www.newphytologist.org 763 Research Blackwell Publishing, Ltd. Palatability of Thlaspi caerulescens for snails: influence of zinc and glucosinolates N. Noret 1,2 , P. Meerts 2 , R. Tolrà 3 , C. Poschenrieder 3 , J. Barceló 3 and J. Escarre 1 1 Centre d’Ecologie Fonctionnelle et Evolutive (CNRS), Route de Mende 1919, F-34293 Montpellier Cedex 05, France; 2 Université Libre de Bruxelles, Laboratoire de Génétique et Ecologie Végétales, Chaussée de Wavre 1850, B-1160 Bruxelles, Belgium; 3 Laboratorio de Fisiología Vegetal, Facultad de Ciencias, Universidad Autónoma de Barcelona, E-08193 Bellaterra, Spain Summary • The hypothesis that zinc (Zn) hyperaccumulation defends Thlaspi caerulescens against herbivores is tested with the snail Helix aspersa. We investigated the effects of leaf zinc, cadmium, glucosinolate, nitrogen and dry matter concentrations on the feeding preferences of snails. • Four T. caerulescens populations from southern France (two from metalliferous and two from normal soils) were grown on low- and high-Zn soils to obtain contrasting leaf Zn concentrations. Plants were also collected in the field, and binary feeding choices involving low- and high-Zn leaves were conducted. • Foliar Zn, Cd, N and dry matter concentrations did not affect the feeding choices of snails, whereas glucosinolate had a significant negative effect on herbivore pre- ferences. Compared with metallicolous plants, nonmetallicolous ones appeared to be better protected against snails, whatever their Zn concentration. • These results do not support the defence hypothesis, as glucosinolates appear to decrease the degree of herbivory when Zn does not. Key words: binary choices, glucosinolates, herbivory, hyperaccumulation, palatability, snails, Thlaspi caerulescens , zinc (Zn). New Phytologist (2005) 165 : 763–772 © New Phytologist (2004) doi : 10.1111/j.1469-8137.2004.01286.x Author for correspondence: N. Noret Tel: +32 2 650 91 60 Fax: +32 2 650 91 70 Email: [email protected] Received: 9 September 2004 Accepted: 1 October 2004 Introduction Few plant species are able to grow on metalliferous soils. Some of these metallophytes accumulate heavy metals in their shoots, up to extremely high concentrations (Brooks, 1998). The evolu- tionary ‘raison d’être’ of such heavy metal hyperaccumulation has been the subject of several hypotheses (Boyd & Martens, 1992; Boyd, 1998). The defence hypothesis, the most com- monly accepted (Pollard, 2000; Assunção et al., 2003), suggests that the metal stored in leaves would have evolved as an elemental defence against herbivores and pathogens (Boyd, 1998). The defence hypothesis has been tested experimentally several times by means of two kinds of experiment. In nonchoice feeding experiments, a herbivore is offered toxic food (e.g. high-metal leaves) or normal food (e.g. low-metal leaves) and its growth on each treatment compared. In Ni hyperaccumulators such feeding experiments (Boyd & Martens, 1994; Martens & Boyd, 1994; Boyd & Moar, 1999) showed that Ni is toxic to herbivores. On the other hand, in binary feeding trials the herbivore is given the choice between foods rich or poor in toxic metal, in order to examine its feeding preference. Some authors (Pollard & Baker, 1997; Huitson & Macnair, 2003; Macnair, 2003) have pointed out that binary choices only could be indicative of a possible herbivore selection pressure. If herbivores prefer to eat the least-accumulating genotypes, they could have promoted the evolution of hyperaccumulation. Two studies tested the defence hypothesis with binary feeding choices involving the Zn/Cd hyperaccumulator Thlaspi caerulescens. Pollard & Baker (1997) used phenotypic plasticity to obtain high- and low-Zn leaves, cultivating T. caerulescens plants on high- and low-Zn culture solutions. Jhee et al. (1999) used the genetic variability of hyperaccumulation existing between ecotypes and between genotypes within the same population to obtain variable foliar Zn concentrations among plants growing in the same medium. Both works concluded that Zn was effective in defending T. caerulescens, as high-Zn leaves were less eaten

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.newphytologist.org
763
Research
Blackwell Publishing, Ltd.
Palatability of
Thlaspi caerulescens
for snails: influence
of zinc and glucosinolates
N. Noret
1,2
, P. Meerts
2
, R. Tolrà
3
, C. Poschenrieder
3
, J. Barceló
3
and J. Escarre
1
1
Centre d’Ecologie Fonctionnelle et Evolutive (CNRS), Route de Mende 1919, F-34293 Montpellier Cedex 05, France;
2
Université Libre de Bruxelles,
Laboratoire de Génétique et Ecologie Végétales, Chaussée de Wavre 1850, B-1160 Bruxelles, Belgium;
3
Laboratorio de Fisiología Vegetal, Facultad de Ciencias,
Universidad Autónoma de Barcelona, E-08193 Bellaterra, Spain
Summary
• The hypothesis that zinc (Zn) hyperaccumulation defends
Thlaspi caerulescens
against herbivores is tested with the snail
Helix aspersa
. We investigated the effectsof leaf zinc, cadmium, glucosinolate, nitrogen and dry matter concentrations on thefeeding preferences of snails.• Four
T. caerulescens
populations from southern France (two from metalliferous andtwo from normal soils) were grown on low- and high-Zn soils to obtain contrastingleaf Zn concentrations. Plants were also collected in the field, and binary feedingchoices involving low- and high-Zn leaves were conducted.• Foliar Zn, Cd, N and dry matter concentrations did not affect the feeding choicesof snails, whereas glucosinolate had a significant negative effect on herbivore pre-ferences. Compared with metallicolous plants, nonmetallicolous ones appeared tobe better protected against snails, whatever their Zn concentration.• These results do not support the defence hypothesis, as glucosinolates appear todecrease the degree of herbivory when Zn does not.
Key words:
binary choices, glucosinolates, herbivory, hyperaccumulation, palatability,snails,
Thlaspi caerulescens
, zinc (Zn).
New Phytologist
(2005)
165
: 763–772
© New Phytologist
(2004)
doi
: 10.1111/j.1469-8137.2004.01286.x
Author for correspondence:
N. Noret
Tel: +32 2 650 91 60
Fax: +32 2 650 91 70
Email: [email protected]
Received:
9 September 2004
Accepted:
1 October 2004
Introduction
Few plant species are able to grow on metalliferous soils. Someof these metallophytes accumulate heavy metals in their shoots,up to extremely high concentrations (Brooks, 1998). The evolu-tionary ‘raison d’être’ of such heavy metal hyperaccumulationhas been the subject of several hypotheses (Boyd & Martens,1992; Boyd, 1998). The defence hypothesis, the most com-monly accepted (Pollard, 2000; Assunção
et al
., 2003), suggeststhat the metal stored in leaves would have evolved as an elementaldefence against herbivores and pathogens (Boyd, 1998).
The defence hypothesis has been tested experimentally severaltimes by means of two kinds of experiment. In nonchoice feedingexperiments, a herbivore is offered toxic food (e.g. high-metalleaves) or normal food (e.g. low-metal leaves) and its growthon each treatment compared. In Ni hyperaccumulators suchfeeding experiments (Boyd & Martens, 1994; Martens & Boyd,1994; Boyd & Moar, 1999) showed that Ni is toxic to herbivores.
On the other hand, in binary feeding trials the herbivore isgiven the choice between foods rich or poor in toxic metal, inorder to examine its feeding preference. Some authors (Pollard& Baker, 1997; Huitson & Macnair, 2003; Macnair, 2003)have pointed out that binary choices only could be indicativeof a possible herbivore selection pressure. If herbivores prefer toeat the least-accumulating genotypes, they could have promotedthe evolution of hyperaccumulation. Two studies tested thedefence hypothesis with binary feeding choices involvingthe Zn/Cd hyperaccumulator
Thlaspi caerulescens
. Pollard &Baker (1997) used phenotypic plasticity to obtain high- andlow-Zn leaves, cultivating
T. caerulescens
plants on high- andlow-Zn culture solutions. Jhee
et al
. (1999) used the geneticvariability of hyperaccumulation existing between ecotypesand between genotypes within the same population to obtainvariable foliar Zn concentrations among plants growing in thesame medium. Both works concluded that Zn was effectivein defending
T. caerulescens
, as high-Zn leaves were less eaten

New Phytologist
(2005)
165
: 763–772
www.newphytologist.org
©
New Phytologist
(2005)
Research764
than low-Zn ones. More recently, Huitson & Macnair (2003)used the Zn/Cd hyperaccumulator
Arabidopsis halleri
to test thedefence hypothesis. These authors failed to find any protectiverole of Zn, and suggested that secondary metabolites could beinvolved in the deterrence of herbivores. To our knowledge, nostudy has yet investigated the possible role of secondary metabolitesas protective compounds in metal-accumulating plants.
The role of secondary metabolites as repellents for phytophag-ous invertebrates has long been known to mediate the relationsbetween plants and herbivores (Ehrlich & Raven, 1964; Rosenthal& Janzen, 1979; Harborne, 1993). The main defence componentsof the Brassicaceae are sulphur metabolites named glucosi-nolates, which are repulsive to generalist herbivores includingsnails and slugs (van Genderen
et al
., 1997; Gebauer, 2002)but attractive to cruciferous specialists (van Loon
et al
., 2002).Furthermore, secondary metabolites often involve metabolic costsfor plants (Mauricio & Rausher, 1997). This led Boyd (1998)to suggest a trade-off between elemental (e.g. metals) and organicdefences in hyperaccumulator species. These, being defendedagainst herbivores by their high metal concentration, could pro-duce lower amounts of defensive secondary metabolites. Indeed,several hyperaccumulators have lower levels of secondarycompounds compared with congeneric nonaccumulator species(Tolrà
et al
., 1998; Davis & Boyd, 2000). Tolrà
et al
. (2001)found lower levels of glucosinolates in
T. caerulescens
leaveswith higher Zn concentrations, which supports the hypothesisof a phenotypic trade-off between organic and elemental defences.
The aim of this study was to test the defence hypothesisusing the Zn hyperaccumulator
T. caerulescens
. In continentalEurope this species has evolved two main ecotypes: one foundon metal-contaminated soils (metallicolous ecotype), and theother on normal soils (nonmetallicolous ecotype) (Meerts& Van Isacker, 1997; Escarré
et al
., 2000). Both ecotypesevolved in different environments, and metallicolous plantscontain more Zn
in natura
than nonmetallicolous ones.According to the trade-off hypothesis (Boyd, 1998), thedefence strategies of both ecotypes could be different, andsome evolutionary questions may be asked.
In our herbivory experiments we not only consideredZn, but also analysed the glucosinolates of leaves as well astheir Cd, N and dry matter concentrations. The followingquestions were addressed:(i) What is the influence of Zn, glucosinolate, Cd, N and drymatter concentrations on the feeding preferences of snails?(ii) As both
T. caerulescens
ecotypes evolved in differentenvironments, did they develop distinct defence systems? Isthere any trade-off between organic vs elemental defences?
Materials and Methods
Plant materials
We studied four populations of
Thlaspi caerulescens
J. & C. Presl(Brassicaceae) established near Montpellier (southern France):
two metallicolous populations (Malines and Avinières) andtwo nonmetallicolous populations (Séranne and Navacelle);for site characteristics see Escarré
et al
. (2000); Frérot
et al
.(2003); Dubois
et al
. (2003). In southern France
T. caerulescens
accumulates up to 2500 µg Zn g
−
1
d. wt on soils with normalionic balance (Reeves
et al
., 2001), and up to 11 000 (Robinson
et al
., 1998) or 18 000 µg Zn g
−
1
d. wt (Reeves
et al
., 2001)on metalliferous soils.
Two experiments were conducted: experiment 1 (2003) usedglasshouse-grown plants, whereas experiment 2 (2004) wascarried out with field-grown plants.
Experiment 1
Seeds were collected
in natura
in 2000 (at least20 families per population); at Séranne seedlings were collectedin autumn 2002.
Experiment 2
In order to test the snails’ preferences withfield-grown individuals,
≈
20 adult plants from three popu-lations (Malines, Avinières, Séranne) were collected
in natura
and brought back to the laboratory with their soils. The Navacellepopulation was not used because of its low population density.For both metallicolous populations, a sample was collected onhighly contaminated and on less-contaminated soils.
Soil treatments
Experiment 1
The seeds were germinated on burnt clay in aglasshouse, except for the population of Séranne, for whichseedlings were collected in the field. Seedlings were thendistributed in two soil treatments in order to obtain glasshouse-grown plants with contrasting foliar Zn concentrations.Seedlings were planted in 1 l pots filled with garden composteither with 2000 mg Zn kg
−
1
d. wt (contaminated treatment,C) or without Zn added (noncontaminated treatment, NC).Zn was added as ZnSO
4
·7H
2
O, which is highly availableto both ecotypes of
T. caerulescens
(Meerts
et al
., 2003). Weobtained 35 replicates of each population in each treatment,a total of 280 plants (35
×
8 sets of plants).
Experiment 2
Plants were brought back to the laboratory withtheir soil of origin, therefore metallicolous and nonmetallicolousplants were on contaminated and normal soils, respectively.
Binary choices
Each binary choice involved one snail and two leaves from twodifferent treatments and/or populations; each feeding choicewas replicated 10 times simultaneously with 10 different snails.Both leaves of a single binary choice had approximately thesame size. For feeding trials involving glasshouse-grown plants,leaves were cut off from 10 plants randomly chosen amongthe 35 replicates available. Feeding trials were sufficientlydistant in time from each other to avoid a possible effect ofglucosinolate induction after cutting. For binary choices using

©
New Phytologist
(2005)
www.newphytologist.org
New Phytologist
(2005)
165
: 763–772
Research 765
plants collected in the field, leaves were cut from 10 of the 20plants of each population. There were no visible differences(hairiness, thickness) between field- and glasshouse-grown leaves,except for their size, field plants being consistently smaller.
Helix aspersa
Müller (Helicidae), a broad dietary generalistsnail (Kerney & Cameron, 1979) that occurs naturally in theregion studied, was used. Snails were bought from a farm inHuesca, Spain (experiment 1) and from a farm near Montpellier,France (experiment 2). They had been fed with syntheticfood and therefore had never previously been in contact withhigh-metal food. Some individuals of
H. aspersa
were usedin different trials, but at least 7 d apart. Binary choices wereperformed in small black plastic boxes (20
×
13
×
6 cm). Theseexperiments were fixed-time food choices (Lockwood, 1998);one night (13 h) was enough for the snails to make a choicewithout having completely eaten both leaves.
Measurements
Leaf consumption
Before each binary choice, the 10 leaves ofeach sort were numbered and photocopied. After the trial eachleaf was photocopied again, and both photocopies (before andafter the trial) were scanned in order to assess the leaf areaeaten (mm
2
;
-
software).
Nitrogen concentration and leaf dry matter content
The leafdry matter content (LDMC) was estimated according to Garnier
et al
. (2001); one leaf from 10 randomly chosen plants washarvested in each population on each treatment.
Leaf N concentration was measured on the same leaves(dry ground material) by means of an elemental CHN analyser(Thermo Electron, Milan, Italy).
Metal analyses
Leaf Zn concentration was measured on uneatenportions of leaves previously used during binary choices, bymeans of the ZINCON method (Macnair & Smirnoff, 1999).The Cd concentration was measured on entire plants. These wereharvested, washed in deionized water, oven-dried at 60
°
Cfor 3 d, then mineralized in a mixture of nitric and perchloricacid. The Cd concentration was determined by inductivelycoupled plasma optical emission spectroscopy (ICP-OES).
Glucosinolate analysis
Experiment 1
Glucosinolates weremeasured on eight glasshouse-grown plants randomly chosenamong the 35 replicates available for each population
×
treatment combination. Total glucosinolate concentrationwas measured following the enzymatic reaction of glucoserelease (adapted from Tolrà, 1997 and from information fromthe Bureau Community of Reference on Certified ReferenceMaterial, CRM-367). About 0.5 g of leaves (f. wt) was harvested,weighed, immediately frozen in liquid N and stored at
−
80
°
Cbefore analysis. The frozen dried leaves were placed in a 20 mlbottle filled with 10 ml hot methanol (70%). The bottle wasthen placed in a bath of warm water (75
°
C) for 3 min. The
leaves were ground for 7 min using an Omni Mixer (OCIInstruments, Waterbury, CT, USA). After 5 min this mixturewas transferred into a centrifugation tube and stored in thefridge (4–8
°
C). The tubes were centrifuged for 3 min at 3000rpm. The supernatant was decanted into a new centrifugationtube with 0.5 ml Pb–Ba acetate (0.5
). The mixture was againcentrifuged (3 min, 3000 rpm), and the supernatant collectedand preserved in the fridge. 0.5 ml of the extract was put ona column filled with 1 ml DEAE Sephadex A-25 diluted inpyridine acetate 0.5
. Myrosinase (250 µl; 2/3 EU ml
−
1
; SigmaT4528) was injected on the column, which had previously beenwashed with 2 ml water. After incubation (15 h) the column waswashed with 2 ml water, which was collected in a 2 ml Eppendorftube. The glucose concentration, stoichiometrically equivalentto the glucosinolate concentration, was then measured with aglucose kit (Diffchamp Group, Västra Frölunda, Sweden).
Experiment 2
The same procedure for glucosinolate analysiswas applied to field-grown plants. Because of their smaller size,we pooled leaves of two or three individuals per sample in orderto obtain sufficient material for the analyses. The number ofreplicates was lower (
n
= 3–4).The extraction procedure was first tested on a certified reference
material (rapeseeds, Bureau Community of Reference, BCR 367;total glucosinolate concentration 99
±
9 mmol kg
−
1
f. wt).The result (101.8
±
8.3 mmol kg
−
1
f. wt; mean
±
SD,
n
= 9)was not significantly different from the reference material(
t
= 0.018; df = 16;
P
> 0.05).
Statistical analyses
Results of binary choice experiments (leaf area eaten) are notindependent data, that is, the level of consumption of one leafwill depend on the palatability of the other. Such data have tobe analysed using paired
t
-tests (Raffa
et al
., 2002).
Glasshouse-grown plants
Zn, Cd, glucosinolate, N and leafdry matter concentrations were analysed by a mixed partiallynested
(SAS, 1999). Data were ln-transformed to meetthe requirements of the
. Population (random factor)was nested within ecotype (fixed), and treatment (fixed) wascrossed with population. Due to missing values, type III sumof squares was used. Given that we were interested in testingthe significant differences between the eight combinationspopulation
× treatment (four populations, two treatments),we performed comparisons of means by a least-squares meanstest (SAS, 1999). Least-squares means are predicted popula-tion margins; that is, they estimate the marginal means overa balanced population. Computed for each effect (in our casepopulation × treatment), the LSMEANS statement performsmultiple comparisons on interactions as well as main effects.
Field-grown plants Foliar Zn, Cd and glucosinolate concen-trations were analysed by a one-way (SAS, 1999). As only

New Phytologist (2005) 165: 763–772 www.newphytologist.org © New Phytologist (2005)
Research766
three populations (Avinières, Malines, Séranne) were available,the ecotype factor could not be used. Data were ln-transformedto meet the requirements of the . Comparisons of meanswere performed by least-squares means tests.
Finally, a proportion test (Fisher’s exact test) was used toexamine whether the proportion of cases where high-Zn leaveswere a deterrent was significantly different from the propor-tion of cases where high-Zn leaves were not a deterrent. Thesame proportion tests were performed for glucosinolate, Cd,N and dry matter concentrations.
Results
Metal concentration of glasshouse-grown plants
Zinc The nested showed significant differences betweensoil treatments (F = 73.31; df = 1,2; P < 0.01). Plants growingon Zn-contaminated (C) soil had significantly higher Znconcentrations (5242 ± 199 µg g−1 d. wt, mean ± SE) than thosegrowing on noncontaminated (NC) soil (2618 ± 120 µg g−1
d. wt) (Table 1). All other effects (ecotype, population andtwo-way interactions) were not significant.
Cadmium There were no significant differences in Cdconcentration between ecotypes (F = 10.71; df = 1, 2; P = 0.08),although metallicolous populations tended to contain moreCd (75.9 ± 7.1 µg g−1 d. wt) than nonmetallicolous ones(30.2 ± 4.4 µg g−1 d. wt) (Table 1). There was a significantinteraction of population × treatment within ecotype (F =
6.43; df = 2,53; P = 0.003). One nonmetallicolous population(Séranne) had a significantly higher Cd concentration on theNC treatment, while the other populations were insensitive tothe treatment. The other effects were not significant.
Metal concentration of field-grown plants
Zinc There were significant differences between populationsfor foliar Zn concentration (F = 10.20; df = 4,39; P < 0.001)(Table 2). It is worth noting that only plants from Avinières(highly contaminated site) exceeded the arbitrary threshold ofZn hyperaccumulation (> 1% leaf d. wt), which is not oftenreached in natura (see Fig. 3b; Assunção et al., 2003; Molitoret al., 2004).
Cadmium There was a significant difference between popu-lations for Cd concentration (F = 169.2; df = 2,10; P < 0.001)(Table 2), all three populations containing significantly differentCd concentrations.
Glucosinolate concentration of glasshouse-grown plants
There were no significant differences between ecotypes, popu-lations or treatments, although nonmetallicolous populationstended to contain more glucosinolates (6.50 ± 0.77 µmol g−1
f. wt, mean ± SE) than metallicolous ones (2.95 ± 0.33µmol g−1 f. wt) (F = 6.06; df = 1,2; P = 0.13). However, therewas a significant interaction between populations (withinecotype) and treatments (F = 7.99; df = 2,59; P < 0.001). In
Table 1 Foliar composition of four Thlaspi caerulescens populations grown on high- and low-Zn soils. (a) Leaf concentrations of glucosinolates, nitrogen and leaf dry matter content (LDMC); (b) elemental concentrations in above-ground biomass
Parameter
Metallicolous Nonmetallicolous
Avinières Malines Navacelle Séranne
C NC C NC C NC C NC
(a)Glucosinolates 1.8a 2.6ab 5.2cd 2.5ab 3.3bc 12.5e 4.2bc 6.2d
(0.4) (0.4) (0.9) (0.3) (0.6) (1.5) (0.7) (0.8)N 0.64a 1.03ab 0.75a 0.84ab 0.90ab 1.08ab 1.14ab 1.47b
(0.04) (0.06) (0.04) (0.08) (0.07) (0.10) (0.23) (0.20)LDMC 21.0b 18.0a 21.7b 20.3ab 21.2b 19.4ab 20.6ab 19.2ab
(0.01) (0.01) (0.01) (0.01) (0.01) (0.01) (0.01) (0.01)
(b)Zn 4760c 2190a 5470c 2840b 5390c 2410ab 5350c 3270b
(422) (183) (413) (270) (343) (182) (433) (310)Cd 60def 50bcd 85ef 102f 23b 34bc 12a 58cde
(10) (12) (14) (14) (2) (8) (3) (11)
(a) Glucosinolates (µmol g−1 f. wt; n = 8–9); N (% d. wt; n = 5–6); LDMC (% d. wt; n = 10).(b) Elemental concentrations (µg g−1 d. wt): Zn (n = 20–29); Cd (n = 7–8).Data are means (SE). Mean values that differ significantly (P < 0.05) are denoted by different superscripts.C, treatment contaminated with 2000 mg Zn kg−1 d. wt; NC, noncontaminated treatment.
© New Phytologist (2005) www.newphytologist.org New Phytologist (2005) 165: 763–772
Research 767
nonmetallicolous populations, individuals growing on NCsoil had significantly more glucosinolates than those growingon the C treatment. For metallicolous populations the gluco-sinolate concentration of Avinières was not affected by thetreatment, while the Malines population had significantlymore glucosinolates on the C treatment (Table 1).
Glucosinolate concentration of field-grown plants
There were significant differences between populations (F =10.09; df = 4,14; P < 0.001) (Table 2). Individuals from Sérannehad the highest values, which were not significantly differentfrom those of the plants of Malines (low-contaminated soil).The other plants showed significantly lower values.
Nitrogen and dry matter concentrations of glasshouse-grown plants
Leaf N concentrations ranged from 0.64 to 1.47%. There wereno significant differences between ecotypes, treatments and theirinteractions. However, nonmetallicolous populations tendedto contain more N than metallicolous ones (Tables 1; F = 7.05;df = 1,2; P = 0.12). Plants on the NC treatment also tendedto have more N compared with those growing on the Ctreatment (Table 1; F = 7.25; df = 1,2; P = 0.11). For the popu-lations of Séranne and Avinières, plants from the NC treatmenthad significantly higher N concentrations compared with theC treatment (Table 1).
The LDMC ranged from 18.1 to 21.7% depending on popu-lation and treatment. Leaves from C soil had a significantlyhigher LDMC (21.1 ± 0.03, mean ± SE) compared with NCones (19.2 ± 0.03) (Table 1; F = 22.1; df = 1,2; P = 0.04). Onlythe population of Avinières showed significant differencesbetween treatments (Table 1).
N and LDMC were not measured for field-grownplants.
Binary choices
Within-population feeding choices Four within-populationfeeding choices were conducted, which involved leaves ofhigh- and low-Zn plants of the same population. Fig. 1 showsthe results of snail feeding preferences in relation to the Znconcentration and total glucosinolate concentration of leaves.Foliar Zn concentration does not consistently control thefeeding choices of snails, as high-Zn leaves were less eaten onlyin the Malines choice. The differences in glucosinolate concen-trations between leaves could explain the feeding preferencesin the Malines and Avinières choices. However, glucosinolatescannot explain the snails’ behaviour in the Navacelle andSéranne trials. It is worth noting that, considering absolutequantities, leaves of nonmetallicolous populations were lesseaten than leaves of metallicolous populations, particularly forthe NC treatment.
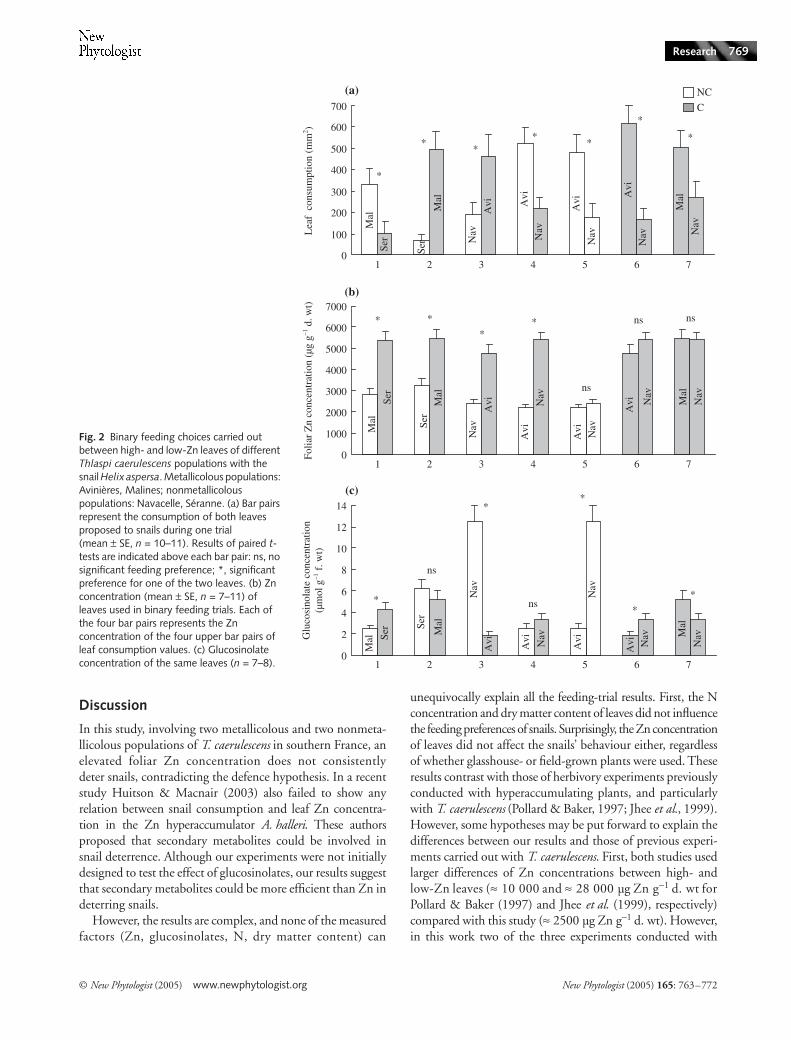
Between-population feeding choices Trials with different Zn concentrations In four other feeding-choice experiments(Fig. 2, #1–4) the snails were offered a high- and a low-Znleaf originating from two different populations. Fig. 2 showsthe results of snail feeding preferences in relation to foliarZn and total glucosinolate concentrations. Snails preferredlow-Zn leaves in choices #1 and 4, whereas high-Zn leaveswere more eaten in choices #2 and 3. However, in trials #1and 3 low-glucosinolate leaves were eaten more. It is worthnoting that in these four choices, the leaf eaten less alwaysbelongs to the nonmetallicolous ecotype, whatever the level ofmetals.
Trials with similar Zn concentrations Three binary choices (Fig. 2,#5–7) were conducted with leaves of similar Zn concentrations.Snails showed clear preferences despite close Zn levels: low-glucosinolate leaves were preferred in trials #5 and 6, whereashigh-glucosinolate leaves were chosen in trial #7.
Table 2 Analysis of field-grown Thlaspi caerulescens plants. (a) Foliar concentration of glucosinolates; (b) elemental concentrations of leaves
Parameter
Metallicolous Nonmetallicolous
Avinières Malines Séranne
High Zn Low Zn High Zn Low Zn
(a)Glucosinolates 2.1a (0.3) 1.4a (0.1) 2.1a (0.3) 3.2b (0.3) 4.2b (0.7)
(b)Zn 13 500c (1860) 5290b (408) 6880b (1120) 2430a (239) 2510a (295)Cd 1710c (125) – 675b (115) – 13a (1)
(a) Glucosinolates (µmol g−1 f. wt; n = 7–8).(b) Elemental concentrations (µg g−1 d. wt): Zn (n = 7–11); Cd (n = 3–5).Data are means (SE). Mean values that differ significantly (P < 0.05) are denoted by different superscripts. Avinières and Malines populations were both collected at two different places, with high and low Zn.

New Phytologist (2005) 165: 763–772 www.newphytologist.org © New Phytologist (2005)
Research768
Feeding trials with field-grown plants Three feeding trialswere also performed with leaves cut from field-grown plants.Fig. 3 shows the results of snail preferences in relation to thefoliar Zn and glucosinolate concentrations. Zinc concentrationdid not affect feeding choices, as snails preferred to eat high-
Zn leaves in choices #8 and 9. In trial #10 snails exhibited nopreference, whereas both leaves differed significantly in theirZn and glucosinolate concentrations. On the other hand,differences in glucosinolate concentrations between leavescould explain snails’ preferences in trials #8 and 9.
Proportion tests In four of the 14 feeding trials (28.5%)snails preferred high-Zn leaves; in three trials (21.4%) high-Zn leaves were less consumed; in three others there were nodifferences in Zn concentration (Table 3). In the last threeruns (Avinières, Navacelle and Séranne within-populationtrials) there were no feeding preferences. The number of caseswhere high-Zn leaves were a deterrent is not significantlydifferent from the number of cases where high-Zn leaves werepreferred (Fisher’s exact test: P = 1.00). Therefore a higher Znconcentration does not appear to protect leaves from snailconsumption. Similarly, a higher Cd concentration does not detersnails either, as high-Cd leaves were significantly preferred(Fisher’s exact test: P = 0.016).
On the other hand, high-glucosinolate leaves were preferredin one trial (7.1%), significantly less consumed in seven trials(50%), and in three trials there were no significant differencesin glucosinolate concentration (Table 3). In the last three runsthe snails exhibited no feeding preferences (Avinières, Navacelleand Séranne within-population trials). The number of trials wherehigh-glucosinolate leaves were a deterrent is significantly differentfrom the number of trials where high-glucosinolate leaves werepreferred (Fisher’s exact test: P = 0.03).
As Zn and glucosinolates may interact, we re-examinedthe cases where high-Zn leaves were less consumed. In two ofthese three cases, leaves with more Zn also had the highestconcentration of glucosinolates. Therefore, leaves with higherZn concentration were less consumed in only one trial. Inthe same way, for glucosinolates there were five trials in whichhigh-glucosinolate leaves were less consumed.
We performed the same proportion tests with N concen-tration and dry matter content of leaves as possible attractiveor deterrent factors, respectively. None of these appears tohave affected the feeding preferences of snails (Fisher’s exacttests: N, P = 0.48; LDMC, P = 1.00).
Fig. 1 Snails (Helix aspersa) were presented with binary feeding choices between high- and low-Zn leaves of the same population of Thlaspi caerulescens. Metallicolous populations: Avinières, Malines; nonmetallicolous populations: Navacelle, Séranne. (a) Zn concentration (mean ± SE, n = 20–29) of leaves used in binary feeding choices. Each of the four bar pairs represents the Zn concentration of the four upper bar pairs of leaf consumption values. (b) Bar pairs represent the consumption of both leaves of one trial (mean ± SE, n = 10–11). Results of paired t-tests are indicated above each bar pair: ns, no significant feeding preference; *, significant preference. (c) Glucosinolate concentration of the same leaves (n = 7–8).
Table 3 Characteristics (levels of Zn and glucosinolates) of the preferred Thlaspi caerulescens leaves in pairwise feeding choices where snails exhibited a significant preference
Zn level
Glucosinolate level
High Low Not different
High 3 (#3, #8, #9) 1 (#2)Low 2 (Malines, #1) 1 (#4)Not different 1 (#7) 2 (#5, #6)
For each level of Zn and glucosinolate the number of trials in the corresponding cases is shown.
© New Phytologist (2005) www.newphytologist.org New Phytologist (2005) 165: 763–772
Research 769
Discussion
In this study, involving two metallicolous and two nonmeta-llicolous populations of T. caerulescens in southern France, anelevated foliar Zn concentration does not consistentlydeter snails, contradicting the defence hypothesis. In a recentstudy Huitson & Macnair (2003) also failed to show anyrelation between snail consumption and leaf Zn concentra-tion in the Zn hyperaccumulator A. halleri. These authorsproposed that secondary metabolites could be involved insnail deterrence. Although our experiments were not initiallydesigned to test the effect of glucosinolates, our results suggestthat secondary metabolites could be more efficient than Zn indeterring snails.
However, the results are complex, and none of the measuredfactors (Zn, glucosinolates, N, dry matter content) can
unequivocally explain all the feeding-trial results. First, the Nconcentration and dry matter content of leaves did not influencethe feeding preferences of snails. Surprisingly, the Zn concentrationof leaves did not affect the snails’ behaviour either, regardlessof whether glasshouse- or field-grown plants were used. Theseresults contrast with those of herbivory experiments previouslyconducted with hyperaccumulating plants, and particularlywith T. caerulescens (Pollard & Baker, 1997; Jhee et al., 1999).However, some hypotheses may be put forward to explain thedifferences between our results and those of previous experi-ments carried out with T. caerulescens. First, both studies usedlarger differences of Zn concentrations between high- andlow-Zn leaves (≈ 10 000 and ≈ 28 000 µg Zn g−1 d. wt forPollard & Baker (1997) and Jhee et al. (1999), respectively)compared with this study (≈ 2500 µg Zn g−1 d. wt). However,in this work two of the three experiments conducted with
Fig. 2 Binary feeding choices carried out between high- and low-Zn leaves of different Thlaspi caerulescens populations with the snail Helix aspersa. Metallicolous populations: Avinières, Malines; nonmetallicolous populations: Navacelle, Séranne. (a) Bar pairs represent the consumption of both leaves proposed to snails during one trial (mean ± SE, n = 10–11). Results of paired t-tests are indicated above each bar pair: ns, no significant feeding preference; *, significant preference for one of the two leaves. (b) Zn concentration (mean ± SE, n = 7–11) of leaves used in binary feeding trials. Each of the four bar pairs represents the Zn concentration of the four upper bar pairs of leaf consumption values. (c) Glucosinolate concentration of the same leaves (n = 7–8).

New Phytologist (2005) 165: 763–772 www.newphytologist.org © New Phytologist (2005)
Research770
field-grown plants involved differences in Zn concentrationsbetween leaves as high as 4500 and 11 000 µg Zn g−1 d. wt,and high-Zn leaves still did not deter snails. Binary choicesusing small differences in Zn concentration may, arguably,best mimic natural situations. Assuming that herbivores are
the evolutionary force at the origin of the evolution of hyper-accumulation, these should be capable of discriminating therelatively small variations in Zn concentration existing withinpopulations. The apparent feeding indifference of snails towardsleaf Zn concentration may suggest that they are simply not ableto detect Zn. However, this hypothesis needs to be confirmed byexperiments using synthetic food with varying Zn concentration.
In addition, we found a significant proportion (50%) oftrials where high-glucosinolate leaves deterred snails. It is thusnot impossible that low-Zn leaves used by Pollard & Baker(1997) and Jhee et al. (1999) also had a low glucosinolateconcentration. The effects of Zn and glucosinolates could thushave been confounded in those studies. Another possibilitycould be that these populations had overall low glucosinolatelevels and, in this case, a high Zn concentration could havebeen a deterrent for herbivores.
It is worth noting that, in pairwise choice experiments involv-ing leaves from different ecotypes, leaves of nonmetallicolousplants were very often less consumed than those of metalli-colous plants (in eight out of nine cases). This result can berelated to the tendency of nonmetallicolous populations tohave more glucosinolates in glasshouse- and field-grown plants(see Results). As suggested by the trade-off hypothesis (Boyd,1998), we thus found lower constitutive glucosinolate con-centrations in metallicolous populations. However, as the latterare definitely less protected against snails, we cannot concludethat elemental and organic defences are equally effective towardsherbivores. In addition, preliminary field experiments suggestthat the intensity of herbivory is lower on metalliferous sites(Noret, 2003). Therefore metallicolous populations could havereduced their production of costly organic defences (glucosi-nolates and/or other defence compounds). This would explainwhy metallicolous leaves are less defended against herbivores.
In conclusion, this work does not support the hypothesisthat Zn hyperaccumulation evolved as a defence againstherbivores in T. caerulescens. The complexity of our results warnsagainst simplistic explanations of feeding preferences, andhighlights the importance of taking into account secondarycompounds, in addition to metals, in herbivory studies withhyperaccumulating plants. In Mediterranean populations ofT. caerulescens Zn does not play a key role in the palatabilityof leaves to snails. Secondary compounds, such as glucosi-nolates, could be more efficient than metals in deterringgeneralist herbivores. Further work considering both metals andsecondary metabolites should help us understand their respec-tive roles in plant defence. Finally, the inadvertent uptakehypothesis, which has not yet been tested, should hold theattention of researchers interested in the evolution of metalhyperaccumulation.
Acknowledgements
We greatly thank Claude Lefèbvre for critically readingthe manuscript and Pierre-Olivier Cheptou for constructive
Fig. 3 Snails (Helix aspersa) were presented with pairwise feeding trials with leaves of field-grown plants of different Thlaspi caerulescens populations. Metallicolous populations: Avinières, Malines; nonmetallicolous population: Séranne. (a) Bar pairs represent the consumption of both leaves proposed to snails during one trial (mean ± SE, n = 7–9). Results of paired t-tests are indicated above each bar pair: ns, no significant feeding preference; *, significant preference for one of the two leaves. (b) Zn concentration (mean ± SE, n = 7–10) of leaves used in binary feeding trials. Each of the four bar pairs represents the Zn content of the four upper bar pairs of leaf consumption. (c) Glucosinolate concentration of the same leaves (n = 3–4).

© New Phytologist (2005) www.newphytologist.org New Phytologist (2005) 165: 763–772
Research 771
comments on an earlier draft. We are also grateful to BrunoBuatois, Christian Collin and Anabelle Dos Santos for technicalassistance. We also thank Dominique Bordat (CIRAD). Thisresearch was partially supported by the ‘Environnement, Vieet Société’ and ‘Biodiversité’ CNRS programs, by a ResearchFellow grant of the FNRS (Belgium), by FRFC (contract2.4565.02) and by the Spanish DGICYT (project BFI 2001-2475-CO2-01).
ReferencesAssunção AGL, Schat H, Aarts MGM. 2003. Thlaspi caerulescens, an
attractive model species to study heavy metal hyperaccumulation in plants. New Phytologist 159: 351–360.
Boyd RS. 1998. Hyperaccumulation as a plant defensive strategy. In: Brooks RR, ed. Plants that Hyperaccumulate Heavy Metals. Wallingford, UK: CAB International, 181–201.
Boyd RS, Martens SN. 1992. The raison d’être of metal hyperaccumulation by plants. In: Baker AJM, Proctor J, Reeves RD, eds. The Vegetation of Ultramafic (Serpentine) Soils. Andover, UK: Intercept, 279–289.
Boyd RS, Martens SN. 1994. Nickel hyperaccumulated by Thlaspi montanum var. montanum is acutely toxic to an insect herbivore. Oikos 70: 21–25.
Boyd RS, Moar WJ. 1999. The defensive function of Ni in plants: response of the polyphagous herbivore Spodoptera exigua (Lepidoptera: Noctuidae) to hyperaccumulator and accumulator species of Streptanthus (Brassicaceae). Oecologia 118: 218–224.
Brooks RR, ed. 1998. Plants that Hyperaccumulate Heavy Metals. Wallingford, UK: CAB International.
Davis MA, Boyd RS. 2000. Dynamics of Ni-based defence and organic defences in the Ni hyperaccumulator, Streptanthus polygaloides (Brassicaceae). New Phytologist 146: 211–217.
Dubois S, Cheptou PO, Petit C, Meerts P, Poncelet M, Vekemans X, Lefèbvre C, Escarré J. 2003. Genetic structure and mating systems of metallicolous and nonmetallicolous populations of Thlaspi caerulescens. New Phytologist 157: 633–641.
Ehrlich PR, Raven PH. 1964. Butterflies and plants: a study in coevolution. Evolution 18: 586–608.
Escarré J, Lefèbvre C, Gruber W, Leblanc M, Lepart J, Rivière Y, Delay B. 2000. Zinc and Cd hyperaccumulation by Thlaspi caerulescens from metallicolous and nonmetallicolous sites in the Mediterranean area: implications for phytoremediation. New Phytologist 145: 429–437.
Frérot H, Petit C, Lefèbvre C, Gruber W, Collin C, Escarré J. 2003. Zinc and cadmium accumulation in controlled crosses between metallicolous and nonmetallicolous populations of Thlaspi caerulescens (Brassicaceae). New Phytologist 157: 643–648.
Garnier E, Shipley B, Roumet C, Laurent G. 2001. A standardized protocol for the determination of specific leaf area and leaf dry matter concentration. Functional Ecology 15: 688–695.
Gebauer J. 2002. Effects of different nitrogen concentrations in wheat leaves on the food choice and feeding activity of the grey field slug, Deroceras reticulatum (Müller) under laboratory conditions. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz 109: 421–429.
van Genderen H, Schoonhoven LM, Fuchs A. 1997. Chemisch–ecologische flora van Nederland en België. Utrecht, the Netherlands: KNNV.
Harborne JB. 1993. Introduction to Ecological Biochemistry. London, UK: Academic Press.
Huitson SB, Macnair MR. 2003. Does zinc protect the zinc hyperaccumulator Arabidopsis halleri from herbivory by snails? New Phytologist 159: 453–459.
Jhee EM, Dandridge KL, Christy AM, Pollard AJ. 1999. Selective herbivory on low-zinc phenotypes of the hyperaccumulator Thlaspi caerulescens (Brassicaceae). Chemoecology 9: 93–95.
Kerney MP, Cameron RAD. 1979. A Field Guide to the Land Snails of North-West Europe. London, UK: Collins & Sons.
Lockwood JR. 1998. On the statistical analysis of multiple-choice feeding preference experiments. Oecologia 116: 475–481.
van Loon JJA, Wang CZ, Nielsen JK, Gols R, Qiu YT. 2002. Flavonoids from cabbage are feeding stimulants for diamondback moth larvae additional to glucosinolates: chemoreception and behaviour. Entomologia Experimentalis et Applicata 104: 27–34.
Macnair MR. 2003. The hyperaccumulation of metals by plants. Advances in Botanical Research 40: 63–105.
Macnair MR, Smirnoff N. 1999. Use of zincon to study uptake and accumulation of zinc by zinc tolerant and hyperaccumulating plants. Communications in Soil Science and Plant Analysis 30: 1127–1136.
Martens SN, Boyd RS. 1994. The ecological significance of nickel hyperaccumulation: a plant chemical defense. Oecologia 98: 379–384.
Mauricio R, Rausher MD. 1997. Experimental manipulation of putative selective agents provides evidence for the role of natural enemies in the evolution of plant defense. Evolution 51: 1435–1444.
Meerts P, Van Isacker N. 1997. Heavy metal tolerance and accumulation in metallicolous and non-metallicolous populations of Thlaspi caerulescens. Plant Ecology 133: 221–231.
Meerts P, Duchêne P, Gruber W, Lefèbvre C. 2003. Metal accumulation and competitive ability in metallicolous and non-metallicolous Thlaspi caerulescens fed with different Zn salts. Plant and Soil 249: 1–8.
Molitor M, Dechamps C, Gruber W, Meerts P. 2004. Thlaspi caerulescens on non metalliferous soil in Luxembourg: ecological niche and genetic variation in mineral element composition. New Phytologist, In press.
Noret N. 2003. Contribution à l’étude de la signification écologique et évolutive de l’hyperaccumulation du Zn chez Thlaspi caerulescens (Brassicaceae): test de l’hypothèse d’une défense contre les herbivores. Mémoire de DEA. Thesis, Université de Montpellier II, Montpellier, France.
Pollard AJ. 2000. Metal hyperaccumulation: a model system for coevolutionary studies. New Phytologist 146: 179–181.
Pollard AJ, Baker AJM. 1997. Deterrence of herbivory by zinc hyperaccumulation in Thlaspi caerulescens (Brassicaceae). New Phytologist 135: 655–658.
Raffa KF, Havill NP, Nordheim EV. 2002. How many choices can your test animal compare effectively? Evaluating a critical assumption of behavioral preference tests. Oecologia 133: 422–429.
Reeves RD, Schwartz C, Morel JL, Edmondson J. 2001. Distribution and metal-accumulating behavior of Thlaspi caerulescens and associated metallophytes in France. International Journal of Phytoremediation 3: 145–172.
Robinson BH, Leblanc M, Petit D, Brooks RR, Kirkman JH, Gregg PEH. 1998. The potential of Thlaspi caerulescens for phytoremediation of contaminated soils. Plant and Soil 203: 47–56.
Rosenthal GA, Janzen DH, eds. 1979. Herbivores. Their Interaction with Secondary Plant Metabolites. London, UK: Academic Press.
SAS. 1999. SAS/STAT User’s Guide, Version 8.0. Cary, NC, USA: SAS Institute.Tolrà RP. 1997. Mecanismos de tolerancia a la toxicidad por Zn en
Thlaspi caerulescens. PhD thesis, Universidad Autónoma de Barcelona, Spain.
Tolrà RP, Gomis M, Barceló J, Poschenrieder C. 1998. Influence of metal hyperaccumulation on glucosinolate concentration. In: Proceedings of the VII National–III Iberian Symposium on Plant Mineral Nutrition. Madrid, Spain: Autonomous University Press, 471–476.
Tolrà RP, Poschenrieder C, Alonso R, Barceló D, Barceló J. 2001. Influence of zinc hyperaccumulation on glucosinolates in Thlaspi caerulescens. New Phytologist 151: 621–626.

New Phytologist (2005) 165: 763–772 www.newphytologist.org © New Phytologist (2005)
Research772
About New Phytologist
• New Phytologist is owned by a non-profit-making charitable trust dedicated to the promotion of plant science, facilitating projectsfrom symposia to open access for our Tansley reviews. Complete information is available at www.newphytologist.org.
• Regular papers, Letters, Research reviews, Rapid reports and Methods papers are encouraged. We are committed to rapidprocessing, from online submission through to publication ‘as-ready’ via OnlineEarly – the 2003 average submission to decision timewas just 35 days. Online-only colour is free, and essential print colour costs will be met if necessary. We also provide 25 offprintsas well as a PDF for each article.
• For online summaries and ToC alerts, go to the website and click on ‘Journal online’. You can take out a personal subscription tothe journal for a fraction of the institutional price. Rates start at £109 in Europe/$202 in the USA & Canada for the online edition(click on ‘Subscribe’ at the website).
• If you have any questions, do get in touch with Central Office ([email protected]; tel +44 1524 592918) or, for a localcontact in North America, the USA Office ([email protected]; tel 865 576 5261).
Related Documents











