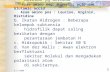
Pembentukan Anti gen dan Antibodi : Mekanisme / Proses Pembentukan Antigen dan Antibodi : Mekanisme / Proses - Di dalam tubuh manusia, antibodi dihasilkan oleh organ limfoid sentral yang terdiri atas sumsum tulang dan kelenjar timus, terutama oleh sel-sel limfosit. Ada dua macam sel limfosit, yaitu sel limfosit B dan sel limfosit T. Kedua sel ini bekerja sama untuk menghasilkan antibodi dalam tubuh. Baik antibodi maupun antigen keduanya mempunyai hubungan spesifi k yang sangat khas. Keadaan ini terlihat sewaktu antigen masuk ke dalam tubuh. Saat itu, dengan seketika sel limfosit T mendeteksi karakteristik dan jenis antigen. Kemudian sel limfosit T bereaksi cepat dengan cara mengikat antigen tersebut me lalui permukaan reseptornya. Setelah itu, sel limfosit T membelah dan membentuk klon. Sementara pada permukaan membrannya menghasilkan immunoglobulin monomerik. Berikutnya, molekul antigen dan molekul antibodi saling berikatan dan ikatan kedua molekul ini ditempatkan pada makrofaga. Secara berurutan, makrofaga menghadirkan antigen pada sel limfosit B. Lantas, sel limfosit B berpoliferasi dan menjadi dewasa, sehingga mampu membentuk antibodi untuk masing - masing antigen. Akan lebih jelas kalian simak Gambar 1. Gambar 1. Reaksi antigen dan antibodi Sementara itu, pembuangan antigen setelah diikat antibodi dapat menggun akan berbagai cara, yakni netralisasi, aglutinasi, presipitas i, dan fiksasi komplemen. Perhatik an Gambar 2.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript
Pembentukan Antigen dan Antibodi : Mekanisme / ProsesPembentukan Antigendan Antibodi : Mekanisme / Proses -Di dalam tubuh manusia, antibodi dihasilkan oleh organ limfoidsentral yang terdiri atas sumsum tulang dan kelenjar timus, terutamaoleh sel-sel limfosit. Ada dua macam sel limfosit, yaitu sel limfosit Bdan sel limfosit T. Kedua sel ini bekerja sama untuk menghasilkanantibodi dalam tubuh.Baik antibodi maupun antigen keduanyamempunyai hubungan spesifi k yang sangat khas.Keadaan ini terlihat sewaktu antigen masuk kedalam tubuh. Saat itu, dengan seketika sel limfositT mendeteksi karakteristik dan jenis antigen. Kemudiansel limfosit T bereaksi cepat dengan caramengikat antigen tersebut melalui permukaanreseptornya. Setelah itu, sel limfosit T membelahdan membentuk klon. Sementara pada permukaanmembrannya menghasilkan immunoglobulinmonomerik.Berikutnya, molekul antigen dan molekul antibodisaling berikatan dan ikatan kedua molekulini ditempatkan pada makrofaga. Secara berurutan,makrofaga menghadirkan antigen pada sellimfosit B. Lantas, sel limfosit B berpoliferasi danmenjadi dewasa, sehingga mampu membentukantibodi untuk masing - masing antigen. Akan lebih jelas kalian simakGambar 1.
Gambar 1.Reaksi antigen dan antibodi
Sementara itu, pembuangan antigen setelah diikat antibodi dapatmenggunakan berbagai cara, yakni netralisasi, aglutinasi, presipitasi, danfiksasi komplemen. Perhatikan Gambar 2.
Gambar 2.Mekanisme pelenyapan antigen
Netralisasi merupakan carayang digunakan antibodi untuk berikatan dengan antigen supaya aktivitasnyaterhambat. Sebagai contoh, antibodi melekat pada molekul yang akandigunakan virus untuk menginfeksi inangnya. Pada proses ini, antibodi danantigen dapat mengalami proses opsonisasi, yakni proses pelenyapan bakteriyang diikat antibodi oleh makrofaga melalui fagositosis.Cara pelenyapan antigen berikutnya adalah aglutinasi. Aglutinasi ataupenggumpalan merupakan proses pengikatan antibodi terhadap bakteriatau virus sehingga mudah dinetralkan dan diopsonisasi. Misalnya, IgGyang berikatan dengan dua sel bakteri atau virus secara bersama-sama.Mekanisme yang sama juga terjadi pada cara berikutnya yaknipresipitasi. Presipitasi atau pengendapan merupakan pengikatan silangmolekul-molekul antigen yang terlarut dalam cairan tubuh. Setelah diendapkan,antigen tersebut dikeluarkan dan dibuang melalui fagositosis.Selain berbagai cara tersebut, pembuangan antigen dapat melaluifiksasi komplemen. Fiksasi komplemen merupakan pengaktifanrentetan molekul protein komplemen karena adanya infeksi. Prosesnyamenyebabkan virus dan sel - sel patogen yang menginfeksi bagian tubuhmenjadi lisis.
MATURASI LIMFOSIT TPerkembangan T cell precursor dimulai di dalam sumsum tulang. T cell precursor bermigrasi ke dalam organ Thymus dan proses maturasi terjadi. Di dalam Thymus bagian subkapsular, T cell precursor menjadi timosit imature dan terjadi diferensiasi dan proliferasi dengan proses pembentukan gen TCR ( T cell Receptor ), CD8+ dan CD4+ dan diekpresikan (Double Positif Tymocyte), sebagian besar timosit imature mati dan sisanya terus berdeferensiasi. Pada daerah cortex di sel -sel epitel ( Thymic Epithelial cell) terjadi proses seleksi positif. Seleksi positif terjadi dengan cara reseptor tersebut mengenali MHC yang dipresentasikan oleh APC. Apabila MHC class I dikenali oleh CD8+ dan MHC class II dikenali oleh CD4+ kemudian menempel pada TCR maka timosit imature tetap hidup bila tidak mengenali APC tersebut maka akan mati atau mengalami apaptosis kemudian difagosit oleh makrofag Selanjutnya dilakukan seleksi negative, yaitu, timosite imature diuji dengan self antigen atau antigen tubuh sendiri. Bila mengenali atau pengenalan self reactive cell maka timosit imature akan mati. Timosite mature/nave melewati dinding venule postkapilar mencapai sirkulasi sistemik dan menempati organ limfoid perifer.
Pengenalan TCRBerfungsi dalam proses inflamasi dan memperbaiki jaringan yang rusak. Fungsi sel Th1 adalah pertahanan terhadap infeksi mikroba intraseluler yang mengaktifkan sel efektor. Kerjasama antara sel T dan fagosit merupakan kerja antarsel nonspesifik yang terjadi melalui sitokin. yang meningkatkan imunitas selular fagosit berupa makrofag. Makrofag yang diaktifkan oleh IFN-Timosite mature/nave sel T dapat mengenal antigen yang dipresentasikan oleh APC. Sel Th memberikan respon terhadap antigen dengan menghasilkan sitokin. Sel Tc memberikan respon terhadap antigen yang berkembang menjadi sel CTL yang dapat memusnahkan sel sendiri. Aktivasi sel T membutuhkan sinyal yang direspon oleh reseptor TCR, adanya molekul stimulatori, dan sitokin. Reseptor sel T hanya mengenal dan akan mengikat fragmen yang berhubungan dengan MHC. Sel T CD4+ yang berdiferensiasi menjadi Th2, mensekresikan IL-4 dan IL-5. IL-4 merangsang sel B untuk memproduksi IgE yang berikatan dengan sel mast. IL-5 mengaktifkan eosinofil sebagai respon terhadap patogen yang berupa cacing. Sitokin yang dihasilkan oleh Th2 menghambat aktivasi makrofag dan reaksi Th1. Sel CD4+ berdiferensiasi menjadi sel efektor Th1 yang mensekresikan IFN-
MATURASI LIMFOSIT BPrekursor sel B berkembang menjadi sel B immature di dalam sumsum tulang kemudian terjadi proliferasi dan deferensiasi yang ditandai dengan pembentukan BCR atau BCR somatic gen rearrangement, yaitu pembentukan reseptor yang dipresentasikan pada permukaan membrane. Setelah terbentuk IgM dalam tahap immature dalam sumsum tulang, sel B immature bermigrasi ke limpa atau disebut dengan sel B transisional, kemudian mengalami deferensiasi menjadi limfosit B mature. Pengembangan sel B terjadi melalui beberapa tahapan, setiap tahap mewakili perubahan genom pada lokus antibodi. Antibodi ini terdiri dari dua rantai identik Light (L)atau light chain dan dua rantai identik heavy (H) atau heavy chain, dan gen-gen ditemukan di daerah 'V' (Variable) dan daerah 'C' (Constant). Dalam heavy chain, daerah 'V' memiliki tiga segmen; V, D dan J, yang dikombinasikan secara acak, dalam proses yang disebut rekombinasi VDJ, untuk menghasilkan sebuah variabel unik imunoglobulin domain di masing - masing sel B. Sebagian besar sel B immature mati serta sisanya terus berdeferensiasi Setelah itu terjadi seleksi negative yaitu bila reseptor mengenali self antigen atau self reactive cell maka sel B immatur akan mati, bila tidak mengenali, maka akan tetap hidup. Sel B mature/nave melewati dinding venule postkapilar mencapai sirkulasi sistemik dan menempati organ limfoid perifer. Seleksi positif bila sel B mampu masuk ke organ sekunder tersebut.Ketika sel B mengalami kegagalan dalam setiap langkah dari proses maturasi, sel B akan mati melalui mekanisme apoptosis, dalam hal ini disebut clonal deletion. Jika telah mengenali self-Antigen selama proses maturasi, sel B akan mengalami apoptosis ( seleksi negatif ). Setelah teraktivasi, sel B akan terbentuk menjadi sel B memori sebagai bagian dari sistem imun adaptif.
AKTIVASI LIMFOSIT DAN PRODUKSI ANTIBODI
Aktivasi limfosit T, khususnya limfosit Th dari interaksi antara reseptor sel T + kompleks antigen-MHC kelas II yang terdapat di permukaan APC. Selain menyajikan antigen, APC juga memproduksi interleukin-1 yang mampu merangsang pertumbuhan sel T. Interaksi ini merangsang berbagai reaksi biokimia di dalam sel T, diantaranya adalah perombakan fosfatidil-inositol dan peningkatan konsentrasi ion Ca++serta aktivasi protein kinase-C yang diperlukan sebagai katalisator pada fosforilasi berbagai jenis protein. Reaksi-reaksi diatas mengakibatkan serangkaian reaksi-reaksi yang menghasilkan ekspresi reseptor IL-2 dan roduksi IL-2 yang diperlukan untuk proliferasi sel selanjutnya (Grey dkk, 1989; Abbas dkk, 1991; Roitt dkk, 1993).Sebagian dari sel T selanjutnya akan berfungsi sebagai sel T helper-inducer untuk membantu sel B, sebagian lagi akan kembali dalam keadaan istirahat menjadi sel memori. Aktivasi sel B dapat terjadi atas rangsangan antigen T-independen tipe Im antigen T-independen tipe II dan antigen T-dependen. Antigen T-dependen memerlukan bantuan sel Th. Antigen T-independen tipe I dalam konsentrasi tinggi mampu merangsang sel B secara poliklonal tanpa mengindahkan spesifsitas reseptor permukaan sel B. Contoh antigen seperti ini adalah lipopolisakarida pada permukaan sel bakteri. Tetapi pada konsentrasi rendah sel B dengan sIg spesifik sebagai reseptor dapat menangkap antigen sehingga sel teraktivasi.Antigen T-independen tipe II adalah antigen yang tidak segera dirombak didalam tubuh misalnyapolisakarida pneumokokus, polimer polivinilpirolidon(PVP) yang mampu merangsang sel B tanpa banuan sel Th. Antigen dapat melekat dengan aviditas kuat pada permukaan sel B dengan ikatan multivalen melalui sIg. Pada umumnya antigen T-independen merangsang pembentukan IgM. Sebagian besar antigen adalah T-dependen yang berarti respon pada sel B baru dapat terjadi atas rangsangan sel T. Agar sel B apat dirangsang oleh sel T maka MHC kelas II pada permukaan kedua sel harus sesuai. Hal ini penting untuk interaksi antara sel T dengan sel B dalam keadaan istirahat (resting B cells). Dilain pihak sel B yang sudah teraktivasi oleh kompleks antigen-MHC yang relevan.Sel T yang diaktivasi oleh antigen akan memproduksi interleukin-2 (IL-2) yang diperlukan untuk proliferasi sel T sendiri, disampign itu sel T juga memproduksi berbagai faktor atau limfokin yang dapat merangsang perubahan pada berbagai jenis sel antara lain sel B, sel T sitotoksik, makrofag dan lain-lain karenanya sel itu disebut sel T inducer (Grey dkk, 1989; Hendrik, 1989; Vitetta dkk, 1989).Berbagai jenis limfokin yang diproduksi oleh sel T dan dipergunakan untuk merangsang sel B adalah:B-cell stimulatory factor (IL 4),B-cell growth factor(II-6),B-cell differentiation factor-u (BCDF-u) danBCDF-gamma serta gamma interferon.Dengan rangsangan limfokin diatas sel B berproliferasi dan berdiferensiasi lebih lanjut menjadi sel plasma dan memproduksi imunoglobulin. BCDF-umerangsang produksi IgM yang diproduksi menjadi IgG dan selanjutnya akan terjadi sintesa dan sekresi immunoglobulin oleh sel plasma (Abbs dkk, 1991; Kresno, 1991).Selain berkembang menjadi sel plasma yang memproduksi imunoglobulin, stimulasi sel B perawan menyebabkan terbetuknya klon sel B yang perlahan-lahan kembali leleadaan istirahat dan menjadi sel memori. Sel ini seringkali mengekspresikan reseptor yang mengalami mutasi dan menunjukkan afinitas yang lebih tinggi. Sel B memori maupun sel T memori akan meninggalkan kelenjar limfe, limpa atau jaringan limfoid lain kemudian masuk kedalam pembuluh limfe dan pembuluh darah untuk melakukansurveillance(Bellanti, 1985; Subowo, 1993; Kresno, 1991).Respon imun sekunder pada umumnya timbul lebih cepat dan lebih kuat dibandingkan dengan respon primer. Hal ini disebabkan oleh karena adanya sel T dan sel B memori seta antibodi yang tersisa. Antigen dapat dikenal oleh sel B spesifik secara lebih efisien. Dalam hal ini sel B bertindak sebgai APC. Karena jumlah sel T dan sel B spesifik lebih banyak, kemungkinan untuk berinteraksi dengan antigen lebih besar, sehingga titer antibodi juga cepat meningkat. Disamping itu antibodi yang tersisa juga dapat bereaksi dengan antigen sehingga kompleks antigen antibodi lebih mudah ditangkap oleh APC dan diproses dan selanjutnya akan terjadi stimulasi sel T dan sel B seperti halnya pada respons imun tetapi dengan kecepatan efisiensi lebih tinggi (Bellanti, 1985;Roitt dkk, 1993).
A. SISTEM KOMPLEMENSistem komplemen adalah suatu sistem yang terdiri dari seperangkat kompleks protein yang satu dengan lainnya sangat berbeda. Ada 9 komponen dasar komplemen yaitu C1 sampai C9 yang bila diaktifkan, dipecah menjadi bagian-bagian yang besar dan kecil (C3a, C4a dan sebagainya). Fragmen yang besar dapat berupa enzim tersendiri dan mengikat serta mengaktifkan molekul lain. Fragmen tersebut dapat juga berinteraksi dengan inhibitor yang menghentikan reaksi selanjutnya. Komplemen sangat sensitif terhadap sinyal kecil, misalnya jumlah bakteri yang sangat sedikit sudah dapat menimbulkan reaksi beruntun yang biasanya menimbulkan reaksi lokal (Baratawidjaja, 2004).Unsur pokok sistem komplemen diwujudkan oleh sekumpulan komponen protein yang terdapat di dalam serum. Protein-protein ini dapat dibagi menjadi protein fungsional yang menggambarkan elemen dari berbagai jalur, dan protein pengatur yang menunjukkan fungsi pengendalian. Komplemen sebagian besar disintesis di dalam hepar oleh sel hepatosit, dan juga oleh sel fagosit mononuklear yang berada dalam sirkulasi darah. Komplemen C l juga dapat di sintesis oleh sel epitel lain diluar hepar. Komplemen yang dihasilkan oleh sel fagosit mononuklear terutama akan disintesis ditempat dan waktu terjadinya aktivasi. Komponen C3 mempunyai fungsi sangat penting pada aktivasi komplemen, baik melalui jalur klasik maupun jalur alternatif. Konsentrasi C3 jauh lebih besar dibandingkan dengan fraksi lainnya, hal ini menempatkan C3 pada kedudukan yang penting dalam pengukuran kadar komplemen di dalam serum. Penurunan kadar C3 di dalam serum dapat dianggap menggambarkan keadaan konsentrasi komplemen yang menurun. Juga penurunan kadar C3 saja dapat dipakai sebagai gambaran adanya aktivasi pada sistem komplemen (Judarwanto, 2009).Aktivasi komplemen menghasilkan sejumlah molekul efektor antara lain anafilaktoisin, kemotaksin, adherens imun, opsonin dan membrane attack complex yang mempunyai efek biologik (Baratawidjaja, 2004).B. AKTIVASI KOMPLEMENa. Aktivasi Komplemen melalui Jalur KlasikJalur ini diawali dengan stimulasi dari kompleks antigen-antibodi yang kemudian mengaktivasi C1q, C1r, C1s, ketiga komponen ini menghasilkan komponen enzimatik yang menstimulasi C4, C4 menghasilkan komponen enzimatik yang menstimulasi C2, komponen C2 ini kemudian menghasilkan komponen enzimatik dan menstimulasi C3 Convertase (pusat katalitik sistem komplemen).
(Gambar :Tiga Jenis Jalur Aktivasi Komplemen)
b. Aktivasi Komplemen melalui Jalur AlternatifJalur ini diawali oleh stimulasi dari permukaan patogen yang mengandung LPS (Lipopolisakarida) yang kemudian langsung menstimulasi C3, C3 menghasilkan komponen enzimatik yang menstimulasi faktor B, faktor B menghasilkan komponen enzimatik yang menstimulasi fakator D, faktor D kemudian menghasilkan komponen enzimatik yang akhirnya mensimulasi C3 convertase.c. Aktivasi Komplemen melalui Jalur LektinJalur ini diawali oleh stimulasi dari kompleks manosa binding protein pada permukaan patogen yang kemudian menstimulasi MBL, MASP-1, MASP-2. Ketiga komponen ini kemudian mnghasilkan komponen enzimatik yang menstimulasi C4, (seperti halnya pada jalur klasik) C4, C4 menghasilkan komponen enzimatik yang menstimulasiC2, komponen C2 ini kemudian menghasilkan komponen enzimatik dan menstimulasi C3 convertase (pusat katalitik sistem komplemen).
Setelah ketiga jalur tersebut mengaktivasi C3 Convertase, C3 convertase ini kemudian menghasilkan C3a, C5a dan C3b. C3a, C5a kemudian menstimulasi peptida mediator untuk inflamasi dan menstimulasi rekrutmen sel fagositik. C3b kemudian berikatan dengan reseptor komplemen pada sel fagositik dan kemudian menstimulasi opsonisasi dan penghilangan kompleks imun. Selain itu, C3b juga menstimulasi komponen terminal komplemen yang kemudian terjadi reaksi cascade : menstimulasi C5b,C6,C7,C8,C9 dan akhirnya membentuk Membran attack complex dan menyebabkan lisis pada patogen.C. RESEPTOR KOMPLEMENAda beberapa reseptor spesifik yang berikatan dengan komponen komplemen dan fragmen pada komplemen. CR1 dan CR3 adalah komponen penting dalam menginduksi proses fagositosis bakteri. CR2 terutaman ditemukan pada sel B yaitu kompleks sel B-koreseptor dan menjadi reseptor terhadap virus Epstein-Barr yang menyebabkan infeksi mononukleusis. CR1 dan CR2 saling membagi struktur komplemen-regulatory protein yang mengikat C3b dan C4b. CR3 dan CR4 saling berintegrasi, CR3 untuk proses migrasi dan adesi leukosit, sedangkan CR4 untuk respon fagositosis. Reseptor C5a dan C3a adalah bagian dari tujuh pasang pada untaian G protein. FDC sebagai sel folikular dendritik tidak terlibat dalam imunitas bawaan (innate immunity) (Janeway et all, 2001).Anafilaksis dan kemotaksisC3a, C4a dan C5a disebut anafilatoksin oleh karena dapat memacu sel mast dan sel basofil untuk melepaskan mediator kimia yang dapat meningkatkan permeabilitas dan kontraksi otot polos vaskular. Reseptor C3a dan C4a terdapat pada permukaan sel mast, sel basofil, otot polos dan limfosit. Reseptor C5a terdapat pada permukaan sel mast, basofil, netrofil, monosit, makrofag, dan sel endotelium.Melekatnya anafilatoksin pada reseptor yang terdapat pada otot polos menyebabkan kontraksi otot polos tersebut. Untuk mekanisme ini C5a adalah yang paling poten dan C4a adalah yang paling lemah. C5a juga mempunyai sifat yang tidak dimiliki oleh C3a dan C4a; oleh karena C5a juga mempunyai reseptor yang spesifik pada permukaan sel-sel fagosit maka C5a dapat menarik sel-sel fagosit tersebut bergerak ke tempat mikroorganisme, benda asing atau jaringan yang rusak; proses ini disebut kemotaksis. Juga setelah melekat C5a dapat merangsang metabolisme oksidatif dari sel fagosit tersebut sehingga dapat meningkatkan daya untuk memusnahkan mikroorganisme atau benda asing tersebut (Judarwanto, 2009).
Opsonisasi dan peningkatan fungsi fagositosisFagositosis yang diperkuat oleh proses opsonisasi C3b dan iC3b yang dibantu oleh IgG atau IgM mungkin merupakan mekanisme pertahanan utama terhadap infeksi bakteri dan jamur secara sistemik.Proses peradanganKombinasi dari semua fungsi yang tersebut diatas mengakibatkan terkumpulnya sel-sel dan serum protein yang diperlukan untuk terjadinya proses dalam rangka memusnahkan mikroorganisme atau benda asing tersebut.Pelarutan dan eliminasi kompleks imunKompleks imun yang beredar mengaktifkan komplemen dan mengaktifkan fragmen C3b yang menempel pada antigen. Kompleks tersebut akan berikatan dengan reseptor pada permukaan eritrosit. Pada waktu sirkulasi eritrosit melewati hati dan limpa, maka sel fagosit dalam limpa dan hati (sel Kupffer) dapat membersihkan kompleks imun yang terdapat pada permukaan sel eritrosit tersebut.D. REGULATORAktivasi komplemen dikontrol melalui tiga mekanisme utama, yaitu 1) komponen komplemen yang sudah diaktifkan biasanya ada dalam bentuk yang tidak stabil sehingga bila tidak berikatan dengan komplemen berikutnya akan rusak, 2) adanya beberapa inhibitor yang spesifik misalnya C1 esterase inhibitor, faktor I dan faktor H, 3) pada permukaan membran sel terdapat protein yang dapat merusak fragmen komplemen yang melekat.FUNGSI KOMPLEMEN1. InflamasiSebagai langkah awal untuk menghancurkan benda asing dan mikroorganisme serta membersihkan jaringan yang rusak. Tubuh mengerahkan elemen - elemen system imun ke tempat benda asing dan mikroorganisme yang masuk ke tubuh atau jaringan yang rusak tersebut. Fagositosis merupakan komponen penting pada inflamasi. Dalam inflamasi, ada 3 hal yang terjadi, yaitu : Peningkatan pasokan darah ke tempat benda asing dan mikrorganisme atau jaringan yang rusak. Peningkatan permeabilitas kapiler yang ditimbulkan oleh pengerutan sel endotel yang memungkinkan molekul yang lebih besar seperti antibody dan fagosit bergerak keluar pembuluh darah menuju ke tempat benda asing ( diapedesis )Mikrorganisme atau jaringan yang rusak. Peningkaan permeabilitas vascular yang local terjadi atas pengaruh anafilatoksin (C3a, C4a, C5a). Aktivasi komplemen C3 dan C5 menghasilkan fragmen kecil C3a dan C5a yang merupakan anafilatoksin yang dapat memacu degranulasi sel mast dan atau basofil melepas histamine. Histamine yang dapat dilepas sel mast atas pengaruh komplemen, meningkatkan permeabilitas vascular dan kontraksi otot polos dan keluarnya plasma yang mengandung banyak antibody, opsonin dan kompnen komplomen ke jaringan.2. KemokinMerupakan molekul yang dapat menarik dan mengerahkan sel-sel fagosit. C3a, C5a dan C5-6-7 merupakan kemokin yang dapat mengerahkans sel-sel fagosit baik mononuclear maupun polimorfonuklear ke tempat terjadi infeksi. C5a adalah kemoatraktan untuk neutrofil yang juga merupakan anafilatoksin. Monosit yang masuk ke jaringan menjadi makrofag, dan fagositosisnya diaktifkan opsonin dan antibody. Makrofag yang diaktifkan melepas berbagai mediator yang ikut berperan dalam reaksi inflamasi.3. Fagositosis opsoninC3b dan C4b mempunyai sifat opsonin. Opsonin adalah molekul yang dapat diikat disatu pihak leh partikel (kuman) dan dilain pihak oleh reseptornya pada fagosit sehingga memudahkan fagositosis bakteri atau sel lain. C3 yang banyak diaktifkan pada aktivasi komplemen merupakan sumber opsonin utama (C3b). Molekul C3b dalam bentuk inaktif (iC3b), juga berperan sebagai opsonin dalam fagositosis oleh karena fagositosis juga memiliki reseptor untuk CiC3b.IgG juga dapat berfungsi sebagai opsonin, bila berikatan dengan reseptor Fc pada permukaan fagosit. Oleh karena fagosit tidak memiliki reseptor Fc untuk IgM, opsonisasi yang dibantu konplemen merupakan hal yang sangat penting selama terjadi respon antibody primer yang didominasi IgM yang merupakan activator komponen poten. CRP juga berfungsi sebagai opsonin.4. Adherens ImunAdherens Imun merupakan fenomena dari partikel antigen yang melekat pada berbagai permukaan (mis: permukaan pembuluh darah), kemudian dilapis antibody dan mengaktifkan komplemen. Akibatkan anigen akan mudah difagositosis. C3b berfungsi dalam adherens imun tersebut.5. Elimiasi kompleks imunC3a atau iC3b dapat diendapkan dipermukaan kompleks imun dan merangsang eleminasi kompleks imun. Baik sel darah merah dan neutrofil memiliki CR1-R dan mengikat C3b dan iC3b. C3 dan C4 ditemukan dalam kompleks imun yang larut dan diikat oleh CR1-R pada sel darah merah yang mengangkutkan ke organ yang mengandung banyak fixed fagosit seperti hati dan limpa. Melalui reseptor komplemen dan Fc, fagosit-fagosit tersebut menyingkirkan dan menghancurkan kompleks imun dari sel darah merah. Pada proses ini, sel darah sendiri tidak rusak.Neutrofil dapat mengeliminasi kompleks imun kecil dalam sirkulasi. Bila antigen tidak larut yang diikat antibody dan dibentuk dalam darah atau jaringan tidak disingkirkan, akan memacu inflamasi dan dapat menimbulkan penyakit kompleks imun. Kompleks besar tidak larut sulit untuk disingkirkan dari jaringan; sejumlah besar C3 yang diaktifkan dapat melarutkan kompleks tersebut.
6. Lisis osmotic bakteriAktivasi C3 (jalur alternative atau klasik) akan mengaktifkan bagian akhir dari kaskade komponen komplemen C5-C9. Aktivasi komplemen yang erjadi dipermukaan sel bakteri akan membentuk Membrane Attack Complex dan akhirnya menimbulkan lisis osmotic sel atau bakteri. C5 dan C6 memiliki aktivasi enzim, yang memungkinkan C7, C8 dan C9 memasuki membrane plasma dari sel sasaran.7. Aktivitas sitolitikEosinofil dan sel polimorfonuklear mempnyai reseptor untuk C3b dan IgG sehingga 3b dapat meningkakan sitotoksisitas sel efektor Antibody Dependent Cell Mediated Cytotoxicity (ADCC) yang kerjanya bergantung pada IgG. Disamping itu sel darah merah yang diikat C3b dapat dihancurkan juga melalui kerusakan kontak. C8-9 merusak membrane membentuk saluran-saluran dalam membrane sel yang menimbulkan lisis osmotic.
RESEPTOR KOMPLEMENAktivasi komplemen jalur alternative dan klasik menghasilkan beberapa fragmen komplemen yang diikat oleh reseptor yang ditemukan pada berbagai jenis sel. C1qR ditemukan pada makrofag yang mengikat C1G dari jaringan kolagen dan berperanan pada elimnasi antigen. CR2 merupakan bagian dari kompleks ko-reseptor sel B dan juga ditemukan pada sel dendritik folikular yang berfungsi dalam fagositosis kompleks imun di center germinal dan dalam perkembangan sel memori. CR3 adalah antegrin (molekul adhesi). Pada fagosit mononukleat, neutrofil dan Sel NK yang fungsinya memudahkan fagositosis kompleks imun dan juga dalam migrasi monosit ke jaringan. CR4 merupakan intergrin yang memupunyai fungsi sama dengan CR3, diekspresikan terutama pada makrofag jaringan.Protein dalam serum yang merupakan komponen pada aktivasi komplemen, baik pada jalur klasik maupun jalur alternative dibentuk oleh hati, makrofag, monosit dan sel epitel intestinal. Bahan-bahan tersebut dilepas kedalam serum dalam bentuk tidak aktif. Pada tiap tahap penglepasan mediator terdapat mekanisme tubuh untuk menetralkan, yang disebut regulator, sehingga tidak akan terjadi reaksi yang berlangsung terus-menerus yang dapat menimbulkan kerusakan jaringan. System enzim yang kompleks ini diatur oleh beberapa penyekat protein yang dapat mencegah aktivasi premature dan aktivitas yang menunjang dari setiap produk. Contohnya adalah penyekat esterase CI (CI INH), penyekat C3b, inaktifator anafilatoksin dan penyekat C4b. Defesiensi bahan - bahan tersebut jarang ditemukan. Penyekat anafilatoksin menginaktifkan C3a dan C5a. penyekat C3b mengikat molekul tersebut dan membuatnya menjadi inaktif.
Related Documents