PDGFR-IgG chimera demonstrates that human 13 PDGFR Ig-like domains 1 to 3 are sufficient for high affinity PDGF BB binding MOHAMMAD A. HEIDARAN, DARUKA MAHAI)EVAN, AND WILLIAM J. LAROCHELLE’ National cancer Institute, National Institutes of Health, Bethesda, Maryland, 20892, USA KESEAHUII UOMMUNIUALIUNS 140 0892-6638/95/0009-0140/$01 .50. © FASEB ABSTRACT To localize human /3 PDGFR binding de- terminants, we constructed a fusion protein comprising /3 PDGFR Ig-like domains 1 to 3 and an IgGi Fc domain (/3 PDGFR-HFc). (3PDGFR-HFc was expressed as a 200 kDa dimeric molecule and contained Fc epitopes as demon- strated by anti-mouse Fc antibody recognition. Scatchard analysis revealed that PDGF BB possessed a dissociation constant of 1.5 nM for (3 PDGFR-HFc. Thus, /3 PDGFR Ig-like domains 1 to 3 are sufficient for high affinity PDGF BB binding. We exploited this fusion protein tech- nology to identify and characterize /3 PDGFR antagonists using a sensitive /3 PDGFR immunosorbent assay. In this assay, j3 PDGFR-HFchalf-maximally bound to PDGF BB with an affinity of around 150 pM. Suramin, as well as bacterially expressed and refolded human a PDGFR do- mains 1-3, inhibited /3 PDGFR-HFc binding to PDGF BB half-maximally at 25 M and 10 nM respectively. Therefore, a PDGFR D1-3, like /3 PDGFR D1-3, are sufficient for high affinity PDGF BB binding. Further- more, the /3 PDGFR-HFc immunosorbent assay will be useful to identify (3 PDGFR antagonists as well as to study a and 13 PDGFR substitution mutants which further map receptor binding determinants.- Heidaran, M. A., Mahadevan, D., LaRochelle, W. J. /3 PDGFR-IgG chimera demonstrates that human /3 PDGFR Ig-like domains 1 to 3 are sufficient for high affinity PDGF BB binding. FASEBJ. 9, 140-145 (1995) Key Words: fusion proteins platelet-derived growth factor recep- tor suramin PLATELET-DERIVED GROWTH FACTOR (PDGF)2 is a major se- rum mitogen for cells of mesenchymal origin such as glial cells, fibroblasts, and smooth muscle cells (1, 2). In addition to roles in wound healing and development, PDGF has also been implicated in numerous pathological states such as ne- oplasia (3, 4, 5) and arteriosclerosis (6, 7). Biochemical evi- dence has established the existence in vivo of three PDGF isoforms; PDGF AA, PDGF BB, and PDGF AB (8). The PDGF BB chain is a homolog of the v-sis oncogene product (9-11). Cell responsiveness to PDGF depends on signal transduc- tion through cell surface a or /3 PDGF receptors (PDGFRs) (12-14). The a PDGFR is triggered by all PDGF isoforms, while the /3 PDGFR is activated by PDGF BB and to a lesser extent by PDGF AB (15-17). Both a and 13 PDGFRs, in ad- dition to a split tyrosine kinase domain, possess five Ig-like domains (D1-D5) within their extracellular region. Com- puter modeling predicts that the a and /3 PDGF receptors share remarkable structural organization with im- munoglobulins in the size of these domains, spacing of cys- teine residues, and placement of characteristic tryptophan and tyrosine residues (18). Recent studies have demonstrated that the soluble extracellular region of the /3 PDGFR is capa- ble of high affinity PDGF binding and dimerization (19, 20). However, the structure(s) within the 3 PDGFR extracellular domain that confer PDGF BB binding are poorly understood. PDGFR mapping studies utilizing a and /3 PDGFR chimeras have demonstrated that a PDGFR Ig-like domains 1-3 specify PDGF AA interaction (21, 22). However, by this strategy we were unable to distinguish receptor domains that specified PDGF BB interaction because both a and /3 PDGFRs bound PDGF BB. Further mapping studies rev- ealed that deletions of a PDGFR Ig-like domains 4 or 5 had no effect on binding of either PDGF (23). Complementary immunochemical approaches demonstrated that the epitope of an a PDGFR neutralizing monoclonal antibody (mAb) (24) that blocked both PDGF AA and PDGF BB binding mapped to Ig-like domain 2 (22). In the present report, we map /3 PDGFR binding determinants and describe an im- munochemically based fusion protein assay for screening and identification of 3 PDGFR antagonists. EXPERIMENTAL PROCEDURES Construction of MMTneo /3 PDGFR-HFc The MMTneo-HFc expression vector containing an Xho I cloning site up- stream and adjacent to the mAb sis 1 (25) eDNA’s encoded hinge and Fe do- main (HFc) was engineered as described (W. J. LaRochelle, submitted for publication). 3 PDGFR eDNA, including its signal peptide to 29 amino acid residues beyond the 3’ encoded cysteine residue of Ig-like domain 3, was am- plified by PCR with Sal I compatible ends, digested with restriction en- donuclease, and cloned into MMTneo-HFc in frame with the IgG HFc en- coded eDNA. Transfection and immunoprecipitation MMTneo I PDGFR-HFc eDNA was transfected into NIII 3T3 cells by the calcium phosphate precipitation technique (26). Colony formation following selection in G418 containing medium was used as an internal marker of transfection efficiency. For immunoprecipitation analysis, /3 PDGFR-HFc transfectants were washed and incubated for 30 mm in methionine- and cysteine-free Dul- becco’s modified Eagle’s minimal essential medium containing 25 tM zinc ‘To whom correspondence should be addressed, at: National Cancer Institute, Building 37, Room 1E24, National Institutes of Health, Bethesda, MD 20892. ‘Abbreviations: PDGF, platelet-derived growth factor; PDGFR, platelet-derived growth factor receptor; HFc, IgG Fc domain in- cluding the hinge; Hc, IgG heavy chain; mAb, monoclonal anti- body; PAGE, polyacrylamide gel electrophoresis; PBS, phosphate- buffered saline; TTBS, 20 mM Tris pH 7.4, 0.15 M sodium chlo- ride, 0.05% Tween; BSA, bovine serum albumin; TAPS, 1% BSA, PBS, 0.05% Tween 20, 1.0% BSA, .02% sodium azide.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PDGFR-IgG chimera demonstrates that human 13 PDGFR
Ig-like domains 1 to 3 are sufficient for high affinity PDGF
BB bindingMOHAMMAD A. HEIDARAN, DARUKA MAHAI)EVAN, AND WILLIAM J. LAROCHELLE’
National cancer Institute, National Institutes of Health, Bethesda, Maryland, 20892,USA
KESEAHUII UOMMUNIUALIUNS
140 0892-6638/95/0009-0140/$01 .50. © FASEB
ABSTRACT To localize human /3 PDGFR binding de-terminants, we constructed a fusion protein comprising /3PDGFR Ig-like domains 1 to 3 and an IgGi Fc domain (/3PDGFR-HFc). (3PDGFR-HFc was expressed as a 200 kDadimeric molecule and contained Fc epitopes as demon-strated by anti-mouse Fc antibody recognition. Scatchardanalysis revealed that PDGF BB possessed a dissociationconstant of 1.5 nM for (3 PDGFR-HFc. Thus, /3 PDGFRIg-like domains 1 to 3 are sufficient for high affinityPDGF BBbinding. We exploited this fusion protein tech-nology to identify and characterize /3 PDGFR antagonistsusing a sensitive /3 PDGFR immunosorbent assay. In thisassay, j3 PDGFR-HFchalf-maximally bound to PDGF BBwith an affinity of around 150 pM. Suramin, as well asbacterially expressed and refolded human a PDGFR do-mains 1-3, inhibited /3 PDGFR-HFc binding to PDGFBB half-maximally at 25 M and 10 nM respectively.Therefore, a PDGFR D1-3, like /3 PDGFR D1-3, aresufficient for high affinity PDGF BB binding. Further-more, the /3 PDGFR-HFc immunosorbent assay will beuseful to identify (3 PDGFR antagonists as well as to studya and 13PDGFR substitution mutants which further mapreceptor binding determinants.- Heidaran, M. A.,Mahadevan, D., LaRochelle, W. J. /3 PDGFR-IgG chimerademonstrates that human /3 PDGFR Ig-like domains 1 to3 are sufficient for high affinity PDGF BB binding.FASEBJ. 9, 140-145 (1995)
Key Words: fusion proteins platelet-derived growth factor recep-tor suramin
PLATELET-DERIVED GROWTH FACTOR (PDGF)2 is a major se-rum mitogen for cells of mesenchymal origin such as glialcells, fibroblasts, and smooth muscle cells (1, 2). In additionto roles in wound healing and development, PDGF has alsobeen implicated in numerous pathological states such as ne-oplasia (3, 4, 5) and arteriosclerosis (6, 7). Biochemical evi-dence has established the existence in vivo of three PDGFisoforms; PDGF AA, PDGF BB, and PDGF AB (8). ThePDGF BB chain is a homolog of the v-sis oncogene product(9-11).
Cell responsiveness to PDGF depends on signal transduc-tion through cell surface a or /3 PDGF receptors (PDGFRs)(12-14). The a PDGFR is triggered by all PDGF isoforms,while the /3 PDGFR is activated by PDGF BB and to a lesserextent by PDGF AB (15-17). Both a and 13 PDGFRs, in ad-dition to a split tyrosine kinase domain, possess five Ig-likedomains (D1-D5) within their extracellular region. Com-puter modeling predicts that the a and /3 PDGF receptorsshare remarkable structural organization with im-munoglobulins in the size of these domains, spacing of cys-
teine residues, and placement of characteristic tryptophanand tyrosine residues (18). Recent studies have demonstratedthat the soluble extracellular region of the /3PDGFR is capa-ble of high affinity PDGF binding and dimerization (19, 20).However, the structure(s) within the 3 PDGFR extracellulardomain that confer PDGF BB binding are poorly understood.
PDGFR mapping studies utilizing a and /3 PDGFRchimeras have demonstrated that a PDGFR Ig-like domains1-3 specify PDGF AA interaction (21, 22). However, by thisstrategy we were unable to distinguish receptor domains thatspecified PDGF BB interaction because both a and /3PDGFRs bound PDGF BB. Further mapping studies rev-ealed that deletions of a PDGFR Ig-like domains 4 or 5 hadno effect on binding of either PDGF (23). Complementaryimmunochemical approaches demonstrated that the epitopeof an a PDGFR neutralizing monoclonal antibody (mAb)(24) that blocked both PDGF AA and PDGF BB bindingmapped to Ig-like domain 2 (22). In the present report, wemap /3 PDGFR binding determinants and describe an im-munochemically based fusion protein assay for screeningand identification of 3 PDGFR antagonists.
EXPERIMENTAL PROCEDURES
Construction of MMTneo /3 PDGFR-HFc
The MMTneo-HFc expression vector containing an Xho I cloning site up-
stream and adjacent to the mAb sis 1 (25) eDNA’s encoded hinge and Fe do-
main (HFc) was engineered as described (W. J. LaRochelle, submitted forpublication). 3 PDGFR eDNA, including its signal peptide to 29 amino acidresidues beyond the 3’ encoded cysteine residue of Ig-like domain 3, was am-plified by PCR with Sal I compatible ends, digested with restriction en-donuclease, and cloned into MMTneo-HFc in frame with the IgG HFc en-coded eDNA.
Transfection and immunoprecipitation
MMTneo I PDGFR-HFc eDNA was transfected into NIII 3T3 cells by thecalcium phosphate precipitation technique (26). Colony formation following
selection in G418 containing medium was used as an internal marker oftransfection efficiency.
For immunoprecipitation analysis, /3 PDGFR-HFc transfectants werewashed and incubated for 30 mm in methionine- and cysteine-freeDul-becco’s modified Eagle’s minimal essential medium containing 25 tM zinc
‘To whom correspondence should be addressed, at: NationalCancer Institute, Building 37, Room 1E24, National Institutes ofHealth, Bethesda, MD 20892.
‘Abbreviations: PDGF, platelet-derived growth factor; PDGFR,platelet-derived growth factor receptor; HFc, IgG Fc domain in-cluding the hinge; Hc, IgG heavy chain; mAb, monoclonal anti-body; PAGE, polyacrylamide gel electrophoresis; PBS, phosphate-buffered saline; TTBS, 20 mM Tris pH 7.4, 0.15 M sodium chlo-ride, 0.05% Tween; BSA, bovine serum albumin; TAPS, 1% BSA,PBS, 0.05% Tween 20, 1.0% BSA, .02% sodium azide.

IgG Hc _fl(_I PDGFR-HFc
C D
200 #{149} 200 -
97 97 -
12
200 -
97 -
4 5
SURAMIN AND SOLUBLE a PDGRF EFFECTS IN /3 PDGFR ANTAGONIST ASSAY 141
RESEARCH COMMUNICATIONS
chloride followed by metabolic labeling for 3.5 hr with [“S]methionine (125
tCi/ml) and [“S]cysteine (125 sCi/ml). After collection, conditionedmedium was immunoprecipitated with anti-mouse Fe followed by Staphylococctzs
aureus protein A Sepharose CL-4B. The immunoprecipitated proteins wereresolved by SDS-polyacrylamide gel electrophoresis (PAGE) and visualizedafter fluorography.
Enrichment of /3 PDGFR-HFc
Conditioned medium was collected from /3 PDGFR-HFc transfectants andpurified using anti-mouse IgG agarose (Sigma, St. Louis, Mo.). Briefly, 500ml of conditioned medium was filtered, made 0.1% in TX-lOU, and loadedovernight on a 2-mI column (packed volume). The column was washed with10 column volumes of phosphate-buffered saline (PBS), 0.1% TXIOO, fol-lowed by twenty column volumes of PBS. The /3PDGFR-HFc was eluted
with 200 mM glycine, HC1 pH 2.5. Fractions were immediately neutralized,
pooled, and concentrated. The concentrated eluate was then dialyzed intoPBS and stored at -70#{176}C.
Enrichment was assessed by silver stain as well as Western blot analysisafter SDS-PAGE using 8% gels. Western blotting was performed as
described after blocking with 20 mM Tris pH 7.4, 0.15 M sodium chloride,0.05% Tween 20 (TTBS), 3.0% milk for 2 h. The filters were incubated withanti-mouse Fe (Pierce) in TTBS, 4% bovine serum albumin (BSA) over-night and washed five times with TTBS, 4% BSA. Bound anti-mouse Fe wasdetected with “51-labeled Protein A in TTBS, 4% BSA followed by TTBSwashing. Filters were visualized after autoradiography.
Ligand binding assay
To determine the concentrationof “I-PDGF BB required for saturation,20 ng /3PDGFR-HFc was incubatedwith increasingconcentrationsof “I-labeled PDGF BB in 200 1d PBS, 0.05% Tween 20, 1.0% BSA, .02% sodiumazide (TAPS, 1% BSA) for four hours. To the reaction mixture, 500 ng anti-mouse Fe (Pierce) was added for 2 h followed by 100 sl Protein A Sepharose(1:1 slurry; Pharmacia) for I h. Immunocomplexes were washed four timeswith PBS, 0.1% TX-100. For Scatchardanalysis,20 ng /3PDGFR-HFc wasincubated with 5 ng “51-labeledPDGF BB and the unlabeledPDGF BBconcentrationvaried as indicated. Precipitated“-labeled PDGF BB wasquantitatedusing a Beckman gamma counter.
Receptor mediated immunosorbent assay BFalcon 3912 microtiter plates were coated overnight with 45 ng PDGF BBin PBS, 0.02% sodium azide. Microtiter wells were then blocked for 30 mmwith PBS, 0.02% sodium azide, 4% BSA. /3PDGFR-HFc, dilutedinTAPS,1% BSA was then added at the indicated concentration and incubated for
16 h. For competition experiments, suramin or a PDGFR D1-3 was addedat the indicated concentrations in TAPS, 1% BSA followed by either the /3PDGFR-HFc (8 nM) or PDGF antibody for 16 h. The wells were washedfive times with TAPS followed by either goat anti-mouse Fe (1:300, Sigma)or rabbit anti-goat antibody (1:300, Sigma) conjugated to alkaline phospha-
tase in TAPS, 1% BSA for 2 h. The wells were washed five times with TAPS.Alkaline phosphatase substrate 104 (1 mg/mI, Sigma) was resuspended in100 mM sodium bicarbonate, 1 mM magnesium chloride pH 9.8 and thenadded to each well. After development of the color reaction, absorbance wasmeasured at 405 nm.
RESULTS AND DISCUSSION
In these studies we determined by an alternative and directstrategy, based on our knowledge of PDGF AA binding to aand j3 PDGFR chimeras, which (3 PDGFR domains bindPDGF BB. We engineered a fusion protein in wh.ich the i3PDGFR eDNA encoding Ig-like D1-3 was recombined withthe HFc portion of the mouse immunoglobulin IgGs heavychain eDNA at the hinge region (Fig. 1A). Using PCR, we
amplified the /3 PDGFR sequence encoding Ig-like D1-3 withSal I compatible ends and subcloned the PCR product intothe MMTneo HFc vector in frame with the IgG1 HFc do-main. The grafting of the IgGj Fe domain to the /3 PDGFRconfers certain advantages such as a means for rapididentification, purification, and quantitation. We further ra-tionalized that deletion of the variable and CH1 domainsfrom the heavy chain would facilitate dimer secretion (27)and that grafting 3 PDGFR D1-3 to the random structure ofthe hinge would allow maximal adaptability consistent withpresentation of growth factor receptor biologic function.
MMTneo 3 PDGFR-HFc was transfected into NIH 3T3cells to investigate the immunochemical and biochemicalproperties of its translational product. Marker selected masscultures were radiolabeled with [‘5S]methionine and[35S]cysteine. Culture fluids were collected and incubatedwith anti-mouse Fe antibody followed by immunoprecipita-tion with protein A Sepharose. As shown in Fig. 1B, a dis-tinct p200 species was revealed. In contrast, this species wasnot found in immunoprecipitates of conditioned mediumfrom mock NIH 3T3 transfectants. In further experiments,protein A alone detected the chimeric gene product althoughthe signal was approximately 20-fold greater when amplifiedwith anti-mouse Fe (data not shown).
For biochemical analysis, /3 PDGFR-HFc was purifiedfrom culture fluids by anti-mouse IgG agarose immunoaffinitychromatography. The enriched /3 PDGFR-HFc was analyzedby silver staining and immunoblotting. As shown in Fig. 1C,(3PDGFR-HFc migrated as a p200 species that immunoblot-ted with anti-mouse Fe (Fig. 1D). Since /3 PDGFR-HFc pos-sesses the IgG1 heavy chain hinge region known to cause IgG
A3 PDGFR
Figure 1. Construction, expression, and purification of (3 PDGFR-HFc. PanelA. /3 PDGFR-HFc construction. The human (3 PDGFRand IgG, heavy chain (He) are depicted with Ig-like or Ig domainsrespectively. The chimeric /3 PDGFR-HFc, which contains /3PDGFR Ig-like domains 1-3 is illustrated. CH refers to the Heconstant regions 1, 2, and 3. The hinge region is depicted using theletter “H” Panel B. NIH 3T3 mock (lane 1) or (3PDGFR-HFc (lane
2) transfectants were metabolically labeled as described in Ex-perimental Procedures. Conditioned medium from each transfec-tant was immunoprecipitated with anti-mouse Fe, followed by pro-tein A-Sepharose CL-4B. The immunocomplexes were analyzed bySDS-PAGE under nonreducing conditions and the results werevisualized by fluorography. Panels C & D. Conditioned mediumfrom NIH 3T3 (3 PDGFR-HFc transfectants was purified by anti-mouse IgG agarose immunoaffinity chromatography. After concen-tration and dialysis, the eluate was subjected to silver stain (panelC) or anti-mouse Fe Western blot (panel D) analysis after SDS-PAGE under nonreducing (lane 4) or reducing (lane 5) conditionsas described in Experimental Procedures.
2.0
1.0
0.0
0 500
0)0)
0.04
0.00
50 100
142 Vol. 9 January 1995 The FASEB Journal HEIDARAN El AL.
RESEARCH COMMUNICATIONS
dimer formation, we sought to determine whether /3PDGFR-HFc is a disulfide-linked dimer. Addition of 100mM dithiothreitol prior to SDS-PAGE was associated withincreased mobility of an apparent 100 kDa species after im-munoblotting with anti-mouse Fe (Figs. IC and D). There-fore, /3 PDGFR-HFc was potently secreted as a disulfide-linked dimer possessing structural determinants of an im-munoglobulin Fe domain.
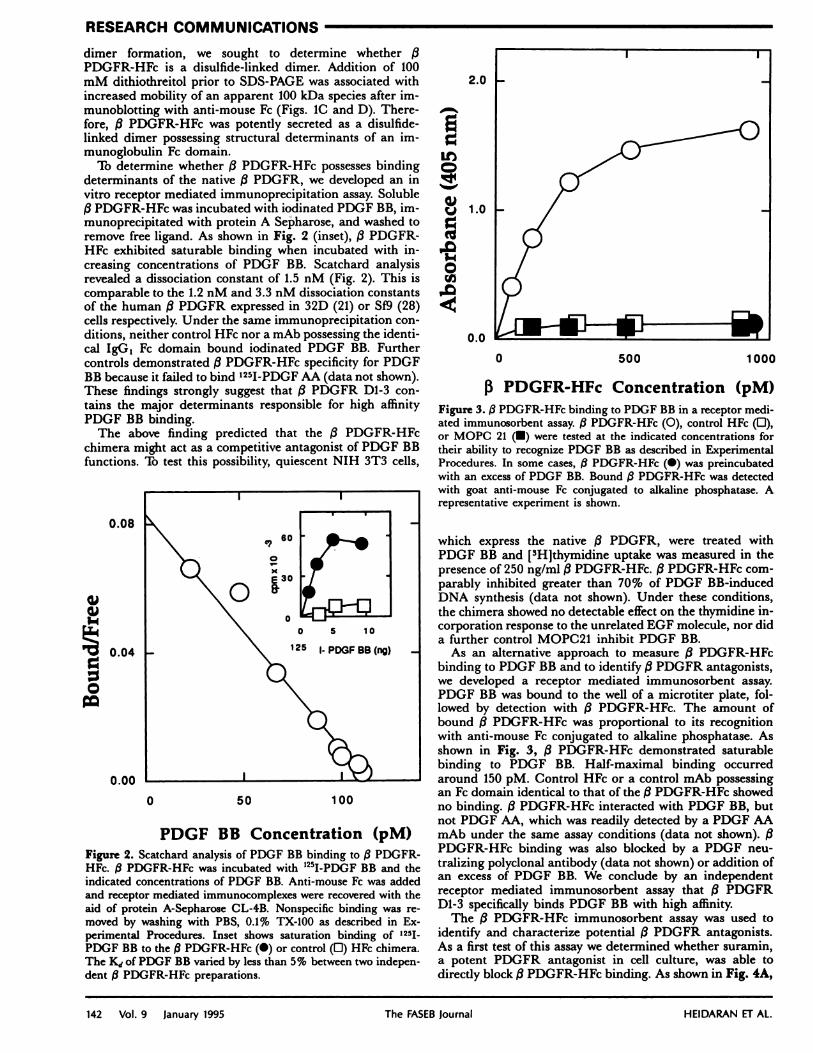
To determine whether /3 PDGFR-HFc possesses bindingdeterminants of the native /3 PDGFR, we developed an invitro receptor mediated immunoprecipitation assay. Soluble/3 PDGFR-HFc was incubated with iodinated PDGF BB, im-munoprecipitated with protein A Sepharose, and washed toremove free ligand. As shown in Fig. 2 (inset), /3 PDGFR-HFc exhibited saturable binding when incubated with in-creasing concentrations of PDGF BB. Scatchard analysisrevealed a dissociation constant of 1.5 nM (Fig. 2). This iscomparable to the 1.2 nM and 3.3 nM dissociation constantsof the human /3 PDGFR expressed in 32D (21) or 519 (28)cells respectively. Under the same immunoprecipitation con-ditions, neither control HFc nor a mAb possessing the identi-cal IgG1 Fe domain bound iodinated PDGF RB. Furthercontrols demonstrated /3 PDGFR-HFc specificity for PDGFBB because it failed to bind ‘251-PDGF AA (data not shown).These findings strongly suggest that /3 PDGFR D1-3 con-tains the major determinants responsible for high affinityPDGF BB binding.
The above finding predicted that the /3 PDGFR-HFcchimera might act as a competitive antagonist of PDGF BBfunctions. To test this possibility, quiescent NIH 3T3 cells,
0.08
0
PDGF BB Concentration (pM)Figure 2. Scatchard analysis of PDGF BB binding to /3 PDGFR-HFc. /3 PDGFR-HFc was incubated with “5I-PDGF BB and theindicated concentrations of PDGF BB. Anti-mouse Fe was addedand receptor mediated immunocomplexes were recovered with theaid of protein A-Sepharose CL-4B. Nonspecific binding was re-moved by washing with PBS, 0.1% TX-100 as described in Ex-perimental Procedures. Inset shows saturation binding of 1251
PDGF BB to the /3 PDGFR-HFc (#{149})or control (0) HFc chimera.The K1 of PDGF BB varied by less than 5% between two indepen-dent /3PDGFR-HFc preparations.
It)
0)cJ
0
1000
13 PDGFR-HFc Concentration (pM)Figure 3. /3 PDGFR-HFc binding to PDGF BB in a receptor medi-ated immunosorbent assay. 3 PDGFR-HFc (0), control HFc (0),or MOPC 21 (#{149})were tested at the indicated concentrations fortheir ability to recognize PDGF BB as described in ExperimentalProcedures. In some cases, /IPDGFR-HFc (#{149})was preincubatedwith an excess of PDGF BB. Bound /3 PDGFR-HFc was detectedwith goat anti-mouse Fe conjugated to alkaline phosphatase. Arepresentative experiment is shown.
which express the native /3 PDGFR, were treated withPDGF BB and [3H]thymidine uptake was measured in thepresence of 250 ng/ml /3PDGFR-HFc. /3 PDGFR-HFc com-parably inhibited greater than 70% of PDGF BB-inducedDNA synthesis (data not shown). Under these conditions,the chimera showed no detectable effect on the thymidine in-corporation response to the unrelated EGF molecule, nor dida further control MOPC21 inhibit PDGF RB.
As an alternative approach to measure /3 PDGFR-HFcbinding to PDGF BB and to identify /3 PDGFR antagonists,we developed a receptor mediated immunosorbent assay.PDGF BB was bound to the well of a microtiter plate, fol-lowed by detection with (3 PDGFR-HFc. The amount ofbound /3 PDGFR-HFc was proportional to its recognitionwith anti-mouse Fe conjugated to alkaline phosphatase. Asshown in Fig. 3, /3 PDGFR-HFc demonstrated saturablebinding to PDGF BB. Half-maximal binding occurredaround 150 pM. Control HFc or a control mAb possessingan Fe domain identical to that of the (3PDGFR-HFc showedno binding. /3 PDGFR-HFc interacted with PDGF BB, butnot PDGF AA, which was readily detected by a PDGF AAmAb under the same assay conditions (data not shown). (3PDGFR-HFc binding was also blocked by a PDGF neu-tralizing polyclonal antibody (data not shown) or addition ofan excess of PDGF BB. We conclude by an independentreceptor mediated immunosorbent assay that /3 PDGFRD1-3 specifically binds PDGF RB with high affinity.
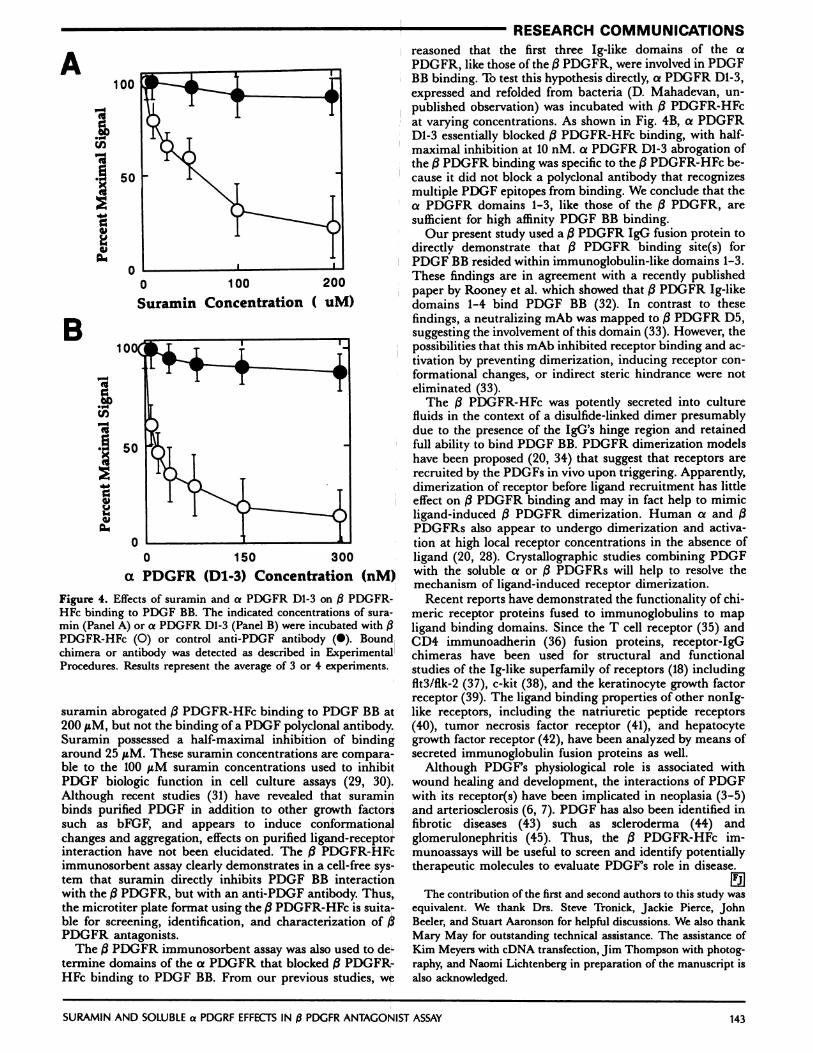
The /3 PDGFR-HFc immunosorbent assay was used toidentify and characterize potential /3 PDGFR antagonists.As a first test of this assay we determined whether suramin,a potent PDGFR antagonist in cell culture, was able todirectly block /3 PDGFR-HFc binding. As shown in Fig. 4A,
100
50
0
(5
0 100
Suramin Concentration (
200
uM)
10(
50
A
B
0 150 300
a PDGFR (D1-3) Concentration (nM)
Figure 4. Effects of suramin and a PDGFR D1-3 on /3 PDGFR-HFe binding to PDGF RB. The indicated concentrations of sura-mm (Panel A) or a PDGFR Dl-3 (Panel B) were incubated with j3PDGFR-HFc (0) or control anti-PDGF antibody (#{149}).Boundchimera or antibody was detected as described in Experimental
Procedures. Results represent the average of 3 or 4 experiments.
suramin abrogated /3 PDGFR-HFc binding to PDGF RB at200 sM, but not the binding of a PDGF polyclonal antibody.Suramin possessed a half-maximal inhibition of bindingaround 25 geM. These suramin concentrations are compara-ble to the 100 tM suramin concentrations used to inhibitPDGF biologic function in cell culture assays (29, 30).Although recent studies (31) have revealed that suraminbinds purified PDGF in addition to other growth factorssuch as bFGF, and appears to induce conformationalchanges and aggregation, effects on purified ligand-receptorinteraction have not been elucidated. The /3 PDGFR-HFcimmunosorbent assay clearly demonstrates in a cell-free sys-tem that suramin directly inhibits PDGF BR interactionwith the /3 PDGFR, but with an anti-PDGF antibody. Thus,the microtiter plate format using the (3PDGFR-HFc is suita-ble for screening, identification, and characterization of j3PDGFR antagonists.
The /3 PDGFR immunosorbent assay was also used to de-termine domains of the a PDGFR that blocked (3PDGFR-HFc binding to PDGF BB. From our previous studies, we
reasoned that the first three Ig-like domains of the aPDGFR, like those of the /3 PDGFR, were involved in PDGFBB binding. To test this hypothesis directly, a PDGFR Dl-3,expressed and refolded from bacteria (D. Mahadevan, un-published observation) was incubated with /3 PDGFR-HFcat varying concentrations. As shown in Fig. 4B, a PDGFRD1-3 essentially blocked /3 PDGFR-HFc binding, with half-maximal inhibition at 10 nM. a PDGFR D1-3 abrogation ofthe /3 PDGFR binding was specific to the /3 PDGFR-HFc be-cause it did not block a polyclonal antibody that recognizesmultiple PDGF epitopes from binding. We conclude that thea PDGFR domains 1-3, like those of the /3 PDGFR, aresufficient for high affinity PDGF RB binding.
Our present study used a /3 PDGFR IgG fusion protein todirectly demonstrate that /3 PDGFR binding site(s) forPDGF BR resided within immunoglobulin-like domains 1-3.These findings are in agreement with a recently publishedpaper by Rooney et al. which showed that /3 PDGFR Ig-likedomains 1-4 bind PDGF BB (32). In contrast to thesefindings, a neutralizing mAb was mapped to /3 PDGFR D5,suggesting the involvement of this domain (33). However, thepossibilities that this mAb inhibited receptor binding and ac-tivation by preventing dimerization, inducing receptor con-formational changes, or indirect steric hindrance were noteliminated (33).
The /3 PDGFR-HFc was potently secreted into culturefluids in the context of a disulfide-linked dimer presumablydue to the presence of the IgG’s hinge region and retainedfull ability to bind PDGF BB. PDGFR dimerization modelshave been proposed (20, 34) that suggest that receptors arerecruited by the PDGFs in vivo upon triggering. Apparently,dimerization of receptor before ligand recruitment has littleeffect on /3 PDGFR binding and may in fact help to mimicligand-induced /3 PDGFR dimerization. Human a and /3PDGFRs also appear to undergo dimerization and activa-tion at high local receptor concentrations in the absence ofligand (20, 28). Crystallographic studies combining PDGFwith the soluble a or 3 PDGFRs will help to resolve themechanism of ligand-induced receptor dimerization.
Recent reports have demonstrated the functionality of chi-meric receptor proteins fused to immunoglobulins to mapligand binding domains. Since the T cell receptor (35) andCD4 immunoadherin (36) fusion proteins, receptor-IgGchimeras have been used for structural and functionalstudies of the Ig-like superfamily of receptors (18) includingflt3/flk-2 (37), c-kit (38), and the keratinocyte growth factorreceptor (39). The ligand binding properties of other nonlg-like receptors, including the natriuretic peptide receptors(40), tumor necrosis factor receptor (41), and hepatocytegrowth factor receptor (42), have been analyzed by means ofsecreted immunoglobulin fusion proteins as well.
Although PDGF’s physiological role is associated withwound healing and development, the interactions of PDGFwith its receptor(s) have been implicated in neoplasia (3-5)and arteriosclerosis (6, 7). PDGF has also been identified infibrotic diseases (43) such as seleroderma (44) andglomerulonephritis (45). Thus, the /3 PDGFR-HFc im-munoassays will be useful to screen and identify potentiallytherapeutic molecules to evaluate PDGF’s role in disease.
The contribution of the first and second authors to this study wasequivalent. We thank Drs. Steve Tronick, Jackie Pierce, JohnBeeler, and Stuart Aaronson for helpful discussions. We also thankMary May for outstanding technical assistance. The assistance ofKim Meyers with cDNA transfection, Jim Thompson with photog-raphy, and Naomi Lichtenberg in preparation of the manuscript isalso acknowledged.
RESEARCH COMMUNICATIONS
SURAMIN AND SOLUBLE a PDGRF EFFECTS IN (3PDGFR ANTAGONIST ASSAY 143
-
.-
U,
(5
I

144 Vol. 9 January 1995 The FASEB Journal HEIDARAN ET AL.
RESEARCH COMMUNICATIONS
REFERENCES
1. Aaronson, S. A. (1991) Growth factors and cancer. Science254, 1146-1153
2. Heldin, C-H., and Westermark, B. (1991) Platelet-derived growth factorand autocrine mechanisms of oncogenic processes. Grit. Rev. Oncog. 2,109-124
3. Nister, M., Claesson-Welsh, L., Eriksson, A., Heldin, C-H., andWestermark, B. (1991) Differential expression of platelet-derived growth
factor receptor in human malignant glioma cell lines. j Biol. Chem. 266,16755-16763
4. Fleming, T. P., Matsui, T., Heidaran, M. A., Molloy, C. J., Artrip, J.,and Aaronson, S. A. (1992) Amplification and/or overexpression of
platelet-derived growth factor receptors and epidermal growth factorreceptor in human glial tumors. Cancer Res. 52, 4550-4553
5. Fleming, T. P., Saxena, A., Clark, W. C., Robertson, J. T., Oldfield,E. H., Aaronson, S. A., and Ali, 1. U. (1992) Demonstration of an acti-vated platelet-derived growth factor autocrine pathway and its role inhuman tumor cell proliferation in vitro. Oncogene 7, 1355-1359
6. Ross, R., Masuda, J., Raines, E. W., Gown, A. M., Katsuda, S., Sasa-
hara, M., Maiden, L. T., Masuko, H., and Sato, H. (1990) Localization
of PDGF-B protein in macrophages in all phases of atherogenesis. Science248, 1009-1012
7. Ferns, G. A. A., Raines, E. W., Sprugel, K. H., Motani, A. S., Reidy,
M. A., and RDss, R. (1991) Inhibition of neointimal smooth muscle ac-cumulation after angioplasty by an antibody to PDGF. Science 253,
1129-11328. Johnsson, A., Heldin, C-H., Westermark, B., and Wasteson, A. (1982)
Platelet-derived growth factor: identification of constituent polypeptidechains. Biochem. Biophys. Res. Commun. 104, 66-71
9. Devare, S. G., Reddy, E. P., Law, D. J., Robbins, K. C., and Aaronson,S. A. (1983) Nucleotide sequence of the simian sarcoma virus genome:demonstration that its acquired cellular sequences encode the putativetransforming gene product p28”. Proc. Nail.Acad. &i. USA 80, 731-735
10. Doolittle, R. F., Hunkapiller, M. W., Hood, L. E., Devare, S. G., Rob-
bins, K. C., Aaronson, S. A., and Antoniades, H. M. (1983) Simian sar-
coma virus oncogene, v-sis, is derived from the gene or genes encodinga platelet-derived growth factor. Science 221, 275-277
II. Waterfield, M. D., Scrace, G. T., Whittle, N., Stroobant, P., Johnsson,
A., Wasteson, A., Westermark, B., Heldin, C-H., Huang, J. S., andDeuel, T. F. (1983) Platelet-derived growth factor is structurally related
to the putative transforming protein p28” of simian sarcoma virus.Nature (London) .304, 35-39
12. Yarden, Y.,Escobedo, J. A., Kuang, W.-J., Yang-Feng, T. L., Daniel,
T. 0., Tremble, P. M., Chen, E. Y., Ando, M. E., Harkins, R. N.,Franke, U., Friend, V. A., Ullrich, A., and Williams, L. T (1986) Struc-
ture of the receptor for platelet-derived growth factor helps define a fa-
mily of closely related growth factor receptors. Nature (London) 323,226-232
13. Matsui, T., Heidaran, M., Miki, T., Popescu, N., LaRochelle, W. J.,Kraus, M., Pierce, J., and Aaronson, S. A. (1989) Isolation of a novelreceptor eDNA establishes the existence of two PDGF receptor genes.
Science 243, 800-80414. Claesson-Welsh, L., Eriksson, A., Westermark, B., and Heldin, C-H.
(1989) eDNA cloning and expression of the human A-type platelet-derived growth factor (PDGF) receptor establishes structural similarityto the B-type PDGF receptor. Proc. Nail. Acad. Sci. USA 86, 4917-4921
15. Hart, C. E., Forstrom,J. W., Kelly,J. D., Seifert, R. A., Smith, R. A.,Ross, R., Murray, M. J., and Bowen-Pope, D. F. (1988) Two classes ofPDGF receptor recognize different isoforms of PDGF. Science 240,1529-1531
16. Heldin, C-H., Backstrom, G., Ostman, A., Hammacher, A., Ronn-strand, L., Rubin, K., Nister, M., and Westermark, B. (1988) Binding
of two different dimeric forms of PDGF to human fibroblasts: evidence
for two separate receptor types. EMBOJ. 7, 1387-139317. Matsui, T., Pierce, J., Fleming, T. P., Greenberger, J. S., LaRochelle,
W. J., Ruggiero, M., and Aaronson, S. A. (1989) Independent expres-sion of human a and /3platelet-derived growth factor receptor cDNAsin a naive hematopoietic cell leads to functional coupling with mitogenicand chemotactic signaling pathways. Proc. Nail. Acad. &z. USA 86,8314-8318
18. Williams, A. F., and Barclay, A. N. (1988) The immunoglobulinsuperfamily-Domains for cell surface recognition. Annu. Rev. Immunol.
6, 381-40519. Duan, D.-S.,Pazin,M. J., Fretto, L. J., and Williams, L. T. (1991) A
functional soluble extracellular region of platelet-derivedgrowth factor(PDGF) /3-receptor antagonizes PDGF-stimulated responses. J. Biol.Chem. 266, 413-418
20. Herren, B.,Rooney, B.,Weyer, K. A.,Schmid, G., and Pech, M. (1993)Dimerizationofextracellulardomains ofplatelet-derivedgrowth factorreceptors. J. Biol. Chem. 268, 15088-15095
21. Heidaran, M. A., Yu, J.-C., Jensen, R. A., Matsui, T., and Aaronson,S. A. (1990) Chimeric a- and /3-platelet-derived growth factor (PDGF)receptors define three immunoglobulin-like domains of the a-PDGF
receptor that determine PDGF-AA binding specificity. j Biol. Chem.265, 18741-18744
22. Yu, J.-C., Mahadevan, D., LaRochelle, W. J., Pierce, J. H., and Heida-ran, M. A. (1994) Structural coincidence of aPDGFR epitopes bindingto platelet-derived growth factor-AA and a potent neutralizing
monoclonal antibody. J. Biol. C/tern. 269, 10668-1067423. Heidaran, M. A., Yu, J.-C., Jensen, R. A., Pierce, J. H., and Aaronson,
S. A. (1992) A deletion in the extracellular domain of the a platelet-derived growth factor (PDGF) receptor differentially impairs PDGF-AAand PDGF-BB binding affinities. j Biol. C/tern. 267, 2884-2887
24. LaRochelle, W. J., Jensen, R. A., Heidaran, M. A., May-Siroff, M.,Wang, L.-M., Aaronson, S. A., and Pierce, J. H. (1993) Inhibition of
platelet-derived growth factor autocrine growth stimulation by amonoclonal antibody to the human a platelet-derived growth factorreceptor. Cell Growth Duff 4, 547-553
25. LaRochelle, W. J., Robbins, K. C., and Aaronson, S. A. (1989) Im-munochemical localization of the epitope for a monoclonal antibodythat neutralizes human platelet-derived growth factor mitogenic ac-tivity. Mol. Cell. Biol. 9, 3538-3542
26. Wigler, M., Silverstein, S., Lee, L.-S., Pellicer, A., Cheng, Y.-C., andAxel,R. (1977)Transferof purified herpes virus thymidine kinase gene
to cultured mouse cells. Cell 11, 223-23227. Cogne, M., Silvain, C., Khamlichi, A. A., and Preud’homme, J.-L.
(1992) Structurally abnormal immunoglobulins in human immunopro-liferative disorders. Blood 79, 2181-2195
28. Jensen, R. A., Reeler, J. F.,Heidaran, M. A., and LaRochelle, W. J.(1992) Characterization of baculovirus expressed human a and /3PDGFreceptors. Biochemistry 31, 10887-10892
29. Betsholtz, C., Johnsson, A., Heldin, C-H., and Westermark, B. (1986)Efficient reversion of simian sarcoma virus-transformation and inhibi-
tion of growth factor-induced mitogenesis by suramin. Proc. Nail. Acad.
Sci. USA 83, 6440-644430. Fleming, T. P., Matsui, T., Molloy, C. J., Robbins, K. C., and Aaron-
son, S. A. (1989) Autocrine mechanism for v-sis transformation requirescell surface localization of internally activated growth factor receptors.Proc. Nail. Acad. Sci. USA 86, 8063-8067
31. Middaugh, C. R., Mach, H., Burke, C.J., Volkin, D. B., Dabora,J. M.,Tsai, P. K., Bruner, M. W., Ryan, J. A., and Marfia, K. E. (1992) Na-
ture of the interaction of growth factors with suramin. Biochemistry 31,9016-9024
32. Rooney, B. C., Hosang, M., and Hunziker, W. (1994)Production ofplatelet-derived growth factor receptor (PDGFR-/3) in E. coli. FEBS Leii.339, 181-184
33. Ramakrishnan, V., Escobedo, M.-A., Fretto, L. J., Seroogy, J. J., Tom-linson, J. E., and Wolf, D. L. (1993) A novel monoclonal antibody de-
pendent on domain 5 of the platelet-derived growth factor beta receptorinhibits ligand binding and receptor activation. Growth Factors 8,253-265
34. Seifert, R. A., Hart, C. A., Phillips, P. E., Forstrom, J. W., Ross, R.,Murray, M. J., and Bowen-Pope, D. F. (1989) Two different subunits as-sociate to create isoform-specific platelet-derived growth factor recep-tors. j Biol. C/tern. 264, 8771-8778
35. Gascoigne, N.J. R., Goodnow, C., Dudzik, K. I., Oi, V. T., and Davis,M. M. (1987) Secretion of a chimeric T-cell receptor-immunoglobulinprotein. Proc. Nail. Acad. Sci. USA 84, 2936-2940
36. Capon, D. J., Chamow, S.M., Mordenti, J., Marsters,S. A., Gregory,T., Mitsuya, H., Byrn, R. A., Lucas, C., Wurm, F. M., Groopman,J. E., Broder, S., and Smith, D. H. (1989)Designing CD4 immunoadhe-sins for AIDS therapy. Nature (London) 337, 525-.531
37. Lyman, S. D., James, L., Vanden Bos, T, de Vries, P., Brasel, K., GIi-niak, B., Hollingsworth, L. T., Picha, K. S., McKenna, H. J., Sp}ett,R. R., Fletcher, F. A., Maraskovsky, E., Farrah, T, Foxwerthe, D., Wil-liams, D. E., and Beckmann, M. P. (1993) Molecular cloning ofaligandfor the fIt3/fik-2 tyrosine kinase receptor: A proliferative factor forprimitive hematopoietic cells. Cell 75, 1157-1167
38. Liu, Y.-C., Kawagishi, M., Kameda, R., and Ohashi, H. (1993) Charac-terization of a fusion protein composed of the extracellular domain of
c-kit and the Fe region of human IgG expressed in a baculovirus system.Biochern. Biophys. Res. Co,nrnun. 197, 1094-1102
39. Cheon, H.-G., LaRochelle, W. J., Bottaro, D. P., Burgess, W. H., andAaronson, S. A. (1994) High-affinity binding sites for related fibroblastgrowth factor ligands reside within different receptor immunoglobulin-like domains. Proc. Nail. Acad. Sci. USA 91, 989-993
40. Bennett, B. D., Bennett, G. L., Vitangeol, R. V., Jewett, J. R. S.,Bur-flier,J., Henzel, W., and Lowe, D. G. (1991)Extracellulardomain-IgGfusion proteins for three human natriuretic peptide receptors. j Biol.Chern. 266, 23060-23067
41. Howard, 0. M. Z.,Clouse,K. A., Smith, C.,Goodwin, R. G.,and Far-

SURAMIN AND SOLUBLE a PDGRF EFFECTS IN /3PDGFR ANTAGONIST ASSAY 145
RESEARCH COMMUNICATIONSrar, W. L. (1993) Soluble tumor necrosis factor receptor: Inhibition ofhuman immunodeficiency virus activation. Proc. Nail. Acad. Sci. USA 90,2335-2339
42. Mark, M. R., Lokker, N. A., Zioncheck, T. F., Luis, E. A., and Godowski,P. J. (1992) Expression and characterization of hepatocyte growth factorreceptor-IgG fusion proteins. j Biol. Chern. 267, 26166-26171
43. Smits, A., Funa, K., Vassbotn, F. S., Beausang-Linder, M., af Eken-stam, F., Heldin, C-H., Westermark, B., and Nister, M. (1992) Expres-sion of platelet-derived growth factor and its receptors in proliferativedisorders of fibroblastic origin. Am. J Pat/wI. 140, 639-648
44. Gay, S., Jones, R. E., Guo-qiang, H., and Gay, R. E. (1989) Immuno-
histologic demonstration of platelet-derived growth factor (PDGF) and
sis-oncogene expression in scleroderma. j Invest. Dermatol. 92, 301-30345. lida,H., Seifert,R.,Alpers, C. E.,Gronwald, R. G. K., Phillips,P.E.,
Pritzl, P., Gordon, K., Gown, A. M., Ross, R., Bowen-Pope, D. F., andJohnson, R. J. (1991) Platelet-derived growth factor (PDGF) and PDGFreceptor are induced in mesangial proliferative nephritis in the rat. Proc.Nail. Acad. Sci. USA 88, 6560-6564
Received for publication June 17, 1994Accepted for publication October 19, 1994
Erratum
The caption to figure 1 [Chambers, D. A., Reid, K. B. M., and Cohen, R. L. (1994) DNA: the double helix and the biomedi-cal revolution at 40 years. FASEBJ 8, 1219-1226] incorrectly identified some of those pictured. The authors’ corrected cap-tion appears below.
Figure 1. Speakers at the DNA conference. Kneeling: M. Meselson. Seated left to right: I. Klotz, L. Hood, J. Baxter, A. Rich, J. Watson,
G. Stent, D. Chambers (chair), J. Lederberg, D. Baltimore, F. Jacob, R. Doll, H. Temin. Standing left to right: S. Tonegawa, D. Jackson,H. Judson, M. Nirenberg, I. Pastan, R. Hotchkiss, P. Heller, J. Solaro (partially obscured), C. Tiekell, E. Kandel, K. Davies, R. Storti,H. Slavkin, D. Weatherall, W. Bodmer, S. Ross. Not present: H. Bourne, R. Davidson, W. Gilbert, A. KIug, M. McCarty, R. Mulligan,H. Varmus, J. Walton, R. Weinberg.
Related Documents











