Botanical Journal of the Linnean Society (1998), 127: 207–223. With 5 figures Article ID: bt970133 Patterns and implications of floral nectar secretion, chemical composition, removal effects and standing crop in Mandevilla pentlandiana (Apocynaceae) CAROLINA TORRES AND LEONARDO GALETTO* Instituto Multidisciplinario de Biologı ´a Vegetal, Casilla de Correo 495, 5000 Co ´rdoba, Argentina Received February 1997; accepted for publication September 1997 Flower morphology, nectar features (chemical composition, production pattern, removal effects, and standing crop) and floral visitors are analysed in an Argentine population of Mandevilla pentlandiana. Nectar variability was examined during the lifetime of a single flower, over the course of the flowering season, and at different times of the day. Nectar is sucrose dominant. There were some variations in the proportions of sugar throughout both the flower lifespan and the flowering season. Flowers produced most nectar during bud-stage. Nectar secretion ceased near the end of the first day after flower opening. Nectar quantity varied as a function of flower age due to a combination of nectar secretion, cessation, and resorption periods. Overall sugar production was increased by nectar removal. Standing crop data showed that each open flower and inflorescence offers c. 2 and 11 mg of sugar respectively at any time of the flowering season. There was higher nectar availability at the beginning of the flowering season compared with the rest of the period. Flowers were visited by bumblebees, honeybees and hummingbirds. The greater the number of open flowers and the nectar variance, the more the mean reward quantity per flower available in the inflorescence. The sources of nectar variability in M. pentlandiana seem to be linked with both the female function (nectar resorption, nectar cessation) and the male one (early and comparatively large nectar availability, variation in nectar production as the flower ages, nectar secretion stimulation by nectar removals). 1998 The Linnean Society of London ADDITIONAL KEY WORDS:—flower age – flower structure – hummingbirds – Hymenoptera – visitors. CONTENTS Introduction . . . . . . . . . . . . . . . . . . . . . . . 208 Material and methods . . . . . . . . . . . . . . . . . . . 209 Plants . . . . . . . . . . . . . . . . . . . . . . . 209 Nectar . . . . . . . . . . . . . . . . . . . . . . . 209 Floral visitors . . . . . . . . . . . . . . . . . . . . . 210 Data analysis . . . . . . . . . . . . . . . . . . . . . 211 * Correspondence to Dr L. Galetto. E-mail: [email protected] 207 0024–4074/98/070207+17 $30.00/0 1998 The Linnean Society of London

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Botanical Journal of the Linnean Society (1998), 127: 207–223. With 5 figures
Article ID: bt970133
Patterns and implications of floral nectarsecretion, chemical composition, removal effectsand standing crop in Mandevilla pentlandiana(Apocynaceae)
CAROLINA TORRES AND LEONARDO GALETTO∗
Instituto Multidisciplinario de Biologıa Vegetal, Casilla de Correo 495, 5000 Cordoba,Argentina
Received February 1997; accepted for publication September 1997
Flower morphology, nectar features (chemical composition, production pattern, removaleffects, and standing crop) and floral visitors are analysed in an Argentine population ofMandevilla pentlandiana. Nectar variability was examined during the lifetime of a single flower,over the course of the flowering season, and at different times of the day. Nectar is sucrosedominant. There were some variations in the proportions of sugar throughout both theflower lifespan and the flowering season. Flowers produced most nectar during bud-stage.Nectar secretion ceased near the end of the first day after flower opening. Nectar quantityvaried as a function of flower age due to a combination of nectar secretion, cessation, andresorption periods. Overall sugar production was increased by nectar removal. Standingcrop data showed that each open flower and inflorescence offers c. 2 and 11 mg of sugarrespectively at any time of the flowering season. There was higher nectar availability at thebeginning of the flowering season compared with the rest of the period. Flowers were visitedby bumblebees, honeybees and hummingbirds. The greater the number of open flowers andthe nectar variance, the more the mean reward quantity per flower available in theinflorescence. The sources of nectar variability in M. pentlandiana seem to be linked with boththe female function (nectar resorption, nectar cessation) and the male one (early andcomparatively large nectar availability, variation in nectar production as the flower ages,nectar secretion stimulation by nectar removals).
1998 The Linnean Society of London
ADDITIONAL KEY WORDS:—flower age – flower structure – hummingbirds –Hymenoptera – visitors.
CONTENTS
Introduction . . . . . . . . . . . . . . . . . . . . . . . 208Material and methods . . . . . . . . . . . . . . . . . . . 209
Plants . . . . . . . . . . . . . . . . . . . . . . . 209Nectar . . . . . . . . . . . . . . . . . . . . . . . 209Floral visitors . . . . . . . . . . . . . . . . . . . . . 210Data analysis . . . . . . . . . . . . . . . . . . . . . 211
∗Correspondence to Dr L. Galetto. E-mail: [email protected]
2070024–4074/98/070207+17 $30.00/0 1998 The Linnean Society of London

C. TORRES AND L. GALETTO208
Results . . . . . . . . . . . . . . . . . . . . . . . . 211Plants . . . . . . . . . . . . . . . . . . . . . . . 211Nectar chemical composition . . . . . . . . . . . . . . . . 212Nectar production . . . . . . . . . . . . . . . . . . . 215Removal effects . . . . . . . . . . . . . . . . . . . . 215Standing crop . . . . . . . . . . . . . . . . . . . . . 216Floral visitors . . . . . . . . . . . . . . . . . . . . . 218
Discussion . . . . . . . . . . . . . . . . . . . . . . . 219Acknowledgements . . . . . . . . . . . . . . . . . . . . 222References . . . . . . . . . . . . . . . . . . . . . . . 222
INTRODUCTION
Of the diverse floral rewards offered by plants, nectar is the most important. Itis composed of sugars which constitute the major energy source for pollinators, andmay contain a variety of other solute compounds, e.g. phenols, amino acids, reducingacids, lipids, proteins, alkaloids (Percival, 1961; Baker & Baker, 1983; Simpson &Neff, 1983; Gottsberger, Schrawen & Linskens, 1984; Bernardello, Galetto & Juliani,1991; Stiles & Freeman, 1993). As a consequence of evolution the rewards arehighly attractive while simultaneously restricting forager access (Heinrich, 1983).Nectar variability is determined by many traits related to each flower. These includenectar concentration, volume, sugar composition, production rate, removal effects,active resorption, flower age, etc., as well as other extrinsic traits such as pollinatorbehaviour, air temperature and humidity, and the presence of nectar robbers (e.g.Corbet, 1978; Baker & Baker, 1983; Zimmerman & Pyke, 1986; Feinsinger, 1987;Murawski, 1987; Waser & Mitchell, 1990; Burquez & Corbet, 1991; Mitchell &Waser, 1992; Rathcke, 1992; Bernardello, Galetto & Rodrıquez, 1994; Galetto,Bernardello & Juliani, 1994; Rivera, Galetto & Bernardello, 1996).
Variation in floral reward can affect many aspects of pollinator behaviour; inturn, the animals determine pollen movement and consequently a plant’s fitnessgain measured in terms of pollen transfer to and from other plants. Plant net fitnessmust be influenced by nectar production assuming it entails some cost to the plant(Zimmerman & Pyke, 1986; Pyke, 1991). Nectar production can be determined bythe resource allocation strategies of plants and can be governed by physiologicalprocesses as well as by nectary morphology (Rathcke, 1992). Nectar secretion andpollinator visitation rates determine the patterns of nectar availability (i.e. standingcrop). Zimmerman & Pyke (1986) summarized the work done on floral nectarproduction and standing crop having found only a few studies which examined alarge number of nectar parameters for a single species.
The present study is the first part of a larger project in which we plan to investigatethe nature of the relationship between nectar production, breeding system, seed setand seedling survival. The study is intended to quantify the net fitness of Mandevillapentlandiana (A.DC.) Woodson; at the same time, the features of nectar productionpresented here will contribute to a better understanding of the reproductive biologyof this species.
We address the following questions. How does nectar chemical composition varyboth during the lifetime of an individual flower and over the course of the floweringseason? Which are the visitors to M. pentlandiana? What is the pattern of nectarsecretion and what are the effects of nectar removal? How does the standing crop

NECTAR OF MANDEVILLA PENTLANDIANA 209
pattern change through the flowering season? What are the implications of nectarvariability on the plant’s reproductive biology?
The basic floral structure of the Apocynaceae is relatively fixed (Fallen, 1986).The pollination mechanism in the family has been described in general by Schick(1980, 1982) and Fallen (1986), and in particular for M. pentlandiana by Galetto(1997). This species has a voluminous floral nectary surrounding the ovary, whichis supplied by phloem and xylem bundles. Around 40 stomata can be detected onthe nectary surface of each flower (Galetto, 1997).
Mandevilla Lindl. comprises 110 species of lianas or herbs that grow in warmregions of Central and South America (Woodson, 1933). Mandevilla pentlandiana, aliana, is one of eight species recorded in Argentina. Populations are found in themountain habitats of central and northern Argentina, Bolivia, and the south ofBrazil (Ezcurra, 1981).
MATERIAL AND METHODS
Plants
Around 60 plants of a native population from Argentina (Cordoba province,Colon department, Cabana) were studied during 1994–5. A voucher specimen isdeposited in CORD (Galetto & Torres 320).
Flower age was determined by tagging buds and making detailed observationsuntil corolla abscission. Flower odour emission was recorded by smelling theinflorescences in the field at different times of day. The osmophores were locatedby staining flowers with neutral red (Kearns & Inouye, 1993).
The material to be sectioned was fixed in FAA. Dehydration was carried outthrough an ethyl alcohol-xylol series and the flowers were embedded in paraffin.Longitudinal sections were cut at 10 lm intervals, mounted serially, and stainedwith haematoxylin-safranin-fast green (Conn, Darrow & Emmel, 1960). Drawingswere made using a camera lucida.
Nectar
Nectar was extracted with capillary glass tubes without removing the flowers fromthe plant. Extreme care was taken to avoid damage to the nectaries and to theother flower structures by contaminating them with either latex or pollen. Twovariables were immediately measured: volume (ll) using graduated micropipettes,and sugar concentration (% of sucrose, wt/wt) with an Atago pocket refractometer.The amount of sugar produced was expressed in mg after Kearns & Inouye (1993).
Chemical compositionNectar samples were collected in order to compare them at eight different stages
in the lifetime of a flower and at different periods of the flowering season. Tests foramino acids, lipids, phenols, alkaloids, and reducing acids were performed afterBaker & Baker (1975). A histidine scale suggested by Baker & Baker (1975)was used to quantify amino acids. Sugar separation was accomplished by gas

C. TORRES AND L. GALETTO210
chromatography. Nectar was lyophilized and silylated according to Sweeley et al.(1963). The derivatives were then injected into a Konik KNK 3000-HRGS gaschromatograph equipped with a Spectra-Physics SP 4290 data integrator, a flameionization detector, and a OV 101 3% column (2 m long) on Chromosorb G/AW-DMCS mesh 100–120. Nitrogen was the carrier gas (30 ml min−1) and the followingtemperature programme was used: 208°C for 1 min, 1°C min−1 until 215°C, 10°Cmin−1 until 280°C for 2 min. Carbohydrate standards (Sigma Chem.) were preparedusing the same method. Sucrose ratio (r) and hexose ratio (hr) were calculated asfollows: sucrose (frustose+glucose)−1 and glucose frustose−1, respectively.
ProductionRandomly-chosen inflorescences in bud stage were bagged using paper bags to
prevent pollinator visits. Individual buds were tagged for identification. The presenceof nectar was checked and quantified. Nectar production was determined by usingten flower sets of 15 flowers each. The study period covers the 4 days of the flowerlifetime. Data were taken once for each set allowing the nectar to accumulate untilthe measurement could be taken, using an untouched new set for each one. Netnectar production rate (NNPR) per hour was calculated as follows: mg of sugarproduced between measurements/hours between them (mg h−1), while net nectarresorption rate (NNRR) per hour as: mg of sugar resorbed between measurements/hours between them (mg h−1).
Removal effectsNectar was removed and measured from the same flower repeatedly during the
first 36 hours after opening. The determined interval time comprises the entireactive nectar secretion period. Sets were subjected to 0–5 removals according to thefollowing scheme: set 5=all removals (5) starting from 20:00 h of the first day; set4=four removals starting from 8:30 h of the second day; set 3=three removalsstarting from 12:00 h of the second day; set 2=two removals starting from 15:30 hof the second day; set 1=one removal starting from 19:00 h of the second day; set0 (control)=nectar was allowed to accumulate until 8:30 h of the third day andmeasured at the end of the experiment.
Standing cropThis was evaluated measuring the volume and concentration of the available
nectar (volume, concentration, and mg of sugar in the way described above) fromflowers that had been exposed to pollinators for their entire life. Data were collectedat random for individual flowers and for entire inflorescences between Decemberand February (1994–5) including all flower ages. These measurements were takenin the morning and afternoon throughout the flowering period.
Floral visitors
Data on visitors were obtained from the population by observations throughoutthe flowering season. Qualitative observations were made primarily during thefollowing time periods: 08:00–12:00, 15:00–19:00 and 20:00–24:00 h, a total of

NECTAR OF MANDEVILLA PENTLANDIANA 211
154 h of observation distributed over 34 days from December to the end of February.Floral visitors were captured and/or photographed for identification.
Data analysis
Statistical analyses were performed using methods described in Sokal & Rohlf(1995). All distributions were tested for randomness of nominal data (Runs test),homogeneity of variances (Levene test), and departures from normality (Kolmogorov–Smirnov for goodness of fit test). The effects of nectar removal in the total mg ofsugar produced by each set of flowers were compared with one-way analysis ofvariance (ANOVA) and with the Tukey–Kramer’s post hoc test for multiple com-parisons among pairs of means. Bud and open-flowers standing crops were comparedby a Mann–Whitney test. Flower nectar standing crops through the flowering seasonand at different times of the day were compared by Friedman two-way ANOVA,and the inflorescence standing crops throughout the flowering season by Kruskal–Wallis one-way ANOVA. The Wilcoxon rank test was used for the post hoc analyses.The variable number of open flowers per inflorescence was square root transformedbefore regression analysis to meet normality. Untransformed data are reported forthe sake of clarity.
RESULTS
Plants
Mandevilla pentlandiana is abundant in the Cordoba sierras, central Argentina.When this species is found under forest trees it ascends through them reaching thecanopy (e.g. Celtis tala Planchon (Ulmaceae), Prosopis alba Griseb., Acacia spp., Geoffroeadecorticans (Hook. et Arn.) Burk. (Fabaceae) ), but when it grows in open habitats theplant has branches of c. 1–2 m and is erect or rests on other plant species.
The inflorescences are simple and lateral, constituting dense racemes 6–25 cmlong (Fig. 1A). The flowers are greenish-white, tubular, perfect, and measure c.1.6 cm (Fig. 2A). The tubular corolla has epipetalous stamens which are composedof short filaments and connivent anthers. They form a cone around the style-headand dehisce introrsely by longitudinal slits before anthesis. The receptive stigmaticzone is located at the base of the style-head. The nectary can be found surroundingthe bicarpellary ovary and is fused to the receptacle (Fig. 3). The osmophores arelocated on both the corrola and the calyx. Floral scents, as can be perceived by thehuman sense of smell, emanate from the flowers only during daylight.
The corolla lobes remain folded over at the bud stage and spread out duringanthesis or as a result of pollinator activity at an earlier stage than naturally (Fig.1C). Thus, the time of flower opening can be difficult to determine. The bloomingtime spans December to February. One to three flowers per inflorescence openeach day, with each flower lasting 3–4 days. Consequently, 2–11 flowers in anthesisper inflorescence (x=4.94±2.02, n=65) can be found during most of this 3-monthperiod (Fig. 1A). The development of the fruits starts in the last days of Decemberand the last flowers are found in the middle of February (Fig. 1B).
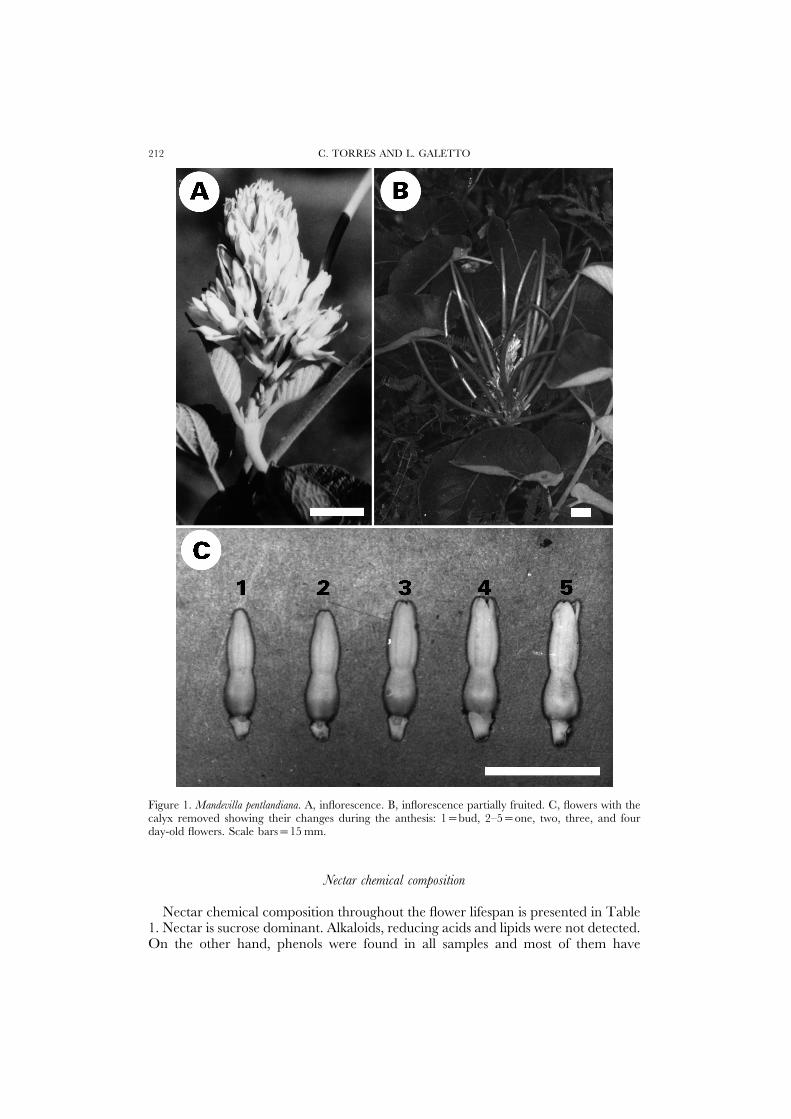
C. TORRES AND L. GALETTO212
Figure 1. Mandevilla pentlandiana. A, inflorescence. B, inflorescence partially fruited. C, flowers with thecalyx removed showing their changes during the anthesis: 1=bud, 2–5=one, two, three, and fourday-old flowers. Scale bars=15 mm.
Nectar chemical composition
Nectar chemical composition throughout the flower lifespan is presented in Table1. Nectar is sucrose dominant. Alkaloids, reducing acids and lipids were not detected.On the other hand, phenols were found in all samples and most of them have

NECTAR OF MANDEVILLA PENTLANDIANA 213
Figure 2. Mandevilla pentlandiana. A, four day-old flower. B–D, flower visitors. B, Apis mellifera. C, Bombusmorio. D, B. opifex. Scale bars=5 mm.

C. TORRES AND L. GALETTO214
a
sh
rs
f
c
ko n
Figure 3. Longitudinal section of flower of Mandevilla pentlandiana. Abbreviations: a=connivent anthers,c=corolla, f=stamen filament, k=calyx, n=nectary, o=ovary, rs=receptive stigmatic zone, sh=style-head. Scale bar=2.5 mm. Lower shaded area=nectar-secreting tissue. Upper shaded area=androecium.
T 1. Nectar chemical composition of Mandevilla pentlandiana throughout flower anthesis. Ab-breviations: hao=number of hours after flower opening that flowers were sampled, for more detailssee Material and Methods; n=number of samples; aa (HS)=amino acids (histidine scale value); ph=phenols; r=sugar ratio; hr=hexose ratio; +=positive reaction; −=negative reaction (not detected)
Sugar compositionFlower age (%, values are mean±1 SD)(hao) n Sucrose Fructose Glucose aa(HS) ph r hr
1 (4) 3 73.99±3.93 15.68±3.29 10.18±1.06 +(0) + 2.861 0.6492 (16) 3 64.32±4.45 18.87±2.62 16.79±1.88 − + 1.803 0.8893 (28) 2 62.99±0.46 22.87±0.44 14.13±0.79 +(0) + 1.702 0.6174 (40) 2 58.71±0.75 19.17±1.00 22.41±0.67 +(1) + 1.411 1.1695 (52) 3 62.75±0.30 17.40±0.51 19.63±0.06 − + 1.694 1.1286 (64) 3 60.91±2.17 18.67±1.00 20.41±1.15 +(0) + 1.558 1.0937 (76) 2 61.40±0.20 16.78±1.06 21.81±0.86 +(0) + 1.591 1.2998 (88) 3 61.79±2.18 15.85±0.95 22.35±1.22 +(0) + 1.617 1.410
Overall mean 21 64.90±5.91 18.07±2.99 16.99±4.86 1.851 0.940Range (58.18–78.61) (11.27–23.32) (8.91–23.22)

NECTAR OF MANDEVILLA PENTLANDIANA 215
T 2. Nectar sugar composition of Mandevilla pentlandiana throughout the flowering period. Ab-breviations: n=number of samples; r=sugar ratio; hr=hexose ratio
Sugar compositionSampling (%, values are mean±1 SD)date n Sucrose Fructose Glucose r rh
Dec 27, 1994 2 78.24±2.21 13.35±2.12 7.89±0.82 3.683 0.591Jan 18, 1995 2 66.91±0.73 24.48±0.95 8.59±1.68 2.023 0.351Jan 27, 1995 2 65.62±1.85 18.65±0.83 15.71±1.03 1.909 0.842Feb 12, 1995 2 61.81±2.44 15.56±1.28 22.61±1.15 1.619 1.453Feb 28, 1995 2 60.88±1.17 17.82±0.94 21.28±0.25 1.557 1.194
Overall mean 10 66.69±6.67 17.97±4.06 15.21±6.53 2.009 0.846Range (60.05–79.80) (11.87–25.16) (7.31–23.43)
amino acids in very low concentrations (Ζ7.6 lg ml−1, i.e. 0–1 in the histidinescale). There are some variations in the nectar sugar proportions throughout theflower lifespan (Table 1). Recently opened flowers have higher values of sucrosecompared with the remaining stages. The hexose ratio (hr) varies as the flower ages.One- and 2-day old flowers have more fructose than glucose whereas the inversepattern is observed in 3- and 4-day old flowers.
The nectar chemical composition data of samples taken through the floweringseason are presented in Table 2. Variations in the proportions of sugars were alsodetected here.
Nectar production
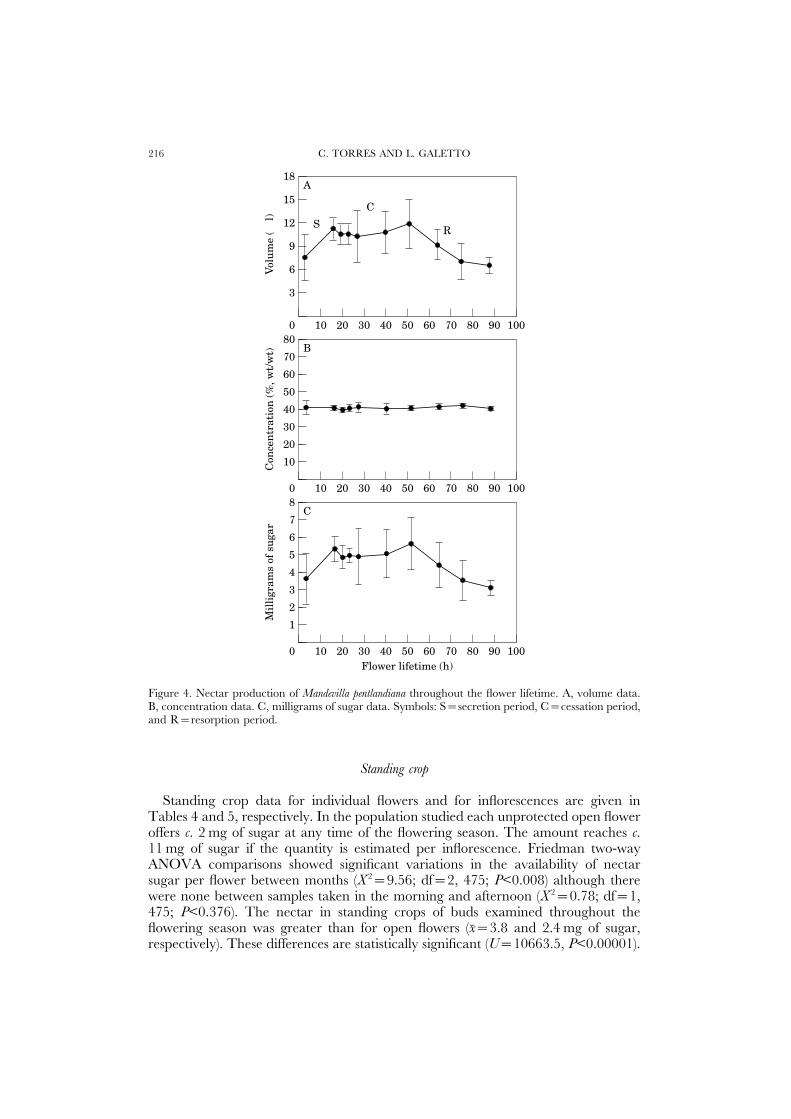
Flowers produced most nectar during bud-stage, while nectar secretion activityceased towards the end of the first day after flower opening. Production throughoutthe life of the flower is shown in Figure 4. Sugar concentration was quite constantduring anthesis (Fig. 4B), consequently, the patterns of volume and mg of sugarproduced were comparable (Fig. 4A, C). Recently opened flowers contained morethan 60% of the total nectar sugar production (Fig. 4C). Active nectar secretioncontinued during the first hours after the flower opening reaching c. 5–6 mg ofsugar at the end of secretion period (x=5.3±0.7 mg). The NNPR was constant(0.14 mg h−1) throughout; a nectar cessation period was inferred because cumulativetotal nectar did not change during the following 35 h. Finally, the nectar wasresorbed (NNRR=0.09 mg h−1) until corolla abscission starting from the third dayafter flower opening (Fig. 4A, C).
Removal effects
Data on nectar removal during the secretion period are included in Table 3.There was a noteworthy increase (c. 40%) in the total sugar produced by a flowerafter nectar removal. ANOVA comparisons showed significant differences [F[5,31]=4.63; P<0.004] between the control set (0) and the sets which were near the end ofthe most prolific secretion period when the removal schedule began (3 and 4).
C. TORRES AND L. GALETTO216
100
8C
Flower lifetime (h)
Mil
ligr
ams
of s
uga
r
50
4
3
2
1
10 20 30 40 60 70 80 900
5
6
7
100
80B
Con
cen
trat
ion
(%
, wt/
wt)
50
40
30
20
10
10 20 30 40 60 70 80 900
50
60
70
100
18A
Vol
um
e (
l)
50
12
3
9
6
10 20 30 40 60 70 80 900
15
S
C
R
Figure 4. Nectar production of Mandevilla pentlandiana throughout the flower lifetime. A, volume data.B, concentration data. C, milligrams of sugar data. Symbols: S=secretion period, C=cessation period,and R=resorption period.
Standing crop
Standing crop data for individual flowers and for inflorescences are given inTables 4 and 5, respectively. In the population studied each unprotected open floweroffers c. 2 mg of sugar at any time of the flowering season. The amount reaches c.11 mg of sugar if the quantity is estimated per inflorescence. Friedman two-wayANOVA comparisons showed significant variations in the availability of nectarsugar per flower between months (X 2=9.56; df=2, 475; P<0.008) although therewere none between samples taken in the morning and afternoon (X 2=0.78; df=1,475; P<0.376). The nectar in standing crops of buds examined throughout theflowering season was greater than for open flowers (x=3.8 and 2.4 mg of sugar,respectively). These differences are statistically significant (U=10663.5, P<0.00001).

NECTAR OF MANDEVILLA PENTLANDIANA 217
T
3.N
ecta
rre
mov
alpe
rfor
med
atth
etim
eof
the
secr
etio
npe
riod
ofM
ande
villa
pent
land
iana
flow
ers.
Dat
aar
em
eans±
1SD
ofne
ctar
volu
me
(ll),
conc
entr
atio
n(%
,w
t/w
t),an
dm
illig
ram
sof
suga
r(m
g)pe
rflo
wer
(n=
5).
Sym
bols:∗=
conc
entr
atio
nva
lue
not
reco
rded
due
toth
ela
ckof
nect
arvo
lum
e
Set#
Sam
plin
gtim
eT
otal
amou
ntpr
oduc
ed1s
tda
y2n
dda
y3r
dda
y20
:00
h8:
30h
12:0
0h
15:3
0h
19:0
0h
8:30
h
mg
3.8±
1.1
1.7±
0.6
00
00.
3±0.
65.
5±0.
45
ll8.
2±2.
23.
2±1.
30
00
0.6±
1.3
11.4±
1.1
%39
.4±
4.2
45.2±
3.2
∗∗
∗42
.0±
0.0
mg
5.3±
0.6
0.2±
0.3
0.3±
0.6
00.
4±0.
56.
3±1.
34
ll11
.2±
1.3
0.4±
0.5
0.6±
1.3
01.
0±1.
413
.2±
3.0
%40
.4±
1.5
48.5±
2.1
42.0±
0.0
∗34
.0±
0.7
mg
4.8±
0.6
0.7±
0.8
00.
8±0.
96.
4±1.
23
ll10
.5±
1.2
1.5±
1.7
01.
7±2.
013
.7±
2.9
%39
.2±
1.5
43.0±
1.4
∗39
.5±
2.1
mg
4.9±
0.4
0.3±
0.6
0.6±
1.0
5.7±
1.3
2ll
10.5±
1.2
0.6±
0.9
1.3±
2.3
12.1±
2.0
%40
.2±
1.8
49.0±
0.0
40.0±
0.0
mg
4.8±
0.8
1.4±
1.3
5.7±
0.6
1ll
10.4±
1.8
3.0±
2.9
12.2±
1.1
%39
.4±
0.8
42.5±
0.7
mg
4.3±
0.5
4.3±
0.5
0ll
9.0±
1.1
9.0±
1.1
%41
.1±
1.6
C. TORRES AND L. GALETTO218
T 4. Flower nectar standing crop throughout the flowering period in Mandevilla pentlandiana. Valuesare mean±1 SD. Abbreviations: n=number of sampled flowers
December January February
Morning mg of sugar 2.2±1.86 2.5±1.82 2.5±1.99ll of nectar 4.8±3.94 6.0±4.22 5.1±4.26
concentration (%) 39.7±3.92 35.6±4.73 41.1±4.30(n=84) (n=72) (n=81)
Afternoon mg of sugar 3.3±2.24 1.6±1.71 1.9±2.18ll of nectar 6.2±4.36 3.3±3.54 3.5±4.21
concentration (%) 44.2±2.73 42.1±4.51 46.7±5.13(n=84) (n=69) (n=86)
T 5. Nectar standing crop (mg) throughout the flowering period in Mandevilla pentlandiana
Mean±SD Minimum Maximum Sample size
Buds 3.83±1.70 1.02 9.00 78Open flowers 2.30±2.04 0.0 10.52 476Inflorescences 10.88±9.55 0.0 39.33 65
December 15.63±9.99 2.32 34.96 24January 8.48±3.67 0.70 13.89 20February 7.73±11.00 0.0 39.32 21
The results also show that, in general, there is a higher nectar availability at thebeginning of the flowering season than later (H=13.78; df=2, 64; P<0.001). Themean number of open flowers per inflorescence found in December is greatercompared with January and February (x=6.12±2.11, x=4.17±1.15, x=4.20±1.88, respectively). Significance was confirmed by Kruskal–Wallis one-wayANOVA and by the Wilcoxon’s post hoc test (H=12.93; df=2, 64; P<0.001).
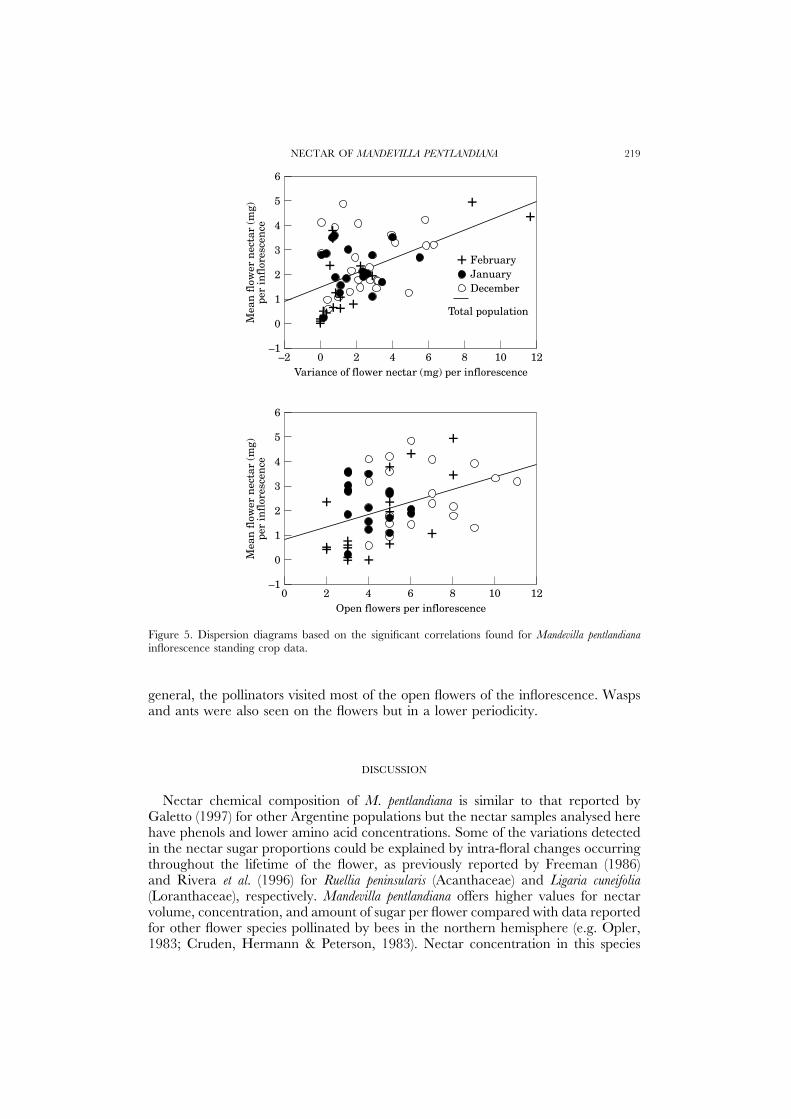
Regression analyses were performed between the mean sugar quantity per flowerin each inflorescence, the variance, and the open-flower number per inflorescence(n=62). The three combinations showed positive significant correlation: mean/variance (y=0.99+0.87 x, r2=0.49, P<0.0001), mean/open flowers (y=−0.54+1.20 x, r2=0.40, P<0.0013), variance/open flowers (y=−0.61+0.85 x,r2=0.50, P<0.00001). The greater the number of open flowers and the nectarvariance, the more the mean reward quantity per flower available in the inflorescence(Fig. 5).
Floral visitors
No flower visitors were noticed during a total of 33 h of nocturnal observationsspread over nine days of the flowering period. Diurnal insects frequented the flowersof this species, including bumblebees (Bombus morio Swed and B. opifex Sm.; Fig. 2C,D) and honeybees (Apis mellifera L.; Fig. 2B). The principal flower visitor was A.mellifera, followed by both species of bumblebees. They arrived at the inflorescenceand selected flowers exclusively seeking nectar. Bumblebees were more abundantduring cloudy days whereas honeybees were always present. Hummingbirds(Chlorostilbon aureoventris D’Orbigny & Lafresnaye) occasionally visited the flowers. In
NECTAR OF MANDEVILLA PENTLANDIANA 219
12
6
–10
Open flowers per inflorescence
Mea
n f
low
er n
ecta
r (m
g)
per
infl
ores
cen
ce 4
5
3
2
1
0
2 4 6 8 10
+
++
12
6
–1–2
Variance of flower nectar (mg) per inflorescence
Mea
n f
low
er n
ecta
r (m
g)
per
infl
ores
cen
ce 4
5
3
2
1
0
2 4 6 8 10
+++++
++
++
++
+
+
+
0
+++ ++++
++
++
+++
++ +
+ FebruaryJanuaryDecember
Total population
Figure 5. Dispersion diagrams based on the significant correlations found for Mandevilla pentlandianainflorescence standing crop data.
general, the pollinators visited most of the open flowers of the inflorescence. Waspsand ants were also seen on the flowers but in a lower periodicity.
DISCUSSION
Nectar chemical composition of M. pentlandiana is similar to that reported byGaletto (1997) for other Argentine populations but the nectar samples analysed herehave phenols and lower amino acid concentrations. Some of the variations detectedin the nectar sugar proportions could be explained by intra-floral changes occurringthroughout the lifetime of the flower, as previously reported by Freeman (1986)and Rivera et al. (1996) for Ruellia peninsularis (Acanthaceae) and Ligaria cuneifolia(Loranthaceae), respectively. Mandevilla pentlandiana offers higher values for nectarvolume, concentration, and amount of sugar per flower compared with data reportedfor other flower species pollinated by bees in the northern hemisphere (e.g. Opler,1983; Cruden, Hermann & Peterson, 1983). Nectar concentration in this species

C. TORRES AND L. GALETTO220
appears not to be related to the influence of the tubular corolla, since otherApocynaceae species with more variability in their nectar sugar concentrations havecomparatively larger corolla tubes (cf. Galetto, 1997).
In terms of pattern of production, M. pentlandiana can be characterized as a slow-producer following the definition of Cruden et al. (1983). It secretes only at thebeginning of anthesis until a critical amount has accumulated (c. 10 ll); thereafter,secretion ceases. Nevertheless, removals cause the resumption of nectar secretion,so that the flower can offer some nectar to further visitors after it is depleted. Theobserved decrease in nectar solute in 3-day old bagged flowers can be inferred asresorption based upon the constancy of nectar concentration throughout anthesis.In addition, this process may be facilitated because nectar remains in contact withnectary stomata throughout the flower lifetime. There are some species of otherfamilies with a similar nectar secretion pattern, with nectary stomata, and withflowers which last 4 days (Combretaceae: Bernardello et al., 1994; Fabaceae: Isele,1996; Loranthaceae: Rivera et al., 1996), all of them visited by a great variety ofpollinators.
Some attributes, such as tubular corolla, greenish-white flowers, nocturnal nectarsecretion and sucrose-dominant nectar, may well have evolved in response tosphingid visitors. Our field observations, on the other hand, do not support thishypothesis, because M. pentlandiana flowers are visited by a wide array of visitors. Inpractice, the floral traits that characterize syndromes are generally of little use inpredicting the pollinators of a given plant species (Herrera, 1996 and citationstherein), and do not always represent adaptations to flower visitors.
Rathcke (1992) suggested that NNPR is probably a better estimator of nectaravailability than computing from the size of a standing crop. Nevertheless, theproduction pattern of M. pentlandiana indicates that its NNPR is a function of bothflower age and nectar removal, as has been demonstrated for other plant species(e.g. Bernardello et al., 1994; Rivera et al., 1996). Therefore, a standing crop stillprovides the best picture of resource availability in this species.
The distribution of nectar within a standing crop is mainly a function of plantnectar manipulations (secretion, cessation, resumption and resorption) and pollinatorforaging behaviour. According to the literature (e.g. Pleasants & Zimmerman, 1983;Zimmerman & Pyke, 1986), the relationship between the mean nectar volume andits standard deviation indicates both the frequency of pollinator visitation and thepattern of the plant nectar production. The standing crop mean nectar volume andthe standard deviation found for M. pentlandiana (x=4.83±4.24 ll/flower; i.e. x=2.30±2.04 mg/flower from Table 5) are not consistent with its nectar productionpattern, because the standing crop pattern in which the mean nectar volumeapproximately equals the SD of the distribution corresponds to a constant nectarproduction throughout the flower lifespan (Zimmerman & Pyke, 1986).
The distribution and presentation of nectar to insects has been demonstrated toinfluence pollinator visitation rates, pollinator movements, and consequently plantfitness (e.g. Cruden et al., 1983; Zimmerman & Pyke, 1986; Real & Rathcke, 1988;Mitchell & Waser, 1992). In particular, bumblebees choosing higher rewarding floraltypes over less rewarding ones, have been shown to exhibit discerning foragingbehaviour, and seem to prefer the plant species with the less variable reward (e.g.Real, 1981; Real, Ott & Silverfine, 1982; Harder & Real, 1987; but also see Real& Rathcke, 1988). These insects are the main visitors of M. pentlandiana and contraryto assumptions, the plant shows numerous ways to increase nectar variability (e.g.

NECTAR OF MANDEVILLA PENTLANDIANA 221
nectar production varies as a function of flower age, nectar removal generatesresumption of nectar production, nectar resorption in old-flowers causes a dimin-ishment in nectar availability) and the bumblebees forage on most of the openflowers of each inflorescence. This behaviour could be explained by: (a) thecomparatively few open flowers per inflorescence that M. pentlandiana displays throughthe flowering season, (b) the fact that most (75%) open flowers present some nectar,and (c) the foraging ‘rules of movement’ that bumblebees seem to follow (e.g. Pyke,1978; Waddington & Heinrich, 1979; Zimmerman, 1981). According to Waddington& Heinrich (1979), the rule of visiting the closest flower resulted in the bees makingseveral visits per inflorescence rather than skipping about between flowers of differentinflorescences.
Variability in the amounts of nectar presented to pollinators will be greaterin species producing no nectar during the bud stage relative to those whoseflowers open with a little nectar already present in them (e.g. Pleasants, 1983;Zimmerman & Pyke, 1986). This could be an additional way to increase thevariation in nectar reward by the plant. In M. pentlandiana nectar can be foundin buds; the larger amount offered by this flowering stage when compared tothe open flower standing crop (66.5% more mg of sugar) is a main source ofvariation. Thus, the implications of bud nectar on standing crop variability willbe conditioned or determined both by the plant nectar production pattern andthe pollinator foraging pressure.
Naturally, M. pentlandiana tends to cause nectar variability within inflorescences.This assertion is made by considering that the largest mean flower standing cropvalues which have greatest variances are similar to those of the unvisited flowers.In addition, variability is enhanced as the inflorescence increases the number ofopen-flowers. On the contrary, the pollinators when foraging on most open-flowersof each inflorescence decrease nectar standing crop variability.
The large values found for December nectar standing crops can be explained bya lower frequency of pollinators foraging on M. pentlandiana at this flowering timeor by a distinctive pattern of nectar presentation by the plant population. Theformer argument is not considered because bud standing crops indicate that periodicpollinator visits occurred since the beginning of the flowering season. This could beinferred due to the higher values of bud standing crops compared to any of thesampled open-flower periods. On the other side and in view of the latter argument,the large number of open-flowers that inflorescences have at the beginning of theblooming can affect December standing crops.
What are the implications for plant reproductive biology in analysing M.pentlandiana nectar variability? Some speculations can be made considering thefemale and male functions separately. Pyke (1991) has demonstrated that nectarproduction entails a cost to the plant in terms of the number of seeds produced(female function). On the other hand and considering the Bateman’s principle,hermaphroditic plants also have evolved traits to increase the transfer of pollento conspecific stigmas, emphasising the male function at anthesis. The sourcesof nectar variability in M. pentlandiana indicate that some of them seem to bemainly linked with the female function (nectar resorption, nectar cessation) whileothers with the male one (early and comparatively large nectar availability,variation in nectar production as the flower ages, resumption of nectar secretionafter removals). A variable but renewed reward would determine additional visitsto the flower maximizing the amount of pollen removed by way of multiple

C. TORRES AND L. GALETTO222
insect probes. On the contrary, nectar ‘saving’ would allow the plant to reallocateresources for reproduction: e.g. number of matured fruits, seeds per fruit, seedquality etc. The latter topic is currently under investigation as an attempt toclarify the links among nectar production features, pollinator behaviour, breedingsystem, and plant reproductive success.
ACKNOWLEDGEMENTS
The authors sincerely thank Arthur Davis for valuable and constructive suggestionson the final draft, G. Bernardello for useful comments on early versions of thismanuscript, Arnaldo Mangeaud for helpful collaboration with statistical analysis,Claudio Sosa for insect identification, ‘Consejo de Investigaciones Cientıficas yTecnicas de la Provincia de Cordoba (CONICOR)’, ‘Consejo Nacional de In-vestigaciones Cientıficas y Tecnicas’, and ‘Secretarıa de Ciencia y Tecnica de laUniversidad Nacional de Cordoba’ for financial support, and CONICOR for thefellowship to CT.
REFERENCES
Baker HG, Baker I. 1975. Studies of nectar-constitution and pollinator-plant coevolution. In: GilbertLE, Raven PH, eds. Coevolution of animals and plants. Austin: University of Texas Press, 100–140.
Baker HG, Baker I. 1983. Floral nectar sugar constituents in relation to pollinator type. In: JonesCE, Little RJ, eds. Handbook of experimental pollination biology. New York: Van Nostrand Reinhold,117–141.
Bernadello LM, Galetto L, Juliani HR. 1991. Floral nectar, nectary structure and pollinators insome Argentinean Bromeliaceae. Annals of Botany 67: 401–411.
Bernadello LM, Galetto L, Rodrıguez IG. 1994. Reproductive biology, variability of nectarfeatures, and pollination of Combretum fruticosum (Combretaceae) in Argentina. Botanical Journal of theLinnean Society 114: 293–308.
Burquez A, Corbet SA. 1991. Do flowers reabsorb nectar? Functional Ecology 5: 369–379.Conn HJ, Darrow MA, Emmel VM. 1960. Staining procedures. Baltimore: Williams & Wilkins.Corbet SA. 1978. Bee visits and the nectar of Echium vulgare. In: Richards AJ, ed. The pollination of
flowers by insects. London: Academic Press, 21–30.Cruden RW, Hermann S, Peterson S. 1983. Patterns of nectar production and plant pollinator
coevolution. In: Bentley B, Elias T, eds. The biology of nectaries. New York: Columbia UniversityPress, 80–125.
Ezcurra C. 1981. Revision de Apocinaceas de la Argentina. Darwiniana 23: 367–474.Fallen ME. 1986. Floral structure in the Apocynaceae: morphological, functional, and evolutionary
aspects. Botanische Jahrbucher fur Systematik 106: 245–286.Feinsinger P. 1987. Approaches to nectarivore-plant interactions in the New World. Revista Chilena
de Historia Natural 60: 285–319.Freeman CE. 1986. Nectar-sugar composition in an individual of Ruellia peninsularis (Acanthaceae).
Madrono 33: 300–302.Galetto L. 1997. Flower structure and nectar chemical composition in three Argentine Apocynaceae.
Flora 192: 197–207.Galetto L, Bernardello LM, Juliani HR. 1994. Nectar secretion pattern and removal and
concentration–volume variations effects in Pyrostegia venusta (Bignoniaceae). New Phytologist 127:465–471.
Gottsberger G, Schrawen J, Linskens HF. 1984. Amino acids and sugars in nectar, and theirputative evolutionary significance. Plant Systematics and Evolution 145: 55–77.
Harder LD, Real L. 1987. Why are bumblebees risk averse? Ecology 68: 1104–1108.

NECTAR OF MANDEVILLA PENTLANDIANA 223
Heinrich B. 1983. Insect foraging energetics. In: Jones CE, Little RJ, eds. Handbook of experimentalpollination biology. New York: Van Nostrand Reinhold, 187–214.
Herrera CM. 1996. Floral traits and plant adaptation to insect pollinators: a Devil’s advocateapproach. In: Lloyd DG, Barrett SCH, eds. Floral biology: studies on floral evolution in animal-pollinatedplants. New York: Chapman & Hall, 65–87.
Isele IC. 1996. Estudios sobre la biologıa reproductiva del Ceibo (Erythrina crista-galli, Papilionoideae-Fabaceae). Unpublished Undergraduate Thesis, Universidad Nacional de Cordoba, Argentina.
Kearns CA, Inouye DW. 1993. Techniques for pollination biologists. Niwot: University Press of Colorado.Mitchell RJ, Waser NM. 1992. Adaptive significance of Ipomopsis aggregata nectar production:
pollination success of single flowers. Ecology 73: 633–638.Murawski DA. 1987. Floral resource variation, pollinator response, and potential pollen flow in
Psiguria warscewiczii. Ecology 68: 1273–1282.Opler PA. 1983. Nectar production in a tropical ecosystem. In: Bentley B, Elias T, eds. The biology of
nectaries. New York: Columbia University Press, 30–79.Percival MS. 1961. Types of nectars in angiosperms. New Phytologist 60: 235–281.Pleasants JM. 1983. Nectar production patterns in Ipomopsis aggregata (Polemoniaceae). American Journal
of Botany 70: 1468–1475.Pleasants JM, Zimmerman M. 1983. The distribution of standing crop of nectar: what does it
really tell us. Oecologia (Berlin) 57: 412–414.Pyke GH. 1978. Optimal foraging: movements patterns of bumblebees between inflorescences.
Theoretical and Population Biology 13: 72–98.Pyke GH. 1991. What does it cost a plant to produce floral nectar? Nature 350: 58–59.Rathcke BJ. 1992. Nectar distributions, pollinator behavior, and plant reproductive success. In:
Hunter MD, Ohguishi T, Price PW, eds. Effects of resource distribution on animal–plant interactions. NewYork: Academic Press, 113–138.
Real L. 1981. Uncertainty and pollinator–plant interactions: the foraging behavior of bumblebeesand wasps at artificial flowers. Ecology 62: 20–26.
Real L, Ott JR, Silverfine E. 1982. On the trade-off between the mean and variance in foraging:effects of spatial distribution and colour preference. Ecology 63: 1617–1623.
Real L, Rathcke BJ. 1988. Patterns of individual variability in floral resources. Ecology 69: 728–735.Rivera GL, Galetto L, Bernardello L. 1996. Nectar secretion pattern, removal effects, and breeding
system of Ligaria cuneifolia (Loranthaceae). Canadian Journal of Botany 74: 1996–2001.Schick B. 1980. Untersuchungen uber die Biotechnik der Apocynaceenblute. I. Morphologie und
Funktion des Narbenkopfes. Flora 170: 394–432.Schick B. 1982. Untersuchungen uber die Biotechnik der Apocyceenblute. I. Bau und Funktion des
Bestaubungsapparates. Flora 172: 347–371.Simpson BB, Neff JL. 1983. Evolution and diversity of floral rewards. In: Jones CE, Little RJ, eds.
Handbook of experimental pollination biology. New York: Van Nostrand Reinhold, 142–159.Sokal RR, Rohlf FJ. 1995. Biometry. San Francisco, California: W.H. Freeman.Stiles FG, Freeman CE. 1993. Patterns in floral nectar characteristics of some bird-visited plant
species from Costa Rica. Biotropica 25: 191–205.Sweeley EC, Bentley R, Makita M, Wells WW. 1963. Gas liquid chromatography of trimethylsilyl
derivatives of sugars and related substances. Journal of the American Chemical Society 85: 2497–2507.Waddington KD, Heinrich B. 1979. The foraging movements of bumblebees on vertical ‘in-
florescences’: an experimental analysis. Journal of Comparative Physiology 134: 113–117.Waser NM, Mitchell RJ. 1990. Nectar standing crops in Delphinium nelsonii flowers: spatial auto-
correlation among plants? Ecology 71: 116–123.Woodson RE. 1933. Studies in Apocynaceae IV. The American genera of Echitoideae. Annals of the
Missouri Botanical Garden 20: 605–790.Zimmerman M. 1981. Patchiness in the dispersion of nectar resources: probable causes. Oecologia
(Berlin) 49: 154–157.Zimmerman M, Pyke GH. 1986. Reproduction in Polemonium: patterns and implications of floral
nectar production and standing crops. American Journal of Botany 73: 1405–1415.
Related Documents











