-Tip AP
Fing APS-Tip ML
Ove
rall
Coh
eren
ce
Fig. 6. Changes with practice in group mean overall coherence betweencrossed axes of the finger and the stick tip movement. Left, ML finger position(input) and AP stick tip position (output); right, AP finger position (input) andML stick tip position (output). *P � 0.05, significant difference with practice.
164 PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

torque and angle for each toppling cycle (Fig. 7). The additiveinverse of the gravitational torque indicates the minimummagnitude of active torque that the participant must generate toprevent the stick from falling under gravity. The ratio betweenthe magnitude of the active torque and the gravitational torqueis termed the “k ratio” (Lee et al. 2012), which represents theparticipant’s strategy to control the rotational dynamics of thestick. The mean ratios ranged from 4.3 (�0.4) to 4.8 (�0.5)and did not vary significantly with either practice [F(1, 8) �2.70, P � 0.14] or toppling direction (e.g., M¡L vs. M¢L)[F(1, 8) � 1.84, P � 0.21] (Table 2). Hence, the participantsutilized only a narrow range of (greater-than-gravity) torques torotate the stick. On the other hand, the ratio for the ML axis(4.64 � 0.45) was slightly greater than that for the AP axis(4.43 � 0.47) irrespective of practice [F(1, 8) � 5.55, P �0.05]. It can be seen that the excursion of the torque and angletraces was reduced after practice, reflecting smaller oscillationsof the stick, and was slightly more linear and closely centeredabout the origin (0, 0). Regardless of these changes with
practice, however, the slopes of the torque-angle lines re-mained almost constant and indicate that the stick behavior wassimilar to a mass-spring system where the displacement of thespring is proportional to the magnitude of the force applied inthe opposite direction.
Phase portrait of the rotational dynamics. The phase por-traits similarly decreased their excursion but showed littlequalitative change with practice other than the trajectoriesbecoming more symmetrical and more congruent with thetheoretical pendulum model. Time-normalized and averagedrelationships between angular velocity and angle are depictedin Fig. 8A to generate an average phase portrait of the stickdynamics in both the ML and AP axes. The direction of thearrow for each trajectory indicates the toppling cycle direction.Similar to normal pendulum dynamics, when the angularvelocity reaches its peak, the stick becomes upright (i.e., itstoppling angle reaches zero) and when the magnitude of thestick angle becomes maximum, as indicated by the verticaldashed lines in the figure, the stick stops rotating (i.e., its
-5 -4 -3 -2 -1 0 1 2 3 4 5
-0.02
0
0.02
-5 -4 -3 -2 -1 0 1 2 3 4 5
-0.02
0
0.02
AP Angle (deg)ML Angle (deg)
Act
ive
torq
ue (N
m)
PRE POST
M L toppling
M L toppling
A P toppling
A P toppling
0.01
-0.01
0.01
-0.01
Fig. 7. Time-normalized prepractice (gray) andpostpractice (black) active torque vs. angle rela-tionships for M¡L and M¢L (left) and A¡Pand A¢P (right) toppling cycles, averagedacross participants for all trials. The straight graylines show the gravitational torque line, whereasthe straight dashed black lines show its additiveinverse. Lateral and posterior angles are posi-tive; medial and anterior angles are negative.
AP Angle (deg)ML Angle (deg)
Ang
ular
Vel
ocity
(deg
/sec
)
M L toppling
M L toppling
A P toppling
A P toppling
-5 -4 -3 -2 -1 0 1 2 3 4 5
-50
-25
0
25
50
-5 -4 -3 -2 -1 0 1 2 3 4 5
-50
-25
0
25
50
0 2 4 6-2-4-6
PRE
POSTM
A
correction angle: -2.5 degA
B
Correction Angle (deg)
*
*
*
*
L
P
M L A P
Fig. 8. A: angular velocity vs. angle relation-ship for the ML and AP axes for prepractice(large continuous curves) and postpractice(small continuous curves) balancing, aver-aged across participants for all trials. Theblack curves indicate M¡L and A¡P top-pling, whereas the gray curves indicate M¢Land A¢P toppling. The negative and positivecorrection angles are indicated by the verticaldashed lines. The black dashed ellipses ineach plot show the phase portrait of the stickpivoting at the radius of gyration (rg) from thecenter of mass for the observed mean correc-tion angles. The background gray dashedlines show these phase portraits for a range ofdifferent values of the correction angle. B:group mean and SD of the negative and pos-itive correction angles pre- and postpractice.*P � 0.05, significant difference with prac-tice.
165PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from
angular velocity reaches zero). These maximum values of theangles in the cycle are defined as the correction angles wherethe stick trajectory away from the vertical is halted and re-versed. The black dashed lines in Fig. 8A show the theoreticalpendulum dynamics from Lee et al. (2012) for the stickpivoting at the rg from the CoM for the mean pre- andpostpractice correction angles. The background gray dashedlines show these theoretical phase portraits for a range ofcorrection angles larger and smaller than those observed. Theobserved mean trajectories decreased in size with practice andbecame more congruent with the model. The trajectories werealso shifted laterally (ML) and posteriorly (AP) before practicebut became symmetrical with practice.
Both the ML and AP correction angles decreased signifi-cantly with practice [F(1, 8) � 5.55, P � 0.05] (Fig. 8B). TheAP correction angles were larger than the ML angles irrespec-tive of practice [F(1, 8) � 6.32, P � 0.05] but reduced morewith practice [F(1, 8) � 5.49, P � 0.05], as confirmed by apost hoc Tukey test, so that the angles became closer inamplitude. There were also greater reductions with practice inM¡L than M¢L toppling and in A¡P than A¢P toppling[F(1, 8) � 22.32, P � 0.05], as confirmed by post hoc Tukeytests. These effects are in accordance with the improvements insymmetry in the phase portraits.
Pendulum length. Support for the pendulum model was alsoprovided by the PL values. Using the mean k ratios measuredfor each participant for each toppling direction, the PL valuesbefore and after practice were calculated via the model of thebalanced stick as angular simple harmonic motion of a normalpendulum (Lee et al. 2012). The notable finding here again wasthe lack of change with practice in the PL values, with mean(�SD) pre- and postpractice values for the four directions ofstick toppling all within a very narrow range of 0.40–0.42(�0.01–0.02) m [F(1, 8) � 2.70, P � 0.14]. Moreover, a PLof 0.41 m placed the pivot point of the pendulum at the rg fromthe CoM of the stick.
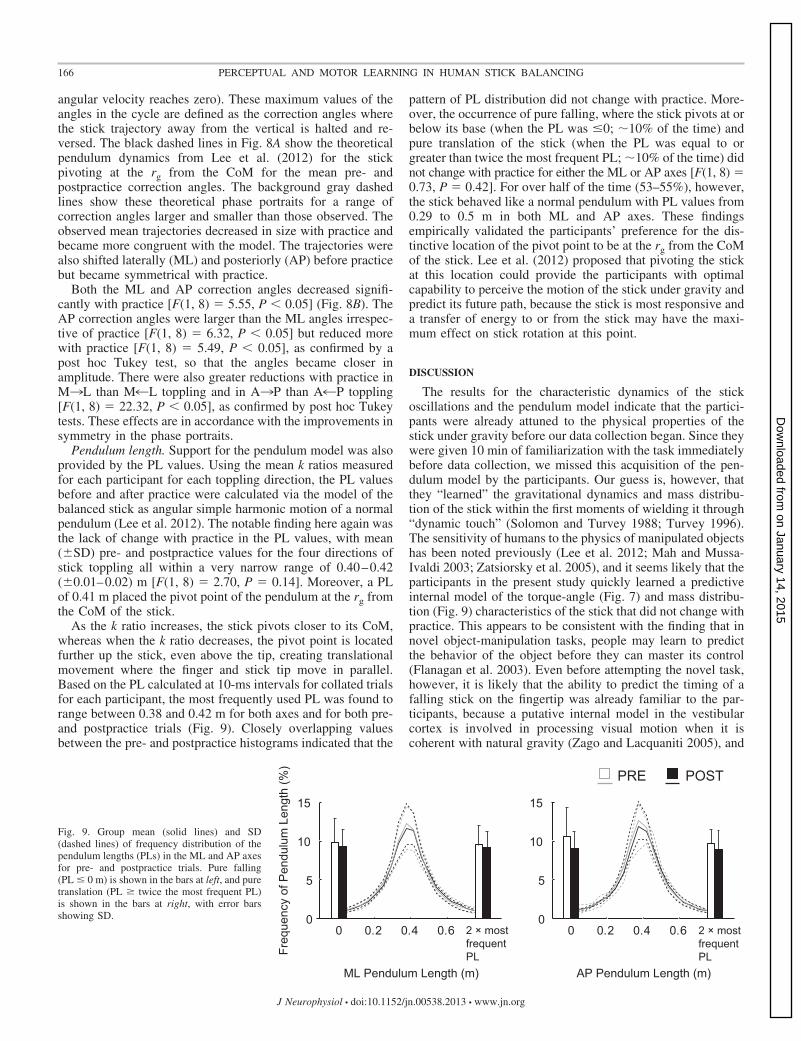
As the k ratio increases, the stick pivots closer to its CoM,whereas when the k ratio decreases, the pivot point is locatedfurther up the stick, even above the tip, creating translationalmovement where the finger and stick tip move in parallel.Based on the PL calculated at 10-ms intervals for collated trialsfor each participant, the most frequently used PL was found torange between 0.38 and 0.42 m for both axes and for both pre-and postpractice trials (Fig. 9). Closely overlapping valuesbetween the pre- and postpractice histograms indicated that the
pattern of PL distribution did not change with practice. More-over, the occurrence of pure falling, where the stick pivots at orbelow its base (when the PL was �0; �10% of the time) andpure translation of the stick (when the PL was equal to orgreater than twice the most frequent PL; �10% of the time) didnot change with practice for either the ML or AP axes [F(1, 8) �0.73, P � 0.42]. For over half of the time (53–55%), however,the stick behaved like a normal pendulum with PL values from0.29 to 0.5 m in both ML and AP axes. These findingsempirically validated the participants’ preference for the dis-tinctive location of the pivot point to be at the rg from the CoMof the stick. Lee et al. (2012) proposed that pivoting the stickat this location could provide the participants with optimalcapability to perceive the motion of the stick under gravity andpredict its future path, because the stick is most responsive anda transfer of energy to or from the stick may have the maxi-mum effect on stick rotation at this point.
DISCUSSION
The results for the characteristic dynamics of the stickoscillations and the pendulum model indicate that the partici-pants were already attuned to the physical properties of thestick under gravity before our data collection began. Since theywere given 10 min of familiarization with the task immediatelybefore data collection, we missed this acquisition of the pen-dulum model by the participants. Our guess is, however, thatthey “learned” the gravitational dynamics and mass distribu-tion of the stick within the first moments of wielding it through“dynamic touch” (Solomon and Turvey 1988; Turvey 1996).The sensitivity of humans to the physics of manipulated objectshas been noted previously (Lee et al. 2012; Mah and Mussa-Ivaldi 2003; Zatsiorsky et al. 2005), and it seems likely that theparticipants in the present study quickly learned a predictiveinternal model of the torque-angle (Fig. 7) and mass distribu-tion (Fig. 9) characteristics of the stick that did not change withpractice. This appears to be consistent with the finding that innovel object-manipulation tasks, people may learn to predictthe behavior of the object before they can master its control(Flanagan et al. 2003). Even before attempting the novel task,however, it is likely that the ability to predict the timing of afalling stick on the fingertip was already familiar to the par-ticipants, because a putative internal model in the vestibularcortex is involved in processing visual motion when it iscoherent with natural gravity (Zago and Lacquaniti 2005), and
0 0.2 0.4 0.60
5
10
15
0 0.2 0.4 0.60
5
10
15
ML Pendulum Length (m) AP Pendulum Length (m)
Freq
uenc
y of
Pen
dulu
m L
engt
h (%
)
PRE POST
2 × most frequent PL
2 × most frequent PL
Fig. 9. Group mean (solid lines) and SD(dashed lines) of frequency distribution of thependulum lengths (PLs) in the ML and AP axesfor pre- and postpractice trials. Pure falling(PL � 0 m) is shown in the bars at left, and puretranslation (PL � twice the most frequent PL)is shown in the bars at right, with error barsshowing SD.
166 PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

such a gravity model (internalized Newton’s laws) wouldoperate from the earliest stages of learning. Miller et al. (2008)have proposed that an early stage of integration of high-levelvisual analysis with gravity-related motion information occursin the vestibular nuclei and posterior cerebellar vermis regions,which are reciprocally connected with the vestibular cortex.
Notwithstanding these issues of how and when they learnedthe physics of the stick, however, it is clear from the initialrecorded trials that the pendulum model, although presumablynecessary, was not sufficient to enable the participants tobalance the stick (see Table 1). This suggests, in line with thefindings of Flanagan et al. (2003), that the participants tookmuch longer to fine-tune the inverse dynamics model that isrequired to stabilize and control the stick. The question thenarises, how did they learn to stabilize the stick and reduce itsmotion? The mechanisms underlying this acquisition of skillwill now be explored.
Mechanisms of Learning
The trajectory of motion of the stick when it falls undergravity is familiar and predictable from the moment the motionbegins, but the precise moment and direction of falling areunpredictable. Oscillating the stick like a pendulum followingsimple harmonic motion imposes a significant degree of pre-dictability on its rotational trajectory, even despite the super-imposed translations of varying magnitude and direction. Thisoscillatory strategy, evident from the first trials, maintained thesame duration (0.605 s) and periodicity (1.65 Hz) of the swaycycle throughout practice as the magnitude of stick motiondecreased, thereby almost halving the average velocity of stickmotion (Fig. 2B). A more slowly translating stick is easier tostabilize, as evidenced after practice by the much smallercorrection angles, which identify where the stick trajectoryaway from the vertical was halted and reversed (Fig. 8). Slowermovements are likely to reduce the uncertainty in signals ofmotor output, efference copy, and afferent feedback comparedwith fast movements because of the effects of fixed feedbackdelays in afferent input, slower changes of signals with time,and lower signal-dependent noise (Gritsenko et al. 2007).Combined with the constant rotational period, this slowing ofthe translational motion may explain why no change wasnecessary, or detected, in the timing relations between move-ment of the finger and the stick to improve balancing skill.Hence, no direct evidence of a change in ability to predict themotion of the stick was observed. Again, however, this begsthe question of how the participants learned to slow andstabilize the motion of the stick.
The motion of the stick is determined by the active torqueapplied by the participant and the torque due to gravity. Themagnitude ratio between these torques, the k ratio, sets thesway cycle period, as well as the pivot point of the pendulum(Lee et al. 2012). Greater-than-gravity torque reduces theperiod of a pendulum relative to that under gravitational torquealone. Any increase or decrease in active torque applied by theparticipants, and hence in the value of k, would respectivelydecrease or increase the period and shift the most frequentpivot point away from its location at the rg from the CoM. Thisunderscores the importance of the sensitivity to the torque dueto gravity (Zatsiorsky et al. 2005) in balancing the stick.Consequently, Lee et al. (2012) proposed that the k ratio is the
major control input parameter in this task. The three parame-ters, k ratio, cycle period, and pivot point location, were heldconstant irrespective of practice. This was the most surprisingfinding of the study, raising the question, how did the subjectsimprove their skill without a change in the rotational dynam-ics? The answer lies in the finding that although the slope ofthe torque-angle relation was unchanged, the amplitude of thesignals was substantially reduced (Figs. 7 and 8). Hence, thetorque-angle gain was unchanged, but the torque and anglesignals were smaller. This demonstrates that the motor com-mands changed with practice. Smaller torques will mean lowersignal-dependent noise (Harris and Wolpert 1998; Meyer et al.1988; Schmidt et al. 1979), whereas smaller and slower move-ments point to more precise perceptual signals (Gritsenko et al.2007). From this starting point, two factors of perception andaction can be identified that were correlated with increasedbalancing skill.
Perceptual Factors
The toppling angle of the stick as measured with respect tothe markers on the participants’ forehead provides an indica-tion of how the stick moved relative to the participants’ visualfield. The time-normalized histograms of this toppling angleshow the relative amount of time spent at each topplingdirection, independent of its magnitude (Fig. 4). The histo-grams revealed that before practice, the stick toppled threetimes more frequently in the APrel direction than the MLreldirection. Since the precision of human detection of motion indepth (the AP direction) is about half of that in the mediolateral(ML) direction (Sumnall and Harris 2002; Tyler 1971; Wes-theimer 1990), the participants could be expected to makemore errors and spend more time correcting errors in the lessaccurately perceived direction, thereby creating the patternbefore practice shown in Fig. 4.
A substantial decrease with practice was observed in thelength of the major axis of the ellipse fitted to the histogramsof the stick toppling angle so that the frequency of toppling inthe APrel direction was then only about twice that in the MLreldirection. This ratio after practice matches the reported relativeprecision of human detection of motion in depth versus themediolateral direction (Sumnall and Harris 2002; Tyler 1971;Westheimer 1990). The reduction in the length of the majoraxis, therefore, is consistent with perceptual learning of depthdiscrimination, and it has been shown that learning of depthperception can indeed occur with practice (Gantz et al. 2007;O’Toole and Kersten 1992; Ramachandran 1976). Further-more, �50% of the variance of the measures of balancingperformance could be accounted for by their correlation withthis perceptual measure (Table 3). Hence, better visual discrim-ination of depth was associated with more skilled balancingperformance. The corollary of this association is that lesssensitive visual perception of the stick toppling in depth islikely to be a primary deficit in the unskilled balancer.
Another significant change in the time-normalized histo-gram of the toppling angle was the shift in the centroid locationof the ellipse in the APrel direction (Fig. 4B). Whereas thefrequency of M¡Lrel and M¢Lrel stick toppling was symmet-rical irrespective of practice, A¡Prel toppling occurred signif-icantly more often than A¢Prel toppling before practice. Thischange could have been due to the proprioceptive localization
167PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

ability of the hand in the depth direction. Van Beers et al.(1998) studied the precision of proprioceptive position senseand found that as the hand moved further away from theshoulder, their participants’ localization ability decreased ac-cordingly (i.e., the ability to pinpoint the location of the handin space without the aid of vision). Since the finger is furtheraway from the body during A¡Prel stick toppling (and thevisual attention of the participant is focused primarily on the tipof the stick so that they cannot simultaneously see the finger),it would therefore be expected that the movement would not beas accurate as during A¢Prel toppling, when the finger iscloser to the body. The reduced proprioceptive accuracy indepth would contribute to more errors in the A¡Prel topplingdirection, resulting in the stick toppling in that direction moreoften and leading to the pattern of asymmetry seen in Fig. 4B.With practice, the toppling frequency in the APrel directionbecame far more symmetrical, although a slight degree ofasymmetry remained. As with visual discrimination of depth,therefore, proprioceptive localization in depth may have im-proved with practice, consistent with the observed reduction inasymmetry. Furthermore, the centroid location in the AP axiswas correlated with both the duration of successful trials andthe 3-D length of path of the finger (Table 3). Hence, moreskilled balancing performance also appears to be associatedwith better proprioceptive discrimination of depth. The impor-tance of proprioceptive learning in stick balancing similarlywas emphasized by Cabrera and Milton (2012), who noted thatthe observation that expert stick balancers adopt a character-istic posture, namely, elbow held in flexion with wrist andindex finger extended in line with the forearm, suggests thatproprioceptive information from the shoulder contributes toskill.
The question whether perceptual learning also occurred inthe ML axis with practice must be considered. The histogramsof the toppling angles in the relative coordinate space (Fig. 4)were time normalized and so provided information on relativerather than absolute frequency of the directions of stick top-pling. The lack of change in the minor axis of the fitted ellipsetherefore does not necessarily mean that there was no reductionin stick toppling in the ML axis with practice. In fact, thecorrection angle in the ML axis reduced by close to one-halfwith practice (Fig. 8B). That perceptual learning is likely tohave contributed to this reduction is shown by recent demon-strations of perceptual learning in vision, audition, somatosen-
sation, and olfaction (see Fahle 2005 for review). In vision,perceptual features of widely differing complexity can belearned, ranging from straightforward orientation discrimina-tion to detecting complex patterns (Fahle 2005; Fine andJacobs 2002). Furthermore, even early sensory cortices, con-ventionally considered as hard wired, can be involved in theunderlying changes of the central nervous system (Fahle 2005;Paz et al. 2004). Perceptual learning improves performance innearly all tasks investigated so far, is often quite specific for theexact task trained, and does not lead to conscious insights thatcan be easily communicated; hence, it is of the procedural orimplicit type (Fahle, 2005), consistent with the phenomenologyof stick balancing. In light of this research, it seems likely thatperceptual learning did also occur in the ML axis.
Motor Factors
Cross talk between the orthogonal ML and AP axes of fingerand stick tip movements (finger ML vs. stick tip AP, finger APvs. stick tip ML) represents an important source of error incontrol of the stick (Fig. 6). Consider for instance what hap-pens if the stick topples toward the participant in the A¡Pdirection and the participant produces an opposing movementthat is not wholly within the V � AP plane of movement thatwould return the stick directly to the vertical, but insteaddeviates slightly from the AP direction. The inaccurate oppos-ing movement will oppose toppling in the direction of theopposing movement, and this may return the stick partiallytoward the vertical, but it will now deviate from the vertical ina new direction due to the out-of-plane or “off-axis” error. Thisdeviation must then be corrected, and if further error is not tobe created, the corrective movement again must remain whollywithin the plane that returns the stick to the vertical. Hence,skillful balancing depends on precisely accurate stick move-ments that do not cause the stick to move in off-axis directionsbetween the ML and AP axes of finger and stick tip movement.Conversely, the process of unskilled balancing can be viewedas a vicious cycle where cross talk between orthogonal axes offinger and stick tip movement result in an accumulation oferrors, causing a progressive increase in the toppling angle thatleads to increased movement, at increased speed, of the fingerand stick. Since a larger toppling angle entails both a largergravitational torque on the stick and less time to avert a fall, theparticipants must generate more torque more rapidly on the stickto maintain balance. These factors will amplify the inaccuracy ofcorrective movements due to signal-dependent noise and exacer-bate the error in accordance with the speed-accuracy trade-off(Carlton and Newell 1993; Fitts 1954; Harris and Wolpert 1998;Meyer et al. 1988; Schmidt et al. 1979), hence the vicious cycle ofunskilled balancing.
The reductions with practice in overall coherence betweenorthogonal axes of finger and stick tip movements (Fig. 6)therefore reflect a reduction in movement error such that theML direction of finger movement became less coupled with theAP direction of the stick tip movement, and vice versa. More-over, this measure of cross talk between orthogonal axes washighly correlated with both the duration of successful trials andthe 3-D LoP of the finger (Table 3). These correlations showthat this source of error was closely linked to successfulbalancing performance and suggest that reduced cross talkcontributed to increased balancing skill.
Table 3. Relation of stick-balancing performance with perceptualand motor measures
Length of APrel
Axis of Ellipse
CentroidLocation inAPrel Axis
Cross TalkBetween ML and
AP axes
Balancing duration 0.44 0.44 0.723-D finger LoP 0.54 0.58 0.54
Data indicate variance [r2: F(1, 14) � 10.68, P � 0.05] of pre- andpostpractice measures of stick-balancing performance (duration, normalizedlength of finger path) accounted for by their relation with perceptual measures(length of APrel axis, centroid location in APrel axis) and motor measures (crosstalk: mean of coherences for finger ML vs. stick tip AP and for finger AP vs.stick tip ML). Note that an ellipse could not be fitted to the toppling angle datafor participant 5 due to the small number and short duration of successfulprepractice trials, so the participant number is reduced to 8 (pre- and postprac-tice combined � 16) for these analyses.
168 PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

Perceptual-Motor Interactions
We propose therefore that the mechanisms underlying theacquisition of skill in balancing a stick comprise, first, percep-tual learning, which provides the participant with more accu-rate visual localization of the stick and proprioceptive local-ization of the finger, and second, motor learning of the direc-tional accuracy of finger movements, which reduces off-axiserrors in finger movements. Perceptual learning and motorlearning alike will improve both the precision and direction ofmovement, leading to more accurate action matched to moreaccurate perception, the major benefit of which is the avoid-ance of accumulating errors that lead to loss of stability. Thisproposal is supported by recent evidence that motor learningand perceptual learning are linearly correlated (Vahdat et al.
2011). Vahdat and colleagues identified a network involvingsecond somatosensory cortex, ventral premotor cortex, andsupplementary motor cortex whose activation was specificallyrelated to perceptual learning, and a network comprising cer-ebellar cortex, primary motor cortex, and dorsal premotorcortex that was linked to motor learning. Subjects who showedgreater changes in functional resting-state connectivity relatedto motor learning showed greater changes related to perceptuallearning.
Successful balancing is determined by motion of the stick inboth the ML and AP directions (Fig. 10A). Therefore, measuresof perception of stick motion in the AP direction alone wouldnot be expected to account for all of the variance in perfor-mance measures (cf., Table 3). However, the potential forinteraction between the ML and AP axes in balancing makes
L M
PP
A
L M
PP
A
L M
PP
A
Finger action in the correct plane
Inaccurate perception of stick tip location and its coupled finger action
F Resultant motion of stick tip due to inaccurate perception or action of the finger
L M
PP
A
Inaccurate action of the finger with correct perception of stick tip location
PAL M
Motion of stick tip in ML axis Motion of stick tip in AP axis
A
B C
D E
Fig. 10. Schematic illustration of hypothetical percep-tual and motor errors. All panels depict motion rela-tive to the finger. A: stick tip falls in M¡L and A¢Pdirections; the actual plane of falling is shown byshaded semicircle in A–C. Accurate corrective actionof finger (filled arrow) will result in corrective motionof stick tip (open arrow) within plane of falling. B:hypothetical inaccurate perception of location of sticktip results in corrective action of finger (filled arrow)in M¡L direction only. C: hypothetical accurate per-ception but inaccurate movement execution results incorrective action of finger (filled arrow) in M¡Ldirection only. D: from B and/or C, action of finger inM¡L direction only will result in motion of stick tipin M¢L direction only (open arrow). E: from Band/or C, stick motion in A¢P direction is uncor-rected, and hence stick tip will fall further in A¢Pdirection (open arrow). F: resultant motion of stick tip(open arrow) is outside plane of falling (shown bylight shaded semicircle with dashed line) and createscross-coupling between ML and AP axes.
169PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

the contribution to performance from perception of depthdifficult to estimate. Errors in perception create inaccurateestimates of the location of the hand or stick tip in space. Suchinaccurate estimates will often result in off-axis errors in thecorrective finger movements, thereby affecting movement inboth axes, as illustrated in Fig. 10, B and D–F. Similarly, motorexecution errors will often result in off-axis errors, therebyaffecting movement in both axes, as illustrated in Fig. 10, C–F.Hence, perceptual (visual and proprioceptive) and motor errorsinteract in their effect on balancing performance, and so bothtypes of error contribute to cross talk between orthogonal axesof finger and stick tip movement. A similar situation applies incontrol of human standing, where a majority of studies haveshown that sensorimotor noise or inappropriate torque modu-lation is the primary source of standing sway (Kiemel et al.2011; Loram et al. 2001; Loram and Lakie 2002; Maurer andPeterka 2005; Mergner et al. 2002). Increased skill in stickbalancing therefore depends on both perceptual and motorlearning, and recent research suggests that similar neuronalmechanisms may govern learning across sensory and motordomains (Paz et al. 2004; Poggio and Bizzi 2004). As noted byPaz et al. (2004), improved encoding of information is equallyimportant for motor and sensory systems, because the signalsgenerated from both systems must be shared with other brainareas in controlling movement. In the present study, it was notthe internal model of the rotational dynamics that changed withpractice but the precision of the signals flowing through themodel. The model was not sufficient without precisely accuratesensory and motor signals, which depend on learning.
Finally, it should be noted that the findings of the presentstudy regarding perceptual learning of depth discriminationcannot be applied where the stick-balancing tasks employedare in one (mediolateral) dimension (e.g., Foo et al. 2000;Reeves et al. 2013; Treffner and Kelso 1999) or virtual (e.g.,Mehta and Schaal 2002). Since visual and proprioceptivesignals of depth are not available to the participant in suchapproaches, they can have no role in the acquisition of balanc-ing skill.
GRANTS
This work was supported in part by Sport Knowledge Australia.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
K.-Y.L., N.O., M.H., and R.S. conception and design of research; K.-Y.L.and M.H. performed experiments; K.-Y.L., N.O., and M.H. analyzed data;K.-Y.L., N.O., M.H., and R.S. interpreted results of experiments; K.-Y.L. andN.O. prepared figures; K.-Y.L. drafted manuscript; K.-Y.L., N.O., M.H., andR.S. edited and revised manuscript; K.-Y.L., N.O., M.H., and R.S. approvedfinal version of manuscript.
REFERENCES
Ada L, O’Dwyer NJ, Neilson PD. Improvement in kinematic characteristicsand coordination following stroke quantified by linear systems analysis.Hum Mov Sci 12: 137–153, 1993.
Barto AG, Sutton RS, Anderson CW. Neuronlike adaptive elements that cansolve difficult learning control-problems. IEEE Trans Syst Man Cybern 13:834–846, 1983.
Bendat JS, Piersol AG. Random Data: Analysis and Measurement Proce-dures. Hoboken, NJ: Wiley InterScience, 1986.
Bizzi E, Kalil RE, Morasso P. Two modes of active eye-head coordination inmonkeys. Brain Res 40: 45–48, 1972.
Cabrera JL, Milton JG. On-off intermittency in a human balancing task.Phys Rev Lett 89: 1–4, 2002.
Cabrera JL, Milton JG. Human stick balancing: tuning Levy flights toimprove balance control. Chaos 14: 691–698, 2004a.
Cabrera JL, Milton JG. Stick balancing: on-off intermittency and survivaltimes. Nonlinear Stud 3: 305–317, 2004b.
Cabrera JL, Milton JG. Stick balancing, falls and Dragon-Kings. Eur PhysJ Spec Top 205: 231–241, 2012.
Carlton LG, Newell KM. Force variability and characteristics of forceproduction. In: Variability and Motor Control, edited by Newell KM,Corcos DM. Champaign, IL: Human Kinetics, 1993, p. 15–36.
Cluff T, Balasubramaniam R. Motor learning characterized by changingLevy distributions. PLoS One 4: 1–7, 2009.
Cluff T, Boulet J, Balasubramaniam R. Learning a stick-balancing taskinvolves task-specific coupling between posture and hand displacements.Exp Brain Res 213: 15–25, 2011.
Cluff T, Manos A, Lee TD, Balasubramaniam R. Multijoint error compen-sation mediates unstable object control. J Neurophysiol 108: 1167–1175,2012.
Cluff T, Riley MA, Balasubramaniam R. Dynamical structure of handtrajectories during pole balancing. Neurosci Lett 464: 88–92, 2009.
Connell ME, Utgoff PE. Learning to control a dynamic physical system.Comput Intell 3: 330–337, 1987.
Fahle M. Perceptual learning: specificity versus generalization. Curr OpinNeurobiol 15: 154–160, 2005.
Fine I, Jacobs RA. Comparing perceptual learning across tasks: a review. JVis 2: 190–203, 2002.
Fitts PM. The information capacity of the human motor system in controllingthe amplitude of movement. J Exp Psychol 47: 381–391, 1954.
Flanagan JR, Vetter P, Johansson RS, Wolpert DM. Prediction precedescontrol in motor learning. Curr Biol 13: 146–150, 2003.
Foo P, Kelso JAS, de Guzman GC. Functional stabilization of unstable fixedpoints: human pole balancing using time-to-balance information. J ExpPsychol Hum Percept Perform 26: 1281–1297, 2000.
Freedman EG. Coordination of the eyes and head during visual orienting. ExpBrain Res 190: 369–387, 2008.
Gander W, Golub GH, Strebel R. Least-squares fitting of circles and ellipses.BIT 34: 558–578, 1994.
Gantz L, Patel SS, Chung STL, Harwerth RS. Mechanisms of perceptuallearning of depth discrimination in random dot stereograms. Vision Res 47:2170–2178, 2007.
Gawthrop P, Lee KY, Halaki M, O’Dwyer N. Human stick balancing: anintermittent control explanation. Biol Cybern 107: 637–652, 2013.
Gritsenko V, Krouchev NI, Kalaska JF. Afferent input, efference copy,signal noise, and biases in perception of joint angle during active versuspassive elbow movements. J Neurophysiol 98: 1140–1154, 2007.
Guez A, Selinsky J. A neuromorphic controller with a human teacher. IEEEInt Conf Neural Networks 2: 595–602, 1988.
Guitton D, Volle M. Gaze control in humans: eye-head coordination duringorienting movements to targets within and beyond the oculomotor range. JNeurophysiol 58: 427–459, 1987.
Halliday D, Resnick R, Walker J. Fundamentals of Physics (6th ed.).Hoboken, NJ: Wiley, 2001.
Harris CM, Wolpert DM. Signal-dependent noise determines motor plan-ning. Nature 394: 780–784, 1998.
Kiemel T, Zhang Y, Jeka J. Identification of neural feedback for uprightstance in humans: stabilization rather than sway minimization. J Neurosci31: 15144–15153, 2011.
Koza JR, Keane MA. Genetic breeding of non-linear optimal control strate-gies for broom balancing In: Lecture Notes in Control and InformationSciences, edited by Bensoussan A, and Lions JL. Berlin: Springer, 1990, vol.144, p. 47–56.
Lakie M, Caplan N, Loram ID. Human balancing of an inverted pendulumwith a compliant linkage: neural control by anticipatory intermittent bias. JPhysiol 551: 357–370, 2003.
Lay BS, Sparrow WA, Hughes KM, O’Dwyer NJ. Practice effects oncoordination and control, metabolic energy expenditure, and muscle activa-tion. Hum Mov Sci 21: 807–830, 2002.
Lee C. Eye and head coordination in reading: roles of head movement andcognitive control. Vision Res 39: 3761–3768, 1999.
170 PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from

Lee CC, Berenji HR. An intelligent controller based on approximate reason-ing and reinforcement learning. Proc IEEE Int Symp Intell Control 4thAlbany 1989, p. 200–205.
Lee KY, O’Dwyer N, Halaki M, Smith R. A new paradigm for human stickbalancing: a suspended not an inverted pendulum. Exp Brain Res 221:309–328, 2012.
Loram ID, Kelly SM, Lakie M. Human balancing of an inverted pendulum:Is sway size controlled by ankle impedance? J Physiol 532: 879–891, 2001.
Loram ID, Lakie M. Human balancing of an inverted pendulum: positioncontrol by small, ballistic-like, throw and catch movements. J Physiol 540:1111–1124, 2002.
Mah CD, Mussa-Ivaldi FA. Evidence for a specific internal representation ofmotion–force relationships during object manipulation. Biol Cybern 88:60–72, 2003.
Maurer C, Peterka RJ. A new interpretation of spontaneous sway measuresbased on a simple model of human postural control. J Neurophysiol 93:189–200, 2005.
Mergner T, Maurer C, Peterka RJ. Sensory contributions to the control ofstance: a posture control model. Adv Exp Med Biol 508: 147–152, 2002.
McIntyre J, Zago M, Berthoz A, Lacquaniti F. Does the brain modelNewton’s laws? Nat Neurosci 4: 693–694, 2001.
Mehta B, Schaal S. Forward models in visuomotor control. J Neurophysiol88: 942–953, 2002.
Meyer DE, Abrams RA, Kornblum S, Wright CE, Smith JE. Optimality inhuman motor performance: Ideal control of rapid aimed movements. Psy-chol Rev 95: 340–370, 1988.
Miller WL, Maffei V, Bosco G, Iosa M, Zago M, Macaluso E, LacquanitiF. Vestibular nuclei and cerebellum put visual gravitational motion incontext. J Neurophysiol 99: 1969–1982, 2008.
Milton J. The delayed and noisy nervous system: implications for neuralcontrol. J Neural Eng 8: 065005, 2011.
Milton JG, Cabrera JL, Ohira T. Unstable dynamical systems: delays, noiseand control. Europhys Lett 83: 48001, 2008.
Milton J, Townsend JL, King MA, Ohira T. Balancing with positivefeedback: the case for discontinuous control. Philos Trans A Math Phys EngSci 367: 1181–1193, 2009a.
Milton JG, Ohira T, Cabrera JL, Fraiser RM, Gyorffy JB, Ruiz FK,Strauss MA, Balch EC, Marin PJ, Alexander JL. Balancing with vibra-tion: a prelude for “drift and act” balance control. PLoS One 4: e7427,2009b.
Neilson PD. Speed of response or bandwidth of voluntary system controllingelbow position in intact man. Med Biol Eng 10: 450–459, 1972.
O’Toole AJ, Kersten DJ. Learning to see random-dot stereograms. Percep-tion 21: 227–243, 1992.
Oytam Y, Neilson PD, O’Dwyer N. Degrees of freedom and motor planningin purposive movement. Hum Mov Sci 24: 710–730, 2005.
Payne VG, Isaacs LD. Human Motor Development: A Lifespan Approach.New York: McGraw-Hill, 2008.
Paz R, Wise SP, Vaadia E. Viewing and doing: similar cortical mechanismsfor perceptual and motor learning. Trends Neurosci 27: 496–503, 2004.
Poggio T, Bizzi E. Generalization in vision and motor control. Nature 431:768–774, 2004.
Porges SW, Bohrer RE, Cheung MN, Drasgow F, McCabe PM, Keren G.New time-series statistic for detecting rhythmic co-occurrence in the fre-
quency domain: the weighted coherence and its application to psychophys-iological research. Psychol Bull 88: 580–587, 1980.
Ramachandran VS. Learning-like phenomena in stereopsis. Nature 262:382–384, 1976.
Reeves NP, Pathak P, Popovich JM Jr, Vijayanagar V. Limits in motorcontrol bandwidth during stick balancing. J Neurophysiol 109: 2523–2527,2013.
Schmidt RA, Zelaznik H, Hawkins B, Frank JS, Quinn JT. Motor-outputvariability - theory for the accuracy of rapid motor acts. Psychol Rev 86:415–451, 1979.
Senot P, Zago M, Lacquaniti F, McIntyre J. Anticipating the effects ofgravity when intercepting moving objects: differentiating up and downbased on nonvisual cues. J Neurophysiol 94: 4471–4480, 2005.
Smith R, Rattanaprasert U, O’Dwyer N. Coordination of the ankle jointcomplex during walking. Hum Mov Sci 20: 447–460, 2001.
Solomon HY, Turvey MT. Haptically perceiving the distances reachable withhand-held objects. J Exp Psychol Hum Percept Perform 14: 404–427, 1988.
Stevens DS, Anderson DI, O’Dwyer NJ, Williams AM. Does self-efficacymediate transfer effects in the learning of easy and difficult motor skills?Conscious Cogn 21: 1122–1128, 2012.
Sumnall JH, Harris JM. Minimum displacement thresholds for binocularthree-dimensional motion. Vision Res 42: 715–724, 2002.
Tipler PA. Physics for Scientists and Engineers (4th ed.) London: W. H.Freeman, 1999.
Treffner PJ, Kelso JA. Dynamic encounters: long memory during functionalstabilization. Ecol Psychol 11: 103–137, 1999.
Turvey MT. Dynamic touch. Am Psychol 51: 1134–1152, 1996.Tyler CW. Stereoscopic depth movement–two eyes less sensitive than one.
Science 174: 958–961, 1971.Vahdat S, Darainy M, Milner TE, Ostry DJ. Functionally specific changes
in resting-state sensorimotor networks after motor learning. J Neurosci 31:16907–16915, 2011.
Van Beers RJ, Sittig AC, van der Gon JJD. The precision of proprioceptiveposition sense. Exp Brain Res 122: 367–377, 1998.
Wang X, O’Dwyer N, Halaki M, Smith R. Identifying coordinative structureusing principal component analysis based on coherence derived from linearsystems analysis. J Motor Behav 45: 167–179, 2013.
Westheimer G. Detection of disparity motion by the human observer. OptomVis Sci 67: 627–630, 1990.
Widrow B. The original adaptive neural net broom-balancer. Proc IEEE IntSymp Circuits Syst Philadelphia 1987, p. 351–357.
Winter DA, Patla AE. Signal Processing and Linear Systems for the Move-ment Sciences. Waterloo, ON, Canada: Waterloo Biomechanics, 1997.
Zago M, Lacquaniti F. Visual perception and interception of falling objects:a review of evidence for an internal model of gravity. J Neural Eng 2:S198–S208, 2005.
Zago M, Bosco G, Maffei V, Iosa M, Ivanenko YP, Lacquaniti F. Internalmodels of target motion: expected dynamics overrides measured kinematicsin timing manual interceptions. J Neurophysiol 91: 1620–1634, 2004.
Zago M, McIntyre J, Senot P, Lacquaniti F. Internal models and predictionof visual gravitational motion. Vision Res 48: 1532–1538, 2008.
Zatsiorsky VM, Gao F, Latash ML. Motor control goes beyond physics:differential effects of gravity and inertia on finger forces during manipula-tion of hand-held objects. Exp Brain Res 162: 300–308, 2005.
171PERCEPTUAL AND MOTOR LEARNING IN HUMAN STICK BALANCING
J Neurophysiol • doi:10.1152/jn.00538.2013 • www.jn.org
on January 14, 2015D
ownloaded from
Related Documents











