Palaeontological evidence of communication between the Central Paratethys and the Mediterranean in the late Badenian/early Serravalian Miloš Bartol a, ⁎, Vasja Mikuž a,b , Aleksander Horvat a a Ivan Rakovec Institute of Palaeontology, Ljubljana, Slovenia b Faculty of Natural Sciences and Engineering, Department of Geology, Ljubljana, Slovenia abstract article info Article history: Received 19 April 2013 Received in revised form 23 November 2013 Accepted 8 December 2013 Available online 15 December 2013 Keywords: Slovenian Corridor Calcareous nannoplankton Diatoms Pereiraea gervaisi In the Middle Miocene the area of present-day eastern Slovenia was situated at the eastern mouth of the Slovenian Corridor – a hypothetical seaway connecting the Central Paratethys and the Mediterranean. The exact location and time of closure of a seaway between the two neighbouring marine realms are still a matter of debate. We studied Badenian (late Langhian and early Serravallian) calcareous nannoplankton assemblages from eastern Slovenia for their palaeogeographical significance by comparing them to contemporaneous Central Paratethyan and Mediterranean assemblages and the assemblages described in selected ODP and IODP reports. Assemblage composition and the sequence of biostratigraphic events observed in Slovenia were very similar to that in several Mediterranean sites. Comparison with the World Ocean suggests that this similarity reflects com- munication between the Mediterranean and Paratethyan populations rather than a universal trend of nanno- plankton evolution. Late Badenian assemblages from different localities within the Central Paratethys exhibit considerable variation with respect to the presence of discoasters (considered as typical of the Mediterranean) and the presence of Rhabdosphaera poculi and Hayella challengeri (considered as typical of the Eastern Paratethys). Our results suggest the mixing of Mediterranean and Eastern Paratethyan influences throughout the entire Central Paratethys during the late Badenian. The observed variability explains how opposing views on the status of the marine connection between the Central Paratethys and the Mediterranean in the late Badenian can arise from palaeontological stud- ies of fossil assemblages considering only a single site or a small area. The gastropod species Pereiraea gervaisi migrated from the Mediterranean to the Central Paratethys – where it is only known from the immediate proximity of the hypothesised Slovenian Corridor. This supports the thesis that the communication between the two seas took place via the Slovenian Corridor and not some other marine con- nection like the hypothetical Axios (Vardar) Trench. © 2013 Elsevier B.V. All rights reserved. 1. Introduction 1.1. Geological setting During the Middle Miocene the area of present-day eastern Slovenia represented the western margin of the Pannonian Basin sys- tem of the Central Paratethys. In recent years the Middle Miocene sed- iments of eastern Slovenia were the subject of two independent studies. The Paratethyan sedimentary sequences from the Mura De- pression (western part of the Mura–Zala Basin) were studied for cal- careous nannoplankton (Bartol, 2009) while a study of the Krka Basin deposits was concerned with siliceous phytoplankton and mol- luscs (Horvat, 2004; Mikuž, 2009). The Mura Depression (western part of the Mura–Zala Basin in Fodor et al., 2002; Márton et al., 2002) was a smaller sub-basin of the Pannonian Basin system in the western Central Paratethys. The South Burgenland Swell and the Kungota fault separated this basin from the Styrian Basin in the north; Pohorje and Kozjak lay on its western coast, while its southern boundary ran near the Donat fault (Márton et al., 2002; Gosar, 2005). The temporally variable occurrences of Miocene marine sediments indicate that the Miocene sedimentary basins in the area of the present-day Slovenia did not represent a single interconnected region (Fig. 1). Furthermore, contemporaneous sediments from various locali- ties, often display a great variety of facies even in close proximity to one another, which indicates a highly complex sedimentary environment (e. g., Bartol, 2009). Sedimentation took place in narrow sedimentary areas (troughs) of varying dynamics of extensional subsidence, separat- ed by intermediate thresholds or submarine highs (Pleničar and Nosan, 1958). On Mt. Pohorje and Kozjak, north of the Periadriatic lineament, deep marine sedimentation started already in Karpatian and continued through the entire Badenian (Fodor et al., 2002). The basins south of the Periadriatic and Donat fault zone (Fig. 1) were flooded during the lower Badenian transgression (Jelen et al., 2008; Pavšič and Horvat, 2009) while in the area of the southernmost Kbasin – the Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157 ⁎ Corresponding author at: Department of Earth Sciences, Palaeobiology, Uppsala, Sweden. E-mail addresses: [email protected], [email protected] (M. Bartol). 0031-0182/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.palaeo.2013.12.009 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Palaeontological evidence of communication between the CentralParatethys and the Mediterranean in the late Badenian/early Serravalian
Miloš Bartol a,⁎, Vasja Mikuž a,b, Aleksander Horvat a
a Ivan Rakovec Institute of Palaeontology, Ljubljana, Sloveniab Faculty of Natural Sciences and Engineering, Department of Geology, Ljubljana, Slovenia
⁎ Corresponding author at: Department of Earth SciSweden.
E-mail addresses: [email protected], Milos.Barto
0031-0182/$ – see front matter © 2013 Elsevier B.V. All rihttp://dx.doi.org/10.1016/j.palaeo.2013.12.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 19 April 2013Received in revised form 23 November 2013Accepted 8 December 2013Available online 15 December 2013
Keywords:Slovenian CorridorCalcareous nannoplanktonDiatomsPereiraea gervaisi
In the Middle Miocene the area of present-day eastern Slovenia was situated at the eastern mouth of theSlovenian Corridor – a hypothetical seaway connecting the Central Paratethys and the Mediterranean. Theexact location and time of closure of a seaway between the two neighbouring marine realms are still a matterof debate. We studied Badenian (late Langhian and early Serravallian) calcareous nannoplankton assemblagesfrom eastern Slovenia for their palaeogeographical significance by comparing them to contemporaneous CentralParatethyan and Mediterranean assemblages and the assemblages described in selected ODP and IODP reports.Assemblage composition and the sequence of biostratigraphic events observed in Slovenia were very similar tothat in several Mediterranean sites. Comparisonwith theWorld Ocean suggests that this similarity reflects com-munication between the Mediterranean and Paratethyan populations rather than a universal trend of nanno-plankton evolution.Late Badenian assemblages from different localities within the Central Paratethys exhibit considerable variationwith respect to the presence of discoasters (considered as typical of the Mediterranean) and the presence ofRhabdosphaera poculi andHayella challengeri (considered as typical of the EasternParatethys). Our results suggestthemixing ofMediterranean and Eastern Paratethyan influences throughout the entire Central Paratethys duringthe late Badenian. The observed variability explains how opposing views on the status of the marine connectionbetween the Central Paratethys and theMediterranean in the late Badenian can arise frompalaeontological stud-ies of fossil assemblages considering only a single site or a small area.The gastropod species Pereiraea gervaisimigrated from theMediterranean to the Central Paratethys –where it isonly known from the immediate proximity of the hypothesised Slovenian Corridor. This supports the thesis thatthe communication between the two seas took place via the Slovenian Corridor and not some othermarine con-nection like the hypothetical Axios (Vardar) Trench.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
1.1. Geological setting
During the Middle Miocene the area of present-day easternSlovenia represented the western margin of the Pannonian Basin sys-tem of the Central Paratethys. In recent years theMiddle Miocene sed-iments of eastern Slovenia were the subject of two independentstudies. The Paratethyan sedimentary sequences from the Mura De-pression (western part of the Mura–Zala Basin) were studied for cal-careous nannoplankton (Bartol, 2009) while a study of the KrkaBasin deposits was concerned with siliceous phytoplankton and mol-luscs (Horvat, 2004; Mikuž, 2009). The Mura Depression (westernpart of the Mura–Zala Basin in Fodor et al., 2002; Márton et al., 2002)was a smaller sub-basin of the Pannonian Basin system in the western
ences, Palaeobiology, Uppsala,
[email protected] (M. Bartol).
ghts reserved.
Central Paratethys. The South Burgenland Swell and the Kungota faultseparated this basin from the Styrian Basin in the north; Pohorje andKozjak lay on its western coast, while its southern boundary ran nearthe Donat fault (Márton et al., 2002; Gosar, 2005).
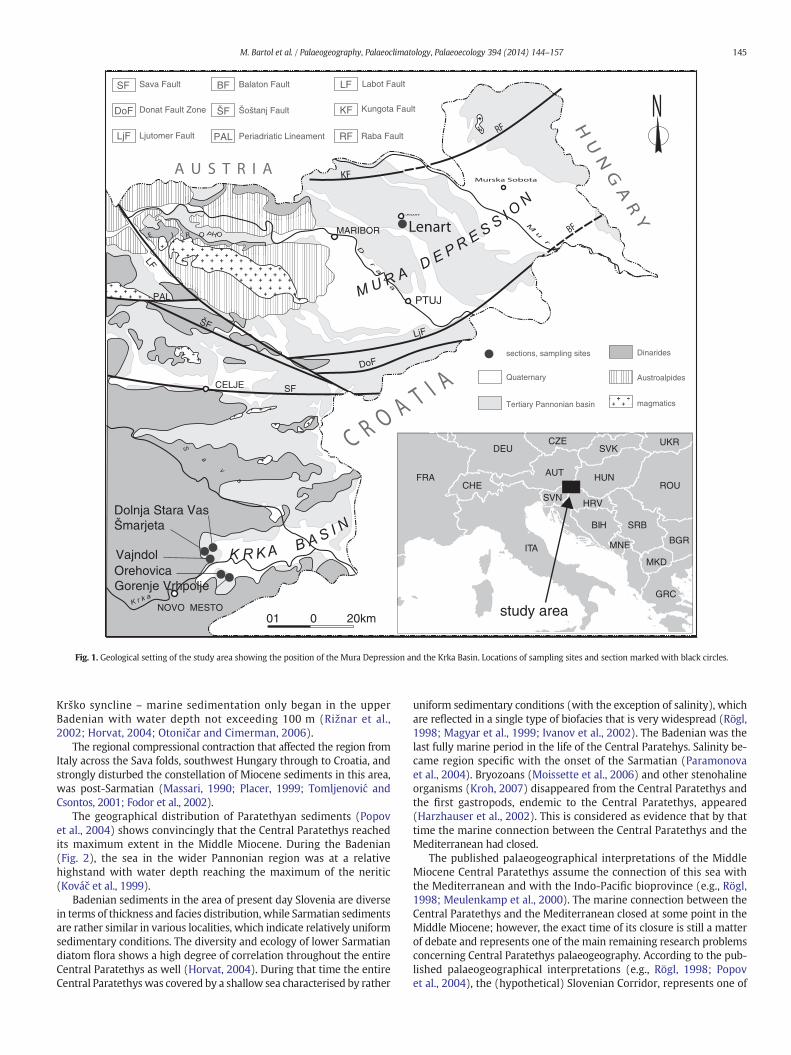
The temporally variable occurrences of Miocene marine sedimentsindicate that the Miocene sedimentary basins in the area of thepresent-day Slovenia did not represent a single interconnected region(Fig. 1). Furthermore, contemporaneous sediments from various locali-ties, often display a great variety of facies even in close proximity to oneanother, which indicates a highly complex sedimentary environment(e. g., Bartol, 2009). Sedimentation took place in narrow sedimentaryareas (troughs) of varying dynamics of extensional subsidence, separat-ed by intermediate thresholds or submarine highs (Pleničar and Nosan,1958). On Mt. Pohorje and Kozjak, north of the Periadriatic lineament,deep marine sedimentation started already in Karpatian and continuedthrough the entire Badenian (Fodor et al., 2002). The basins south ofthe Periadriatic and Donat fault zone (Fig. 1) were flooded duringthe lower Badenian transgression (Jelen et al., 2008; Pavšič andHorvat, 2009) while in the area of the southernmost Kbasin – the
MARIBOR
NOVO MESTO
CELJE
PTUJ
LjF
PAL
SF
DoF
ŠF
LF
01 0 20km
SVNHRV
BIH
AUT
MNE
SRB
HUN
ITA
MKD
GRC
CHE
SVKCZE
study area
FRA
UKRDEU
ROU
BGR
Sava FaultSF
Donat Fault ZoneDoF
Ljutomer FaultLjF
Balaton FaultBF
Šoštanj FaultŠF
Periadriatic LineamentPAL
Labot FaultLF
Kungota FaultKF
Raba FaultRF
sections, sampling sites
Quaternary
Tertiary Pannonian basin
Dinarides
Austroalpides
magmatics
Gorenje Vrhpolje
Dolnja Stara VasŠmarjeta
VajndolOrehovica
Fig. 1. Geological setting of the study area showing the position of the Mura Depression and the Krka Basin. Locations of sampling sites and section marked with black circles.
145M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
Krško syncline – marine sedimentation only began in the upperBadenian with water depth not exceeding 100 m (Rižnar et al.,2002; Horvat, 2004; Otoničar and Cimerman, 2006).
The regional compressional contraction that affected the region fromItaly across the Sava folds, southwest Hungary through to Croatia, andstrongly disturbed the constellation of Miocene sediments in this area,was post-Sarmatian (Massari, 1990; Placer, 1999; Tomljenović andCsontos, 2001; Fodor et al., 2002).
The geographical distribution of Paratethyan sediments (Popovet al., 2004) shows convincingly that the Central Paratethys reachedits maximum extent in the Middle Miocene. During the Badenian(Fig. 2), the sea in the wider Pannonian region was at a relativehighstand with water depth reaching the maximum of the neritic(Kováč et al., 1999).
Badenian sediments in the area of present day Slovenia are diversein terms of thickness and facies distribution,while Sarmatian sedimentsare rather similar in various localities, which indicate relatively uniformsedimentary conditions. The diversity and ecology of lower Sarmatiandiatom flora shows a high degree of correlation throughout the entireCentral Paratethys as well (Horvat, 2004). During that time the entireCentral Paratethyswas covered by a shallow sea characterised by rather
uniform sedimentary conditions (with the exception of salinity), whichare reflected in a single type of biofacies that is very widespread (Rögl,1998; Magyar et al., 1999; Ivanov et al., 2002). The Badenian was thelast fully marine period in the life of the Central Paratehys. Salinity be-came region specific with the onset of the Sarmatian (Paramonovaet al., 2004). Bryozoans (Moissette et al., 2006) and other stenohalineorganisms (Kroh, 2007) disappeared from the Central Paratethys andthe first gastropods, endemic to the Central Paratethys, appeared(Harzhauser et al., 2002). This is considered as evidence that by thattime the marine connection between the Central Paratethys and theMediterranean had closed.
The published palaeogeographical interpretations of the MiddleMiocene Central Paratethys assume the connection of this sea withthe Mediterranean and with the Indo-Pacific bioprovince (e.g., Rögl,1998; Meulenkamp et al., 2000). The marine connection between theCentral Paratethys and the Mediterranean closed at some point in theMiddle Miocene; however, the exact time of its closure is still a matterof debate and represents one of the main remaining research problemsconcerning Central Paratethys palaeogeography. According to the pub-lished palaeogeographical interpretations (e.g., Rögl, 1998; Popovet al., 2004), the (hypothetical) Slovenian Corridor, represents one of

NN5
NN4
M I
O C
E N
E
LAN
GH
IAN
SARMATIANs.s.
BA
DE
NIA
N
KONKIAN
KARAGANIAN
TARKHANIAN
Age
(M
a)
Epo
ch
3rd order sequences
CHOKRAKIAN
SE
RR
A V
ALL
IAN
Stage
Med
iter
-ran
ean
Paratethys
LATE
MIDDLE
EARLY
Central Eastern
13
14
15
12
Calcareous nannoplankton zones(Martini, 1971; Fornaciari et al., 1996)and selected events (Abdul Aziz et al.,
2005; Abels et al., 2005; Turco et al., 2002)
SARMATIANs.l.
(VOLHYNIAN)NN6
MNN6a
MNN6b
MNN5a
MNN5b
MNN4b
FCO R. pseudoumbilicus L
LO S. heteromorphus
LCO Cy. floridanus
FCO H. walbersdorfensis
LCO H. waltrans
16MNN4a
al., 2008Rögl et al.,
2007Lena
rtse
ctio
n
TB2. 6
TB2. 5
TB2. 4
TB2. 3
FCO Cal. macityrei
Fig. 2. Stratigraphical correlation ofMiddleMioceneCentral Paratethys andMediterranean stages (Hilgen et al., 2012), standard andMediterranean nannoplankton biozones and 3rd ordereustatic cycles.
146 M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
the most likely seaways connecting the Central Paratethys and theMediterranean.
In the Middle Miocene, our study area (Fig. 1) was situated at theeastern mouth of this hypothetical seaway. The goal of our study wasto investigate if and how the existence of the Slovenian Corridor isreflected in upper Middle Miocene sediments of eastern Slovenia. Westudied calcareous nannoplankton from the Mura Depression and sili-ceous phytoplankton and molluscs from the Krka Basin. The palaeo-geographical position of our research area and the time intervalstudied make our material a particularly relevant source of informationconcerning the existence, position and time of closure of theSlovenian Corridor. In this contribution we present the results of ourpalaeontological investigations and discuss them in the context of rel-evant publications in an attempt to answer this palaeogeographicalquestion.
1.2. Stratigraphic correlation of sea-level fluctuations in the CentralParatethys during the Badenian
From theOligocene until the end of theMiddleMiocene, the time res-olution of isotopic events in the Central Paratethys region is comparablewith global trends (Abreu andHaddad, 1998), which suggests good com-munication between the Central Paratethys and the neighbouring seas.During the Badenian, the Middle Miocene Central Paratethys stage, thisepicontinental sea communicated with the Mediterranean and theEastern Paratethys (and the Indo-Pacific) (Rögl, 1998; Meulenkampand Sissingh, 2003; Goncharova et al., 2004; Báldi, 2006).
The Badenian was correlated with three 3rd order sedimentary cy-cles. Though the correlation of individual sedimentary sequences ofthe Central Paratethys with global sea-level cycles is hampered bystrong regional influences (Fig. 2) it has been largely explained as a re-sult of extensive research (e.g., Kováč et al., 2007; 2008; Rögl et al.,2007). The early Badenian can be correlated with the early Langhianof the Mediterranean and the global sea level cycle TB2.3 above theBur5/Lan1 sequence boundary (Piller et al., 2007). Studying the Viennabasin, Rögl et al. (2007) correlated the Badenian with three global 3rdorder eustatic cycles: TB2.3, TB2.4 and TB2.5, which correspond to theearly, middle and late Badenian. Alternatively, Kováč et al. (2007;2008) described several 3rd and 4th order cycles for the CentralParatethys, which cannot be directly correlated to the 3rd order se-quences of Haq et al. (1988), and subdivided the Badenian into only
the early (corresponding roughly to TB.3 and TB.4 cycles) and the lateBadenian (corresponding to the TB2.5 cycle).
The transgression of the TB2.5 eustatic cycle begins at the NN5/NN6boundary at 13.65 Ma (Abels et al., 2005), shortly after the Langhian/Serravallian boundary dated at 13.82 Ma (Hilgen et al., 2009). This sed-imentary cycle ends at the Ser3 sequence boundary (Hardenbol et al.,1998) which is correlated with the Badenian/Sarmatian boundaryclose to 12.8 Ma by de Leeuw et al. (2012) or more recently at12.624 Ma as suggested by new results from Romania (Bartol et al.,2013).
The close correlation of sea-level fluctuations within the CentralParatethys and global eustatic changes in the late Badenian is the conse-quence of a decrease in the intensity of subsidence (Fodor et al., 2002).The bathymetric changes in the Central Paratethys during that time in-terval were thus controlled primarily by global eustatic fluctuations andnot by regional factors (Horvat, 2004; Báldi, 2006; Kováč et al., 2007).
The Ser2 sequence boundary, correlated with the advanced stage ofthe regression at the base of the late Badenian, was the time of deposi-tion of evaporites in the Carpathian Foredeep and the TransylvanianBasin (e.g., Peryt, 2006; Cendón et al., 2004). In Slovenia, the regressionbetween themiddle and upper Badenian is reflected by a hiatus in somelocalities (Pavšič and Aničić, 1998; Oblak, 2003). Fresh water influencesare noticeable in some other localities in marginal areas of depositionalbasins (Pezelj and Sremac, 2007), while some localities, presumably lo-cated in the deeper parts of sedimentary basins, remained more or lessunaffected by this eustatic sea-level drop (Bartol, 2009).
According to the prevailing view this sea-level lowstand marksthe end of communication between the Central Paratethys and theMediterranean (Rögl, 1998). On the other hand, some authors (e.g.,Kováčová et al., 2009; Studencka et al., 1998; Bartol et al., 2012)argue that an active connection of the two marine realms persisteduntil the end of the TB2.5 eustatic cycle and the end of the Badenian.
1.3. Opposing views on the status of the Slovenian Corridor in the lateBadenian and the palaeontological evidence supporting them
So when did the Central Paratethys and the Mediterranean sepa-rate? Belowwe present the different answers to this question publishedin various scientific publications.
One of the main arguments in favour of the separation of theMediterranean and the Central Paratethys before the late Badenian

147M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
comes from the Vienna Basin. The calcareous nannoplankton and forami-niferal assemblages in upper Badenian sections near Walbersdorf (Rögland Müller, 1976) are distinctly different from the Mediterranean ones:they contain the species Hayella (former genus name Nannocorbis)challengeri and Rhabdosphaera poculi (which do not occur in the con-temporaneous Mediterranean assemblages) and are virtually free ofdiscoasters (that are usually common in the contemporaneous Mediter-ranean assemblages). The Walbersdorf nannoplankton assemblageswere considered as typical of the Central Paratethys, leading the re-searchers (Rögl et al., 1978; Rögl, 1998) to the conclusion that the con-nection between the Central Paratethys and the Mediterranean did notexist during the late Badenian. A similar conclusion was reached byDumitrica, 1978a,b by studying late Badenian silicoflagellate and radio-larian assemblages from Radiolarian Shales overlying the evaporitic de-posits along the Carpathian Foredeep and in the Transylvanian Basin.According to his research early and middle Badenian assemblages showclose affinities with the Mediterranean radiolarians, while late Badenianassemblages are poorer – and therefore more difficult to use to supportany interpretation – but more alike to the Indo-Pacific assemblages thanto the Mediterranean ones. Consequently, the isolation of the CentralParatethys from the Mediterranean following the regression at the endof the middle Badenian was proposed in the palaeographical maps ofSteiniger et al. (1978, p. 113, Abb. 26), Rögl and Steininger (1983),Hámor and Halmai (1988, map 4) and Rögl (1998). According to thispalaeogeographical concept the late Badenian transgression caused thereestablishment of communication between the Central Paratethys, theEastern Paratethys and the Indo-Pacific bioprovince, while the direct ma-rine connection with the Mediterranean never reopened.
On the other hand, several palaeontological studies support the exis-tence of a marine connection between the Central Paratethys and theMediterranean during the late Badenian as well. In contrast to thepalaeogeographical interpretation of Rögl et al. (1978), Nagymarosyand Müller (1988) presume that the faunal connection of the CentralParatethys and the Mediterranean probably persisted until the end ofthe Badenian, but do not mention any concrete findings to supportthis general assumption with.
According to several authors, the Paratethyan mollusc assemblagesshow close affinities to the Mediterranean ones. Studencka et al. (1998),for instance, report of the simultaneous appearance of 26 bivalve speciesin the Atlantic, the Mediterranean, and the Central Paratethys during thelate Badenian transgression which indicate an active communication ofthe Central Paratethys with the Atlantic and the Mediterranean duringthe late Badenian. On the other hand they find that the late Badenianfauna of the Eastern Paratethys ismarked by a significantly lower diversi-ty (316:97 species) than the Central Paratethyan one and that thecontemporaneous bivalve fauna in the Central Paratethys shows noIndo-Pacific affinity, which contradicts the idea of good communicationbetween theCentral and theEasternParatethys. The existence of an activeseaway between theMediterranean and the Central Paratethys in the lateBadenian is also supported by the comparative study of gastropods(Turritellidae family) found in late Badenian sediments in the KrkaBasin (Mikuž, 2009). Slovenian turritelid assemblages of Badenian ageare more similar to the assemblages found within the Central Paratethysand Mediterranean realm (Austria 42% of common taxa, Hungary 29%,Czech Republic 15%, Poland 25%), and Western Paratethys (Italy 21%,Greece 21%, France 17%), than to those of the Eastern Paratethys(Romania 10%, Bulgaria 8%, Ukraine 10%), which contradicts the idea ofan exclusive late Badenian connection of the Central Paratethys with theIndo-Pacific bioprovince through the Eastern Paratethys. Kókay (1985)describes the simultaneous existence of mollusc assemblages reflectingnormal and lowered salinity in the late Badenian sediments studied invarious localities in the vicinity of Budapest. He assumes that during thelate Badenian the Central Paratethys communicated with the Mediterra-nean as well as the Eastern Paratethys.
According to Bitner (1990), the late Badenian brachipod fauna of theCentral Paratethys shows more affinity with the Miocene to recent
Mediterranean brachiopod fauna than with that of the Indo-Pacificbioprovince. The Badenian echinoids of the Central Paratethys are alsoreportedly more similar to the Mediterranean assemblages than to theIndo-Pacific Middle Miocene fauna (Ali, 1983a,b; Ali and Maczynska,1986; Mikuž and Horvat, 2003a,b).
The results of our comparative study of late Badenian calcareousnannoplankton from the Mura Depression with the contemporaneousMediterranean nannoplankton assemblages seem to agree with thesefindings and support the existence of an active connection betweenthe Central Paratethys and the Mediterranean persisting until the endof the Badenian.
1.4. Slovene Corridor or some other seaway?
Another crucial palaeogeographical question is whether the com-munication between the neighbouring realms took place across theSlovenian Corridor or through some other seaway. In his palaeo-geographical reconstruction, Rögl (1998) assumes that the connectionof the Central Paratethys with the Mediterranean crossed the territoryof the present-day Slovenia through the so-called Slovenian (orTranstethyan) Corridor. The location of this marine connection is oftenreproduced in various scientific publications and is also suggested byPopov et al. (2004) in the most detailed palaeogeographical maps of theCentral Paratethys produced to date. However, the Slovenian Corridor,crossing the area of the present-day Slovenia, is not the only possible lo-cation of a marine connection between the Central Paratethys and theMediterranean. Studencka et al. (1998) for example, are of theopinion that in the early Badenian, the Central Paratethys and theMediterranean communicated through the Slovenian Corridor, whilein the late Badenian this seaway had closed and the communication ofthe two realms took place across the Axios (Vardar) Trench. There is nocontinuous stretch of Miocene age sediments between the Paratethyanand Mediterranean realms along the hypothetical Slovenian Corridor orthe Axios (Vardar) Trench, as they were presumably eroded during theLate Miocene and/or later. This means that there is little evidence to sup-port any location of the seaway apart from the geographic constellation ofthe Alps and the Dinarides.
During the study of Middle Miocene gastropods from the Krka Basinan interesting pattern of the palaeogeographical distribution of a gastro-pod species Pereiraea gervaisiwasobserved.Weuse this pattern as a valu-able piece of information supporting the existence of the SlovenianCorridor.
2. Materials and methods
The study of calcareous nannoplankton of NE Slovenia includedanalyses of several sections in the Mura Depression. In this paper wewill concentrate on the Lenart section in Lenart (Fig. 1), a 26 m thicksedimentary sequence composed mostly of marls with intercalatedsands and sandstones in a 2 m thick interval. The sediments studied inthis section were deposited in a deeper part of the Mura Depression atthe eastern mouth of the supposed Slovenian Corridor. Nannoplanktonassemblages were diverse and well preserved, several biostratigraphicevents were observed (the LO of Sphenolithus heteromorphus, the LCOand LO of Cyclicargolithus floridanus the FO and FCO of Reticulofenestrapseudoumbilicus N7 μm, FO of Calcidiscus macityrei), which allowed thesediments to be assigned biostratigraphically to the upper part of NN5and lower part of NN6 standard nannoplankton zones (upper part ofthe middle and upper Badenian). The sands and sandstones werefound to occur at the level of NN5/NN6 boundary, corresponding tothe TB2.4/TB2.5 regression (for a log of the section, position of samplesand full assemblages in each sample see Bartol, 2009). The section wassampled in 10 cm intervals, according to the sedimentation rates esti-mated on the basis of ages of biostratigraphic events in Di Stefanoet al. (2008), each such interval corresponds to approximately 15000 yr.

148 M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
To determine the palaeogeographical implications of the nannoplank-ton assemblages studied in the Lenart section,we compared the results ofour biostratigraphical analyses to several published studies on calcareousnannoplankton in the corresponding time interval. Firstly we consideredthe assemblages from Lenart as model Central Paratethyan assemblages(much like the assemblages from Walbersdorf were used by Rögl(1998) in his palaeogeographical considerations) and compared themto contemporaneous Mediterranean nannoplankton assemblages as de-scribed in available scientific publications. Then we compared the sameassemblages to the ones described in ODP and IODP reports from aroundthe globe to place ourfindings into a global context. Finally,we comparedthe assemblages from Lenart to several other Central Paratethyan assem-blages described from late Badenian sequences to determine the variabil-ity within the Central Paratethys itself.
Molluscs and siliceous phytoplankton was studied in several locali-ties near Šentjernej (Ivanji Dol, Dolnja Stara Vas, Orehovica, GorenjeVrhpolje) in the southern part of Krka Basin and in the Šmarjeta sectionin the northern part of the Krka Basin (Fig. 1). The land in this entirearea is in agricultural use or is covered by constructions, so no propergeological sectionswere accessible. In all sampling sites Pereiraea fossilswere found in dark grey to grey marls known also as the “BadenianTegel” (Keferstein, 1828), which represents a typical Badenian forma-tion of the Central Paratethys. Siliceous phytoplankton analyses concen-trated on beds of carbonate siltstone containing a rich macrofauna ofturritellids and Pereiraeas. Probe samples were prepared from freshrock and frommaterial attached to themacrofauna.Material from Ivanjidol and Šmarjeta contained amoderately well-preserved and abundantdiatomaceous flora, silicoflagellates, ebridians, endoskeletal dinoflagel-lates, sponge spicules were also rather common. The diatoms andsilicoflagellates from the Ivanji Dol locality indicate a Late Badenianage (Horvat, 2004). In terms of macrofauna, these localities were dom-inated by the Pereiraea-Protomamollusc assemblage (Mikuž, 2009). Thematrix attached to the fossil gastropods was also studied for nanno-plankton in an attempt to determine their age as accurately as possible.
3. Calcareous nannoplankton—a comparative study
3.1. Comparing the Lenart section and the Mediterranean
The presence of marker species (Discoaster braarudii, Coronocyclusnitescens, Calcidiscus macintyrei, etc.) and observed biostratigraphicevents (LO of S. heteromorphus, LCO and LO of C. floridanus, FO andFCO of R. pseudoumbilicus N7 μm) allowed the assignment of the Lenart
Fig. 3. Biostratigraphically and palaeoecologically significant calcareous nannoplankton species fro(Discoaster spp.) from interval, enriched with discoasters, at the NN5/NN6 boundary. A: Calcidis(N7 μm) (Gartner) Gartner. C: Sphenolithus heteromorphus Deflandre. D: Cyclicargolithus floridanuBukry. G: Discoaster braarudii Bukry. H: Discoaster exilisMartini & Bramlette. I: Discoaster variabil
section to the upper part of the NN5 and lower part of NN6 biozones ofMartini (1971). The most important biostratigraphic maker species arepresented in Fig. 3; selected events are marked on Fig. 4. For the com-plete assemblage in individual samples and a stratigraphic log of thesection see Bartol (2009).
A great similarity was found between the late Badenian nannoplank-ton assemblages from the Lenart section and contemporaneous (MNN5b,MNN6a, MNN6b) Mediterranean assemblages (Ellis and Lohman, 1978;Fornaciari et al., 1996; Di Stefano et al., 2008). The assemblage composi-tion is virtually identical with all the species listed as elements of anormal assemblage in Fornaciari et al. (1996) also constituting the assem-blages found in Lenart (Bartol, 2009), and the biostratigraphic events ob-served taking place in the samemanner (described in detail in Fornaciariet al., 1996) and the same order of succession (Fig. 4).
In the Lenart section the LO of S. heteromorphus is followed by theLCO of C. floridanus and the FCO of the large form of R. pseudoumbilicus(N7 μm, R. pseudoumbilicus L hereafter). The FO of R. pseudoumbilicus Ltakes place below the LO of S. heteromorphus, the LO of C. floridanus oc-curs slightly above the FCO of R. pseudoumbilicus L. The same events inthe same sequence were observed in the Mediterranean sites DSDPSite 372, Karraba and Serravalle Scrivia (Fornaciari et al., 1996; DiStefano et al., 2008). The occurrence of the first rare and scattered spec-imens of C. macintyrei is also noted in all four Mediterranean sites andthe Lenart section within this short stratigraphic interval (Fig. 4). Toavoid the ambiguity its stratigraphic position, related to different taxo-nomic concepts used by various authors, we only considered specimens≥11 μm in diameter (Fig. 6A). This is in accordancewith the definitionsof Perch-Nielsen (1985), Young (1998), andWise et al. (2002) and alsowith the definition of the species used by Raffi and Flores (1995),Fornaciari et al. (1996), and Turco et al. (2002).
No other biostratigraphical events were recorded in any of the sec-tions within this particular time slice.
The observed similarity between the Lenart section (CentralParatethys) and the Mediterranean sites can be explained either as a re-sult of an active connection between the two realms in the upperBadenian (Serravallian) or as a consequence of a universal global trendof nannoplankton floral turnover, common at least to areas of similarlatitude.
3.2. Comparing the Lenart section and the World Ocean
To resolve this question we compared the sequence of biostrati-graphic events observed in the Lenart section (and the Mediterranean)
mthe Lenart section. Position of events including species A–Dmarked in Fig. 4. Species in E–Icus macintyrei (Bukry & Bramlette) Loeblich & Tappan. B: Reticulofenestra pseudoumbilicuss (Hay et al.) Bukry. E: Discoaster stelullus Gartner, emend. Jiang &Wise. F: Discoaster mooreiisMartini & Bramlette.

A:
B:
C: D:
Fig. 4. Biostratigraphic correlation of biostratigraphic events (calcareous nannoplankton) observed in the Lenart section (Central Paratethys) and three Mediterranean sites (afterFornaciari et al., 1996).
A B
Fig. 5. Biostratigraphic events observed in the Lenart section in global comparison. A: Events observed at the NN5/NN6 transition in the Lenart section and matching events described inODP andDSDP reports. B: Events observed at theNN5/NN6 boundary in variousODP andDSDP sites that donot occur in the Central Paratethys or theMediterranean. Data fromMiller et al.(1985), Parker et al. (1985), Lohman (1986), Olafsson (1989), Fornaciari et al. (1990), Gartner (1992), Jansen et al. (1996), Fornaciari (2000), Kameo and Bralower (2000), Muza (2000),Lirer et al. (2002), Sprovieri et al. (2002), and Zachos et al. (2004).
149M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157

150 M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
to ODP and DSDP reports discussing the same time interval in the Med-iterranean and the World Ocean (Fig. 5).
The results have shown that the LOof S. heteromorphus is indeed glob-ally well correlated, assigned various ages between 13.4 and 13.6 Ma(compare Turco et al., 2002; Sblendorio-Levy and Howe, 1998; Backmanand Raffi, 1997; Raffi and Flores, 1995; Abdul Aziz et al., 2008), but noneof the other events considered.
The FO of R. pseudoumbilicus L (Fig. 5A) is considered to be a latitudi-nally controlled event (e.g., Fornaciari et al., 1993; Raffi et al., 1995), i.e., ittakes place in the tropics before it does in mid and high latitudes. Thetiming of the LO of C. floridanus (Fig. 5A) depends on local ecological con-ditions and has been assigned various ages ranging between 13.19 Ma(Raffi and Flores, 1995) and 12.2–11.2 Ma (Wei and Wise, 1992).
The FO of C. macintyrei (Fig. 5A) is considered to be a poor strati-graphic marker (Fornaciari et al., 1993), dated between 13.34 Ma inthe tropical Pacific (Raffi and Flores, 1995) and 12.41 Ma in theMediter-ranean (Hilgen et al., 2003). Though the Mediterranean FO datum isyounger than the interval considered in this study, it is very importantto note, that the first scattered specimens of C. macintyrei in DSDP Site372, Serravale Scrivia and Karraba sections as described by Fornaciariet al. (1996)were noticedwell below the pointwhere the FO of the spe-cies was pinpointed (marked in Fig. 4). The pattern of discontinuouspresence of very rare specimens is the same as the situation observedin the Lenart section, so this too can be considered as a similarity be-tween the Mediterranean and the NW Central Paratethys.
The events recorded in the investigated time slice in the Mediterra-nean and the Paratethyan Lenart sectionwere not observed in the samesequence anywhere else. Moreover, several other events are reportedfrom the World Ocean during the studied time intervals that do nottake place in the Mediterranean or the Central Paratethys (Fig. 5B).
3.3. Comparing the Lenart section and other localities within the CP
In contrast to the very similar composition of calcareous nannofossilassemblages and succession of biostratigraphic events in differentMed-iterranean sites during the lower Serravallian, the contemporaneousupper Badenian assemblages from different localities within the CentralParatethys exhibit considerable variation. Due to different taxonomicand (bio)stratigraphic concepts, language barriers and limited availabil-ity of publication resulting in parallel studies, and pronounced localcharacter of assemblages, studies of Paratethyan nannofossils are ratherdifficult to compare. For this reason we picked out two prominent as-semblage characteristics, whichwewere able to track accurately in var-ious publications andwhich – according to Rögl andMüller (1976) – aretypical of the Mediterranean or the Central Paratethys (Fig. 6):
- The presence of R. poculi and H. (old generic name Nannocorbis)challengeri, typical of Paratethyan Miocene nannofossil assemblages(e. g., Ćorić and Hohenegger, 2008) and not known from the Medi-terranean during that time,
- The presence of discoasters, supposedly absent from late BadenianParatethyan assemblages but usually quite common in contempora-neous Mediterranean assemblages.
An enrichment with several species of discoasters (Fig. 3E–I) at theboundary of NN5/NN6 is one of the most prominent characteristics ofnannoplankton assemblages studied in samples from the Lenart section(Bartol and Pavšič, 2005). None of the samples from the Lenart sectioncontained R. poculi orH. challengeri, in fact, the two speciesweremissingin all Badenian nannofossil assemblages (from 17 sections) from NESlovenia (Bartol, 2009).
The assemblages and nannoplankton events studied in the Lenartsection are most similar to those described in Serbia by Mihajlovićand Knežević (1989), with the LO (LCO in the Lenart section) ofS. heteromorphus followed by an enrichment with discoasters, whichin turn are replaced by assemblages rich in sphenoliths. The same lo-cality is also marked by the occurrence of very rare specimens of
C. macintyrei in the bottompart of NN6 – as is the case in Lenart sectionand all Mediterranean sites considered. An enrichment of the top oflate Badenian Paratethyan assemblages with discoasters is alsoknown from W Slovakia (Ozdínová, 2008); discoasters occur in lateBadenian assemblages in the Transylvanian Basin (Chira and Malacu,2008).
The late Badenian nannoflora fromWalbersdorf in E Austria (Rögl andMüller, 1976), which represents one of the key arguments supporting thelack of communication between the Central Paratethys and the Mediter-ranean, is notably different than the contemporaneous assemblagesfound in the Lenart section. It contains R. poculi and H. challengeri and isvirtually free of discoasters and sphenoliths. Rare and scattered speci-mens of R. poculi and H. challengeri were detected in E Slovakian lateBadenian deposits (Ozdínová, personal communication) and in NCroatia (Galović, 2009). R. poculi is present in Badenian deposits of theTransylvanian Basin (Chira and Malacu, 2008) and in some other locali-ties (see Fig. 6).
4. Palaeogeographic significance of P. gervaisi
4.1. Biostratigraphic dating of P. gervaisi
The prominent gastropod species P. gervaisi (Fig. 7) is known in theMediterranean Tethys from the middle Burdigalian to the Tortonian. Inthe Central Paratethys the species is only mentioned in sediments ofBadenian age. The significant environmental changes in the CentralParatethys area at the end of the Badenian apparently caused the localextinction of the species – it is not known from the Sarmatian.
In the Central Paratethys the species is only known from a handful oflocalities in Croatia (Gorjanović-Kramberger, 1896), Hungary (Vadász,1960; Kókay, 1966; Strausz, 1966) and Slovenia (Mikuž, 1999, 2009).
In the samples from the P. gervaisii type locality of Ivanji Dol nearŠentjernej 84 forms of diatoms were described (Horvat, 2003). Accord-ing to the biostratigraphycally significant species (Fig. 8): Actinocyclusingens, A. tsugaruensis, Chasea tortonica, Coscinodiscus vetustissimus,Denticulopsis hustedti, D. lauta, Dimerogramma tortonica, Liradiscusplicatulus, Rhaphoneis gratiosa and Stephanopyxis multispinosa the de-scribed assemblage could be compared with the other Badenian assem-blages in Central Paratethys (Řeháková, 1978; Hajós, 1986). The FOD ofD. hustedti is dated at 14.2 Ma (Barron, 1985; Bustillo and Lopez García,1997; Censarek and Gersonde, 2002), so the assemblage could beassigned to the interval between the end of middle Badenian (upperpart of NN5 biozone) and lower Sarmatian.
The diatom assemblages are dominated by the resting spores ofChaetoceros spp. and Thalassionema nitzschioideswhich usually indicatestrong coastal upwelling conditions and transgression. The variationsobserved in the diatom assemblages are consistent with nutrient-enriched waters, which were transported laterally from the upwellingcentre to the west of the study area, presumably from the zone ofmixing of warm proto-Mediterranean and cool Atlantic water masses.It seems that the origin of the transgressive polyhalobous diatomite fa-cies was associated with coastal upwelling as suggested by a eustaticmodel of opal-rich facies formation and anti-estuarine circulation pat-tern of water masses (Horvat, 2012).
When correlating our biostratigraphic results with the sequencestratigraphy data for the Central Paratethys (Fig. 2), the transgressivecharacter of the diatom assemblage could support a correlation withyounger Badenian transgression (sensu Kováč et al., 2008) or tentative-ly support a correlation with the transgression of the late BadenianTB2.5 cycle (Rögl et al., 2007).
To refine the biostratigraphic dating of P. gervaisi specimens, weperformed additional micropalaeontological studies of calcareous nan-noplankton of the marl matrix filling the specimens found in DolnjaStara Vas, Orehovica and Gorenje Vrhpolje. The nannoplankton assem-blages foundwere rather poor andmoderately preserved, but neverthe-less provided some biostratigraphical information. The biostratigraphic

A
B
Fig. 6. Palaeogeographic consideration of Late Badenian calcareous nannoplankton taxa in the Central Paratethys. A: Presence/absence of Discoaster spp. B: Presence/absence ofH. challenger and/or R. poculi. Data from Bóna and Kernerné Sümegi (1966), Chira et al. (2000), Brânzilă and Chira (2005), and Mărunţeanu (2006).
20 mm
Fig. 7. Pereiraea gervaisi Vézian, specimen from the Krka Basin.
151M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
age could be assigned most convincingly to the specimen found inOrehovica. The matrix from a specimen from this site contained 11nannoplankton species including Helicosphaera waltrans andH. walbersdorfensis (Fig. 9). The former has a short stratigraphicrange limited to the middle part of NN5 (Theodoridis, 1984), whilethe latter has its FCO in the middle of NN5 (see Fig. 2 for their abso-lute ages). The assemblage from Orehovica is also interesting fromthe palaeoecological perspective – it is very rich in the speciesBraarudosphaera bigelowii. Enrichments with this species are usuallyassociated with lowered salinity (Bartol et al., 2008 and the refer-ences therein). The B. bigelowi pentaliths found in the sample fromOrehovica were rather variable in size (Fig. 9), but all belonged to asingle morphotype with sharp corners and trapezoidal segments.
The assemblage studied in the matrix attached to the macrofossilfrom Dolnja Stara Vas was of little use biostratigraphically. However, asample collected a few metres below the horizon with the macrofossils
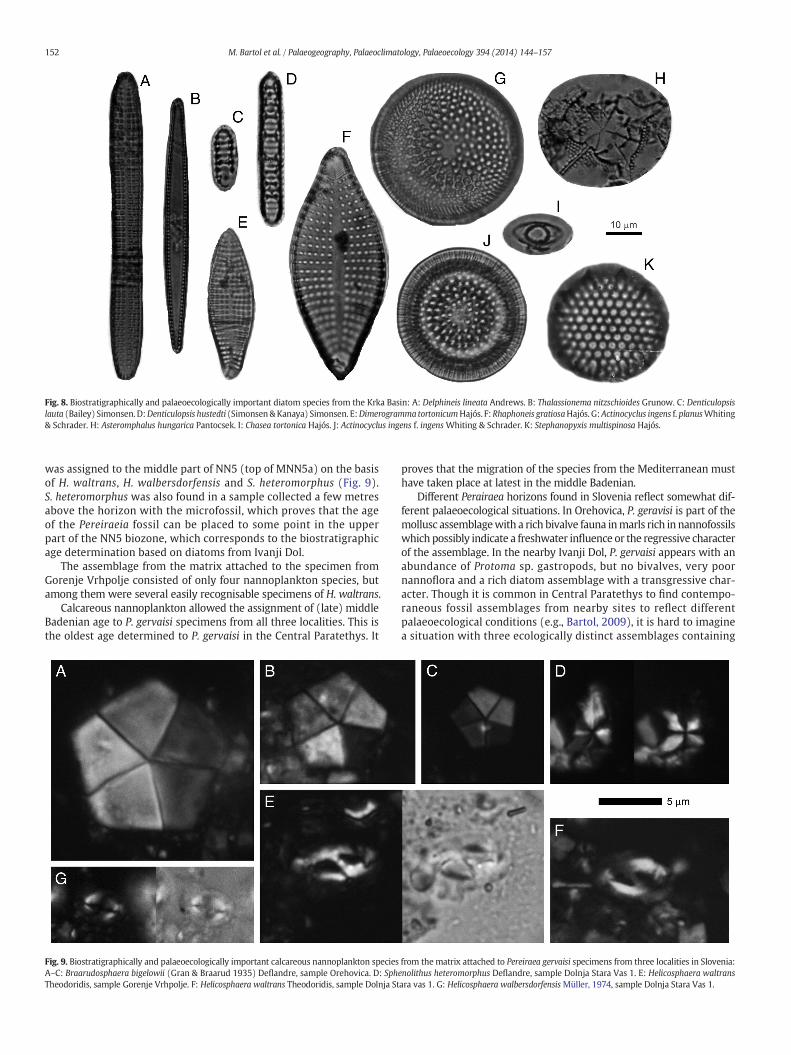
Fig. 8. Biostratigraphically and palaeoecologically important diatom species from the Krka Basin: A: Delphineis lineata Andrews. B: Thalassionema nitzschioides Grunow. C: Denticulopsislauta (Bailey) Simonsen. D:Denticulopsis hustedti (Simonsen&Kanaya) Simonsen. E:Dimerogramma tortonicumHajós. F:Rhaphoneis gratiosaHajós. G:Actinocyclus ingens f. planusWhiting& Schrader. H: Asteromphalus hungarica Pantocsek. I: Chasea tortonica Hajós. J: Actinocyclus ingens f. ingensWhiting & Schrader. K: Stephanopyxis multispinosa Hajós.
152 M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
was assigned to the middle part of NN5 (top of MNN5a) on the basisof H. waltrans, H. walbersdorfensis and S. heteromorphus (Fig. 9).S. heteromorphus was also found in a sample collected a few metresabove the horizon with the microfossil, which proves that the ageof the Pereiraeia fossil can be placed to some point in the upperpart of the NN5 biozone, which corresponds to the biostratigraphicage determination based on diatoms from Ivanji Dol.
The assemblage from the matrix attached to the specimen fromGorenje Vrhpolje consisted of only four nannoplankton species, butamong them were several easily recognisable specimens of H. waltrans.
Calcareous nannoplankton allowed the assignment of (late) middleBadenian age to P. gervaisi specimens from all three localities. This isthe oldest age determined to P. gervaisi in the Central Paratethys. It
Fig. 9. Biostratigraphically and palaeoecologically important calcareous nannoplankton speciesA–C: Braarudosphaera bigelowii (Gran & Braarud 1935) Deflandre, sample Orehovica. D: SphTheodoridis, sample Gorenje Vrhpolje. F: Helicosphaera waltrans Theodoridis, sample Dolnja St
proves that the migration of the species from the Mediterranean musthave taken place at latest in the middle Badenian.
Different Perairaea horizons found in Slovenia reflect somewhat dif-ferent palaeoecological situations. In Orehovica, P. geravisi is part of themollusc assemblagewith a rich bivalve fauna inmarls rich in nannofossilswhich possibly indicate a freshwater influence or the regressive characterof the assemblage. In the nearby Ivanji Dol, P. gervaisi appears with anabundance of Protoma sp. gastropods, but no bivalves, very poornannoflora and a rich diatom assemblage with a transgressive char-acter. Though it is common in Central Paratethys to find contempo-raneous fossil assemblages from nearby sites to reflect differentpalaeoecological conditions (e.g., Bartol, 2009), it is hard to imaginea situation with three ecologically distinct assemblages containing
from thematrix attached to Pereiraea gervaisi specimens from three localities in Slovenia:enolithus heteromorphus Deflandre, sample Dolnja Stara Vas 1. E: Helicosphaera waltransara vas 1. G: Helicosphaera walbersdorfensisMüller, 1974, sample Dolnja Stara Vas 1.

153M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
P. gervaisi lying in the same sedimentary basin less than 1 km fromone other. Therefore it seems more likely that the gastropods appearin (slightly) different stratigraphical horizons within the time inter-val between the upper part of the middle Badenian and the lateBadenian.
4.2. Palaeoecology of P. gervaisi
P. gervaisi is an extinct gastropod species of the Strombidae family.All extant representatives of this family are herbivores and occasionaldetritivores and live in tropical or subtropicalmarine environments. Ac-cording to Abbott andDance (1991) they inhabitwarm shallow seas liv-ing in water depths up to 45 m. In Hungary, P. gervaisi have been foundin marine sediments of Badenian age in the corbulid horizon – alumachelle rich in bivalves of the species Corbula basteroti, Saxolucina(Plastomiltha) suessi and Acanthocardia paucicostata (Kókay, 1966). Ex-tant species of the Corbula genus live in water depths from 22 to55 m, those of the Lucina genus inhabit sea floor atwater depths around20 m,while most acanthocardians live in water depths around 30 m. InOrehovica Corbula (Carycorbula) basteroti and Anadara diluvii clams arevery common in the P. gervaisi horizon. The extant species of the Andaragenus live attached to plants or other substrates in warm shallow seas.We believe that the above allows the consideration of P. gervaisi as ashallow water marine species.
In some localities in Slovenia (Ivanji Dol, Šmarjeta) P. gervaisi speci-mens are very abundant and dominate assemblages in association withProtoma sp. (Mikuž, 2009) and very few other mollusc taxa. This indi-cates the existence of shallow water environments in the southernpart of our study area (Fig. 1).
P. gervaisi fossils were found in several sites in the Mediterranean,the Western Paratethys and the Central Paratethys. The geographicaldistribution of the species in the Central Paratethys is very interesting(Fig. 10). Despite comprehensive publications on Paratethyan molluscfauna diversity and geographical distribution, P. gervaisi was only re-ported from the northern margin of the Central Paratethys. The south-ernmost localities in the Central Paratethys are in northern Croatiaand in the Krka Basin (Mikuž, 1999 and references therein) in immedi-ate proximity to the hypothesised position of the Slovenian Corridor.
Fig. 10. Palaeogeographical map of the Central Paratethys and the neighbouring seas in the Lategervaisi (simplified after Popov et al., 2004). Data fromVézian (1856), Crosse (1867), Kinkelin (1et al. (1954), Zbyszewski (1957), Crosse (1968), and Harzhauser et al. (2003).
5. Discussion
5.1. A reconsideration of micropalaeontological arguments supporting theseparation of the Central Paratethys and the Mediterranean in the lateBadenian
The presence of R. poculi and H. challengeri in the assemblages fromthe Walbersdorf locality in the Vienna Basin (Rögl and Müller, 1976) isconsidered as a typical characteristic of Paratethyan nanofossil assem-blages. This information was particularly influential in the palaeo-geographic interpretations of Steiniger et al. (1978) and Rögl et al.(1978), which have been repeated and cited ever since their publica-tion. The assemblage found in the Lenart locality (Bartol, 2009), howev-er, was very different from the one found in Walbersdorf (Rögl andMüller, 1976). A comparison of contemporaneous nannoplankton as-semblages studied in different sites within the Central Paratethys re-vealed a great deal of variability (see chapter 3.3). Our analyses show,that – according to our criteria – an individual late Badenian assemblageismore similar to the one described from Lenart (and contemporaneousMediterranean assemblages) or to the one from Walbersdorf (and pre-sumably Eastern Paratethyan assemblages) or contains certain charac-teristics of both. This means that it is difficult to consider any givenCentral Paratethyan nannoplankton assemblage to be typical of the en-tire Central Paratethys.
In the samemanner we can challenge another palaeontological ar-gument in favour of separation of the Central Paratethys and theMed-iterranean in the late Badenian, which comes from the late Badenianradiolarian flora (Dumitrica, 1978a,b). Firstly, the palaeogeographicalinterpretation of Dumitrica (1978a,b) is based on relatively poorsilicoflagellate and radiolarian assemblages. Secondly, the materialfor these studies only comes from sediments deposited in a part ofpresent-day Romania, which are considered as representative of thelate Badenian flora of the entire Central Paratethys. We find this as-sumption to be rather questionable.
The comparison of late Badenian biostratigraphic events as observedin the calcareous nannoplankton assemblages from the Lenart sectionand contemporaneous assemblages from the Mediterranean revealeda very close resemblance between them, both in terms of assemblagecomposition and the sequence of observed biostratigraphic events.
Badenianwith the palaeogeographical distribution the gastropod species Pereiraea gervaisi892), Hilber (1893), Hörnes (1895), Brives (1897), Sacco (1905), Šuklje (1929), Dartevelle

154 M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
The comparison with the World Ocean showed that only the LO ofS. heteromorphus is a globallywell correlated event. All other nannoplank-ton events considered are reportedly diachronous or missing in othersites considered in the study. In our opinion the observed parallelism be-tween our reference Central Paratethyan locality and the Mediterraneansites represents a strong argument in favour of a marine connection be-tween the two marine realms during the late Badenian.
5.2. Palaeogeographical significance of P. gervaisi
The micropalaeontological analyses of P. gervaisi macrofossils inSlovenia revealed their middle (possibly late) Badenian age. The migra-tion of the species from the Mediterranean had already taken place bythe middle Badenian, so dating P. gervaisi fossils does not tell muchabout the connection of the Mediterranean and the Central Paratethysin the late Badenian. However, the palaeogeographic distribution ofP. gervaisi in the Central Paratethys is most interesting as an indicatorof the exact position of the seaway connecting the Central Paratethysand the Mediterranean. P. gervaisi fossils were only found in Slovenia,Croatia and Hungary (Fig. 10). This represents a convincing argumentin favour of the thesis that the speciesmigrated from theMediterraneanto the Central Paratethys through the Slovenian Corridor and not acrosssome other marine connection (e. g., through the hypothetical Axios(Vardar) Trench, the existence of which is postulated by some authors(e.g., Studencka et al., 1998; Hámor, 2002).
5.3. Palaeoecological complexity of the Central Paratethys and itswater circulation
Considering the palaeoecological preferences of P. gervaisi, the pres-ence of large populations of this gastropod species found in several sitesin present day Slovenia proves the existence of shallow water environ-ments in the southern margin of the area of the hypothesised SlovenianCorridor in the upper part of the middle Badenian. Diverse nannoplank-ton assemblages found in sediments of the same age in the nearbyMura Depression (Bartol, 2009), on the other hand, indicate the simulta-neous existence of deeper marine environments in the NE part of thehypothesised Slovenian Corridor. The coexistence of shallow water anddeepermarine environments is also evident fromvariousmollusc assem-blages studied in different parts of the Pannonian Basin (Kókay, 1966),but here the difference is attributed to varying salinity.
The incredible range of palaeoecological complexity characterisingmarine environments in the middle/late Badenian Central Paratethysis also reflected in contemporaneous nannoplankton assemblagesexisting in close proximity, e. g. in the SE Karpathians (Bartol et al.,2013; unpublished data) or in the adjacent Vienna and Danube Basins(Rögl and Müller, 1976; Halasova, personal communication; Ozdínová,2008; Bartol, 2009). This situation nicely complements the findings ofKováčová et al. (2009), who recognise various degrees of communica-tion with the Mediterranean in the northern and southern part of theVienna Basin.
Analysing the microfossil assemblages from Walbersdorf, we findthat some interbeds in the Walbersdorf sequence are enriched withB. bigelowii andMicrantolithus vesper as well as with benthic foraminif-era of the Bulimina and Bolivina genera (Rögl andMüller, 1976). The au-thors interpret this pattern as a consequence of changing circulation.Alternatively, these assemblage shifts are also consistent with episodesof lowered salinity. This alternative interpretation corresponds well tothe hypothesis proposed by Kókay (1985). His circulation model is atwo-layered circulation system within the Central Paratethys withbrackish waters flowing in from the Eastern Paratethys and the normalsalinity currentflowing in from theMediterranean. A similar– estuarine– circulation model for the late Badenian was also proposed by Báldi(2006). Kókay (1985) traces the effects of reduced salinity as far eastas the Tapolca Basin (north of Lake Balaton), where the “eastern” faunalelements requiring lowered salinity disappear from the assemblages.
Interestingly enough, Tapolca lies approximately half way betweenthe Lenart section and the Walbersdorf locality. While the nannofossilassemblages in the Lenart section exhibit strong Mediterranean affini-ties and are consistent with normal salinity, the nannofossil in theWalbersdorf section are markedly distinct from the Mediterraneanones and indicate an environment of changing salinity. This situationcould be explained with a slight modification of Kókay's rough palaeo-current scheme (Fig. 6A) according to which the northern branch ofthe normal salinity current flowing in from the Mediterranean did notcontinue towards the Vienna Basin, as it was blocked by the BurgenlandSwell. This ridge of metamorphic rocks represents a boundary betweentheMura Depression and the Styrian Basin andwhich existed already atthe birth of the basins and it is still more elevated than the basin itself(Márton et al., 2002). Instead, the Vienna Basin was under the influenceof the brackish current from the Eastern Paratethys, which continuedfurther north from the vicinity of the Tapolca Basin. This complicatedcirculation scheme can explain the mixed Mediterranean and EasternParatethys affinities of the nannoplankton assemblages from theDanube Basin (Ozdínová, 2008) and the Transylvanian Basin (Chiraand Vulc, 2003) as well as strong similarity between the nannofossilcontent of upper Badenian deposits in Slovenia and Serbia (compareBartol, 2009 and Mihajlović and Knežević, 1989).
A certain type of nannoplankton assemblage (in terms of similarityto the Mediterranean or the Eastern Paratethys, se chapter 3.3) is obvi-ously not limited to a particular part of the Central Paratethys as assem-blages with Mediterranean and Eastern Paratethyan characteristicshave been reported from virtually this entire marine realm (Fig. 6).This suggests that themixing ofMediterranean and Eastern Paratethyaninfluences was characteristic of the entire Central Paratethys during thelate Badenian times.
Our data shows that the Slovenian Corridor remained open andallowed a certain degree of communication between the CentralParatethys and the Mediterranean until the end of the Badenian.The complex palaeoecological situation characterising the CentralParatethys in the late Badenian and the contradicting results of var-ious palaeontological studies suggest that the Mediterranean influ-ences did not spread throughout the entire Central Paratethys.Apart from the complex situation in terms of nannoplankton assem-blage composition, this is also indicated by the fact that the easilyrecognisable P. gervaisi has only been found near the NW margin ofthe Central Paratethys, despite the fact that this landlocked systemof interconnected basins with a complex shoreline and numerous is-land archipelagos surely provided an abundance of suitable marinehabitats for the thriving of the species in terms of water depth andfeeding habitats. Apparently, various parts of the Central Paratethys ex-perienced various degrees of isolation from theMediterranean after theend of themiddle Badenian. Themosaic distribution of various fossil as-semblages reported from this entiremarine realmprobably results fromthe mixing of Eastern Paratethyan andMediterranean influences and isassociated with the complex configuration of water and land in theCentral Paratethys.
6. Conclusions
Palaeontological studies of calcareous nannoplankton, diatoms andmolluscs in Badenian sedimentary successions in eastern Slovenia bringnew insights into the status of the Slovenian Corridor – a hypothesisedseaway linking the Central Paratethys and the Mediterranean – in thelate Badenian (early Serravalian).
The palaeogeographical distribution of P. gervaisi in the CentralParatethys is limited to its NW part, to the immediate vicinity of thehypothesised Slovenian Corridor. This represents a strong argument in fa-vour of the thesis that the central Paratethys and theMediterranean com-municated through this seaway and not some other marine connection.
Late Badenian nannoplankton assemblages studied in NE Slovenia(Central Paratethys) are very similar to the contemporaneous

155M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
Mediterranean ones. A comparative study of nannoplanktonbiostratigraphical events observed in this time interval throughoutthe World Ocean suggests that this similarity reflects communica-tion between the Mediterranean and Paratethyan populationsrather than a universal trend of nannoplankton evolution.
The palaeogeographical interpretation according to which theSlovenian Corridor was closed in the late Badenian is based largely onpalaeontological evidence froma single site (nannoplankton) or a hand-ful of sites in a relatively small area in the E Central Paratethys (radiolar-ians, silicoflagellates). Our findings call this interpretation into question.Namely, late Badenian assemblages from individual sites were found tobe surprisingly variable with respect to “Mediterranean” and “EasternParatethyan” affinities throughout virtually this entire marine realm.Thuswe find that it is difficult to consider any given Central Paratethyanassemblage to be typical of the Central Paratethys as a whole.
The mosaic distribution of two distinct types of nannofossil assem-blages reported from the Central Paratethys supports the idea of mixingcurrentsflowing into theCentral Paratethys from the Eastern Paratethysand the Mediterranean. In our opinion, the character of the assemblagein a particular Central Paratethyan site is determined by its position inthe complex configuration of water and land characterising this marinerealm.
Acknowledgement
We would like to thank Marijan Grm for producing Fig. 1 presentedin this paper. We are grateful to the three anonymous reviewers and toJorijntje Henderiks for their suggestions that enabled us to improve thequality of themanuscript.Wewould also like to thank GrahamBudd forreading the manuscript and helping us to improve the level of English.
References
Abbott, R.T., Dance, S.P., 1991. Compendium of Seashells: A Colour Guide to More Than4200 of the World's Marine Shells, 9. Charles Letts & Ltd., London (411 pp.).
Abdul Aziz, H., et al., 2008. Integrated stratigraphy and 40Ar/39Ar chronology of early Mid-dle Miocene sediments from DSDP Leg 42A, Site 372 (Western Mediterranean).Palaeogeogr. Palaeoclimatol. Palaeoecol. 257, 123–138.
Abels, H.A., et al., 2005. Long-period orbital control on middle Miocene global cooling: in-tegrated stratigraphy and astronomical tuning of the Blue Clay Formation on Malta.Paleoceanography 20, 1–17.
Abreu, V.S., Haddad, G.A., 1998. Glacioeustatic fluctuations: the mechanism linking stableisotope events and sequence stratigraphy from the Early Oligocene to Middle Mio-cene. In: Graciansky, P.-C.-d., Hardendol, J., Jacquin, T., Vail, P.R. (Eds.), Mesozoicand Cenozoic Sequence Chronostratigraphic Framework of European Basins. SpecialPublication. SEPM, Tulsa, pp. 245–259.
Ali, M.S.M., 1983a. Tertiary echinoids and the time of collision between Africa and Eurasia.Neues Jb. Geol. Paläontol. Monat. 4, 213–227.
Ali, M.S.M., 1983b. The paleogeographic distribution of the Clypeaster (Echinoidea) dur-ing Cenozoic Era. Neues Jb. Geol. Paläontol. Monat. 8, 449–464.
Ali, M.S.M., Maczynska, S., 1986. Middle Miocene echinoids in the Tethys (Egypt) and theParatethys (Poland). Neues Jb. Geol. Paläontol. Monat. 10, 577–586.
Backman, J., Raffi, I., 1997. Calibration of Miocene nannofossil events to orbitally tunedcyclostratigraphies from Ceara Rise. In: Shackleton, N.J., Curry, W.B. (Eds.), Proceed-ings of the Ocean Drilling Program. Scientific Results, College Station, pp. 83–99.
Báldi, K., 2006. Paleoceanography and climate of the Badenian (Middle Miocene, 16.4–13.0 Ma) in the Central Paratethys based on foraminifera and stable isotope (d18Oand d13C) evidence. Int. J. Earth Sci. 95, 119–142.
Barron, J.A., 1985. Miocene to Holocene planktic diatoms. In: Bolli, H.M., Saunders, J.B.,Perch-Nielsen, K. (Eds.), Plankton Stratigraphy: Planktic Foraminifera, CalcareousNannofossils and Calpionellids. Cambridge University Press, pp. 763–809.
Bartol, M., 2009. Middle Miocene calcareous Nannoplankton of NE Slovenia (westernCentral Paratethys). ZRC Publishing, Ljubljana (136 pp.).
Bartol, M., Pavšič, J., 2005. Badenian discoasters from the section in Lenart (NortheastSlovenia, Central Paratethys). Geophys. J. R. Astron. Soc. 48, 211–223.
Bartol, M., Pavšič, J., Dobnikar, M., Bernasconi, S.M., 2008. Unusual Braarudosphaerabigelowii and Micrantholithus vesper enrichment in the Early Miocene sedimentsfrom the Slovenian Corridor, a seaway linking the Central Paratethys and the Medi-terranean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 267 (1–2), 77–88.
Bartol, M., Mikuž, V., Horvat, A., 2012. Paleontological evidence of communication be-tween the Central Paratethys and the Mediterranean during the Late Badenian/Serravallian. In: Stoica, M., Melinte-Dobrinescu, M.C., Palcu, D. (Eds.), RCMNS InterimColloquium, Bucharest.
Bartol, M., Tulbure, M., Palcu, D., Krijgsman, W., 2013. The Badenian–Sarmatian boundaryin the Getic Depression: preliminary results of integrating biostratigraphy and
magnetostratigraphy. In: Bybell, L.M. (Ed.), INA14. International Nannoplankton As-sociation, Reston, Virginia, p. 21.
Bitner, M.A., 1990. Middle Miocene (Badenian) brachiopods from the Roztocze Hills,south-eastern Poland. Acta Geol. Pol. 40 (3–4), 129–157.
Bóna, J., Kernerné Sümegi, K., 1966. Mikropaleontologiae vizsgalotok a tekeres 1. sz.Foldtani alapfuras Miocen kepzodmenyein. Magyar Allami Foldtani Intezet. EviJelentese 1964, 113–137.
Brânzilă, M., Chira, C., 2005. Microfossils assemblages from the Badenian/Sarmatianboundary in boreholes from the Moldavian Platform. Acta Paleontologica Romaniae5, 17–26.
Brives, A., 1897. Fossiles Miocenes. Premiere partie. Matériaux pour la Carte Géologiquede L'Algérie. Paléontologie. Monographies, 3. Service de la carte géologique, Alger(1-38 pp.).
Bustillo, M.A., Lopez García, M.J., 1997. Age, distribution and composition of Miocene dia-tom bearing sediments in the Guadalquivir Basin, Spain. Geobios 30, 335–350.
Cendón, D.I., Peryt, T.M., Ayora, C., Pueyo, J.J., Taberner, C., 2004. The importance of recyclingprocesses in the Middle Miocene Badenian evaporite basin (Carpathian foredeep):palaeoenvironmental implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 212,141–158.
Censarek, B., Gersonde, R., 2002. Miocene diatom biostratigraphy at ODP Sites 689, 690,1088, 1092 (Atlantic sector of the Southern Ocean). Mar. Micropaleontol. 45,309–356.
Chira, C., Malacu, A., 2008. Biodiversity and paleoecology of the Miocene calcareous nan-noplankton from Sibiu area (Transylvania, Romania). Acta Paleontologica Romaniae6, 17–28.
Chira, C., Vulc, A.N.A.M., 2003. The Miocene calcareous nannofossils from Bistriţa area(Transylvania, Romania). Studia Universitatis Babeş-Bolyai Geologia 48, 67–80.
Chira, C., Szabo, E., Ianoliu, C., 2000. Badenian (Middle Miocene) calcareous nannofossilsfrom Paglisa (Cluj district): biostratigraphical importance. Studia UniversitatisBabeş-Bolyai Geologia 45, 21–32.
Ćorić, S., Hohenegger, J., 2008. Quantitative analyses of calcareous nannoplankton assem-blages from the Baden–Sooss section (Middle Miocene of Vienna Basin, Austria).Geol. Carpath. 59, 447–460.
Crosse, 1867. Gasteropodes dos depositos terciarios, etc. (Gasteropodes dos dépôts tertiaresdu Portugal), par Pereira da Costa, F. A. Journal de Conchyliologie 3, 464–465.
Crosse, H., 1968. Description du nouveau genre Pereiraea. Journal de Conchyliologie 3 (T. 8),191–194.
Dartevelle, E., Roger, J., Daci, A., 1954. Contribution a la connaissance de la faune duMiocen de l'Angola. Communicações da Commissão do Serviço Geológico dePortugal, Lisboa (90 pp.).
de Leeuw, A., et al., 2012. Paleomagnetic and chronostratigraphic constraints on the Mid-dle to Late Miocene evolution of the Transylvanian Basin (Romania): implications forCentral Paratethys stratigraphy and emplacement of the Tisza–Dacia plate. Glob.Planet. Chang. http://dx.doi.org/10.1016/j.gloplacha.2012.04.008.
Di Stefano, A., et al., 2008. Calcareous plankton high resolution biomagnetostratigraphyfor the Langhian of the Mediterranean area. Riv. Ital. Paleontol. Stratigr. 114 (1),51–76.
Dumitrica, P., 1978a. Badenian Radiolaria from Central Paratethys. In: Brestenska, E. (Ed.),Chronostratigraphie and Neostratotypen. Miozän der Zentralen Paratethys. MiozänM4-Badenien. VEDA, Bratislava, pp. 231–261.
Dumitrica, P., 1978b. Badenian silicoflagellates from Central Paratethys. Chronostratigraphieand Neostratotypen. Miozän der Zentralen Paratethys. Miozän M4-Badenien, 6,pp. 207–230.
Ellis, C.H., Lohman, W.H., 1978. Neogene calcareous nannoplankton biostratigraphy inEastern Mediterranean Deep-Sea sediments (DSDP Leg 42A, Sites 375 and 376).Mar. Micropaleontol. 4, 61–84.
Fodor, L., et al., 2002. Miocene to Quaternary deformation, stratigraphy and paleogeogra-phy in Northeastern Slovenia and Southwestern Hungary. Geophys. J. R. Astron. Soc.45 (1), 103–114.
Fornaciari, E., 2000. Calcareous nannofossil biostratigraphy of the California margin. In:Lyle, M., Koizumi, I. (Eds.), Proceedings of the Ocean Drilling Program. Scientific re-sults, College Station, pp. 3–40.
Fornaciari, E., et al., 1990. Quantitative distribution patterns of Oligocene and Miocenecalcareous nannofossils from the western equatorial Indian Ocean. In: Duncan, R.A.,Backman, J., Peterson, L.C. (Eds.), Proceedings of the Ocean Drilling Program. ScientificResults, pp. 237–254.
Fornaciari, E., Backman, J., Rio, D., 1993. Quantative distribution patterns of selected Lowerto Middle Miocene calcareous nannofossils from the Ontong Java Plateau. In: Berger,W.H., Kroenke, L.W. (Eds.), Proceedings of the Ocean Drilling Program. Scientific Re-sults, College Station, pp. 245–256.
Fornaciari, E., Di Stefano, A., Rio, D., Negri, A., 1996. Middle Miocene quantitative calcare-ous nannofossil biostratigraphy in the Mediterranean region. Micropaleontology 42,37–63.
Galović, I., 2009. Middle Miocene (Sarmatian) Calcareous Nannoplankton, Silicoflagellates,and Diatoms of the Southwestern Part of the Paratethys. University of Ljubljana,Ljubljana (211 pp.).
Gartner, S., 1992. Miocene nannofossil chronology in the North Atlantic, DSDP Site 608.Mar. Micropaleontol. 18 (4), 307–313 (317–331).
Goncharova, I.A., Shcherba, I.G., Khondkarian, S.O., 2004. Map 5: Early MiddleMiocene (Langhian, Early Badenian, Chokrakian) 16–15 Ma. In: Popov, S.V., etal. (Eds.), Lithological-Paleogeographic Maps of Paratethys. Schweizerbart'scheVerlagsbuchhandlung, Frankfurt am Mein, pp. 19–21.
Gorjanović-Kramberger, D., 1896. Über das Vorkommen der Pereiraia gervaisii Vez. sp.Verhandlungen der Geologischen Reichsanstalt 1–18.
Gosar, A., 2005. Geophysical and structural characteristics of the pre-Tertiary basement oftheMura Depression (SWPannonian Basin, NE Slovenia). Geol. Carpath. 56, 103–112.

156 M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
Hajós, M., 1986. Stratigraphy of Hungary's Miocene diatomaceous earth deposits. Fasc.Inst. Geol. Hungariae Series Paleontologica 49, 1–339.
Hámor, G., 2002. Significance of the Neo-Vardar Tectonic Zone in the Tertiary structuralevolution and paleogeographic changes of the Carpathian Basin. Acta Geol. Hung.45, 63–77.
Hámor, G. and Halmai, J., 1988. Neogene palaeogeographic atlas of Central and EasternEurope: scale 1:3.000.000. Hungarian Geological Survey.
Haq, B.U., Hardenbol, J., Vail, P.R., 1988. Mesozoic and Cenozoic chronostratigraphy andcycles of sea-level change. In: Wilgus, C.K., et al. (Eds.), Sea-Level Changes. SEPM,Tulsa, pp. 71–108.
Hardenbol, J., et al., 1998. Cenozoic sequence biochronostratigraphy. In: Graciansky, P.-C.-d.,Hardendol, J., Jacquin, T., Vail, P.R. (Eds.), Mesozoic and Cenozoic Sequence Stratigraphyof European Basins. SEPM, Tulsa (pp. Chart 2).
Harzhauser, M., Piller, W.E., Steininger, F.F., 2002. Circum-Mediterranean Oligo-Miocenebiogeographic evolution—the gastropods' point of view. Palaeogeogr. Palaeoclimatol.Palaeoecol. 183 (1), 103–133.
Harzhauser, M., Mandic, O., Zuschin, M., 2003. Changes in Paratethyan marine molluscs atthe Early/Middle Miocene transition: diversity, palaeogeography and palaeoclimate.Acta Geol. Pol. 53 (4), 323–339.
Hilber, V., 1893. Fauna der Pereiraia-Schichten von Bartelmae in Unter-Krain.Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftlichen Classe 1, 1005–1032.
Hilgen, F.J., Abdul Aziz, H., Krijgsman, W., Raffi, I., Turco, E., 2003. Integrated stratigraphyand astronomical tuning of the Serravallian and lower Tortonian at Monte dei Corvi(Middle–Upper Miocene, northern Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol.199, 229–264.
Hilgen, F.J., et al., 2009. The global stratotype section and point (GSSP) of the SerravallianStage (middle Miocene). Episodes 32 (3), 152–166.
Hilgen, F.J., Lourens, L.J., Van Dam, J.A., 2012. The Neogene Period. In: Gradstein, F.M., Ogg,J.G., Schmitz, M., Ogg, G. (Eds.), The Geologic Time Scale. Elsevier, Oxford, pp. 923–978.
Hörnes, R., 1895. Pereiraia gervaisii Véz. von Ivandol bei St. Bartelmae in Unterkrain.Annalen des Naturhistorischen Museums 10, 1–16.
Horvat, A., 2003. Badenian (Middle Miocene) diatoms (Bacillariophyta) from the westernpart of Central Paratethys. Dissertationes Academiae Scientarium et ArtiumSlovenica, Classis IV. Historia Naturalis, 44, pp. 5–55.
Horvat, A., 2004. Middle Miocene siliceous algae of Slovenia. ZRC Publishing, Ljubljana(255 pp.).
Horvat, A., 2012. Middle Miocene opal-rich facies of the Paratethys —evidence of upwell-ing or increased continental runoff stimulated productivity? In: Missoni, S., G.H. J.(Eds.), 29th IAS Meeting of Sedimentology, Manchester, p. 536.
Ivanov, D., Ashraf, A.R., Mosbrugger, V., Palamarev, E., 2002. Palynological evidence forMiocene climate change in the Forecarpathian Basin (central Paratethys, NWBulgaria). Palaeogeogr. Palaeoclimatol. Palaeoecol. 178 (1), 19–37.
Jansen, E., Raymo, M.E., Blum, P., 1996. Hole 982. In: Dearmont, L. (Ed.), Proceedings of theOcean Drilling Program, Initial report, pp. 91–138.
Jelen, B., Rifelj, H., Skaberne, D., Poljak, M., Kralj, P., 2008. Slovenian Paratethys basins. In:McCann, T. (Ed.), The Geology of Central Europe: Mesozioc and Cenozoic. GeologicalSociety Pub. House, London, pp. 1098–1102.
Kameo, K., Bralower, T.J., 2000. Neogene calcareous nanofossil biostratigraphy of Sites998, 999, and 1000, Caribbean Sea. In: Leckie, R.M., Sigurdsson, H. (Eds.), Proceedingsof the Ocean Drilling Program, Scientific Results. College Station, pp. 3–17.
Keferstein, C., 1828. BeobachtungenundAnsichtenüber die geognostischenVerhältnisse dernördlichen Kalk-Alpenkette in Oesterreich und Baiern. Teutschland Geognostisch-Geologisch Dargestellt 5, 1–425.
Kinkelin, F., 1892. Neogenbildungen westlich von St. Barthelmae in Unterkrain. Jahrbuchder Kaiserlich-Königlichen Geologischen Reichsanstalt, 41, pp. 401–414.
Kókay, J., 1966. A Herend-Márkói barnaköszénterület földtani és öslénytani vizsgálata.Geol. Hung. 36 (1), 1–149.
Kókay, J., 1985. Central and eastern Paratethyan interrelations in the light of the LateBadenian salinity conditions. Geol. Hung. 48, 7–95.
Kováč, M., Holcová, K., Nagymarosy, A., 1999. Paleogeography, paleobathymetry and rel-ative sea-level changes in the Danube Basin and adjacent areas. Geol. Carpath. 50,325–338.
Kováč, M., et al., 2007. Badenian evolution of the Central Paratethys Sea: paleogeography,climate and eustatic sea-level changes. Geol. Carpath. 58, 579–606.
Kováč, M., Sliva, L., Sopková, B., Hlavatá, J., Škulová, A., 2008. Serravallian sequence stratig-raphy of the northern Vienna Basin: high frequency cycles in the Sarmatian sedimen-tary record. Geol. Carpath. 59, 545–561.
Kováčová, P., Emmanuel, L., Hudáčková, N., Renard, M., 2009. Central Paratethyspaleoenvironment during the Badenian (Middle Miocene): evidence from foraminif-era and stable isotope (δ13C and δ18O) study in the Vienna Basin (Slovakia). Int.J. Earth Sci. 98, 1109–1127.
Kroh, A., 2007. Climate changes in the Early to Middle Miocene of the Central Paratethysand the origin of its echinoderm fauna. Palaeogeogr. Palaeoclimatol. Palaeoecol. 253(1–2), 169–207.
Lirer, F., et al., 2002. Astrochronological calibration of the Upper Serravallian/LowerTortonian sedimentary sequence at Tremiti Islands (Adriatic Sea, Southern Italy).Riv. Ital. Paleontol. Stratigr. 108, 241–398.
Lohman,W.H., 1986. Calcareous nannoplankton biostratigraphy of the southern Coral sea,and southwester Pacific Ocean, Deep Sea Drilling Project Leg 90: Neogene andQuarternary. In: Kennett, J.P., von der Borch, C.C. (Eds.), Initial Reports of the DeepSea Drilling Project. U.S. Government Printing Office, Washington D.C., pp. 763–793.
Magyar, I., Geary, D.H., Muller, P., 1999. Paleogeographic evolution of the Late Miocene LakePannon in Central Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 147 (3–4), 151–167.
Martini, E., 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation.Proc. 2nd Int. Conf. Planktonic Microfossils Roma: Rome, pp. 739–785.
Márton, E., et al., 2002. Miocene to Quaternary deformation in NE Slovenia: complex pa-leomagnetic and structural study. J. Geodyn. 34 (5), 627–651.
Mărunţeanu, M., 2006. Nanoflora depozitelor Neogen superioare din Bazinul Dacic şiimplicaţiile ei paleoambientale. In: Panin, N., Oaie, G., Jipa, D. (Eds.), Influenţafactorilor globali (clima, tectonica, eustatism) asupra evoluţiei Bazinului Dacic(Neogen Superior), Bucharest, pp. 230–261.
Massari, F., 1990. The foredeeps of the Northern Adriatic margin: evidence of diachroneityin deformation of the Southern Alps. Riv. Ital. Paleontol. Stratigr. 96, 351–379.
Meulenkamp, J.E., Sissingh, W., 2003. Tertiary palaeogeography and tectonostratigraphicevolution of the Northern and Southern peri-Tethys platforms and the intermediatedomains of the African–Eurasian convergent plate boundary zone. Palaeogeogr.Palaeoclimatol. Palaeoecol. 196, 209–228.
Meulenkamp, J.E., et al., 2000. Early Langhian (16.4–15.5 Ma). In: Dercourt, J., Gaetani,B., Vrielynck, B. (Eds.), Atlas Peri-Tethys, Palaeogeographical Maps. CCGM/CGMW,Paris.
Mihajlović, D., Knežević, S., 1989. Calcareous nannoplankton from Badenian andSarmatian deposits at Višnjica and Karaburma in Belgrade. Geološki analiBalkanskoga poluostrva 53, 343–366.
Mikuž, V., 1999. Pereiraea gervaisi (Vezian) from Miocene beds south of Šentjernej inLower Carniola, Slovenia. Geophys. J. R. Astron. Soc. 42, 123–140.
Mikuž, V., 2009. Miocene gastropods from the vicinity of Šentjernej and from other local-ities in the Krka basin, Slovenia. Folia biologica et geologica 50, 5–70.
Mikuž, V., Horvat, A., 2003a. Paleobiogeografija klipeastrov (Echinoidea). Geološkizbornik 17, 119–123.
Mikuž, V., Horvat, A., 2003b. Miocenski morski ježki Slovenije. Geološki zbornik, 17,pp. 115–119.
Miller, K.G., et al., 1985. Oligocene–Miocene biostratigraphy, magnetostratigraphy, andisotopic stratigraphy of the western North Atlantic. Geology 13, 257–261.
Moissette, P., Dulai, A., Muller, P., 2006. Bryozoan faunas in the Middle Miocene ofHungary: biodiversity and biogeography. Palaeogeogr. Palaeoclimatol. Palaeoecol.233 (3–4), 300–314.
Müller, C., 1974. Nannoplankton aus dem Mittel-Miozän von Walbersdorf (Burgenland).Senckenb. Lethaea 55, 389–405.
Muza, J.P., 2000. Calcareous nannofossil biostratigraphy from a 15-km transect (Cocosplate to Caribbean plate) across the Middle America Trench, Nicoya Peninsula,Costa Rica. In: Silver, E.A., Kimura, G. (Eds.), Proceedings of the Ocean Drilling Pro-gram, Scientific Results. College Station, pp. 1–63.
Nagymarosy, A. and Müller, P., 1988. Some aspects of Neogene biostratigraphy in thePannonian Basin. In: L.H. Royden and F. Horváth (Editors), The Pannonian Basin: astudy in basin evolution. AAPG Memoir. AAPG Memoir. AAPG & The Hungarian Geo-logical Society, Tulsa, Budapest, pp. 69–77
Oblak, K., 2003. Upper Oligocene and Miocene Foraminifera from Plohov Breg nearPodčetrtek (Slovenia). Dissertationes Academiae Scientarium et Artium Slovenica,Classis IV. Historia Naturalis, 44, pp. 240–283.
Olafsson, G., 1989. Quantitative calcareous nannofossil biostratigraphy of upper Oligoceneto middle Miocene sediment from ODP Hole 667 A and middle Miocene sedimentfrom DSDP Site 574. In: Ruddiman, W., Sarnthein, M. (Eds.), Proceedings of theOcean Drilling Program, Scientific Results. College Station, pp. 3–13.
Otoničar, B., Cimerman, F., 2006. Facialna analiza, biostratigrafija in depozicijski modelsrednjemiocenskih karbonatnih kamnin med Krško vasjo in Obrežjem. In: Režun,B., Eržen, U., Petrič, M., Gantar, I. (Eds.), slovenski geološki kongres, Idrija, 2, p. 71.
Ozdínová, S., 2008. Badenian calcareous nanofossils from Smerovce ŠV-8 and Cifer-1boreholes (Danube Basin). Mineralia Slovaca 40, 141–150.
Paramonova, N.P., Shcherba, I.G., Khondkarian, S.O., 2004. Map 7: Late Middle Miocene(Late Seravallian, Sarmatian s.s., Middle Sarmatian s.l.). In: Popov, S.V., et al.(Eds.), Lithological–Paleogeographic Maps of Paratethys. Schweizerbart'scheVerlagsbuchhandlung, Frankfurt am Mein, pp. 27–30.
Parker, M.E., Clark, M., Sherwood, W.W., 1985. Calcareous nannofossils of Deep Sea Dril-ling Project Sites 558 and 563, North Atlantic Ocean: biostratigraphy and the distribu-tion of Oligocene braarudosphaerids. In: Bougault, H., Cande, C.S. (Eds.), Initial reportsof the Deep Sea Drilling Project. Government Printing Office, Washington,pp. 559–589.
Pavšič, J., Aničić, B., 1998. Nannoplankton stratigraphy of Oligocene andMiocene strata onPlohov Breg near Podčetrtek (Eastern Slovenia). Dissertationes AcademiaeScientarium et Artium Slovenica, Classis IV. Historia Naturalis, 34, pp. 55–79.
Pavšič, J., Horvat, A., 2009. The Eocene, Oligocene and Miocene in Central and EasternSlovenija. In: Pleničar, M., Ogorelec, B., Novak, M. (Eds.), The Geology of Slovenia.Geological Survey of Slovenia, Ljubljana, pp. 373–426.
Perch-Nielsen, K., 1985. Cenozoic calcareous nannofossils. In: Bolli, H.M., Saunders,J.B., Perch-Nielsen, K. (Eds.), Plankton Stratigraphy: Planktic Foraminifera, Cal-careous Nannofossils and Calpionellids. Cambridge University Press,pp. 427–554.
Peryt, T.M., 2006. The beginning, development and termination of the Middle Mio-cene Badenian salinity crisis in Central Paratethys. Sediment. Geol. 188,379–396.
Pezelj, Đ., Sremac, J., 2007. Badenian marginal marine environment in the MedvednicaMt. (Croatia). Joannea - Geologie und Paläontologie 9, 83–84.
Piller, W.E., Harzhauser, M., Mandic, O., 2007. Miocene Central Paratethys stratigraphy—current status and future directions. Stratigraphy 4, 71–88.
Placer, L., 1999. Contribution to the macrotectonic subdivision of the border region be-tween Southern Alps and External Dinarides. Geophys. J. R. Astron. Soc. 41 (1998),223–255.
Pleničar, M., Nosan, A., 1958. Paleogeografija panonskega obrobja v Sloveniji. Geophys.J. R. Astron. Soc. 4, 94–110.
Popov, S.V., et al., 2004. Lithological–paleogeographic maps of Paratethys, 250.Schweizerbart'sche Verlagsbuchhandlung (46 pp.).

157M. Bartol et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 394 (2014) 144–157
Raffi, I., Flores, J.A., 1995. Pleistocene through Miocene calcareous nannofossils fromeastern equatorial Pacific Ocean (Leg 138). In: Pisias, N.G., Mayer, L.A. (Eds.),Proceedings of the Ocean Drilling Program, Scientific Results. College Station,pp. 233–286.
Raffi, I., Rio, D., d'Atri,, A., Rochetti, S., 1995. Quantitative distribution patterns andbiomagnetostratigraphy of Middle and late Miocene calcareous nannofossils fromequatorial Indian and Pacific Oceans (Legs 115, 130, and 138). In: Pisias, N.G.,Mayer, L.A., et al. (Eds.), Proceedings of the Ocean Drilling Program, Scientific Results.College Station, pp. 479–502.
Řeháková, Z., 1978. Die marine Diatomeen flora des Badenien in der Zentralen Paratethys.In: Brestenská, E. (Ed.), Chronostratigraphie und Neostratotypen, Miozan M4.Bratislava, Badenien, pp. 533–553.
Rižnar, I., Miletić, D., Verbič, T., Horvat, A., 2002. Middle Miocene sediments on the north-ern part of Gorjanci between Čatež and Kostanjevica (SE Slovenia). Geophys. J. R.Astron. Soc. 45, 531–536.
Rögl, F., 1998. Palaeogeographic considerations for Mediterranean and Paratethys sea-ways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien 99,279–310.
Rögl, F., Müller, C., 1976. Das Mittelmiozän und die Baden-Sarmat Grenze inWalbersdorf (Burgenland). Annalen des Naturhistorischen Museums in Wien80, 221–232.
Rögl, F., Steininger, F.F., 1983. Vom Zerfall der Tethys zu Mediterran und Paratethys.Annalen des Naturhistorischen Museums in Wien 85, 135–163.
Rögl, F., Steininger, F.F., Müller, C., 1978. Middle Miocene salinity crisis and paleogeogra-phy of the Paratethys (Middle and Eastern Europe). Initial Rep. Deep Sea Drill. Proj.42 (1), 985–990.
Rögl, F., et al., 2007. Cyclostratigraphy and transgressions at the Early/Middle MioceneKarpatian/Badenian) boundary in the Austrian Neogene Basins (Central Paratethys).Scripta Facultatis scientiarum naturalium Universitatis Masarykianae Brunensis. Ge-ology, 36, pp. 7–13.
Sacco, F., 1905. Sopra un Pereiraia del Miocene della Sardegna. Riv. Ital. Paleontol. Stratigr.11, 112.
Sblendorio-Levy, J., Howe, R.W., 1998. Calcareous nannofossil biostratigraphy of Site953. Canary Basin, northeast North Atlantic. In: Weaver, P.P.E., Schmincke, H.-U.,et al. (Eds.), Proceedings of the Ocean Drilling Program, Scientific Results. CollegeStation, pp. 83–96.
Sprovieri, R., et al., 2002. An integrated calcareous plankton biostratigraphic scheme andbiochronology for the Mediterranean Middle Miocene. Riv. Ital. Paleontol. Stratigr.108, 337–353.
Steiniger, F., Rögl, F., Müller, C., 1978. Geodynamik und paläogeographische Entwicklungdes Badenien. In: Brestenská, E. (Ed.), Chronostratigraphie und Neostratotypen,Miozän M4. VEDA, Bratislava, Badenien, pp. 110–127.
Strausz, L., 1966. Die miozän-mediterranen Gastropoden Ungarns. Akedémiai Kiadó,Budapest (692 pp.).
Studencka, B., Goncharova, I.A., Popov, S.V., 1998. The bivalve faunas as a basis for recon-struction of theMiddleMiocene history of Paratethys. Acta Geol. Pol. 48 (3), 285–342.
Šuklje, F., 1929. Mediteranska fauna Zaprešić brijega u Samoborskoj gori. VijestiGeološkog zavoda 3, 1–52.
Theodoridis, S., 1984. Calcareous nannofossil biozonation of the Miocene and revision ofthe helicoliths and discoasters. Utrecht micropaleontological bulletins 32, 271.
Tomljenović, B., Csontos, L., 2001. Neogene–Quaternary structures in the border zone be-tween Alps, Dinarides and Pannonian basin (Hrvatsko Zagorje and Karlovac basins,Croatia). Int. J. Earth Sci. 90 (3), 560–578.
Turco, E., et al., 2002. Middle Miocene high-resolution calcareous plankton biostratigra-phy at Site 926 (Leg 154, equatorial Atlantic Ocean): palaeoecological andpalaeobiogeographical implications. Geobios 35, 257–276.
Vadász, E., 1960. Magyarország földtana: második, átdolgozott és bövített kiadás.Akadémiai Kiadó, Budapest 1–646.
Vézian, A., 1856. Mollusques et Zoophytes des Terrains Nummulitique et Tertiaire marin dela province de Barcelone. Faculté des Sciences de Montpellier, Montpellier (72 pp.).
Wei, W., Wise Jr., S.W., 1992. Oligocene–Pleistocene calcareous nannofossils from South-ern Ocean sites 747, 748, and 751. In: Wise Jr., S.W., Schlich, R. (Eds.), Proceedings ofthe Ocean Drilling Program, Scientific Results, pp. 509–522.
Wise, S.W., Covington, M., Ladner, B.C., Wei, W., 2002. NannoWare/BugCam Cenozoic Dig-ital-Image Catalog.
Young, J.R., 1998. Neogene. In: Bown, P.R. (Ed.), Calcareous nannofossil biostratigraphy.British Micropalaeontological Society Publication Series. Chapman and Hall; KluwerAcademic, London, pp. 225–265.
Zachos, J.C., Kroon, D., Bölum, P., et al., 2004. Site 1264. In: Nevill, H. (Ed.), Proceedings ofthe Ocean Drilling Program, Initial Reports, pp. 1–73.
Zbyszewski, G., 1957. Le Burdigalien de Lisbone. Communicações da Commissão doServiço Geológico de Portugal 38, 91–215.
Related Documents











