Patching up the crypt Innate immune cells orchestrate intestinal regeneration Mónica Romera Hernández

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Patching up the cryptInnate immune cells orchestrate
intestinal regeneration
Mónica Romera Hernández


Patching up the crypt: Innate immune cells orchestrate

....Copyright © 2018
All rights reserved.
in any form or by any means without permission from the author. The copyright of
Layout: Egied Simons Cover: Iliana Boshoven-Gkini
The work described in this thesis was performed at the Department of Hematology at the
the Netherlands.

Patching up the crypt:
Innate immune cells orchestrate
aangeboren afweer systeem
op gezag van de
Prof.dr. H.A.P. Pols
De openbare verdediging zal plaatsvinden op
3 mei 2018 om 11:30 uur
door
Mónica Romera Hernández

DOCTORAL COMMITEE
Supervisor: Prof.dr. J.J. Cornelissen
Other members: Prof.dr. I.P. Touw
Prof.dr. F. Koning
Co-supervisor: Dr. T. Cupedo

A mi iaio, de extrema honra y dura proeza


CONTENTS
Chapter 1: Aims and outline of this thesis 26
of Group 3 innate lymphoid cells
English summary

1

GENERAL INTRODUCTION
HARNESSING THE IMMUNE SYSTEM TO ENHANCE INTESTINAL BARRIER FUNCTION
Based in part on: Damage control: Ror

ABSTRACT
1
epithelium by local stem cells, that are contained within a niche in the crypts of Lieberkühn.

11
INTRODUCTION
2,3. Key physical components
self-contained enteric nervous system.
as well as interspersed between epithelial cells, and these contain cell types belonging to
A dynamic balance between the immune system and the epithelium is vital to ensure
4–6.
environment.
.
H , yet
1

12
The family of ILCs can be subdivided in three main types based on their master
+ + ILC3 subtypes
11,12
ILCs depend on c13,14 but in
-lymphoid progenitors ( + integrin 4+-
4+
Id2hi
and ILCP progenitors are able to generate ILC1, ILC2 and NKp46+ + ILC3s . Id2+ -
4+ ILC3s12,20. The
21
is not known22–24
.
mirroring TH cell subsets . ILC1s express T-bet, produce IFN-
H . GATA-3hi
IL-33, as TH
.
t for their development.
Chapter 1

13
-lymphoid progenitor (
.
CD8+ T cells due to its developmentally dependence on T-bet and Eomes as well as their ability to secrete IFN- , granzyme and perforins . In contrast to ILC1s, NK cells are localized
+ ILC3s can produce IFN as they adopt an ILC1 phenotype by t . Also, NKp46+ +
ILC3san ILC1-like phenotype .
1

14
61 have been characterized in humansresident human ILC precursors, using humanized mice21. Hopefully, further analysis in
ILC development62,63.
64
imprint their gene expression are established very early during development compared to
. In line with this, human
been shown that ILCs are generated in the bone-marrow or fetal liver and then migrate to
466 + ILC3s migrate to
T cells68
resident and maintained by self-renewal in various microenvironments, but hematogenous . Therefore, it is
. Similarly, human adult CD34+ t and an
21. However, the signals that
Chapter 1

cells. Accordingly, mice lacking t .
. Post-birth, LTi cell counterparts also interact with mesenchymal stromal cells to form discrete ILC3 clusters termed cryptopatches . PP are
81 82,83. In more detail, the primary events leading
1 2+ LTi cells and LT + mesenchymal cells, termed
Tumor necrosis factor-a (TNFPP development . Also, PP are absent in mice with simultaneous defects in both CXCL13-
. Cryptopatches predominantly contain adult LTi cells and DCs organized around stromal
cells drives the expression of CCL20 and cells in CPsincluding CCL20 , which recruit B cells , which further grow into mature B cell follicles with the help of TNF produced by DCs and macrophages in response to bacterial compounds
. Mature ILFs generate IgA-producing B cells that target the microbiota, controlling the bacterial community during homeostasis
1 2 by LTi cells . These organized structures provide
1

16
response to damage.
. ILC3s produce 44,100,101. ILC3s are subdivided by the mutually exclusive expression of
+
SILTs . NKp46+
+
as well as HLA+ ILC3s in humans, and these cells are able to regulate CD4+ T cell responses 102.
103
+ ILC3s, through 104
circadian rhythms in the epithelium to control fat storage and other host metabolic
primarily IL-23 and IL-1
. This,
110,111
Chapter 1

Enterococous faecalis and Salmonella spp and 112–114.
. ILC3-derived IL-22 contributes to immunity in a diverse range of clinically-relevant
116, Rotavirus , 118, as well . Extensive research has demonstrated that ILC3s prevent morbidity
in the early phase of cell responses
123,124. Salmonella, for example, has evolved to
.+ and
.
and GM-CSF, in an
manner .
68. MHCII+
+
68,102,130. MHCII+
102,131.
and ILC3s can promote immune responses132
ILC3s .
+ ILC3s and have elevated 133.
responses or aberrant B cell responses, even long before disease onset.
1

18
induces IL-22 expression on ILC3s in the skin. Both 134. It
NKp44 in the skin and blood . ,
138, systemic sclerosis , experimental autoimmune 140
141
ILC3s have also been implicated in the development of cancer, such as in the gut where a 142.
Microbiota-induced IL-1GM-CSF143. GM-CSF maintains CD103+
144. In turn, GM-CSF modulates mononuclear phagocyte 143
128.
by ILC3s , likely through myeloid cells.
Similarly, a new emerging area is the study of ILC3-enteric nervous axis. It has been
of adult ILC3s.
cell-mediated killing in the context of GvHD. And in this thesis, we show that ILC3s contribute
Chapter 1

Lieberkühn
secretory cells .
absorb nutrients and water. Goblet cells secrete mucus-associated molecules that generate a mucus layer to keep microorganisms separated from the epithelium
metabolism
160. Paneth cells are also a source of + ISCs in in vitro cultures, thus being
considered a pivotal component of the ISC niche161
host-defense.
characterized by the expression of the Wnt pathway target gene leucine-rich repeat-+ ISCs
1

20
162
to CBCs, Bmi1+ ISCs are indispensable for epithelial maintenance at steady-state163. They 164 and label-retaining cells or
, mTert , Lrig . However, these genes are broadly expressed by numerous cells in
they have also been termed reserve ISCs
+ cells and Prox1+ cells, as well as enteroendocrine lineage cells possess . In
parallel, another study has reported that Bmi1-GFP+ cells and other secretory cells, including + + + stem
.
.
Gradients of Wnt ligands exist with high expression in crypts and lower expression towards the villi
-catenin signaling pathway. By knocking-out secreted Wnt antagonists, such as , , and the Wnt target gene 180–183
Chapter 1

21
184 Fzd7in vivo
ligands186. Mesenchymal cells, that underlie the epithelium of the crypts, have also been reported to enhance Wnt signaling pathway in ISCs. Even though there is also redundancy when compared to Paneth cells, it is thought that non-epithelial Wnt signals could provide a secondary physiological source of Wnt .
growth factor--catenin suppression. Consequently, BMP levels are higher in
188. At the base of the crypt, mesenchymal-derived . Noggin-1 and Noggin-2, as well as
chordin-like 1 are also highly expressed in the submucosa region by the mesenchyme to suppress BMP signaling .
of epithelial progenitors towards the secretory cell lineage .
and
.
EGF signaling regulates cellular processes that include cell-cycle progression, survival and
demonstrated in the underlying mesenchymal cells and in Paneth cells161. EGF is currently
.
1

22
.
As discussed in the previous paragraphs, there are intrinsic epithelial mechanisms that
cells sense the external forces and respond to the environment200–202
neurons, intraepithelial lymphocytes, smooth muscle cells of the muscularis mucosa, lymph and vascular endothelial cells and a variety of mesenchymal cells, such as pericryptal
203,204
.
.
Chapter 1

23
donor marrow did not have any impact on GvHD-associated organ pathology.
- + + t+
host-derived ILCs appeared to be targeted by the GvHD as their numbers subsided during . Combined
GvHD-induced cell death.
strengthened by the fact that a similar subset of radioresistant NKp46- + + t+ 210.
210. Conversely, absolute levels of IL-22 in the thymus were increased 2 to 3-fold in experimental models of sublethal-TBI without HSCT, lethal TBI and syngeneic HSCT, and T cell-depleted
intrathymic IL-22 was inversely correlated with the thymic cellularity210.
TBI led to an enhancement of thymopoiesis. Thus, the thymic ILC3s are not responsible for
1

24
210
The mechanisms that regulated ILC-derived IL-22 also showed overlap with those found
in IL-23 producing CD103+
The skin also forms a highly organized physical and immunological barrier against the external environment and the microbiota. It is well established that a correct recruitment of
assessed. In an elegant study, ILC3s were shown to favor wound healing by enhancing the
During wound repair, epidermal Notch1 signaling is indispensable for wound closure ,
mediated the recruitment of ILC3s into wounded areas of the skin. ILC3 were required to 211.
and endothelial cells in lymph node anlagen.
Chapter 1
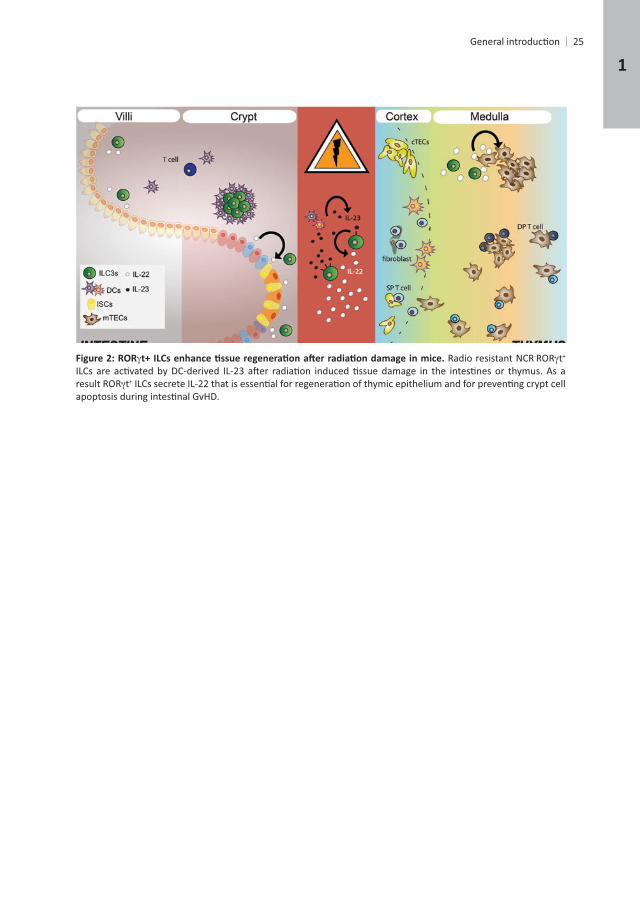
- t+
t+
1

26
AIMS AND OUTLINE OF THE THESIS
The
through which this is achieved.Chapter 1
In
stem cell maintenance post-damage.In chapter 3
+
unknown. Chapter 4
which resembled responses occurring during epithelial damage. This chapter proposes a model that supports a direct link between ILC3s in cryptopatches and the maintenance of
In chapter 5
Finally, a general discussion of the thesis is presented in chapter 6
Chapter 1

REFERENCES
1. Hanash, A. M. 37,
management of gut barrier leaking: The emerging role for mucosal barrier protectors. European Review for 19,
Gut Mesenchymal Stromal Cells in Immunity.
159,
19,
homeostasis. 17,
8. Walker, J. a, Barlow, J. L. & McKenzie, A. N. J. Innate lymphoid cells--how did we miss them? 13,
Innate lymphoid cells--a proposal for uniform nomenclature. 13,
cells. 41,
15,
cell lineage. 17,
NKp46 + cell subsets from Id2-dependent precursors.
cell lineages. 157,
lymphoid cells. Nature
factor NFIL3 directs the development of a common innate lymphoid cell precursor. Elife 3,
20. Chea, S. Lymphoid Progenitors to Notch Signaling. 14,
Aims and outline of this thesis
1

28
21. Lim, A. I.
cell progenitors in adult murine bone marrow. 31,
BM. Blood 117,
Blood
innate lymphoid cells.
191,
Lymphoid Cells. 37,
30. Yang, Q.
17,
Nature 517,
Lymphoid Cells. 37,
36. Moro, K. Nature 463,
Nature 464,
virus.
dependent on and independent of Notch. 13,
40. Yang, Q. 16,
Chapter 1

43,
42. Yu, Y.
34,
44. Klose, C. S. N. Nature 494,
M. A human natural killer cell subset provides an innate source of IL-22 for mucosal immunity. Nature 457,
of gut innate lymphocytes. 4,
Provide Innate Mucosal Immune Defense.
33,
14,
196,
Transforming growth factor- and Notch ligands act as opposing environmental cues in 9,
Cells in Salivary Glands. 44,
17,
2 innate lymphoid cells in the lungs. 17,
17,
165,
13,
43,
innate lymphoid cells. 41,
1

30
17,
167,
64. Yu, Y. Nature 539,
Shaped by the Microbiome. 166,
Blood 115,
mucosal draining lymph nodes. 6,
lymphoid and nonlymphoid organs.
41,
Lymphoid Cell Subsets. 44,
5,
7,
peripheral lymphoid organs and natural killer cells depends on the helix-loop-helix inhibitor Id2. Nature 397,
5,
Chapter 1

31
murine embryo. 9,
82. Hamada, H.
progenitors develop.
84. Cao, X.
3,
signaling through the lymphotoxin beta receptor. 9,
Systems. 5,
Nature 456,
Science
179,
7,
engender lymph nodes and Peyer’s patches. 17,
45,
1

32
Nature 464,
homeostasis at barrier surfaces by IL-22.
Nature
103. van de Pavert, S. A. immunity. Nature
104. Gomez de Aguero, M. The maternal microbiota drives early postnatal innate immune development. 351,
3,
36,
196,
108. Spencer, S. P. Barrier Immunity. 343,
35,
111. Sano, T. 163,
112. Pham, T. A. N. 16,
113. Pickard, J. M. sickness. Nature 514,
345,+
16,
Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance 39,
9,
14,
Chapter 1

33
37,
Complementarity and redundancy of IL-22-producing innate lymphoid cells. 17,
+
Monocyte Crosstalk Promotes Klebsiella Pneumoniae Clearance. 165,
128. Pearson, C. Elife 5,
33,
130. Goto, Y., Panea, C., Nakato, G., Cebula, A., Lee, C., Diez, M. G., Laufer, T. M., Ignatowicz, L. & Ivanov, I. I.
+ T cells. TL - 348. Science
132. von Burg, N. 111,
134,
2112–2118
13,
138. Ciccia, F.
74,
1

34
196,
13,
142. Chan, I. H. 7,
143. Mortha, A., Chudnovskiy, A., Hashimoto, D., Bogunovic, M., Spencer, S. P., Belkaid, Y. & Merad, M. Microbiota- 343,
9,
BioEssays
Methods in Enzymology
73,
Gastroenterology 116,
353,
Nature 449,
71,
11,
36,
Nature
Chapter 1

69,
154,
161. Sato, T. Nature 469,
14,
transgenic mice. 6,
Stem 3,
Gastroenterology 151,
Biology
Science 334,
16,
tumor suppressor. 149,
Gastroenterology 149,
Nature 495,
4,
1

36
Dickkopf-1.
c-Myc.
183. van Es, J. H. Dll1+ secretory progenitor cells revert to stem cells upon crypt damage. 14,
+ stem cells. Reports 4,
Gastroenterology
188. Kosinski, C.
catenin signaling. 36,
Reports
cells. Gastroenterology
columnar stem cells. Development 139,
morphogenesis.
Gastroenterology
Gastroenterology 139,
Nature 519,
Nature
Nature 539,
Science
Chapter 1

propria. 73,
37,
206. Lindemans, C. A. Nature
208. Pickert, G.
210. Dudakov, J. A. 336,
7,
1

2

TYPE 3 INNATE LYMPHOID CELLS MAINTAIN INTESTINAL EPITHELIAL STEM CELLS
AFTER TISSUE DAMAGE
Patricia Aparicio-Domingo , Julien J. Karrich1, Ferry Cornelissen1, Natalie Papazian1, Dicky Lindenbergh-Kortleve2, James A. Butler3,
Louis Boon4, Mark C. Coles3, Janneke N. Samsom2 and Tom Cupedo1
1
2
3Centre for Immunology 4Bioceros,
These authors contributed equally

40
ABSTRACT
understood.
severely impaired in the absence of ILC3s or the ILC3 signature cytokine IL-22. These data
cells..
Chapter 2

41
INTRODUCTION
1. Damage sustained by
2 3 and a 4.
epithelium
and in mice, upon DSS-.
12–14
al epithelial cells ex vivo16.
defense against enteric pathogens and containment of microbiota
. Most Nkp46+
21 + ILC3s are located in close 22.
mediated killing23.

42
MATERIALS AND METHODS
Mice t , Ncr1
whenever possible. Thy1+
g every other day, for 2 weeks.
Methotrexate
the last MTX dose.
6 t mice.
exposed to MTX.
Chapter 2

43
and centrifuge at 600 rpm for 3 min to separate the crypts from single cells. Crypts were
+ cells was performed blinded by at least two independent analysts. Pathology score was obtained as previously described (de
and
hours. Protein content of supernatants was determined by enzyme-linked immunosorbent

44
+Lin-NK1.1- + + NKp46
by Flow Cytometry and NKp46+ +
+
+ +
2
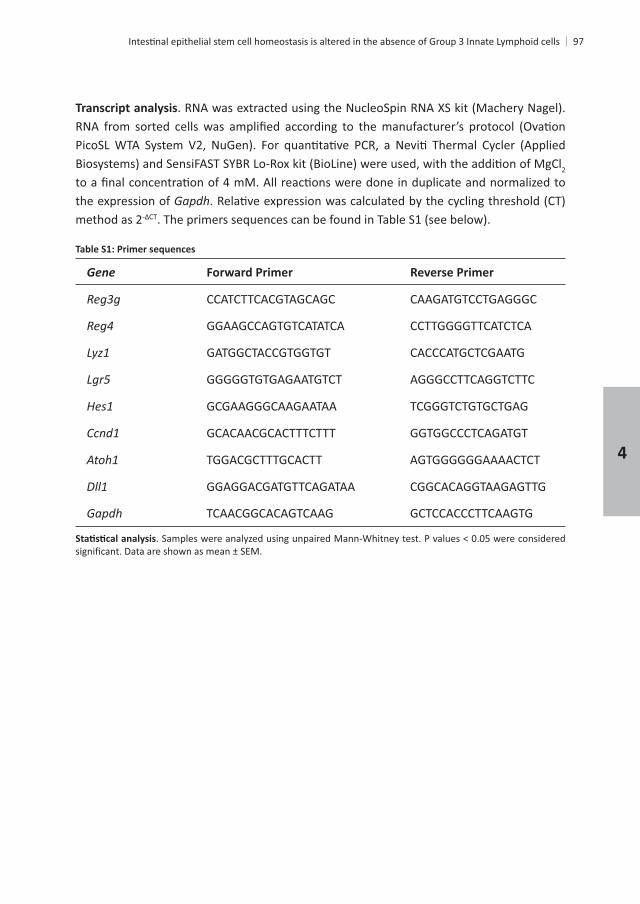
Gene Forward Primer Reverse Primer
Gapdh TCAACGGCACAGTCAAG GCTCCACCCTTCAAGTG
GGAGCCCCTTTGTAGACT CGGGAAACTTGCTGTG
CTCCCC CAGTCAGACAG CAATCGCCTTGATCTCTC
GACCCGCCTGAAGATATT ATCCGCATAGGTGGT AAC T
GGGGGCTTCCAGAACT GGGCCATAGAACTGATGAG
CCCCCTGAAACTGGTAGTA GTGGCAGTCTTCAGTTGG
CTTGGCGCAAAAGTGA TTGCTGGATGAGAACAGAA
CAAAAGGATGGTGACATGA GGGTTGTTGACCTCAAACT
Rorc GTGGGGACAAGTCATCTG CGGCCAAACTTGACAG
ACGTCGGGTTGAGAAGA GGATGTGGAGGCTAGATTC
Bax AAGGCCCTGTGCACTAA GAGGCGGTGAGGACTC
CGTGGCCTTTTTCTCC GGCTGCTGCATTGTTC
CGTGGACGCTAGAGAAAG AGCCAGACACTCCATGAC
± SEM.
Chapter 2

RESULTS AND DISCUSSION
cells in S-phase24
. To visualize damage responses by epithelial cells we assessed the number of cycling
+ crypt cells were reduced at day 1 but returned to baseline levels by day 4 . Conversely, transcripts for the
28
in mucosal wound healing8
peaking at day 1 of Stat3 had returned to baseline . These data show that MTX-induced small
. To
are indeed epithelial cell intrinsic or whether immune cells are involved, we administered MTX to mice pre-treated with Thy1-
mice treated with
at day 4
+ mice strongly reduced the recovery of cycling cells at day 4
Stat3 target gene and
mice
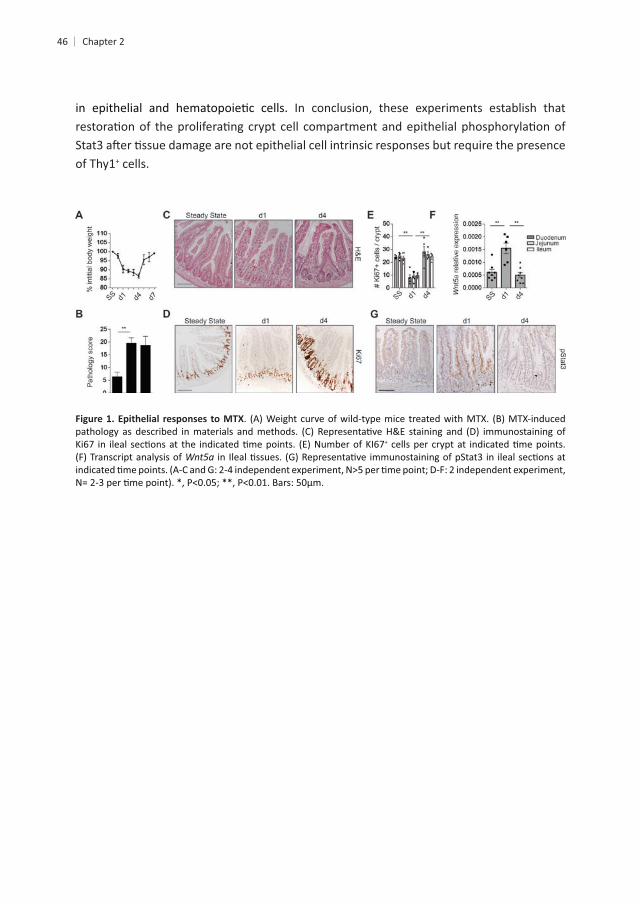
46
In conclusion, these experiments establish that
of Thy1+ cells.
+
Chapter 2
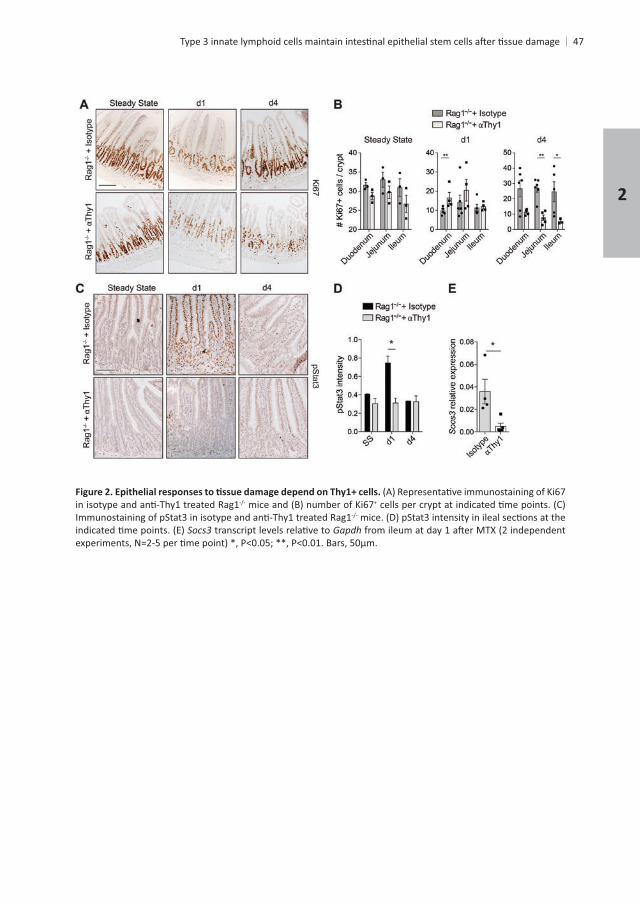
+
Gapdh

48
+ cells led us to hypothesize that Thy1+
+ and NKp46+
. Compared + , Rorc, , and , but
and . NKp46+
, Rorc, , , and . These data show that + +
damage by neutralizing IFN mice treated with either neutralizing IFN
. In line with
protein and transcript level . We next exposed Ncr1
pathology. However, the overall pathology in Ncr1
in vivo mice in the presence or absence of Thy1+ cells.
clearly showed that the presence of Thy1+
for , , , and , as well as protein for both IL-22 and GM-CSF
Chapter 2
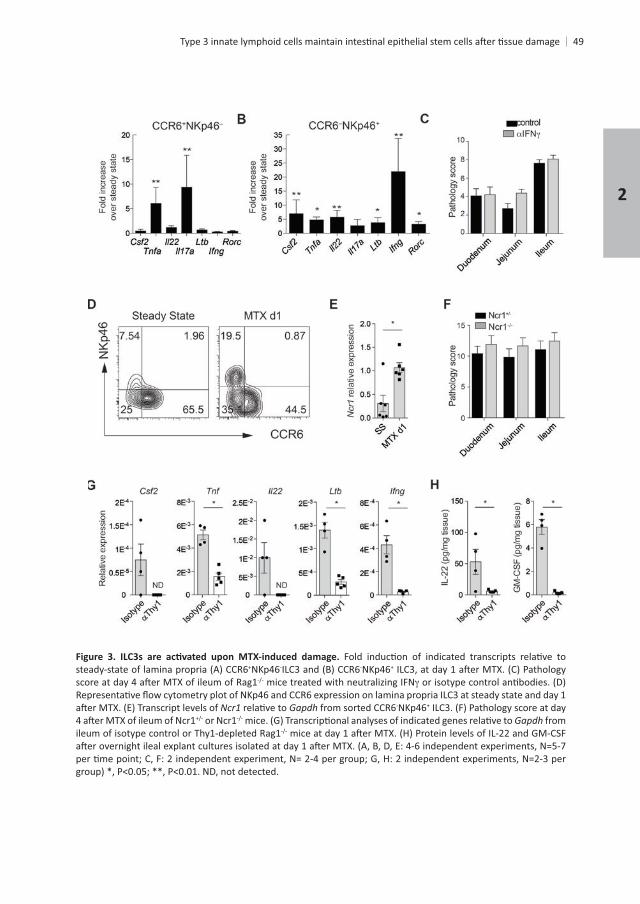
+NKp46- -NKp46+
mice treated with neutralizing IFN
Gapdh -NKp46+
or Ncr1 Gapdh from

+
tt mice showed impaired recovery of the cycling crypt
t .
Consequently, Stat3 signaling was reduced as evidenced by lower levels of transcripts
.t mice revealed
tt mice at day 4
.
t mice compared to control mice . The extent
t . Survival of epithelial cells
30 t Bax
t Bax-
absence of ILC3s.t+
Chapter 2
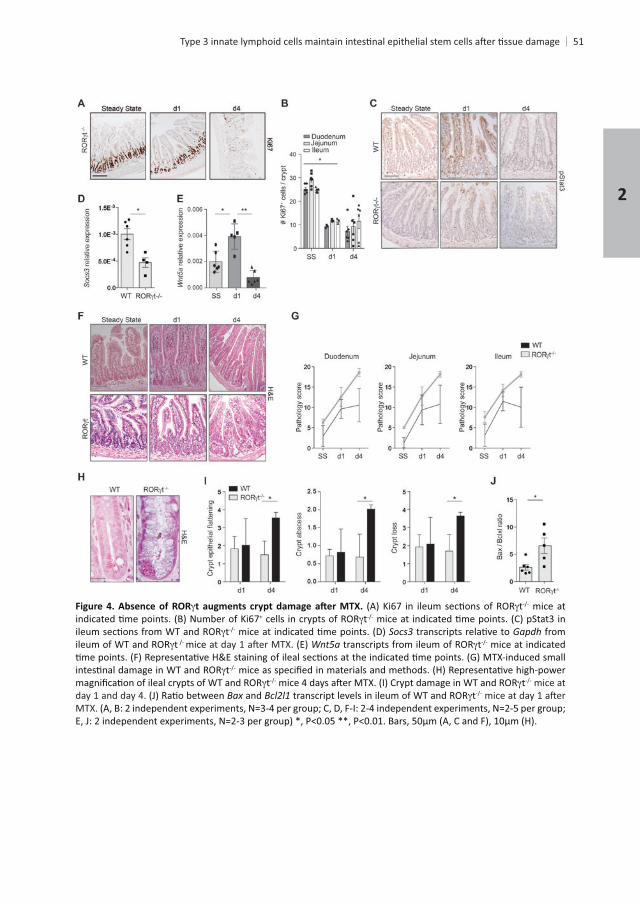
Absence of ROR t mice at + t
t Gapdh from t mice t mice at indicated
tt t mice at
day 1 and day 4. Bax and tMTX.

31. Since t
+ ISCs in response to MTX. In order to determine whether +
the percentage of GFP+
+
4 and numbers normalized by day 6 t-expressing
t+ t+
tpathology compared to WT chimeric mice as characterized by crypt
, resembling the pathology t+ lymphocytes on ISCs
+ cells were comparable between both groups
+ ISCs within the EpCAM-1+
+
+
t+ ILC3s as
epithelial cells23
. These data indicate that IL-22 is one of the factors involved in stem
+ t+
Chapter 2
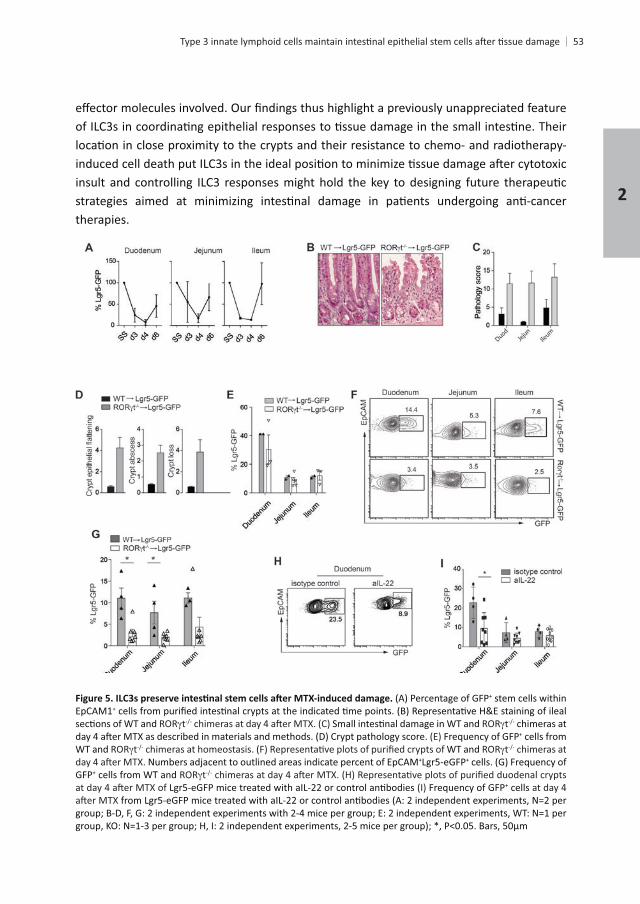
therapies.
+ stem cells within EpCAM1+
t t chimeras at Frequency of GFP+ cells from
WT and t WT and t chimeras at + +
GFP+ cells from WT and tFrequency of GFP+ cells at day 4
(A: 2 independent experiments, N=2 per

of the manuscript. Ncr1
-
Chapter 2

REFERENCES
14,
17,
17,
15,
5, 1–8
15,
10. Anderson, C. A. 43,
19,
12. Gunther, C., Neumann, H., Neurath, M. F. & Becker, C. Apoptosis, necrosis and necroptosis: cell death Gut
Cells. 11,
Nature
Nature 459,
crypt stem cell survival.
Nature 517,
20. Sawa, S., Cherrier, M., Lochner, M., Satoh-Takayama, N., Fehling, H. J., Langa, F., Di Santo, J. P. & Eberl, G.
lymphoid cells. 41,

progenitors develop.
23. Hanash, A. M. 37,
194,
28. Miyoshi, H.
Nature 494,
and disease. 17,
Chapter 2


3

GROUP 3 INNATE LYMPHOID CELLS DIRECT
INTESTINAL CRYPTS AFTER TISSUE DAMAGE
Mónica Romera-Hernández1 1 1, 1 1 1,
and Tom Cupedo1
1
2

ABSTRACT
hi
hi ISCs fail to downregulate Wnt and Notch
immune system.
60 Chapter 3

61
INTRODUCTION
1
2,3. + ISC frequencies during in vivo
4. + .
6–8
12,13
.
ILC3s. We show that IL-22 signaling is dispensable for crypt recovery but that in the absence +
3

62
MATERIALS AND METHODS
Mice t-GFP, IL-22
mice every 2 days,
i.p to WT mice at day-1 and day 0 for MTX d1 analysis and at day-1, day 0 and day 2 for MTX d4 analysis.
18.
centrifuge at 200g for 2 min to separate the crypts from single cells. Crypts were incubated
Chapter 3

63
Cells were
each gene. Principle component analysis was performed on the fragment counts using the
20.
3

64
+
least two independent analysts. Pathology was scored as previously described21
Chapter 3
+ ILC3s were sorted as indicated.
2
the expression of Gapdhmethod as 2
Primer sequences
Gene Forward Primer Reverse primer
GCCTGATGCTCTTATCTC AAGGCATAGCAGTAGGAG
CCACAAAACAGATGAAGAGAC TCTTTTGCAGCCTCTTATTC
CCATCTTCACGTAGCAGC CAAGATGTCCTGAGGGC
Gapdh TCAACGGCACAGTCAAG GCTCCACCCTTCAAGTG
. Samples were analyzed using unpaired Mann-Whitney test. p values <
3

66
RESULTS
t+ ILC3s but are not con-
tt
whereas in IL-22controls
t mice but not of IL-22.
mice during MTX treatment. In these two models,
and STAT3 signaling for stem cell maintenance + . These data show that
t+
t t , IL-22 and IL-22 mice four days
t t , IL-22 and IL-22 mice. t t , IL-22 and IL-22
+
GFP+ mice treated with
+
+
Chapter 3
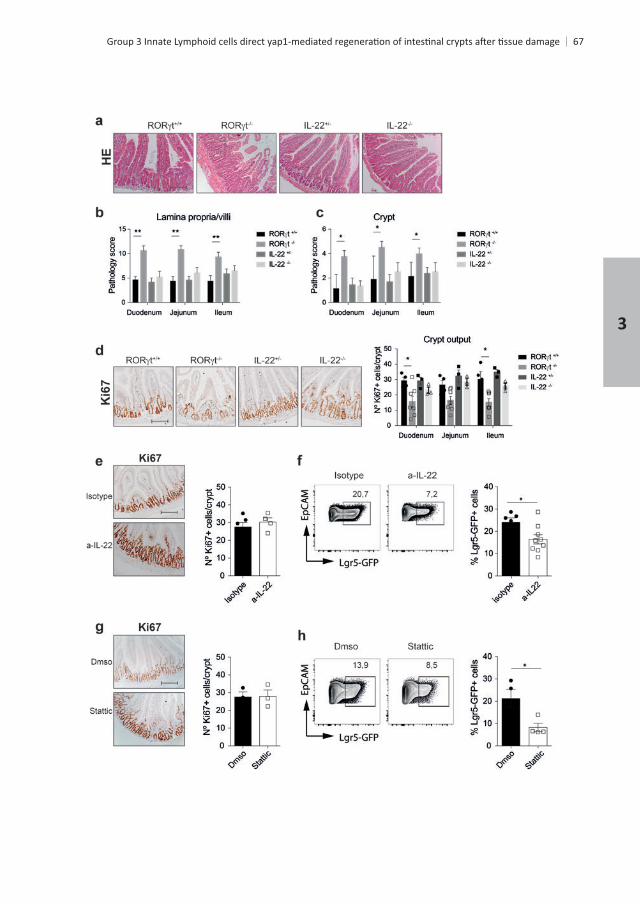
3
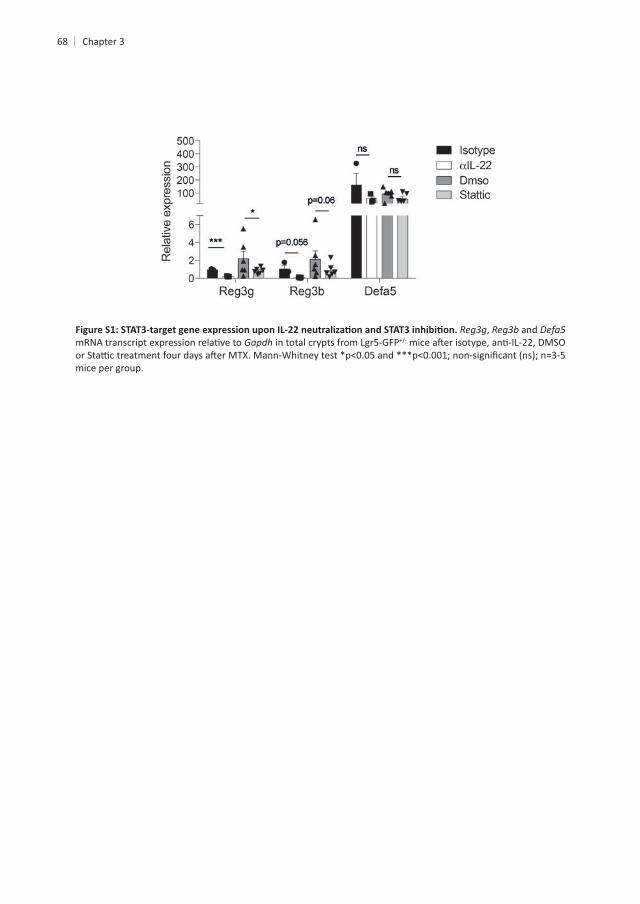
68
, and Gapdh
mice per group.
Chapter 3

hi
MTX d1, including , , Areg, and
treatment, Wnt target genes, such as , , , , , and were
for . This suggests that Yap1 suppresses Wnt signaling, in line with previous reports22. Stem cell
target gene hi
. In line with , 4 . Expression of the receptor Notch ligands and
Besides preserving stemness, Notch also determines cell fate decisions during
+ , and Kitlat MTX d1, and and Kitl further decreased at MTX d4 . Mex3a labels a
+
insults. In our analysis, hi ISCs . Finally, the expression of other
3

which ( + ISCs
and were hi
as , Sct, , , , and . Paneth cell genes, such as , , , , , , Spdef, , Kit and
and Goblet cell genes , , , , and were all upregulated
and returned to steady-state frequencies at MTX day 4, whereas Paneth cell frequencies Alpi
hi
of Bax, , and hi
+
hi
to MTX-induced damage, concomitant to a repression of the Wnt signaling pathway. The
are upregulated, coinciding with an increased frequency of enteroendocrine cells in small
Chapter 3
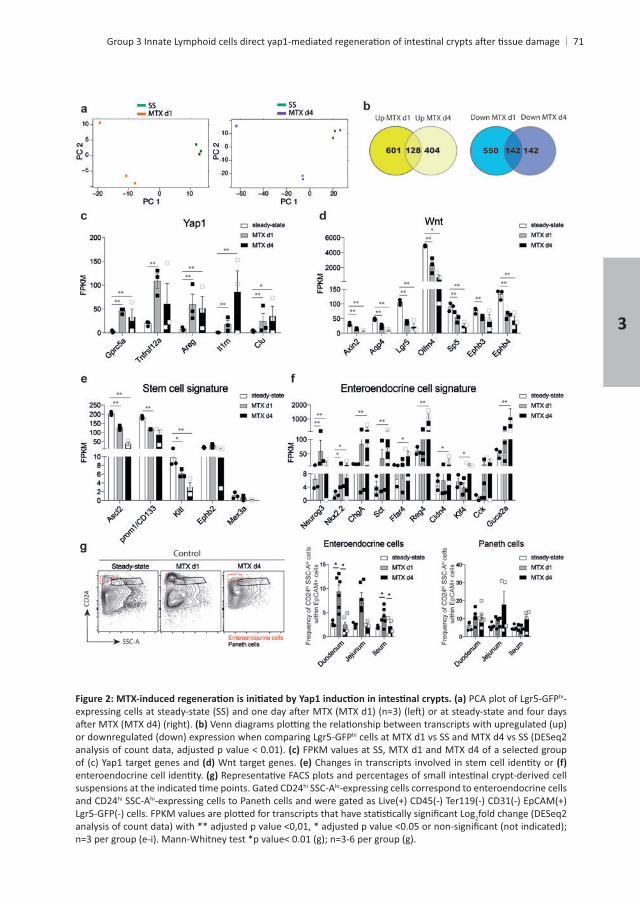
hi-
hi cells at MTX d1 vs SS and MTX d4 vs SS (DESeq2 FPKM values at SS, MTX d1 and MTX d4 of a selected group
Wnt target genes.
hi SSC-Alo-expressing cells correspond to enteroendocrine cells and CD24hi SSC-Ahi
2fold change (DESeq2
3
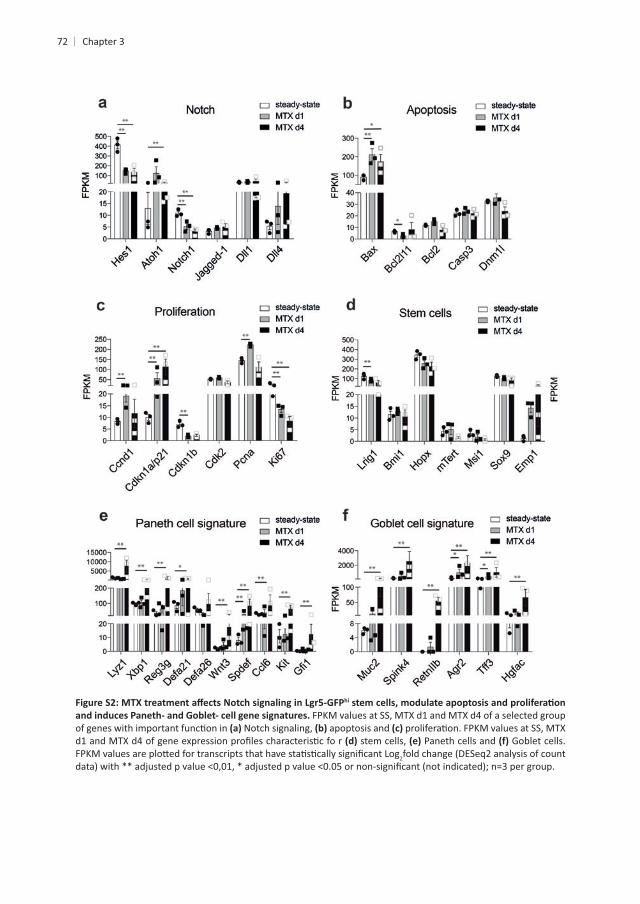
Chapter 3
hi
FPKM values at SS, MTX d1 and MTX d4 of a selected group Notch signaling, apoptosis and
stem cells, Paneth cells and Goblet cells. 2fold change (DESeq2 analysis of count

MTX of epithelial stem cells and that Yap1 is important during epithelial remodeling. To test
in vivo we blocked Yap1-
of Yap1 signaling lead to a general increase in pathology . This increased
crypts3
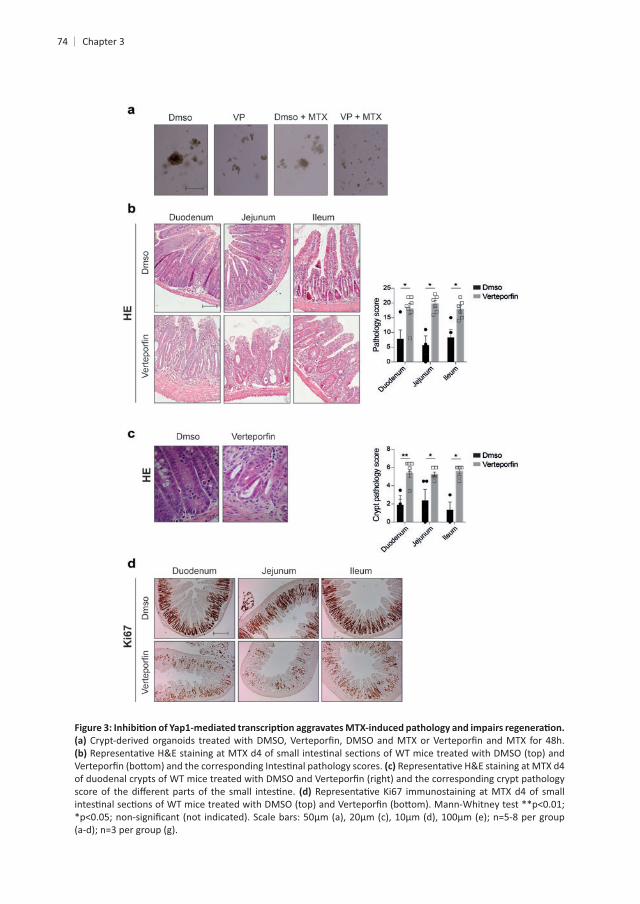
Chapter 3

dependent
thi
hi ISCs display a remarkable divergent gene expression signature in the presence or t+
hi t mice t
. Moreover, comparing the ISC transcriptome at hi ISCs showed 1421 hi t .
hi
t hi
t hi ISCs . Focusing on Yap1 signaling and
, , Areg, and were increased in controls .
hi
t mice t mice .
t t The percentage of crypts t .
hi
t+ cells. , Kitl and were not
t mice t ISCs . Finally,
genes associated with the secretory lineage, including enteroendocrine, Paneth and Goblet
3
t ISCs compared to controls and
hi t, we analyzed the frequencies of
t mice, enteroendocrine cell frequencies were not increased in response to damage .
hi
t-/-
MTX d4
Up Down Down
77 100 22
177 37
hi t t mice 2Fold change values, meaning higher gene expression levels in
2
Control t Control tDown Down Down Down
284 1 2
3
hi ISCs from t t 2Fold change values, meaning increased
2
Chapter 3
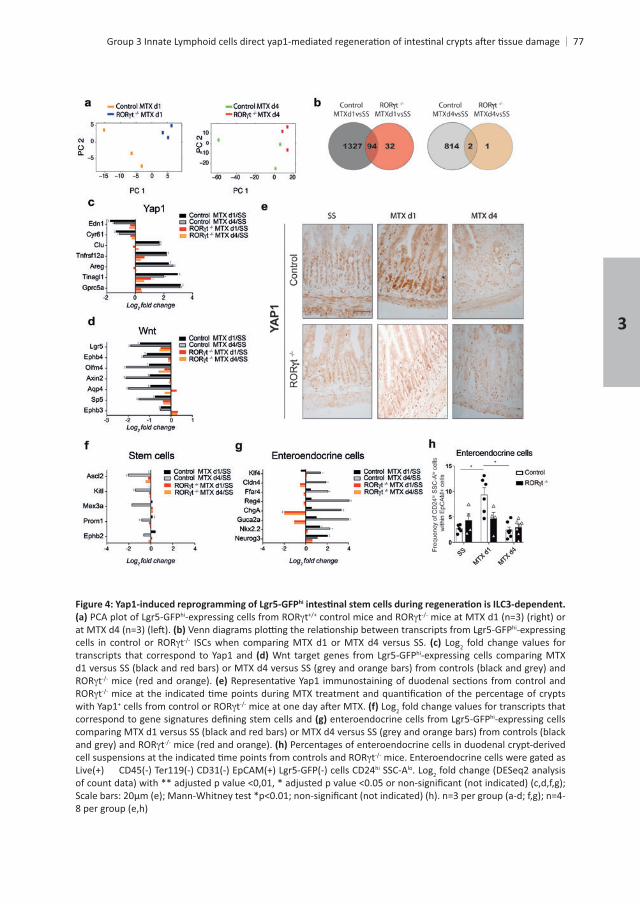
hi
hi t thi-expressing
t ISCs when comparing MTX d1 or MTX d4 versus SS. Log2 fold change values for transcripts that correspond to Yap1 and hi-expressing cells comparing MTX
tt
with Yap1+ t Log2 fold change values for transcripts that hi-expressing cells
t Percentages of enteroendocrine cells in duodenal crypt-derived t mice. Enteroendocrine cells were gated as
hi SSC-Alo. Log2 fold change (DESeq2 analysis
3
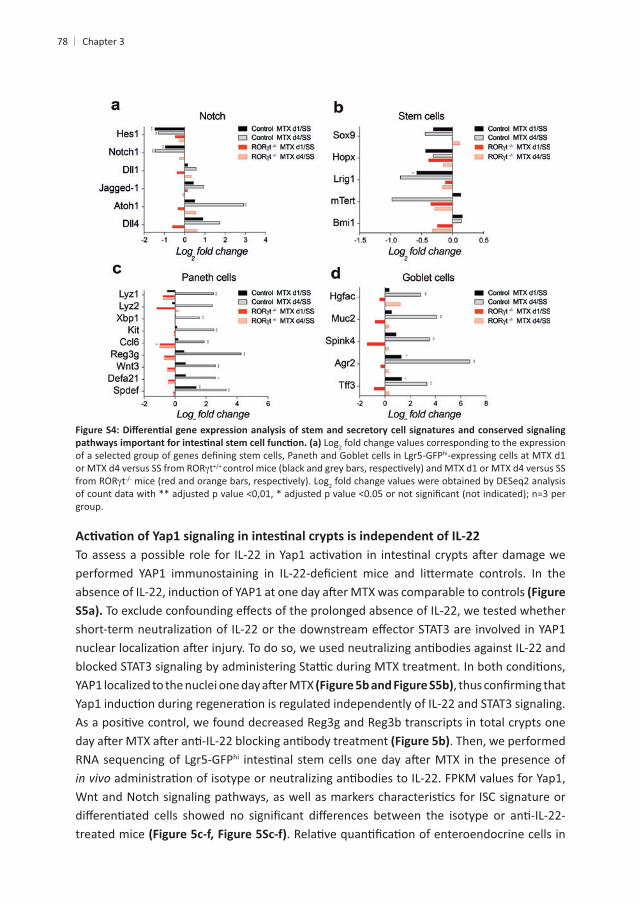
Log2 fold change values corresponding to the expression hi-expressing cells at MTX d1
tt 2 fold change values were obtained by DESeq2 analysis
group.
. Then, we performed hi
in vivo
treated mice
Chapter 3
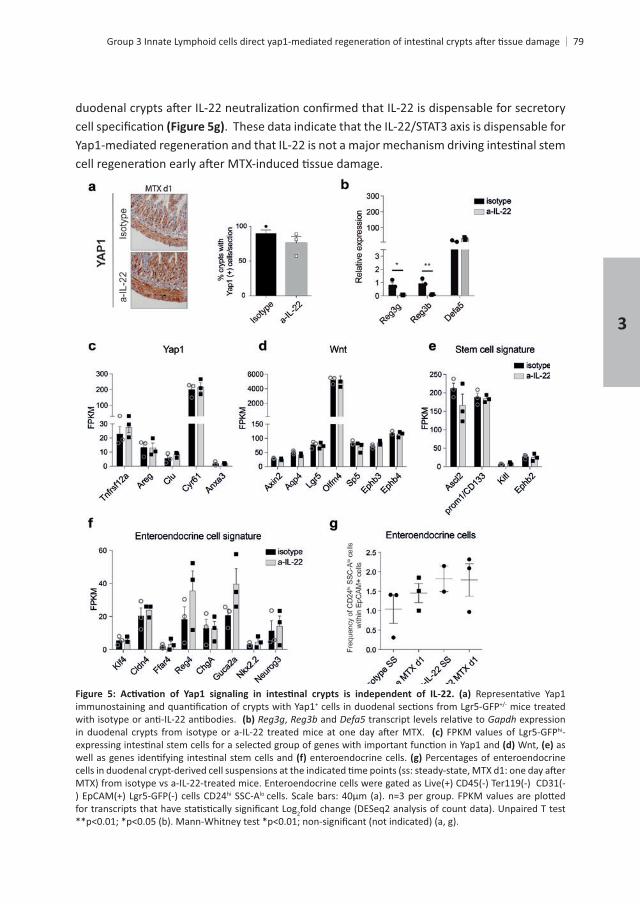
+ mice treated , and Gapdh expression
hi- Wnt, as
enteroendocrine cells. Percentages of enteroendocrine
hi SSC-Alo
2
3
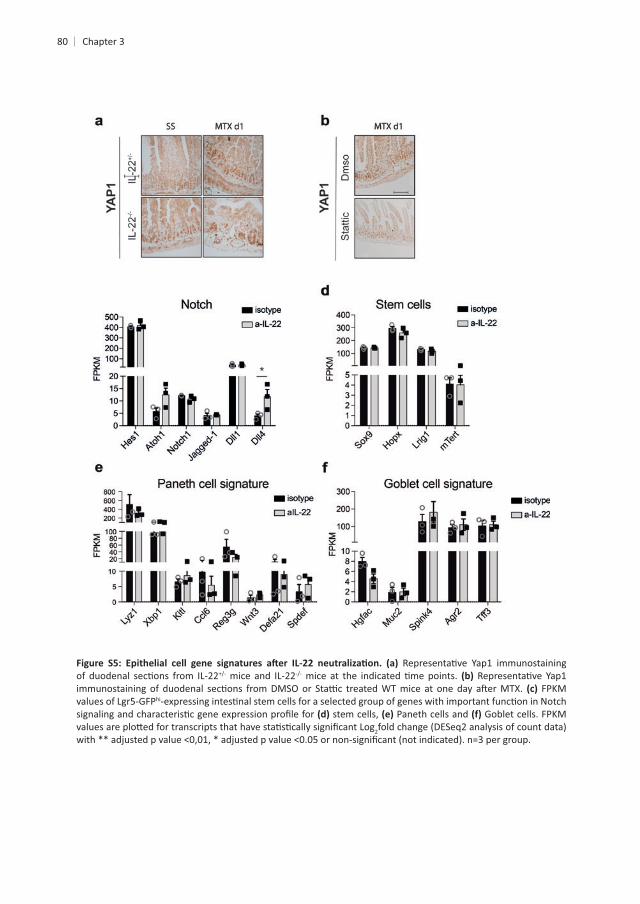
80
mice and IL-22FPKM
hi
stem cells, Paneth cells and Goblet cells. FPKM 2
Chapter 3

81
of STAT3. We set out to determine whether Src family kinase signaling is needed for Yap1
in vivo lead to increased total and crypt pathology . Moreover, we interrogated the role of
+
hi
il6ra +
+ ILC3s transcribed , , and
. Similarly, the percentage of . Based on
involvement of L-11 signaling. To test this, we made use of a small molecule inhibitor of
, although crypt pathology and
3
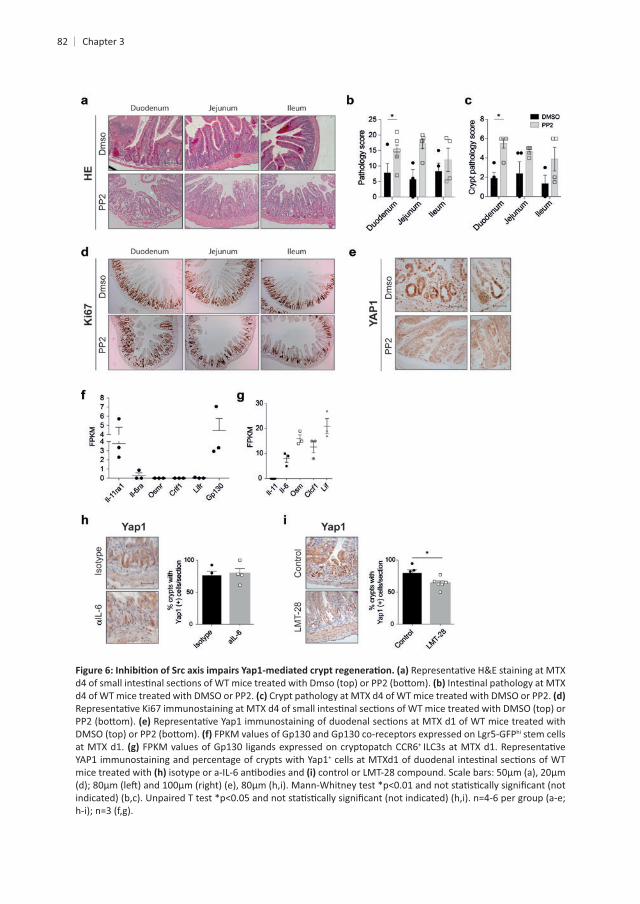
82
hi stem cells at MTX d1. FPKM +
YAP1 immunostaining and percentage of crypts with Yap1+
mice treated with
Chapter 3
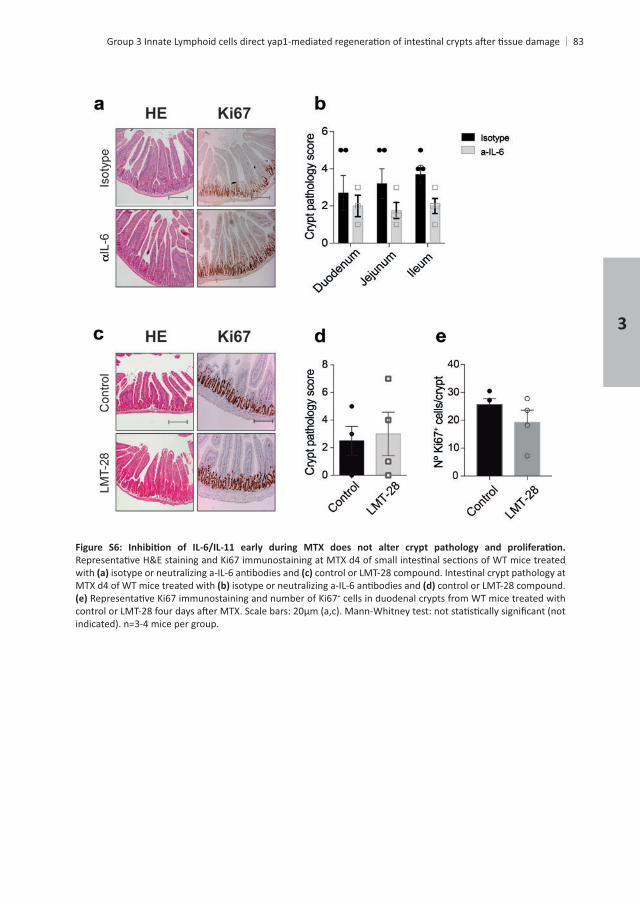
83
with MTX d4 of WT mice treated with control or LMT-28 compound.
+ cells in duodenal crypts from WT mice treated with
3

84
DISCUSSION
toxicity associated with cancer treatments.
immune cells, mesenchymal cells and the enteric nervous system, which together form the ISC niche.
+
cells23,24
+ ISC expansion .In this study, we interrogated the in vivo
+
Chapter 3

hi-expressing ISCs in the absence of ILC3s and in vivo
28
22.
30
22. Later during
and this is controlled by increasing Wnt signalling31,32.
+
+ ISCs by Yap1 depends on presence of ILC3s.
mechanism.
30. In agreement with this, our results derived from in vivo
3

86
Based on our data we propose two possible regulatory pathways by which Yap1 mediates + ISCs transcribed Gp130 and il11ra1. Conversely,
in vivofor a possible role of IL-11 in inducing Yap1, which was strengthen with the results observed
+ ISCs is controlled by innate
Chapter 3

REFERENCES
15,
endocrine cells. Endocrinology 145,
4. Aparicio-Domingo, P. damage.
Nature
6. Collu, G. M., Hidalgo-Sastre, A. & Brennan, K. Wnt-Notch signalling crosstalk in development and disease. 71,
mammals. 43,
19,
Gastroenterology 143,
17,
11. Pellegrinet, L. cells. Gastroenterology
71,
combinatorial control. 7,
Leucine-rich repeat-containing, G protein-coupled receptor 4 null mice exhibit Mol Endocrinol 2241–
Drosophila.
Gastroenterology 134,
18. Barker, N. 6,
157,
20. Subramanian, A.
3

88
21. de Koning, B. A. E.
Nature
23. Naik, S. H. tyrosine kinase 3 ligand bone marrow cultures. 174,
37,
26. Dudakov, J. A. 336,
Nature
17,
30. Taniguchi, K. Nature 519,
31. Ashton, G. H. 19,
Nature
Chapter 3


4

INTESTINAL EPITHELIAL STEM CELL HOMEOSTASIS IS ALTERED IN THE ABSENCE OF GROUP 3 INNATE
LYMPHOID CELLS
Mónica Romera-Hernández1 1 1
1 1 and Tom Cupedo1
1
2

ABSTRACT
+
induced damage, independently of IL-22. However, whether presence of ILC3s or IL-22
t but not in in IL-22
was regulated by ILC3s in an IL-22-independent fashion. Since has
+ cells with an enteroendocrine cell phenotype in the absence of ILC3s. When mice were exposed to
+
+ t+ ISC resembling
enteroendocrine cells.
Chapter 4

INTRODUCTION
and consequently shape microbial diversity1
2.
3,4.
+
. ILC3s are an important source of IL-226, which acts mainly on epithelial cells and mesenchymal stromal
of epithelial cells+
10
11. Finally, IL-22 also can act on + 12.
3,13. They are characterized
+ ISCs vulnerable to genotoxic damage14
4

+
such as Bmi1, mTert, Lrig1 or Hopx16–20 + CBCs are completely dispensable
and maintenance of normal crypt-villus architecture 21,22. However, because Hopx and Bmi1
stem cell type remains unknown16–18,21,23
secretory progenitor features.
24. Similarly, recent data suggest that subsets of secretory enteroendocrine +
.
in vivo.
crypt loss. We demonstrate that ILC3s instruct crypt epithelial cells via IL-22 dependent and
hi lo
lo
Chapter 4

MATERIALS AND METHODS
Mice t-GFP, IL-22 t-GFP
were used whenever possible.
Methotrexate
mice every 2 days,
centrifuge at 200g for 2 min to separate the crypts from single cells. Crypts were incubated
and counted by using a microscope.
4

genes were performed with DESeq2
each gene. Principle component analysis was performed on the fragment counts using the
30.
performed blinded by at least two independent analysts. Pathology was scored as previously described31
Chapter 4
2
the expression of Gapdhmethod as 2
Gene Reverse Primer
CCATCTTCACGTAGCAGC CAAGATGTCCTGAGGGC
GGAAGCCAGTGTCATATCA CCTTGGGGTTCATCTCA
GATGGCTACCGTGGTGT CACCCATGCTCGAATG
GGGGGTGTGAGAATGTCT AGGGCCTTCAGGTCTTC
GCGAAGGGCAAGAATAA TCGGGTCTGTGCTGAG
GCACAACGCACTTTCTTT GGTGGCCCTCAGATGT
TGGACGCTTTGCACTT AGTGGGGGGAAAACTCT
GGAGGACGATGTTCAGATAA CGGCACAGGTAAGAGTTG
Gapdh TCAACGGCACAGTCAAG GCTCCACCCTTCAAGTG
4

RESULTS
ROR
degree of bleeding. It also comprised several parameters related to the appearance of the stem cell-containing crypt compartment including degree of crypt loss, crypt abscesses and
stress related transcripts and
controls mice were similar , showing that ILC3 absence does not alter steady-state crypt output. Finally, we examined the in vitro ex vivo isolated
the average number of mature organoids that grew per well . Together these
Chapter 4
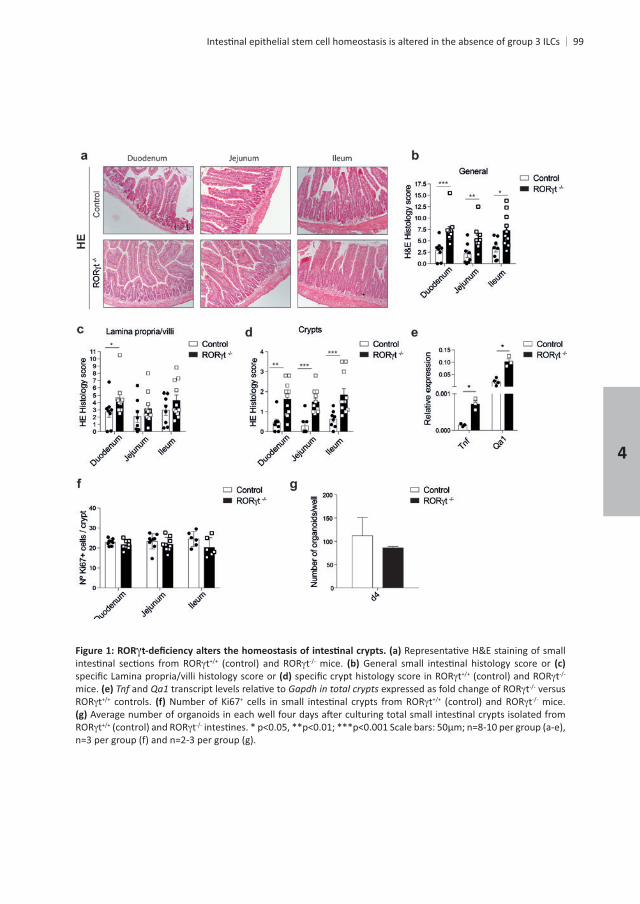
t t mice. t t
mice. and Gapdh in total crypts t versus t controls. + t t mice.
t t
4
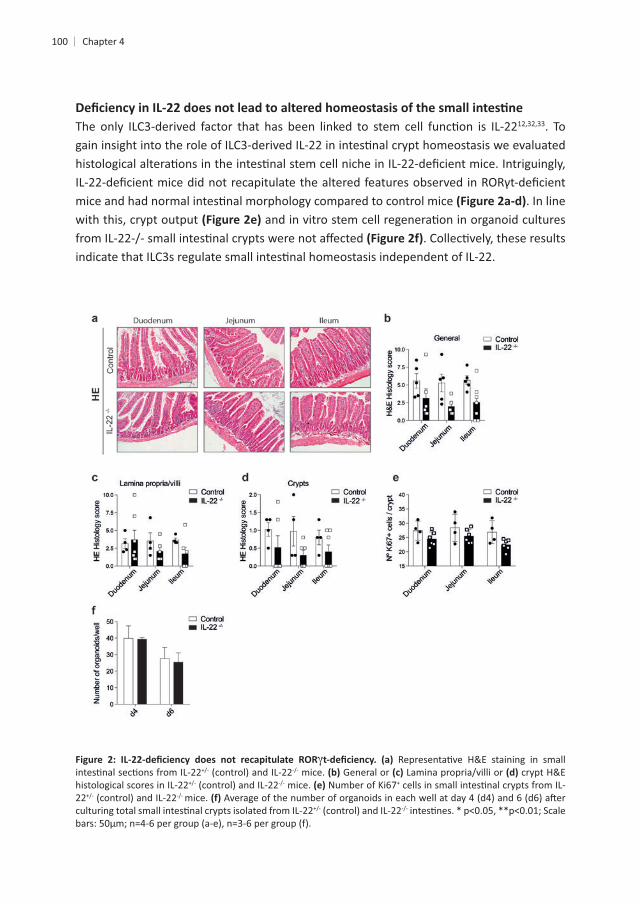
100 Chapter 4
12,32,33. To
. In line with this, crypt output
mice. General or crypt H&E histological scores in IL-22 mice. +
22 mice.

+
enterocytes are derived from crypt progenitors, we tested whether the presence or absence
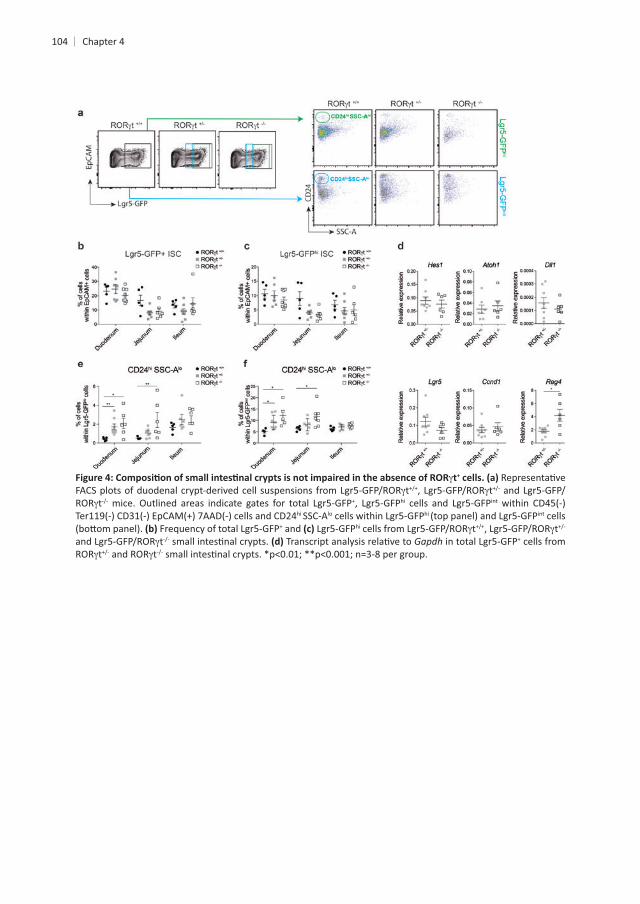
+ ISCs express intermediate levels of the cell adhesion molecule CD24 . Hence, we used intermediate CD24 levels (CD24med + ISCs in wild-type B6
t and IL-22hi SSC-Alo hi SSC-Ahi
these three main cellular subsets in absence of ILC3s or IL-22 . Next, we tested + cells at steady-state was controlled by IL-22. To assess
+ ISCs and of the other cell subsets present in
. Even though the frequency of PC was not altered by the absence
t or IL-22 mice. As expected, t and IL-22 PC when compared to WT controls . This is in line with
epithelial cells. In contrast, transcript levels of a marker of both PC and EE cells34 as t mice but not in IL-22 PC
Next, we set out to determine changes in ISC markers and genes important for ISC
med
and the Notch-target gene The Notch-target gene transcribed in CD24med
Notch signaling within crypt epithelial cells enriched for ISCs.
101
4
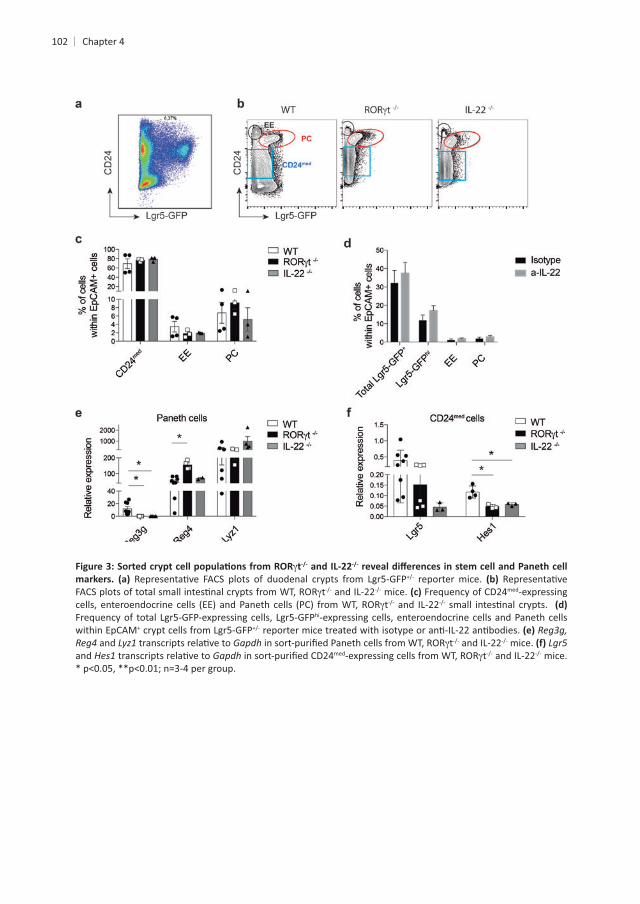
102
t-/- -/-
reporter mice. t and IL-22 mice. Frequency of CD24med-expressing
t and IL-22 hi-expressing cells, enteroendocrine cells and Paneth cells
within EpCAM+
and Gapdh t and IL-22 mice. and Gapdh med t and IL-22 mice.
Chapter 4

103
+ hi SSC-A
+
t + hi
int cells within total EpCAM-1+
. In line with published reports,
expressing cells t + hi t t mice
t t mice. Genes involved in Notch signaling, such as , and +
t t mice . transcript levels seemed slightly +
+ ISCs expressed similar levels of the regulator of cyclin-dependent kinase, Together these analyses revealed
+
PC, + t t mice
lineage cells (CD24hi SSC-Alo hi int
cytometry t duodenal crypts contained increased frequencies of CD24hi SSC-Alo cells that co-expressed GFP This was the
or intermediate levels . + ISC pool in the absence of
with features of both enteroendocrine cells (CD24hi SSC-Alo +
4
104
t+
t tt + hi int
hi SSC-Alo hi int cells + and hi t t
t Gapdh + cells from t t
Chapter 4

hi
t mice, + t
int
hi cells. In order to gather evidence for either of these hi t t duodenal crypts by
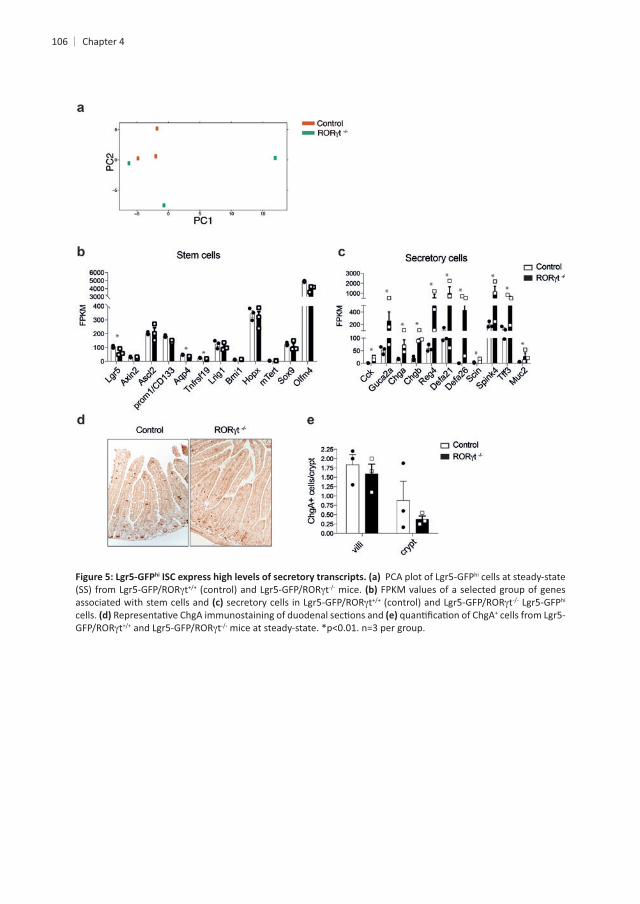
hi ISCs and in line with this high degree of similarity, PCA revealed a poor hi hi t ISCs
hi t crypts when compared to control cells. First, we established the expression of genes involved in controlling stemness, to determine
cells. GFPhi , and
of ILC3s. This indicates that the GFPhi tcell genes. Subsequently we focused on genes associated with mature EE cells, in support
hi cells were enriched for EE cell-associated genes, such as , , , , , Scin,
, and . The increase in EE genes was not due to altered EE cell
enteroendocrine cells in villi or crypts in the absence of ILC3s GPFhi ISCs in the absence of ILC3s co-express secretory genes and stem cell genes, which
int EE precursors.
4
106
hi hi cells at steady-state t t mice. FPKM values of a selected group of genes
associated with stem cells and t t hi cells. +
t t
Chapter 4
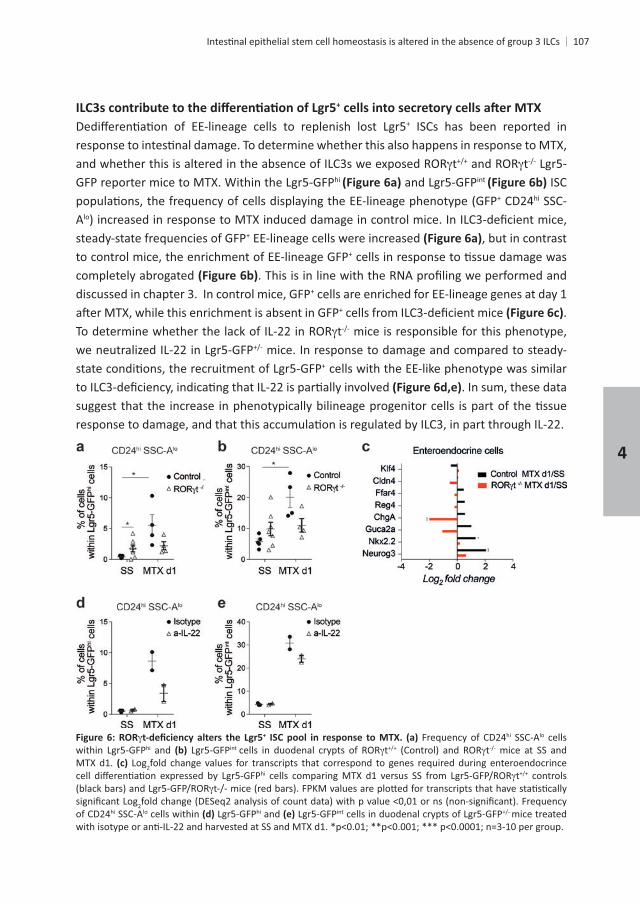
+
+ ISCs has been reported in
t thi int ISC
+ CD24hi SSC-Alo
steady-state frequencies of GFP+ EE-lineage cells were increased , but in contrast to control mice, the enrichment of EE-lineage GFP+
completely abrogated discussed in chapter 3. In control mice, GFP+ cells are enriched for EE-lineage genes at day 1
+ . t mice is responsible for this phenotype,
mice. In response to damage and compared to steady-+ cells with the EE-like phenotype was similar
. In sum, these data
+ Frequency of CD24hi SSC-Alo cells hi and int t t mice at SS and
MTX d1. Log2fold change values for transcripts that correspond to genes required during enteroendocrince hi t controls
2 Frequency of CD24hi SSC-Alo cells within hi and int mice treated
4

108
DISCUSSION
stem cells. ILC3s contribute to epithelial renewal in response to damage12,32,33. The cellular
expressing crypt epithelial stem by ILC3s. The reduced levels of
36
of CD24hi SSC-Alo hi int
GFPhi tcells (
, and but similar levels of , , and , among others hi ISCs and mature epithelial cell types,
t under
hi ISC pool to maintain normal crypt output, raising the possibility that survival or hi
possibility, double-immunostaining for secretory and stem cell markers, such as ChgA and
hi ex vivo
for maintenance of crypt homeostasis. IL-22 Paneth cells had no detectable transcript levels of
t and IL-22 mice are explained by
t
Chapter 4

and + + cells in the
in vivo.hi
enteroendocrine cells was increased. These cells acquired secretory cell transcripts and maintained the expression of some stem cell genes. All together this suggests that subsets
int hi
This study contributes to current views on how innate lymphoid cells regulate crypt
+
4

110
REFERENCES
14,
36,
3. Barker, N. Nature 449,
4. Sato, T.
Seminars in
14,
10. Sato, T. Nature 469,
11. Turner, J. E., Stockinger, B. & Helmby, H. IL-22 Mediates Goblet Cell Hyperplasia and Worm Expulsion in 9,
12. Lindemans, C. A. Nature
markers. 31,
14. Tao, S. DNA damage. 34,
14,
tumor suppressor. 149,
Science 334,
Gastroenterology 151,
Chapter 4

111
Nature
14,
Lineage Daughters.
Am J
157,
30. Subramanian, A.
31. de Koning, B. A. E.
32. Hanash, A. M. 37,
33. Aparicio-Domingo, P. damage.
34. Sasaki, N.
Nature
36. Jensen, J. Control of endodermal endocrine development by Hes-1.
4

5

DENDRITIC CELLS ASSOCIATE WITH GROUP 3 INNATE LYMPHOID CELLS TO CONTROL INTESTINAL REGENERATION
Mónica Romera-Hernández1 1
1 1, Louis Boon 3 and Tom Cupedo1
1
2
3

114
ABSTRACT
+
CD11c+
that SILTs act as innate control centers where ILC3 and DC interact and translate environmental
+
+ cells contribute to epithelial responses
remain elusive. Next,
+ MHCII+ CD11b- CD103- Plet1+ cells, expressing high levels of and
with ILC3s and able to regulate ILC3. It suggests that SILT-DCs can sense epithelial damage

INTRODUCTION
The mucosal immune system needs to protect the single layer of epithelial cells which is
1,2. DCs induce
the murine small bowel3,4 +CD11b+
. Single CD103+CD11b- cells (CD24+ + CD8 + +
Th1 responses . Another subset of DCs, the CD103-CD11b+ cells are able of inducing both 4 -CD103- cells is a largely
t-
and
by ILC3s is regulated by LT + DCs11
of ILC3 subsets. For instance, DCs expressing CXCL16 retain NKp46+ ILC3 in the LP due to 12,13
.
structures are developmentally dependent on ILC3s and on LT -LT 18
LT and LT mice.
5

116
+ ILC3, DCs and non-
+ ILC3 in SILTs in

MATERIALS AND METHODS
Mice t-GFP and hCD2-GFPmice were bred in the animal facility
mice were bred at Icahn School of Medicine at Mount Sinai Hess Center for Science and Medicine
mice. When DT was combined with MTX, DT
as well as in mesenteric lymph nodes.
from hCD2-GFP
in vitro Spleen were excised into small
pressed through the strainer using the plunger end of a syringe. Cells were washed with red
+ cells were
Cells were cultured at 0.3x106
14
5

118
in vitro 6 splenic DCs or BMDC were cultured
Cells were

genes were performed with DESeq2
each gene. Principle component analysis was performed on the fragment counts using the
20.
+
m cell strainers previous
for ILC3s.
5
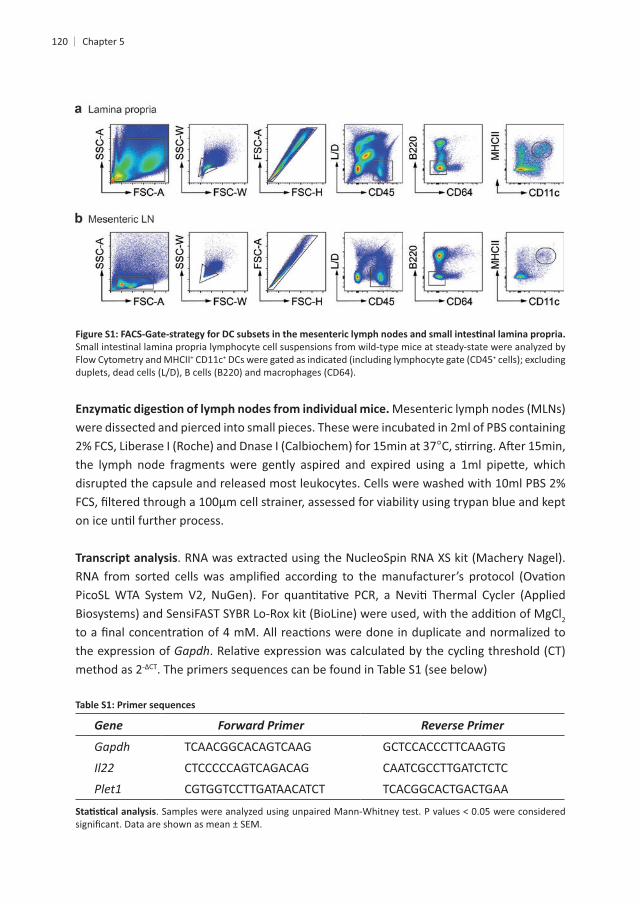
120
Flow Cytometry and MHCII+ CD11c+ +
were dissected and pierced into small pieces. These were incubated in 2ml of PBS containing
2
the expression of Gapdhmethod as 2
Gene Forward Primer Reverse Primer
Gapdh TCAACGGCACAGTCAAG GCTCCACCCTTCAAGTG
CTCCCCCAGTCAGACAG CAATCGCCTTGATCTCTC
CGTGGTCCTTGATAACATCT TCACGGCACTGACTGAA

121
RESULTS
CD11c+
induced stem cell damage. We hypothesize that the CD11c+ CP-DCs are involved in this +
all CD11c+
+ cells in the lamina
+ cells in both the LP and MLNs + cells slightly increased,
+
being recruited from bone marrow-derived progenitors. This strategy allowed us to further +
treatment, hypothesizing that CD11c+ cells and ILC3s in cryptopatches are required to
with MTX in mice lacking CD11c+
+ cells per crypt. In the absence of CD11c+
This +
We assessed whether CD11c+ + mice treated with DT or PBS at steady-state. First, we determined +ILC3s within the LP of the SI and found to be unchanged
. Second, we analyzed the level of expression of the signature cytokine IL-22. CD11c+ cell +
that CD11c+
+
5
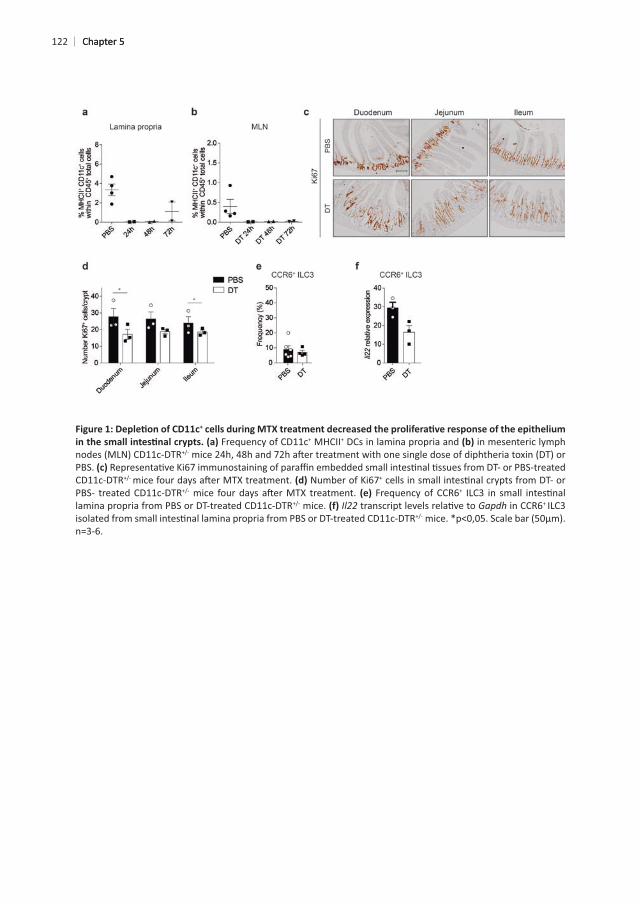
122
+
Frequency of CD11c+ MHCII+ DCs in lamina propria and in mesenteric lymph
PBS. +
+
mice. Gapdh + ILC3
n=3-6.

123
CD11c+ - -
+ cells that lacked CD103 but expressed Plet1 . We have previously + CD11b+
- Plet1+
t mice. This revealed that CD103- Plet1+ t mice
SILT-DCs. Next, we made use of hCD2-GFP+ reporter mice in which SILTs can be micro dissected using a stereomicroscope due to high expression of hCD2-GFP in ILC3 and B cells. Comparing cell suspensions derived from micro dissected GFP+ SILT regions or GFP- non-SILT lamina propria regions showed that CD103- Plet1+ DCs were only present in GFP+
and absent from non-SILT lamina propria + Plet1- phenotype to isolate CP-DCs and perform phenotypic
macrophages and B cells, and gated as CD11c+ MHCII+
+ DCs are CD11b+CD103+ there is another small subset of CD11b-CD103-Plet1+ DCs
Plet1 . CD103- Plet1+ cells were further examined for the expression of CD11b, CD8
t mice showed decreased frequencies of
+ CD11b- CD103- Plet1+ DCs as SILT-resident cells. 5
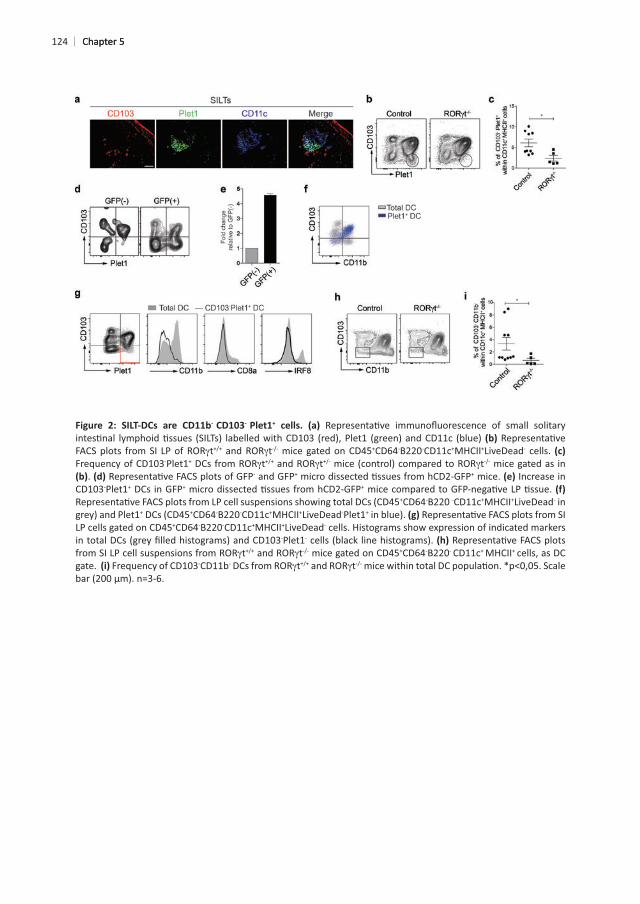
124
- - +
t t +CD64-B220-CD11c+MHCII+LiveDead- cells. Frequency of CD103-Plet1+ t t t mice gated as in
. of GFP- and GFP+ + mice. Increase in CD103-Plet1+ DCs in GFP+ +
+CD64-B220 -CD11c+MHCII+LiveDead- in + +CD64-B220-CD11c+MHCII+LiveDead-Plet1+
+CD64-B220-CD11c+MHCII+LiveDead- cells. Histograms show expression of indicated markers -Plet1-
t t +CD64-B220- CD11c+ MHCII+ cells, as DC gate. Frequency of CD103-CD11b- t t
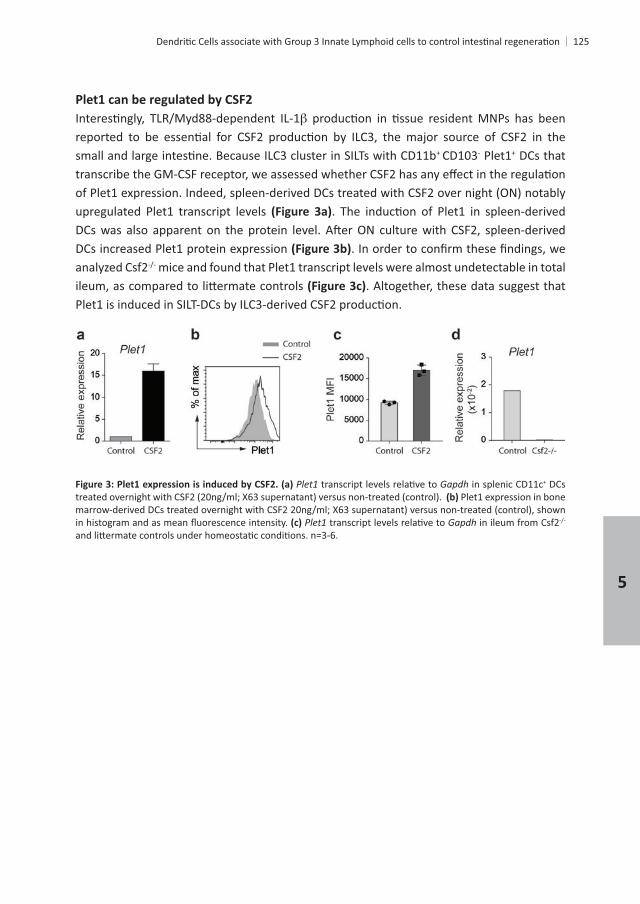
+ CD103- Plet1+ DCs that
upregulated Plet1 transcript levels
DCs increased Plet1 protein expression analyzed Csf2 mice and found that Plet1 transcript levels were almost undetectable in total
. Altogether, these data suggest that
Gapdh in splenic CD11c+ DCs Plet1 expression in bone
Gapdh in ileum from Csf2
5

126
- Plet1+ DC and compared their transcriptome with that of CD11b+ CD103+
regulated in SILT-DCs when compared with CD11b+ CD103+
were found to be down-regulated . Despite the fact that CD11b- CD103- cells
+ DCs are contained within CD11b- CD103-
DCs express lower levels of and + DC development . expression was similar between the two DC subset analyzed. However, SILT-
DCs expressed higher levels of and , which have also been associated with CD11b+
CD103+ and CD11b- CD103+ . Likewise, SILT-DCs expressed and enriched in SILT-DCs, such as and for and +
CD11b- CD8 +
, and . Furthermore, SILT-DCs were enriched for , , , and , whereas they lacked and
. SILT-DCs also expressed high levels of , which indicates that SILT-DCs might be able to migrate to gut-draining lymph nodes. Nonetheless, CD80, CD86 and CD40 were transcribed at similar levels in SILT-DCs and CD11b+ CD103+
. Strikingly, SILT-DCs were enriched for transcripts encoding and ,
DC were and , while and
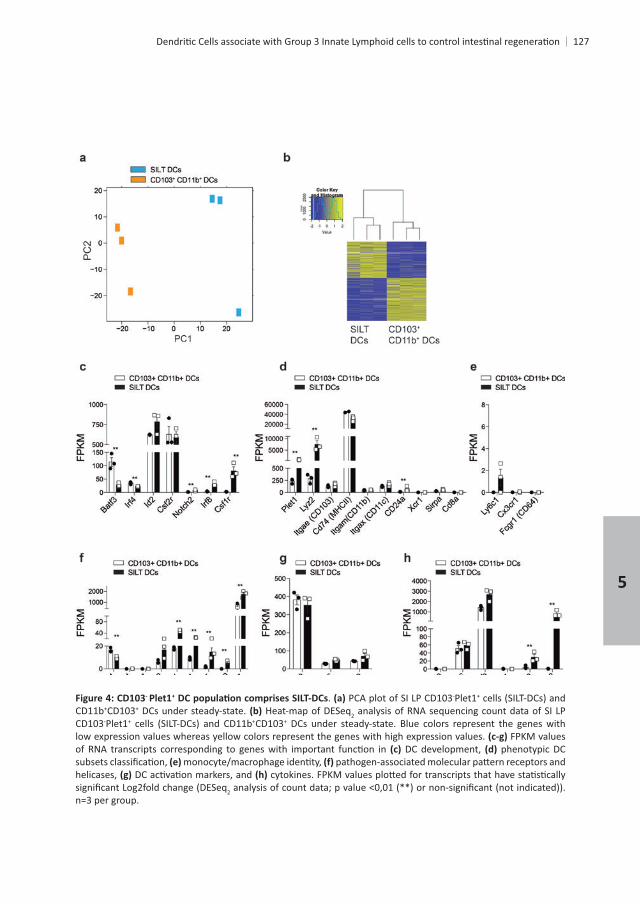
- + . PCA plot of SI LP CD103-Plet1+
CD11b+CD103+ DCs under steady-state. Heat-map of DESeq2CD103-Plet1+ +CD103+ DCs under steady-state. Blue colors represent the genes with low expression values whereas yellow colors represent the genes with high expression values. FPKM values
DC development, phenotypic DC
helicases, 2
n=3 per group.
5

128
Because CD11c+
MTX-induced stem cell damage
, and , were increased in response to MTX, and the immature DC marker was downregulated
. ,
and , we hypothesized that SILT-DCs might emigrate
CD103- Plet1+ DCs within CD11c+ MHCII+
. Further experiments are in progress to fully demonstrate that SILT-DCs are able to migrate
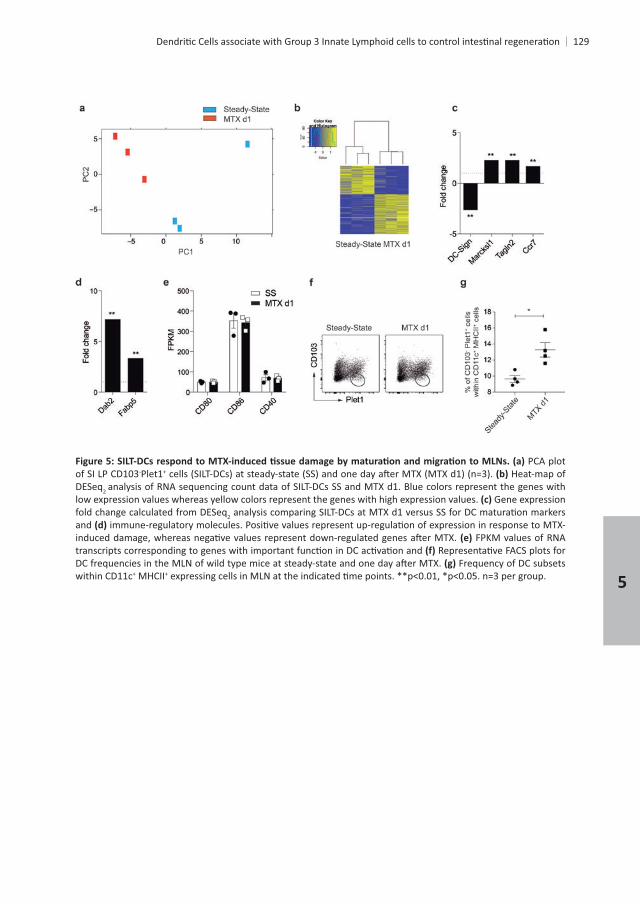
PCA plot of SI LP CD103-Plet1+ Heat-map of DESeq2 low expression values whereas yellow colors represent the genes with high expression values. Gene expression fold change calculated from DESeq2and
Frequency of DC subsets within CD11c+ MHCII+
5

130
DISCUSSION
+
+
21. To determine whether similar mechanisms
CD103- CD11b-
whether Plet1+
CD103- CD11b- t mice suggests that SILT-DCs are only present when their cryptopatches are formed.
+ CD103- CD11c+
, and , and showed that DC-expressing Plet1 express Csf2-producing ILC3s, likely occurring in cryptopatches.
order to fully elucidate whether SILT-DCs are DC precursors or an intermediate state during

131
22. There is
MLN to induce tolerance - CD11b-
for these diseases.
5

132
REFERENCES
innate immune system.
3. Persson, E.
6,
Cell Homeostasis. 44,
Provide Innate Mucosal Immune Defense.
lymphoid cells. 41,
mucosal draining lymph nodes. 6,
Science
5,
lymphotoxin alpha and the lymphotoxin beta receptor. 173,
157,

133
20. Subramanian, A.
6,
Bacterial Flagellin Enhances Mucosal Innate Immune Defense. 36,
programs. Nature 539,
lymph nodes. 191,
191,
5

6

GENERAL DISCUSSION


SUMMARY
The overall goal of this thesis was to underpin the role of Group 3 Innate Lymphoid cells
+
+
+ ISCs, but
+ ISCs from
+ ISCs likely occurs indirectly via another cell type. We propose that
t
+ ISCs expressing enteroendocrine-like markers, closely resembling a reserve ISC
+
+
+ cells co-localizing with ILC3s in solitary lymphoid follicles and found that they transcribe high levels of
, and
General discussion
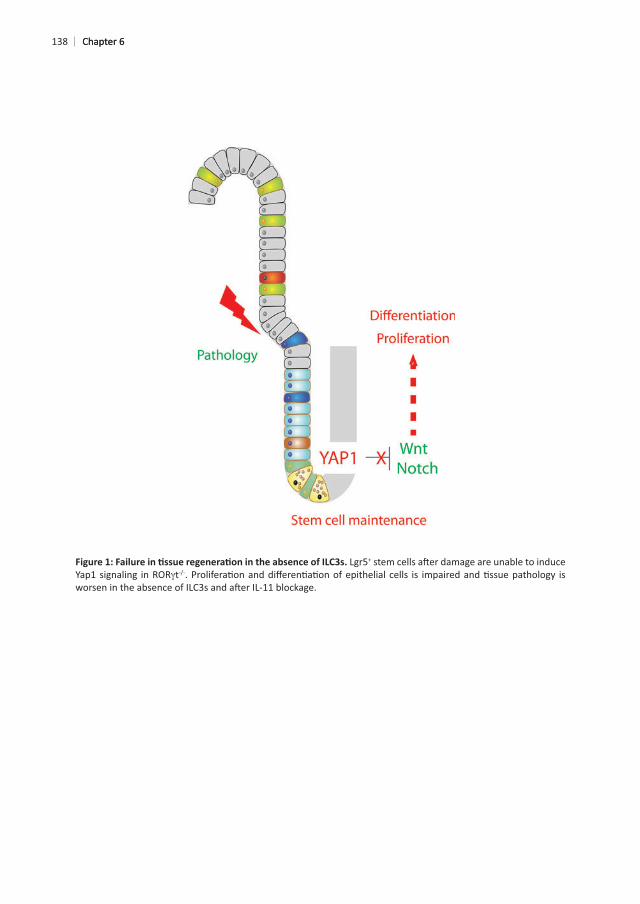
138
+
t
Chapter 6Chapter 6
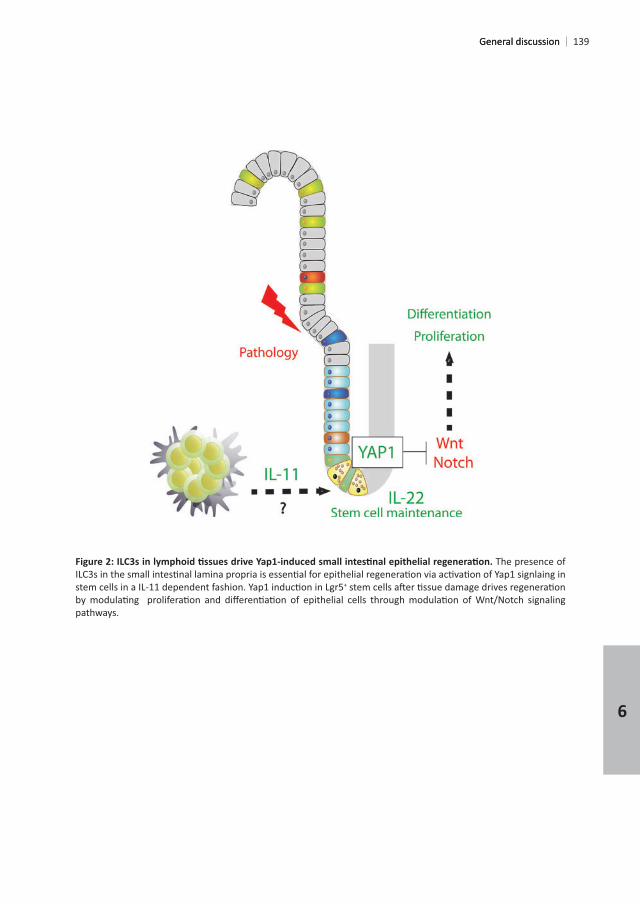
The presence of
+
pathways.
General discussionGeneral discussion
6

140
factors. In the gut mucosa, immune cells reinforce the epithelial barrier and induce tolerance against food or commensal bacteria. Among these immune cells, a recently discovered type of lymphocytes, innate lymphoid cells or ILCs, orchestrates immune responses and maintains
1–3
three types, ILC1s, ILC2s and ILC3s4
IL-22 2-LT 6,8.
.ILC3s also interact with macrophages to establish tolerance towards the commensal
in LysM+
10
towards the commensal microbiota11–14. ILC2s contribute to responses against helminths.
cell death. Epithelial cells sense the danger signals released by the dying cells and produce
18
. However, ILCs can
and ILC3s, which can acquire the ability to make IFN
contribute to gut pathology20,21
from maintenance of barrier, tolerance against commensals and immune responses against
Chapter 6

141
appreciated. ILC3s contribute to skin wound healing22
insult23 24. In this thesis, we
cells, which are crucial for epithelial self-renewal. As a result, epithelial damage is a dose-
lamina propria NKp46+ +
cells , enteric neurons28 and group 3 innate lymphoid cells
+
+
+ CBCs. The cells that + cells, Prox1+
30–34. The actual
General discussion
6

142
+
24.
important results are discussed below. In general, our work provides novel insights into the
vs
. High-dose
In a series of experiments, we demonstrate that ILC3s are necessary to induce a number of
t to demonstrate a role for ILC3s in
wound healingthe peak of damage occurs, which indicates that epithelial responses are induced very
signaling remain to be determined. First, it would be necessary to assess the phenotype of the pSTAT3+
36
enteroendocrine cells (Claudin-4
associated with survival and apoptosis should be assessed. Finally, DNA damage responses
Chapter 6

143
IL-22 induces pSTAT3, as we found that NKp46+
IL-22 neutralized mice vs isotype-treated mice as well as in IL-22
38
+ CBCs mediates stem cell
40.
t
Wnt, Notch, BMP, EGF. During homeostasis, in vitro
General discussion
6

144
t but not IL-22
+ cells leads to a decrease in crypt output, +
t + cell subset that we think interact with ILC3s, therefore it is not a good model to address this. Co-cultures with CD11c+
+ CBCs
+
+ CBCs caused by blocking IL-22 or STAT3 signaling does not translate in reduced
+ CBCs, as supported by previous published reports30,41–43. It might also be possible that de novo +
+
+ CBC maintenance (Chapter 2 and
+
of the stem cell compartment in in vitroIL-22 . However, blocking cytokine signaling or administering recombinant cytokines in vivo or in vitro
+ CBCs.
+ CBCs + CBCs downregulate
Chapter 6

+
damage44 +
reduced, Yap1 is not completely absent in mice lacking ILC3s.
agreement with previous reports that highlighted the importance of Yap1 in crypt epithelial 44–46
epithelial Yap1+ cells in vivo cannot be ruled out.
+ CBCs
important to unequivocally show that Yap1 signaling is regulated in an IL22-independent
. This could
+ CBCs but not
results48.
+ CBCs expressed high levels of transcripts encoding and , and low levels of . Because cryptopatch ILC3s expressed but not , we blocked IL-6 during
General discussion
6

146
IL-6 alone, Yap1+
then be via an indirect mechanism through mesenchymal or epithelial cell-derived IL-11. It t
.
, and
and in response to damage and whether they migrate to lymph nodes to interact with lymph
propose that cryptopatches have emerged in mammals in order to regulate the magnitude
+
Chapter 6

+ .
+ stromal
whether cryptopatches are sites from where epithelial responses are orchestrated.
. Also,
, skin or hair follicles , corneal epithelium60 and lung pulmonary alveoli and bronchi61 + stem cells and can contain ILC3s
6
General discussion

148
necessary. In parallel, in vitro and in vivo experiments could help to elucidate the signals
molecules are produced and signal through membrane-bound receptors. Finally, the candidate molecules, will be also need to be tested both in vitro and in vivo.
Chapter 6

REFERENCES
4. Mortha, A., Chudnovskiy, A., Hashimoto, D., Bogunovic, M., Spencer, S. P., Belkaid, Y. & Merad, M. Microbiota- 343,
Elife 5,
+ T cells. TL - 348. Science
Innate lymphoid cells--a proposal for uniform nomenclature. 13,
8. Balzola, F., Bernstein, C., Ho, G. T. & Lees, C. Innate lymphoid cells drive interleukin-23-dependent innate 11,
Mucosal
14,
Bacterial Flagellin Enhances Mucosal Innate Immune Defense. 36,
13. Macho-Fernandez, E., Koroleva, E. P., Spencer, C. M., Tighe, M., Torrado, E., Cooper, A. M., Fu, Y.-X. &
by epithelial cells.
14. Atarashi, K. 331,
Science 341,
16. Furusawa, Y. cells. Nature
Metabolites produced by commensal bacteria promote peripheral regulatory T-cell Nature
Nature 464,
General discussion
6

Nature 463,
MHCII-mediated dialog between group 2 innate lymphoid cells and CD4+ T cells 41,
21. Nussbaum, J. C. Type 2 innate lymphoid cells control eosinophil homeostasis. Nature
23. Bernink, J. H. 14,
7,
26. Dudakov, J. A. 336,
37,
Development 141,
7,
364,
31. Ibiza, S. Glial-cell-derived neuroregulators control type 3 innate lymphoid cells and gut defence. Nature 535,
32. Aparicio-Domingo, P. damage.
Lineage Daughters.
Nature 495,
Dll1+ secretory progenitor cells revert to stem cells upon crypt damage. 14,
36. Yan, K. S.
38. Bollrath, J. 15,
Chapter 6

41. Lindemans, C. A. Nature
42. Aden, K. 5,
Nature
Genes 7,
13,
Nature 539,
48. Taniguchi, K. Nature 519,
+
13,
Cells. 11,
review.
Mucosal
celiac disease. 356,
PLoS One
39,
17,
64,
General discussion
6

mice.
long-lived, hair follicle stem cells.
adenocarcinoma. 5,
Chapter 6


A

ADDENDUM


ILCs: innate lymphoid cellsILC1s: Group 1 innate lymphoid cellsILC2s: Group 2 innate lymphoid cellsILC3s: Group 3 innate lymphoid cells
GALT: gut-associated lymphoid folliclesLP: lamina propriaBM: bone-marrowPPs: Peyer PatchesCPs: Cryptopatches
LNs: lymph nodesMLNs: mesenteric lymph nodes
CLPs: common lymphoid progenitorsCHILPs: common helper Innate Lymphoid Cell progenitor
SAA: serum amyloid protein
A

Addendum
MNP: mononuclear phagocytes
Plet1: placenta-expressed transcript 1 proteinECM: extracellular matrix
BMT: bone marrow transfer
MTX: methotrexateDSS: dextran sulfate sodium SS: steady-state
YAP1: yes-associated protein 1EGF: epidermal growth factor
Ihh: indian hedgehogGC: Goblet cellPC: Paneth cellEE: Enteroendocrine cellDT: diphtheria toxin
i.p: intraperitoneal

ENGLISH SUMMARY
which cellular and molecular mechanisms are involved in such crosstalk. Second, we also
during these processes.Chapter 1
In , we demonstrated in an experimental model of chemotherapy-induced
+
by ILC3-derived IL-22 in response to epithelial insult in chapter 3. By blocking IL-22 and
damage. This revealed a previously unacknowledged layer of immune cell-mediated control
In chapter 4
t mice at steady-
English summary
A

in chapter 5
Indeed, when we depleted CD11c+
- Plet1+ DCs are present in SILTs and
Finally, chapter 6
to progress this research.
suggest that SILTs are anatomical sites from where innate immune cells orchestrate epithelial
Addendum160

161
Hoofdstuk 1
In beschadiging van de dunne darm aangetoond dat de aanwezigheid van ILC3s in cryptopatches
het epitheel en het repareren van de schade. Dit model van schade aan de dunne darm
zoals bestraling en chemotherapie. In dit model hebben we kunnen aantonen dat ILC3s
+ darm stamcellen na weefselschade.In hoofdstuk 3
is om darm stamcellen te behouden. Echter, de afwezigheid van IL-22 had geen invloed op
van entero-endocriene cellen. Tevens hebben we kunnen aantonen dat de cytokines IL-11
A

162
schade. Deze controle van darm epitheliale cellen door cellen van het immuun systeem was nooit eerder beschreven.
hoofdstuk 4 hebben we bepaald of dit een gevolg was
ook in de afwezigheid van darmschade. Deze veranderingen bestonden onder andere uit crypt-epitheelafvlakking, een verhoogde expressie van stress en ontstekings genen, en een
die, in mindere mate, ook waargenomen werd na toediening van MTX, wat zou kunnen
hoofdstuk 5
+
+
hoofdstuk 6 een algemene discussie over de meest relevante bevindingen
immuun cellen. Het ontrafelen van de mechanismen die zorgen voor ILC3-gemedieerd
AddendumAddendum

163
immuun cellen de homeostase van het epitheel en het herstel van het weefsel controleren. ILC3s zouden daarom in de toekomst wellicht klinisch gemanipuleerd kunnen worden om
te verbeteren.
A


Science and Technology at IES Guillem de Berguedà high school she obtain her cum laude
Fellowship within the Europe’s Erasmus Exchange Programme in order to perform an
PhD candidate in the research group of Dr. Tom Cupedo at the Department of Hematology
th Framework Programme for research and coordinated by Dr. Mark Coles and Prof.dr. Paul Kaye at the Centre for
and the infrastructure of the immune system and collaborated with industrial and academic
Honors:th
Curriculum vitae
A


LIST OF PUBLICATIONS
Romera-Hernandez M,
Mueller CG.
2016.
YH, Mokry M, Romera-Hernandez M, Cupedo T, Dow L, Nieuwenhuis EE, Shroyer NF, Liu C,
damage. Aparicio-Domingo P, Romera-Hernandez M, Karrich JJ, Cornelissen F, Papazian N, Lindenbergh-Kortleve DJ, Butler JA, Boon L, Coles MC, Samsom JN, Cupedo T.
Helder B, Papazian N, Romera-Hernandez M, Tak PP, Cupedo T, Tas SW.
Romera-Hernandez M, Aparicio-Domingo P, Cupedo T.
A

PhD PORTFOLIO
Name PhD student: Erasmus MC Department:
PhD Period:
Promotor:
Supervisor:
Year ECTS
• Imaging course.201320132014
4.244
• Photoshop and Illustrator CS6 Workshop.• The Microscopic Image Analysis course.• Galaxy for NGS.
20132014
0.34
• Work discussions.• Erasmus Hematology Lectures.• PhD lunch with seminar speaker.
• Literature discussions.2012-2016
10
2
2014, 2016
2014
0.61
1.41.41.4
0.3
A
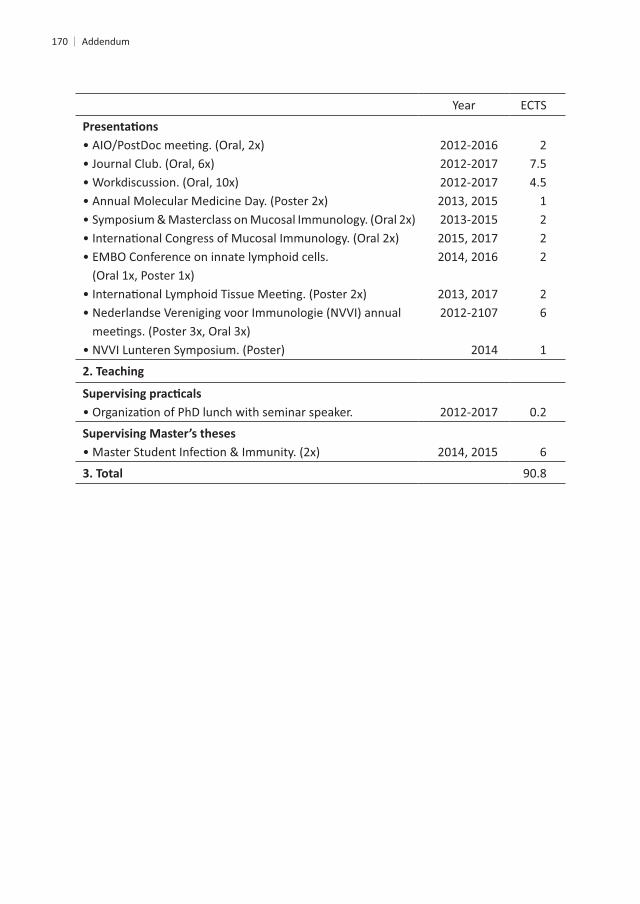
Addendum
Year ECTS
2012-2016
2014, 2016
2014
2
1222
26
1
0.2
6

Acknowledgements
First of all, I would like to thank all the people who have supported me taking this very important step in my career as a PhD student at the Erasmus Medical Center. I have learnt a
discussions. I thank you for teaching me how important is our fundamental research to
To Dr. Tom Cupedo, thanks so much for your great guidance and mentoring as my co-promotor and supervisor. Without your courage and love for science, all this work would
period. I am extremely thankful to you for being always there, so approachable and full of
I want to specially thank Dr. Janneke Samsom for sharing all your knowledge in science,
to convey science into a crystal-clear message.
I cannot be more grateful to any other person in the lab rather than to Natalie. You know it,
Thanks for making me grow as a person next to you. Honestly, I will always have your coco-chocolate sweets as a memory of your special soul.
A

To Ferry, thanks so much for teaching me the importance of being so precise in the lab. I
I am especially thankful to Dicky for showing me everything about immuno-histochemistry.
th
and Linda, it has been a great pleasure to meet you and share experiences in the lab and outside. Thanks for your input during work discussions and your support through all these
Luckily, we will have the chance to discover other parts of the world together, even though
AddendumAddendum

knowledge with me during work discussions and helping me with machines, reagents and protocols in the lab.
de Bospoort and Jan van Kapel for their support. Also, thanks to Egied for his help in the
Thanks to everyone working at the EDC for taking the best care of our precious mice.
Thanks to my friends and colleagues Emmanuele, Cansu, Almira, Tim, Sophie, Adil, Melissa,
beginning you included me as part of your team and I cannot be happier for that. Thanks for opening your arms to me and show how crazy-fun is life. I will always appreciate you all
years.
for their training during my internship and their guidance and encouragement to start my
Acknowledgements
A

career as a PhD.
as an early stage researcher.
shared since my Erasmus in 2011. Jordi, Elena, Lucia, Guillem, Ander, Xabi, Pietro, Joana,
I especially would like to thank my best friend Carla, with who I started this amazing
also to travel around the world with me and always be there when I was sad. I would need
AddendumAddendum

in Canada.
And last but not least, I am really thankful to the Netherlands that has manage to make me
and specially to your tolerant spirit, which makes possible the coexistence of an amazing
Acknowledgements
A

Related Documents