Yeast Yeast 2003; 20: 881–892. Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/yea.994 Research Article Parallel analysis of tagged deletion mutants efficiently identifies genes involved in endoplasmic reticulum biogenesis† Robin Wright 1 *, Mark L. Parrish 1,2 , Emily Cadera 1‡ , Lynnelle Larson 1§ , Clinton K. Matson 1 , Philip Garrett-Engele 2 , Chris Armour 2 , Pek Yee Lum 2 and Daniel D. Shoemaker 2 1 University of Minnesota, Department of Genetics, Cell Biology and Development, 321 Church Street, 6-160 Jackson Hall, Minneapolis, MN 55455, USA 2 Rosetta Inpharmatics, 12040 115th Avenue NE, Kirkland, WA 98034, USA *Correspondence to: Robin Wright, University of Minnesota, Department of Genetics, Cell Biology and Development, 321 Church Street, 6-160 Jackson Hall, Minneapolis, MN 55455. E-mail: [email protected] † The first four authors contributed equally to this work. ‡ Current address: University of California, Department of Molecular and Cellular Biology, 401 Barker Hall 3202, University of California, Berkeley, CA 94720-202, USA. § Current address: Western Washington University, Woodring College of Education, MH 206E, 516 High Street, Bellingham, WA 98225-9090, USA. Received: 25 November 2002 Accepted: 28 January 2003 Abstract Increased levels of HMG-CoA reductase induce cell type- and isozyme-specific proliferation of the endoplasmic reticulum. In yeast, the ER proliferations induced by Hmg1p consist of nuclear-associated stacks of smooth ER membranes known as karmellae. To identify genes required for karmellae assembly, we compared the composition of populations of homozygous diploid S. cerevisiae deletion mutants following 20 generations of growth with and without karmellae. Using an initial population of 1557 deletion mutants, 120 potential mutants were identified as a result of three independent experiments. Each experiment produced a largely non- overlapping set of potential mutants, suggesting that differences in specific growth conditions could be used to maximize the comprehensiveness of similar parallel analysis screens. Only two genes, UBC7 and YAL011W, were identified in all three experiments. Subsequent analysis of individual mutant strains confirmed that each experiment was identifying valid mutations, based on the mutant’s sensitivity to elevated HMG-CoA reductase and inability to assemble normal karmellae. The largest class of HMG-CoA reductase-sensitive mutations was a subset of genes that are involved in chromatin structure and transcriptional regulation, suggesting that karmellae assembly requires changes in transcription or that the presence of karmellae may interfere with normal transcriptional regulation. Copyright 2003 John Wiley & Sons, Ltd. Keywords: HMG-CoA reductase; endoplasmic reticulum; karmellae; DNA microar- ray; Saccharomyces cerevisiae Introduction Increased levels of certain endoplasmic reticu- lum (ER) membrane proteins induce dramatic changes in the amount and organization of the ER. A well-characterized example of such protein- induced ER biogenesis occurs in cells express- ing increased levels of HMG-CoA reductase, a key enzyme in sterol biosynthesis (Brown and Goldstein, 1980; Anderson et al., 1983; Wright et al., 1988, 1990; Goldstein and Brown, 1990), for example a 10-fold increase in the level of the HMG-CoA reductase isozymes, Hmg1p or Hmg2p, induces biogenesis of smooth ER membrane arrays in Saccharomyces cerevisiae that are morpholog- ically and possibly functionally distinct (Lorenz and Parks, 1987; Koning et al., 1996). Hmg1p induces assembly of a nuclear-associated array of smooth ER membranes called karmellae (Wright et al., 1988; Lum and Wright, 1995; Parrish et al., 1995). Hmg2p induces assembly of shorter stacks of smooth ER membranes that may be located in Copyright 2003 John Wiley & Sons, Ltd.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

YeastYeast 2003; 20: 881–892.Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/yea.994
Research Article
Parallel analysis of tagged deletion mutants efficientlyidentifies genes involved in endoplasmic reticulumbiogenesis††
Robin Wright1*, Mark L. Parrish1,2, Emily Cadera1‡, Lynnelle Larson1§, Clinton K. Matson1,Philip Garrett-Engele2, Chris Armour2, Pek Yee Lum2 and Daniel D. Shoemaker2
1University of Minnesota, Department of Genetics, Cell Biology and Development, 321 Church Street, 6-160 Jackson Hall, Minneapolis, MN55455, USA2Rosetta Inpharmatics, 12040 115th Avenue NE, Kirkland, WA 98034, USA
*Correspondence to:Robin Wright, University ofMinnesota, Department ofGenetics, Cell Biology andDevelopment, 321 ChurchStreet, 6-160 Jackson Hall,Minneapolis, MN 55455.E-mail: [email protected]
†The first four authorscontributed equally to this work.
‡Current address: University ofCalifornia, Department ofMolecular and Cellular Biology,401 Barker Hall 3202,University of California, Berkeley,CA 94720-202, USA.
§Current address: WesternWashington University, WoodringCollege of Education, MH 206E,516 High Street, Bellingham,WA 98225-9090, USA.
Received: 25 November 2002Accepted: 28 January 2003
AbstractIncreased levels of HMG-CoA reductase induce cell type- and isozyme-specificproliferation of the endoplasmic reticulum. In yeast, the ER proliferations inducedby Hmg1p consist of nuclear-associated stacks of smooth ER membranes knownas karmellae. To identify genes required for karmellae assembly, we compared thecomposition of populations of homozygous diploid S. cerevisiae deletion mutantsfollowing 20 generations of growth with and without karmellae. Using an initialpopulation of 1557 deletion mutants, 120 potential mutants were identified as aresult of three independent experiments. Each experiment produced a largely non-overlapping set of potential mutants, suggesting that differences in specific growthconditions could be used to maximize the comprehensiveness of similar parallelanalysis screens. Only two genes, UBC7 and YAL011W, were identified in all threeexperiments. Subsequent analysis of individual mutant strains confirmed that eachexperiment was identifying valid mutations, based on the mutant’s sensitivity toelevated HMG-CoA reductase and inability to assemble normal karmellae. Thelargest class of HMG-CoA reductase-sensitive mutations was a subset of genesthat are involved in chromatin structure and transcriptional regulation, suggestingthat karmellae assembly requires changes in transcription or that the presence ofkarmellae may interfere with normal transcriptional regulation. Copyright 2003John Wiley & Sons, Ltd.
Keywords: HMG-CoA reductase; endoplasmic reticulum; karmellae; DNA microar-ray; Saccharomyces cerevisiae
Introduction
Increased levels of certain endoplasmic reticu-lum (ER) membrane proteins induce dramaticchanges in the amount and organization of theER. A well-characterized example of such protein-induced ER biogenesis occurs in cells express-ing increased levels of HMG-CoA reductase, akey enzyme in sterol biosynthesis (Brown andGoldstein, 1980; Anderson et al., 1983; Wrightet al., 1988, 1990; Goldstein and Brown, 1990),
for example a 10-fold increase in the level of theHMG-CoA reductase isozymes, Hmg1p or Hmg2p,induces biogenesis of smooth ER membrane arraysin Saccharomyces cerevisiae that are morpholog-ically and possibly functionally distinct (Lorenzand Parks, 1987; Koning et al., 1996). Hmg1pinduces assembly of a nuclear-associated array ofsmooth ER membranes called karmellae (Wrightet al., 1988; Lum and Wright, 1995; Parrish et al.,1995). Hmg2p induces assembly of shorter stacksof smooth ER membranes that may be located in
Copyright 2003 John Wiley & Sons, Ltd.

882 R. Wright et al.
the cytoplasm or associated with the nucleus orplasma membrane (Hampton et al., 1996; Koninget al., 1996). Analysis of HMG-CoA reductase-induced membrane biogenesis provides opportu-nities to explore the regulation of ER biogenesis(Wright, 1993), for example an important compo-nent of the karmellae-inducing signal in Hmg1p iscontained within a single loop of the membranedomain that is located within the ER lumen (Pro-fant et al., 2000). Sequences on the cytoplasmicside of the ER are also important, suggesting thatthe process of ER biogenesis requires interactionsor processes in both the cytoplasm and the ERlumen (Profant et al., 1999).
To complement our studies of the karmellae-inducing signal in Hmg1p, we have conducted sev-eral genetic screens to identify trans-acting factorsneeded for karmellae assembly. A screen of 2500randomly generated temperature-sensitive mutantsfor their ability to assemble karmellae identifiedvacuole mutants with karmellae-assembly defects,but did not identify mutants with karmellae-dependent growth defects (Koning et al., 2002).It is likely that these vacuole biogenesis genesfunction indirectly in the cell’s ability to assem-ble karmellae. In another screen, potential karmel-lae assembly mutants were identified based ontheir inability to grow on galactose, which inducedkarmellae assembly via expression of HMG1 fromthe GAL1 promoter. However, subsequent analysisof these potential mutants revealed that the screenwas efficiently identifying mutations in galactosemetabolism, but not in karmellae assembly (unpub-lished observations).
The recent availability of a complete set ofoligonucleotide-tagged deletion mutants, producedby the Saccharomyces Genome Deletion Project,enables a systematic examination of non-essentialgenes involved in a process or pathway of interest(Winzeler and Davis, 1997; Giaever et al., 1999;Winzeler et al., 1999, 2000; Barth and Thumm,2001; Bianchi et al., 2001; Giaever et al., 2002;Oshiro et al., 2002). We report that analysis ofa population of tagged deletion mutants had ahigh success rate for identifying genes requiredfor karmellae assembly and robust growth of cellsexpressing increased levels of Hmg1p, but was noteffective for identifying mutations whose growthdefects were suppressed by increased levels ofHmg1p. In addition, we found that this approachwas sensitive to specific experimental parameters,
enabling subtle differences in experimental designto yield different classes of mutants.
Materials and methods
Creation of deletion mutant populationEach of 1557 different homozygous deletion mut-ants in the S288C derivative BY4743 (Mata/αhis3�1/his3�1 leu2�0/leu2�0 ura3�0/ura3�0met15�0/+ lys2�0/+) was independently patchedonto solid YPD medium (2% each yeast extract,proteose peptone, dextrose and agar) containing150 mg/l G418. The identity of all mutant strainsin this population can be obtained by request.Equal amounts of each mutant were harvested andcombined to generate a mixed mutant populationin which each individual strain was present inapproximately equal abundance. This mutant pop-ulation was suspended in 15% glycerol and storedat 80 ◦C. To transform the population, portions ofthe frozen culture were scraped into liquid YPDmedium to a final concentration of 0.25 OD600/ml.After the cells had resumed growth and completedone doubling time (about 8 h), the mutant pop-ulation was harvested and made competent fortransformation by washing three times in 0.1 M
LiCl, followed by an overnight incubation in 0.1M LiCl. Aliquots of the competent yeast weretransformed with URA3-containing plasmids usingminor modifications of the transformation protocolfrom the Saccharomyces Genome Deletion Project(Multiwell Transformation Protocol, http://www-sequence.stanford.edu/group/yeast deletion pro-ject/deletions3.html). Transformants were selectedon rich minimal medium lacking uracil (0.17%yeast nitrogen base, 0.5% ammonium sulphate,2% casamino acids, 2% agar, with 30 µg/ml ade-nine, 20 µg/ml histidine, 40 µg/ml lysine, 40 µg/mlleucine, 20 µg/ml methionine, 30 µg/ml tryptophanand 20 µg/ml tyrosine). All of the resulting trans-formant colonies (approximately 30 000–140 000independent transformants depending on the exper-iment) were pooled, rinsed once by centrifugationand resuspension in distilled water, resuspended in15% glycerol at a concentration of 3 OD600/ml, andfrozen in 1 ml aliquots.
Competitive growth conditionsAn aliquot of the transformed mutant populationcontaining 3 × 107 cells was thawed and used
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

Parallel analysis of tagged deletion mutants in ER biogenesis 883
to inoculate 50 ml rich minimal medium broth.The initial inoculation was to a cell density of0.05 OD600/ml and cells were grown at 26 ◦C,28 ◦C, or 30 ◦C (depending on the experiment)in a shaking water bath until they reached acell density of approximately 0.2 OD600/ml. Atthis point, 25 ml of culture containing approx-imately 106 cells were removed and 25 ml offresh medium were added to the remaining cul-ture. The cycles of growth and dilution contin-ued, maintaining cells in log-phase growth, untilthe culture had undergone 20 doublings (approx-imately 48 h for cultures grown with glucose ascarbon source and 50 h for cultures grown withgalactose as carbon source). Three different com-petition experiments were carried out (Table 1).The first experiment compared the compositionof the same mutant population transformed withpAK266 (URA3, CEN6 with HMG1 under controlof GAL1 promoter; Koning et al., 1996) following20 generations of growth on 2% sucrose-containingmedium (no karmellae induction) vs. 2% galac-tose plus 2% sucrose containing medium (karmel-lae induction). The second experiment, which wasrepeated three times, compared growth of themutant population transformed with the vector con-trol pBM258 (URA1, CEN4, GAL1/10 promotersequences, equivalent to pBM150; Johnston andDavis, 1984) vs. the mutant population transformed
with the karmellae-inducing plasmid pAK266; bothpopulations were grown on rich minimal mediumcontaining 2% galactose (karmellae-induction). Thethird experiment compared growth of the mutantpopulation transformed with a control plasmidthat expresses high levels of HMG-CoA reduc-tase activity but no karmellae (pRH127-3, a 2µURA3 plasmid containing the Hmg1p catalyticdomain, Hmg1pcat, expressed from the constitu-tive TDH3 promoter; Donald et al., 1997, pro-vided by Randy Hampton, UC, San Diego) vs.the mutant population transformed with a plasmidthat expressed the entire Hmg1p, thus inducinghigh HMG-CoA reductase activity and also karmel-lae assembly (pLL716, a CEN6, URA3 plasmidcontaining HMG1 expressed from the constitutiveTDH3 promoter; created by replacing the GAL1promoter in pAK266 with the TDH3 promoterencoded on a PCR product using routine plasmidgap-repair strategy, as described by Orr-Weaveret al., 1988; Muhlrad et al., 1992). In this exper-iment, expression of Hmg1p and Hmg1pcat wasconstitutive, so that both populations were grownon rich minimal medium containing 2% glucose.
Oligonucleotide tag amplification andmicroarray analysisGenomic DNA was prepared from 3 OD600 units ofeach culture using a modification of the MasterPure
Table 1. Experimental growth conditions
Experiment Control culture Experimental culture Notes
1 Deletion mutant populationcontaining the galactose-inducibleHMG1 gene on pAK266; grown insucrose to repress karmellaeassembly
Deletion mutant populationtransformed with thegalactose-inducible HMG1 gene onpAK266; grown in galactose-sucroseto induce karmellae assembly1
Growth in rich minimal medium at26 ◦C; 18 strains identified aspotential karmellae-sensitivemutants2
2 Deletion mutant populationtransformed with pBM258 vector;grown in galactose
Deletion mutant populationtransformed with thegalactose-inducible HMG1 gene onpAK266; grown in galactose toinduce karmellae assembly1
Growth in rich minimal medium at28 ◦C; 60 strains identified aspotential karmellae-sensitive mutants
3 Deletion mutant populationtransformed with constitutivelyexpressed HMG1 catalytic domainon pRH127-3; grown in glucose;expresses high levels of HMG-CoAreductase activity but does notassemble karmellae
Deletion mutant populationtransformed with constitutivelyexpressed HMG1 on pLL713; grownin glucose; expresses high levels ofHMG-CoA reductase activity andassembles karmellae
Growth in rich minimal medium at30 ◦C; 61 strains identified aspotential karmellae-sensitive mutants
1 This strain also expressed increased HMG-CoA reductase activity under these growth conditions.2 ‘Karmellae-sensitive’ is defined as a strain in which the hybridization signal from the experimental culture is at least five-fold lower than thatfrom the control culture.
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

884 R. Wright et al.
Yeast DNA protocol (Epicentre, Madison, WI)that included addition of 0.5 mm diameter glassbeads to the meniscus of the resuspended cellsample and use of a Mini-Beadbeater (BioSpecProducts, Bartlesville, OK) instead of vortexing.Samples that were examined included the pre-transformation mutant population and the post-transformation mutant populations at time zero,and post-transformation mutant populations after20 doublings under experimental or control condi-tions. The unique 20 bp oligonucleotide sequences(‘uptags’ and ‘downtags’) that mark each mutantstrain were separately amplified from genomicDNA, using primers to the tag priming sites thatare common to all deletions (B-U2-comp and U1 orB-D2-comp and D1). In the first experiment, bothprimer pairs were used. However, because no sig-nificant differences were observed in data obtainedwith different primer pairs, subsequent experimentsused only probes prepared from the downtags. ThePCR-amplified tags from control and experimentalcultures were simultaneously hybridized, as previ-ously described, to a DNA microarray containingoligonucleotides complementary to each individualmutant uptag or downtag sequence (Winzeler et al.,1999). The arrays were examined by laser scanningmicroscopy and Affymetrix GeneChip software, aspreviously described (Winzeler et al., 1999).
Results
Three different competition experimentsproduced three different sets of potentialkarmellae-sensitive mutants
To identify genes involved in karmellae assem-bly, we sought to identify deletion mutants inwhich karmellae assembly resulted in an observ-able change in cell division rate. To this end,we compared the composition of a population of1557 different deletion mutants following growthfor 20 generations under control vs. karmellae-inducing conditions. The deletion population con-tained 637 deletion strains corresponding to knowngenes, with the remainder corresponding to ORFsof unknown function (list available on request).Following the competitive growth period, genomicDNA was isolated from the two cultures and usedas the template to amplify the unique oligonu-cleotide tags that mark each individual mutantstrain. The primers for the PCR reaction were
labelled with either Cy-3 or Cy-5, allowing PCRproducts from experimental and control popu-lations to be differentially labelled and simul-taneously hybridized to a DNA oligonucleotidemicroarray representing every mutant in the popu-lation. If a particular mutant divides at the samerate in both conditions, equal PCR product willbe present in both the experimental and controlDNA samples, producing a hybridization signalratio of 1. However, if a particular mutant dividesat a decreased rate when karmellae are induced (i.e.is karmellae-sensitive), less PCR product represent-ing that mutant will be present in the karmellae-induced culture than in the control. Hybridizationof these PCR products to the microarray producesa hybridization signal ratio of experimental vs.control DNA that is less than 1. If a particularmutant divides at a greater rate when karmellaeare induced than in the control culture (i.e. iskarmellae-suppressed), more PCR product repre-senting that mutant will be present in the karmellae-containing culture than in the control. Hybridiza-tion of these PCR products produces a hybridiza-tion signal ratio of experimental vs. control DNAthat is greater than 1.
Three separate competition experiments werecarried out (Table 1). In the first experiment, themutant population, transformed with a galactose-inducible HMG1 gene (pAK266), was grownunder inducing (galactose/sucrose) or non-inducing(sucrose) conditions. This experiment was con-ducted at 26 ◦C to allow survival of potentialtemperature-sensitive mutants that are defective forkarmellae assembly, such as certain vacuole bio-genesis mutants (Koning et al., 2002). The secondexperiment compared the growth of the mutantpopulation transformed with a control plasmid(pBM258) to the mutant population transformedwith a karmellae-inducing plasmid (pAK266). Bothpopulations were grown at 28 ◦C on galactosemedium, minimizing differences in growth ratedue to differences in carbon source. Finally, thethird experiment compared growth of the mutantpopulation transformed with a plasmid that pro-duced high HMG-CoA reductase activity, but nokarmellae (pRH127-3), to the mutant populationtransformed with a plasmid that produced bothhigh HMG-CoA reductase activity and karmellae(pLL716). Because expression of the Hmg1p pro-teins from these plasmids was constitutive, both
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

Parallel analysis of tagged deletion mutants in ER biogenesis 885
cultures were grown on glucose-containing richminimal medium.
Among the three experiments, 120 potentialkarmellae-sensitive mutants were identified thatdisplayed five-fold or greater decreases in the ratioof hybridization signal in the karmellae-inducedculture vs. the control (Table 2). Based on the over-all doubling rate of the mutant population, we esti-mate that this decreased signal ratio corresponds toan average increase in doubling time from 2.5 hto 2.7 h in galactose-containing medium or from2.4 h to 2.6 h in glucose-containing medium. Ofthe 120 potential mutants, only two, yal011w�and ubc7�, were identified in all three experi-ments; 15 additional mutants were identified askarmellae-sensitive in two of the experiments.Thus, most of the potential mutants were identi-fied in only one of the experiments, indicating thateach experiment produced a largely unique set ofmutants.
Most prospective mutants displayed anHMG-CoA reductase-dependent slow growthphenotype when grown clonally and assembledabnormal karmellae
In theory, the competition experiment identi-fied strains with only subtle growth differencesunder karmellae-inducing conditions (on average,a ∼12 min or greater increase in doubling time).
However, this difference in growth rate occurredwhen the mutant was growing in liquid mediumin competition with more than 1500 other strains.To test whether such subtle growth differencesin a population would also be observable in themutant strains growing clonally on solid medium,we examined the growth of all seventeen strainsthat were identified as karmellae-sensitive in twoor more of the experiments. Each individual taggeddeletion mutant strain was transformed indepen-dently with control or karmellae-inducing plasmidsand the growth of serially diluted liquid culturesplated on solid rich minimal medium was examinedat 16 ◦C, 26 ◦C, 30 ◦C and 37 ◦C (Figure 1). Elevenout of 17 multiply-hit strains showed observ-able decreases in growth on solid medium underkarmellae-inducing vs. non-inducing conditions atone or more temperatures. Thus, for most potentialkarmellae-sensitive mutants, slow growth was not aconsequence of competition with other mutants orof growth in liquid medium, but instead reflected ageneralized slowing of growth rate upon karmellaeinduction.
Expression of Hmg1p from the GAL1 pro-moter results in an approximately 10-fold eleva-tion in Hmg1p. This increase has at least twoconsequences: induction of karmellae assemblyand induction of a 10-fold increase in HMG-CoA reductase activity. To determine whether or
Table 2. Genes predicted by microarray analysis to result in karmellae sensitivity1 when deleted
Genes identified in Genes ORFS
All experiments UBC7 YAL011WExperiments 1 and 2 GOS1 YML013W; YML035C-AExperiments 1 and 3 CUE1; HTZ1; JNM1; YIM1 YOL054WExperiments 2 and 3 CKA2; DRS2; HIR2; HPC2; NUP53; RPS6B YMR153C-AExperiment 1 EFT2; GSG1; SIP1; STE20; VPS61 YAL048C; YCL005W; YNL334CExperiment 2 ARD1; ARG1; CCR4; CHA4; CIN1; CIN2; ECM18; GPD2; ILV1;
NIP100; OPI1; RPL15B; RPL18B; RPS0A; RVS167; SAT4; SGS1;SHE4; SIC1; SNF2; SPT21; RPN10; SXM1; XDJ1
YCL060C; YDR359C; YDR386W; YEL007W;YEL031W; YER056C-A; YGR181W; YGR182C;YLR061W; YLR065C; YLR109W; YLR111W;YML010W-A; YML033W; YMR156C; YMR191W;YMR230W; YNL246W; YNL273W; YOR295W;YOR308C; YOR359W; YPL202C; YPL226W
Experiment 3 ARP1; ARP6; CIN8; CKB2; COT1; CPR6; FOB1; GIM4; GRX3;HAC1; ICY1; MKK2; MLH1; OYE2; RGP1; RPL18A; RTG1;STA3; SWI5; THP2; UBP6; VIK1; VPS30; YEN1
YDR049W; YDR120C; YDR383C; YEL001C;YER068C-A; YER079W; YFR007W; YGR164W;YHL023C; YHR115C; YHR126C; YHR133C;YHR156C; YLR190W; YLR217W; YMR044W;YMR162C; YMR166C; YMR299C; YOR051C;YOR066W; YOR314W; YOR342C
1 A potential karmellae-sensitive mutant is operationally defined as one that displays a five-fold or greater decrease in hybridization ratio ofPCR-amplified DNA from that strain grown under karmellae-inducing vs. control conditions.
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

886 R. Wright et al.
Glucose(control)
Galactose(HMG1 induced)
gos1∆30°C
yml035c-a∆37°C
ypl120w∆37°C
ylr 085c∆26°C
pBM258
pAK266
pDP304
pBM258
pAK266
pDP304
pBM258
pAK266
pDP304
pBM258
pAK266
pDP304
Figure 1. Serial dilution growth assay of potentialHMG-CoA reductase-sensitive mutants. This figure showsexamples of the growth assay used to test the validity ofprospective mutants identified in the competition assays.Each mutant was transformed with a vector control(pBM258), a plasmid that induces both karmellae assemblyand increased HMG-CoA reductase activity (pAK266),and a plasmid that induces karmellae but no increasedHMG-CoA reductase activity (pDP304). The gos1� mutanttransformed with all three plasmids grew similarly ongalactose, indicating that it did not have a karmellaeor HMG-CoA reductase-sensitive phenotype. The otherthree mutants displayed increasing sensitivity to HMG-CoAreductase activity (poor growth on galactose with pAK266).All mutants were assayed at three different temperatures;this figure shows only the single temperature at which themost severe phenotype was observed
not the slow-growth phenotype of the prospectivekarmellae-sensitive mutants was due to sensitivityof the mutant to karmellae, to increased HMG-CoA reductase activity, or both, we compared thegrowth of strains expressing wild-type Hmg1p orcatalytically-inactive Hmg1p in which the essen-tial histidine at position 1020 has been replacedwith glutamine (pDP304; Profant et al., 1999). Thismutant protein induces normal levels of karmellae,but produces no measurable increase in HMG-CoAreductase catalytic activity. All but one (yl013w�)of the potential karmellae-sensitive mutants grewnormally when they expressed catalytically inac-tive Hmg1p, indicating that, rather than beingkarmellae-sensitive, these strains were sensitive toelevated HMG-CoA reductase activity (Table 3).
Table 3. Growth and karmellae assembly phenotype ofpotential HMG-CoA reductase-sensitive mutants identifiedin multiple experiments1
Growth KarmellaeGene or
locus
Karmellae-sensitive Abnormal YML013W
HMG-CoA Abnormal CKA2reductase-sensitive CUE1
DRS2GOS1HIR2HTZ1UBC7YAL011WYMR153C-A
Normal HPC2NUP53RPS6BYOL054W
Normal Abnormal JNM1YIM1
1 Data represent clonal analysis of all mutants that were identified intwo or more of the three experiments.
One possibility is that the sensitivity of themutants to HMG-CoA reductase activity wascoincident with abnormal karmellae assembly. Ifso, a cell’s poor growth in response to elevatedHMG-CoA reductase activity might be due toits inability to properly organize karmellae mem-branes. To test this hypothesis, we used DiOC6staining to examine the morphology of karmellae inthe mutants. Eleven of the 17 mutants tested assem-bled karmellae membranes that were highly disor-ganized or less abundant than in wild-type controls,including four of the mutants that grew nor-mally with elevated HMG-CoA reductase. Thus,the majority (11/16) of the putative ‘karmellae-sensitive’ mutants identified in the screen weredefective for karmellae assembly, whether or notthey displayed a clonal slow-growth phenotype.
This high success rate for identifying mutantswith karmellae assembly defects might simplymean that many of the deletion mutants arekarmellae-defective, whether or not they have akarmellae-dependent growth defect. To test thispossibility, we randomly selected 10 mutants fromthe population that were not identified in any of thethree experiments (arg4�, asn2�, cwh36�, fre6�,ntg2�, pau2�, pet100�, rps7A�, spa2�, zrt2�).Two of these mutants (cwh36� and pet100�) did
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

Parallel analysis of tagged deletion mutants in ER biogenesis 887
not grow on galactose and consequently couldnot be assayed for karmellae assembly using agalactose-inducible HMG1 gene. The remainingeight mutants assembled normal karmellae andgrew normally with elevated HMG-CoA reduc-tase. Thus, HMG-CoA reductase sensitivity andabnormal karmellae assembly appear to be rareamong the mutant strains in this population andthe microarray data effectively identified mutantswith karmellae assembly defects.
No direct quantitative relationship wasobserved between the log(hybridization ratio)and measured growth rates of the mutants inclonal cultures
To determine whether the differences in hybridiza-tion ratio observed via the microarray approachcorrelated closely with the actual differences indoubling times of the individual mutant strains, wecompared the growth rates of all multiply-identifiedmutants in control and karmellae-inducing condi-tions (Table 4). Fourteen of the 17 strains examinedshowed the expected karmellae-dependent increase
in doubling time, ranging from 4 to 99 min.However, no obvious direct correlation betweenthe log(hybridization ratio) data obtained fromthe microarray and the measured doubling timewas apparent, for example, the log(hybridizationratio) of the yol054w� mutant was −1.78, indi-cating a 60-fold decrease in the abundance ofthis mutant following competitive growth for 20generations under karmellae-inducing conditionsvs. the control. However, when cultured clonally,the yol054w� had a doubling time increase ofonly 26 min. In contrast, a 91 min increase indoubling time was observed for htz1� mutants,which had a log(hybridization ratio) in the com-petition experiment of −1.31, indicating a 20-fold decrease in the abundance of this mutant.Finally, the doubling time of the rps6b� mutantwas not affected by karmellae assembly, in spiteof the log(hybridization ratio) of −0.78. Thus,the hybridization ratio appeared to predict karmel-lae sensitivity with reasonable accuracy, but didnot predict the degree of sensitivity of individualmutants in clonal growth. Thus, differences in the
Table 4. Hybridization ratio and doubling times
Doubling time (min)
Mutation
Log (hybridization ratio,experimental/
control)1Vector-transformed
controlpAK266
transformed Change
WT 214 210 −4yol054w� −1.78 139 164 26yml035c-a� −1.67 256 320 65fun36� −1.49 170 173 3ubc7� −1.38 140 225 84htz1� −1.31 138 230 91yml013w� −1.31 274 281 8drs2� −1.22 196 236 40cue1� −0.97 133 158 25jnm1� −0.97 216 315 99yim1� −0.89 230 278 47gos1� −0.87 219 244 25cka22� −0.81 147 177 29rps6b� −0.78 474 470 −4hpc22� −0.77 154 196 42ymr153c-a� −0.76 155 146 −9hir2� −0.75 216 315 98nup53� −0.66 287 365 79AVERAGE(for mutants) −1.08 208 250 44
1 Hybridization ratio is the log of the ratio of the signal produced by the PCR product representing the individual mutantstrain in the experimental culture vs. the PCR product representing that gene in the control culture.
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

888 R. Wright et al.
log(hybridization signal) did not have predictivevalue for ordering the severity of mutant pheno-type.
Mutants identified in only one of theexperiments were also enriched for strainswith HMG-CoA reductase sensitivity andabnormal karmellae assembly
To determine whether or not potential mutantsidentified in only one of the experiments werealso of interest, we selected the 15 mutants fromExperiment 3 that displayed the highest negativelog(hybridization ratio). In addition, we randomlychose three mutants identified in only Experiment1 and four mutants identified in only Experi-ment 2. Each of these strains was transformedindependently with karmellae-inducing and con-trol plasmids. The ability of the transformants toassemble karmellae and to grow on medium thatinduced elevated expression of Hmg1p was exam-ined (Table 5). Most (14/19) of these potentialmutants displayed HMG-CoA reductase-sensitivegrowth, abnormal karmellae assembly, or both.Thus, although the number of false positives washigher than that observed in potential mutants iden-tified in multiple experiments, these data were alsocontained valid mutants of interest.
The microarray data were not useful foridentifying karmellae- or HMG-CoAreductase-suppressed mutants
The microarray data from Experiment 3 identi-fied both potential HMG-CoA reductase-sensitivemutants, which grew more slowly with increasedHMG-CoA reductase levels, and HMG-CoA reduc-tase-suppressed mutants, which grew more rapidlywith increased HMG-CoA reductase levels. Weexamined the growth and karmellae assembly phe-notypes of 24 of the mutants that grew more rapidlywith increased Hmg1p (Table 6). When grownclonally on solid medium, none of these strainsgrew better with increased levels of HMG-CoAreductase expression. In contrast, seven grew moreslowly, indicating they were actually HMG-CoAreductase-sensitive. Indeed, two of these mutants(ard1� and sgs1�) had been identified as potentialHMG-CoA reductase sensitive mutants in Exper-iment 2. In addition, only six strains among the24 had a karmellae-assembly defect. Thus, the
Table 5. Growth and karmellae assembly phenotype ofpotential HMG-CoA reductase-sensitive mutants identifiedin only one experiment1
Growth KarmellaeGene or
locus
Karmellae and HMG-CoA Abnormal ARD1reductase-sensitive SGS1
HMG-CoA Abnormal ARP6reductase-sensitive GIM4
THP2VIK1YDR383C
Normal ARP1CKB2CPR6SPT21VPS30
Normal Abnormal GRX3YFR007WYOR051C
Normal EFT2HAC1YEN1
1 Data represent only the subset of mutants identified in onlyone of the experiments that were retested for clonal growth andkarmellae assembly.
expected growth phenotype of the potential sup-pressed mutants was not reproduced in clonalgrowth. However, mutants of interest (HMG-CoA reductase sensitive and/or karmellae assemblydefective) were more common among these strainsthan in the general mutant population.
Discussion
Several observations suggest that, in addition toincreased expression of HMG1, karmellae assem-bly requires the action of other genes, for example,if only Hmg1p were needed for karmellae assem-bly, expression of Hmg1p in mammalian cellsshould also lead to assembly of karmellae. How-ever, Hmg1p instead induces assembly of crystal-loid ER rather than karmellae, suggesting that cell-specific factors are required (Wright et al., 1990).In addition, mutants that are defective in karmel-lae assembly can assemble normal Hmg2p-inducedmembranes, suggesting that the processes may begenetically distinguishable (Koning et al., 2002).With the knowledge that other genes were likely
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

Parallel analysis of tagged deletion mutants in ER biogenesis 889
Table 6. Growth and karmellae assembly phenotype ofprospective ‘HMG-CoA reductase-suppressed’ mutants1
Growth KarmellaeGene or
locus
Karmellae and HMG-CoA Abnormal ARD12
reductase-sensitive SGS13
HMG-CoA Abnormal BNI1reductase-sensitive Normal ESC1
ESC2TYE7YLR184W
Normal Abnormal KRH1PHO8YMR193C-A
Normal DPL1EFT2ECM14GRE3MSI1RIT1RPL36ASAL6SPT213
VPS29YPL102CYPL208W
Petite Not tested GLN3MSS51YOL027C
1 Defined as mutants that have a five-fold or greater increase inhybridization ratio in culture with high HMG-CoA reductase levels vs.control in Experiment 3.2 Also identified and confirmed as potential HMG-CoA reductase-sensitive mutant in Experiment 2.3 Also identified as a potential karmellae-sensitive mutant inExperiment 1.
to be involved in karmellae assembly, we under-took a variety of genetic approaches over the past13 years, each of which was at best marginallysuccessful. The availability of the tagged deletionmutants provided an opportunity to approach theissue more systematically.
The logic of the competitive growth screendescribed in this paper is based on the factthat karmellae do not confer a growth pheno-type upon wild-type cells (Wright et al., 1988).We hypothesized that the ability of normal cellsto grow robustly with karmellae required the cellto properly organize karmellae membranes, forexample, in normal cells that contain karmel-lae, nuclear pores are no longer distributed uni-formly but instead are concentrated in the region
of the nuclear envelope opposite from the nucle-olus (Wright et al., 1988). If karmellae assemblywere abnormal, nuclear pore clustering might becompromised, resulting in abnormal nuclear porefunction and decreased growth rate or death. Con-sistent with this hypothesis, we were able to iden-tify mutations that grew slowly when Hmg1p wasexpressed at increased levels and the majority ofthese mutants (7 of the 11 mutants identified inmultiple experiments and 6 of 10 of the mutantsidentified in single experiments) also assembledabnormal karmellae. However, further experimentsdemonstrated that the slow growth phenotype wasnot directly due to abnormal karmellae but insteadresulted from sensitivity of the mutants to increasedlevels of HMG-CoA reductase activity. Expressionof very high levels of the Hmg1p catalytic domainleads to slow growth in wild-type cells, possiblydue to the presence of excess squalene (Donaldet al., 1997). Thus, these mutants may be particu-larly sensitive to even moderately increased levelsof squalene that might accompany the, 10-fold ele-vation in Hmg1p levels.
If the mutants identified in these screens weresimply sensitive to increased HMG-CoA reductaseactivity, we would not expect to find such a highfrequency of abnormal karmellae assembly amongthem, i.e. if the two processes were completelyunlinked, we would expect most cells would beeither karmellae-defective or HMG-CoA reductase-sensitive. Nine of the 36 mutants that we studied indetail were sensitive to HMG-CoA reductase activ-ity but assembled karmellae that appeared normalat the light microscope level. These observationsare consistent with the hypothesis that karmellaeassembly and HMG-CoA reductase activity areseparate processes. However, the majority of theHMG-CoA reductase-sensitive mutants also wereunable to assemble normal karmellae, suggestingthat the two phenotypes may be linked. Indeed, itis possible that the ‘normal’ karmellae in the sub-set of HMG-CoA reductase-sensitive mutants mayhave defects that are not visible at the light micro-scope level.
These observations lead us to hypothesize thatproper organization of karmellae membranes mayhelp to regulate HMG-CoA reductase activity, e.g.enclosure of the Hmg1p protein within the inte-rior layers of karmellae may inhibit access of
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.
890 R. Wright et al.
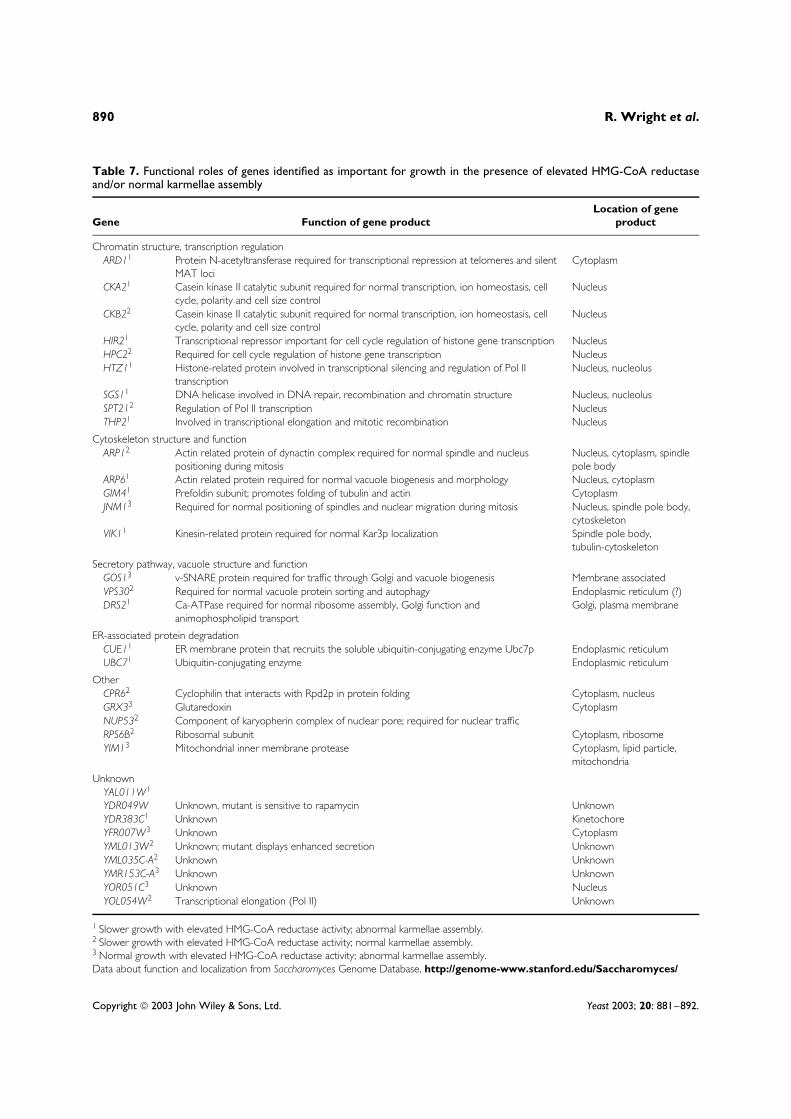
Table 7. Functional roles of genes identified as important for growth in the presence of elevated HMG-CoA reductaseand/or normal karmellae assembly
Gene Function of gene productLocation of gene
product
Chromatin structure, transcription regulationARD11 Protein N-acetyltransferase required for transcriptional repression at telomeres and silent
MAT lociCytoplasm
CKA21 Casein kinase II catalytic subunit required for normal transcription, ion homeostasis, cellcycle, polarity and cell size control
Nucleus
CKB22 Casein kinase II catalytic subunit required for normal transcription, ion homeostasis, cellcycle, polarity and cell size control
Nucleus
HIR21 Transcriptional repressor important for cell cycle regulation of histone gene transcription NucleusHPC22 Required for cell cycle regulation of histone gene transcription NucleusHTZ11 Histone-related protein involved in transcriptional silencing and regulation of Pol II
transcriptionNucleus, nucleolus
SGS11 DNA helicase involved in DNA repair, recombination and chromatin structure Nucleus, nucleolusSPT212 Regulation of Pol II transcription NucleusTHP21 Involved in transcriptional elongation and mitotic recombination Nucleus
Cytoskeleton structure and functionARP12 Actin related protein of dynactin complex required for normal spindle and nucleus
positioning during mitosisNucleus, cytoplasm, spindlepole body
ARP61 Actin related protein required for normal vacuole biogenesis and morphology Nucleus, cytoplasmGIM41 Prefoldin subunit; promotes folding of tubulin and actin CytoplasmJNM13 Required for normal positioning of spindles and nuclear migration during mitosis Nucleus, spindle pole body,
cytoskeletonVIK11 Kinesin-related protein required for normal Kar3p localization Spindle pole body,
tubulin-cytoskeleton
Secretory pathway, vacuole structure and functionGOS13 v-SNARE protein required for traffic through Golgi and vacuole biogenesis Membrane associatedVPS302 Required for normal vacuole protein sorting and autophagy Endoplasmic reticulum (?)DRS21 Ca-ATPase required for normal ribosome assembly, Golgi function and
animophospholipid transportGolgi, plasma membrane
ER-associated protein degradationCUE11 ER membrane protein that recruits the soluble ubiquitin-conjugating enzyme Ubc7p Endoplasmic reticulumUBC71 Ubiquitin-conjugating enzyme Endoplasmic reticulum
OtherCPR62 Cyclophilin that interacts with Rpd2p in protein folding Cytoplasm, nucleusGRX33 Glutaredoxin CytoplasmNUP532 Component of karyopherin complex of nuclear pore; required for nuclear trafficRPS6B2 Ribosomal subunit Cytoplasm, ribosomeYIM13 Mitochondrial inner membrane protease Cytoplasm, lipid particle,
mitochondria
UnknownYAL011W1
YDR049W Unknown, mutant is sensitive to rapamycin UnknownYDR383C1 Unknown KinetochoreYFR007W3 Unknown CytoplasmYML013W2 Unknown; mutant displays enhanced secretion UnknownYML035C-A2 Unknown UnknownYMR153C-A3 Unknown UnknownYOR051C3 Unknown NucleusYOL054W2 Transcriptional elongation (Pol II) Unknown
1 Slower growth with elevated HMG-CoA reductase activity; abnormal karmellae assembly.2 Slower growth with elevated HMG-CoA reductase activity; normal karmellae assembly.3 Normal growth with elevated HMG-CoA reductase activity; abnormal karmellae assembly.Data about function and localization from Saccharomyces Genome Database, http://genome-www.stanford.edu/Saccharomyces/
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

Parallel analysis of tagged deletion mutants in ER biogenesis 891
the enzyme to its substrate, thus decreasing fluxthrough the sterol biosynthetic pathway and min-imizing squalene accumulation. If so, the primarydefect in mutants that exhibit HMG-CoA reductasesensitivity may actually be karmellae assembly.
Several specific results of this analysis helpus understand the function of as yet unidentifiedORFs, for example, the NUP53 gene overlaps atits 3′ end with the 3′ end of an open readingframe on the opposite strand, YMR053C-A. Bothgenes were identified in the screen as HMG-CoA reductase-sensitive, but only ymr052c-a�
strains assembled abnormal karmellae. This resultsuggests that loss of NUP53 may cause HMG-CoAreductase-sensitivity because of the collateral lossof the carboxyl terminus of the YMR053C-A geneproduct. It also indicates that YMR053C-A probablyencodes a valid gene.
Genes that were not identified in the screens canalso be informative, e.g. CKA2 and CKB2 wereidentified in the screens as HMG-CoA reductase-sensitive. However, the related casein kinase gene,CKA1, was represented by a deletion mutant inthe population but was not identified in any ofthe screens. Thus, the two kinases must havedistinct roles with regard to resistance to ele-vated HMG-CoA reductase. An additional exam-ple involves ARP1. Loss of ARP1 makes cellssensitive to elevated levels of HMG-CoA reduc-tase but arp1� mutants assemble karmellae thatappear normal by light microscopy. Arp1p interactsphysically with Arp10p, Jnm1p, Jsn1p, Nip100p,Nup1p, Srp1p, Ubc6p and YJR008Wp and genet-ically with BEM1, BIM1, BNI1, CIN8, KAR9 andSTT4 (see the Saccharomyces Genome Database,http://biodata.mshri.on.ca/grid/servlet/SearchResults?keywords=YHR129C). Of these interact-ing genes, the mutant population contained bem1�,bim1�, cin8� and jnm1�. Among these mutants,bni1�, jmn1� and cin8� were identified as HMG-CoA reductase sensitive in the screens but bim1was not, suggesting that these proteins may havedifferent roles in the cell’s response to elevationsin HMG-CoA reductase activity.
The largest class of genes in which mutationsproduced both HMG-CoA reductase sensitivity anddefective karmellae assembly were involved inchromatin structure and/or transcriptional regula-tion (Table 7). This observation may indicate thateither changes in gene expression dependent onthese genes (ARD1, CKA2, HIR1, HTZ1, SGS1
and THP2) are important in karmellae assemblyand/or HMG-CoA reductase resistance, or that thepresence of karmellae alters the structure of thenucleus in a manner that necessitates the functionof these previously non-essential genes, for exam-ple changes in the levels or function of histonesmay be important for maintaining proper chromatinfunction in nuclei with karmellae-induced changesin nuclear morphology or nuclear pore distribu-tion. Alternatively, increased activity of HMG-CoAreductase may result in changes in ER membranefluidity that disrupt normal associations betweenchromatin and the inner nuclear membrane, neces-sitating the function of chromatin remodelling pro-teins for the maintenance of cell growth.
All three competition experiments were designedto identify karmellae-sensitive mutants and wereexpected to identify a largely overlapping set ofmutants. Indeed, isolation of the same mutant inmultiple experiments is frequently a good indica-tion that the gene represented by the mutation has avalid role in the process under investigation. Thisexpectation was not supported by the results. Infact, most genes were identified in only one ofthe experiments. Nevertheless, the non-overlappingsets of genes identified by each experiment wereenriched for genes with roles in karmellae biogen-esis and/or resistance to high levels of HMG-CoAreductase. Thus, in considering a comprehensivefunctional analysis of genes involved in a particularprocess using competition analysis, identification ofall genes involved in a process may require com-pletion of a variety of rather subtle experimentalpermutations and variations in cell growth condi-tions. These results underscore the fact that the roleof a particular gene product in a cellular processmay be greatly influenced by the physiological stateof the cell.
Acknowledgement
This work was supported by NSF Grant 0078287 to RW.
References
Anderson RG, Orci L, Brown MS, Garcia-Segura LM, Gold-stein JL. 1983. Ultrastructural analysis of crystalloid endoplas-mic reticulum in UT-1 cells and its disappearance in responseto cholesterol. J Cell Sci 63: 1–20.
Barth H, Thumm M. 2001. A genomic screen identifies AUT8 asa novel gene essential for autophagy in the yeast Saccharomycescerevisiae. Gene 274: 151–156.
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.

892 R. Wright et al.
Bianchi MM, Ngo S, Vandenbol M, et al. 2001. Large-scalephenotypic analysis reveals identical contributions to cellfunctions of known and unknown yeast genes. Yeast 18:1397–1412.
Brown MS, Goldstein JL. 1980. Multivalent feedback regulationof HMG CoA reductase, a control mechanism coordinatingisoprenoid synthesis and cell growth. J Lipid Res 21: 505–517.
Donald KA, Hampton RY, Fritz IB. 1997. Effects of overproduc-tion of the catalytic domain of 3-hydroxy-3-methylglutaryl co-enzyme A reductase on squalene synthesis in Saccharomycescerevisiae. Appl Environ Microbiol 63: 3341–3344.
Giaever G, Chu AM, Ni L, et al. 2002. Functional profiling of theSaccharomyces cerevisiae genome. Nature 418: 387–391.
Giaever G, Shoemaker DD, Jones TW, et al. 1999. Genomicprofiling of drug sensitivities via induced haploinsufficiency.Nature Genet 21: 278–283.
Goldstein JL, Brown MS. 1990. Regulation of the mevalonatepathway. Nature 343: 425–430.
Hampton RY, Koning A, Wright R, Rine J. 1996. In vivoexamination of membrane protein localization and degradationwith GFP. Proc Natl Acad Sci USA 93: 828–833.
Johnston M, Davis RW. 1984. Sequences that regulate the diver-gent GAL1–GAL10 promoter in Saccharomyces cerevisiae. MolCell Biol 4: 1440–1448.
Koning AJ, Larson LL, Cadera EJ, Parrish ML, Wright RL.2002. Mutations that affect vacuole biogenesis also inhibitproliferation of the endoplasmic reticulum in Saccharomycescerevisiae. Genetics (in press).
Koning AJ, Roberts CJ, Wright RL. 1996. Different subcellularlocalization of Saccharomyces cerevisiae HMG-CoA reductaseisozymes at elevated levels corresponds to distinct endoplasmicreticulum membrane proliferations. Mol Biol Cell 7: 769–789.
Lorenz RT, Parks LW. 1987. Regulation of ergosterol biosynthesisand sterol uptake in a sterol-auxotrophic mutant. J Bacteriol 169:3707–3711.
Lum PY, Wright R. 1995. Degradation of HMG Co-A reductase-induced membranes in the fission yeast, Schizosaccharomycespombe. J Cell Biol 131: 81–94.
Muhlrad D, Hunter R, Parker R. 1992. A rapid method for local-ized mutagenesis of yeast genes. Yeast 8: 79–82.
Orr-Weaver TL, Nicolas A, Szostak JW. 1988. Gene conversionadjacent to regions of double-strand break repair. Mol Cell Biol8: 5292–5298.
Oshiro G, Wodicka LM, Washburn MP, et al. 2002. Parallelidentification of new genes in Saccharomyces cerevisiae.Genome Res 12: 1210–1220.
Parrish ML, Sengstag C, Rine JD, Wright RL. 1995. Identificationof the sequences in HMG-CoA reductase required for karmellaeassembly. Mol Biol Cell 6: 1535–1547.
Profant DA, Roberts CJ, Koning AJ, Wright RL. 1999. The roleof the 3-hydroxy 3-methylglutaryl co-enzyme A reductasecytosolic domain in karmellae biogenesis. Mol Biol Cell 10:3409–3423.
Profant DA, Roberts CJ, Wright RL. 2000. Mutational analysis ofthe karmellae-inducing signal in Hmg1p, a yeast HMG-CoAreductase isozyme. Yeast 16: 811–827.
Winzeler EA, Davis RW. 1997. Functional analysis of the yeastgenome. Curr Opin Genet Dev 7: 771–776.
Winzeler EA, Liang H, Shoemaker DD, Davis RW. 2000. Func-tional analysis of the yeast genome by precise deletion andparallel phenotypic characterization. Novartis Found Symp 229:105–111.
Winzeler EA, Shoemaker DD, Astromoff A, et al. 1999. Func-tional characterization of the S. cerevisiae genome by genedeletion and parallel analysis. Science 285: 901–906.
Wright R. 1993. Insights from inducible membranes. Curr Biol 3:870–873.
Wright R, Basson M, D’Ari L, Rine J. 1988. Increased amountsof HMG-CoA reductase induce ‘karmellae’: a proliferation ofstacked membrane pairs surrounding the yeast nucleus. J CellBiol 107: 101–114.
Wright R, Keller G, Gould SJ, Subramani S, Rine J. 1990. Cell-type control of membrane biogenesis induced by HMG-CoAreductase. New Biol 2: 915–921.
Copyright 2003 John Wiley & Sons, Ltd. Yeast 2003; 20: 881–892.
Related Documents











