www.newphytologist.org 65 Research Blackwell Publishing Ltd Patterns of asymmetry in the twining vine Dalechampia scandens (Euphorbiaceae): ontogenetic and hierarchical perspectives Christophe Pélabon 1 , Thomas F. Hansen 2,3 , Matthew L. Carlson 4 and W. Scott Armbruster 1,5,6 1 Department of Biology, Norwegian University of Science and Technology (NTNU), 7491 Trondheim, Norway; 2 Department of Biological Science, Florida State University, Tallahassee, FL 32306, USA; 3 Centre for Ecological and Evolutionary Synthesis, Department of Biology, University of Oslo, Norway; 4 Alaska Natural Heritage Program, Environment and Natural Resources Institute, University of Alaska, Anchorage, AK 99501, USA; 5 Institute of Arctic Biology, University of Alaska, Fairbanks, AK 99775, USA; 6 School of Biological Sciences, University of Portsmouth, Portsmouth, Hants PO1 2DY, UK Summary • We studied patterns of fluctuating asymmetry (FA) in leaves of four populations of the neotropical vine Dalechampia scandens to obtain insight into the origin of leaf FA and the level at which it is controlled. We analysed correlations in signed and unsigned asymmetry at different organizational levels. We also analysed the ontogeny of FA during leaf expansion to test whether asymmetry is regulated during cell expansion, and whether fast-expanding leaves are more or less asymmetrical. • Signed asymmetry was negatively correlated between successive leaves, that is, when the right side of a leaf was larger than the left side, the next leaf on the shoot tended to show the opposite pattern. The magnitude of FA, however, was very weakly correlated among successive leaves or among leaves measured on different shoots. • The direction of asymmetry did not change during leaf expansion, but the relative asymmetry, that is, asymmetry corrected for difference in trait size, decreased during expansion. We found a weak negative relationship between leaf expansion rate and relative asymmetry on the fully expanded leaves. • These results suggest that leaf asymmetry in Dalechampia originates from pertur- bations in cell proliferation in the stem, creating asymmetries in opposite directions in successive leaves. These asymmetries persist during leaf expansion, but tend to be reduced by unknown mechanisms. Key words: fluctuating asymmetry, developmental stability, leaf expansion, Dalechampia scandens , Euphorbiaceae. New Phytologist (2006) 170 : 65–74 © The Authors (2006). Journal compilation © New Phytologist (2006) doi : 10.1111/j.1469-8137.2005.01636.x Author for correspondence: Christophe Pélabon Tel: +47 73596282 Fax: +47 73596100 Email: [email protected] Received: 27 June 2005 Accepted: 7 November 2005 Introduction Developmental stability is the ability of an organism to buffer nonadaptive phenotypic variation resulting from stochastic perturbations during development (developmental noise) (Waddington, 1957; Palmer & Strobeck, 1986; Debat & David, 2001; Nijhout & Davidowitz, 2003). Variation in developmental stability will affect the ability of organisms to reach their target phenotype (the phenotype that would be reached from a given genetic and environmental background without noise of any kind: Nijhout & Davidowitz, 2003). Therefore developmental stability is an important variational property related to fitness (Tracy et al., 2003; see Clarke, 2003 for discussion). However, measuring developmental stability remains difficult because it consists of estimating within- individual variance around an optimal value, which is unknown in most cases. Because both sides of bilateral characters depend on the same genes and share the same environment, the measure of small, directionally random differences between the two sides, referred to as fluctuating asymmetry, has been suggested

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.newphytologist.org
65
Research
Blackwell Publishing Ltd
Patterns of asymmetry in the twining vine
Dalechampia scandens
(Euphorbiaceae): ontogenetic and hierarchical
perspectives
Christophe Pélabon
1
, Thomas F. Hansen
2,3
, Matthew L. Carlson
4
and W. Scott Armbruster
1,5,6
1
Department of Biology, Norwegian University of Science and Technology (NTNU), 7491 Trondheim, Norway;
2
Department of Biological Science, Florida
State University, Tallahassee, FL 32306, USA;
3
Centre for Ecological and Evolutionary Synthesis, Department of Biology, University of Oslo, Norway;
4
Alaska
Natural Heritage Program, Environment and Natural Resources Institute, University of Alaska, Anchorage, AK 99501, USA;
5
Institute of Arctic Biology,
University of Alaska, Fairbanks, AK 99775, USA;
6
School of Biological Sciences, University of Portsmouth, Portsmouth, Hants PO1 2DY, UK
Summary
• We studied patterns of fluctuating asymmetry (FA) in leaves of four populationsof the neotropical vine
Dalechampia scandens
to obtain insight into the origin of leafFA and the level at which it is controlled. We analysed correlations in signed andunsigned asymmetry at different organizational levels. We also analysed the ontogenyof FA during leaf expansion to test whether asymmetry is regulated during cellexpansion, and whether fast-expanding leaves are more or less asymmetrical.• Signed asymmetry was negatively correlated between successive leaves, that is,when the right side of a leaf was larger than the left side, the next leaf on the shoottended to show the opposite pattern. The magnitude of FA, however, was very weaklycorrelated among successive leaves or among leaves measured on different shoots.• The direction of asymmetry did not change during leaf expansion, but the relativeasymmetry, that is, asymmetry corrected for difference in trait size, decreased duringexpansion. We found a weak negative relationship between leaf expansion rate andrelative asymmetry on the fully expanded leaves.• These results suggest that leaf asymmetry in
Dalechampia
originates from pertur-bations in cell proliferation in the stem, creating asymmetries in opposite directionsin successive leaves. These asymmetries persist during leaf expansion, but tend tobe reduced by unknown mechanisms.
Key words:
fluctuating asymmetry, developmental stability, leaf expansion,
Dalechampia scandens
, Euphorbiaceae.
New Phytologist
(2006)
170
: 65–74
© The Authors (2006). Journal compilation ©
New Phytologist
(2006)
doi
: 10.1111/j.1469-8137.2005.01636.x
Author for correspondence:
Christophe Pélabon Tel: +47 73596282 Fax: +47 73596100 Email: [email protected]
Received:
27 June 2005
Accepted:
7 November 2005
Introduction
Developmental stability is the ability of an organism to buffernonadaptive phenotypic variation resulting from stochasticperturbations during development (developmental noise)(Waddington, 1957; Palmer & Strobeck, 1986; Debat &David, 2001; Nijhout & Davidowitz, 2003). Variation indevelopmental stability will affect the ability of organisms toreach their target phenotype (the phenotype that would bereached from a given genetic and environmental background
without noise of any kind: Nijhout & Davidowitz, 2003).Therefore developmental stability is an important variationalproperty related to fitness (Tracy
et al
., 2003; see Clarke, 2003for discussion). However, measuring developmental stabilityremains difficult because it consists of estimating within-individual variance around an optimal value, which is unknownin most cases. Because both sides of bilateral characters dependon the same genes and share the same environment, the measureof small, directionally random differences between the twosides, referred to as fluctuating asymmetry, has been suggested

New Phytologist
(2006)
170
: 65–74
www.newphytologist.org
© The Authors (2006). Journal compilation ©
New Phytologist
(2006)
Research66
to reflect the composite effect of developmental noise anddevelopmental stability (Van valen, 1962; Palmer & Strobeck,1986).
Although fluctuating asymmetry, as a measure of develop-mental stability, has been studied intensively over the past twodecades, both theoretically and empirically (see Polak, 2003for review), the relationship between developmental stabilityand fluctuating asymmetry remains poorly understood. Partof the problem resides in a lack of understanding of the pro-cesses that control the development of both sides of bilateralcharacters, and whether these processes have organism-wideor more localized effects.
Analysis of the correlation pattern between fluctuatingasymmetry measurements on different characters or repeatedmodules, such as leaves or flowers, provides valuable informa-tion on the level at which fluctuating asymmetry, and possiblydevelopmental stability, are regulated. Indeed, correlation inunsigned fluctuating asymmetry (independent of the directionof asymmetry) among traits should reflect the existence of anorganism-wide regulation of developmental stability (Leamy,1993; Polak
et al
., 2003). However, correlation in signedasymmetry among traits might also result from the effects ofdevelopmental noise shared among structurally or develop-mentally related traits (Van Dongen
et al
., 1999; Klingenberg
et al
., 2001; Polak
et al
., 2003).Another series of questions underlying the relationship
between fluctuating asymmetry and developmental stabilityconcerns the origin and possible control of fluctuating asym-metry during ontogeny. Several models have been put forwardto describe the ontogeny of fluctuating asymmetry (Emlen
et al
., 1993; Swaddle & Witter, 1997; Aparicio, 1998; seeKellner & Alford, 2003; Klingenberg, 2003 for reviews).Distinction between these different models depends on whetherasymmetry is regulated during ontogeny, and whether thisregulation involves feedback mechanisms between the twosides (Kellner & Alford, 2003). Data concerning the ontogenyof fluctuating asymmetry, such as the level of changes inmagnitude and direction of asymmetry that occur during traitdevelopment, are therefore crucial for a better understandingof the mechanisms that control/affect developmental stability.
Plants are particularly useful for studying the ontogeny offluctuating asymmetry and correlation patterns in the level offluctuating asymmetry across traits. Their modular structureprovides additional organizational levels at which fluctuatingasymmetry can be measured and compared. Additionally,measurements on leaves and flowers provide the opportunityto test for correlation in fluctuating asymmetry amonghomologous traits within individuals.
Using observations in a twining vine,
Dalechampia scandens
(Euphorbiaceae), we address questions related to the develop-mental origin of fluctuating asymmetry. We first tested whetherthe observed asymmetry resulted from the twining behaviourof the vine, or from the phenotypic expression of develop-mental noise. We further tested whether fluctuating asymmetry
reveals perturbations in developmental processes at the level ofthe leaf, shoot or plant. To answer this question, we analysedthe pattern of correlation in signed and unsigned asymmetryat these different levels of organization. We then analysed theasymmetry pattern during leaf expansion to test whether cellexpansion amplified already existing asymmetries producedduring cell division, or whether regulatory processes mightdecrease the level of asymmetry. Finally, we analysed therelationship between fluctuating asymmetry and the rate ofleaf expansion, to test whether fast-expanding leaves are lessasymmetrical, as predicted if fluctuating asymmetry is nega-tively related to fitness (fitness-indicator model: Teather, 1996;Valkama & Kozlov, 2001); or more asymmetrical, as predictedif a trade-off occurs between growth rate and the ability tocorrect developmental errors (developmental-homeostasismodel, Martel
et al
., 1999; Lempa
et al
., 2000).
Materials and Methods
Study organism and breeding conditions
Dalechampia scandens
L. is an alternate-leaved, neotropicalvine producing unisexual flowers aggregated into bisexualpseudanthial inflorescences or blossoms (Webster & Webster,1972; Webster & Armbruster, 1991). Both blossoms and leavesare bilaterally symmetrical (Fig. 1; Pélabon
et al
., 2004a, 2004b,2005). Individual plants measured in this study came fromseeds collected at two locations in Venezuela and two inMexico (see Hansen
et al
., 2000 for location of populations).Blossoms from the two Mexican populations (Tulum andChetumal) have large, resin-producing glands; those from thetwo Venezuelan populations (Tovar and Caracas) have smallglands (Hansen
et al
., 2003a). Leaves from all populations arevery similar. The only difference observed in the glasshousewas the more frequent occurrence of leaves with five lobes inthe Venezuelan populations (we did not measure the five-lobedleaves in this study).
Infructescences (comprising three pistillate flowers andcontaining up to nine seeds) were collected from separateindividuals in each population in early 1998. Several seedsfrom each infructescence were germinated in March–May1998 in the glasshouse of the Biology Department, NTNU(Trondheim, Norway). Leaf measurements were taken onindividuals from this first glasshouse generation for all popu-lations except Tulum. For the Tulum population, individualsfrom the first glasshouse generation were crossed in a dialleldesign to estimate the genetic variance in floral traits (Hansen
et al
., 2003a, 2003b; Pélabon
et al
., 2004a). For this popula-tion, measurements were therefore taken from the secondglasshouse generation. Measurements on growing leaves weremade on individuals originating from the Chetumal popula-tion in 1999, and from the Tovar population in 2003.
Conditions in the glasshouse were maintained as constantas possible during the whole study with an average temperature

© The Authors (2006). Journal compilation ©
New Phytologist
(2006)
www.newphytologist.org
New Phytologist
(2006)
170
: 65–74
Research 67
of 28
°
C by day and 22
°
C by night, 60–80% humidityand 13 : 11 light : dark light regime. Plants were fertilizedweekly.
Measurements of asymmetry and fluctuating asymmetry
Leaf asymmetry was defined as the difference in lengthbetween the left and right lobes (Fig. 1). We also measuredasymmetry in the width of the central lobe as the differencein distance between the left and right margins and the centralvein (Fig. 1). We defined (signed) asymmetry as the signeddifference
L
−
R
. Fluctuating asymmetry (FA) corresponds tothe absolute value |
L
−
R
|. Relative asymmetry and relative FAwere calculated as: relative asymmetry = 100
×
[ln(
L
)
−
ln(
R
)];and relative FA = 100
×
| ln(
L
)
−
ln(
R
) |, respectively, where lnis the natural log-transformed character value (Clarke, 1998).The relative asymmetry and relative FA can be read as measuresof asymmetry in percentage of the trait size. The length of thecentral lobe was used as a measure of overall leaf size. Allmeasurements were performed with a digital calliper to thenearest 0.01 mm using an optical binocular magnifier (
×
5magnification) by a single observer (C.P.).
Asymmetry pattern on successive leaves
We measured asymmetry on two successive leaves on the sameshoot, one shoot per plant, on 29–309 plants per population,across four populations (Tulum
n
= 309; Chetumal
n
= 31;Caracas
n
= 30; Tovar
n
= 29). Measurements were made onleaves borne by long shoots projecting out from the crownof the plant. The two successive leaves shared similar lightconditions, as did leaves measured on different plants. In thesemeasurements we did not include the first leaf after the shootbranched, which was studied separately, because it usuallyexhibited a different pattern of development. We collected thecomplete shoot and measured two successive leaves flattenedunder transparent sheet within few minutes of collection. Themean length of these leaves was comparable with that of fullyexpanded leaves measured at the same time, in the studyinvestigating leaf expansion.
We also measured the first leaf on the branch (mostproximal leaf ) for 73 individual plants from the Tulumpopulation. These data were used to test the hypothesis that leafasymmetry results from the twining pattern of the shoot. Wepredicted that, if leaf asymmetry results from twining, thenasymmetries measured at a constant position (here the first
Fig. 1 (a) Sketch of a shoot of Dalechampia scandens. (b) Diagram of the traits measured on leaves of D. scandens. The illustration shows a normal, three-lobed leaf. Measurements: LCL, length of central lobe; LRL and LLL, length of right and left lobe, respectively; wl and wr, left and right width of central lobe, respectively. Widths of central lobe left and right of the mid-vein were measured perpendicular to the central vein where the lobe was the widest.

New Phytologist
(2006)
170
: 65–74
www.newphytologist.org
© The Authors (2006). Journal compilation ©
New Phytologist
(2006)
Research68
leaf of the shoot) should display a pattern of directionalasymmetry. We also measured one additional leaf on thesame shoot for comparison of the size and fluctuatingasymmetry level.
We measured two successive leaves on two different shoots(four leaves per plant) in the same 73 individuals from theTulum population, to test the correlation in developmentalstability across shoots from the same plant.
Fluctuating asymmetry and rate of leaf expansion
Ontogeny of leaf asymmetry during leaf expansion wasstudied first on a set of 22 leaves on 11 plants (two leaves perplant on different shoots) from the Chetumal population(Mexico). We measured the length of each of the three lobeson each leaf, as described in Fig. 1, every 5 d from when leaveswere approximately 20 mm long until the final size was reached(approx. 30 d later). Measurements were stopped when noincrease in leaf length was observed between two consecutivemeasurements. Fully grown leaves were collected and measuredimmediately after being flattened under a transparent sheet.The relationship between expansion rate and final asymmetrywas further analysed on individuals from the distinct populationof Tovar (Venezuela). From early September to October 2003,we measured the expansion rate and the final asymmetry onone leaf from each of 31 individuals.
Expansion rate was measured on actively expanding leavesthat were 44% (SE = 2.6%) of their final size in the Chetumalpopulation, and from 17.2% (SE = 0.6%) of their final size inthe Tovar population. The difference in percentage of the finalsize at which initial measurements were made has two differentcauses. First, the initial measurements on the leaves of theTovar population were made on younger (smaller) leavesbecause we did not record the length of both lateral lobes,which would have implied measurements on unfolded leaves.Additionally, Tovar leaves achieved a larger size when fullyexpanded than the leaves from the Chetumal population,presumably because of unknown changes in the glasshouseenvironment between the first series of measurements in 1999and the second series in 2003.
The expansion rate of each leaf was estimated by fitting thedata with an asymptotic exponential model:
Y
=
A
−
Be
−
Ct
where
Y
is the length of the central lobe and
t
the age of theleaf in days after the first measurement; and
A
,
B
and
C
are theparameters of the model. Models were fitted for each leafindependently using nonlinear least-squares analysis (nls in
S
-
PLUS
; Crawley, 2002). We considered
C
as an index of the leafexpansion rate and analysed the relationship between
C
andthe final asymmetry. Because two leaves were measured perindividual plant in the Chetumal data set, we analysed therelationship between the expansion rate and the final fluctuating
asymmetry using a linear mixed model, where individualidentity was entered as a random factor nested into population,also entered as random factor.
Statistics and measurement errors
Measurement error biases the estimation of fluctuatingasymmetry (Palmer, 1994). Measurement error on unexpandedleaves was estimated by repeated measurements on 17 youngleaves of average size (SD) of 23.15 (3.73) mm (range 14–36 mm). Measurement error on fully expanded leaves wasestimated from the last two measurements on each leaf (
n
= 22)when expansion had ceased. Repeatability of FA measurementon both young and adult leaves was high (young leaves:
R
=0.98,
F
16,17
= 80.49; adult leaves:
R
= 0.91,
F
21,22
= 21.39).Additionally, the means and variances of FA were correctedfor measurement error (see Table 1 and Pélabon
et al
., 2004afor further explanation).
The size of the samples used to test the different hypothesesvaries substantially in this study, as does the power of the tests.We therefore focused on the effect size of the patterns,especially when similar effect sizes (
r
2
) were observed in testswith different sample sizes. All statistical analyses were done in
S
-
PLUS
(Venables & Ripley, 2002).
Results
Asymmetry: general pattern
The twining pattern of
D. scandens
was identical in thefour populations observed, the stem showing a similar right-handed rotation (clockwise; see Hashimoto, 2002 fordefinition of right-handed sense of rotation). Descriptivestatistics for leaf size and asymmetries in the four populationsare reported in Table 1. We found no evidence of directionalasymmetry (zero included in the 95% CI of the mean signedasymmetry in all populations; Table 1) or antisymmetry (nonegative kurtosis of the distribution of signed asymmetries inall populations; Table 1). Therefore the asymmetry pattern isconsistent with fluctuating asymmetry (Palmer & Strobeck,1986).
There were no significant differences between the fourpopulations in leaf size or in the level of leaf FA or relative FA(Table 1). Fluctuating asymmetry increased weakly with leafsize (FA in lobe length vs leaf size,
r
2
= 0.03,
P
= 0.005; nointeraction effect with population, Table 1). Signed asymmetryin the width of the central lobe was positively, althoughweakly, correlated with the signed asymmetry in the length oflateral lobes in all populations (average
r
2
= 0.08; Table 1),indicating that leaf fluctuating asymmetry results partlyfrom developmental disturbance between the two halveson each side of the central vein. However, the low
r
2
of thisrelationship, and the larger relative asymmetry in the widthof the central lobe compared with the asymmetry in lobe

© The Authors (2006). Journal compilation ©
New Phytologist
(2006)
www.newphytologist.org
New Phytologist
(2006)
170
: 65–74
Research 69
length (mean relative FA, width central lobe = 14.21%, lobelength = 5.76%), suggest that these different measurementsof leaf asymmetry reflect partly independent sources ofdevelopmental disturbance.
As reported earlier (Webster & Webster, 1972), the mostproximal leaf on a
D. scandens
shoot (first leaf after the shootbranches off ) displays a different pattern of developmentfrom the other leaves. Although we did not notice a systematicchange in the number of lobes, as reported by Webster &Webster (1972), the most proximal leaves measured in theTulum population were, on average, smaller than more distalleaves on the same shoot [mean leaf length (SD): most proximalleaf = 61.33 (15.10) mm; distal leaf = 77.36 (13.65) mm;paired
t
-test
=
−8.55, n = 73, P < 0.001]. Furthermore, theseleaves displayed a greater level of asymmetry than more distalleaves [mean relative FA (SD) in lobe length: proximal leaf= 6.74 (6.04); distal leaf = 4.05 (3.37); paired t-test = 3.74,n = 73, P < 0.001]. However, we did not observe directionalasymmetry on these leaves (mean signed asymmetry = 0.47;95% CI = −0.50; 1.44). This suggests a random origin of theside of the asymmetry.
Correlations in signed and unsigned FA
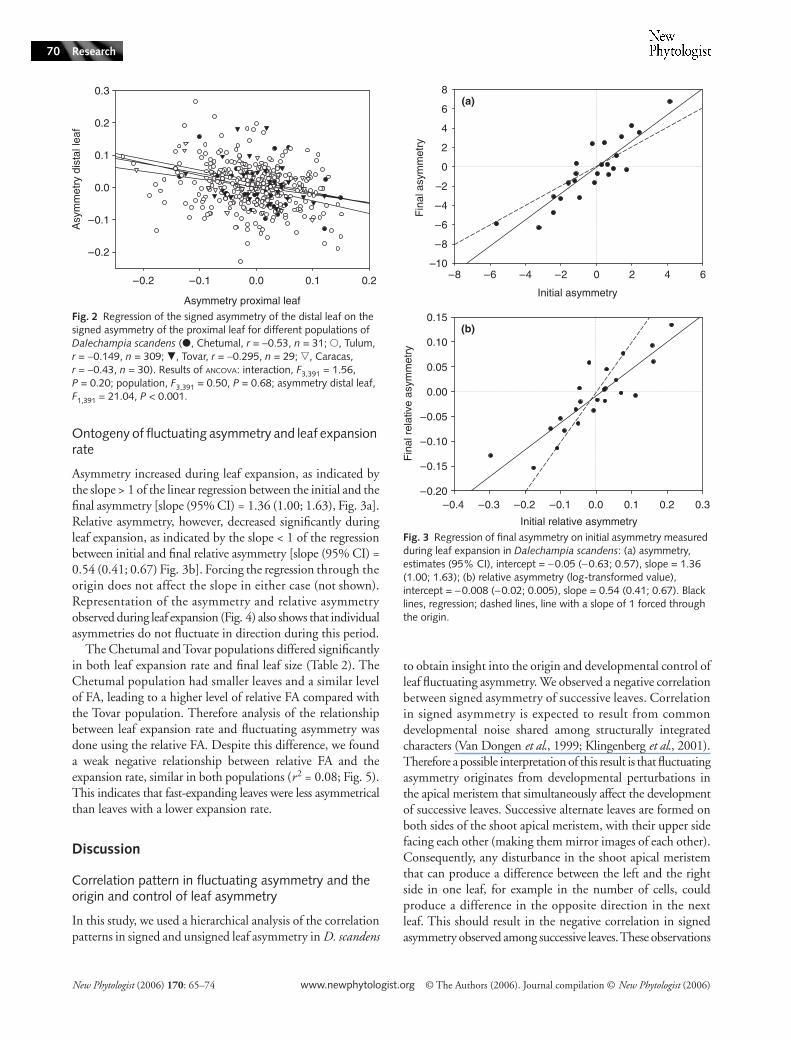
The signed asymmetries of two successive leaves on a shoot werenegatively correlated; this pattern was similar in the four popu-lations. Although correlations between the two successive leavesranged from −0.15 to −0.50, the slopes of the regression analyseswere very similar (Fig. 2). Thus, when the right side of a leafwas larger than the left side, the next leaf on the shoot tendedto show the opposite pattern. Analysis of asymmetry in the widthof the central lobe gave similar results (average r = −0.16).
Fluctuating asymmetry was, at most, weakly correlatedamong successive leaves (r ranging from −0.20 to 0.36 in thefour populations, average r = 0.09), and there was no significantcorrelation in FA among leaves measured on two differentshoots on the same plant (mean r2 = 0.005, n = 73). Weconducted a variance component analysis to estimate theproportion of variance in FA expressed at the different levels:individuals; shoots within individuals; and leaf within shoot.This analysis revealed that the vast majority of the variancein FA (98%) was caused by differences among leaves, theremaining 2% being caused by differences among individuals.
Table 1 Descriptive statistics for trait size and asymmetry in adult leaves in four populations of Dalechampia scandens. Two successive leaves were measured on a single shoot per individual. The size of both leaves is reported, but only the leaf in proximal position is taken into account to estimate descriptive statistics in FA. Data are in mm except for relative FA which is in percentage of trait size. The 95% confidence intervals were estimated using bootstrapping methods. Mean and variance in FA and relative FA are corrected for measurement error as follows:
FAcorr = where FAobs is the observed FA, and is the measurement variance calculated as Var(m1 − m2), where m1 and
m2 are the signed asymmetry calculated from the first and second measurements. Variance in FA corrected for measurement error was obtained by removing (1 − 2/π) from the observed variance in FA (see Pélabon et al. (2004a) for further details)
Chetumal Tulum Tovar Caracass
N 31 309 29 30Leaf size (SD)a
Proximal leaf 57.04 (11.31) 57.60 (12.67) 54.95 (8.24) 60.33 (9.58)Distal leaf 55.35 (8.47) 56.18 (11.29) 57.50 (11.32) 57.32 (10.37)
Signed asymmetryMean 0.37 −0.06 0.57 −0.27(95% CI) (−0.55; 2.26) (−0.25; 0.08) (−0.03; 2.43) (1.53; 0.89)Kurtosis 1.62 2.37 0.29 1.92Skew −0.001 0.55 −0.16 0.56
FAb
Mean 2.35 2.56 1.76 2.80Variance 5.58 5.72 1.98 7.33CV 1.01 0.93 0.80 0.97
Correlation FA, sizer 0.23 0.14 0.15 −0.08(95% CI) (0.08; 0.55) (0.09; 0.25) (−0.05; 0.57) (−0.44; 0.21)
Unsigned relative FAb
Lobe length 4.95 5.44 3.61 5.53Lobe width 15.59 14.08 12.69 14.50
Correlation between relative asymmetries in lobe length and widthr 0.25 0.19 0.32 0.39(95% CI) (−0.22; 0.62) (0.10; 0.23) (−0.06; 0.68) (−0.07; 0.73)
aTest for difference in leaf size among populations: F3,398 = 0.995, P = 0.395.bTest for differences in fluctuating asymmetry (FA) among populations: ANOVA on square-root transformed data: FA, F3,395 = 0.84, P = 0.47; relative-FA, F3,395 = 0.73, P = 0.54.
FAobs2 2 /− 2sm π sm
2
sm2
New Phytologist (2006) 170: 65–74 www.newphytologist.org © The Authors (2006). Journal compilation © New Phytologist (2006)
Research70
Ontogeny of fluctuating asymmetry and leaf expansion rate
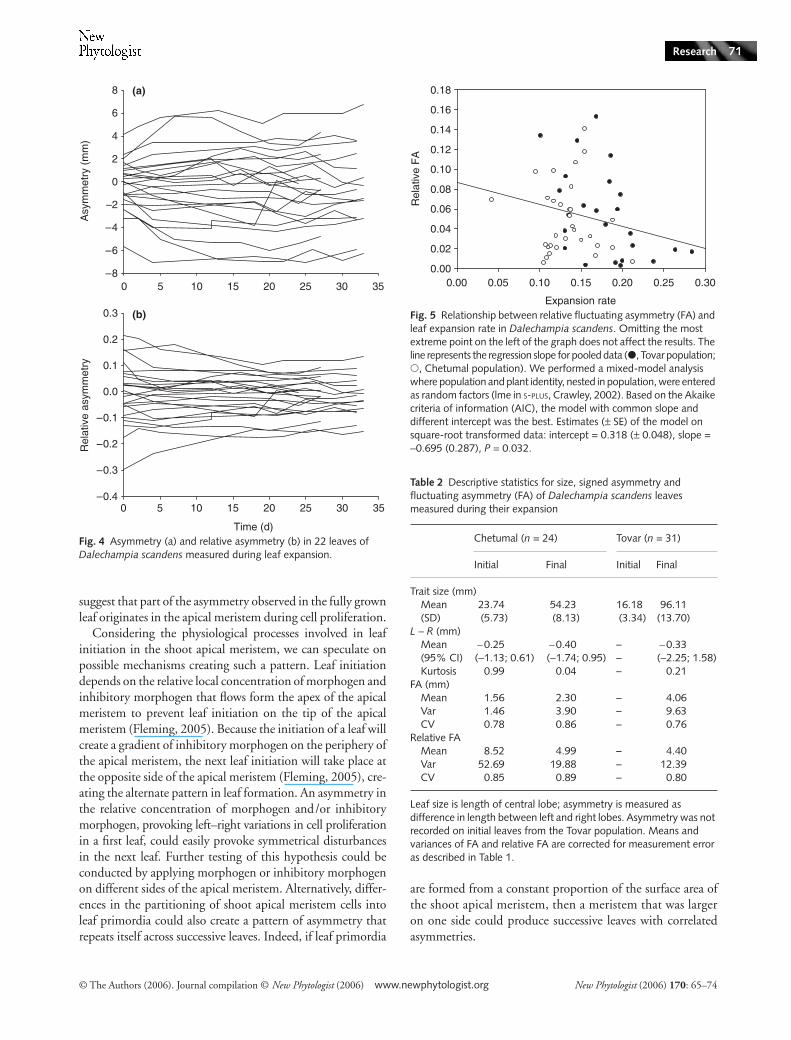
Asymmetry increased during leaf expansion, as indicated bythe slope > 1 of the linear regression between the initial and thefinal asymmetry [slope (95% CI) = 1.36 (1.00; 1.63), Fig. 3a].Relative asymmetry, however, decreased significantly duringleaf expansion, as indicated by the slope < 1 of the regressionbetween initial and final relative asymmetry [slope (95% CI) =0.54 (0.41; 0.67) Fig. 3b]. Forcing the regression through theorigin does not affect the slope in either case (not shown).Representation of the asymmetry and relative asymmetryobserved during leaf expansion (Fig. 4) also shows that individualasymmetries do not fluctuate in direction during this period.
The Chetumal and Tovar populations differed significantlyin both leaf expansion rate and final leaf size (Table 2). TheChetumal population had smaller leaves and a similar levelof FA, leading to a higher level of relative FA compared withthe Tovar population. Therefore analysis of the relationshipbetween leaf expansion rate and fluctuating asymmetry wasdone using the relative FA. Despite this difference, we founda weak negative relationship between relative FA and theexpansion rate, similar in both populations (r2 = 0.08; Fig. 5).This indicates that fast-expanding leaves were less asymmetricalthan leaves with a lower expansion rate.
Discussion
Correlation pattern in fluctuating asymmetry and the origin and control of leaf asymmetry
In this study, we used a hierarchical analysis of the correlationpatterns in signed and unsigned leaf asymmetry in D. scandens
to obtain insight into the origin and developmental control ofleaf fluctuating asymmetry. We observed a negative correlationbetween signed asymmetry of successive leaves. Correlationin signed asymmetry is expected to result from commondevelopmental noise shared among structurally integratedcharacters (Van Dongen et al., 1999; Klingenberg et al., 2001).Therefore a possible interpretation of this result is that fluctuatingasymmetry originates from developmental perturbations inthe apical meristem that simultaneously affect the developmentof successive leaves. Successive alternate leaves are formed onboth sides of the shoot apical meristem, with their upper sidefacing each other (making them mirror images of each other).Consequently, any disturbance in the shoot apical meristemthat can produce a difference between the left and the rightside in one leaf, for example in the number of cells, couldproduce a difference in the opposite direction in the nextleaf. This should result in the negative correlation in signedasymmetry observed among successive leaves. These observations
Fig. 2 Regression of the signed asymmetry of the distal leaf on the signed asymmetry of the proximal leaf for different populations of Dalechampia scandens (�, Chetumal, r = −0.53, n = 31; �, Tulum, r = −0.149, n = 309; �, Tovar, r = −0.295, n = 29; �, Caracas, r = −0.43, n = 30). Results of ANCOVA: interaction, F3,391 = 1.56, P = 0.20; population, F3,391 = 0.50, P = 0.68; asymmetry distal leaf, F1,391 = 21.04, P < 0.001.
Fig. 3 Regression of final asymmetry on initial asymmetry measured during leaf expansion in Dalechampia scandens: (a) asymmetry, estimates (95% CI), intercept = −0.05 (−0.63; 0.57), slope = 1.36 (1.00; 1.63); (b) relative asymmetry (log-transformed value), intercept = −0.008 (−0.02; 0.005), slope = 0.54 (0.41; 0.67). Black lines, regression; dashed lines, line with a slope of 1 forced through the origin.
© The Authors (2006). Journal compilation © New Phytologist (2006) www.newphytologist.org New Phytologist (2006) 170: 65–74
Research 71
suggest that part of the asymmetry observed in the fully grownleaf originates in the apical meristem during cell proliferation.
Considering the physiological processes involved in leafinitiation in the shoot apical meristem, we can speculate onpossible mechanisms creating such a pattern. Leaf initiationdepends on the relative local concentration of morphogen andinhibitory morphogen that flows form the apex of the apicalmeristem to prevent leaf initiation on the tip of the apicalmeristem (Fleming, 2005). Because the initiation of a leaf willcreate a gradient of inhibitory morphogen on the periphery ofthe apical meristem, the next leaf initiation will take place atthe opposite side of the apical meristem (Fleming, 2005), cre-ating the alternate pattern in leaf formation. An asymmetry inthe relative concentration of morphogen and/or inhibitorymorphogen, provoking left–right variations in cell proliferationin a first leaf, could easily provoke symmetrical disturbancesin the next leaf. Further testing of this hypothesis could beconducted by applying morphogen or inhibitory morphogenon different sides of the apical meristem. Alternatively, differ-ences in the partitioning of shoot apical meristem cells intoleaf primordia could also create a pattern of asymmetry thatrepeats itself across successive leaves. Indeed, if leaf primordia
are formed from a constant proportion of the surface area ofthe shoot apical meristem, then a meristem that was largeron one side could produce successive leaves with correlatedasymmetries.
Fig. 4 Asymmetry (a) and relative asymmetry (b) in 22 leaves of Dalechampia scandens measured during leaf expansion.
Fig. 5 Relationship between relative fluctuating asymmetry (FA) and leaf expansion rate in Dalechampia scandens. Omitting the most extreme point on the left of the graph does not affect the results. The line represents the regression slope for pooled data (�, Tovar population; �, Chetumal population). We performed a mixed-model analysis where population and plant identity, nested in population, were entered as random factors (lme in S-PLUS, Crawley, 2002). Based on the Akaike criteria of information (AIC), the model with common slope and different intercept was the best. Estimates (± SE) of the model on square-root transformed data: intercept = 0.318 (± 0.048), slope = −0.695 (0.287), P = 0.032.
Table 2 Descriptive statistics for size, signed asymmetry and fluctuating asymmetry (FA) of Dalechampia scandens leaves measured during their expansion
Chetumal (n = 24) Tovar (n = 31)
Initial Final Initial Final
Trait size (mm)Mean 23.74 54.23 16.18 96.11(SD) (5.73) (8.13) (3.34) (13.70)
L − R (mm)Mean −0.25 −0.40 – −0.33(95% CI) (−1.13; 0.61) (−1.74; 0.95) – (−2.25; 1.58)Kurtosis 0.99 0.04 – 0.21
FA (mm)Mean 1.56 2.30 – 4.06Var 1.46 3.90 – 9.63CV 0.78 0.86 – 0.76
Relative FAMean 8.52 4.99 – 4.40Var 52.69 19.88 – 12.39CV 0.85 0.89 – 0.80
Leaf size is length of central lobe; asymmetry is measured as difference in length between left and right lobes. Asymmetry was not recorded on initial leaves from the Tovar population. Means and variances of FA and relative FA are corrected for measurement error as described in Table 1.

New Phytologist (2006) 170: 65–74 www.newphytologist.org © The Authors (2006). Journal compilation © New Phytologist (2006)
Research72
Surprisingly, we observe only a weak correlation in signedasymmetry between traits measured on the same leaf (asym-metry in central lobe width and asymmetry in the length ofthe lateral lobes), and this is not larger than the correlation insigned asymmetry observed between successive leaves. Thisindicates a relatively weak integration of the leaf, where growthof the lateral lobes is somewhat independent of growth ofthe central lobe. Such a lack of integration may occur if thegrowth in lobe width depends on the activity of lateralprimordia, while the growth in lobe length depends primarilyon initial cell distribution in the leaf primordium.
Despite the correlation in signed asymmetry observed amongsuccessive leaves, we found only a weak correlation in fluctu-ating asymmetry among these leaves, and no correlation at allamong leaves measured on different shoots on the same plant.Correlation in unsigned asymmetry among traits indicatesorganism-wide control of developmental stability (Leamy,1993). Considering that the correlation in fluctuating asym-metry among successive leaves is probably caused by an effectof common developmental noise, as suggested by the negativecorrelation in signed asymmetries, our results suggest that nooverall control of developmental stability, or at most a weak,undetectable control, occurs in D. scandens at either the indi-vidual or the shoot level. This is confirmed by the variancecomponent analysis, which indicates that most variability inFA is confined at the leaf level.
Ontogeny of asymmetry
Asymmetry measurement on small leaves is difficult andimprecise without damaging the leaf. Therefore the firstasymmetry measurements on leaves were made when theywere an average of 40% of final size. Although lateral primordiamay contribute to leaf growth via cell division during laterstages of leaf ontogeny, the increase in leaf size observed in thisstudy essentially corresponds to cell expansion (Maksymowych,1973).
We observed an increase in fluctuating asymmetry from theinitial measurement on young leaves to the stage of full leafexpansion, without a change in the direction of asymmetry.This pattern is consistent with the pattern observed by Møller& Van Dongen (2003) on expanding leaves of Ulmus glabra,and fits the persistent asymmetry model, where asymmetriesdetermined during early ontogeny persist during growth(Kellner & Alford, 2003). The origin of asymmetry duringcell division in the apical meristem, suggested by the negativecorrelation in signed asymmetry between successive leaves,also fits this hypothesis. However, a simple magnification ofearly determined asymmetries by cell expansion later duringleaf ontogeny should result in the maintenance of the relative(size-corrected) asymmetry during leaf expansion. We observedthat relative asymmetry decreased during leaf expansion,suggesting a regulation of symmetry during this part of theleaf ontogeny.
Fluctuating asymmetry in the leaves therefore appears toresult primarily from developmental disturbances during celldivision, and while regulatory mechanisms can partly reducethe level of relative fluctuating asymmetry, the absoluteasymmetry is magnified by later cell expansion. Because of thedifficulties in analysing fluctuating asymmetry in leaf prim-ordium, occurrence of similar regulatory mechanisms duringthe stage of cell division remains unknown.
Fluctuating asymmetry and leaf expansion rate
Both positive and negative relationships between growthrate and fluctuating asymmetry have been suggested. Thedevelopmental-homeostasis hypothesis suggests that rapidgrowth beyond the selected optimum may negatively affectdevelopmental stability because of a trade-off between growthrate and the ability to correct developmental errors (Calow,1982; Arendt, 1997). In this case rapid growth, such ascompensatory growth, should be positively correlated withfluctuating asymmetry (Martel et al., 1999). Note that thesimultaneous effects of some stressful events, such as defoliationor plant damage, on growth and developmental stability couldproduce the same positive correlation between growth rateand fluctuating asymmetry without any direct relationshipbetween growth rate and developmental stability (Kozlov,2003). Alternatively, rapid growth may be the consequenceof a good individual condition (genetic or phenotypic), andif there is a positive relationship between condition anddevelopmental stability, a negative relationship is expectedbetween growth rate and developmental instability (Valkama& Kozlov, 2001). These two hypotheses are not mutuallyexclusive. Indeed, the negative relationship between growthand developmental stability is expected only when the growthrate is beyond the selected optimum. Below the optimal level,a positive relationship can occur. If we take rate of leaf expansionas a proxy for leaf growth, the observed negative correlationbetween leaf expansion rate and fluctuating asymmetry inDalechampia supports the second hypothesis. Interestingly,the effect size observed in this relationship is similar to theeffect size observed by Valkama & Kozlov (2001) on leavesfrom mountain birch (Betula pubescens). Note, however, thatWilsey & Saloniemi (1999) did not find any relationshipbetween leaf fluctuating asymmetry and shoot growth inmountain birches.
Leaf development is highly dependent on environmentalfactors such as light and water availability (Van Volkenburgh,1999). Furthermore, Montalvo (1994) provided evidencethat leaf expansion rate could reflect the fitness of the plant.Therefore, in principle, a negative relationship betweenfluctuating asymmetry and leaf expansion rate suggests thatfluctuating asymmetry reflects individual fitness. But the smalleffect sizes observed in this study show that a relationshipbetween fluctuating asymmetry and fitness must be very weak.Furthermore, in a study of the genetic basis of developmental

© The Authors (2006). Journal compilation © New Phytologist (2006) www.newphytologist.org New Phytologist (2006) 170: 65–74
Research 73
stability in blossoms and leaves of D. scandens, we found noevidence of additive genetic variance for developmental stability,and we found also no consistent effect of inbreeding oroutbreeding on the level of developmental stability (Pélabonet al., 2004a, 2004b). Therefore the positive correlationbetween leaf expansion rate and developmental stability is notexpected to reflect individual genetic quality, but appearsmore likely to result from covariation in the effects of the envi-ronment on both growth rate and developmental stability. Inconclusion, a component of fluctuating asymmetry appears tooriginate during early ontogeny via disturbance of cell prolif-eration in the leaf primordium. Although partially correctedby some regulating mechanisms, these asymmetries are notcompletely eliminated in expanding leaves. Whether regula-tion is achieved by compensatory growth or targeted growthremains unresolved, and requires additional study. The absenceof a correlation in FA among leaves from the same plant sug-gests that there is no detectable variation in overall control ofdevelopmental stability at the plant level. Furthermore, theweak correlation in fluctuating asymmetry observed in succes-sive leaves seems to result essentially from the sharing ofcommon developmental noise, not from a common develop-mental stability at the shoot level. Finally, although a negativecorrelation was found between fluctuating asymmetry andexpansion rate, the weakness of this correlation, and theapparent absence of genetic variation in fluctuating asymme-try observed in this species (Pélabon et al., 2004a), suggestthat such correlation is unimportant for fitness and is purelyphenotypic.
Acknowledgements
The authors thank L. Antonsen, L. Dalen and T. Berge forseed collection in the field, and G. Fyhn-Hanssen for glasshouseassistance. We also thank M. Kozlov and two anonymousreferees who provided very valuable comments on an earlierdraft of this manuscript.
References
Aparicio JM. 1998. Patterns of fluctuating asymmetry in developing primary feathers: a test of the compensational growth hypothesis. Proceedings of the Royal Society of London, B 265: 2353–2357.
Arendt JD. 1997. Adaptive intrinsic growth rates: an integration across taxa. Quarterly Review of Biology 72: 149–177.
Calow P. 1982. Homeostasis and fitness. American Naturalist 120: 416–419.
Clarke GM. 1998. The genetic basis of developmental stability. IV. Individual and population asymmetry parameters. Heredity 80: 553–561.
Clarke GM. 2003. Developmental stability – fitness relationships in animals: some theoretical considerations. In: Polak M, ed. Developmental Instability: Causes and Consequences. Oxford, UK: Oxford University Press, 187–195.
Crawley MJ. 2002. Statistical Computing: An Introduction to Data Analysis Using S-PLUS. New York, USA: John Wiley and Sons.
Debat V, David P. 2001. Mapping phenotypes: canalization, plasticity and developmental stability. Trends in Ecology and Evolution 16: 555–561.
Emlen JM, Freeman DC, Graham JH. 1993. Nonlinear growth dynamics and the origin of fluctuating asymmetry. Genetica 89: 77–96.
Fleming AJ. 2005. The control of leaf development. New Phytologist 166: 9–20.
Hansen TF, Armbruster WS, Antonsen L. 2000. Comparative analysis of character displacement and spatial adaptations as illustrated by the evolution of Dalechampia blossoms. American Naturalist 156: S17–S34.
Hansen TF, Pélabon C, Armbruster WS, Carlson ML. 2003a. Evolvability and genetic constraint in Dalechampia blossoms: components of variance and measures of evolvability. Journal of Evolutionary Biology 16: 754–766.
Hansen TF, Armbruster WS, Carlson ML, Pélabon C. 2003b. Evolvability and constraint in Dalechampia blossoms: genetic correlation and conditional evolvability. Journal of Experimental Zoology 296B: 23–39.
Hashimoto T. 2002. Molecular genetic analysis of left–right handedness in plants. Philosophical Transactions of the Royal Society of London, B 357: 799–808.
Kellner JR, Alford RA. 2003. The ontogeny of fluctuating asymmetry. American Naturalist 161: 931–947.
Klingenberg CP. 2003. A developmental perspective on developmental instability: therory, models, and mechanisms. In: Polak M, ed. Developmental Instability: Causes and Consequences. Oxford, UK: Oxford University Press, 14–34.
Klingenberg CP, Badyaev AV, Sowry SM, Beckwith NJ. 2001. Inferring developmental modularity from morphological integration: analysis of individual variation and asymmetry in bumblebee wings. American Naturalist 157: 11–23.
Kozlov MV. 2003. Are fast growing birch leaves more asymmetrical? Oikos 101: 654–658.
Leamy L. 1993. Morphological integration of fluctuating asymmetry in the mouse mandible. Genetica 89: 139–153.
Lempa K, Martel J, Koricheva J, Haukioja E, Ossipov V, Ossipova S, Pihlaja K. 2000. Covariation of fluctuating asymmetry, herbivory and chemistry during birch leaf expansion. Oecologia 122: 354–360.
Maksymowych R. 1973. Analysis of Leaf Development. Cambridge, UK: Cambridge University Press.
Martel J, Lempa K, Haukioja E. 1999. Effects of stress and rapid growth on fluctuating asymmetry and insect damage in birch leaves. Oikos 86: 208–216.
Møller AP, Van Dongen S. 2003. Ontogeny of asymmetry and compensational growth in elm Ulmus glabra leaves under different environmental conditions. International Journal of Plant Sciences 164: 519–526.
Montalvo AM. 1994. Inbreeding depression and maternal effects in Aquilegia caerulea, a partially selfing plant. Ecology 75: 2395–2409.
Nijhout HF, Davidowitz G. 2003. Developmental perspective on phenotypic variation, canalization, and fluctuating asymmetry. In: Polak M, ed. Developmental Instability: Causes and Consequences. Oxford, UK: Oxford University Press, 3–13.
Palmer AR. 1994. Fluctuating asymmetry analyses: a primer. In: Markow A, ed. Development Instability: Its Origins and Evolutionary Implications. Dordrecht, the Netherlands: Kluwer Academic, 335–364.
Palmer AR, Strobeck C. 1986. Fluctuating asymmetry: measurement, analysis, patterns. Annual Review of Ecology and Systematic 17: 391–421.
Pélabon C, Carlson ML, Hansen TF, Armbruster WS. 2005. Effects of crossing distance on offspring fitness and developmental stability in Dalechampia scandens (Euphorbiaceae). American Journal of Botany 92: 842–851.
Pélabon C, Hansen TF, Carlson ML, Armbruster WS. 2004a. Variational and genetic properties of developmental stability in Dalechampia scandens. Evolution 58: 504–514.
Pélabon C, Carlson ML, Hansen TF, Yoccoz NG, Armbruster WS. 2004b. Consequences of inter-population crosses on developmental stability and canalization of floral traits in Dalechampia scandens (Euphorbiaceae). Journal of Evolutionary Biology 17: 19–32.

New Phytologist (2006) 170: 65–74 www.newphytologist.org © The Authors (2006). Journal compilation © New Phytologist (2006)
Research74
Polak M, ed. 2003. Developmental Instability: Causes and Consequences. Oxford, UK: Oxford University Press.
Polak M, Møller AP, Gangestad SW, Kroeger DE, Manning JT, Thornhill R. 2003. Does an individual asymmetry parameter exist? A meta-analysis. In: Polak M, ed. Developmental Instability: Causes and Consequences. Oxford, UK: Oxford University Press, 81–96.
Swaddle JP, Witter MS. 1997. On the ontogeny of developmental stability in a stabilized trait. Proceedings of the Royal Society of London, B 264: 329–334.
Teather K. 1996. Patterns of growth and asymmetry in nestling Tree Swallows. Journal of Avian Biology 27: 302–310.
Tracy M, Freeman DC, Duda JJ, Miglia KJ, Graham JH, Hough RA. 2003. Developmental instability: an appropriate indicator of plant fitness components?. In: Polak M, ed. Developmental Instability: Causes and Consequences. Oxford, UK: Oxford University Press, 196–212.
Valkama J, Kozlov MV. 2001. Impact of climatic factors on the developmental stability of mountain birch growing in a contaminated area. Journal of Applied Ecology 38: 665–673.
Van Dongen S, Sprengers E, Lofstedt C. 1999. Correlated development,
organism-wide asymmetry and patterns of asymmetry in two moth species. Genetica 105: 81–91.
Van Valen L. 1962. A study of fluctuating asymmetry. Evolution 16: 125–142.
Van Volkenburgh E. 1999. Leaf expansion – an integrating plant behaviour. Plant, Cell & Environment 22: 1463–1473.
Venables WN, Ripley BD. 2002. Modern Applied Statistics with S-PLUS. Berlin, Germany: Springer-Verlag.
Waddington CH. 1957. The Strategy of the Genes. New York, USA: Macmillan.
Webster GL, Armbruster WS. 1991. A synopsis of the neotropical species of Dalechampia (Euphorbiaceae). Biological Journal of the Linnean Society 105: 137–177.
Webster GL, Webster BD. 1972. The morphology and relationships of Dalechampia scandens (Euphorbiaceae). American Journal of Botany 59: 573–586.
Wilsey BJ, Saloniemi I. 1999. Leaf fluctuating asymmetry in tree-line mountain birches, Betula pubescens ssp. tortuosa: genetic or environmentally influenced? Oikos 87: 341–345.
About New Phytologist
• New Phytologist is owned by a non-profit-making charitable trust dedicated to the promotion of plant science, facilitating projectsfrom symposia to open access for our Tansley reviews. Complete information is available at www.newphytologist.org.
• Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged.We are committed to rapid processing, from online submission through to publication ‘as-ready’ via OnlineEarly – the 2004 averagesubmission to decision time was just 30 days. Online-only colour is free, and we provide 25 offprints as well as a PDF for eacharticle.
• For online summaries and ToC alerts, go to the website and click on ‘Journal online’. You can take out a personal subscription tothe journal for a fraction of the institutional price. Rates start at £125 in Europe/$232 in the USA & Canada for the online edition(click on ‘Subscribe’ at the website).
• If you have any questions, do get in touch with Central Office ([email protected]; tel +44 1524 594691) or, for a localcontact in North America, the US Office ([email protected]; tel +1 865 576 5261).
Related Documents











