Paleoceanographic, and paleoclimatic constraints on the global Eocene diatom and silicoflagellate record John A. Barron a, ⁎, Catherine E. Stickley b , David Bukry a a U.S. Geological Survey, MS910, Menlo Park, CA 94025, USA b Evolution Applied Ltd., 50 Mitchell Way, Upper Rissington, Cheltenham GL54 2PL, UK abstract article info Article history: Received 18 August 2014 Received in revised form 13 January 2015 Accepted 18 January 2015 Available online 24 January 2015 Keywords: Diatom Silicoflagellate Eocene paleoclimate Paleoceanography Sea level Ocean gateway Eocene diatom and silicoflagellate biostratigraphy are summarized and correlated with the most recent geologic time scale as well as with the global oxygen isotope and eustatic sea level curves. The global distribution of Eocene diatom/silicoflagellate-bearing sediments varies considerably, reflecting changing oceanic gateways and paleoceanography with changing patterns that are punctuated by four major depositional events. Event 1 (~49 million years ago, Ma), at the end of the Early Eocene Climatic Optimum (EECO), saw the cessation of diatom/silicoflagellate deposition in epicontinental regions of the North Sea region and in the northern Russia and the onset of biosilica deposition in the Arctic. Event 2 (~46 Ma), which coincided with intensification of the Middle Eocene cooling trend, marked the widespread expansion of diatom/silicoflagellate deposition in both the North and South Atlantic. A shift of diatom/silicoflagellate deposition from the Atlantic to the Pacific began at Event 3, at the end of the Middle Eocene Climatic Optimum (MECO) (~40 Ma), that was likely tied to the initial opening of the Drake Passage between Antarctica and South America. Event 4 (~39 Ma) coincided with a major sea level fall and a widespread deep-sea hiatus in the latest Middle Eocene. Late Eocene diatom/silicoflagellate deposition became more concentrated in middle-to-high latitude regions and coastal upwelling regions, partic- ularly in the Pacific Ocean. Tabulation of the first and last occurrences of 132 biostratigraphically-important diatoms suggests increased species turnover during the latest Paleocene to earliest Eocene that may be in part due to a monographic effect. An increasing rate of evolution of new diatom species between ~46 and 43 Ma and after ~40 Ma coincides respec- tively with the widespread expansion of diatom deposition in the Atlantic and with an increased pole-to-equator thermal gradient that witnessed the expansion of diatoms in high latitude oceans and coastal upwelling settings. Published by Elsevier B.V. 1. Introduction The Eocene (~56 to 34 Ma), the longest and most climatically diverse epoch of the Cenozoic, spans from the warmest period of the Cenozoic, the Early Eocene, through a long-term cooling trend during the Middle and Late Eocene that culminated in the major expansion of the Antarctic Ice Sheet in the earliest Oligocene (Zachos et al., 2001, 2008). Although marine diatoms constitute a major group of phytoplankton responsible for up to 45% of ocean's primary productivity, knowledge of their Eocene history is largely fragmentary and pieced together from numerous, geo- graphically separated and often stratigraphically discontinuous, on-land and deep-sea stratigraphic sections. Diatoms, unicellular heterokont algae, produce a bipartite external skeleton (frustule) composed of opaline silica (hydrated silicon dioxide), which is ornately sculpted and forms the basis of diatom taxonomic clas- sification. The size of diatom frustules ranges from less than a few micrometers (μm) to more than 1000 μm, but most frustules range in size from 10 to 40 μm(Scherer et al., 2007). Diatoms are common virtu- ally everywhere there is water and light, in the ocean, freshwater, soils, and on damp surfaces. As the oceans are typically undersaturated in silica, lightly silicified diatom frustules are typically dissolved after death of the organism during descent to the seafloor. Diatom frustules are preferen- tially preserved in seafloor sediments beneath areas of high surface water production, typically areas of upwelling, where seawater is silica- saturated. Even then, preservation in the older fossil record may be hin- dered by the instability of opaline diatom frustules to survive diagenesis, as diatomaceous silica (opal-A) is converted to amorphous cristobalite and eventually to chert upon deep burial and exposure to temperatures in excess of 50 °C. Where fossil diatoms are preserved, however, they provide a wealth of paleoenvironmental, paleoceanographic and biostrat- igraphic information, particularly in regions where calcareous microfos- sils are in low abundance, e.g., the polar regions. Marine surface waters and sediments containing diatoms typically also contain silicoflagellates. Silicoflagellates are unicellular marine algae that are entirely planktonic. They produce internal tubular skele- tons of opaline silica that are generally 30–100 μm in diameter, and Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100 ⁎ Corresponding author. E-mail addresses: [email protected] (J.A. Barron), [email protected] (C.E. Stickley), [email protected] (D. Bukry). http://dx.doi.org/10.1016/j.palaeo.2015.01.015 0031-0182/Published by Elsevier B.V. Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Paleoceanographic, and paleoclimatic constraints on the global Eocenediatom and silicoflagellate record
John A. Barron a,⁎, Catherine E. Stickley b, David Bukry a
a U.S. Geological Survey, MS910, Menlo Park, CA 94025, USAb Evolution Applied Ltd., 50 Mitchell Way, Upper Rissington, Cheltenham GL54 2PL, UK
⁎ Corresponding author.E-mail addresses: [email protected] (J.A. Barron),
(C.E. Stickley), [email protected] (D. Bukry).
http://dx.doi.org/10.1016/j.palaeo.2015.01.0150031-0182/Published by Elsevier B.V.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 August 2014Received in revised form 13 January 2015Accepted 18 January 2015Available online 24 January 2015
Keywords:DiatomSilicoflagellateEocene paleoclimatePaleoceanographySea levelOcean gateway
Eocene diatom and silicoflagellate biostratigraphy are summarized and correlated with the most recent geologictime scale as well as with the global oxygen isotope and eustatic sea level curves. The global distribution ofEocene diatom/silicoflagellate-bearing sediments varies considerably, reflecting changing oceanic gatewaysand paleoceanography with changing patterns that are punctuated by four major depositional events.Event 1 (~49 million years ago, Ma), at the end of the Early Eocene Climatic Optimum (EECO), saw the cessationof diatom/silicoflagellate deposition in epicontinental regions of the North Sea region and in the northern Russiaand the onset of biosilica deposition in the Arctic. Event 2 (~46 Ma), which coincided with intensification of theMiddle Eocene cooling trend, marked the widespread expansion of diatom/silicoflagellate deposition in both theNorth and South Atlantic. A shift of diatom/silicoflagellate deposition from the Atlantic to the Pacific began atEvent 3, at the end of the Middle Eocene Climatic Optimum (MECO) (~40 Ma), that was likely tied to the initialopening of the Drake Passage between Antarctica and South America. Event 4 (~39 Ma) coincided with a majorsea level fall and a widespread deep-sea hiatus in the latest Middle Eocene. Late Eocene diatom/silicoflagellatedeposition became more concentrated in middle-to-high latitude regions and coastal upwelling regions, partic-ularly in the Pacific Ocean.Tabulation of the first and last occurrences of 132 biostratigraphically-important diatoms suggests increasedspecies turnover during the latest Paleocene to earliest Eocene that may be in part due to a monographic effect.An increasing rate of evolution of newdiatom species between~46 and 43Maand after ~40Ma coincides respec-tivelywith thewidespread expansion of diatom deposition in the Atlantic andwith an increased pole-to-equatorthermal gradient that witnessed the expansion of diatoms in high latitude oceans and coastal upwelling settings.
Published by Elsevier B.V.
1. Introduction
The Eocene (~56 to 34Ma), the longest andmost climatically diverseepoch of the Cenozoic, spans from the warmest period of the Cenozoic,the Early Eocene, through a long-term cooling trend during the Middleand Late Eocene that culminated in themajor expansion of theAntarcticIce Sheet in the earliest Oligocene (Zachos et al., 2001, 2008). Althoughmarine diatoms constitute a major group of phytoplankton responsiblefor up to 45% of ocean's primary productivity, knowledge of their Eocenehistory is largely fragmentary and pieced together fromnumerous, geo-graphically separated and often stratigraphically discontinuous, on-landand deep-sea stratigraphic sections.
Diatoms, unicellular heterokont algae, produce a bipartite externalskeleton (frustule) composed of opaline silica (hydrated silicon dioxide),which is ornately sculpted and forms the basis of diatom taxonomic clas-sification. The size of diatom frustules ranges from less than a few
micrometers (μm) to more than 1000 μm, but most frustules range insize from 10 to 40 μm (Scherer et al., 2007). Diatoms are common virtu-ally everywhere there is water and light, in the ocean, freshwater, soils,and on damp surfaces. As the oceans are typically undersaturated in silica,lightly silicified diatom frustules are typically dissolved after death of theorganism during descent to the seafloor. Diatom frustules are preferen-tially preserved in seafloor sediments beneath areas of high surfacewater production, typically areas of upwelling, where seawater is silica-saturated. Even then, preservation in the older fossil record may be hin-dered by the instability of opaline diatom frustules to survive diagenesis,as diatomaceous silica (opal-A) is converted to amorphous cristobaliteand eventually to chert upon deep burial and exposure to temperaturesin excess of 50 °C. Where fossil diatoms are preserved, however, theyprovide awealth of paleoenvironmental, paleoceanographic and biostrat-igraphic information, particularly in regions where calcareous microfos-sils are in low abundance, e.g., the polar regions.
Marine surface waters and sediments containing diatoms typicallyalso contain silicoflagellates. Silicoflagellates are unicellular marinealgae that are entirely planktonic. They produce internal tubular skele-tons of opaline silica that are generally 30–100 μm in diameter, and

86 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
they are commonly found in the same samples (both living and fossil)as diatoms but at much reduced concentrations (generally b10%)(Perch-Nielsen, 1985). Silicoflagellates are commonly considered instudies of diatom biostratigraphy and paleoecology.
Themodern seafloor contains threemain belts of diatomaceous sed-iments: (1) a southern belt around Antarctica concentrated between45° and 65°S, (2) a northern belt that dominates in the Pacific north ofabout 40°N but is more restricted in the North Atlantic, and (3) anequatorial belt occurring in the nutrient-rich eastern areas of the Pacific,Atlantic, and Indian Oceans (Fig. 1). These belts coincide with areas ofupwelling and subpolar regions where high surface water nutrient(nitrogen, phosphorus, silica) concentrations fuel abundant populationsof diatoms. Each region contains distinctive diatom assemblages, whichare associated with distinctive water mass properties relating to seasurface temperature (SST), nutrient concentration, and salinity, for ex-ample. These regional differences intensifiedwith the onset of Antarcticglaciation and deep-water cooling in the earliest Oligocene (Fenner,1985; Scherer et al., 2007) and intensified further during the LateMiocene and Pliocene high-latitude cooling steps (Barron, 2003;Lazarus et al., 2014).
1.1. Eocene paleoclimatology
The Middle Eocene and Late Eocene are characterized by a ~15 mil-lion year-long cooling trend in middle to high latitudes that began at~49 Ma following the Early Eocene Climatic Optimum (EECO), thewarmest period of the Cenozoic (Zachos et al., 2001, 2008). In contrastto the current “icehouse” interval of extensive Antarctic glacial ice thatbegan in the earliest Oligocene (~34 Ma), and to the Early Eocene
AFRICA
ASIA
ANTARCTIC<100100-250
250-500>500
0°
30°N
60°N
30°S
60°S
0° 60°E 120°E
Fig. 1. Global variation in the extraction of dissolved silica (g/m2/yr) by phytoplankton in neathe dominant phytoplankton group in the oceans. The distribution of diatom production correla1972).
“greenhouse” interval of significant polar warmth (~56–49 Ma), thisMiddle to Late Eocene cooling has sometimes been termed the “doubthouse” interval in reference to the likelihood of small (likely ephemeral)high-topography ice sheets on Antarctica (Miller et al., 1991; Wilsonet al., 2013) and in the Arctic region (Stickley et al., 2009). The doubthouse was a period of climate instability reflected, for example, in theonset of Arctic sea ice that fluctuated episodically between seasonaland perennial regimes (Stickley et al., 2009; Darby, 2014); and in thepunctuation of the cooling trend by a brief warming at ~40 Ma termedthe Middle Eocene Climatic Optimum (MECO) (Bohaty et al., 2009).
Cramer et al. (2009) link variations in interbasinal deep water δ18Ogradients during theMiddle Eocene to Oligocenewith the increasing in-fluence of wind-driven surfacewatermixing associatedwith the gradu-al tectonic opening of Southern Ocean passages and the initiation andstrengthening of the Antarctic Circumpolar Current. The opening ofthe Tasmanian Gateway between Australia and Antarctica and theDrake Passage between South America and Antarctica were both criti-cal. Bijl et al. (2013) recently hypothesized that the opening of thesouthern end of the Tasmanian Gateway commenced at the EarlyEocene–Middle Eocene boundary (ca. 50–49 Ma) and may havebeen responsible for the onset of the Antarctic Circumpolar Current.Comparison of Pacific and Atlantic Pacific neodymium (Nd) isotopic ra-tios by Thomas (2004), Scher and Martin (2004), and Thomas et al.(2008) suggests that the flow of Pacific water through the DrakePassage began near the Middle–Late Eocene boundary. In support ofthese Nd studies, Borrelli et al. (2014) cite benthic foraminiferal isotopeevidence in arguing that thermal differentiation between NorthernComponent Water (NCW) and Southern Component Water (SCW)was initiated at ~38.5 Ma during the initial stages of Drake Passageopening.
AMERICA
NORTH
A
180°E 240°E
r-surface ocean waters. This represents relative diatom production, because diatoms aretes well with belts of diatomaceous sediments on the sea floor (Reproduced from Lisitzin,

87J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
Recent improvements in the Eocene geologic time scale (Gradsteinet al., 2004, 2012) necessitate new age calibration for many of theseEocene paleoclimatic and paleoceanographic events in these studies.However, many of these published Eocene proxy records of paleoclimat-ic and paleoceanographic change are referenced only against geologictime (Ma) rather than against the constraining microfossil (mainly cal-careous nannofossil) biostratigraphies, making it difficult to compare de-tailed results. The purpose of this paper is fourfold: 1) we present thefirst comprehensive stratigraphic summary of the key global Eocenediatom- and silicoflagellate-rich deposits, 2) update the age calibrationof Eocene diatom and silicoflagellate biostratigraphy to the geologicaltimescale of Gradstein et al. (2012), 3) place the global record ofdiatom/silicoflagellate deposition into a paleoceanographic and paleocli-matic framework, which allows us to 4) suggest controls on the globaldistribution of Eocene diatoms and silicoflagellates. All ages cited inthis paper refer to those in Gradstein et al. (2012) with integration ofthe astrochronology byWesterhold et al. (2012) for the Early Paleogenethat includes the Early Eocene. We acknowledge that there remainproblems with the assignment of accurate numerical ages to the midPaleogene (Westerhold et al., 2012) and that the Paleogene timescale re-mains in a state of transition (Gradstein et al., 2012).
C17
C16
C15
C13
Late
Mid
dle
C18
Oligo.
CH
RO
N
PO
LAR
ITY
Ma EP
OC
H /
SU
BE
PO
CH
TropicalDiatom Zone
Eoc
ene
Ear
ly
C21
C22
C23
C24
C25
C20
C19 Hemiaulus
gondolaformis
Hemiaulus alatus
Triceratium kanayae
Cr. oblongus
Cr. undulatus
Pyxillagracilis
Hemiaulus
incurvus
34
36
38
40
42
44
46
48
50
52
54
56
58
Paleoc.
CP
13
CP
14
a
c
b
a
b
a
CP
12
CP
11
CP
10
ba
CP7
CP
8C
P9
b
a
CP
15
CP
16 b
a
NannoZone
Pyxillacaputavis
NP21
NP19 -20
NP18
NP16
NP
15
NP
14
NP13
NP12
NP11
NP10
NP9
Cestodiscus pulchellusnovazeal.
Cestodiscus fennerae
Cestodiscus antarcticus
Ces. robustus
Planktonic Foram Zone
a
b
bNP17 Triceratium inconspicuum
Rad.Zone
RP13
RP14
RP15
RP16
RP17
RP18
RP19
RP12
RP8
?
P. o
ligoc
aeni
ca te
nue
NP8
RP20
Bip
ala
oam
arue
nsis
M. u
ral.
T. v
ent.
C. p
ay.
E4
E3
E2E1P5
P4
E5
E6
E7
E8
E9
E10
E11
E12
E13
E14
E15
E16
O1
RP11
RP9
c
b
a
b
a
ba
?
Cos
cino
disc
us e
xcav
atus
Ces
todi
scus
robu
stus
Ces
todi
scus
ant
arct
icus
Ces
todi
scus
fenn
eraeC
esto
disc
us tr
ochu
sA
zpei
tia o
ligoc
enic
aC
esto
disc
us c
onve
xus
Ryl
ands
ia b
iradi
ata
Eur
aisa
in P
latfo
rm
Cr. oblongus -Br. imperfecta
Dia
?
?
Pyx
illa
grac
ilis
Fig. 2. Eocene diatom and silicoflagellate zones and ranges of key taxa referenced against the Gradsee Table 1. Planktonic foraminiferal zones after Berggren and Pearson (2005); calcareous nan(1980)(right); radiolarian zones after Nigrini et al. (2005); diatom zones (Fenner, 1985; Barroand Radionova and Khokhlova, 1994. Abbreviations: T. vent. = Trinacria ventriculosa;M. ural. =
2. Diatom and silicoflagellate biostratigraphy
Diatom biostratigraphy utilizes evolutionary events (appearancesand extinctions) of mainly planktonic species, as these are more wide-spread ecologically and geographically than benthic species (Fenner,1985; Scherer et al., 2007). As summarized by Fenner (1985), pre-Oligocene diatom biostratigraphy lacks refinement, due to the scarcityof continuous reference sections. Eocene biostratigraphy has beenpieced together mostly through study of deep-sea sections that havebeen calibrated to the geologic time scale through calcareous nanno-fossil biostratigraphy (Fenner, 1984a,b, 1985; Fenner and Mikkelsen,1990).
Fig. 2 summarizes Eocene diatom biostratigraphy and the ranges ofkey biostratigraphic taxa referenced against the geologic time scale ofGradstein et al. (2012)(Table 1). In most cases for the Eocene, the agecalibration of these ranges is indirect through the correlation of diatomzones to other microfossil zonations that are tied directly with paleo-magnetic stratigraphy. The tropical diatom zonation of Fenner (1985)with modifications by Scherer et al. (2007) is updated by the substitu-tion of the zonation for the Late Eocene of Ocean Drilling Program(ODP) Site 1090 in the southeast Atlantic Ocean by Barron et al.
Ces
todi
scus
pul
chel
lus
v. no
vaze
alan
dica
Ast
erol
ampr
a m
aryl
andi
caB
right
wel
lia im
perfe
cta
Hem
iaul
us g
ondo
lafo
rmis
Hem
iaul
us a
latu
sP
yxill
a ca
puta
vis
Cra
sped
odis
cus
undu
latu
sP
yxill
a gr
acili
s
Moi
ssee
via
ural
ensi
s
Cos
cind
iscu
s pa
yeri
Azp
eitia
tube
rcul
ata
Trin
acria
ven
ticul
osa
Hem
iaul
us in
curv
us
Cra
sped
odis
cus
oblo
ngus
Pyx
illa
olig
ocae
nica
Tric
erat
ium
inco
nspi
cuum
v. t
rilob
ata
Fene
stre
lla a
ntiq
ua
Tric
erat
ium
kan
ayae
Silicoflagellate Zone
Dictyochahexacantha
Corb
isem
a apic
ula
taN
avic
ulo
psis
fo
liace
a
N
av.
robu
sta
Dic
tyo
ch
a s
pin
osa
Naviculopsis constricta
Dc. deflandre
iN
. tr
ispin
osa s
z
tom Ranges
Dic
tyoc
ha h
exan
cant
a
Dic
tyoc
ha s
pino
sa
Bac
hman
noce
na o
amar
uens
is
Nav
icul
opsi
s bi
apic
ualta
s.s
.D
icty
ocha
def
land
rei s
.s.
Nav
icul
opsi
s tri
spin
osa
Nav
icul
opsi
s ro
bust
aN
avic
ulop
sis
folia
cea
Bac
hman
noce
na a
picu
lata
s.s
.
Nav
icul
opsi
s as
pera
Nav
icul
opsi
s co
nstri
cta
Cor
bise
ma
apic
ualta
Dic
tyoc
ha e
long
ata
Cor
bise
ma
bim
ucro
nata
#
stein et al. (2012) Geologic Time Scale. #, base of zone is younger in high latitude sections –nofossil zones after Martini (1970)(left) and Bukry (1973a, 1975a) and Okada and Bukryn et al., 2014); Eurasian platform (mainly Russia) diatom zones after Strelnikova (1992)Moisseevia uralensis; C. pay. = Coscinodiscus payeri.

Table 1Names, markers, authors, age estimates, and source of correlation of diatom and silicoflagellate zones.
Base Author Age (Ma) Constraint Source of correlation
Tropical diatomCestodiscus robustus FO C. robustus Barron et al. (2014) 33.7 PM Barron et al. (2014)Cestodiscus antarcticus FO C. antarcticus Barron et al. (2014) 34.9 PM Barron et al. (2014)Cestodiscus fennerae FO C. fennerae Barron et al. (2014) 36.9 PM Barron et al. (2014)Cestodiscus pulchellusvar. novazelandica LO Triceratium inconspicuum Barron et al. (2014) ~39.0 N Barron et al. (2014)Triceratium inconspicuum LO Craspedodiscus oblongus McLean and Barron (1988) ~40.4 N Fenner (1984a); mod*Craspedodiscus oblongus-Brightwellia imperfecta FO B. imperfecta Fenner (1984a): mod* ~41.8 N Fenner (1984a)Hemiaulus gondolaformis FO H. gondolaformis Fenner (1984a) ~43.6 N Fenner (1984a)Hemiaulus alatus FO H. alatus Fenner (1984a) ~44.4 N Fenner (1984a)Pyxilla caputavis FO P. caputavis Fenner (1984a) ~45.6 N Fenner (1984a)Triceratium kanayae FO T. kanayae Fenner (1984a) ~48.4 N Fenner (1984a)Craspedodiscus oblongus FO C. oblongus Fenner (1984a) ~50.8 N Fenner (1984a)Craspedodiscus undulatus FO C. undulatus Gombos (1982); Fourtanier (1991) ~52.3 N Fenner (1984a)Pyxilla gracilis FO P. gracilis Fourtanier (1991) ~54.0 PM Fourtanier (1991)Hemiaulus incurvus FO H. incurvus Fourtanier (1991) ~60.5 PM, N Fourtanier (1991)
Russian platform diatomBipala oamaruensis FO Bipala oamaruensis Radionova and Khokhlova (1994) ~45 N Radionova et al. (2003)Pyxilla oligoaenica var. tenue FO P. oligocaenica Strelnikova (1992) ~50.6 N Oreshkina (2012)Pyxilla gracilis FO P. gracilis Strelnikova (1992) ~54.0 N Oreshkina (2012)Coscindiscus payeri FO C. payeri Strelnikova (1992) ~55.3 N Oreshkina (2012)Moissevia uralensis FO M. uralensis Strelnikova (1992) 57.2 N Oreshkina (2012)Trinacria ventriculosa FO T. venticulosa Strelnikova (1992) ~58.5 N Oreshkina (2012)
SilicoflagellateCorbisema apiculata LO Dictyocha hexacantha Perch-Nielsen (1985); Bukry (1978a) 37.0 PM Barron et al. (2014)Dictyocha hexacantha FO Dictyocha hexacantha Bukry & Foster (1974); Bukry (1977) ~43.6
(~40.0 highlatitude)
N Perch-Nielsen (1985):Kim and Glezer (2007)
Dictyocha spinosa sz+ FO Corbisema spinosa Bukry (1977) (DSDP 39) ~50.8 N Perch-Nielsen (1985)Naviculopsis robusta sz+ FO Naviculopsis foliacea Bukry (1978a) (DSDP 43) ~54.7 N Perch-Nielsen (1985)Naviculopsis constricta FO Naviculopsis constricta Perch-Nielsen (1975) ~59.0 N Perch-Nielsen (1985)
Notes: mod* = modified herein; + = subzone of Naviculopsis foliacea Zone; PM = paleomagnetic stratigraphy; N = nannofossil stratigraphy.
88 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
(2014). A separate zonation for sediments deposited on the Eurasianepicontinental platform (mainly Russia but including parts of theUkraine and northern Kazakhstan) is included for the Early Eoceneafter Strelnikova (1992). These deposits occur in the threemain regions:Volga-North Peri-Caspian (Central Russia plate), West Siberia lowland(Trans Uralia) and Eastern Peri-Caspian region (T. Oreshkina, writtencomm., 2014). Fenner (1984a, b, 1985) and Fenner and Mikkelsen(1990) detail the diatom biostratigraphy of a number of key Deep SeaDrilling Project (DSDP) sections, many of which have calcareousnannofossil, planktonic foraminifer, and/or radiolarian biostratigraphy.
Early Eocene diatomassemblages are dominated by relatively robustgenera such as Pyxilla, Hemiaulus, Stephanopyxis, Trinacria, andTriceratium that were gradually replaced during the late Middle andLate Eocene by assemblages dominated bymore lightly silicified centricdiatoms such as Asterolampra, Azpeitia, Cestodiscus, and Coscinodiscus(Fenner, 1985; Strelnikova, 1990; Barron et al., 2014). This trend towardmore lightly silicified taxa coincided with middle-to-high latitudecooling and increased mixing rates of surface waters.
Eocene silicoflagellate biostratigraphy is less advanced than diatombiostratigraphy and relies on events of Naviculopsis and Dictyocha, twomostly warm-water genera (Bukry, 1981; Perch-Nielsen, 1985). Similarto diatom biostratigraphy, Cenozoic silicoflagellate biostratigraphy hasbeen pieced together during the past 40+ years through study of tem-porally limited and often geographically isolated deep-sea sections(Table 1). A summary of themost widely applied Eocene silicoflagellatezones and their marker taxa is included in Fig. 2.
Calibration of Eocene diatom and silicoflagellate zones to geologictime has mainly been through calcareous nannofossil zones of Bukry(1973a, 1975a): Okada and Bukry (1980) and Martini (1970) and theradiolarian zones of Nigrini et al. (2005), which, in turn, have been cal-ibrated with the geomagnetic polarity time scale by Berggren andPearson (2005), Nigrini et al. (2005), and Lyle et al. (2010). The plank-tonic foraminiferal zones of Berggren and Pearson (2005) are includedin Fig. 2 for comparison. Unfortunately, direct correlation of Eocene dia-tom biostratigraphywith robust paleomagnetic stratigraphy is available
in very few sections globally; these include ODP 752A in the easternIndian Ocean (Fourtanier, 1991), ODP 1090A in the southeast Atlantic(Barron et al., 2014), and the Shaldril core hole off the Antarctic Penin-sula (Bohaty et al., 2011). Correlation of diatom-bearing biosiliceous in-tervals are also available for ODP 1172 south of Australia (Stickley et al.,2004a; Bijl et al., 2011, 2013) and indirectly through comparison withearlier biostratigraphic studies for DSDP 338 and ODP 913B in theNorwegian-Greenland Sea (Eldrett et al., 2004). An updated correlationof diatom and silicoflagellate zones and othermicrofossil zoneswith thegeologic time scale of Gradstein et al. (2012) is presented in Fig. 2.
3. Eocene diatom-bearing sections
Table 2 lists important Eocene reference sections that have beenstudied for diatom and silicoflagellate biostratigraphy along with thebasis of their chronological age constraints. Onshore Eocene sectionsthat have been analyzed in detail include: the Fur Formation inDenmark (Homann, 1991; Fenner, 1994), sections in the North Seaarea (Mitlehner, 1996); the Kellogg Shale of California (Barron et al.,1984), the Oamaru Diatomite of New Zealand (Edwards and DSIRGeology and Geophysics, 1991), and numerous sections from theEurasian epicontinental platform, mainly from the Crimea, Caucasus,and western Siberia (Glezer, 1994, 1996a,b; Oreshkina and Oberhänsli,2003; Radionova et al., 2003; Oreshkina, 2012). Other important on-shore diatomaceous sections that have had other microfossil biostratig-raphies published but have yet to have been thoroughly studied fordiatoms include the Oceanic Formation of the Barbados (Saunderset al., 1984), the Kreyenhagen Formation of California (Dumoulin,1984; McLean and Barron, 1988), and onshore sequences in Peru(Marty et al., 1988; Dunbar et al., 1990). The potential importance of Eo-cene diatomaceous sections in Kamchatka has recently been demon-strated by the diatom biostratigraphic studies of Tsoy (2003, 2011)and Gladenkov (2013) who record isolated assemblages from both theMiddle and Late Eocene that are similar in age to diatomassemblages re-corded from theKellogg Shale and Kreyenhagen Formation of California.
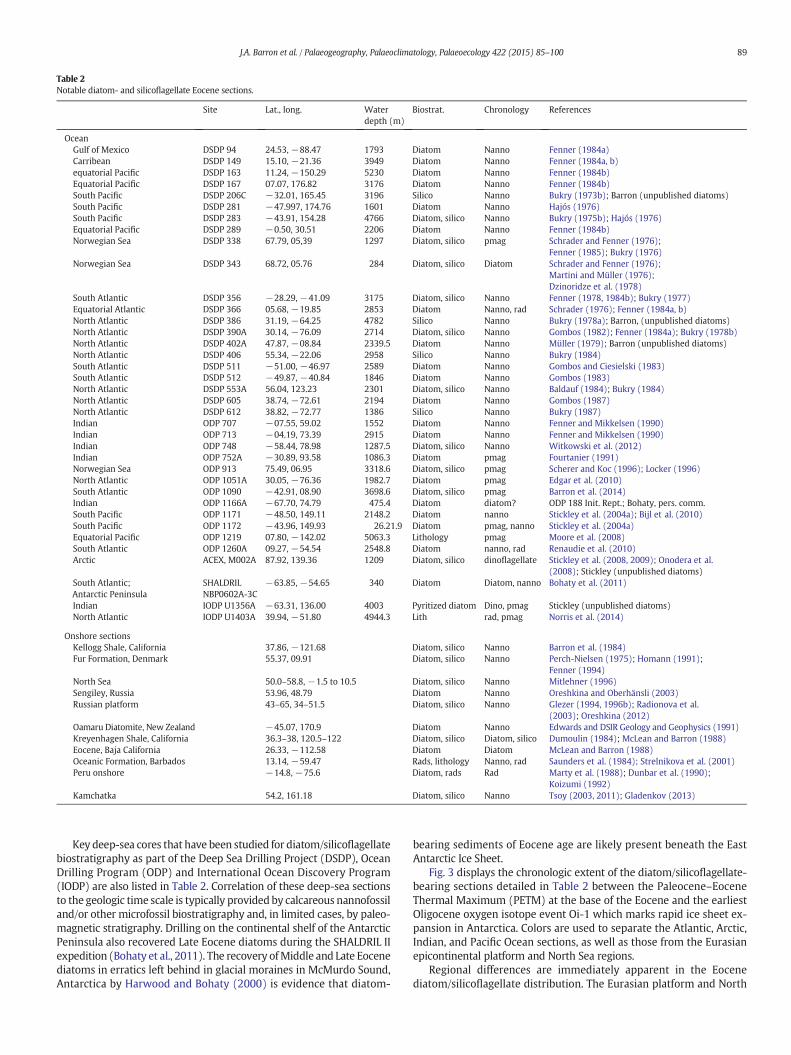
Table 2Notable diatom- and silicoflagellate Eocene sections.
Site Lat., long. Waterdepth (m)
Biostrat. Chronology References
OceanGulf of Mexico DSDP 94 24.53, −88.47 1793 Diatom Nanno Fenner (1984a)Carribean DSDP 149 15.10, −21.36 3949 Diatom Nanno Fenner (1984a, b)equatorial Pacific DSDP 163 11.24, −150.29 5230 Diatom Nanno Fenner (1984b)Equatorial Pacific DSDP 167 07.07, 176.82 3176 Diatom Nanno Fenner (1984b)South Pacific DSDP 206C −32.01, 165.45 3196 Silico Nanno Bukry (1973b); Barron (unpublished diatoms)South Pacific DSDP 281 −47.997, 174.76 1601 Diatom Nanno Hajós (1976)South Pacific DSDP 283 −43.91, 154.28 4766 Diatom, silico Nanno Bukry (1975b); Hajós (1976)Equatorial Pacific DSDP 289 −0.50, 30.51 2206 Diatom Nanno Fenner (1984b)Norwegian Sea DSDP 338 67.79, 05,39 1297 Diatom, silico pmag Schrader and Fenner (1976);
Fenner (1985); Bukry (1976)Norwegian Sea DSDP 343 68.72, 05.76 284 Diatom, silico Diatom Schrader and Fenner (1976);
Martini and Müller (1976);Dzinoridze et al. (1978)
South Atlantic DSDP 356 −28.29, −41.09 3175 Diatom, silico Nanno Fenner (1978, 1984b); Bukry (1977)Equatorial Atlantic DSDP 366 05.68, −19.85 2853 Diatom Nanno, rad Schrader (1976); Fenner (1984a, b)North Atlantic DSDP 386 31.19, −64.25 4782 Silico Nanno Bukry (1978a); Barron, (unpublished diatoms)North Atlantic DSDP 390A 30.14, −76.09 2714 Diatom, silico Nanno Gombos (1982); Fenner (1984a); Bukry (1978b)North Atlantic DSDP 402A 47.87, −08.84 2339.5 Diatom Nanno Müller (1979); Barron (unpublished diatoms)North Atlantic DSDP 406 55.34, −22.06 2958 Silico Nanno Bukry (1984)South Atlantic DSDP 511 −51.00, −46.97 2589 Diatom Nanno Gombos and Ciesielski (1983)South Atlantic DSDP 512 −49.87, −40.84 1846 Diatom Nanno Gombos (1983)North Atlantic DSDP 553A 56.04, 123.23 2301 Diatom, silico Nanno Baldauf (1984); Bukry (1984)North Atlantic DSDP 605 38.74, −72.61 2194 Diatom Nanno Gombos (1987)North Atlantic DSDP 612 38.82, −72.77 1386 Silico Nanno Bukry (1987)Indian ODP 707 −07.55, 59.02 1552 Diatom Nanno Fenner and Mikkelsen (1990)Indian ODP 713 −04.19, 73.39 2915 Diatom Nanno Fenner and Mikkelsen (1990)Indian ODP 748 −58.44, 78.98 1287.5 Diatom, silico Nanno Witkowski et al. (2012)Indian ODP 752A −30.89, 93.58 1086.3 Diatom pmag Fourtanier (1991)Norwegian Sea ODP 913 75.49, 06.95 3318.6 Diatom, silico pmag Scherer and Koc (1996); Locker (1996)North Atlantic ODP 1051A 30.05, −76.36 1982.7 Diatom pmag Edgar et al. (2010)South Atlantic ODP 1090 −42.91, 08.90 3698.6 Diatom, silico pmag Barron et al. (2014)Indian ODP 1166A −67.70, 74.79 475.4 Diatom diatom? ODP 188 Init. Rept.; Bohaty, pers. comm.South Pacific ODP 1171 −48.50, 149.11 2148.2 Diatom nanno Stickley et al. (2004a); Bijl et al. (2010)South Pacific ODP 1172 −43.96, 149.93 26.21.9 Diatom pmag, nanno Stickley et al. (2004a)Equatorial Pacific ODP 1219 07.80, −142.02 5063.3 Lithology pmag Moore et al. (2008)South Atlantic ODP 1260A 09.27, −54.54 2548.8 Diatom nanno, rad Renaudie et al. (2010)Arctic ACEX, M002A 87.92, 139.36 1209 Diatom, silico dinoflagellate Stickley et al. (2008, 2009); Onodera et al.
(2008); Stickley (unpublished diatoms)South Atlantic;Antarctic Peninsula
SHALDRILNBP0602A-3C
−63.85, −54.65 340 Diatom Diatom, nanno Bohaty et al. (2011)
Indian IODP U1356A −63.31, 136.00 4003 Pyritized diatom Dino, pmag Stickley (unpublished diatoms)North Atlantic IODP U1403A 39.94, −51.80 4944.3 Lith rad, pmag Norris et al. (2014)
Onshore sectionsKellogg Shale, California 37.86, −121.68 Diatom, silico Nanno Barron et al. (1984)Fur Formation, Denmark 55.37, 09.91 Diatom, silico Nanno Perch-Nielsen (1975); Homann (1991);
Fenner (1994)North Sea 50.0–58.8, −1.5 to 10.5 Diatom, silico Nanno Mitlehner (1996)Sengiley, Russia 53.96, 48.79 Diatom Nanno Oreshkina and Oberhänsli (2003)Russian platform 43–65, 34–51.5 Diatom, silico Nanno Glezer (1994, 1996b); Radionova et al.
(2003); Oreshkina (2012)Oamaru Diatomite, New Zealand −45.07, 170.9 Diatom Nanno Edwards and DSIR Geology and Geophysics (1991)Kreyenhagen Shale, California 36.3–38, 120.5–122 Diatom, silico Diatom, silico Dumoulin (1984); McLean and Barron (1988)Eocene, Baja California 26.33, −112.58 Diatom Diatom McLean and Barron (1988)Oceanic Formation, Barbados 13.14, −59.47 Rads, lithology Nanno, rad Saunders et al. (1984); Strelnikova et al. (2001)Peru onshore −14.8, −75.6 Diatom, rads Rad Marty et al. (1988); Dunbar et al. (1990);
Koizumi (1992)Kamchatka 54.2, 161.18 Diatom, silico Nanno Tsoy (2003, 2011); Gladenkov (2013)
89J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
Key deep-sea cores that have been studied for diatom/silicoflagellatebiostratigraphy as part of the Deep Sea Drilling Project (DSDP), OceanDrilling Program (ODP) and International Ocean Discovery Program(IODP) are also listed in Table 2. Correlation of these deep-sea sectionsto the geologic time scale is typically provided by calcareous nannofossiland/or other microfossil biostratigraphy and, in limited cases, by paleo-magnetic stratigraphy. Drilling on the continental shelf of the AntarcticPeninsula also recovered Late Eocene diatoms during the SHALDRIL IIexpedition (Bohaty et al., 2011). The recovery ofMiddle and Late Eocenediatoms in erratics left behind in glacial moraines in McMurdo Sound,Antarctica by Harwood and Bohaty (2000) is evidence that diatom-
bearing sediments of Eocene age are likely present beneath the EastAntarctic Ice Sheet.
Fig. 3 displays the chronologic extent of the diatom/silicoflagellate-bearing sections detailed in Table 2 between the Paleocene–EoceneThermal Maximum (PETM) at the base of the Eocene and the earliestOligocene oxygen isotope event Oi-1 which marks rapid ice sheet ex-pansion in Antarctica. Colors are used to separate the Atlantic, Arctic,Indian, and Pacific Ocean sections, as well as those from the Eurasianepicontinental platform and North Sea regions.
Regional differences are immediately apparent in the Eocenediatom/silicoflagellate distribution. The Eurasian platform and North

C17
C16
C15
C13
Late
Mid
dle
C18
Oligo.
CH
RO
N
PO
LAR
ITY
Ma EP
OC
H /
SU
BE
PO
CH
TropicalDiatom Zone
Eoc
ene
Ear
ly
C21
C22
C23
C24
C25
C20
C19 Hemiaulus
gondolaformis
Hemiaulus alatus
Triceratium
kanayae
Cr. oblongus
Cr. undulatus
Pyxillagracilis
Hemiaulus
incurvus
34
36
38
40
42
44
46
48
50
52
54
56
58
Paleoc.
CP
13
CP
14
a
c
b
a
b
a
CP
12
CP
11
CP
10
ba
CP7
CP
8C
P9
b
a
CP
15
CP
16 b
a
NannoZone
Pyxillacaputavis
NP21
NP19 -20
NP18
NP16
NP
15
NP
14
NP13
NP12
NP11
NP10
NP9
Cr. oblongus -Br. imperfecta
Cestodiscus pulchellusnovazeal.
Cestodiscus fennerae
Cestodiscus antarcticus
Ces. robustus
TropicalSilicoflagellate Zone
Dictyochahexacantha
Co
rbis
em
a
ap
icu
lata
Navic
ulo
psis
fo
liace
a
Nav.
robu
sta
Dic
tyo
ch
a s
pin
osa
Naviculopsis constricta
Dc. deflandre
iN
. tr
ispin
osa s
z
*
283
356
386
553A
Kellogg
149
366
?
343
406
? 4
02A
206C
281
748
126
0A
Planktonic Foram Zone
a
b
Fur
*33
8
*752
A
94
bNP17 Triceratium inconspicuum
512
605
70
7 &
71
3
289
163
167
Rad.Zone
RP13
RP14
RP15
RP16
RP17
RP18
RP19
RP12
RP8
Pacific biogenic silica accumulationevents (ESAE) of Moore et al. (2008)* = paleomagnetic stratigraphy
?
P. o
ligoc
aeni
ca te
nue
Atlantic Ocean Pacific OceanIndianOceanA
rcti
c
*109
0
AC
EX
NP8
*91
3
?
390
A
RP20
Pyx
illa
grac
ilis
M. u
ral.
T v
ent.
C. p
ay.
wid
espr
ead
cher
ts
Oce
anic
For
mat
ion
-Bar
bado
s
* U
1403
A
Event 1
Event 2
E4
E3
E2E1P5
P4
E5
E6
E7
E8
E9
E10
E11
E12
E13
E14
E15
E16
O1
RP11
RP9
c
b
a
b
a
ba
?
511
Nor
th S
ea
Event 3
Per
u on
shor
e
Cal
iforn
ia K
reye
nhag
en
*105
1
MECO
South -tropics- North NG.
612
612
South --tropics------ North
*12
19
Pla
tfo
rm
SH
ALD
RIL
3C
117
211
711166
? Oam
aru
hiatus of Wade & Kroon (2002)
PETM
Oi-1 Event
?
Eur
asia
n pl
atfo
rm s
ectio
ns
hiat
us?
U13
56A
?
Sen
gile
y
Bip
ala
oam
arue
nsis
?
?
Eur
aisa
n P
latfo
rm
Kam
chat
ka o
ccur
renc
es
Event 4
?
?
Fig. 3. Chronologic extent and distribution of key diatom/silicoflagellate-bearing sections (see Table 2). Colors are used to separate the Atlantic (green), Arctic (brown), Indian (purple),and Pacific (light blue) Ocean sections, aswell as those from the Eurasian epicontinental platform andNorth Sea (pink) regions. Dashed bars indicate sectionswith poor stratigraphic con-trol. Bold type and an asterisk indicate sections with published paleomagnetic stratigraphy. Blue bar in radiolarian zones refers to the Eocene Silica Accumulation Events (ESAE) of Mooreet al. (2008) in the equatorial Pacific. Key events indicated by gray shading: PETM Paleocene–Eocene Thermal Maximum; Diatom Depositional (DD) Event 1; DD Event 2; DD Event 3, theMiddle Eocene Climatic Optimum (MECO); DD Event 4; Oi-1, the earliest Oligocene oxygen isotope event associated with ice sheet expansion in Antarctica. (For interpretation of the ref-erences to color in this figure legend, the reader is referred to the web version of this article.)
90 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
Sea sections are mainly restricted to the Early Eocene, with limited oc-currences in the Middle Eocene (see Radionova et al. (2003) andOreshkina (2012)). Atlantic diatom/silicoflagellate-bearing sequencesoccur throughout the Eocene but are most numerous in the MiddleEocene interval between ~46 and 40 Ma. Widespread cherts occurin the North Atlantic during the Early Eocene and early MiddleEocene; however, their origin is not likely associated with diatomdeposition (Mutoni and Kent, 2007). Central Arctic continuousdiatom/silicoflagellate oozes appear to be largely concentrated in theearly Middle Eocene interval between ~49 (base C22n) and 46 Ma(ages updated), but biosilica occurs until ~36 Ma (early Late Eocene)(Dell'Agnese and Clark, 1994; Onodera et al., 2008; Stickley et al.,2008, 2009; Poirier and Hillaire-Marcel, 2011). Indian Ocean Eocenediatom/silicoflagellate-bearing sequences are few, restricted to theEarly Eocene and early part of the Middle Eocene (ODP 752 and IODPU1356A (poorly-preserved pyritized specimens), the Middle Eocene(ODP Sites 707, 713, and 748) and the Late Eocene of the Antarctic mar-gin in Prydz Bay (ODP 1166). Pacific diatom/silicoflagellate-bearing sec-tions aremost common in the lateMiddle and Late Eocene interval after41 Ma. A notable exception is the South Pacific ODP Site 1172, whichbears an abundant, well-preserved and long record of diatoms from
the early Middle Eocene (~45 Ma) through the Late Eocene, includingthe Middle Eocene Climatic Optimum (MECO), across the Eocene–Oli-gocene transition and throughout the Neogene (Stickley et al., 2004a,b; updated by Bijl et al. (2010)). Comparable Early Eocene through ear-liest Oligocene pyritized assemblages are preserved at IODP SiteU1356A on the Wilkes Land Margin (Escutia et al., 2011; Stickley, un-published data) and as opal-A in the Neogene (Escutia et al., 2011).
Ideally, diatom and silicoflagellate relative abundances in sedimentsshould be expressed quantitatively in percent biogenic silica (opal);however, such compilations have not yet been completed for the Eo-cene oceans with the exception of that of Moore et al. (2008) for thetropical Pacific Ocean. Moore et al. (2008) document Eocene biogenicsilica accumulation events (ESAE) (dark blue bars shown in radiolarianzones in Fig. 3) in the tropical Pacific that may coincide with enhanceddiatom deposition. However, Moore et al. (2008) argue that Eocene ra-diolarians likely benefitted from enriched silica concentrations in deepwaters that were as much as 10 °C warmer than those of the Neogenecoupledwith reduced silica utilization by diatoms in the relatively poor-ly mixed surface waters of the warm Eocene oceans.
The global pattern of Eocene diatom/silicoflagellate deposition ap-pears to have been punctuated by at least four events (Table 3), which

Table 3Description, biostratigraphic markers, and climate/tectonic expression of major Eocene diatom deposition events.
Event Age (Ma) Description Nanno Zone Diatom marker Climate/sea level Tectonic
4 ~39 Completion of shift of diatomdeposition from Atlantic toPacific and upwelling areas
UppermostCP14b
LO Tr. inconspicuum(middle latitudes)
Major sea level fall; widespreaddeep-sea hiatus
Noticable opening of Drake Passage
3 ~40 Onset of shift of diatom depositionfrom Atlantic to Pacific andupwelling areas; end of MECO
lower CP14b ~LO Cr. oblongus Cooling event at end of MECO;coastal upwelling increases
Initial opening of Drake Passage?
2 ~46–47 Onset widespread Atlanticdiatom deposition
CP13a ~FO Az. tuberculata Intensified cooling; end of widespreadAtlantic cherts; onset intensified oceancirculation; CCD rise in Atlantic; increasedradiolarian deposition equat. Pacific
Onset of drift deposition in NewFoundland area; enhanced oceancirculation IODP 342
1 ~49–50 Decline in North Sea and EurasianPlatform diatom deposition,onset of Arctic diatom deposition
CP12a FO Tr. inconspicuum Start of long-term cooling trend; closing ofTurgai Strait via sea level fall?
Isolated Arctic; North AtlanticReflector A, extensive erosion
91J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
are indicated by gray horizontal bars in Fig. 3. Depositional Event 1 oc-curs within calcareous nannofossil subzone CP12a (at ~49 Ma) whereit marks the cessation of diatom/silicoflagellate deposition on epiconti-nental regions of the North Sea and central Russia and the onset ofdiatom/silicoflagellate deposition in the central Arctic. Event 2 at~46 Ma occurs within calcareous nannofossil subzone CP13a where itcoincides with the onset of widespread diatom deposition in both theNorth and South Atlantic Oceans. Event 2 also appears to approximatea decline in diatom/silicoflagellate deposition in the Arctic (Onoderaet al., 2008; Stickley et al., 2008). Event 3 corresponds with the end ofthe Middle Eocene Climatic Optimum (MECO; Bohaty and Zachos,2003), occurring at ~40 Ma in the uppermost part of calcareousnannofossil subzone CP14a. Event 3 coincides with an abrupt declinein Atlantic diatom/silicoflagellate deposition and the onset of increasedequatorial and North Pacific diatom/silicoflagellate deposition. Event 4ismarked by awidespread deep-sea hiatus in the uppermost part of cal-careous nannofossil subzone CP14b at ~39 Ma. In middle latitudediatom-bearing sections Event 4 often coincides with the last occur-rence of the cosmopolitan diatom Triceratium inconspicuum, althoughthis diatom has its last occurrence at the termination of the MECO(Event 3) at low latitude ODP Site 1051 (Witkowski et al., 2014). AfterEvent 4, Late Eocene diatom/silicoflagellate deposition became moreconcentrated in middle-to-high latitude regions and coastal upwellingregions, particularly in the Pacific Ocean.
4. Paleoceanographic controls on Eocene diatom/silicoflagellatedistribution
Fig. 4 superimposes the oxygen isotope curve of Zachos et al. (2008)and the sea level curve of Kominz et al. (2008) (both updated to the2012 geologic time scale) on the Eocene diatom/silicoflagellate-bearingsection distributionfigure (Fig. 3). Eocene sea level generally follows thetrend of the oxygen isotope curve, high during the warm Early Eoceneand decreasing during the Middle and Late Eocene, but punctuated byfluctuations. Radionova et al. (2003) point out that gaps in Eocenediatom-bearing sections on the Eurasian epicontinental platform coin-cide with drops in global sea level.
4.1. The Early Eocene
The Eocene began with the Paleocene–Eocene Thermal Maximum(PETM), a geologically brief (~170 kyr duration) intervalwherein globaltemperatures rose by ~6 °C in both high and low latitudes and in thedeep ocean (Westerhold et al., 2008). The PETMwasmarked by a signif-icant negative excursion of carbon stable isotopes (δ13C),which coincid-ed with marked shoaling of the ocean's carbonate compensation depththat suggests that a massive amount of 13C-depleted CO2 entered thehydrosphere or atmosphere. Although a massive extinction of benthicforaminifers accompanied the PETM, planktonic foraminifers and dino-flagellates appear to have flourished. Fourtanier (1991) reported no
significant diatom assemblage change at ODP Site 752 in the easternIndian Ocean, the only diatom-bearing deep-sea sequence that crossesthe PETM. Oreshkina (2012) analyzed well-preserved diatom assem-blages across the PETM in the middle and northern Ural Mountain re-gion of western Siberia. She correlated assemblages from the upperpart of the Trinacria ventriculosa Zone with the pre-PETM, assemblagesof the Hemiaulus proteus Zone with the PETM itself, and assemblagesof the Moisseevia uralensis Zone with the post-PETM. Oreshkina(2012) noted that the PETM interval was characterized by transgressivediatomites containing high taxonomic diversity.
During the Early Eocene, a reduced pole-to-equator temperaturegradient resulted in warm surface and deep waters that were poorlymixed by relatively weak zonal winds. Diatom deposition was in shal-low seas such as on the Eurasian epicontinental platform and in theNorth Sea region. Mitlehner (1996) argued that Early Eocene diatoma-ceous units were deposited in a highly restricted sub-basin within a re-stricted North Sea characterized by wide range of diatom preservation.Mitlehner (1996) proposed that low salinity surface layerswith upwell-ing of nutrient-richwaters occurred in the area of present day Denmark.
Citing the arguments of Yool and Tyrell (2005), Mutoni and Kent(2007) argue that the concentration of cherts in the North Atlantic dur-ing calcareous nannofossil zones NP10 to early NP15 (~55–46 Ma,Fig. 4) was due to basin–basin fractionation. They point out that the dis-tribution of Early Eocene cherts in the North Atlantic does not coincidewith oceanic upwelling belts. Mutoni and Kent (2007) propose that theconcentration of dissolved silica in the relatively small North Atlantic es-tuarine basin may have increased significantly during the Early Eocene,leading to the formation of the widespread cherts. They argue that bot-tom waters warmed over ~10 myr of the Late Paleocene and EarlyEocene to peak temperatures of ~14 °C during the Early Eocene ClimaticOptimum (EECO) at ~50Ma, with chert occurrences reaching their max-imum during calcareous nannofossil zones NP11–NP15a (~54–46 Ma).Mutoni and Kent (2007) suggest that warmer bottom water tempera-tures and enhanced weathering of silicates due to an accelerated hydro-logical cycle led to increased buildup of dissolved silica in the deeperparts of the North Atlantic. They hypothesize that silica precipitation ofcherts during the EECO received an inorganic assist from clay minerals,which can take up dissolved silica and form more siliceous phases.Mutoni and Kent (2007) also speculate that a reverse conveyor belt cir-culation pattern existed, with a small North Atlantic estuarine basinlying at the end of far-traveled flow of deep waters from other basins.Hence, McGowran's (1989) argument that the widespread occurrenceof cherts in the Early Eocene oceanswas due to extensive volcanism dur-ing the earliest Eocene in the North Atlantic does not seem likely.
4.2. Diatom depositional Event 1
Diatom depositional Event 1 at ~49 Ma coincides with the onset ofthe long-term Eocene cooling trend, that followed the EECO. A declinein diatom deposition in epicontinental regions of the North Sea and

C17
C16
C15
C13
Late
Mid
dle
C18
Oligo.
CH
RO
N
PO
LAR
ITY
Ma EP
OC
H /
SU
BE
PO
CH
TropicalDiatom Zone
Eoc
ene
Ear
ly
C21
C22
C23
C24
C25
C20
C19 Hemiaulus
gondolaformis
Hemiaulus alatus
Triceratium
kanayae
Cr. oblongus
Cr. undulatus
Pyxillagracilis
Hemiaulus
incurvus
34
36
38
40
42
44
46
48
50
52
54
56
58
Paleoc.
CP
13
CP
14
a
c
b
a
b
a
CP
12
CP
11
CP
10
ba
CP7
CP
8C
P9
b
a
CP
15
CP
16 b
a
NannoZone
Pyxillacaputavis
NP21
NP19 -20
NP18
NP16
NP
15
NP
14
NP13
NP12
NP11
NP10
NP9
Cr. oblongus -Br. imperfecta
Cestodiscus pulchellusnovazeal.
Cestodiscus fennerae
Cestodiscus antarcticus
Ces. robustus
TropicalSilicoflagellate Zone
Dictyochahexacantha
Corb
isem
a apic
ula
taN
avic
ulo
psis
folia
cea
N
av.
robusta
Dic
tyocha s
pin
osa
Naviculopsis constricta
Dc. deflandre
iN
. tr
ispin
osa s
z*
?
Planktonic Foram Zone
a
b
bNP17 Triceratium inconspicuum
Rad.Zone
RP13
RP14
RP15
RP16
RP17
RP18
RP19
RP12
RP8
?
P. o
ligoc
aeni
ca te
nue
Atlantic Ocean Pacific Ocean
Arc
tic
NP8
?
RP20
Pyx
illa
grac
ilis
M. u
ral.
T v
ent.
C. p
ay.
Event 1
Event 2
E4
E3
E2E1P5
P4
E5
E6
E7
E8
E9
E10
E11
E12
E13
E14
E15
E16
O1
RP11
RP9
c
b
a
b
a
ba
?
Event 3
South -tropics- North NG. South --tropics----- North Pla
tfo
rm
?
0+3 +2 +1δ18O
0-50 50 100 150 200
Zachos et al. (2008) isotope curve (updated)
Kominz et al. (2008) sea level (updated by Ken Miller, 2013)
Sea Level Relative to Modern (m)
PETM
Oi-1 Event
Indian Ocean
MECO
?
?
Bip
ala
oam
arue
nsis
Eur
aisn
Pla
tform
?
?
Event 4
?
?
Fig. 4.Chronologic extent of keydiatom-bearing sections (Fig. 3) comparedwith the oxygen isotope curve of Zachos et al. (2008) and the global sea level curve of Kominz et al. (2008)(bothupdated to the 2012 Geologic Time Scale). Key events indicated by gray shading: PETMPaleocene–Eocene ThermalMaximum;DiatomDepositional (DD) Event 1; DDEvent 2; DDEvent 3,the Middle Eocene Climatic Optimum (MECO); DD Event 4; Oi-1, the earliest Oligocene oxygen isotope event associated with ice sheet expansion in Antarctica. (For interpretation of thereferences to color in this figure legend, the reader is referred to the web version of this article.)
92 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
central Russia marks the closure of the Turgai Strait to the Arctic Oceanat this time and appears to be associated with falling sea levels(Radionova et al., 2003).
Paleogene biosilica preservation began in the Arctic Ocean at diatomdepositional Event 1. The Arctic was largely isolated from the World'socean at this time and therefore not affected by changes in deep-water circulation. Although global cooling may have been a major con-tributory factor for enhancing preservation of opal-A on the seafloor atthis time, a combination of local factors were likely responsible for in-tensifying production and preservation.
The Eocene central Arctic was a semi-enclosed, seasonally stratifiedbasin (e.g., Backman and Moran, 2009), which acted as a nutrient andsediment trap to create a rich sediment archive that records seasonaland sub-seasonal changes in extraordinary detail. For example, thebiosiliceous sediments of Lithostratigraphic Unit 2 and subunit 1.6 ofthe IODP ACEX drillsites M0002A and M0004A on the LomonosovRidge represent one of the most important discoveries of Paleogene si-liceous microfossils of the last decade (Onodera et al., 2008; Stickleyet al., 2008, 2009). Abundant, diverse and well-preserved diatoms,silicoflagellates, ebridians and chrysophyte cysts are preserved in ca.100 m of sediment. The diatoms are a mix of marine and certain seaice taxa, whereas the ebridians indicate brackish waters, and the
chrysophytes largely freshwater. During the growing season, thesegroups were stratified in the water column (see Stickley et al. (2008;2012) for summaryfigures). It is possible that certain taxonomic diatomgroups were adapted to stratified conditions, such as Hemiaulus, andproduced frustules throughout the growing season in the deep chloro-phyll maximum. Repeated seasonal productivity-flux episodes intosilica-rich water allowed preservation of fine laminations, which reflectseasonal and sub-seasonal changes (Brinkhuis et al., 2006; Stickley et al.,2012). At the onset of biosilica preservation inUnit 2, diatomproductionmay have been stimulated by an intensification of the hydrological cycleleading to increased humic-river run-off, which contained high concen-trations of bio-available Fe, an important biolimiting nutrient for dia-toms. But from ca. 47–46.5 Ma (updated age), the influence ofseasonal sea ice (Stickley et al., 2009, 2012) played a major role in con-trolling biosilica production and preservation. The preservation of highabundances of the finely silicified sea ice diatom taxa Synedropsis spp.is unprecedented in pre-Holocene sediments and attests to the extraor-dinary environmental conditions that favored the preservation of opal-A in the Arctic at this time. Laminated Middle Eocene sediments on theAlpha Ridge (Dell'Agnese and Clark, 1994) indicate that the preserva-tion of Middle Eocene biosilica was basin-wide, and that both ridgeswere bathed in waters rich in silicic acid at this time. Stickley et al.

93J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
(2008) hypothesize that periodic flushing of Arctic deeper waters withsilica-rich waters from the North Atlantic allowed increased Si concen-trations at depth, enabling such excellent preservation.
Event 1 closely approximates seismic Reflector A (super c) in thewestern North Atlantic, which formed shortly after the end of theEarly Eocene between 48 and 49 Ma (Norris et al., 2001). Reflector Ais correlative with a sequence of unconformities present in nearlyevery part of the global ocean from the shallow shelf to the deep sea,suggesting that this time interval is associated with a global change inocean circulation, including a major sea-level lowstand (Norris et al.,2001). Hohbein et al. (2012) argue that the onset of deep-water over-flow from the Norwegian-Greenland Sea into the North Atlantic andthe onset of a modern-style North Atlantic Deep Water (NADW) masscommenced close to the Early Eocene–Middle Eocene boundary.Hohbein et al. (2012) note that dating of the onset of the “Judd FallsDrift” in the Faeroe–Shetland Basin of the North Atlantic is constrainedto have occurred between 51.5 and 48.5 Ma, but they comment thatthese dates represent maximum ages for the onset of deep overflowwater because the onset of the deep current flow likely erodedpreexisting sediments. Stoker et al. (2013), however, cite sedimentolog-ic and structural evidence in arguing that this event does not representthe onset of NADW production.
4.3. Diatom depositional Event 2
Diatomdepositional Event 2 at ~46Ma is followed by thewidespreadexpansion of diatom/silicoflagellate-bearing sections in the Atlantic andthe onset of increased biosilica accumulation in the equatorial Pacific(Moore et al., 2008). Event 2 coincides with intensification in the longterm Eocene cooling trend (Fig. 4) and possibly marks an increase inthe intensity of global deep-water circulation. Event 2 also coincideswith the end of peak chert accumulation in the North Atlantic (Mutoniand Kent, 2007), signaling a major change in silica fractionation in thedeep sea. Diatom deposition begins at about this time (~45 Ma) at ODPSite 1172 near Antarctica (Stickley et al., 2004 a, b; updated by Bijlet al., 2011). On the Eurasian platform there is a gap or hiatus at thistime (calcareous nannofossil Subzone CP13a) in diatom/silicoflagellate-bearing sections that Radionova et al. (2003) argue was due to a fall inglobal sea level.
In the Arctic ACEX corehole, there is evidence for intensification of(seasonal) sea ice- and glacial-ice rafting (St. John, 2008; Stickleyet al., 2009) as well as for Milankovitch-scale cyclicity in productivityand sedimentation (Sangiorgi et al., 2008). This time also marks thestart of an interesting phase of fluctuation between seasonal and peren-nial sea ice cover, indicating climate instability during this time (Darby,2014). The ACEX sediments during this fluctuation phase are thereforedifficult to interpret but may indicate short episodes of limited surfacewater exchange between the North Atlantic and the Arctic (Onoderaet al., 2008; Stickley, unpublished data), interspersed with sedimentsindicating heavy reworking of older material (our interpretation). Thisphase of climate instability may have been apparent in the Arctic untilthe late Middle–early Late Eocene (~36–37 Ma) based on diatom bio-stratigraphy (second author's unpublished data), after which perennialsea ice dominated the Arctic (Darby, 2014).
In the northwest North Atlantic, IODP Expedition 342 Scientists(2012) interpret the beginning of relatively clay-rich, high accumula-tion-rate (N3 cm/kyr.) sedimentation, as the onset of sediment driftdeposition, placing its age just above the NP14/NP15 calcareousnannofossil boundary, an interval correlated with the upper part of pa-leomagnetic chron C21n at both the J-Anomaly Ridge and the SoutheastNewfoundland Ridge. This correlation closely matches diatom deposi-tional Event 2 at ~46 Ma (Figs. 3, 4). The Expedition 342 Scientists(2012) argue that although the onset of drift deposition to the north(Labrador Sea andNortheast Atlantic) and to the south (Blake, Hatteras,and Chesapeake Drifts) have been thought to have occurred no earlierthan Middle to Late Eocene (~40 Ma) (Mountain and Tucholke, 1985;
Arthur et al., 1989), erosion associated with a significant regional un-conformity during the Eocene–Oligocene transition (Reflector Au)may have removed sediment possibly associated with Early to MiddleEocene drifts along the eastern US margin according to Mountain andTucholke (1985).
4.4. Diatom dispositional Event 3 (MECO)
The Middle Eocene Climatic Optimum (MECO) represents an inter-ruption in the long-term Middle Eocene cooling trend at ~40.5 Mathat was characterized by a globally uniform 4 to 6 °C warming ofboth surface and deep oceans, rise in atmospheric CO2 concentrations,prolonged carbonate dissolution in the deep oceans, and biotic changeduring a ~500 kyr-long period (Bohaty et al., 2009; Bijl et al., 2010;Sluijs et al., 2013). TheMECO occurs in the lowermost part of calcareousnannofossil subzone CP14b and lowermost paleomagnetic ChronC18.2n to uppermost C18.2r (Bohaty et al., 2009) (Fig. 4).
Moore et al. (2008) commented that radiolarian zone RP15 through19 are better preserved in the Pacific and Indian Oceans, whereas zonesRP6 through 13 are better preserved in the Atlantic–Caribbean. Theseauthors speculated that these preservational differences may havebeen due to basin–basin fractionation of nutrients, and they cited theNd isotope studies of Thomas (2004) in arguing that a basin-to-basinshift fromAtlantic to Pacific deposition of biogenic silica (opal) occurredduring radiolarian zones RP14 and 15 (~43–40 Ma, Fig. 3).
An increase of diatom/silicoflagellate deposition in coastal upwellingsettings appears to have occurred immediately after the MECO, namelyin the Oceanic Formation of Barbados (Saunders et al., 1984), theKreyenhagen Shale of California (McLean and Barron, 1988), and in on-shore Peru (Marty et al., 1988). Bosboom et al. (2014) document an in-crease in aridification in continental Asia immediately after the MECO,which supports an increase in pole-to-equator temperature gradientsthat should lead to increased winds and coastal upwelling of nutrientsthat triggered diatom blooms.
A number of detailed studies of diatom assemblages across theMECO have recently been published. Renaudie et al. (2010) studiedquantitative changes in diatom and silicoflagellate assemblages duringa Middle Eocene warming event that they attributed to the early partthe MECO warming event at ODP Site 1260A from the Demerara Risein the western tropical Atlantic off South America. They documentedthe replacement of a cosmopolitan diatom flora dominated byTriceratium by a new and more diverse diatom assemblage during thisevent that was accompanied by a steady increase in biogenic silica.However, thewarm event studied by Renaudie et al. (2010) has recent-ly been correlated to paleomagentic Chron 19r (Westerhold and Röhl,2013) and nearly 2 myr older than the MECO event.
Witkowski et al.'s (2012) high resolution, quantitative study ofbiosiliceous sedimentation at ODP Sites 748 and 749 on Kerguelen Pla-teau (Southern Ocean) also revealed that a significant increase inbiosilica sedimentation was associated with the MECO; however,these authors note that this increase in biosilica comprised mainlyebridians and radiolarians. Witkowski et al. (2012) observed that allbiosiliceous groups experienced peak MECO warming by the influx ofcosmopolitan, tropical and sub-tropical species. Further studies byWitkowski et al. (2014) at subtropical eastern North Atlantic ODP Site1051 revealed relatively low rates of pelagic siliceous phytoplanktonsedimentation accompanied the peak MECO warming during a~70 kyr interval centered at ~40 Ma. Immediately following the termi-nation of theMECO, a 200 kyr-long interval of increased siliceous plank-ton abundance was recorded.
Witkowski et al. (2012) document commonoccurrences of the large,distinctive silicoflagellate Dictyocha grandis immediately prior to theMECO in Kerguelen Plateau ODP Site 748. Shaw and Ciesielski (1983)record many varieties of D. grandis throughout calcareous nannofossilsubzone CP14a at DSDP Site 512 on the Falkland Plateau. Perch-Nielsen (1975) illustrates D. grandis as D? sp. 6 from DSDP 277 in the

94 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
southwest Pacific, a likely reworked occurrence of this distinctive Mid-dle Eocene silicoflagellate in Oligocene sediments. It would appearthat D. grandis might be an excellent biostratigraphic marker in theSouthern Ocean for the CP14a interval and the MECO.
4.5. Diatom depositional Event 4 ~39 Ma
Diatom depositional Event 4 occurs at the CP14/CP15 (= NP17/NP18) calcareous nannofossil boundary in the uppermost part of paleo-magnetic chron C18.1n at ~39 Ma (Figs. 3, 4). It coincides with a majorsea level fall (Fig. 4) and with a widespread deep-sea hiatus document-ed by Keller et al. (1987) and Wade and Kroon (2002). Dawber andTripati (2011) detail a N+0.5‰ increase in benthic δ18O values in paleo-magnetic chron C18.1n in equatorial Pacific ODP Site 1209, which theyrelate to Antarctic glaciation. We update their ages of 39.0–38.5 Mafor this event to ~39.3–38.8 Ma. Egan et al. (2013) argue that openingof the Drake Passage and Tasman Gateway beginning in the Late Eoceneand intensifying in the earliest Oligocene lead to increased diatomabundance in the circum Antarctic that contributed to pCO2 drawdownand global cooling that possibly triggered expandedAntarctic glaciation.Borrelli et al. (2014) cite oxygen and carbon isotopic data from benthicforaminifers in arguing that the onset of Northern Component Wateroccurred at ~38.5 Ma. Eldrett et al. (2007) document stratigraphicallyextensive ice-rafted debris in northern Norwegian-Greenland (NG)Sea ODP Site 913 that begins in uppermost paleomagnetic chronC18n.1n (38.8 Ma updated age) and extends into the Lower Oligocene(~30 Ma). They argue that the provenance of the igneous clasts foundin the Upper Eocene section of ODP 913 is consistent with a sourcefrom the Paleogene flood basalts of East Greenland, implying the pres-ence of glacial ice on that coast.
We argue (see Backman et al. (2008) and discussion in Poirier andHillaire-Marcel (2011)) that ACEX Subunit 1/6 is latest Middle to earlyLate Eocene in age (~38.4–36.4 Ma, updated ages) based on correlationof its diatom and silicoflagellate assemblages with ODP 913 in the north-ern NG Sea. In particular, the diatoms Actinoptychus irregularis andCoscinodiscus sp. aff. C. tenerrimus and the silicoflagellate Dictyochahexacantha are not present in middle-to-high latitude assemblagesolder than about 38 Ma. If such a correlation is correct, it is possible thata hiatus (or period of extremely slow deposition) exists on LomonosovRidge between diatom depositional Events 2 and 4 (~46–39 Ma).
Diekmann et al. (2004) completed detailed studies on both the bio-genic proxies (carbonate, opal, and organic carbon) and the lithogenicfraction between the Middle Eocene and late Early Miocene at ODPSite 1090 in the southeast South Atlantic. They recorded a Late Eocene(~37.5–33.5 Ma, ages updated) interval of increased opal values (withweight percent values ranging from ~20 to N60%), which they sug-gested was due to an increase in upwelling along topographic highs.This pulse in opal deposition equates to a distinct increase in biogenicaccumulation rates andmay be indicative of a firstmassive opal deposi-tion in the northern parts of the Southern Ocean.
5. The role of ocean gateways
The modern distribution of oceanic diatom and silicoflagellate-richsediments (Fig. 1) is a function of the present day locations of continentsand oceanic gateways, a strong pole-to-equator temperature gradientwhich results in intensified equatorward winds that lead to enhancedcoastal upwelling and diatom blooms along the eastern margin ofocean basins, and the presence of thermohaline circulation which en-hances biosilica preservation in the Pacific compared to the Atlanticdue to basin–basin fractionation (Berger, 1970).
Most of these conditions were not present in the Eocene, and theglobal distribution of diatom/silicoflagellate-bearing sediments wasquite different (e.g., see reconstructions of Barron and Baldauf (1989)and Baldauf and Barron (1990)). Fig. 5 plots the distribution of Eocenediatom/silicoflagellate-bearing sections on plate tectonic reconstruction
maps for 50 and 40 Ma (after the reconstructions of Hay et al. (1999);http://www.odsn.de/odsn/services/paleomap/paleomap.html). LowerEocene to lower Middle Eocene (58–45 Ma) sections are plotted onthe 50 Ma reconstruction map, whereas upper Middle and UpperEocene sections are plotted on the 40 Ma map. Dashed circles indicatemajor oceanic gateways, including the Drake Passage (DP), TasmanGateway (TG), Arctic Gateway to the North Atlantic (AG), and TurgaiStrait (TS) entrance to the Arctic. The widespread global distributionof diatom/silicoflagellate-bearing sections at 40 Ma contrasts with themore restricted distribution at 50 Ma.
5.1. Arctic–Atlantic gateway
The Arctic gateway to the Atlantic involves a complex geological his-tory. During the Cretaceous and Paleogene, the Arctic was largely isolat-ed from the World Ocean; the proto-Norwegian Greenland Sea was along and narrow, shallow seaway that partially connected the Arcticwith the Atlantic through surface-water exchange. For example, MiddleEocene surface-water exchange is indicated by (albeit rare) occurrencesof Pyxilla oligocaenica and Brightwellia hyperborea, in the Arctic ACEXmaterial. These are age-diagnostic diatoms from the NG and NorthAtlantic respectively (Dzinoridze et al., 1978; Gombos, 1982, 1987;Fenner, 1985). Further evidence for Arctic isolation is provided byhigh abundances of the freshwater fern Azolla spp. in the same sectionof core (Brinkhuis et al., 2006). The wider and deeper modern-day ex-change with the Atlantic evolved through a series of tectonic stepswhich can be broadly summarized into two phases: (1) the timing ofthe opening/deepening of the Fram Strait—the narrow conduit betweenthe Arctic and the proto-NG sea in the north, and (2) that of the south-ern end of the proto-sea with the Atlantic. The southern end likelyopened/deepened first. For example, Hohbein et al. (2012) argue foran Early Eocene–Middle Eocene boundary (~50–49 Ma) onset ofdeep-water overflow from the Norwegian-Greenland Sea into theAtlantic, which they interpret as a modern-style North Atlantic DeepWater (NADW)mass. As the Fram Strait is the only deep-water connec-tion between the Arctic and the world ocean, the timing of the forma-tion of this passage is critical for global circulation models andunderstanding the consequences of bottom water exchange betweenthe Arctic and the world ocean via the (proto-) Norwegian-GreenlandSea. Several tectonic reconstructions have been put forward for thetiming of the opening of the Fram Strait ranging from Early Oligoceneto Late Miocene, culminating in the most recent study by Jakobssonet al. (2007) who indicate Fram Strait deepening and widening by17.5 Ma (Early Miocene) that allowed a complete change from anoxygen-poor isolated basin to full Arctic ventilation and exchange ofdeep waters with the proto-NG sea and enhanced Atlantic influence.
5.2. The Southern Ocean
Both the opening of the TasmanianGateway between Antarctica andAustralia and the Drake Passage between Antarctica and South Americawere critical to the development of the Southern Ocean and the biogen-ic silica belt (Fig. 1).
The Tasmanian Gateway opened and widened in a series of stepswith the main event occurring 2 myr prior to Early Oligocene Antarcticglaciation (Stickley et al., 2004b). The influence of themainly eastboundthrough-flow (or proto-ACC) did not act to cool Antarctica but rather to(temporarily) warm it along the East Antarctic Margin in the EarlyOligocene (Huber et al., 2004; Stickley et al., 2004b). The initiation ofthis early countercurrent coincided with both regional surface waterand continental cooling (2–4 °C). Recently, Bijl et al. (2013) arguedthat a westbound circum Antarctic surface water counter-current com-menced near the Early Eocene–Middle Eoceneboundary (ca. 50–49Ma)during the opening of the southern end of the Tasmanian Gateway.
Livermore et al. (2007) review the tectonic and paleoceanographicevidence for the opening of the Drake Passage. Tectonic evidence

180˚
180˚
210˚
210˚
240˚
240˚
270˚
270˚
300˚
300˚
330˚
330˚
0˚
0˚
30˚
30˚
60˚
60˚
90˚
90˚
120˚
120˚
150˚
150˚
180˚
180˚
−90˚ −90˚
−60˚ −60˚
−30˚ −30˚
0˚ 0˚
30˚ 30˚
60˚ 60˚
90˚ 90˚
40.0 Ma Reconstruction
180˚
180˚
210˚
210˚
240˚
240˚
270˚
270˚
300˚
300˚
330˚
330˚
0˚
0˚
30˚
30˚
60˚
60˚
90˚
90˚
120˚
120˚
150˚
150˚
180˚
180˚
−90˚ −90˚
−60˚ −60˚
−30˚ −30˚
0˚ 0˚
30˚ 30˚
60˚ 60˚
90˚ 90˚
50.0 Ma Reconstruction
cherts
DPTG
AG
AG
390A
94
752A
553
343ACEX
338913
206C
1171, 281
283Oamaru
KamchatkaCalifornia
Baja
Peru
Barbados
748
1219163
167149
356
512 1090A
386
3661260
707
713
402A, 406
289
cherts
U1403A
385
511
605612
1051
SHAL.3C
11721166
U1356A TG
Fur/ North SeaTS
Eurasian Platform
Eurasian Platform
Fig. 5. Global distribution of key Eocene diatom/silicoflagellate-bearing sections (red, ocean cores; blue, onshore) plotted on 50 Ma an 40 Ma plate tectonic reconstructions after http://www.odsn.de/odsn/services/paleomap/paleomap.html. Major oceanic gateways indicated by dashed circles — TG, Tasman Gateway; DP, Drake Passage; AG, Arctic Gateway; TS, TurgaiStrait.
95J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
suggests an increase in seafloor spreading rates in the proto-Drake Pas-sage region at ~50 Ma. The first opening of the gateway was limited towater depth b1000 m over submerged continental shelves. Livermoreet al. (2007) note that crustal stretching and subsidence characterizedthe interval from 50 to 34 Ma. Studies of Nd isotopes in fossil fishteeth fromODP sites 689 and 1090 in the Atlantic sector of the SouthernOcean reveal significant increases in the ratio of radiogenic to non-radiogenic neodymium (143Nd/144Nd) between ~41 and 34 Ma whichScher and Martin (2004, 2006) argue reflects an influx of PacificOcean seawater resulting from the earliest opening of the DrakePassage. Recently, Borrelli et al. (2014) cite oxygen and carbon isotopicdata from the North and South Atlantic and equatorial Pacific in arguingthat the Drake Passage had opened sufficiently at ~38.5 Ma to cause theonset of Northern Component Water and the onset of modern thermo-haline circulation.
In summary, the development of circum-Antarctic surfacewater flowin the Southern Ocean appears to have occurred in stages beginning at~49–50 Ma and increasing between ~41 and 38 Ma that likely affectedthe global distribution of diatom/silicoflagellate-bearing sediments. Thetemporal (Fig. 3) and geographic (Fig. 5) distribution of upper Middleand Upper Eocene diatom and silicoflagellate-bearing sections arguesthat the latter events (~41 to 38 Ma) had a more pronounced effect onthe global distribution of these biosiliceous sediments, a result that issupported by the radiolarian synthesis study of Moore et al. (2008).
The appearance of post 39 Ma biosiliceous sediments at ODP 913(Fig. 3) reflects opening of the northern Norwegian-Greenland Sea(Scherer and Koc, 1996), an important precursor to the establishmentof an Arctic–Atlantic Gateway. However, there is no compelling evi-dence that a more northerly surface water connection with the Arcticexisted at that time. This may be in part to the predominance of peren-nial sea ice conditions (Darby, 2014) and in part to tectonic activity,i.e., stalled Late (?) Eocene–Oligocene submergence of the LomonosovRidge resulting in a suspected long hiatus due to un-roofing and erosionof the ridge crest (e.g., O'Regan et al., 2008), that has hampered a full in-vestigation into basin–basin coeval comparison (e.g., Backman andMoran, 2009).
6. Response of diatom taxa
Howdid diatom taxa respond to Eocene paleoceanographic change?Although quantitative changes in diatom assemblages have largely onlybeen documented for theMECO event, the present compilation of globalEocene diatombiostratigraphydoes allow someadditional observationsto be made. Age estimates for the first and last occurrences of 132 dia-tom taxa useful for Eocene biostratigraphy are compiled on Supplemen-tary Table 1. References for the age constraints of these datum levels aretabulated in Supplementary Table 2. These tables are updated from theBarronDiatomCatalog “BDC”whichwasfirst published as a supplement

96 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
to Lazarus et al. (2014). As with the BDC, the Eocene diatom events listpresented in Supplementary Table 1 has been compiled from a variety(n = 30) of literature sources (Supplementary Table 2), which includeprimary biostratigraphic studies of key sections and syntheses such asFenner (1985) and Scherer et al. (2007). It also includes age estimatesderived from the Neptune database of DSDP and ODP studies (Spencer-Cervato, 1999). The original data was corrected for taxonomic synony-mies and an effort was made to mostly include biostratigrapically-useful taxa suggested by the syntheses of Fenner (1985) and Oreshkina(2012). Consequently, numerous taxa documented only in monographson key stratigraphic sections (e.g., the Fur Formation (Homann, 1991;Fenner, 1994) and Oamaru Diatomite (Edwards and DSIR Geology andGeophysics, 1991)) are not included. Similarly, the largely endemicArctic diatom floras of the ACEX core (Onodera et al., 2008; Suto et al.,2009) are not included in this table. Thus, Supplementary Table 1 of dia-tom species events is both preliminary and limited.
Fig. 6 plots the first and last occurrences of 132 biostratigraphically-important diatoms compiled for 0.5myr-long increments and comparesthemwith theupdated oxygen isotope curve of Zachos et al. (2008). The
C17
C16
C15
C13
Late
Mid
dle
C18
Oligo.
CH
RO
N
PO
LAR
ITY
Ma EP
OC
H /
SU
BE
PO
CH
TropicalDiatom Zone
Eoc
ene
Ear
ly
C21
C22
C23
C24
C25
C20
C19 Hemiaulus
gondolaformis
Hemiaulus alatus
Triceratium kanayae
Cr. oblongus
Cr. undulatus
Pyxillagracilis
Hemiaulus
incurvus
34
36
38
40
42
44
46
48
50
52
54
56
58
Paleoc.
CP
13
CP
14
a
c
b
a
b
a
CP
12
CP
11
CP
10
ba
CP7
CP
8C
P9
b
a
CP
15
CP
16 b
a
NannoZone
Pyxillacaputavis
NP21
NP19 -20
NP18
NP16
NP
15
NP
14
NP13
NP12
NP11
NP10
NP9
Cr. oblongus -Br. imperfecta
Cestodiscus pulchellusnovazeal.
Cestodiscus fennerae
Cestodiscus antarcticus
Ces. robustus
Planktonic Foram Zone
a
b
bNP17 Triceratium inconspicuum
Rad.Zone
RP13
RP14
RP15
RP16
RP17
RP18
RP19
RP12
RP8
?
Pyx
illa
olig
ocae
nica
v. t
enue
NP8
RP20
Pyx
illa
grac
ilis
M. u
ral.
T v
ent.
C. p
ay.
E4
E3
E2E1P5
P4
E5
E6
E7
E8
E9
E10
E11
E12
E13
E14
E15
E16
O1
RP11
RP9
c
b
a
b
a
ba
?
# Diatom
First Occurre
0 2 4 6
Fig. 6. First and last occurrences of biostratigraphically-important diatoms (Supplementary Tabof Zachos et al. (2008). Key climatic events indicated by gray shading: PETMPaleocene–Eocene TEocene Climatic Optimum(MECO); DDEvent 4; Oi-1, the earliest Oligocene oxygen isotope evecolor in this figure legend, the reader is referred to the web version of this article.)
concentration of events that cross the Paleocene–Eocene boundary be-tween ~58 and 55Ma likely reflects both the number of detailed studiesdone on this interval (e.g., Homann, 1991; Fenner, 1994; Radionovaet al., 2003; Oreshkina, 2012) and the environmental effects of thePETM (Oreshkina and Oberhänsli, 2003).
The scarcity of diatom events during the earliest Eocene to earlyMiddle Eocene interval between ~53 and 47 Ma may be an artifact ofthe relatively smaller number of stratigraphic sections (Fig. 3) thathave been studied in detail for this interval (Table 2) as well as thewidespread occurrence of cherts in the deep North Atlantic (Mutoniand Kent, 2007). However, Lazarus et al. (2014) indicate a steadyincrease in global planktonic diatom diversity for this interval.
Diatom first occurrences increase abruptly to six during the 46.5–46.0 Ma interval associated with diatom depositional Event 2 (Fig. 6),possibly reflecting increased diatomdiversity associatedwithmore pro-nounced cooling trend in the oxygen isotope curve. Strelnikova (1990)argued that themajor turnover in Eocenediatom species occurred at theEarly Eocene–Middle Eocene boundary; however, her data was com-piled at much coarser temporal resolution than Fig. 6.
Event 1
Event 2
Event 4
Event 3MECO
δ18O
PETM
Oi-1 Event
Events
nces Last Occurrences
Zachos et al. (2008) isotope curve (updated)
0+1+28 10 12 0 2 4 6 8# Diatom Events
10
le 1) tabulated at 0.5 myr-long intervals compared with the updated oxygen isotope curvehermalMaximum;DiatomDepositional (DD) Event 1;DDEvent 2;DDEvent 3, theMiddlent associatedwith ice sheet expansion in Antarctica. (For interpretation of the references to

97J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
A more regular succession of diatom first and last occurrences after~42 Ma may be partly due to the greater number of stratigraphic sec-tions that have been studied for this interval (Fig. 3, Table 1); however,an increasing pole-to-equator thermal gradient and enhanced coastalupwelling undoubtedly triggered increased diatom diversity and turn-over during the latest Middle and Late Eocene (Lazarus et al., 2014).An increase in diatom last occurrences to four during the 40.0–39.5 Ma interval is consistent with Witkowski et al.'s (2012) observa-tion that diatom diversity peaked during the MECO. Diatom speciesturnover increased markedly at ~35 Ma during the latest Eocene andcontinues through the earliest Oligocene.
7. Conclusions
Global Eocene diatom and silicoflagellate biostratigraphic zones arecalibrated against the geologic time scale of Gradstein et al. (2012) pri-marily through correlation with calcareous nannofossil and radiolarianbiostratigraphy, although a limited number of paleomagnetic strati-graphic studies provide direct calibration in selected intervals. Whencompared with the global oxygen isotope and eustatic sea level curve,the chronologic extent of 48 diatom/silicoflagellate bearing stratigraph-ic sections (on land and deep sea) reveals a distinctly regional pattern ofglobal distribution that responded to changing paleoceanographic andpaleoclimatic conditions and was punctuated by fourmajor deposition-al events.
Warm surface and deepwaters that were poorly mixed by relativelyweak zonal winds characterized the Early Eocene, resulting from re-duced pole-to-equator temperature gradient. Diatom/silicoflagellatedeposition was concentrated on epicontinental platforms in westernSiberia, Russia and in the North Sea region and along the eastern conti-nental margins of North America. Widespread chert deposition in thedeep North Atlantic was likely due to chemical precipitation associatedwith warmer bottom water temperatures and enhanced weathering ofsilicates.
Depositional Event 1 (~49Ma) at the endof the Early Eocene Climat-ic Optimum (EECO), saw the cessation of diatom/silicoflagellate deposi-tion in epicontinental regions of North Sea region and the northernparts of central Russiamost likely because of falling sea level. The abruptonset of biosilica deposition in the Arctic also occurred at Event 1,whichis likely caused by changes in local factors and enhanced seasonalchanges. Although recent studies (Bijl et al., 2013) suggest that the Tas-manian Gateway between Australia and Antarctica may have begun toopen at this time (50–49 Ma), limited evidence (Stickley et al., 2004a,ODP Site 1172) suggests that substantial biogenic silica depositionalong the margin of Antarctica began at ~45 Ma.
Depositional Event 2 (~46Ma), which coincidedwith intensificationof the Middle Eocene cooling trend, marked the widespread expansionof diatom/silicoflagellate deposition in both the North and South Atlan-tic, increased biosilica accumulation in the equatorial Pacific (Mooreet al., 2008), and an apparent increase in diatom deposition in the mar-ginal seas around Antarctica (Stickley et al., 2004a, Fig. 3).
Depositional Event 3 at the end of the Middle Eocene ClimaticOptimum (MECO) (~40 Ma), marked the beginning of a shift ofdiatom/silicoflagellate deposition from the Atlantic to the Pacificthat was likely tied to the opening of the Drake Passage betweenAntarctica and South America. An increase of diatom/silicoflagellate de-position in coastal upwelling settings appears to have occurred immedi-ately after theMECO, namely in the Oceanic Formation of Barbados, theKreyenhagen Shale of California, and in onshore Peru. An increase inpole-to-equator temperature gradients leading to increased winds andcoastal upwelling at this time is supported by Bosboom et al.'s (2014)documentation of enhanced aridification in continental Asia.
Depositional Event 4 (~39 Ma) coincided with a major sea level falland a widespread deep-sea hiatus in the latest Middle Eocene. Dawberand Tripati (2011) detail a N+0.5‰ increase in benthic δ18O values inpaleomagnetic chron C18n1n (updated ages ~39.3–38.5 Ma) in
equatorial Pacific ODP Site 1209, which they relate to Antarctic glacia-tion. Borrelli et al. (2014) cite oxygen and carbon isotopic data frombenthic foraminifers in arguing that the onset of Northern ComponentWater occurred at ~38.5 Ma. The shift of diatom/silicoflagellate deposi-tion fromwidespread areas of the Atlantic to modern coastal upwellingand high-latitude upwelling regions (mostly in the Pacific and SouthernOcean) was essentially completed by Event 4.
Tabulation of thefirst and last occurrences of 132 biostratigraphically-important diatoms reveals that Eocene diatom species turnover in-creased markedly after ~40 Ma likely in response to the cooling ofpolar regions and increased provinciality. Large species turnover oc-curred across the PETM, although it may result in part from a mono-graphic effect that witnessed more detailed studies of key stratigraphicsections during the ~58–54Ma interval. Similarly, the scarcity of diatomevents between ~54 and 46.5Mamay be linkedwith the limited numberof biostratigraphic studies during this interval, although low speciesturnover would be expected during this warm interval of the EarlyEocene Climatic Optimum. Diatom first occurrences increase in numberand regularity beginning at ~46 Ma coincident with intensification ofthe cooling trend of the Eocene oxygen and with diatom depositionalEvent 2 which saw the widespread expansion of diatom deposition inthe world's oceans.
Acknowledgments
The Micropalaeontological Society (UK) inspired the Eocene re-search of the senior author by awarding him the Brady Medal in 2011and then enthusiastically receiving his lecture on the Eocene history ofmarine diatoms. Part of Catherine Stickley's research was partly fundedthrough the Research Council of Norway and Statoil ASA while she wasat the Department of Geology, University of Tromsø and NorwegianPolar Institute, Tromsø, Norway. The research of Barron and Bukrywas funded by the Climate and Land Use, Research and DevelopmentProgram. We thank Ken Miller for providing an updated copy of theKominz et al. (2008) sea level curve. This manuscript benefitted frominitial reviews by Scott Starratt, Andy Gombos, Jakub Witkowski, andTatiana Oreshkina and from the editorial review of Michael Clynne.Jakub and Tatiana contributed new information that improved themanuscript's interpretations. We are also grateful to our journal re-viewers, Andrey Gladenkov and an anonymous reviewer, for their cor-rections and excellent suggestions.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.palaeo.2015.01.015.
References
Arthur, M.A., Srivastava, S., Kaminsky, M., Jarrard, R., 1989. Seismic stratigraphy and his-tory of deep circulation and sediment drift development in Baffin Bay and the Labra-dor Sea. Proceedings of the Ocean Drilling Program, Scientific Results 105 (2),957–988.
Backman, J., Moran, K., 2009. Expanding the Cenozoic paleoceanographic record in thecentral Arctic Ocean: IODP Expedition 302 synthesis. Central Eur. J. Geosci. 1 (2),157–175.
Backman, J., et al., 2008. Age model and core‐seismic integration for the Cenozoic ArcticCoring Expedition sediments from the Lomonosov Ridge. Paleoceanography 23,PA1S03. http://dx.doi.org/10.1029/2007PA001476.
Baldauf, J.G., 1984. Cenozoic diatom biostratigraphy and paleoceanography of the RockallPlateau Region, North Atlantic, Deep Sea Drilling Project Leg 81. Initial Rep. Deep SeaDrill. Proj. 81, 439–478.
Baldauf, J.G., Barron, J.A., 1990. Evolution of biosiliceous sedimentation patterns for theEocene through Quaternary: paleoceanographic response to polar cooling. In:Thiede, J., J., Beil, U. (Eds.), Geologic History of the Polar Oceans: Arctic versusAntarctic. Kluwer Academic Publication Group: Dordrecht, Holland, pp. 575–608.
Barron, J.A., 2003. Appearance and extinction of planktonic diatoms during the past18 m.y. in the Pacific and Southern oceans. Diabetes Res. 18 (2), 203–224.
Barron, J.A., Baldauf, J.G., 1989. Tertiary cooling steps and paleoproductivity as reflected bydiatoms and biosiliceous sediments. In: Berger, W.H., Smetacek, V.S., Wefer, G. (Eds.),

98 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
Productivity of the Ocean: Present and Past, Dahlem Konferenzen. J Wiley and Sons,Ltd., Chichester, UK, pp. 341–354.
Barron, J.A., Bukry, D., Poore, R.Z., 1984. Correlation of the Middle Eocene Kellogg Shale ofnorthern California. Micropaleontology 30, 138–170.
Barron, J.A., Bukry, D., Gersonde, R., 2014. Diatom and silicoflagellate biostratigraphy forthe Late Eocene: ODP 1090 (sub-Antarctic Atlantic). In: Kociolek, J.P. (Ed.), NovaHedwigia, Beihefte 143. J. Cramer, Stuttgart, Germany, pp. 1–31.
Berger, W.H., 1970. Biogenous deep-sea sediments: fractionation by deep-sea circulation.Bull. Geol. Soc. Am. 81, 1385–1402.
Berggren, W.A., Pearson, P.N., 2005. A revised tropical to subtropical Paleogene planktonicforaminiferal zonation. J. For. Res. 35, 279–298.
Bijl, P.K., Houben, A.J.P., Schouten, S., Bohaty, S.M., Sluijs, A., Reichart, G.-J., SinnigheDamsté, J.S., Brinkhuis, H., 2010. Transient Middle Eocene atmospheric CO2 and tem-perature variations. Science 330, 819–821.
Bijl, P.K., Pross, J., Warnaar, J., Stickley, C.E., Huber, H., Guerstein, R., Houben, A.J.P., Sluijs,A., Visscher, H., Brinkhuis, H., 2011. Environmental forcings of Paleogene SouthernOcean dinoflagellate biogeography. Paleoceanography 26. http://dx.doi.org/10.1029/2009PA001905.
Bijl, P.K., Bendle, J.A.P., Bohaty, S.M., Pross, J., Schouten, S., Tauxe, L., Stickley, C.E., McKay,R.M., Röhl, U., Olney, M., Sluijs, A., Escutia, C., Brinkhuis, H., 2013. Eocene coolinglinked to early flow across the Tasmanian Gateway. Proc. Natl. Acad. Sci. U. S. A.110 (24), 9645–9650.
Bohaty, S.M., Zachos, J.C., 2003. Significant Southern Ocean warming event in the lateMiddle Eocene. Geology 31, 1017–1020.
Bohaty, S.M., Zachos, J.C., Florindo, F., Delaney, M.L., 2009. Coupled greenhouse warmingand deep-sea acidification in the Middle Eocene. Paleoceanography 24, PA2207.http://dx.doi.org/10.1029/2008PA001676.
Bohaty, S.M., Kulhanek, D.K., Wise Jr., S.W., Jemison, K., Warny, S., Sjunneskog, C., 2011.Age Assessment of Eocene–Pliocene Drill Cores Recovered During the SHALDRIL IIExpedition, Antarctic Peninsula, pp. 63–113. In: Anderson, J.B., Wellner, J.S. (Eds.),Tectonic, Climatic, and Cryospheric Evolution of the Antarctic Peninsula. AGU,Washington, D. C. http://dx.doi.org/10.1029/SP063 (218 pp.).
Borrelli, C., Cramer, B.S., Katz, M.E., 2014. Bipolar Atlantic deep water circulation in themidde–Late Eocene: effects of Southern Ocean gateway openings. Paleoceanography29. http://dx.doi.org/10.1002/2012PA002444.
Bosboom, R.E., Abels, H.A., van den Hoorn, C., Berg, B.C.J., Guo, Z., Dupont-Nivet, G., 2014.Aridification in continental Asia after the Middle Eocene Climatic Optimum (MECO).Earth Planet. Sci. Lett. 389 (2014), 34–42.
Brinkhuis, H., et al., 2006. Episodic fresh surface waters in the Early Eocene Arctic Ocean.Nature 441, 606–609. http://dx.doi.org/10.1038/nature04692.
Bukry, D., 1973a. Low-latitude coccolith biostratigraphic zonation. Initial Rep. Deep SeaDrill. Proj. 15, 685–703.
Bukry, D., 1973b. Coccolith and silicoflagellate stratigraphy, Tasman Sea and southwest-ern Pacific Ocean, Deep Sea Drilling Project Leg 21. Initial Rep. Deep Sea Drill. Proj.21, 885–893.
Bukry, D., 1975a. Coccolith and silicoflagellate stratigraphy, northwestern Pacific Ocean.Initial Rep. Deep Sea Drill. Proj. 32, 677–701.
Bukry, D., 1975b. Silicoflagellate and coccolith stratigraphy, Deep Sea Drilling Project, Leg29. Initial Rep. Deep Sea Drill. Proj. 29, 845–870.
Bukry, D., 1976. Silicoflagellate and coccolith stratigraphy, Norwegian-Greenland Sea,Deep Sea Drilling Project Leg 38. Initial Rep. Deep Sea Drill. Proj. 38, 843–855.
Bukry, D., 1977. Coccolith and silicoflagellate stratigraphy, South Atlantic Ocean, Deep SeaDrilling Project Leg 39. Initial Rep. Deep Sea Drill. Proj. 39, 825–839.
Bukry, D., 1978a. Cenozoic silicoflagellate and coccolith stratigraphy, Northwestern Atlan-tic Ocean, Deep Sea Drilling Project Leg 43. Initial Rep. Deep Sea Drill. Proj. 44,775–805.
Bukry, D., 1978b. Cenozoic coccolith, silicoflagellate, and diatom stratigraphy, Deep SeaDrilling Project Leg 44. Initial Rep. Deep Sea Drill. Proj. 44, 807–863.
Bukry, D., 1981. Synthesis of silicoflagellate stratigraphy for Maastrichtian to Quaternarymarine sediment. SEPM Spec. Publ. 32, 433–444.
Bukry, D., 1984. Cenozoic silicoflagellates from Rockall Plateau, Deep Sea Drilling ProjectLeg 81. Initial Rep. Deep Sea Drill. Proj. 81, 547–563.
Bukry, D., 1987. Eocene siliceous and calcareous phytoplankton, Deep Sea Drilling ProjectLeg 95. Initial Rep. Deep Sea Drill. Program 95, 395–415.
Bukry, D., Foster, J.H., 1974. Silicoflagellate zonation of Upper Cretaceous to lower Mio-cene deep-sea sediment. Journal Research U.S. Geological Survey 2, 303–310.
Cramer, B.S., Toggweiler, J.R., Wright, J.D., Katz, M.E., Miller, K.G., 2009. Ocean overturningsince the Late Cretaceous: inferences from a new benthic foraminiferal isotope com-pilation. Paleoceanography 24, PA4216. http://dx.doi.org/10.1029/2008PA001683.
Darby, D.A., 2014. Ephemeral formation of perennial sea ice in the Arctic Ocean during theMiddle Eocene. Nat. Geosci. 7, 210–213. http://dx.doi.org/10.1038/NGEO2068.
Dawber, C.F., Tripati, A.K., 2011. Constraints in theMiddle Eocene (46–37Ma) from OceanDrilling Program (ODP) Site 1209 in the tropical Pacific Ocean. Paleoceanography 26,PA2208. http://dx.doi.org/10.1029/2010PA002037.
Dell'Agnese, D., Clark, D.L., 1994. Siliceous microfossils from the warm Late Cretaceousand Early Cenozoic Arctic Ocean. J. Paleontol. 68, 31–46.
Diekmann, B., Kuhn, G., R. Gersonde, R., Mackensen, A., 2004. Middle Eocene to EarlyMiocene environmental changes in the sub-Antarctic Southern Ocean: evidencefrom biogenic and terrigenous depositional patterns at ODP Site 1090. Glob. Planet.Chang. 40, 295–313.
Dumoulin, J.A., 1984. Silicoflagellate biostratigraphy and paleoecology of the upperKreyenhagen Formation (Eocene–Oligocene), California. In: Blueford, J.R. (Ed.),Kreyenhagen Formation and related rocks, Pacific Section, Society of EconomicPaleontologists and Mineralogists, pp. 29–49.
Dunbar, R.B., Marty, R.C., Baker, P., 1990. Cenozoic marine sedimentation in the Sechuraand Pisco basins, Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 77 (3-4), 235–261.
Dzinoridze, R.N., Jousé, A.P., Koroleva-Golikova, G.S., Kozlova, G.E., Nagaeva, G.S.,Petrushevskaya, M.G., Strelnikova, N.I., 1978. Diatom and radiolarian Cenozoic stra-tigraphy, Norwegian Basin; DSDP Leg 38. Initial Rep. Deep Sea Drill. Proj. Supplementvolume 38-41, 289–427.
Edgar, K.M., Wilson, P.A., Sexton, P.F., Gibbs, S.J., Roberts, A.P., Norris, R.D., 2010. New bio-stratigraphic, magnetostratigraphic and isotopic insights in the Middle EoceneClimatic Optimumin low latitudes. Palaeogeography, Palaeoclimatology, Palaeoecology297, 670–682. http://dx.doi.org/10.1016/j.palaeo.2010.09.016.
Edwards, A.R., DSIR Geology and Geophysics, 1991. The Oamaru Diatomite. N. Z. Geol.Surv. Paleontol. Bull. 64 (260 pp.).
Egan, K.E., Rickaby, R.E.M., Hendry, K.R., Halliday, A.N., 2013. Opening the gateways for di-atoms primes Earth for Antarctic glaciation. Earth Planet. Sci. Lett. 375, 34–43. http://dx.doi.org/10.1016/j.epsl.2013.04.030.
Eldrett, J.S., Harding, I.C., Firth, J.V., Roberts, A.P., 2004. Magnetostratigraphic calibration ofEocene–Oligocene dinoflagellate biostratigraphy from the Norwegian-Greenland Sea.Mar. Geol. 204, 91–127.
Eldrett, J.S., Harding, I.C., Wilson, P.A., Butler, E., Roberts, A.P., 2007. Continental ice inGreenland during the Eocene and Oligocene. Lett. Nat. 446. http://dx.doi.org/10.1038/nature05591 (8 March 2007).
Escutia, C., Brinkhuis, H., Klaus, A., Expedition 318 Scientists, 2011. Proceedings of theIntegrated Ocean Drilling Program 318. Integrated Ocean Drilling ProgramManagement International, Inc, Tokyo.
Expedition 342 Scientists, 2012. Paleogene Newfoundland sediment drifts. IODP Prelim.Rept. 342. http://dx.doi.org/10.2204/iodp.pr.342.2012.
Fenner, J., 1978. Cenozoic diatom biostratigraphy of the equatorial and southern AtlanticOcean. Initial Rep. Deep Sea Drill. Program 39, 491–623.
Fenner, J., 1984a. Eocene–Oligocene planktic diatom stratigraphy in the low latitudes andthe high southern latitudes. Micropaleontology 30, 319–342.
Fenner, J., 1984b. Middle Eocene to Oligocene planktonic diatom stratigraphy from DeepSea Drilling Sites in the South Atlantic, Equatorial Pacific, and Indian oceans. InitialRep. Deep Sea Drill. Program 75, 1245–1271.
Fenner, J., 1985. Late Cretaceous to Oligocene planktic diatoms. In: Bolli, H.B., Saunders,J.B., Perch-Nielsen, K. (Eds.), Plankton Stratigraphy. Cambridge University Press,Cambridge, pp. 713–763.
Fenner, J., 1994. Diatoms of the Fur Formation, their taxonomy and biostratigraphic inter-pretation. Results from the Harre borehole, Denmark. Aarh. Geosci. 1, 99–163.
Fenner, J., Mikkelsen, N., 1990. Eocene–Oligocene diatoms in the Western Indian Ocean:taxonomy, stratigraphy, and paleoecology. Proc. Ocean Drill. Program Sci. Results115, 433–463.
Fourtanier, E., 1991. Paleocene and Eocene diatom biostratigraphy and taxonomy of east-ern Indian Ocean Site 752. Proc. Ocean Drill. Program Sci. Results 121, 171–187.
Gladenkov, A.Yu., 2013. First finds of Eocene diatoms in the marine Paleogene reference sec-tion in the Ilpinskii peninsula, northeastern Kamchatka. Stratigr. Geol. Correl. 21, 96–106.
Glezer, Z.I., 1994. Correlation of development stages of the Paleogene diatom flora frommarine epicontinental and oceanic basins. Stratigr. Geol. Correl. 2 (1), 95–98.
Glezer, Z.I., 1996a. Evolutionary relationship of the Eocene diatom flora to some abioticfactors. Stratigr. Geol. Correl. 4 (3), 269–276.
Glezer, Z.I., 1996b. Problems of Eocene siliceous phytoplankton zonation (exemplified bythe Caspian Eocene deposits). Stratigr. Geol. Correl. 4 (4), 392–402.
Gombos Jr., A.M., 1982. Early and Middle Eocene diatom evolutionary events. Bacillaria 5,225–242.
Gombos Jr., A.M., 1983. Middle Eocene diatoms from the South Atlantic. Initial Rep. DeepSea Drill. Proj. 71, 565–581.
Gombos Jr., A.M., 1987. Middle Eocene diatoms from the North Atlantic, Deep Sea DrillingProject 605. Initial Rep. Deep Sea Drill. Program 93, 793–799.
Gombos Jr., A.M., Ciesielski, P.F., 1983. Late Eocene to Early Miocene diatoms from thesouthwest Atlantic. Initial Rep. Deep Sea Drill. Program 71, 583–632.
Gradstein, F., Ogg, J., Smith, A., 2004. A Geologic Time Scale 2004. Cambridge UniversityPress, Cambridge (589 pp.).
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M. (Eds.), 2012. The Geologic Time Scale2012. Elsevier, Boston, USA (1144 pp.).
Hajós, M., 1976. Upper Eocene and Lower Oligocene Diatomaceae, Archaeomonadaceae,and Silicoflagellatae in southwestern Pacific sediments, DSDP Leg 29. Initial Rep.Deep Sea Drill. Program 35, 817–883.
Harwood, D.M., Bohaty, S.M., 2000. Marine diatom assemblages fro Eocene and youngererratics, McMurdo Sound, Antarctica. In: Stilwell, J.D. (Ed.), Paleobiology andPaleoenvironments of Eocene Rocks, McMurdo Sound, East Antarctica AntarcticResearch Series 76, pp. 73–98.
Hay, W.W., DeConto, R., Wold, C.N., Wilson, K.M., Voigt, S., Schulz, M., Wold-Rossby, A.,Dullo, W.-C., Ronov, A.B., Balukhovsky, A.N., Soeding, E., 1999. Alternative globalCretaceous paleogeography. In: Barrera, E., Johnson, C. (Eds.), The Evolution ofCretaceous Ocean/Climate Systems. Geological Society of America Special Paper332, pp. 1–47.
Hohbein, M.W., Sexton, P.F., Cartwright, J.A., 2012. Onset of North Atlantic Deep Waterproduction coincident with inception of the Cenozoic global cooling trend. Geology40 (3), 255–258.
Homann, M., 1991. Die Diatomeen der Fur-Formation (Alttertiar) aus dem Limfjord-Gebiet, Nordjutland/Danemark. Geol. Jahrb. Reihe A 123, 3–285.
Huber, M., Brinkhuis, H., Stickley, C.E., Döös, K., Sluija, A., Warnaarm, J., Schellenberg, S.A.,Williams, G.L., 2004. Eocene circulation of the Southern Ocean: was Antarctica keptwarm by subtropical waters? Paleoceanography 19, PA4026. http://dx.doi.org/10.1029/2004PA001014.
Jakobsson, M., Backman, J., Rudels, B., Nycander, J., Frank, M., Mayer, L., Jokat, W.,Sangiorgi, F., O'Regan, M., Brinkhuis, H., King, J., Moran, K., 2007. The Early Mioceneonset of a ventilated circulation regime in the Arctic Ocean. Nature 447, 986–990.http://dx.doi.org/10.1038/nature05924.

99J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
Keller, G., Herbert, T., Dorsey, R., D'Hondt, S., Johnsson, M., Chi, W.R., 1987. Global distri-bution of Late Paleogene hiatuses. Geology 15, 199–203.
Kim, B.I., Glezer, Z.I., 2007. Sedimentary cover of the Lomonosov Ridge: Stratigraphy,structure, deposition history, and ages of seismic facies units. Stratigraphy and Geo-logical Correlation 15 (4), 401–420.
Koizumi, I., 1992. Diatomaceous sediments along the Pacific coastal areas of SouthAmerica and their evaluation. J. Fac. Sci. Hokkaido Univ. Ser. 4 Geol. Mineral. 23 (2),227–245.
Kominz, M.A., Browning, J.V., Miller, K.G., Sugarman, P.J., Misintseva, S., Scotese, C.R., 2008.Late Cretaceous to Miocene sea-level estimates from the New Jersey and Delawarecoastal plain coreholes: an error analysis. Basin Res. 20, 211–226.
Lazarus, D., Barron, J., Renaudie, J., Diver, P., Türke, A., 2014. Cenozoic planktonic marinediatom diversity and correlation to climate change. PLoS ONE 9 (1), 1–18 (January).
Lisitzin, A.P., 1972. Sedimentation in the World Ocean. SEPM Spec. Publ. 17 (Tulsa,Oklahoma).
Livermore, R., Hillenbrand, C.-D., Mereth, M., Eagles, G., 2007. Drake Passage and Cenozoicclimate: an open and shut case? Geochem. Geophys. Geosyst. 8 (1), 1–11. http://dx.doi.org/10.1029/2005GC001224.
Locker, S., 1996. Cenozoic siliceous flagellates from the Fram Strait and the east Greenlandmargin: Biostratigraphic and paleoceanographic results. Proceedings of the OceanDrilling Program, Scientific Results 151, 101–124.
Lyle, M., Pälike, H., Nishi, H., Raffi, I., Gamage, K., Klaus, A., The IODP Expeditions 320/321Scientific Party, 2010. The Pacific equatorial age transect, IODP Expeditions 320 and321: building a 50-million-year-long environmental record of the Equatorial Pacific.Sci. Drill. 9, 4–15. http://dx.doi.org/10.2204/iodp.sd.9.01.2010.
Martini, E., 1970. Standard Tertiary and Quaternary calcareous nannoplankton zonation.In: Farinacci, A. (Ed.), Proceedings II Planktonic Conference, Roma 2, pp. 739–785.
Martini, E., Müller, C., 1976. Eocene to Pleistocene silicoflagellates from the Norwegian-Greenland Sea (DSDP Leg 38). Initial Reports of the Deep Sea Drilling Project 38,857–895.
Marty, R., Dunbar, R., Martin, J.B., Baker, P.A., 1988. Late Eocene diatomite from thePeruvian coastal desert, coastal upwelling in the eastern Pacific, and Pacific circula-tion before the terminal Eocene event. Geology 16, 818–822.
McGowran, B., 1989. Silica burp in the Eocene ocean. Geology 17, 857–860.McLean, H., Barron, J.A., 1988. A late Middle Eocene diatomite in northwestern Baja
California Sur, Mexico: implications for tectonic translation. In: Filewicz, M.V.,Squires, R.L. (Eds.), Paleogene Stratigraphy, West Coast of North America: PacificSection, Society of Economic Paleontologists andMineralogists.West Coast PaleogeneSymposium 58, pp. 1–8.
Miller, K.G., Wright, J.D., Fairbanks, R.G., 1991. Unlocking the Ice House: Oligocene–Miocene oxygen isotopes, eustasy, and margin erosion. J. Geophys. Res. 96 (B4),6829–6848.
Mitlehner, A.G., 1996. Palaeoenvironments in the North Sea Basin around the Paleocene–Eocene boundary evidence from diatoms and other siliceous microfossils. In: Know,R.W.O., Corfiedl, R.M., Dunay, R.E. (Eds.), Correlation of the Early Paleogene inNorthwest Europe. Geological Society Special Publication 101, pp. 255–273.
Moore Jr., T.C., Jarrard, R.D., Olivarez Lyle, A., Lyle, M., 2008. Eocene biogenic silica accu-mulation rates at the Pacific equatorial divergence zone. Paleoceanography 23,PA2202. http://dx.doi.org/10.1029/2007PA001514.
Mountain, G.S., Tucholke, B.E., 1985. Mesozoic and Cenozoic Geology of the U.S. AtlanticContinental Slope and Rise. In: Poag, C.W. (Ed.), Geologic Evolution of the U.S. AtlanticMargin. Van Nostrand Reinhold, Stroudsburg, PA, pp. 293–341.
Müller, C., 1979. Calcareous nannofossil from the North Atlantic (Leg 49). Initial Reportsof the Deep Sea Drilling Project 48, 589–639.
Mutoni, G., Kent, D.V., 2007. Widespread formation of cherts during the Early Eocene cli-mate optimum. Palaeogeogr. Palaeoclimatol. Palaeoecol. 253, 348–362.
Nigrini, C., Sanfilippo, A., Moore Jr., T.J., 2005. Cenozoic radiolarian biostratigraphy: amagnetobiostratigraphic chronology of Cenozoic sequences from ODP Sites 1281,1219, and 1220, equatorial Pacific. Proc. Ocean Drill. Program Sci. Results 199, 1–76.
Norris, R.D., Klaus, A., Kroon, D., 2001. Mid-Eocene deep water, the Late Palaeocene Ther-mal Maximum and continental slope mass wasting during the Cretaceous–Palaeogene impact. In: Kroon, D., Norris, R.D., Klaus, A. (Eds.), Western North AtlanticPaleogene and Cretaceous Paleoceanography. Geol. Soc. Spec. Publ. London 183,pp. 23–48.
Norris, R.D., Wilson, P.A., Blum, P., The Expedition 342 Scientists, 2014. Site U1403.Proceedings of the, Integrated Ocean Drilling Program 342.
O'Regan, M., Moran, K., Backman, J., Jakobsson, M., Sangiorgi, F., Brinkhuis, H., RobPockalny, R., Skelton, A., Stickley, C.E., Koc, N., Brumsack, H.-J., Willard, D., 2008.Mid-Cenozoic tectonic and paleoenvironmental setting of the central Arctic Ocean.Paleoceanography 23, 1–5. http://dx.doi.org/10.1029/2007PA001559 (PA1S20).
Okada, H., Bukry, D., 1980. Supplementary modification and introduction of code num-bers to the low-latitude coccolith biostratigraphic zonation (Bukry, 1973; 1975).Mar. Micropaleontol. 5 (3), 321–325.
Onodera, J., Takahashi, K., Jordan, R., 2008. Eocene silicoflagellate and ebridianpaleoceanography in the central Arctic Ocean. Paleoceanography 23, PA1S15.http://dx.doi.org/10.1029/2007/PA001474.
Oreshkina, T.V., 2012. Evidence of Late Paleocene–Early Eocene hyperthermal events inbiosiliceous sediments of western Siberia and adjacent areas. Aust. J. Earth Sci. 105,145–153.
Oreshkina, T., Oberhänsli, H., 2003. Diatom turnover in the Early Paleogene diatomite ofthe Sengiley section, middle Povolzhie, Russia: a response to the initial EoceneThermal Maximum? In: Wing, S.L. (Ed.), Causes and Consequences of GloballyWarm Climates in the Early Paleogene. Geological Society of America Spec. Pap369. Boulder, Colo, pp. 169–179
Perch-Nielsen, K., 1975. Late Cretaceous to Pleistocene silicoflagellates from the southernsouthwest Pacific, DSDP Leg 29. Initial Rep. Deep Sea Drill. Program 29, 677–721.
Perch-Nielsen, K., 1985. Silicoflagellates. In: Bolli, H.M., Saunders, J.B., Perch-Nielsen, K.(Eds.), Plankton Stratigraphy. Cambridge University Press, Cambridge, pp. 811–846.
Poirier, A., Hillaire-Marcel, C., 2011. Improved Os-isotope stratigraphy of the Arctic Ocean.Geophys. Res. Lett. 38, L14607.
Radionova, E.P., Khokhlova, I.E., 1994. Paleogene episodes of biogenic silica accumulationin the northern Caucasus and adjacent Tethyan regions. Stratigr. Geol. Correl. 2 (5),61–169.
Radionova, E.P., Beniamovski, V.N., Iakovleva, A.I., Muzylöv, N.G., Oreshkina, T.V.,Shcherbinina, E.A., Kozlova, G.E., 2003. Early Paleogene transgressions: stratigraphicaland sedimentological evidence from the northern Peri-Tethys. Geol. Soc. Am. Spec.Pap. 369, 239–261.
Renaudie, J., Danelian, T., Saint Martin, S., Le Callonnec, L., Tribovillard, N., 2010. Siliceousphytoplankton response to a Middle Eocene warming event recorded in the tropicalAtlantic (Demerara Rise, ODP Site 1260A). Palaeogeogr. Palaeoclimatol. Palaeoecol.286, 121–134.
Sangiorgi, F., van Soelen, E.E., Spofforth, D.J.A., Pailike, H., Stickley, C.E., St. John, K., Koc, N.,Schouten, S., Sinninghe Damsté, J.S., Brinkhuis, H., 2008. Cyclicity in the MiddleEocene central Arctic Ocean sediment record: orbital forcing and environmental re-sponse. Paleoceanography 23, PA1S08. http://dx.doi.org/10.1029/2007PA001487.
Saunders, J.B., Bernoulii, D., Müller-Merz, E., Oberhänsli, H., Perch-Nielsen, K., Riedel, W.R.,Sanfilippo, A., Torrini Jr., R., 1984. Stratigraphy of the late Middle Eocene to EarlyOligocene in the Bath Cliff section, Barbados, West Indies. Micropaleontology 30,390–425.
Scher, H.D., Martin, E.E., 2004. Circulation in the Southern Ocean during the Paleogene in-ferred from neodymium isotopes. Earth Planet. Sci. Lett. 228, 391–405.
Scher, H., Martin, E.E., 2006. Timing and climatic consequences of the opening of theDrake Passage. Science 312, 428–430.
Scherer, R., Koc, N., 1996. Late Paleogene diatom biostratigraphy and paleoenvironmentsof the northern Norwegian-Greenland Sea. Proc. Ocean Drill. Program Sci. Results151, 75–100.
Scherer, R.P., Gladenkov, A.Yu., Barron, J.A., 2007. Methods and applications of Cenozoicmarine diatom biostratigraphy. In: Starratt, S.W. (Ed.), Pond Scum to Carbon Sink:Geological and Environmental Applications of the Diatoms. The PaleontologicalSociety Papers 13, pp. 61–83.
Schrader, H.-J., 1976. Cenozoic planktonic diatom biostratigraphy of the southern PacificOcean. Initial Rep. Deep Sea Drill. Program 35, 605–671.
Schrader, H.-J., Fenner, J., 1976. Norwegian Sea Cenozoic diatom biostratigraphy and tax-onomy. Initial Rep. Deep Sea Drill. Program 38, 921–1099.
Shaw, C.A., Ciesielski, P.F., 1983. Silicoflagellate biostratigraphy of Middle Eocene to Holo-cene subantarctic sediments recovered by Deep Sea Drilling Project Leg 71. Initial Re-ports of the Deep Sea Drilling Project 71, 687–736.
Sluijs, A., Zeebe, R.E., Bijl, P.K., Bohaty, S.M., 2013. A Middle Eocene carbon cycle conun-drum. Nat. Geosci. 6, 429–434. http://dx.doi.org/10.1038/ngeo1807.
Spencer-Cervato, C., 1999. The Cenozoic deep sea microfossil record: explorations of theDSDP/ODP sample set using the Neptune database. Palaeontol. Electron. 2 (web).
St John, K., 2008. Cenozoic ice-rafting history of the central Arctic Ocean: terrigenoussands on the Lomonosov Ridge. Paleoceanography 23, PA1S05.
Stickley, C.E., Brinkhuis, H., McGonigal, K.L., Chaproniere, G.C.H., Fuller, M., Kelly, D.C.,Nürnberg, D., Pfuhl, H.A., Schellenberg, S.A., Schoenfeld, J., Suzuki, N., Touchard, Y.,Wei, W., Williams, G.L., Lara, J., Stant, S.A., 2004a. Late Cretaceous–Quaternarybiomagnetostratigraphy of ODP Sites 1168, 1170, 1171 and 1172, TasmanianGateway. Proc. Ocean Drill. Program Sci. Results 189, 1–57.
Stickley, C.E., Brinkhuis, H., Schellenberg, S.A., Sluijs, A., Röhl, U., Fuller, M., Grauert, M.,Huber, M., Warnaar, J., Williams, G.L., 2004b. Timing and nature of the deepening ofthe Tasmanian Gateway. Paleoceanography 19, PA4027. http://dx.doi.org/10.1029/2004PA001022.
Stickley, C.E., Koç, N., Brumsack, H.-J., Jordan, R.W., Suto, I., 2008. A siliceous microfossilview of Middle Eocene Arctic paleoenvironments: a window of biosilica productionand preservation. Paleoceanography 23. http://dx.doi.org/10.1029/2007PA001485.
Stickley, C.E., St. John, K., Koç, N., Jordan, R.W., Passchier, S., Pearce, R.B., Kearns, L.E., 2009.Evidence for Middle Eocene Arctic sea ice from diatoms and ice-rafted debris. Nature460 (16 July), 376–379. http://dx.doi.org/10.1038/nature08163.
Stickley, C.E., Koç, N., Pearce, R.B., Kemp, A.E.S., Jordan, R.W., Sangiorgi, F., St. John, K.,2012. Variability in the length of the sea ice season in the Middle Eocene Arctic.Geology 40, 727–730. http://dx.doi.org/10.1130/G32976.1.
Stoker, M., Leslie, A., Smith, K., Ólavsdóttirm, J., Johnson, H., Laberg, J.S., 2013. Onset ofNorth Atlantic Deep Water production coincident with inception of the Cenozoiccooling trend: COMMENT. Geology e291 (September 2013).
Strelnikova, N.I., 1990. Evolution of diatoms during the Cretaceous and Paleogene periods.In: Simola, H. (Ed.), Proceedings of the Tenth International Diatom Symposium.Koeltz Scientific Books, Koenigstein, Germany, pp. 195–204.
Strelnikova, N.I., 1992. Diatoms of the Paleogene. The University of Saint-Peterburg Press,Saint-Peterburg (312 pp., [In Russian]).
Strelnikova, N.I., Fourtanier, E., Kociolek, J.P., Barron, J.A., 2001. Ultrastructure studies ofCoscinodiscus and Cestodiscus species from the Eocene and Oligocene. In: Jahn,Regine, Kociolek, John P., Witkowski, A., Compère, P. (Eds.), Lange-Bertalot-Fest-schrift: Studies on Diatoms. Koenigstein (Koeltz Scientific Books), pp. 63–96.
Suto, I., Jordan, R.W., Watanabe, M., 2009. Taxonomy of Middle Eocene diatom restingspores and their allied taxa from IODP sites in the central Arctic Ocean (LomonosovRidge). Micropaleontology 55, 259–312.
Thomas, D.J., 2004. Evidence for deep-water production in the North Pacific during theEarly Cenozoic warm interval. Nature 430, 65–68.
Thomas, D.J., Lyle, M., Moore Jr., T.C., Rea, D.K., 2008. Paleogene deep-water masscomposition of the tropical Pacific and implications for thermohaline circulation ina Greenhouse World. Geochem. Geophys. Geosyst. 9, Q02002. http://dx.doi.org/10.1029/2007GC001748.

100 J.A. Barron et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 422 (2015) 85–100
Tsoy, I., 2003. Eocene diatoms and silicoflagellates from the Kronotskii Bay deposits (EastKamchatka). Stratigr. Geol. Correl. 11, 376–390.
Tsoy, I.B., 2011. Silicoflagellates of the Middle–early Late Eocene of the Kronotskii Bay(Eastern Kamchatka). Stratigr. Geol. Correl. 19, 87–101.
Wade, B.S., Kroon, D., 2002. Middle Eocene regional climate instability: evidence from thewestern Atlantic. Geology 30, 1011–1014.
Westerhold, T., Röhl, U., 2013. Orbital pacing of Eocene climate during the Middle EoceneClimatic Optimum and chron 19r event: missing link found in the tropical westernAtlantic. Geochem. Geophys. Geosyst. 14 (11), 4811–4825. http://dx.doi.org/10.1002/ggge.20293.
Westerhold, T., Röhl, U., Raffi, I., Fornaciari, E., Monechi, S., Reale, V., Bowles, J., Evans, H.F.,2008. Astronomical calibration of Paleocene time. Palaeogeogr. Palaeoclimatol.Palaeoecol. 257 (4), 377–403.
Westerhold, T., Röhl, U., Laskar, J., 2012. Time scale controversy: accurate orbital calibra-tion of the Early Paleogene. Geochem. Geophys. Geosyst. 13, Q06015. http://dx.doi.org/10.1029/2012GC004096.
Wilson, D.S., Pollard, D., DeConto, R.M., Jamieson, S.S.R., Luyendyk, B.P., 2013. Initiation oftheWest Antarctic Ice Sheet and estimates of total Antarctic ice volume in the earliest
Oligocene. Geophys. Res. Lett. 40 (16), 4305–4309. http://dx.doi.org/10.1002/grl.50797.
Witkowski, J., Bohaty, S.M., McCartney, K., Harwood, D.M., 2012. Enhanced siliceousplankton productivity in response to Middle Eocene warming at Southern OceanODP Sites 748 and 749. Palaeogeogr. Palaeoclimatol. Palaeoecol. 326–328, 78–94.
Witkowski, J., Bohaty, S.M., Edgar, K.M., Harwood, D.M., 2014. Rapid fluctuations in mid-latitude siliceous plankton production during the Middle Eocene Climatic Optimum(ODP Site 1051, western North Atlantic). Mar. Micropaleontol. 106, 110–129.http://dx.doi.org/10.1016/j.marmicro.2014.01.001.
Yool, A., Tyrell, T., 2005. Implications for the history of Cenozoic opal deposition from aquantitative model. Palaeogeogr. Palaeoclimtol. Palaeoecol. 218, 239–255.
Zachos, J., Pagain, M., Sloan, L., Thomas, E., Billips, K., 2001. Trends, rhythms, and aberra-tions in global climate 65 Ma to present. Science 292, 686–691.
Zachos, J.C., Dickens, G.R., Zeebe, R.E., 2008. An Early Cenozoic perspective on greenhousewarming and carbon-cycle dynamics. Nature 451, 279–283. http://dx.doi.org/10.1038/nature06588.

Supplementary Table 1. Estimated ages in Ma of first (FO) and last ocurrences (LO) of biostratigraphically important diatoms withcitation reference for those ages (reference key in Supplementary Table 2) and ocean area where they appear to be most useful. Tr = tropical; SO = Southern Ocean, RP = Russian epicontinental ptlatform and/or North Sea; NA = North Atlantic, NP = North Pacific
Formal Name FO 2012 LO 2012 OceanAge
Reference Other genusAbas wittii (Grunow) Ross & Sims 1980 53.5 39.5 Tr 1; 2 SyringidiumActinocyclus octonarius Ehrenberg 1837 37.5 post Eocene Tr 14Actinoptychus irregularis Grunow in Van Heurck 1883 38.6 post Eocene NA 13Actinoptychus splendens (Shadbolt) Ralfs in Pritchard 1861 39.9 post Eocene Tr 2 ActinophaeniaAsterolampra affinis Greville 1862 43.7 post Eocene Tr 16Asterolampra grevillei (Wallich) Greville 1860 43.7 post Eocene Tr 16 AsteromphalusAsterolampra insignis Schmidt in Schmidt et al. 1888 46.3 post Eocene Tr 2Asterolampra marylandica Ehrenberg 1844 40.4 post Eocene Tr 8Asterolampra punctifera (Grove in Schmidt et al.) Forti in Tempère & Peragallo 1914 35.2
post EoceneSO 10
Asterolampra vulgaris Greville 1862 46.4 post Eocene SO 2Asteromphalus oligocenicus Schrader & Fenner 1976 33.8 post Eocene SO 2Asteromphalus schmidtii Heiden in Heiden & Kolbe 1928 37.5 post Eocene SO 2Azpeitia crenuloides P.A. Sims in Fryxell, Sims & Watkins 1986 39.4
post EoceneTr 2
Azpeitia oligocenica (Jousé) P.A. Sims in Fryxell, Sims & Watkins 1986 34.3
post EoceneTr 7; 3 Coscinodiscus
Azpeitia oligocenica var. nodosa (Jousé) Sims 34.0 post Eocene Tr 7 CoscinodiscusAzpeitia tuberculata (Greville) P.A. Sims in Fryxell et al.1986 46.2 35.6 Tr #1; 7 CoscinodiscusBaxteriopsis brunii (Van Heurck) Karsten 1928 35.1 post Eocene Tr 7; 9 BaxteriaBrightwellia hyperborea Grunow in Van Heurck 1883 53.0 39.4 Tr 5; 8Brightwellia imperfecta Jousé in Gleser & Jousé 1974 41.8 34.6 Tr 8Cestodiscus antarcticus Fenner 1984 34.9 33.2 SO 7Cestodiscus convexus Castracane 1886 34.4 post Eocene Tr 7; 9Cestodiscus demergitus (Fenner) Fenner & Mikkelsen 1990 34.3 post Eocene Tr 7; 2 CoscinodiscusCestodiscus fennerae Strelnikova & Fourtanier 2001 36.9 35.0 Tr 7Cestodiscus pulchellus var. novazealandica (Grove ex Rattray) Fenner ex Strelnikova & Fourtanier 2001 39.0 35.0 Tr 7Cestodiscus robustus Jousé 1974 33.7 post Eocene Tr 7; 9Cestodiscus sp. 2 Fenner 1981 33.9 33.3 Tr 7; 9Cestodiscus spinifer (Grove & Sturt) Fenner 1981 38.3 35.4 Tr 7Cestodiscus stokesianus Greville 1866 35.0 post Eocene Tr 2Cestodiscus trochus Castracane 1886 34.2 post Eocene Tr 7; 3Chaetoceros asymmetricus Fenner in Fenner & Mikkelsen 1990 35.0
post EoceneTr 2
Coscinodiscus bulliens Schmidt in Schmidt et al. 1878 53.0 post Eocene Tr 17; 15Coscinodiscus cruxii Fenner 1991 Paleocene 56.7 Tr 2Coscinodiscus decrescens Grunow in Schmidt et al. 1878 55.3 33.9 Tr 5, 17Coscinodiscus denarius (Schmidt) Strelnikova & Fourtanier 2001 55.5 33.4 RP, SO 5; 7Coscinodiscus eomonoculus Bukry 1987 50.4 46.0 Tr 46Coscinodiscus excavatus Greville manuscript, Ralfs in Pritchard 1861 33.8
post EoceneTr 9
Coscinodiscus hajosiae Fenner 1984 37.6 post Eocene Tr 7; 9Coscinodiscus payeri Grunow 1884 55.3 48.1 RP 4Coscinodiscus uralensis Jouse in Proschkina-Lavrenko 1949 55.9 53.8 RP 6 MoisseeviaCraspedodiscus möelleri Schmidt in Schmidt et al. 1893 55.9 40.4 RP 6; 8 CoscinodiscusCraspedodiscus oblongus (Greville) Grunow in Schmidt et al. 1881 50.8 40.4 Tr 15; 8 Coscinodiscus; PorodiscusCraspedodiscus umbonatus Greville 1866 44.0 39.7 Tr 2 AbasCraspedodiscus undulatus Gombos 1982 52.3 50.4 Tr 8Ethmodiscus rex (Wallich in Rattray) Hendey in Wiseman & Hendey 1953
37.8 post EoceneTr 2 Coscinodiscus
Eurossia irregularis (Greville) Sims in Mahood, Barron & Sims 1993
38.0 post EoceneSO 7; 10 Triceratium
Fenestrella antiqua (Grun.) Swatman 1948 56.4 50.6 NA 6; 19 JanischiaGrunoviella (Grunowiella) gemmata (Grunow) Van Heurck 1896 Paleocene 50.6 RP 5 SceptroneisGrunoviella (Grunowiella) palaeocaenica Jousé 1951 Paleocene 56.7 RP 2Gyrocylindrus antiquus Strelnikova et Nikolaev 1995 55.4 55.4 RP 6Hemiaulus alatus Greville 1865 44.4 37.9 Tr 8Hemiaulus altar Brun 1896 37.1 post Eocene Tr 2Hemiaulus barbadensis Grunow 1884 33.7 post Eocene Tr 2Hemiaulus caracteristicus Hajós 1976 56.5 post Eocene SO 1; 2Hemiaulus curvatulus Strelnikova 1971 Cretaceous 54.8 RP 19Hemiaulus dubius Grunow 1884 46.2 post Eocene Tr 1Hemiaulus elegans (Heiberg) Grunow emend Homann 1991 Cretaceous 54.8 NA 22 CorinnaHemiaulus exiguus Greville 1865 42.0 post Eocene Tr 1; 2Hemiaulus februatus Heiberg 1863 Cretaceous 55.6 NA 19Hemiaulus gondolaformis Fenner 1984 43.6 37.2 Tr 8Hemiaulus gracilis Fenner in Fenner & Mikkelsen 1990 34.4 34.1 Tr 2Hemiaulus grassus Fenner 1984 45.4 36.3 Tr 8Hemiaulus inaequilaterus Gombos 1976 56.1 48.1 Tr 8

Hemiaulus incisus Hajos 1976 Paleocene 44.1 SO 2Hemiaulus incurvus Schibkova in Krotov & Schibkova 1959 Paleocene 52.0 Tr 1; 3Hemiaulus kristoffersenii Fenner 1991 Paleocene 56.9 Tr 2Hemiaulus lyriformis Greville 1865 44.7 post Eocene Tr 1Hemiaulus nocchiae Fenner 1991 Paleocene 56.9 Tr 2Hemiaulus peripterus var. longispina Fourtanier 1991 55.3 53.4 Tr 3; 1Hemiaulus polycistinorum var. mesolepta Grunow 1884 46.2 post Eocene Tr 1; 10Hemiaulus proteus Heiberg 1863 56.4 41.0 RP 17; 6; #3Hemiaulus rectus var. twista Fenner 1984 55.4 post Eocene SO 23Hemiaulus subactus Grunow 1884 57.1 post Eocene Tr 2Hemiaulus taurus Gombos in Gombos & Ciesielski 1983 34.2 post Eocene SO 2Hyalodiscus ambiguus (Grunow; Grunow in Cleve & Grunow) Tempère & Peragallo 1889 Paleocene 57.5 Tr 2 PodosiraKozloviella subrotunda Fenner 1984 39.8 33.8 Tr 8; 7Navicula udentsevii Schrader & Fenner 1976 34.2 post Eocene NA, SO 7Odontotropis cristata Grunow 1884 Paleocene 55.0 NA 19Pseudotriceratium chenevieri (Meister) Gleser 1974 43.2 post Eocene NA 2Pseudotriceratium radiosoreticulatum (Grunow) Jouse in Dzinoridze, Jouse & Strelnikova 1979
36.8 post Eocene SO 7; 13 Triceratium
Pterotheca carinifera (Grunow) Forti 1909 Paleocene ~35 RP 19 PyxillaPterotheca clavata Strelnikova 1974 Cretaceous 56.9 Tr 25; 2Pterotheca crucifera Hanna 1927 Cretaceous 42.2 NA 26; 21Pterotheca danica (Grunow) Forti 1909 Cretaceous 34.2 SO 22; 23 StephanogoniaPterotheca evermanni (evermannii) Hanna 1927 Cretaceous 54.5 RP 26; 12Pterotheca kittoniana (Grunow in Van Heurck) Forti 1909 Paleocene 57.4 RP 12; 2 PyxillaPyxilla caputavis (caput avis) Brun 1896 45.6 41.8 Tr 8Pyxilla gracilis Tempère & Forti in Forti 1909 54.0 40.4 Tr 3Pyxilla oligocaenica Jousé 1955 50.6 45.5 RP 11; 4Pyxilla reticulata Grove & Sturt 1887 46.6 post Eocene Tr 8; 2Rhizosolenia hebetata J.W. Bailey 1856 55.4 post Eocene RP, NP 12Rhizosolenia interposita Hajós 1976 34.9 post Eocene SO 2Rhizosolenia oligocaenica Schrader 1976 34.0 post Eocene SO 7; 10Rhizosolenia setigera Brightwell 1858 55.4 post Eocene RP, Tr 12Riedelia claviger (A. Schmidt) Schrader & Fenner 1976 41.4 post Eocene Tr 17; 20; 2 HemiaulusRocella praenitida (Fenner) Fenner in Kim & Barron 1986 ~46.5 post Eocene Tr 17;13 CoscinodiscusRouxia granda Scharder in Schrader & Fenner 1976 34.7 post Eocene SO 2Rylandsia biradiata Greville 1861 36.3 34.0 SO 8; 7Rylandsia inaequiradiata Barker & Meakin 1945 35.9 33.8 SO 8Sceptroneis fennerae Scherer et Koc 1996 34.1 post Eocene NA 13Sceptroneis pesplanus Fenner & Schrader in Schrader & Fenner 1976 44.9
post EoceneNA 21; 2
Sceptroneis tenue Schrader & Fenner 1976 34.2 post Eocene NA 13; 2Skeletonemopsis barbadense (Greville) Sims 1994 37.3 post Eocene Tr 7; 2 SkeletonemaStephanopyxis eocenica Hajos 1976 37.0 post Eocene 7Stephanopyxis marginata Grunow 1884 Paleocene post Eocene Tr 2 PyxidiculaStephanopyxis ornata Schulz 1935 35.4 33.8 SO 2Stephanopyxis splendidus (Greville) Harwood 1989 37.5 33.6 SO 23; 18 Thalassiosira cf hydra GombosThalassiosira dubiosa Schrader in Schrader & Fenner 1976 38.6 post Eocene SO 13Thalassiosiropsis wittiana (Pantocsek) Hasle in Hasle & Syvertsen 1985
Cretaceous50.6
RP5 Coscinodiscus
Triceratium areolatum Greville 1861 56.0 42.0 RP 19; 20 SheshukoviaTriceratium barbadense Greville 1861 46.3 40.3 Tr 17; #2Triceratium gombosii Fenner 1991 Paleocene 57.5 Tr 2Triceratium heibergii Grunow in Van Heurck 1883 Paleocene 55.9 Tr 1Triceratium inconspicuum Greville 1861 48.4 39.0 Tr 8; #2 LisitziniaTriceratium kanayae Fenner 1984 48.4 39.0 Tr 8 LisitziniaTriceratium mirabile Jouse in Proschkina-Lavrenko 1949 Paleocene 55.8 Tr 5; 6 SheshukoviaTriceratium polycystinorum sensu Krotov & Schibkova 1959 53.5 45.8 Tr 1Triceratium unguiculatum Greville 1864 37.5 33.8 SO 2; 7Trinacria aries Schmidt in Schmidt et al. 1886 Cretaceous 56.8 Tr 3Trinacria conifera (Brightwell) Grunow 1884 Paleocene 56.9 Tr 2 TriceratiumTrinacria deciusi Hanna 1927 Cretaceous 56.9 Tr 2 MedliniaTrinacria excavata Heiberg 1863 f. tetragona Cretaceous 37.2 SO 24; 7 SheshukoviaTrinacria exsculpta (exculpta) (Heiberg) Hustedt 1930 56.4 54.0 Tr 14; 12 SoliumTrinacria regina Heiberg emend Homann 1991 Paleocene 55.0 Tr 19Trinacria simulacroides Pantocsek 1892 Paleocene 56.7 Tr 2 TriceratiumTrinacria simulacrum Grove & Sturt 1887 Paleocene 44.9 Tr 2Trinacria subcapitata (Greville) Grunow 1884 34.0 post Eocene Tr 7Trinacria ventriculosa Grove & Sturt 1887 Paleocene 56.4 RP 5Trochosira coronata Schrader & Fenner 1976 56.0 35.4 RP, SO 19; 13Trochosira gracillima (Fenner) Fenner 1991 Paleocene 56.6 Tr 19; 2 PterothecaTrochosira marginata Fenner 1991 Paleocene 57.3 Tr 2Trochosira radiata Fenner 1991 Paleocene 55.0 Tr 19; 2Trochosira trochlea Hanna 1927 Cretaceous 45.9 Tr 26; 1 CostopyxisXanthiopyxis acrolopha Forti 1912 34.7 post Eocene SO 2

Supplementary Table 2. Reference detail tp references cited in Supplementary Table 1.
ReferenceReference number
Fenner, J., 1985. Late Cretaceous to Oligocene planktic diatoms In Bolli, H. M., et al. (eds.), Plankton Stratigraphy. Cambridge University Press, Cambridge, United Kingdom, p. 1C. Spencer-Cervato, written comm., 2003. List of diatom FADs and LADS generated from Neptune database 2Fourtanier, E., 1991. Paleocene and Eocene diatom biostratigraphy and taxonomy of eastern Indian Ocean Site 752. Proceedings of the Ocean Drilling Program, Science 3Glezer, Z.I., 1996. Problems of Eocene siliceous phytoplankton zonation(exemplified by the Caspian Eocene deposits. Stratigraphy and Geological Correlation 4(4), 392-402. 4Oreshkina, T.V., 2012. Evidence of Late Paleocene-Early Eocene hyperthermal events in biosiliceous sediments of western Siberia and adjacent areas. Austrian Journal of Earth 5Oreshkina, T. & Oberhänsli, H., 2003. Diatom turnover in the early Paleogene diatomite of the Sengiley section, middle Povolzhie, Russia: A response to the intitla Eocene Thermal Maximum?, in Causes and Consequences of Globally Warm Climates in the Early Paleogene, edited by S. L. Wing et al., Geological Society of America, Spec. Pap. 369, 169- 6Barron, J., Bukry, D., and Gersonde, R., 2014. Diatom and silicoflagellate biostratigraphy for the late Eocene: ODP 1090 (Subantarctic Atlantic), Beihefte zur Nova Hedwigia. 7Scherer, R., Hannah, M., Maffioli, P., Persico, D., Sjunneskog, C., Strong, P., Taviani, M., & Winter, D, 2007. palaeontologic characterization and analysis of ANDRILL core AND-1b. 8Barron, J A., Fourtanier, E., & Bohaty, S., 2004. Oligocene and Earliest Miocene Diatom Biostratigraphy of Site 1220, ODP Leg 199, Equatorial Pacific. Proceedings of the Ocean Drilling Program, Scientific Results, 199, 1-25. 9Scherer, R.P, Bohaty, S, and Harwood, D.M., 2000. Oligocene and lower Miocene siliceous microfossil biostratigraphy of Cape Roberts Project, Core CRP-2/2a, Victoria Land Basin, Ross Sea, Antarctica. Terra Antartica 7(4), 417-442. 10Radionova, E P., Beniamovski, V.N., Iakovleva, A.I., Muzylov, N.G., Oreshkina, T.V., Shcherbinina, E.A., & Kozlova, G.E., 2003. Early Paleogene transgressions: Stratigraphical and sedimentological evidence from the northern Peri-Tethys, in Causes and Consequences of Globally Warm Climates in the Early Paleogene, edited by S. L. Wing et 11Homann, M.. 1991. Die Diatomeen der Fur-Formation (Alttertiar) aus dem Limfjord-Gebiet, Nordjutland/Danemark. Geologisches Jahrbuch, Reihe A, 123, 3-285. 12Scherer, R. & Koc, N., 1996. Late Paleogene diatom biostratigraphy and paleoenvironments of the northern Norwegian-Greenland Sea. Proceedings of the Ocean 13Hajós, M., 1976. Upper Eocene and lower Oligocene Diatomaceae, Archaeomonadaceae, and Silicoflagellatae in southwestern Pacific sediments, DSDP Leg 29, Initial Reports of the Deep Sea Drilling Program 35, 817–883. 14Gombos, A.M., Jr., 1982. Early and middle Eocene diatom evolutionary events, Bacillaria 5, 15Gombos, A.M., Jr., 1983. Middle Eocene diatoms from the South Atlantic Initial Reports of the Deep Sea Drilling Project 71, 565-581. 16Fenner, J., 1978. Cenozoic diatom biostratigraphy of the equatorial and southern Atlantic Ocean. Initial Reports of the Deep Sea Drilling Program 39 (Suppl.), 491-623. 17Wilson, G S., Roberts, A.P., Verosub, K.L., Florindo, F., Sagnotti, L., 1998. Magnetobiostratigraphic chronology of the Eocene—Oligocene transition in the CIROS-1 core, Victoria Land margin, Antarctica: Implications for Antarctic glacial history, Geological 18

Fenner J., 1994. Diatoms of the Fur Formation, their taxonomy and biostratigraphic interpretation. - Results from the Harre borehole, Denmark. Aarhus Geoscience, 1, 99-163. 19Fenner, J. M., & Mikkelsen, N., 1990. Eocene-Oligocene diatom in the western Indian Ocean: Taxonomy, stratigraphy, and paleoecology. Proceedings of the Ocean Drilling 20Schrader, H.-J., and Fenner, J., 1976. Norwegian Sea Cenozoic diatom biostratigraphy and taxonomy. Initial Reports of the Deep Sea Drilling Project 38, 921-1099. 21Hajos, M. & Stradner, H., 1975. Late Cretaceous Archaeomonadaceae, Diatomaceae, and Silicoflagellatae from South Pacific Ocean, Deep Sea Drilling Project, Leg 29, Site 275. Initial Reports of the Deep Drilling Project 29, 913-1009. 22Harwood, D .M. & Bohaty, S.M., 2001. Early Oligocene Siliceous Microfossil Biostratigraphy of Cape Roberts Project Core CRP-3, Victoria Land Basin, Antarctica. Terra Antartica 8, 315- 23Harwood, D.M., 1988. Upper Cretaceous and lower Paleocene diatom and silicoflagellate biostratigraphy of Seymour Island, eastern Antarctic Peninsula Geological Society of 24Strelnikova, N.I., 1974. The Diatoms of the Late Cretaceous of Western Siberia. (Diatomei Pozdnego Mela) Academy of Sciences of USSR. Publishing House Nauka, Moscow., pp. 1- 25Hanna, GD, 1927. Cretaceous Diatoms from California. Occasional Papers of the California Academy of Sciences, 12, 5-49. 26John Barron, unpublished notes #1Steve Bohaty, written communication ODP1218, 2004 #2N.I. Strenikova, 1990; Written communication, 2001 #3
Related Documents











