Accepted Manuscript Title: PCR-sequence characterisation of new adenoviruses found in reptiles and the first successful isolation of a lizard adenovirus Authors: Tibor Papp, Beth Fledelius, Volker Schmidt, Gy˝ oz˝ o L. Kaj´ an, Rachel E. Marschang PII: S0378-1135(08)00338-6 DOI: doi:10.1016/j.vetmic.2008.08.003 Reference: VETMIC 4124 To appear in: VETMIC Received date: 13-5-2008 Revised date: 24-7-2008 Accepted date: 14-8-2008 Please cite this article as: Papp, T., Fledelius, B., Schmidt, V., Kaj´ an, G.L., Marschang, R.E., PCR-sequence characterisation of new adenoviruses found in reptiles and the first successful isolation of a lizard adenovirus, Veterinary Microbiology (2007), doi:10.1016/j.vetmic.2008.08.003 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. peer-00532460, version 1 - 4 Nov 2010 Author manuscript, published in "Veterinary Microbiology 134, 3-4 (2009) 233" DOI : 10.1016/j.vetmic.2008.08.003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Accepted Manuscript
Title: PCR-sequence characterisation of new adenovirusesfound in reptiles and the first successful isolation of a lizardadenovirus
Authors: Tibor Papp, Beth Fledelius, Volker Schmidt, GyozoL. Kajan, Rachel E. Marschang
PII: S0378-1135(08)00338-6DOI: doi:10.1016/j.vetmic.2008.08.003Reference: VETMIC 4124
To appear in: VETMIC
Received date: 13-5-2008Revised date: 24-7-2008Accepted date: 14-8-2008
Please cite this article as: Papp, T., Fledelius, B., Schmidt, V., Kajan, G.L., Marschang,R.E., PCR-sequence characterisation of new adenoviruses found in reptiles and thefirst successful isolation of a lizard adenovirus, Veterinary Microbiology (2007),doi:10.1016/j.vetmic.2008.08.003
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0Author manuscript, published in "Veterinary Microbiology 134, 3-4 (2009) 233"
DOI : 10.1016/j.vetmic.2008.08.003

Page 1 of 27
Accep
ted
Man
uscr
ipt
1
PCR-sequence characterisation of new adenoviruses found in 1
reptiles and the first successful isolation of a lizard adenovirus. 2
3
4
Tibor Papp1, Beth Fledelius2, Volker Schmidt3, Gyızı L. Kaján 4 and Rachel E. 5
Marschang1* 6
7
1 Institut für Umwelt- und Tierhygiene, Hohenheim University, Garbenstr. 30, 70599 8
Stuttgart, Germany 9
2 Small Animal Clinic „Hjortekaer”, 2800 Kgs Lyngby, Denmark 10
3 Bird and Reptile Klinik of Leipzig University, 04103 Leipzig, Germany 11
4 Veterinary Medical Research Institute of the Hungarian Academy of Sciences, Hungária 12
krt. 21., 1143 Budapest, Hungary 13
14
*Corresponding author: Tel.: +49 711 459 22468; fax: +49 711 459 22431. E-mail address: 15
17
Abstract 18
A consensus nested PCR was used to screen diagnostic samples from approximately 70 19
reptiles for the presence of adenoviruses (AdV) in the years 2006-2007. Classical virus 20
isolation methods were also used with all samples. After adenoviruses were detected in a 21
group of helodermatid lizards in a Danish zoo, a follow up study was also carried out on 22
lizards from this group (10 Mexican beaded lizards and 24 Gila monsters) over the period of 23
a year. Adenoviruses were detected in a total of 26 lizards and snakes by PCR. The PCR 24
Manuscriptpe
er-0
0532
460,
ver
sion
1 -
4 N
ov 2
010

Page 2 of 27
Accep
ted
Man
uscr
ipt
2
amplicons from all positive animals were sequenced and the resulting polymerase gene 25
sequences were used for phylogenetic analysis. Altogether six Agamid AdVs were amplified, 26
with a minimal sequence variation between one another and between these and GenBank 27
Agamid AdVs. The sequence obtained from one of the Gila monsters is identical with the 28
GenBank Helodermatid AdV. In a snake collection we have detected a new AdV from an 29
Asp viper. All of the above mentioned adenoviruses cluster in the Atadenovirus genus. 30
However, the sequence from a new Varanid AdV detected in this study clusters outside this 31
genus. On cell culture, viruses were isolated from three of the AdV positive helodermatid 32
lizards (one Mexican beaded lizard and two Gila monsters) and identified as AdVs based on 33
electron microscopy and PCR and sequencing using cell culture supernatant. This is the first 34
report of the successful isolation of a lizard AdV. 35
36
Key words: Adenovirus, Atadenovirus, lizard, PCR, reptile, virus isolation 37
38
Introduction 39
Adenoviruses (AdVs) occur worldwide and have been described from representatives of five 40
classes of the group Vertebrata (Russell & Benkı, 1999). Current taxonomy of the family 41
Adenoviridae (Benkı et al., 2005) suggests a coevolutionary lineage of the viruses with their 42
hosts, and additional host switches. According to this theory classes of vertebrates can be 43
assigned to different genera of the virus family. For mammals this would be the genus 44
Mastadenovirus, for birds the genus Aviadenovirus, for reptiles the genus Atadenovirus, for 45
amphibians the genus Siadenovirus and for fish the proposed genus “Ichtadenovirus”. The 46
genus classification criteria are mainly based on genomic and genetic characteristics of these 47
viruses, and host specificity, other than that described above, has been postulated to indicate 48
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 3 of 27
Accep
ted
Man
uscr
ipt
3
host-switches in the evolutionary past (Harrach, 2000). All reptilian AdVs described so far 49
are members of the genus Atadenovirus (Wellehan et al., 2004; Benkı et al., 2006). 50
In reptiles, AdV infections have been detected by light and electron microscopy (EM) 51
examination or by in situ hybridization (ISH) (Ramis et al., 2000; Perkins et al., 2001) of 52
histopathological sections in a number of different species of the Diapsida class, including 53
one crocodile species from the Archosauria subclass (Jacobson et al., 1984), and several 54
agamid, varanid and chameleonid species as well as 10 snake species from the Squamata 55
order of the Lepidosauria subclass (Essbauer & Ahne, 2001; Wellehan et al., 2004). 56
Associated pathological lesions varied from enterohepatic inflammation (hepatitis, 57
oesophagitis, enteritis,) to splenitis, nephritis, pneumonia or encephalopathy. The primary 58
pathogenic role of these viruses was questioned in many cases in which they were detected 59
without signs of concurrent disease (Jacobson & Kollias, 1986; Jacobson & Gardiner, 1990; 60
Ogawa et al. 1992; Schumacher et al., 1994). However, the pathogenicity of an AdV for 61
reptiles was demonstrated in one case by an experimental transmission study (Jacobson et al., 62
1985). 63
In spite of the numerous detections of reptilian AdV infection by EM, ISH or, more recently, 64
by PCR there are very few reported cases in which the virus was successfully isolated. 65
Jacobson et al. (1985) obtained AdV from a boa constrictor (Boa constrictor) while Ahne and 66
his co-workers isolated an AdV strain from a royal python (Python regius) (Ogawa et al., 67
1992) and from a moribund corn snake (Elaphe guttata) showing clinical signs of pneumonia 68
(Juhasz & Ahne, 1992). This corn snake isolate was later randomly cloned and completely 69
sequenced (Farkas et al. 2002, 2008) and thus serves as a prototype for reptilian AdVs. A 70
sequence comparison of partial IVa2 and polymerase gene sequences of this prototype virus 71
with those of three AdV isolates from other German snakes showed that they were identical 72
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 4 of 27
Accep
ted
Man
uscr
ipt
4
(Marschang et al., 2003). Although adenovirus infections are frequently described in lizards, 73
no virus has been isolated from a lizard in cell culture to date. 74
Wellehan et al. (2004) designed two degenerate primer pairs based on the consensus 75
sequence of the polymerase genes of different adenovirus types from three genera. This 76
nested PCR system has been shown to be an efficient tool for surveying adenovirus infections 77
of all genera in a wide range of animals, among them reptiles (Zsivanovits et al., 2006; Benkı 78
et al., 2006). In spite of the degenerate primers, direct sequencing of the products is possible, 79
and phylogenetic analysis of the sequences can help to determine the virus type. Wellehan 80
and co-workers have used this system to describe 6 novel lizard adenoviruses from seven 81
host species. In both of the above mentioned studies the phylogenetic analysis of the short 82
(ca. 300 bp) polymerase segments clearly clustered all reptilian AdVs within the 83
Atadenovirus genus, giving further support for the coevolution theory (Harrach, 2000). 84
However, the sequence recently obtained from a Sulawesi tortoise using the same PCR 85
(Wellehan, unpublished; GenBank Accession No: EU056826) clusters in the Siadenovirus 86
genus, showing that Testudines AdVs may differ from this scheme. 87
In our diagnostic laboratory we use this consensus nested PCR, together with classical virus 88
isolation methods for the detection of reptilian adenoviruses. The present report describes the 89
use of these methods to detect and characterize adenoviruses from different lizards and 90
snakes, resulting in the description of new reptilian AdVs and the first isolation of a lizard 91
AdV in cell culture. 92
93
Materials and Methods 94
Samples 95
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 5 of 27
Accep
ted
Man
uscr
ipt
5
Routine diagnostic samples from a total of about 60 lizards and snakes with case histories 96
considered suspicious for AdV infection were sent to our laboratory during 2006 and the 97
beginning of 2007. Twelve additional reptilian samples sent for other virus tests were 98
surveyed “blind” for the presence of AdVs as well, using the PCR protocol described later. 99
DNA or RNA extracts of different organs or swabs were pooled for the same animal or same 100
batch and treated as one. The animals that were positive for AdVs are listed in Table 1. 101
In the case of the AdV positive Gila monsters (Heloderma suspectum) and Mexican beaded 102
lizards (Heloderma horidum) (Nos. 7-11 in Table 1), follow-up tests were performed at the 103
Danish zoo in which these animals were kept after the detection of AdV. Five consecutive 104
tests were performed in the group consisting of Gila monsters, Mexican beaded lizards and 105
leopard tortoises (Geochelone pardalis). First intestine, heart, liver and liver-swab samples 106
from a dead Mexican beaded lizard (No.7) were tested when an outbreak started among a 107
group of juveniles. Two weeks later oral and cloacal swabs were taken from some survivors. 108
This time two beaded lizards (No.8) and a Gila monster (No.9) were tested. Another two 109
months later a Gila monster and two leopard tortoises from the same animal group were 110
swabbed (these samples are not listed in Table 1). Six months after the outbreak, two 111
asymptomatic young Gila monsters from a separate enclosure were swabed for a follow up 112
study (No.10). Finally, another half year later 21 newly introduced Gila monsters and 7 fresh 113
Mexican beaded lizards were orally swabbed, before introducing them to the facility (No. 114
11). 115
Virus isolation 116
Isolation of viruses was attempted from all samples on the iguana heart cell line (IgH-2, 117
ATCC: CCL-108) and/or on Russell’s viper heart cells (VH-2, ATCC: CCL-140) and/or 118
Terrapene heart cells (TH-1, ATCC: CCL-50). Small pieces of tissues or the cotton heads of 119
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 6 of 27
Accep
ted
Man
uscr
ipt
6
swabs were sonicated in 3ml Dulbecco’s modified Eagle’s medium (DMEM) (Biochrom AG, 120
Berlin, Germany) supplemented with antibiotics. The samples were centrifuged at low speed 121
(2000xg, 10 min) for the removal of cell-debris and bacteria, then 200 µl of the homogenate 122
was inoculated onto approximately 70% confluent cell monolayers in 30 mm diameter 123
Cellstar® tissue culture dishes (Greiner Bio-One GmbH, Frickenhausen, Germany). After 124
incubating for 2 hours at 28 ºC, 2ml nutrient medium (DMEM supplemented with 2% foetal 125
calf serum, 1 % non-essential amino acids and antibiotics) was added to each dish. Cells were 126
examined for cytopathic effects (CPE) approximately every 3 days with an inverted light 127
microscope (Wilovet, Wetzlar, Germany), and dishes were frozen when extended CPE was 128
seen. Dishes showing no CPE were frozen after 2 weeks of incubation for blind passaging. 129
Two additional passages were performed from each dish after a freeze and thaw cycle and 130
low speed centrifugation. 131
Electron microscopy 132
Cell culture supernatant was negative-stained with 2% potassium phosphotungstate (pH 7.3) 133
on 3.05 mm copper grids (Plano GmbH, Wetzlar, Germany), and examined with a JEM-1011 134
transmission electron microscope (JEOL, Japan) for the presence of viral particles. Electron 135
microscopic examinations were carried out by Valerij Akimkin at the Chemische und 136
Veterinäruntersuchungsamt (CVUA) Stuttgart, Germany. 137
PCR and sequencing 138
DNA was extracted from sample homogenates using the DNAeasy® kit (Qiagen GmbH, 139
Hilden, Germany). Nested PCR was done in 25 µl reaction mixtures, containing 1x 140
concentration of Taq Buffer with (NH4)2SO4, 1.5 mM of MgCl2, 200 µM of each dNTP, and 141
0.6 U of Taq polymerase (all from MBI Fermantas, St Leon-Rot, Germany). 1 µM from both 142
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 7 of 27
Accep
ted
Man
uscr
ipt
7
forward and reverse primers of the outer or inner pairs (Wellehan et al., 2004) were used in 143
the first or second rounds of the reaction respectively. In each round, 45 cycles were carried 144
out. In the first round 1 µl DNA extract, in the second round 2.5 µl first round amplicon 145
served as a template. Products were separated on 1.5 % agarose gels (Bioenzym, Oldendorf, 146
Germany) in TAE puffer containing 0.5 µg/ml ethidium-bromide and visualised under 320 147
nm UV light. 148
Gel purified PCR amplicons (Invisorb Spin DNA Extraction Kit; Ivitek GmbH, Berlin, 149
Germany) were sequenced directly using a Big-Dye terminator kit v.1.1 and analysed on an 150
ABI prism 310 automated DNA sequencer (both Applied Biosystems, Foster City, USA). 151
Analysis of sequences 152
Raw sequences were processed by the ABI Sequence Analysis Programme 5.1.1 (Applied 153
Biosystems, Foster City, USA) then edited, assembled and compared using the STADEN 154
Package version 2003.0 Pregap4 and Gap4 programmes (Bonfield et al., 1995). The 155
sequences were compared to the data in GenBank (National Center for Biotechnology 156
Information, Bethesda, USA) online (www.ncbi.nih.gov) using BLASTN and BLASTX 157
options. Homologous sequences were retrieved from GenBank through the non-redundant 158
AdV database of the Molecular Virology Group at the Veterinary Medical Research Institute, 159
Budapest (www.vmri.hu/~harrach). Multiple alignment of sequences was performed with 160
ClustalW algorithm of the BioEdit Sequence Alignment Editor programme (Hall, 1999) 161
using default settings. This alignment was further used for phylogenetic calculations in the 162
PHYLIP program Package version 3.6. (Felsenstein, 1989) applying various methods 163
(maximum likelihood and distance based) to obtain an optimal tree. With a bootstrap of 100 164
replicates, the Dayhoff matrix algorithm for the protein distance calculations and the Fitch-165
Margoliash method for the tree calculations provided the most robust tree. 166
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 8 of 27
Accep
ted
Man
uscr
ipt
8
167
Results 168
Isolation of the viruses 169
Viruses were isolated from three animals out of the total of 26 that tested adenovirus positive 170
by PCR during the course of the study (Tab. 1). All three of these were Heloderma spp. (one 171
Mexican beaded lizard and two Gila monsters) from a single zoo. During this study and the 172
follow-up investigation at this zoo, a total of 34 Heloderma spp. specimens (10 Mexican 173
beaded lizards and 24 Gila monsters) were tested for adenoviruses. The isolated viruses 174
caused a CPE with rounding and detachment of some cells. The samples from the dead 175
Mexican beaded lizard (No.7) were strongly positive on IgH-2 cells in the first passage. 176
Liver, intestine, liver-swab and heart samples all showed extensive CPE after 4, 14, 24 and 177
30 days post inoculation, respectively (Fig. 1). Oral and clocal swabs of a Gila monster tested 178
6 months later (sample No.10) and one year later (sample No.11) showed a similar CPE on 179
IgH-2 cells. All of the isolates could be passaged. The isolated viruses were identified as 180
AdVs by PCR amplification of virus from the cell culture supernatant as described above as 181
well as by electron microscopic examination of the cell culture supernatants. Non-enveloped 182
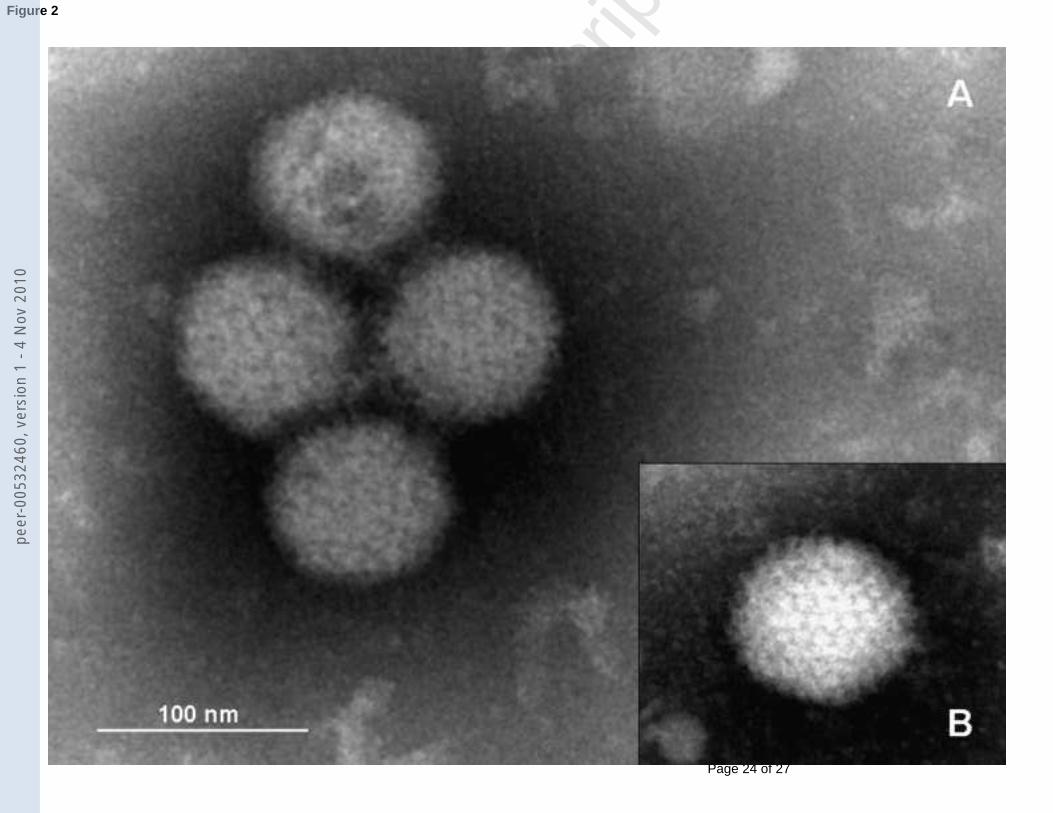
icosahedral particles of approximately 80 nm were detected with negative staining in both the 183
Mexican beaded lizard and the Gila monster isolates (Fig. 2). 184
Consensus nested PCR 185
We have found AdVs in a total of 26 diagnostic samples during 2006 and beginning of 2007. 186
AdVs were detected from a total of 6 bearded dragons. In three cases (No.1, No.2 and No.6) 187
virus was detected from both oral and cloacal swabs, in one case (No.5) only from a cloacal 188
swab, in one case (No.3) from mixed tissue and faeces, and in one case (No.4) from several 189
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 9 of 27
Accep
ted
Man
uscr
ipt
9
different tissues (lung, liver and kidney) (Table 1). In the case of one dead animal (No.4) and 190
a chronically sick one (No.5) we also tested oral and cloacal swabs of the partner dragons 191
from the same enclosure, but they were all negative in the AdV PCR. 192
In the Danish zoo cases, among the Mexican beaded lizards all of the tissues and swabs tested 193
from No.7, and the swabs from No.8 were positive for AdVs by PCR. Oral swabs as solitary 194
probes taken from several Gila monsters from the same enclosure collected some time after 195
the original detection were also all positive (No.8 & No.9). However, the companion beaded 196
lizard and two leopard tortoises from the same enclosure tested two months after the original 197
detection gave negative PCR results (data not shown). The latest sampling, of 28 new 198
quarantined Heloderma spp. specimens 1 year after the outbreak detected 12 positives among 199
the Gila monsters (No.11). 200
The kidney and spleen tissue samples from the emerald monitor (Varanus prasinus) that died 201
with a hepatitis were PCR positive although the liver was negative (No.12). 202
AdVs were also detected in a snake collection with no specific signs indicative of AdV 203
infection. No.13 was a batch of samples from snakes at a collection containing nine 204
specimens from two asp viper subspecies (Vipera aspis aspis, Vipera a. francisciredi) and 205
two striped Aesculapian rat snakes (Elaphe lineata). The solitary AdV positive sample from 206
this batch was an asp viper which died suddenly and organ samples of intestine, liver, kidney 207
and lung were sent in for paramyxovirus (PMV) testing, along with oral and cloacal swabs 208
from each living companion. PMV was detected in three of these survivors but not in the 209
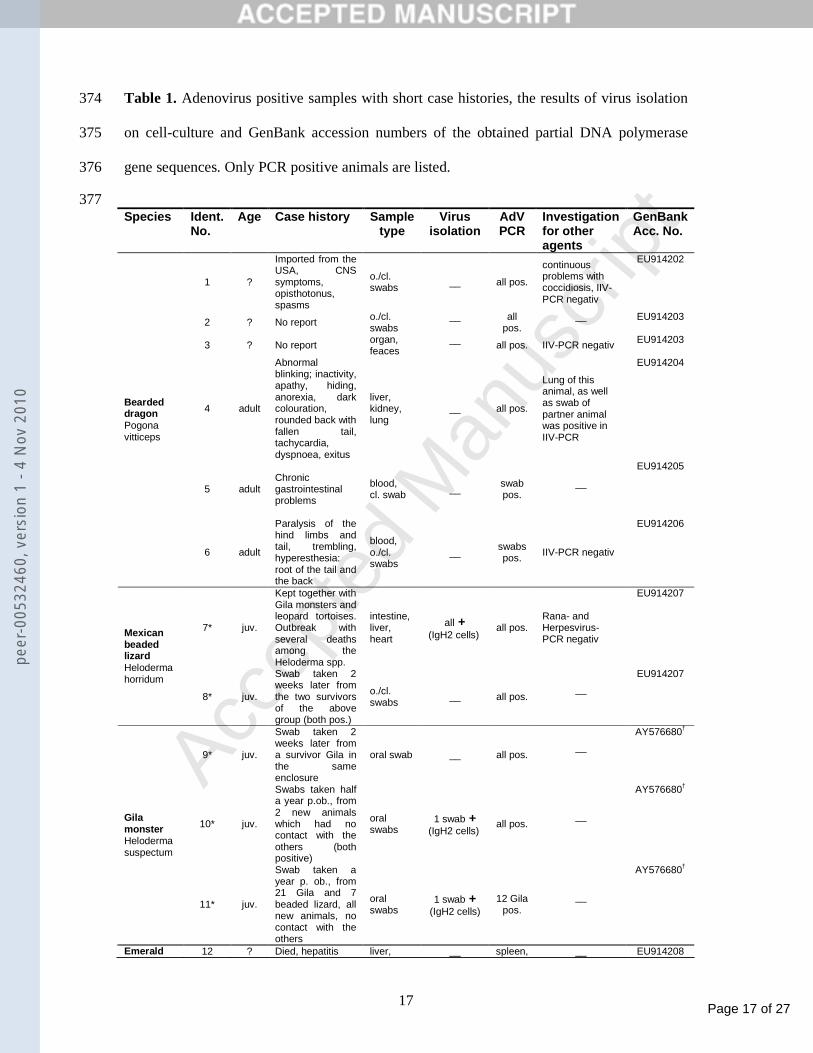
tissues of the dead snake. However, a pooled organ sample of this animal was positive in the 210
AdV consensus nested PCR. 211
In many cases, multiple bands of nonspecific amplicons were visible above the approx. 320 212
bp long second round specific product, making the evaluation more difficult. In the case of 213
No.8, these bands were equally strong as the specific one. 214
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 10 of 27
Accep
ted
Man
uscr
ipt
10
Sequence analysis 215
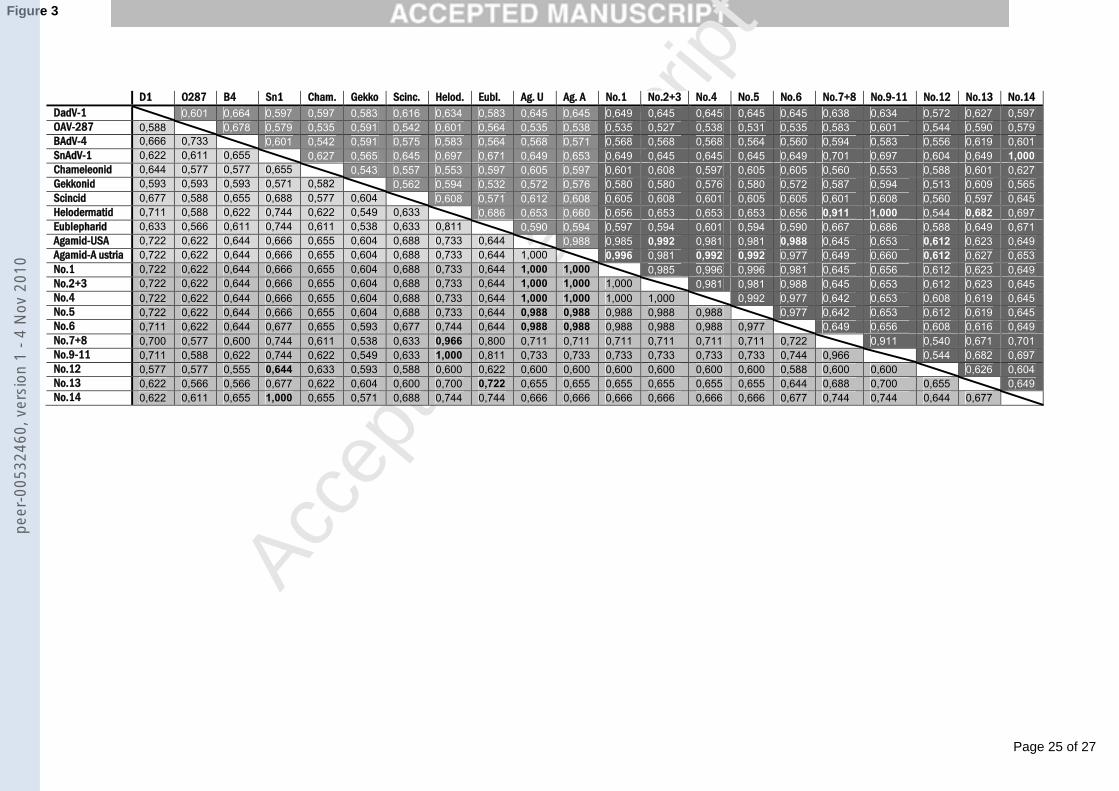
The second round PCR products were all 272 nt long after editing out the primer sequences. 216
These sequences were used for further analysis. An identity matrix of the sequences is shown 217
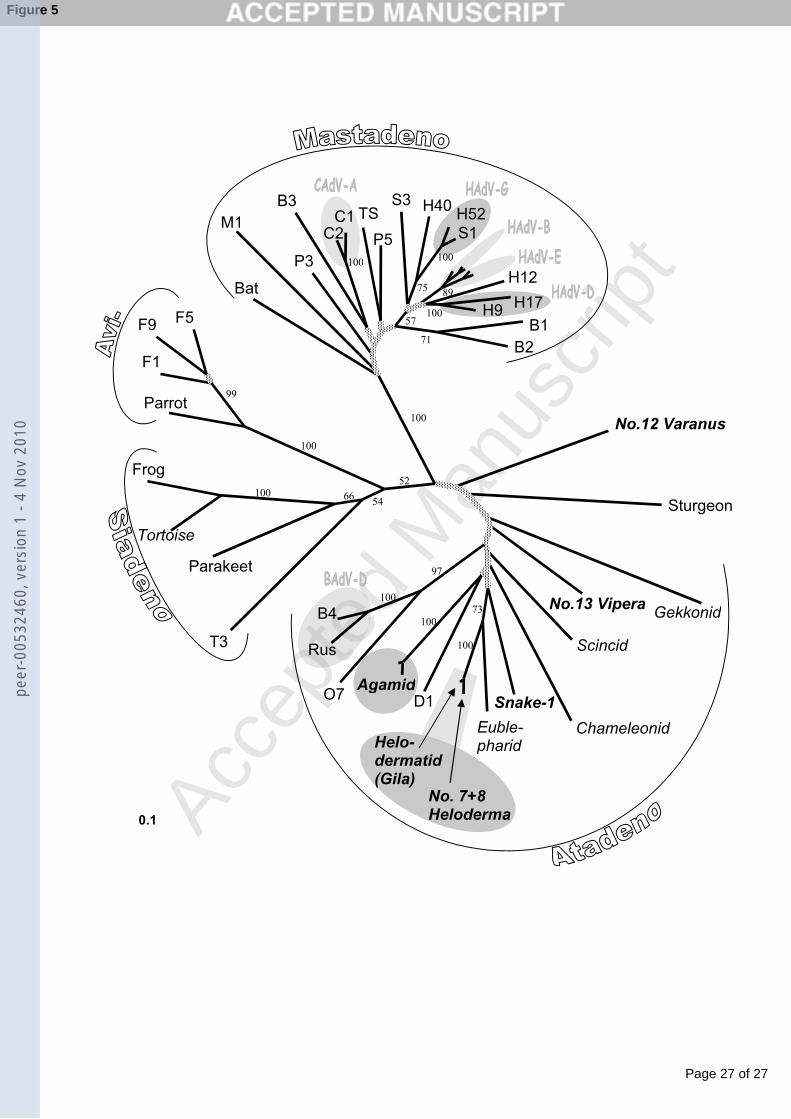
in Fig. 3, an alignment of the amino acid sequences in Fig. 4 and a phylogenetic tree based on 218
this alignment is shown in Fig.5. The sequences were submitted to GenBank and have been 219
assigned the accession numbers EU914202 to EU914209 (Table 1). 220
The six agamid AdVs detected are most similar to one another. There is a maximum of 6 nt 221
differences between the sequences, with at most 2 non-silent mutations. In the data set of the 222
follow-up study from the Heloderma spp., the sequences from the Mexican beaded lizards 223
were always distinctly different from those of the Gila monsters. All Gila sequences in the 224
batches No.10 and No.11 were identical with the corresponding GenBank Helodermatid AdV 225
sequence. Similarly, the sequences of all of the Mexican beaded lizard samples were identical 226
with one another, but show 24 nt differences compared to the Helodermatid AdV sequence. 227
Yet only 3 of these were non-silent mutations. The partial sequence obtained from the 228
emerald monitor AdV No.12 has the highest BLASTX values with the snake AdV sequence, 229
whereas the identity matrix of the nucleotide sequences shows the highest value with the 230
Agamid AdVs. The asp viper AdV (sample No.13) has the greatest similarity to the 231
Eublepharid AdV and the Helodermatid AdV with approx. 65-68 % and 70-72 % similarity 232
values on the nucleotide and amino acid level respectively. The partial polymerase sequence 233
from the AdV isolate from a Boa constrictor used as a positive control in all PCRs (No. 14) 234
was identical to the corresponding region of the SnAdV-1. 235
236
Discussion 237
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 11 of 27
Accep
ted
Man
uscr
ipt
11
This is the first report of the successful isolation of a lizard AdV. Although there have been 238
several papers describing the occurrence of AdVs in eublepharid, geckonid, varanid, 239
chameleonid and agamid Squamates (Essbauer and Ahne, 2001; Farkas et al., 2002; 240
Wellehan et al,. 2004), these viruses have never been isolated. All reptilian isolates to date 241
are from snakes, and there is evidence that these might represent the same virus (Marschang 242
et al., 2003). Isolation of the two different Helodermatid AdVs, in one case from a Mexican 243
beaded lizard and two cases from Gila monsters is therefore an important step in the further 244
study of AdVs of reptiles. PCR amplified partial sequence comparison is possible with field 245
samples, but virus isolation is essential for physico-chemical and ultrastructural 246
characterisation as well as for pathogenicity studies. 247
The comparison of the partial polymerase sequences of our helodermatid sequences with that 248
in GenBank has revealed that the Gila monsters and Mexican beaded lizards posess two 249
different AdVs. The GenBank helodermatid AdV sequence from a Gila monster in the USA 250
is identical to our Danish Gila sequences, whereas our beaded lizard isolate and sample 251
sequences from the same enclosure differ from these. This finding supports the host-252
specificity coevolution-cospeciation theory of AdVs (Harrach, 2000). However, this strict 253
host-specificity is not supported by the identical sequences found in isolates from different 254
snakes (Boa sample No.14 and SnAdV-1 from a corn snake; also in Marschang et al., 2003; 255
python, corn snake & rat snake samples in Benkı et al., 2006). The difference (8.9% aa and 256
3.3% nt) between the gene portions of the two helodermatid viruses are relatively small. 257
Compared to those between other AdVs, these viruses seem to represent the same species. 258
Further sequencing and/or serological testing are needed for further characterization of these 259
isolates. 260
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 12 of 27
Accep
ted
Man
uscr
ipt
12
Our six bearded dragon AdV sequences possess less than 2.5% nt variation in the examined 261
region, and thus, together with data from GenBank and additional sequences from Hungary 262
(Benkı et al, 2006; Farkas, personal communication) they provide further support that a 263
single agamid AdV is circulating in the bearded dragon collections across Europe and the 264
USA. 265
There were several interesting aspects of the AdV found in the emerald monitor (No.12). 266
Although the histological examination of this animal showed hepatitis, no AdV was detected 267
in the liver by PCR. The spleen and kidney were both PCR positive and the sequence 268
obtained from these amplicons revealed that this varanid AdV is very different from every 269
earlier described reptilian AdVs. Using either nucleotide or amino acid sequence in the 270
distance matrix analysis the virus clustered not in but beside the Atadenovirus genus, equally 271
distant to the proposed “Ichtadenovirus” genus and the Mastadenovirus genus. Yet the 272
sequence is short and the bootstrap values at this branching of the tree are under 50 (Fig. 5). 273
Further sequencing is necessary to determine the exact taxonomic position of this virus. 274
The asp viper sample (No.13) is a good example for the successful detection of new AdVs in 275
blind surveys. This snake was the only animal that died in a collection of snakes in which a 276
PMV was detected although only the companion animals were positive in a PMV RT-PCR. 277
The phylogenetic analysis of the polymerase gene portion of the AdV in this animal has 278
clustered this virus into the Atadenovirus genus, but representing a new species: Viperid 279
AdV. 280
Each of the viruses presented above have a balanced A+T content (41.7% - 57.5%) in the 281
sequenced region which might indicate that they have coevolved over a longer period of time 282
together with their hosts (Farkas et al., 2002; Wellehan et al., 2004). 283
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 13 of 27
Accep
ted
Man
uscr
ipt
13
In conclusion, while the majority of our sequence data support the coevolution-cospeciation 284
theory and the reptilian origin of atadenoviruses, a close look at individual reptilian AdV 285
shows that this theory may not explain all of the AdV found in reptiles. One open question is 286
the occurance of the same AdV in different snake hosts. The other is posed by the Varanid 287
AdV which does not seem to belong to the Atadenovirus genus. Additional work is 288
necessary, both to obtain additional taxonomic information on the AdVs described here as 289
well as to characterize further reptilian AdV. 290
291
Acknowledgements 292
We would like to thank Silvia Speck and Christa Shäfer for their excellent technical 293
assistance. Samples were sent to our laboratory by Dr. Marc Hoferer from the Chemisches- 294
und Veterinär Untersuchungsamt (CVUA), Stuttgart-Fellbach; Dr. Karina Mathes and Ms. 295
Günther, Small Animal Clinic of the Veterinary College of Hannover; Dr. Jutta Wiechert, 296
Veterinary Praxis, Mainz; Dr. Bernd Dörsch, Small Animal Praxis, Kiel, and Dr. Michael 297
Pees, Bird and Reptile Klinik of Leipzig University, all in Germany. Their help is gratefully 298
acknowledged. We also thank Valerij Akimkin at the CVUA, Stuttgart-Fellbach, Germany, 299
for his help with the electron microscopic examination and Dr. Balázs Harrach at the 300
Veterinary Medical Research Institute, Budapest, Hungary for his advice in taxonomical 301
questions. Dr. Kaján’s work was partially supported by the National Office for Research and 302
Technology, József Öveges programme, and the Hungarian Scientific Research Fund, grant 303
OTKA K 67781. 304
305
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 14 of 27
Accep
ted
Man
uscr
ipt
14
References 306
Benkı, M., Élı, P., Ursu, K., Ahne, W., LaPatra, S.E., Thomson, D., Harrach, B., 2002. First 307
molecular evidence for the existence of distinct fish and snake adenoviruses. J. Virol. 308
76, 10056-9. 309
Benkı, M., Harrach, B., Both, G.W., Russell, W.C., Adair, B.M., Ádám, É., de Jong, J.C., 310
Hess, M., Johnson, M., Kajon, A., Kidd, A.H., Lehmkuhl, H.D., Li, Q.G., Mautner, 311
V., Pring-Akerblom, P., Wadell, G., 2005. Family Adenoviridae. In: Fauquet, C.M. 312
(Ed.), Virus Taxonomy. Eighth Report of the International Committee on Taxonomy 313
of Viruses, Academic Press, New York, pp. 213-228. 314
Benkı, M., Kaján, G.L., Jánoska, M., Kovács, E.R., Wellehan, J.F.X., Zsivanovits, P., 315
Cough, R.E., Blahak, S., Schirrmeier, H., Bakonyi, T., Harrach, B., 2006. How to find 316
“new” adenoviruses. In: Leitão, A., Martins, C. (Eds.), Proceedings of the 7th 317
International Congress of Veterinary Virology. Ministério da Agricultura, Lisboa, 318
Portugal, p. 103. 319
Bonfield, J.K., Smith, K.F., Staden, R., 1995. A new DNA sequence assembly program. 320
Nucleic Acids Res. 24, 4992-4999. 321
Essbauer, S., Ahne, W., 2001. Viruses of lower vertebrates. J. Vet. Med. B, Infectious 322
Diseases and Veterinary Public Health 48, 403-475. 323
Farkas, S.L., Benkı, M., Élı, P., Ursu, K., Dán, Á., Ahne, W., Harrach, B., 2002. Genomic 324
and phylogenetic analyses of an adenovirus isolated from a corn snake (Elaphe 325
guttata) imply a common origin with members of the proposed new genus 326
Atadenovirus. J. Gen. Virol. 83, 2403-2410. 327
Farkas, S.L., Harrach, B., Benkı, M., 2008. Completion of the genome analysis of snake 328
adenovirus type 1, a representative of the reptilian lineage within the novel genus 329
Atadenovirus. Vir. Res. 132, 132-139. 330
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 15 of 27
Accep
ted
Man
uscr
ipt
15
Felsenstein, J., 1989. PHYLIP - Phylogeny Inference Package. Cladistics 5, 164-166. 331
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis 332
program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 41, 95-98. 333
Harrach, B., 2000. Reptile adenoviruses in cattle? Acta Vet. Hung. 48, 484-490. 334
Jacobson, E.R., Kollias, G.V., 1986. Adenovirus-like infection in a savannah monitor. J. Zoo 335
Anim. Med. 17, 149-151. 336
Jacobson, E.R., Gardiner, C.H., 1990. Adeno-like virus in esophageal and tracheal mucosa of 337
a Jackson’s chameleon (Chamaeleo jacksoni). Vet. Path. 27, 210-212. 338
Jacobson, E.R., Gardiner, C.H., Foggin, C.M., 1984. Adenovirus-like infection in two Nile 339
crocodiles. J. Am.Vet. Med. Assoc. 185, 1421-1422. 340
Jacobson, E.R., Gaskin, J.M., Gardiner, C.H., 1985. Adenovirus-like infection in a boa 341
constrictor. J. Am. Vet. Med. Assoc. 187, 1226-1227. 342
Juhász, A., Ahne, W., 1992. Physico-chemical properties and cytopathogenicity of an 343
adenovirus-like agent isolated from corn snake (Elaphe guttata). Arch. Virol. 130, 344
429-439. 345
Marschang, R.E., Mischling, M., Benkı, M., Papp, T., Harrach, B., Böhm, R., 2003. 346
Evidence for wide-spread atadenovirus infection among snakes. In: Jestin, A., 347
Clement, G. (Eds.), Virus persistence and evolution. Proceedings of the 6th 348
International Congress of Veterinary Virology. ZOO-POLE développement –ISPAIA, 349
Ploufragan, France. p. 152. 350
Ogawa, M., Ahne, W., Essbauer, S., 1992. Reptilian viruses: adenovirus-like agent isolated 351
from royal python (Python regius). J. Vet. Med.. B, Infectious Diseases and 352
Veterinary Public Health 39, 732-736. 353
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 16 of 27
Accep
ted
Man
uscr
ipt
16
Perkins, L.E, Campagnoli, R.P., Harmon, B.G., Gregory, C.R., Steffens, W.L., Latimer, K., 354
Clubb, S., Crane, M., 2001. Detection and confirmation of reptilian adenovirus 355
infection by in situ hybridization. J. Vet. Diagn. Invest. 13, 365-368. 356
Ramis, A., Fernandez-Bellon, H., Majo, N., Martinez-Silvestre, A., Latimer, K., Campagnoli, 357
R., Harmon, B.G., Gregory, C.R., Steffens, W.L., Clubb, S., Crane, M., 2000. 358
Adenovirus hepatitis in a boa constrictor (Boa constrictor). J. Vet. Diagn. Invest. 12, 359
573-576. 360
Russell, W.C., Benkı, M., 1999. Animal adenoviruses. In: Granoff, A., Webster, R.G. (Eds.), 361
Encyclopedia of Virology. Academic Press, New York, pp. 14-21. 362
Schumacher, J., Jacobson, E.R., Burns, R., Tramontin, R.R., 1994. Adenovirus-like infection 363
in two rosy boas (Lichanura trivirgata). J. Zoo and Wildlife Med. 25, 461-465. 364
Wellehan, J.F.X., Johnson, A.J., Harrach, B., Benkı, M., Pessier, A.P., Johnson, C.M., 365
Garner, M.M., Childress. A., Jacobson, E.R., 2004. Detection and analysis of six 366
lizard adenoviruses by consensus primer PCR provides further evidence of a reptilian 367
origin for the atadenoviruses. J Virol. 78, 13366-9. 368
Zsivanovits, P., Monks, D.J., Forbes, N.A., Ursu, K., Raue, R., Benkı, M., 2006. 369
Presumptive identification of a novel adenovirus in a Harris hawk (Parabuteo 370
unicinctus), a Bengal eagle owl (Bubo bengalensis), and a Verreaux’s eagle owl 371
(Bubo lacteus). J. Avian Med. Surgery 20, 105-112. 372
373
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0
Page 17 of 27
Accep
ted
Man
uscr
ipt
17
Table 1. Adenovirus positive samples with short case histories, the results of virus isolation 374
on cell-culture and GenBank accession numbers of the obtained partial DNA polymerase 375
gene sequences. Only PCR positive animals are listed. 376
377 Species Ident.
No. Age Case history Sample
type Virus
isolation AdV PCR
Investigation for other agents
GenBank Acc. No.
1 ?
Imported from the USA, CNS symptoms, opisthotonus, spasms
o./cl. swabs __ all pos.
continuous problems with coccidiosis, IIV-PCR negativ
EU914202
2 ? No report o./cl. swabs
__
all pos.
__
EU914203
3 ? No report organ, feaces
__
all pos. IIV-PCR negativ EU914203
4 adult
Abnormal blinking; inactivity, apathy, hiding, anorexia, dark colouration, rounded back with fallen tail, tachycardia, dyspnoea, exitus.
liver, kidney, lung
__ all pos.
Lung of this animal, as well as swab of partner animal was positive in IIV-PCR
EU914204
5 adult
Chronic gastrointestinal problems
blood, cl. swab __ swab
pos. __
EU914205
Bearded dragon Pogona vitticeps
6 adult
Paralysis of the hind limbs and tail, trembling, hyperesthesia: root of the tail and the back
blood, o./cl. swabs
__ swabs pos. IIV-PCR negativ
EU914206
7* juv.
Kept together with Gila monsters and leopard tortoises. Outbreak with several deaths among the Heloderma spp.
intestine, liver, heart
all + (IgH2 cells)
all pos. Rana- and Herpesvirus-PCR negativ
EU914207
Mexican beaded lizard Heloderma horridum
8* juv.
Swab taken 2 weeks later from the two survivors of the above group (both pos.)
o./cl. swabs
__
all pos.
__
EU914207
9* juv.
Swab taken 2 weeks later from a survivor Gila in the same enclosure
oral swab
__
all pos. __
AY576680†
10* juv.
Swabs taken half a year p.ob., from 2 new animals which had no contact with the others (both positive)
oral swabs
1 swab + (IgH2 cells)
all pos. __
AY576680†
Gila monster Heloderma suspectum
11* juv.
Swab taken a year p. ob., from 21 Gila and 7 beaded lizard, all new animals, no contact with the others
oral swabs
1 swab + (IgH2 cells)
12 Gila pos.
__
AY576680†
Emerald 12 ? Died, hepatitis liver, __ spleen, __ EU914208
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0
Page 18 of 27
Accep
ted
Man
uscr
ipt
18
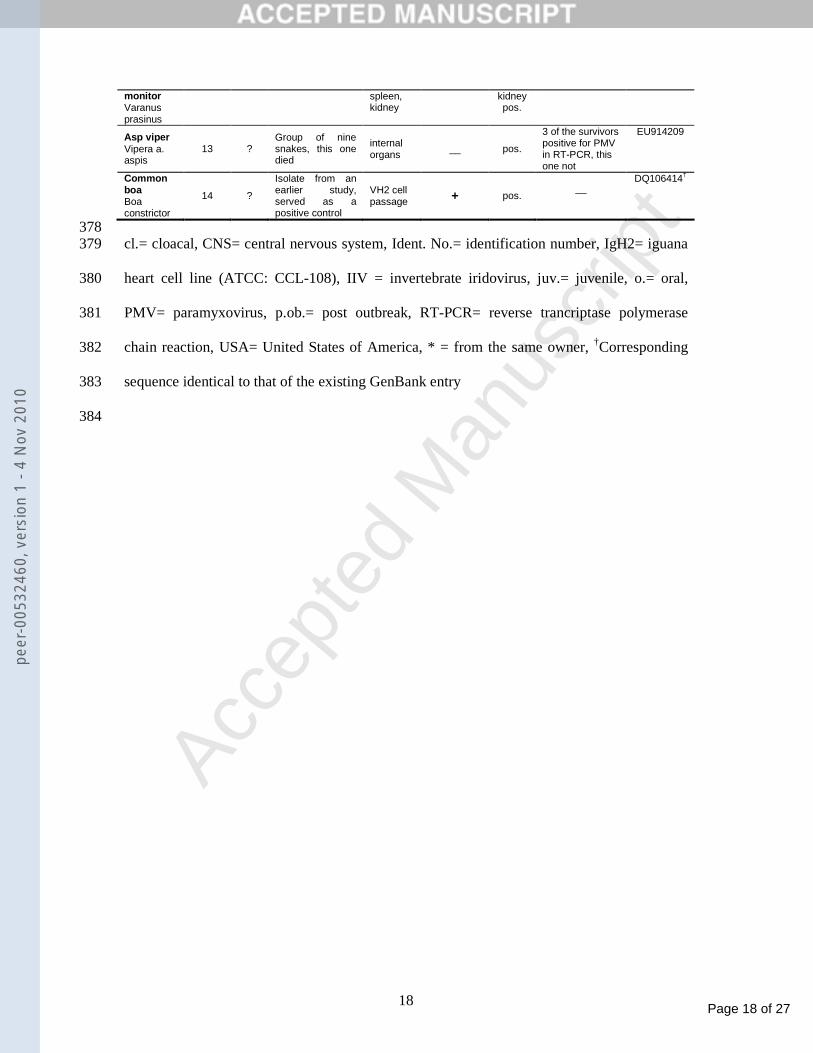
monitor Varanus prasinus
spleen, kidney
kidney pos.
Asp viper Vipera a. aspis
13 ? Group of nine snakes, this one died
internal organs
__ pos.
3 of the survivors positive for PMV in RT-PCR, this one not
EU914209
Common boa Boa constrictor
14 ?
Isolate from an earlier study, served as a positive control
VH2 cell passage + pos.
__
DQ106414†
378 cl.= cloacal, CNS= central nervous system, Ident. No.= identification number, IgH2= iguana 379
heart cell line (ATCC: CCL-108), IIV = invertebrate iridovirus, juv.= juvenile, o.= oral, 380
PMV= paramyxovirus, p.ob.= post outbreak, RT-PCR= reverse trancriptase polymerase 381
chain reaction, USA= United States of America, * = from the same owner, †Corresponding 382
sequence identical to that of the existing GenBank entry 383
384
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 19 of 27
Accep
ted
Man
uscr
ipt
19
Figure captions: 385
386
Fig. 1. Adenovirus isolate from a Gila monster (Heloderma suspectum). Cytopathic effect 387
(CPE) five days after inoculation. A- Negative control IgH-2. B- 3rd passage of an isolate 388
from an oral swab from case No.11 on IgH2 cells 5 days post inoculation. (Magnification: 389
400x) 390
Fig. 2. Electron micrographs of icosahedral non-enveloped virus particles in cell culture 391
supernatant from two Helodermatid isolates. (Both 3rd passage on IgH2 cells 5 days post 392
inoculation). A- Mexican beaded lizard isolate with identification No.7. B- Isolate from an 393
oral swab from Gila monster with identification No.10 394
Fig. 3. Identity matrix of the nucleotide (above diagonal, with white letters) and the deduced 395
amino acid (below diagonal, with black letters) sequences of the amplified DNA-polymerase 396
gene region. Accepted and proposed members of the Atadenovirus genus from GenBank are 397
marked with lighter background colouring compared to the sequences newly reported here 398
with darker grey background. Highest values for the new sequences compared to GenBank 399
sequences are shown in bold. For accession numbers see legend of Fig. 5; Agamid AdV, 400
Austria (AAY83284), Agamid AdV, USA (AY576678). 401
Fig. 4. Alignment of predicted partial DNA-polymerase sequences of the amplified region 402
from the atadenoviruses and the sturgeon adenovirus. (Identical sequences are represented 403
by one selected virus. Start of alignment corresponds to the codon at position 6458 of 404
HAdV-1 whole genomic nucleotide sequence, Accession No.: AF534906.) Sequences of 405
reptilian viruses are printed with capital letters, sequences new to science are highlighted 406
with bold printed names. OAdV-7 type species was taken as a null-sequence for the 407
comparison, identical amino acids are marked with dots in the rest of the alignment. Virus 408
species with more than one serotype are separated with lines and indicated with grey 409
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 20 of 27
Accep
ted
Man
uscr
ipt
20
background shading of the names. Vertical background shading of sequences in black and 410
grey refers to 80% identity and similarity of conserved amino acid positions respectively, 411
regarding the alignment of all available sequences for this region. (helodermatid and 412
agamid isolates show a maximum of 3 amino acid variations between one another, and are 413
therefore also grouped together as they most probably belong to one species.) For 414
abbreviations and GenBank accession numbers see legend of Fig. 5. 415
Fig 5. Phylogenetic distance tree of partial adenovirus DNA-polymerase amino acid 416
sequences. The tree was generated using the Protdist program with Dayhoff matrix, followed 417
by Fitch program with global rearrangements. Bootstrap values above 50 (for 100 418
replications) are shown beside the branches, lower value branchings are shown with dotted 419
lines. Accepted and presumed virus species are indicated with grey circles, genera with half 420
circles. Names of reptilian viruses are printed in italics, and those representing sequences 421
from our study (too), are in bold italics. 422
Abbreviations and GenBank accession numbers (in brackets): B=Bovine, C=Canine, 423
D=Duck, F=Fowl, H=Human, M=Murine, O=Ovine, P=Porcine, Rus=BadV-4 strain Rus, 424
S=Simian, T=Turkey, TS=Tree shrew 425
Agamid AdV (AY576678 and AAY83284), BAdV-1 (YP_094032), BAdV-2 (AP_000006), 426
BAdV-3 (AP_000026), BAdV-4 (AAK13183), BAdV-4 Rus (not yet released), Bat AdV 427
(AB303301), CAdV-1 (AAB05434), CAdV-2 (AAB38716), Chameleonid AdV 428
(AY576679), DAdV-1 (AP_000080), Eublepharid AdV (AY576677), FAdV-1 (AP_000410), 429
FAdV-5 (DQ159938), FAdV-9 (AC_000013), Frog (AAF86924), Gekkonid AdV 430
(AY576681), HAdV-9 (CAI05960), HAdV-12 (CAA51882), HAdV-17 (AP_000143), 431
HAdV-40 (AAC13953), HAdV-52 (ABK35035), HAdV-B=HAdV-3 (ABB17778) + HAdV-432
7 (AP_000539) + HAdV-35 (AAN17476), HAdV-E=HAdV-4 (AAS66917) + SAdV-25 433
(AP_000304), Helodermatid AdV (AAS89696), MAdV-1 (AP_000342), OAdV-7 434
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 21 of 27
Accep
ted
Man
uscr
ipt
21
(AAD45950), PAdV-3 (AB026117), PAdV-5 (AAK26504), Parakeet AdV (EU056825), 435
SAdV-1 (AAX19399), SAdV-3 (AAT84618), Scincid AdV (AY576682), Snake AdV 436
(AAL89790), Sturgeon AdV (not yet released), TAdV-3 (AAC64523), Tortoise AdV 437
(EU056826), TSAdV (YP_068060), 438
439
440
peer
-005
3246
0, v
ersi
on 1
- 4
Nov
201
0

Page 22 of 27
Accep
ted
Man
uscr
ipt
Figure 1Ape
er-0
0532
460,
ver
sion
1 -
4 N
ov 2
010

Page 23 of 27
Accep
ted
Man
uscr
ipt
Figure 1Bpe
er-0
0532
460,
ver
sion
1 -
4 N
ov 2
010
Page 24 of 27
Accep
ted
Man
uscr
ipt
Figure 2pe
er-0
0532
460,
ver
sion
1 -
4 N
ov 2
010
Page 25 of 27
Accep
ted
Man
uscr
ipt
D1 O287 B4 Sn1 Cham. Gekko Scinc. Helod. Eubl. Ag. U Ag. A No.1 No.2+3 No.4 No.5 No.6 No.7+8 No.9-11 No.12 No.13 No.14 DadV-1 0,601 0,664 0,597 0,597 0,583 0,616 0,634 0,583 0,645 0,645 0,649 0,645 0,645 0,645 0,645 0,638 0,634 0,572 0,627 0,597OAV-287 0,588 0,678 0,579 0,535 0,591 0,542 0,601 0,564 0,535 0,538 0,535 0,527 0,538 0,531 0,535 0,583 0,601 0,544 0,590 0,579BAdV-4 0,666 0,733 0,601 0,542 0,591 0,575 0,583 0,564 0,568 0,571 0,568 0,568 0,568 0,564 0,560 0,594 0,583 0,556 0,619 0,601SnAdV-1 0,622 0,611 0,655 0,627 0,565 0,645 0,697 0,671 0,649 0,653 0,649 0,645 0,645 0,645 0,649 0,701 0,697 0,604 0,649 1,000Chameleonid 0,644 0,577 0,577 0,655 0,543 0,557 0,553 0,597 0,605 0,597 0,601 0,608 0,597 0,605 0,605 0,560 0,553 0,588 0,601 0,627Gekkonid 0,593 0,593 0,593 0,571 0,582 0,562 0,594 0,532 0,572 0,576 0,580 0,580 0,576 0,580 0,572 0,587 0,594 0,513 0,609 0,565Scincid 0,677 0,588 0,655 0,688 0,577 0,604 0,608 0,571 0,612 0,608 0,605 0,608 0,601 0,605 0,605 0,601 0,608 0,560 0,597 0,645Helodermatid 0,711 0,588 0,622 0,744 0,622 0,549 0,633 0,686 0,653 0,660 0,656 0,653 0,653 0,653 0,656 0,911 1,000 0,544 0,682 0,697Eublepharid 0,633 0,566 0,611 0,744 0,611 0,538 0,633 0,811 0,590 0,594 0,597 0,594 0,601 0,594 0,590 0,667 0,686 0,588 0,649 0,671Agamid-USA 0,722 0,622 0,644 0,666 0,655 0,604 0,688 0,733 0,644 0,988 0,985 0,992 0,981 0,981 0,988 0,645 0,653 0,612 0,623 0,649Agamid-A ustria 0,722 0,622 0,644 0,666 0,655 0,604 0,688 0,733 0,644 1,000 0,996 0,981 0,992 0,992 0,977 0,649 0,660 0,612 0,627 0,653No.1 0,722 0,622 0,644 0,666 0,655 0,604 0,688 0,733 0,644 1,000 1,000 0,985 0,996 0,996 0,981 0,645 0,656 0,612 0,623 0,649No.2+3 0,722 0,622 0,644 0,666 0,655 0,604 0,688 0,733 0,644 1,000 1,000 1,000 0,981 0,981 0,988 0,645 0,653 0,612 0,623 0,645No.4 0,722 0,622 0,644 0,666 0,655 0,604 0,688 0,733 0,644 1,000 1,000 1,000 1,000 0,992 0,977 0,642 0,653 0,608 0,619 0,645No.5 0,722 0,622 0,644 0,666 0,655 0,604 0,688 0,733 0,644 0,988 0,988 0,988 0,988 0,988 0,977 0,642 0,653 0,612 0,619 0,645No.6 0,711 0,622 0,644 0,677 0,655 0,593 0,677 0,744 0,644 0,988 0,988 0,988 0,988 0,988 0,977 0,649 0,656 0,608 0,616 0,649No.7+8 0,700 0,577 0,600 0,744 0,611 0,538 0,633 0,966 0,800 0,711 0,711 0,711 0,711 0,711 0,711 0,722 0,911 0,540 0,671 0,701No.9-11 0,711 0,588 0,622 0,744 0,622 0,549 0,633 1,000 0,811 0,733 0,733 0,733 0,733 0,733 0,733 0,744 0,966 0,544 0,682 0,697No.12 0,577 0,577 0,555 0,644 0,633 0,593 0,588 0,600 0,622 0,600 0,600 0,600 0,600 0,600 0,600 0,588 0,600 0,600 0,626 0,604No.13 0,622 0,566 0,566 0,677 0,622 0,604 0,600 0,700 0,722 0,655 0,655 0,655 0,655 0,655 0,655 0,644 0,688 0,700 0,655 0,649No.14 0,622 0,611 0,655 1,000 0,655 0,571 0,688 0,744 0,744 0,666 0,666 0,666 0,666 0,666 0,666 0,677 0,744 0,744 0,644 0,677
Figure 3pe
er-0
0532
460,
ver
sion
1 -
4 N
ov 2
010

Page 26 of 27
Accep
ted
Man
uscr
iptHeAdV
AgAdV
BAdV-D
10 20 30 40 50 60 70 80 90 | | | | | | | | | | | | | | | | | | Ovine 7 salthplpygktlnafeanaqidyfqellqr-kekidyfdnsikpmivvadceppsldyldvlpplcskksgklcwsnetlinevltsidlDuck 1 ......m.f.r.edplt.sis.kt..dk.ds-pa.ls..ge.......y...y..p.ehv........r...r...t..p.lg..v.t...Bovine 4 ......m...r...p....ts..em.nm.ds-s.vls...pr..a..........t.e..................t..p....tv.....isol. Rus ......m...r...p....ia..el.sm.ds-svils...sg..a.v........t.e..................t.gp.v..tv.....SNAKE 1 ...S..M...P..SP.DSAVA.AE..RK.DG-QSELS...PD.F.......AF....HC..........R......T..P.LG....TV..CHAMELEON ...S..M.S.T.ESPTD.ALS.A...D..DK-PDQ.S..S-QV.....L...Y..A.AR.........RR......T..V.TA.A..TV..EUBLEPARID ...S..M...T..SP.DSSKAMAS..A..DG-.DCLS...PR.L....KV..F..P.YH..T..........R...T..P.LG..I.TV.IGEKKONID ......M.F.LPCEP.T..IH.RQ..Y..DEVGKP.S...ER......A...F...IKE......M.TR.G.....T..S.HM.I...V..SCINCID ......M.S.IP.DP.TSSIA.RK..NK.DE-PST.S...PD.F..V.....S..P.EQ.............R...T..P.EV.T..T...GILA M. ...S..M...L..SPLD.SVAMAR..DK.DS-T..LSF..KN.L....K...F..P.YH.............R...T..P.LG....TV.IHELOD.No8 ...S..M...L..SPLD.SVAMAR..DK.ES-TG.LSF..KD.L....K...F..P.YH.............R...T..P.LG....TV.IAGAMID ...S..M.C.R..PPLD.SIE.RR..DK.DK-PH..S...PNL.....A...I..P.NE..........A..R...T..P.VG........AGAMA No5 ...S..M.C.R..PPLD.SIE.RR..DK.DK-PH..S...PNL.....A...I..P.NE..........A..R...T..P.AG........AGAMA No6 ...S..M.C.R..PPLD.SVE.RR..DK.DK-PH..S...PNL.....A...I..P.NE..........A..R...T..P.VG........VIPER No13 ...S..M...R..GP.D.AIAMRH..A..DT-QK..S..EPT.N...ITV..F..DILR.........R...R...T..A.LG....TV.IVARAN.No12 ...?..M.S.NPESP.D.AIS.EN.RR..EQ-I.P.....PR......TV.AD...D.Q.........RH..R...T..P.RK....TV.I
Sturgeon ......m...yp.epl..sih.kl.....d.-.e..s..ndtv....lsi.ah..nin...t......rq..r...t..a..s.iv..l.c
HAdV-E
Figure 4pe
er-0
0532
460,
ver
sion
1 -
4 N
ov 2
010
Page 27 of 27
Accep
ted
Man
uscr
ipt
0.1
No.12 Varanus
Sturgeon
GekkonidNo.13 Vipera
Scincid
Chameleonid
Snake-1
Euble-pharidHelo-
dermatid (Gila)
D1Agamid
O7
Rus
B4
T3
Parakeet
Tortoise
Frog
Parrot
F1
F9 F5
Bat
M1
P3
B3
C2C1 TS
P5
S3 H40 H52S1
H12
H17H9B1
B2
No. 7+8 Heloderma
100 100
75 89
100
71
57
100
100
99
54
52
66100
100
97
100
100
73
Figure 5pe
er-0
0532
460,
ver
sion
1 -
4 N
ov 2
010
Related Documents











