of March 6, 2018. This information is current as Activation during Apoptosis but Not for Inflammasome Pannexin-1 Is Required for ATP Release David Hackos and Vishva M. Dixit Gilmour, Salina Louie, James E. Cupp, George R. Dubyak, Yan Qu, Shahram Misaghi, Kim Newton, Laurie L. ol.1100478 http://www.jimmunol.org/content/early/2011/04/20/jimmun published online 20 April 2011 J Immunol average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2011 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on March 6, 2018 http://www.jimmunol.org/ Downloaded from by guest on March 6, 2018 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of March 6, 2018.This information is current as
Activationduring Apoptosis but Not for Inflammasome Pannexin-1 Is Required for ATP Release
David Hackos and Vishva M. DixitGilmour, Salina Louie, James E. Cupp, George R. Dubyak, Yan Qu, Shahram Misaghi, Kim Newton, Laurie L.
ol.1100478http://www.jimmunol.org/content/early/2011/04/20/jimmun
published online 20 April 2011J Immunol
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Pannexin-1 Is Required for ATP Release during Apoptosis butNot for Inflammasome Activation
Yan Qu,* Shahram Misaghi,† Kim Newton,* Laurie L. Gilmour,‡ Salina Louie,*
James E. Cupp,‡ George R. Dubyak,x David Hackos,{ and Vishva M. Dixit*
Apoptotic cell death is important for embryonic development, immune cell homeostasis, and pathogen elimination. Innate immune
cells also undergo a very rapid form of cell death termed pyroptosis after activating the protease caspase-1. The hemichannel
pannexin-1 has been implicated in both processes. In this study, we describe the characterization of pannexin-1–deficient mice.
LPS-primed bone marrow-derived macrophages lacking pannexin-1 activated caspase-1 and secreted its substrates IL-1b and IL-
18 normally after stimulation with ATP, nigericin, alum, silica, flagellin, or cytoplasmic DNA, indicating that pannexin-1 is
dispensable for assembly of caspase-1–activating inflammasome complexes. Instead, thymocytes lacking pannexin-1, but not
the P2X7R purinergic receptor, were defective in their uptake of the nucleic acid dye YO-PRO-1 during early apoptosis. Cell
death was not delayed but, unlike their wild-type counterparts, Panx12/2 thymocytes failed to recruit wild-type peritoneal
macrophages in a Transwell migration assay. These data are consistent with pannexin-1 liberating ATP and other yet to be
defined “find me” signals necessary for macrophage recruitment to apoptotic cells. The Journal of Immunology, 2011, 186: 000–
000.
Several members of the NOD-like receptor (NLR) family ofintracellular adapter proteins expressed in innate immunecells assemble inflammasome complexes for caspase-1 ac-
tivation upon detecting pathogen-associated molecular patterns ordamage-associated molecular patterns (DAMPs) (1, 2). NLRC4(also known as Ipaf) activates caspase-1 in response to bacterialflagellin or infection with bacteria such as Salmonella typhi-murium, Legionella pneumophila, and Shigella flexneri (3–6).NLRP3 (also called cryopyrin and NALP3) activates caspase-1 inresponse to diverse stimuli, which include the DAMPs ATP anduric acid (7, 8), but the mechanism of NLRP3 activation remainsunclear. The hemichannel protein pannexin-1 was suggested toform large, nonselective pores after ATP stimulation of P2X7R,thereby allowing DAMPs and pathogen-associated molecular pat-terns access to cytosolic NLRP3 (9, 10). Pannexin-1–dependentpore formation also has been suggested as a mechanism for IL-1bsecretion after cleavage of pro–IL-1b by caspase-1 (11–17), as IL-1b lacks signal sequences for export via the classical endoplasmicreticulum/Golgi-dependent secretory pathway (18, 19).Pannexin-1 is one of three vertebrate pannexins that show ho-
mology to gap junction-forming invertebrate innexins (20, 21).
Pannexin-1 is expressed broadly, whereas pannexin-2 is expressedlargely in brain, and pannexin-3 is in skin and connective tissues(22). Pannexins and connexins have four transmembrane domains,cytosolic N and C termini, and two extracellular loops (22). De-spite their structural similarities, these two protein families seemto have distinct biological functions. Gap junction channelsformed by connexins mediate metabolic and electrical coupling ofadjacent cells, whereas the sugar moiety on pannexin-1 preventsalignment and docking of adjacent pannexin hemichannels to formgap junctions (23). Thus, evidence for pannexin-1 functioning asa physiologically relevant gap junction channel in vivo is lacking(24). Recent studies suggest that pannexin-1 in brain may con-duct small molecules up to ∼1 kDa, such as ions, ATP, inositoltriphosphate, and amino acids (25, 26). In cultured astrocytes,pannexin-1 was reported to mediate ATP release, and in RBCs, ithas been implicated in the secretion of arachidonic acid andits metabolites (26, 27). A recent study targeting pannexin-1 withpharmacological inhibitors or small interfering RNAs identifiedpannexin-1 as the conduit for ATP release from apoptotic cells(28). Cleavage of pannexin-1 by the executioner caspases 3 and 7appeared to trigger its channel activity, and this was necessary forthe recruitment of phagocytic cells (28, 29). Pannexin-1 was notrequired, however, in macrophages migrating in response to ATP(30).We generated mice lacking pannexin-1 by gene targeting to
explore the contribution of pannexin-1 to inflammasome function.We found that neither caspase-1 activation nor IL-1b secretionmediated by either NLRC4 or NLRP3 was compromised bypannexin-1 loss. Instead, Panx12/2 cells were less permeable thantheir wild-type (WT) counterparts to the nucleic acid dye YO-PRO-1 after apoptosis induction, and the dying Panx12/2 cellswere less effective at recruiting macrophages in a Transwellassay. Our data support previous studies (28, 30) indicating thatpannexin-1 releases “find me” signals from apoptotic cells in needof clearance by phagocytic cells but is dispensable in the mi-grating phagocyte itself. Contrary to previous studies (9, 10), weprovide genetic evidence that pannexin-1 and P2X7R function indistinct signaling pathways.
*Physiological Chemistry Department, Genentech, Inc., South San Francisco, CA94080; †Early Stage Cell Culture Department, Genentech, Inc., South San Francisco,CA 94080; ‡Immunology Department, Genentech, Inc., South San Francisco, CA94080; xDepartment of Physiology and Biophysics, Case Western Reserve UniversitySchool of Medicine, Cleveland, OH 44120; and {Neuroscience Department, Genen-tech, Inc., South San Francisco, CA 94080
Received for publication February 14, 2011. Accepted for publication March 25,2011.
Address correspondence and reprint requests to Dr. Vishva M. Dixit or Dr. DavidHackos, Physiological Chemistry Department, Genentech, Inc., 1 DNA Way, SouthSan Francisco, CA 94080 (V.M.D.) or Neuroscience Department, Genentech, Inc., 1DNA Way, South San Francisco, CA 94080 (D.H.). E-mail addresses: [email protected] (V.M.D.) and [email protected] (D.H.)
Abbreviations used in this article: BMDM, bone marrow-derived macrophage; CHX,cycloheximide; DAMP, damage-associated molecular pattern; NLR, NOD-like re-ceptor; PI, propidium iodide; WT, wild-type.
Copyright� 2011 by The American Association of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1100478
Published April 20, 2011, doi:10.4049/jimmunol.1100478 by guest on M
arch 6, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsReagents
Key reagents and their sources were as follows: Escherichia coli LPSserotype O1101:B4 (List Biological Laboratories); ATP, cycloheximide(CHX), dexamethasone, and nigericin (Sigma); z-VAD-fmk (MBL); Imjectalum (Thermo Scientific); silica (US Silica); S. typhimurium flagellin(InvivoGen); Lipofectamine 2000, pcDNA3.1(+), Pacific blue annexin V,and YO-PRO-1 (Invitrogen); complete protease inhibitor mixture andDOTAP (Roche); Jo2 hamster anti-mouse CD95 and propidium iodide (PI;BD); cleaved (Asp175) and total (8G10) caspase-3 Abs (Cell Signaling);GAPDH, caspase-1 p10, and actin Abs (Santa Cruz); 4B4 caspase-1 p20Ab (Genentech); and 3ZD anti–IL-1b and pro–IL-1b Ab (BiologicalResources Branch, National Cancer Institute). S. enerica serovar typhi-murium strain SL1344 was from D. Monack (Stanford University).
Mice
The strategy used to generate Panx12/2 mice is depicted in Fig. 1A. Ho-mologous recombinant C57BL/6 C2 embryonic stem cells were deletedof the neomycin selection cassette with adenovirus-expressed FLP re-combinase. Deletion of exon 2, which generates a frameshift and pre-mature stop codon, was achieved by breeding to the C57BL/6-Gt(ROSA)26Sortm16(Cre)Arte Cre deleter strain (TaconicArtemis). Mice were geno-typed with PCR primers (59-TGA CCA CAG ACA GCA CTTAAG-39 and59-CGT CTG AGA GCT CCC TGG CG-39) that yield a 651-bp WT DNAfragment and a 335-bp knockout DNA fragment (Fig. 1B). P2rx72/2 micewere generated by Lexicon Pharmaceuticals. The Genentech InstitutionalAnimal Care and Use Committee approved all experiments.
Cells
Bone marrow and thioglycolate-elicited peritoneal macrophages werecultured in DMEM supplemented with 20% L cell-conditioned medium,
10% heat-inactivated FBS, 100 U/ml penicillin, 10 mM L-glutamine, and100 mg/ml streptomycin. Adherent bone marrow-derived macrophages(BMDMs) were obtained after 5–7 d and replated in 6- or 24-well platesfor experiments. Thymocytes were cultured in RPMI 1640 media sup-plemented with 10% heat-inactivated FBS, 100 U/ml penicillin, 100 mg/mlstreptomycin, and 10 mM L-glutamine.
Western blots and ELISAs
BMDMs were routinely seeded in 6-well plates (2 3 106 cells/well) forWestern blot analysis and in 24-well plates for ELISA. Cell supernatantswere concentrated by TCA precipitation for Western blotting (31), andcells were lysed in 150 ml RIPA lysis buffer (1% Nonidet P-40, 0.5%sodium deoxycholate, 0.1% SDS, pH 7.4, in PBS) containing a proteaseinhibitor mixture. Mouse tissues were homogenized in KINEXUS buffer(20 mM HEPES pH 7.2, 2 mM EGTA, 5 mM EDTA, 30 mM NaF, 60 mMb-glycerolphosphate, 20 mM sodium pyrophosphate, 1 mM Na3VO4, and1% Triton X-100) supplemented with protease and Halt Phosphataseinhibitor cocktails (Thermo Scientific Pierce). Total protein concentra-tion was determined by bicinchoninic acid protein assay (Thermo Scien-tific Pierce). IL-1b and IL-18 in culture supernatants were measured byBioplex-23 assay (Bio-Rad).
ATP measurements
Culture supernatants were heated at 98˚C for 5 min and then assayed witha CellTiter-Glo kit (Promega) in an EnVison Multilabel Reader (Perkin-Elmer).
FACS analysis
Thymocytes were washed twice with cold PBS and then stained with Pacificblue-conjugated annexin V, YO-PRO-1, and PI for 30 min at room tem-perature. Cells were analyzed in an LSR-II cytometer (BD).
FIGURE 1. Generation of Panx12/2 mice. A, Strategy for deleting exon 2 of Panx1 in C57BL/6 C2 embryonic stem cells. B, Identification of Panx12/2
mice by PCR. C, Western blot analysis of pannexin-1 protein expression in mouse tissues (70 mg total protein/lane). Variable pannexin-1 migration likely
represents variable glycosylation of the protein. GAPDH is presented to indicate protein loading.
2 PANNEXIN-1 IS DISPENSABLE FOR INFLAMMASOME ACTIVATION
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Migration assay
Thymocytes (2 3 106/ml) were plated on 12-well dishes and left untreatedor stimulated with 0.5 mM dexamethasone in the absence or presence of 25mM zVAD for 4 h. Culture supernatants were collected by centrifugationtwice at 500 3 g for 5 min. Transwell migration assays were performedwith 500 ml apoptotic cell supernatant in the lower chamber and 250 ml ofWT or Panx12/2 thioglycolate-elicited peritoneal macrophages (2 3 106/ml) in the upper chamber/insert of a 24-well plate with 3-mM pore-sizemembrane (Corning) for 90 min at 37˚C. Cells that migrated into the lowerchamber were counted with a hemocytometer. Where indicated, culturesupernatants were supplemented with 1 U/ml apyrase for 5 min at roomtemperature. Separately, 500 ml of the supernatants from these migrationassays were boiled at 98˚C for 5 min prior to ATP measurement.
Cell viability assay
Thymocytes (1 3 106/ml) were plated on 96-well dishes and left untreatedor stimulated with dexamethasone in the absence or presence of zVADfor 4, 24, 30, and 48 h. Cell viability was measured with a CellTiter-GloLuminescent Cell Viability Assay Kit (Promega).
YO-PRO-1 influx
BMDMs (1 3 106/ml) plated on PDL-coated, black 96-well plates werewashed once with PBS and cultured in the following media: 135 mM
NaCl, 5 mM KCl, 10 mM glucose, 10 mM HEPES pH 7.4, 1 mM CaCl2,and 1 mM MgCl2. After addition of YO-PRO-1 (5 mM) and ATP (3 mM),cells were imaged using a 320 Plan Fluor ELWD objective lens andimages collected every 60 s for 80 min. Fluorescence intensity wasrecorded and calculated using Nikon’s NIS Elements software, whichenables regions-of-interest selection and automatic tracking of 20–25 nu-clei per image at each time point throughout the recording time.
ResultsPannexin-1 is dispensable for caspase-1 activation by ATP ornigericin
Panx12/2 mice generated by gene targeting (Fig. 1A, 1B) wereconfirmed to lack pannexin-1 with a polyclonal Ab against theprotein’s C terminus (23). Western blotting of WT tissues revealedseveral pannexin-1 bands between ∼41 and 48 kDa, but thesewere absent in Panx12/2 tissues (Fig. 1C). Pannexin-1 was de-tected readily in kidney, lung, muscle, spleen, brain, and thymus.BMDMs also expressed pannexin-1 but at a much lower level.Prolonged stimulation of the purinergic receptor P2X7R with
ATP is proposed to trigger pannexin-1 pore formation for extra-cellular bacterial products to enter the cell and promote NLRP3-
FIGURE 2. Pannexin-1 is not required for P2X7R-dependent activation of the NLRP3 inflammasome. WT (Panx1+/+), Panx12/2, or P2rx72/2 BMDMs
were primed with 0.5 mg/ml LPS for 4 h and then treated with 5 mM ATP or 10 mM nigericin for the time indicated. Culture supernatants were Western
blotted for IL-1b and the processed caspase-1 subunits p10 and p20. Cell lysates were Western blotted for p45 procaspase-1 (A–C). IL-1b (D) and IL-18 (E)
secreted into the culture supernatant are presented as the mean 6 SD of triplicate samples. Results are representative of three independent experiments.
The Journal of Immunology 3
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

dependent activation of caspase-1 (10, 14–17). We tested thisnotion with WT, Panx12/2, and P2rx72/2 BMDMs (Fig. 2). Thecells were primed with LPS for 4 h and then stimulated with eitherATP or the ionophore nigericin to activate caspase-1. Both WTand Panx12/2 BMDMs cleaved procaspase-1 (p45) into its p10and p20 subunits after 15 min of ATP stimulation (Fig. 2A, 2B),whereas P2rx72/2 BMDMs did not cleave procaspase-1 even after1 h of ATP stimulation (Fig. 2C). Panx12/2 BMDMs also pro-
cessed procaspase-1 in response to nigericin, which triggerscaspase-1 cleavage independently of the P2X7R (Fig. 2A–C).Further confirmation of caspase-1 activation in Panx12/2 BMDMswas the normal processing (Fig. 2A–C) and secretion (Fig. 2D) ofbiologically active IL-1b (p17). Caspase-1–dependent secretionof IL-18 also occurred normally in Panx12/2 BMDMs (Fig. 2E).These data prove that pannexin-1 is dispensable for NLRP3-dependent activation of caspase-1 and secretion of the proin-
FIGURE 3. Pannexin-1 is not required for NLRP3-, NLRC4-, or AIM2-dependent caspase-1 activation and cytokine release. WT (Panx1+/+) (A, C) and
Panx12/2 (B, D) BMDMs were primed with 0.5 mg/ml LPS for 2 h and then treated with 5 mM ATP for 30 min, 500 ng/ml alum, 125 ng/ml silica, 2.5 mg/
ml flagellin, 1 mg/ml pcDNA3.1, or Salmonella (multiplicity of infection = 20) for 6 h. Culture supernatants were Western blotted for the processed caspase-
1 subunits p10 and p20. Cell lysates were Western blotted for p45 procaspase-1 (A–D). IL-1b (E), and IL-18 (F) secreted into the culture supernatant are
shown as the mean 6 SD of triplicate samples. Results are representative of three independent experiments.
4 PANNEXIN-1 IS DISPENSABLE FOR INFLAMMASOME ACTIVATION
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
flammatory cytokines IL-1b and IL-18 in response to LPS/ATP orLPS/nigericin.
Pannexin-1 is not required for NLRP3-, NLRC4-, orAIM2-dependent caspase-1 activation
Next, we determined whether pannexin-1 might contribute tocaspase-1 activation by other stimuli. NLRP3-dependent caspase-1activation in response to alum or silica (32, 33), NLRC4-dependentcaspase-1 activation in response to S. typhimurium or transfectedflagellin (3, 6, 32), and AIM2-dependent caspase-1 activation inresponse to transfected plasmid DNA (34–36) occurred normallyin Panx12/2 BMDMs (Fig. 3A–D). Consistent with these data,WT and Panx12/2 BMDMs secreted equivalent amounts of IL-1band IL-18 (Fig. 3E, 3F). Our results indicate that pannexin-1 is
dispensable for caspase-1 activation by distinct inflammasomecomplexes and the subsequent secretion of IL-1b and IL-18.
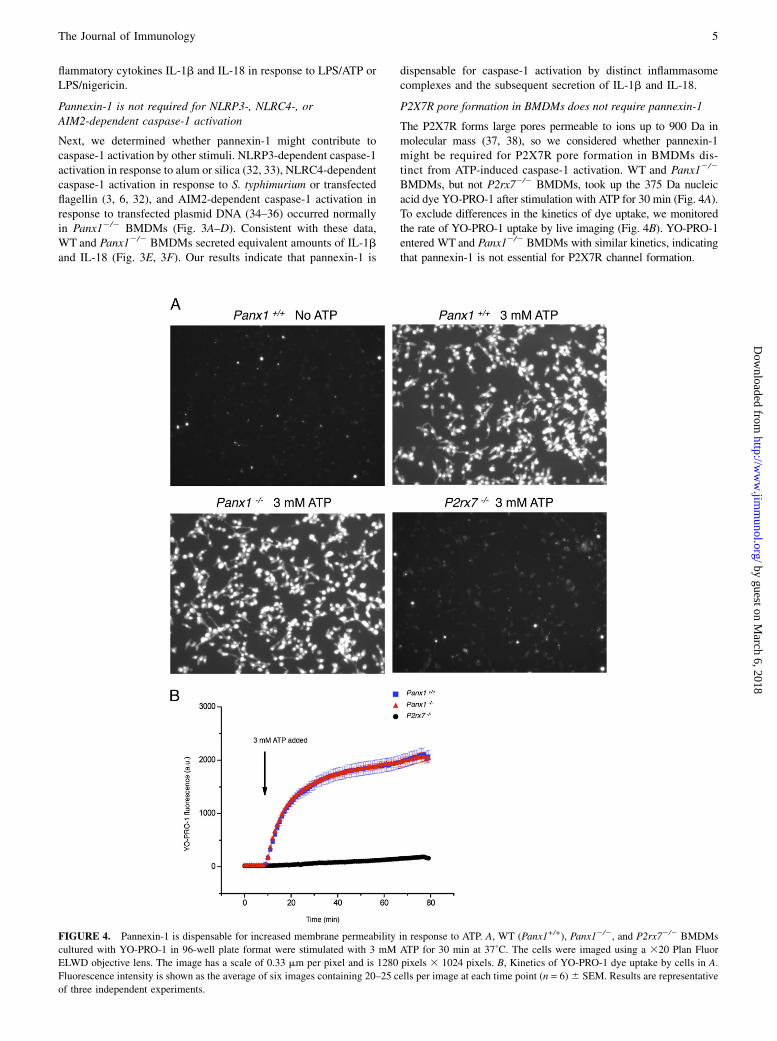
P2X7R pore formation in BMDMs does not require pannexin-1
The P2X7R forms large pores permeable to ions up to 900 Da inmolecular mass (37, 38), so we considered whether pannexin-1might be required for P2X7R pore formation in BMDMs dis-tinct from ATP-induced caspase-1 activation. WT and Panx12/2
BMDMs, but not P2rx72/2 BMDMs, took up the 375 Da nucleicacid dye YO-PRO-1 after stimulation with ATP for 30 min (Fig. 4A).To exclude differences in the kinetics of dye uptake, we monitoredthe rate of YO-PRO-1 uptake by live imaging (Fig. 4B). YO-PRO-1entered WT and Panx12/2 BMDMs with similar kinetics, indicatingthat pannexin-1 is not essential for P2X7R channel formation.
FIGURE 4. Pannexin-1 is dispensable for increased membrane permeability in response to ATP. A, WT (Panx1+/+), Panx12/2, and P2rx72/2 BMDMs
cultured with YO-PRO-1 in 96-well plate format were stimulated with 3 mM ATP for 30 min at 37˚C. The cells were imaged using a 320 Plan Fluor
ELWD objective lens. The image has a scale of 0.33 mm per pixel and is 1280 pixels 3 1024 pixels. B, Kinetics of YO-PRO-1 dye uptake by cells in A.
Fluorescence intensity is shown as the average of six images containing 20–25 cells per image at each time point (n = 6)6 SEM. Results are representative
of three independent experiments.
The Journal of Immunology 5
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Pannexin-1 mediates YO-PRO-1 uptake and ATP release inapoptotic cells
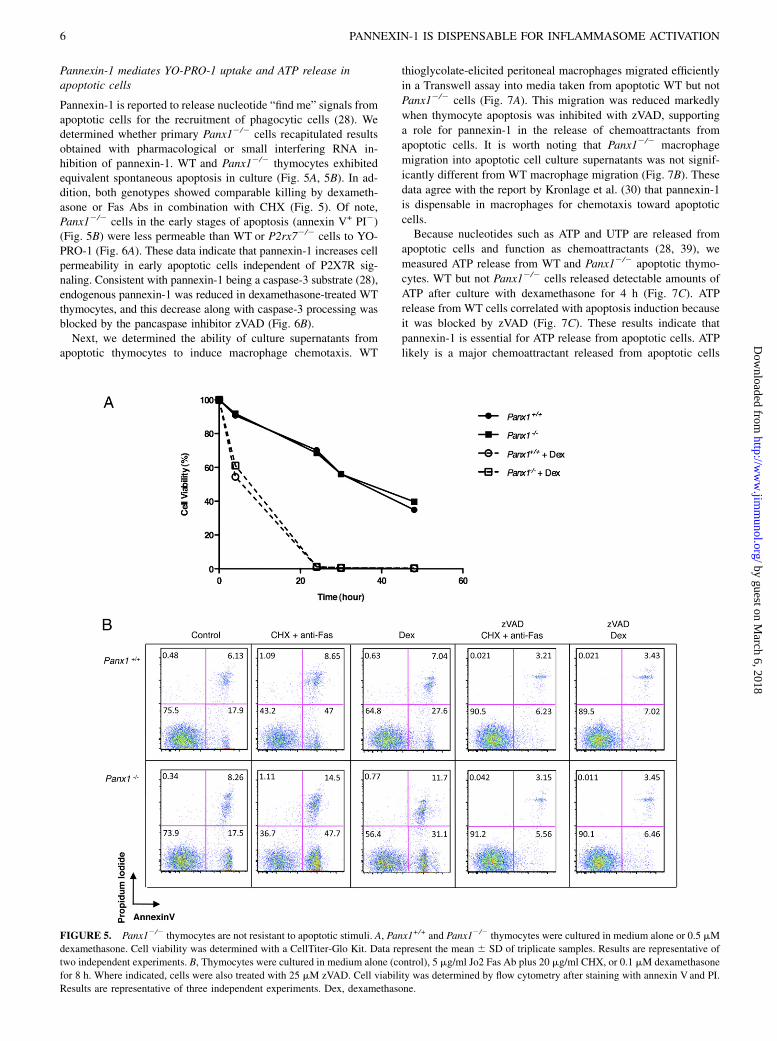
Pannexin-1 is reported to release nucleotide “find me” signals fromapoptotic cells for the recruitment of phagocytic cells (28). Wedetermined whether primary Panx12/2 cells recapitulated resultsobtained with pharmacological or small interfering RNA in-hibition of pannexin-1. WT and Panx12/2 thymocytes exhibitedequivalent spontaneous apoptosis in culture (Fig. 5A, 5B). In ad-dition, both genotypes showed comparable killing by dexameth-asone or Fas Abs in combination with CHX (Fig. 5). Of note,Panx12/2 cells in the early stages of apoptosis (annexin V+ PI2)(Fig. 5B) were less permeable than WT or P2rx72/2 cells to YO-PRO-1 (Fig. 6A). These data indicate that pannexin-1 increases cellpermeability in early apoptotic cells independent of P2X7R sig-naling. Consistent with pannexin-1 being a caspase-3 substrate (28),endogenous pannexin-1 was reduced in dexamethasone-treated WTthymocytes, and this decrease along with caspase-3 processing wasblocked by the pancaspase inhibitor zVAD (Fig. 6B).Next, we determined the ability of culture supernatants from
apoptotic thymocytes to induce macrophage chemotaxis. WT
thioglycolate-elicited peritoneal macrophages migrated efficiently
in a Transwell assay into media taken from apoptotic WT but not
Panx12/2 cells (Fig. 7A). This migration was reduced markedly
when thymocyte apoptosis was inhibited with zVAD, supporting
a role for pannexin-1 in the release of chemoattractants from
apoptotic cells. It is worth noting that Panx12/2 macrophage
migration into apoptotic cell culture supernatants was not signif-
icantly different from WT macrophage migration (Fig. 7B). These
data agree with the report by Kronlage et al. (30) that pannexin-1
is dispensable in macrophages for chemotaxis toward apoptotic
cells.Because nucleotides such as ATP and UTP are released from
apoptotic cells and function as chemoattractants (28, 39), we
measured ATP release from WT and Panx12/2 apoptotic thymo-
cytes. WT but not Panx12/2 cells released detectable amounts of
ATP after culture with dexamethasone for 4 h (Fig. 7C). ATP
release from WT cells correlated with apoptosis induction because
it was blocked by zVAD (Fig. 7C). These results indicate that
pannexin-1 is essential for ATP release from apoptotic cells. ATP
likely is a major chemoattractant released from apoptotic cells
FIGURE 5. Panx12/2 thymocytes are not resistant to apoptotic stimuli. A, Panx1+/+ and Panx12/2 thymocytes were cultured in medium alone or 0.5 mM
dexamethasone. Cell viability was determined with a CellTiter-Glo Kit. Data represent the mean 6 SD of triplicate samples. Results are representative of
two independent experiments. B, Thymocytes were cultured in medium alone (control), 5 mg/ml Jo2 Fas Ab plus 20 mg/ml CHX, or 0.1 mM dexamethasone
for 8 h. Where indicated, cells were also treated with 25 mM zVAD. Cell viability was determined by flow cytometry after staining with annexin V and PI.
Results are representative of three independent experiments. Dex, dexamethasone.
6 PANNEXIN-1 IS DISPENSABLE FOR INFLAMMASOME ACTIVATION
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

because ATP depletion from apoptotic thymocyte culture super-natants with apyrase reduced macrophage migration to that seen incontrol supernatants (Fig. 7A, 7B).
DiscussionPannexin-1 in immune cells has been linked to inflammasome as-sembly and caspase-1 activation (9, 10), secretion of proinflam-matory cytokines (16, 40), P2X7R-dependent dye uptake, and che-
moattractant release during apoptosis (17, 28, 39). Our analyses of
Panx12/2 BMDMs indicate that pannexin-1 is not required for
NLRP3-, NLRC4-, or AIM2-dependent caspase-1 activation, nor is
it needed for caspase-1–dependent secretion of the proinflammatory
cytokines IL-1b and IL-18 (Figs. 2, 3). We also demonstrate that
ATP engagement of the P2X7R increases BMDM membrane per-
meability independent of pannexin-1 (Fig. 4). Consistent with
Chekeni et al. (28), our data reveal that pannexin-1 regulates
FIGURE 6. Pannexin-1 is required for YO-PRO-1 dye uptake during early apoptosis. A, WT, Panx12/2, and P2rx72/2 thymocytes were cultured with 5
mg/ml Jo2 Fas Ab plus 20 mg/ml CHX or 0.5 mM dexamethasone for 8 h. YO-PRO-1 uptake by annexin V+, PI2 apoptotic cells was determined by flow
cytometry. White histograms represent cells cultured with DMSO carrier, whereas gray histograms represent cells cultured with 25 mM zVAD. B, WT
(Panx1+/+) and Panx12/2 thymocytes were treated with dexamethasone and/or 25 mM zVAD for 18 h. Dex, dexamethasone.
The Journal of Immunology 7
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

membrane permeability in early apoptosis (Fig. 6) and the releaseof macrophage chemoattractants such as ATP (Fig. 7). Therefore,P2X7R and pannexin-1 function in distinct signaling pathways.Prior studies concluding pannexin-1 involvement in inflamma-
some activation and cytokine release in BMDMs (9, 10, 16, 17)relied in large part on mimetic blocking peptides, pharmacologicalinhibitors, and/or overexpression approaches in transformed celllines. Possible off-target activities (24) associated with these ex-perimental approaches and the use of transformed cells may ac-
count for the discrepancy with our genetic evidence. Our data thatpannexin-1 regulates the release of chemoattractants for phago-cytic cells during apoptosis while being dispensable for inflam-masome activation is consistent with the notion that apoptosisis an immunologically silent, anti-inflammatory mode of celldeath (41). Panx12/2 mice are viable and fertile indicating thatpannexin-1 is not essential for mouse development. MonitoringPanx12/2 mice from birth to adulthood for 12 mo did not revealany gross defects. Histological and pathology examinations of
FIGURE 7. Release of chemoattractants from apoptotic thymocytes requires pannexin-1. WT (Panx1+/+) and Panx12/2 thymocytes were treated with 0.5
mM dexamethasone in the absence or presence of 25 mM zVAD for 4 h. Culture supernatants were used in Transwell migration assays with thioglycolate-
elicited peritoneal macrophages from WT (A) or Panx12/2 (B) mice. Where indicated, 1 U/ml apyrase was added to culture supernatants for 5 min before
the migration assay. The ATP content in the culture supernatants is shown in C. Data represent the mean 6 SD of triplicate samples. Results are rep-
resentative of three independent experiments. All p values were determined by t test using independent variants.
8 PANNEXIN-1 IS DISPENSABLE FOR INFLAMMASOME ACTIVATION
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

major organs were also normal. Whether clearance of apoptoticcell populations during development or pathogen challenge isdelayed remains to be determined.
DisclosuresY.Q., S.M., K.N., L.L.G., S.L., J.E.C., D.H., and V.M.D. are employees of
Genentech, Inc.
References1. Ting, J. P., and B. K. Davis. 2005. CATERPILLER: a novel gene family im-
portant in immunity, cell death, and diseases. Annu. Rev. Immunol. 23: 387–414.2. Schroder, K., and J. Tschopp. 2010. The inflammasomes. Cell 140: 821–832.3. Mariathasan, S., K. Newton, D. M. Monack, D. Vucic, D. M. French, W. P. Lee,
M. Roose-Girma, S. Erickson, and V. M. Dixit. 2004. Differential activation ofthe inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 430: 213–218.
4. Amer, A., L. Franchi, T. D. Kanneganti, M. Body-Malapel, N. Ozoren, G. Brady,S. Meshinchi, R. Jagirdar, A. Gewirtz, S. Akira, and G. Nunez. 2006. Regulationof Legionella phagosome maturation and infection through flagellin and hostIpaf. J. Biol. Chem. 281: 35217–35223.
5. Miao, E. A., C. M. Alpuche-Aranda, M. Dors, A. E. Clark, M. W. Bader,S. I. Miller, and A. Aderem. 2006. Cytoplasmic flagellin activates caspase-1 andsecretion of interleukin 1beta via Ipaf. Nat. Immunol. 7: 569–575.
6. Franchi, L., A. Amer, M. Body-Malapel, T. D. Kanneganti, N. Ozoren,R. Jagirdar, N. Inohara, P. Vandenabeele, J. Bertin, A. Coyle, et al. 2006. Cy-tosolic flagellin requires Ipaf for activation of caspase-1 and interleukin 1beta insalmonella-infected macrophages. Nat. Immunol. 7: 576–582.
7. Mariathasan, S., D. S. Weiss, K. Newton, J. McBride, K. O’Rourke, M. Roose-Girma, W. P. Lee, Y. Weinrauch, D. M. Monack, and V. M. Dixit. 2006. Cry-opyrin activates the inflammasome in response to toxins and ATP. Nature 440:228–232.
8. Sutterwala, F. S., Y. Ogura, M. Szczepanik, M. Lara-Tejero, G. S. Lichtenberger,E. P. Grant, J. Bertin, A. J. Coyle, J. E. Galan, P. W. Askenase, and R. A. Flavell.2006. Critical role for NALP3/CIAS1/Cryopyrin in innate and adaptive immu-nity through its regulation of caspase-1. Immunity 24: 317–327.
9. Marina-Garcıa, N., L. Franchi, Y. G. Kim, D. Miller, C. McDonald, G. J. Boons,and G. Nunez. 2008. Pannexin-1-mediated intracellular delivery of muramyldipeptide induces caspase-1 activation via cryopyrin/NLRP3 independently ofNod2. J. Immunol. 180: 4050–4057.
10. Kanneganti, T. D., M. Lamkanfi, Y. G. Kim, G. Chen, J. H. Park, L. Franchi,P. Vandenabeele, and G. Nunez. 2007. Pannexin-1-mediated recognition ofbacterial molecules activates the cryopyrin inflammasome independent of Toll-like receptor signaling. Immunity 26: 433–443.
11. MacKenzie, A., H. L. Wilson, E. Kiss-Toth, S. K. Dower, R. A. North, andA. Surprenant. 2001. Rapid secretion of interleukin-1beta by microvesicleshedding. Immunity 15: 825–835.
12. Qu, Y., L. Franchi, G. Nunez, and G. R. Dubyak. 2007. Nonclassical IL-1 betasecretion stimulated by P2X7 receptors is dependent on inflammasome activa-tion and correlated with exosome release in murine macrophages. J. Immunol.179: 1913–1925.
13. Perregaux, D., and C. A. Gabel. 1994. Interleukin-1 beta maturation and releasein response to ATP and nigericin. Evidence that potassium depletion mediated bythese agents is a necessary and common feature of their activity. J. Biol. Chem.269: 15195–15203.
14. Surprenant, A., F. Rassendren, E. Kawashima, R. A. North, and G. Buell. 1996.The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor(P2X7). Science 272: 735–738.
15. Locovei, S., E. Scemes, F. Qiu, D. C. Spray, and G. Dahl. 2007. Pannexin1 is partof the pore forming unit of the P2X(7) receptor death complex. FEBS Lett. 581:483–488.
16. Pelegrin, P., and A. Surprenant. 2007. Pannexin-1 couples to maitotoxin- andnigericin-induced interleukin-1beta release through a dye uptake-independentpathway. J. Biol. Chem. 282: 2386–2394.
17. Pelegrin, P., and A. Surprenant. 2006. Pannexin-1 mediates large pore formationand interleukin-1beta release by the ATP-gated P2X7 receptor. EMBO J. 25:5071–5082.
18. Rubartelli, A., F. Cozzolino, M. Talio, and R. Sitia. 1990. A novel secretorypathway for interleukin-1 beta, a protein lacking a signal sequence. EMBO J. 9:1503–1510.
19. Wewers, M. D. 2004. IL-1beta: an endosomal exit. Proc. Natl. Acad. Sci. USA101: 10241–10242.
20. Bruzzone, R., S. G. Hormuzdi, M. T. Barbe, A. Herb, and H. Monyer. 2003.Pannexins, a family of gap junction proteins expressed in brain. Proc. Natl.Acad. Sci. USA 100: 13644–13649.
21. MacVicar, B. A., and R. J. Thompson. 2010. Non-junction functions ofpannexin-1 channels. Trends Neurosci. 33: 93–102.
22. Shestopalov, V. I., and Y. Panchin. 2008. Pannexins and gap junction proteindiversity. Cell. Mol. Life Sci. 65: 376–394.
23. Penuela, S., R. Bhalla, X. Q. Gong, K. N. Cowan, S. J. Celetti, B. J. Cowan,D. Bai, Q. Shao, and D. W. Laird. 2007. Pannexin 1 and pannexin 3 are gly-coproteins that exhibit many distinct characteristics from the connexin family ofgap junction proteins. J. Cell Sci. 120: 3772–3783.
24. Dubyak, G. R. 2009. Both sides now: multiple interactions of ATP withpannexin-1 hemichannels. Focus on “A permeant regulating its permeation pore:inhibition of pannexin 1 channels by ATP.” Am. J. Physiol. Cell Physiol. 296:C235–C241.
25. Wang, J., M. Ma, S. Locovei, R. W. Keane, and G. Dahl. 2007. Modulation ofmembrane channel currents by gap junction protein mimetic peptides: sizematters. Am. J. Physiol. Cell Physiol. 293: C1112–C1119.
26. Iglesias, R., G. Dahl, F. Qiu, D. C. Spray, and E. Scemes. 2009. Pannexin 1: themolecular substrate of astrocyte “hemichannels.” J. Neurosci. 29: 7092–7097.
27. Jiang, H., A. G. Zhu, M. Mamczur, J. R. Falck, K. M. Lerea, and J. C. McGiff.2007. Stimulation of rat erythrocyte P2X7 receptor induces the release ofepoxyeicosatrienoic acids. Br. J. Pharmacol. 151: 1033–1040.
28. Chekeni, F. B., M. R. Elliott, J. K. Sandilos, S. F. Walk, J. M. Kinchen,E. R. Lazarowski, A. J. Armstrong, S. Penuela, D. W. Laird, G. S. Salvesen, et al.2010. Pannexin 1 channels mediate ‘find-me’ signal release and membranepermeability during apoptosis. Nature 467: 863–867.
29. Idziorek, T., J. Estaquier, F. De Bels, and J. C. Ameisen. 1995. YOPRO-1 per-mits cytofluorometric analysis of programmed cell death (apoptosis) withoutinterfering with cell viability. J. Immunol. Methods 185: 249–258.
30. Kronlage, M., J. Song, L. Sorokin, K. Isfort, T. Schwerdtle, J. Leipziger,B. Robaye, P. B. Conley, H. C. Kim, S. Sargin, et al. 2010. Autocrine purinergicreceptor signaling is essential for macrophage chemotaxis. Sci. Signal. 3: ra55.
31. Qu, Y., L. Ramachandra, S. Mohr, L. Franchi, C. V. Harding, G. Nunez, andG. R. Dubyak. 2009. P2X7 receptor-stimulated secretion of MHC class II-containing exosomes requires the ASC/NLRP3 inflammasome but is in-dependent of caspase-1. J. Immunol. 182: 5052–5062.
32. Eisenbarth, S. C., O. R. Colegio, W. O’Connor, F. S. Sutterwala, andR. A. Flavell. 2008. Crucial role for the Nalp3 inflammasome in the immu-nostimulatory properties of aluminium adjuvants. Nature 453: 1122–1126.
33. Hornung, V., F. Bauernfeind, A. Halle, E. O. Samstad, H. Kono, K. L. Rock,K. A. Fitzgerald, and E. Latz. 2008. Silica crystals and aluminum salts activatethe NALP3 inflammasome through phagosomal destabilization. Nat. Immunol. 9:847–856.
34. Fernandes-Alnemri, T., J. W. Yu, P. Datta, J. Wu, and E. S. Alnemri. 2009. AIM2activates the inflammasome and cell death in response to cytoplasmic DNA.Nature 458: 509–513.
35. Hornung, V., A. Ablasser, M. Charrel-Dennis, F. Bauernfeind, G. Horvath,D. R. Caffrey, E. Latz, and K. A. Fitzgerald. 2009. AIM2 recognizes cytosolicdsDNA and forms a caspase-1-activating inflammasome with ASC. Nature 458:514–518.
36. Burckstummer, T., C. Baumann, S. Bluml, E. Dixit, G. Durnberger, H. Jahn,M. Planyavsky, M. Bilban, J. Colinge, K. L. Bennett, and G. Superti-Furga.2009. An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplas-mic DNA sensor for the inflammasome. Nat. Immunol. 10: 266–272.
37. Virginio, C., A. MacKenzie, F. A. Rassendren, R. A. North, and A. Surprenant.1999. Pore dilation of neuronal P2X receptor channels. Nat. Neurosci. 2: 315–321.
38. Virginio, C., A. MacKenzie, R. A. North, and A. Surprenant. 1999. Kinetics ofcell lysis, dye uptake and permeability changes in cells expressing the rat P2X7receptor. J. Physiol. 519: 335–346.
39. Chekeni, F. B., and K. S. Ravichandran. 2010. The role of nucleotides in apo-ptotic cell clearance: implications for disease pathogenesis. J. Mol. Med. 89: 13–22.
40. Pelegrin, P., C. Barroso-Gutierrez, and A. Surprenant. 2008. P2X7 receptordifferentially couples to distinct release pathways for IL-1beta in mouse mac-rophage. J. Immunol. 180: 7147–7157.
41. Munoz, L. E., K. Lauber, M. Schiller, A. A. Manfredi, and M. Herrmann. 2010.The role of defective clearance of apoptotic cells in systemic autoimmunity. NatRev Rheumatol 6: 280–289.
The Journal of Immunology 9
by guest on March 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents











