Estuarine Lago Mare fauna from the Tertiary Piedmont Basin indicates episodic Atlantic/Mediterranean exchange during the final stage of the Mediterranean Salinity Crisis Patrick Grunert a,b,c, ⁎, Mathias Harzhauser d , Yair Rosenthal b,c , Giorgio Carnevale e a Institute of Earth Sciences, University of Graz, NAWI Graz, Heinrichstrasse 26, A-8010 Graz, Austria b Department of Earth and Planetary Sciences, Rutgers University, 610 Taylor Road, Piscataway, NJ 08854-8066, United States c Department of Marine and Coastal Sciences, Rutgers University, 71 Dudley Road, New Brunswick, NJ 08901, United States d Geological-Paleontological Department, Natural History Museum Vienna, Burgring 7, A-1010 Vienna, Austria e Dipartimento di Scienze della Terra, Università degli Studi Torino, Via Valperga Caluso 35, I-10125 Torino, Italy abstract article info Article history: Received 10 November 2015 Received in revised form 23 May 2016 Accepted 5 June 2016 Available online 6 June 2016 The geochemical signature of fossils from Lago Mare deposits at Moncucco Torinese (NW Italy) indicates a tem- porary marine ingression into the Tertiary Piedmont Basin just before the Miocene/Pliocene boundary. Isotope (δ 18 O, δ 13 C, 87 Sr/ 86 Sr) and trace element (Sr/Ca, Ba/Ca) data from mollusk shells of multiple species and croaker otoliths indicate an upper estuarine environment with considerable seasonal variability in evaporation and fresh- water input. Reconstructed salinities b 10 psu and low 87 Sr/ 86 Sr values indicate the close proximity of a water- shed draining off of Mesozoic ophiolites and carbonates of the Alps and/or Apennines. Communication with an open marine environment is supported by otoliths of fully marine fishes such as lanternfishes, codlets, and slimeheads found in the same assemblage that carry a euhaline signature close to open ocean values. A thorough taphonomic evaluation renders reworking of the otoliths from pre-evaporitic deposits unlikely; instead we ex- plain their presence in the Lago Mare assemblage as related to large predatory marine fish that periodically mi- grated into marginal marine and estuarine environments. The presence of a paralic paleobiotope with thalassogenic water in the Tertiary Piedmont Basin contradicts ideas of the Mediterranean as a brackish lake fed by the Paratethys during the latest stage of the Mediterranean Salinity Crisis. Instead, our data support previous notions of temporary Atlantic/Mediterranean exchange during sea-level highstands preceding the permanent re-connection at the Miocene/Pliocene transition. The establishment of short-lived and disjunct estuarine and lagoonal environments such as in the Tertiary Piedmont Basin during ma- rine ingressions also puts highly variable 87 Sr/ 86 Sr values and occurrences of shallow water benthic foraminifera in Lago Mare deposits of other Mediterranean basins into a new perspective. © 2016 The Authors. Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/). Keywords: Messinian Mediterranean Lago Mare Stable isotopes Sr isotopes Trace elements 1. Introduction Tectonic and climatic forcing increasingly isolated the Mediterra- nean Sea from the open ocean during the early Messinian. Exchange with the Atlantic was finally severed at ~5.97 Ma, marking the onset of the Messinian Salinity Crisis (MSC), and a permanent connection was only re-established by the early Pliocene (Manzi et al., 2013; Hernández-Molina et al., 2014; Roveri et al., 2014a; Van der Schee et al., in press). Isolation resulted in a succession of hyper- and hyposaline conditions in the Mediterranean: widespread deposition of thick evaporitic deposits, progressing stepwise from marginal to deep basins and culminating at 5.6–5.55 Ma, was followed by mostly brackish deposits until 5.33 Ma (Roveri et al., 2014a). The fossil record suggests that these stressful and volatile conditions had a devastating impact on Mediterranean ecosystems and strongly favored opportunistic spe- cies. The peculiar and prevalent late MSC “Lago Mare” fauna, mainly consisting of euryhaline ostracods and mollusks, is often cited as a prime example. Based on its species content and low strontium isotope values the existence of a late MSC brackish lake, probably sourced by Paratethys waters, was inferred (Orszag-Sperber, 2006; Rouchy and Caruso, 2006; Roveri et al., 2014b). However, this interpretation has been challenged by frequent occurrences of foraminifera and marine fish in Lago Mare deposits, and episodic exchange with the Atlantic dur- ing the late MSC has been proposed as an alternative hypothesis (see e.g., Butler et al., 1999; Carnevale et al., 2006b, 2008b; Bache et al., 2012; Mezger, 2012; Zecchin et al., 2013; Roveri et al., 2014a and Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79 ⁎ Corresponding author at: Institute of Earth Sciences, University of Graz, NAWI Graz, Heinrichstrasse 26, A-8010 Graz, Austria. E-mail address: [email protected] (P. Grunert). http://dx.doi.org/10.1016/j.palaeo.2016.06.005 0031-0182/© 2016 The Authors. Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/). Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Estuarine Lago Mare fauna from the Tertiary Piedmont Basin indicatesepisodic Atlantic/Mediterranean exchange during the final stage of theMediterranean Salinity Crisis
Patrick Grunert a,b,c,⁎, Mathias Harzhauser d, Yair Rosenthal b,c, Giorgio Carnevale e
a Institute of Earth Sciences, University of Graz, NAWI Graz, Heinrichstrasse 26, A-8010 Graz, Austriab Department of Earth and Planetary Sciences, Rutgers University, 610 Taylor Road, Piscataway, NJ 08854-8066, United Statesc Department of Marine and Coastal Sciences, Rutgers University, 71 Dudley Road, New Brunswick, NJ 08901, United Statesd Geological-Paleontological Department, Natural History Museum Vienna, Burgring 7, A-1010 Vienna, Austriae Dipartimento di Scienze della Terra, Università degli Studi Torino, Via Valperga Caluso 35, I-10125 Torino, Italy
⁎ Corresponding author at: Institute of Earth Sciences,Heinrichstrasse 26, A-8010 Graz, Austria.
E-mail address: [email protected] (P. Grune
http://dx.doi.org/10.1016/j.palaeo.2016.06.0050031-0182/© 2016 The Authors. Published by Elsevier B.V
a b s t r a c t
a r t i c l e i n f oArticle history:Received 10 November 2015Received in revised form 23 May 2016Accepted 5 June 2016Available online 6 June 2016
The geochemical signature of fossils from Lago Mare deposits at Moncucco Torinese (NW Italy) indicates a tem-porary marine ingression into the Tertiary Piedmont Basin just before the Miocene/Pliocene boundary. Isotope(δ18O, δ13C, 87Sr/86Sr) and trace element (Sr/Ca, Ba/Ca) data frommollusk shells of multiple species and croakerotoliths indicate anupper estuarine environmentwith considerable seasonal variability in evaporation and fresh-water input. Reconstructed salinities b 10 psu and low 87Sr/86Sr values indicate the close proximity of a water-shed draining off of Mesozoic ophiolites and carbonates of the Alps and/or Apennines. Communication with anopen marine environment is supported by otoliths of fully marine fishes such as lanternfishes, codlets, andslimeheads found in the same assemblage that carry a euhaline signature close to open ocean values. A thoroughtaphonomic evaluation renders reworking of the otoliths from pre-evaporitic deposits unlikely; instead we ex-plain their presence in the Lago Mare assemblage as related to large predatory marine fish that periodically mi-grated into marginal marine and estuarine environments.The presence of a paralic paleobiotope with thalassogenic water in the Tertiary Piedmont Basin contradicts ideasof theMediterranean as a brackish lake fed by the Paratethys during the latest stage of theMediterranean SalinityCrisis. Instead, our data support previousnotions of temporary Atlantic/Mediterranean exchangeduring sea-levelhighstands preceding the permanent re-connection at the Miocene/Pliocene transition. The establishment ofshort-lived and disjunct estuarine and lagoonal environments such as in the Tertiary Piedmont Basin during ma-rine ingressions also puts highly variable 87Sr/86Sr values and occurrences of shallowwater benthic foraminiferain Lago Mare deposits of other Mediterranean basins into a new perspective.
© 2016 The Authors. Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Keywords:MessinianMediterraneanLago MareStable isotopesSr isotopesTrace elements
1. Introduction
Tectonic and climatic forcing increasingly isolated the Mediterra-nean Sea from the open ocean during the early Messinian. Exchangewith the Atlantic was finally severed at ~5.97 Ma, marking the onsetof the Messinian Salinity Crisis (MSC), and a permanent connectionwas only re-established by the early Pliocene (Manzi et al., 2013;Hernández-Molina et al., 2014; Roveri et al., 2014a; Van der Scheeet al., in press). Isolation resulted in a succession of hyper- andhyposaline conditions in the Mediterranean: widespread deposition ofthick evaporitic deposits, progressing stepwise from marginal to deep
University of Graz, NAWI Graz,
rt).
. This is an open access article under
basins and culminating at 5.6–5.55Ma,was followed bymostly brackishdeposits until 5.33 Ma (Roveri et al., 2014a). The fossil record suggeststhat these stressful and volatile conditions had a devastating impacton Mediterranean ecosystems and strongly favored opportunistic spe-cies. The peculiar and prevalent late MSC “Lago Mare” fauna, mainlyconsisting of euryhaline ostracods and mollusks, is often cited as aprime example. Based on its species content and low strontium isotopevalues the existence of a late MSC brackish lake, probably sourced byParatethys waters, was inferred (Orszag-Sperber, 2006; Rouchy andCaruso, 2006; Roveri et al., 2014b). However, this interpretation hasbeen challenged by frequent occurrences of foraminifera and marinefish in LagoMare deposits, and episodic exchangewith the Atlantic dur-ing the late MSC has been proposed as an alternative hypothesis (seee.g., Butler et al., 1999; Carnevale et al., 2006b, 2008b; Bache et al.,2012; Mezger, 2012; Zecchin et al., 2013; Roveri et al., 2014a and
the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).

71P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
references therein). In addition, the taxonomic revision of a Lago Maremollusk fauna fromnorthern Italy has cast doubt on biogeographic rela-tions to Paratethyan faunas (Harzhauser et al., 2015; Neubauer et al.,2015).
In this study, we apply geochemical proxy methods to co-occurringbrackish mollusks and marine otoliths from Lago Mare deposits in theTertiary Piedmont Basin (NW Italy) to unravel their taphonomy andhabitats. The paleoenvironmental reconstruction will offer new per-spectives on Atlantic/Mediterranean exchange during the late MSC.
2. Study area
The Tertiary Piedmont Basin (TPB) in NW Italy is a wedge-top basinwithin the arc of the Southwestern and Ligurian Alps that developed inresponse to the collision of the European and Adriatic plates (Fig. 1;Bertotti and Mosca, 2009; Rossi et al., 2009; Maino et al., 2013). TheTPB infill consists of Eocene to Pliocene sediments which are in turncovered by Pliocene to Holocene deposits of the Savigliano and Alessan-dria piggy-back basins (see e.g., Bertotti and Mosca, 2009; Rossi et al.,2009; Irace et al., 2010 for details). The Messinian deposits of the TPBshow a fourfold development characteristic of marginal Mediterraneanbasins. Hemipelagic marls and laminated mudstones of the upperTortonian to lower Messinian Marne di Sant'Agata Formation are un-conformably overlain by selenitic gypsum of the Vena de Gesso Forma-tion (corresponding to the Primary Lower Gypsum, 5.97–5.6Ma; Roveriet al., 2014a). The top of the evaporites is bound by an erosional surface,followed by re-deposited evaporites of the “Valle Versa Chaotic Com-plex” (Dela Pierre et al., 2011). The uppermost Messinian is marked byintercalation of brackish and continental deposits of the “facies àCongeria” with the Conglomerati di Cassano Spinola Formation (Iraceet al., 2010). Marine shales and marls of the Argille Azzurre Formationdocument the re-establishment of normal marine conditions at thebase of the Pliocene (Violanti et al., 2011). At present, Messinian sedi-ments are exposed in two areas along the northern (Torino Hill,Monferrato) and southern (Langhe, Alto Monferrato, Borbera Grue do-mains) margins of the TPB (Fig. 1).
The studied material derives from post-evaporitic upper Messiniandeposits in the gypsum quarry at Moncucco Torinese, situated alongthe southern slope of the Torino Hills (Figs. 1, 2; Trenkwalder et al.,2008; Angelone et al., 2010; Alba et al., 2014; Colombero et al., 2014,
Fig. 1. Geographic and geological setting of the Tertiary Piedmont Basin and the Po Plain,showing the position of the Moncucco Torinese section. Sediment distribution in grey;black lines indicate boundaries between major tectonic units (adopted from Harzhauseret al., 2015).
2015). The brackish and continental “facies à Congeria” overlies the“Valle Versa Chaotic Complex” and comprises up to 6.5 m of beige togreen-blue clayey marls with occasional, locally constrained calcareouspaleosol interbeds. Several depositional units have been distinguishedwithin the “facies à Congeria” that vary laterally in thickness and fossilcontent (Angelone et al., 2010). The basal unit consists of sandy claysand contains ostracods and rare brackish mollusks representing theLago Mare biofacies (M1 in Angelone et al., 2010). Conglomeratesmark an erosional surface at the top of the basal layer. Overlying unitsM2–8 consist of sandy clays with occasional, locally constrained con-glomerates and paleosols. Fossil assemblages consist of mixed terres-trial and reworked brackish fossils; the latter are shells of dreissenidand limnocardiid bivalves alongwithmelanopsid and hydrobiid gastro-pods (Harzhauser et al., 2015). Ostracods andmammals allow a correla-tion with upper Messinian mammal zone MN 13 and theLoxocorniculina djafarovi Zone (5.40 to 5.33 Ma; Trenkwalder et al.,2008; Angelone et al., 2010; Colombero et al., 2014).
3. Material and methods
A variety of terrestrial and aquaticmollusk taxa has been selected foranalyses from beds M3 and M4 (Table 1; Fig. 2). Whenever possible,portions of the same shell fragment have been used for geochemicalanalyses and SEM documentation. A variety of marine and estuarineotolith taxa have been recovered from the “facies à Congeria”, ofwhich 5 taxa from units M3-M5 have been selected for isotope analyses(Tables 2, 3).
3.1. Stable isotopes (δ18O, δ13C)
With the exception of Limax, mollusk shells have not been drilleddue to their small size, but small pieces have been broken off close tothe aperture. For some larger specimens, measurements have been per-formed on several fragments to observe internal variability. Shell frag-ments and otoliths have been ultrasonically cleaned with H2Odest andMethanol for several minutes to remove clay. Isotopic analyses havebeen performed using an automatic Kiel II preparation line and aFinniganMAT Delta Plusmass spectrometer at the Institute of Earth Sci-ences, University of Graz. Samples were reacted with 100% phosphoricacid at 70 °C. Analytical precision, based on replicate analysis of interna-tional standards NBS-19 and NBS-18 and an internal laboratory stan-dard is b0.04‰ for δ13C and b0.08‰ for δ18O. Results are reported inconventional δ-notation relative to the Vienna Pee Dee Belemnite stan-dard (VPDB) in ‰ units.
3.2. Trace metals
Shell fragments (weight: 170–450 μg) of thirteen terrestrial andaquatic gastropods have been analyzed for Sr/Ca and Ba/Ca. Adherentclays were removed by repeated wash with distilled, deionized (dd)H2O and Methanol. Organic matter was removed with an oxidative re-agent (200 μL H2O2 + 20mL dd H2O+ 10mL NH4OH). Finally, a diluteacid rinse with 250 μL of 0.001 N HNO3was applied. Following cleaning,the samples were dissolved in 100–150 μL of 0.065 N HNO3. After10 min of centrifugation, 90 μL of the dissolved sample were added to300 μL of 0.5 N HNO3.
Trace metal analysis was performed at the Department of MarineandCoastal Sciences, RutgersUniversity, on a ThermoScientific Element2 inductively coupled plasma mass spectrometer (ICP-MS). Repeatedanalyses of standard solutions with constant element ratios (El/Ca)but changing Ca concentrations ([Ca] = 1.5 mM–8 mM) were used toquantify and correct for matrix changes. Precision is b0.5% for Sr/Caand b1% for Ba/Ca (RSD).

Fig. 2. Stratigraphic sketch of the Moncucco Torinese section. Important biostratigraphictie-points and position of sampled beds in the “facies à Congeria” are indicated.Sampling range for mollusk (MO) and otolith (OT) fossils are indicated by grey boxes.For further details see Angelone et al. (2010) and Colombero et al. (2014).
72 P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
3.3. Sr isotopes (87Sr/86Sr)
87Sr/86Sr analysis has been performed on 8 gastropod and 4 otolithsamples at the Department of Earth and Planetary Sciences, RutgersUniversity. Adherent clays were removed by successive washes withdd H2O and Methanol. 800–9100 μg of the cleaned material werecrushed and homogenized with a mortar for each sample. The powderhas been dissolved in 1.5 N HCl, Sr has been separated using standardion exchange techniques (Hart and Brooks, 1974).
The samples were analyzed on an Isoprobe T Multicollector TIM. In-ternal precision for the data set averaged 0.000007, the external preci-sion is approximately ±0.000008 (based on replicate analyses ofstandards). NBS 987 is measured for these analysis at 0.710241 normal-ized to 86Sr/88Sr of 0.1194.
3.4. SEM documentation
Preservation of mollusk shells has been screened with a Zeiss DSM982 Gemini scanning electron microscope (SEM) at the Institute ofEarth Sciences, University of Graz. Images have been taken at 5–10 kVaccelerating voltage by using the DISS 5 software package (version5.9.9.1). The SEM images reveal well-preserved mollusk shells with in-ternal layers of differently orientated aragonite crystals intact (Fig. 3).No signs of overgrowth have been observed.
4. Results
4.1. Stable isotopes
Terrestrial and aquatic mollusks are clearly separated in their isoto-pic values (Table 1; Fig. 4). δ13C values of terrestrial gastropods are thelowest of the entire data-set, and fall into two distinctive groups: valuesof Parmacella sp. fall ~4‰ below those of helicids and Limax fossilis. δ18Ovalues are negative and belowmost aquaticmollusks exceptMelanoidescurvicosta and individual Melanopsis specimens.
Aquatic mollusks show a range of ~3.5‰ (−1.18‰ to 2.30‰) forδ13C (not including M. curvicosta outlier, MO 4) with mostly positivevalues, and a considerably wider range of ~7.6‰ in δ18O. Scatter ofδ13C is large in all taxa except Prososthenia and most bivalves, bothalso showing lowest δ13C. Two slightly overlapping groups of dataform along the δ18O-gradient. The first group comprises melanopsidsand neritids and exhibits mainly negative values. M. curvicosta consis-tently shows lowest δ18O overlapping with terrestrial taxa. Melanopsisfusulatina, M. narzolina and Theodoxus spp. show comparably enrichedδ18O, the latter two taxa being well separated from M. curvicosta andpartly overlapping with Saccoia spp. The second group compriseshydrobiid gastropods and dreissenid and cardiid bivalves and showsprimarily positive values. Highest δ18O values occur in Prosostheniawhich also shows the smallest scatter.
Repeated measurements on different pieces of the same molluskshell show an average offset of ~0.5‰ for δ18O (total range: 0.02–1.83‰) and ~0.4‰ for δ13C (total range: 0.01–1.54‰).
δ13C of marine otoliths generally falls below aquatic mollusks withthe exception of Hoplostethus cf. mediterraneus, δ18O falls in the upperrange of aquatic mollusks (Table 3). The isotopic signal of the brackishcroaker Sciaenidarum sp. is distinctively different from marine taxa byits severely depleted isotopic values.
4.2. Sr isotopes
87Sr/86Sr of gastropods and otoliths form twowell-separated groups(average Δ87Sr/86Sr = 0.00015; Tables 1, 4; Fig. 5). Mollusk values(0.708745–0.708805) are well below those of otoliths, and no system-atic trend between melanopsids and hydrobiids is apparent withinthis narrow range. The variability of the enriched otolith 87Sr/86Sr(0.708889–0.708940) shows a similar range.
4.3. Trace metals
Lowest Sr/Ca ratios of the data-set separate terrestrial from aquaticgastropods (Table 1). Helicid Sr/Ca ratios fall slightly below aquaticmol-lusks and Ba/Ca ratios are comparable to Saccoia spp. andM. curvicosta.In strong contrast, Parmacella sp. shows extremely low values in bothratios.
Amongst aquatic gastropods,Melanopsis spp. stand out with highestSr/Ca and Ba/Ca ratios of the data-set. Low Sr/Ca ratios occur in Saccoiaspp., Prososthenia spp. and M. curvicosta share intermediate values. Ba/Ca show considerable variability. They are lowest in M. curvicosta,Prososthenia spp. and Saccoia spp. share intermediate values.
Mn/Ca, Fe/Ca, and Ti/Ca ratios arewell below limits of contaminationfor all samples, only Al/Ca levels of MO 2 (Saccoia sp. 1) and MO 23
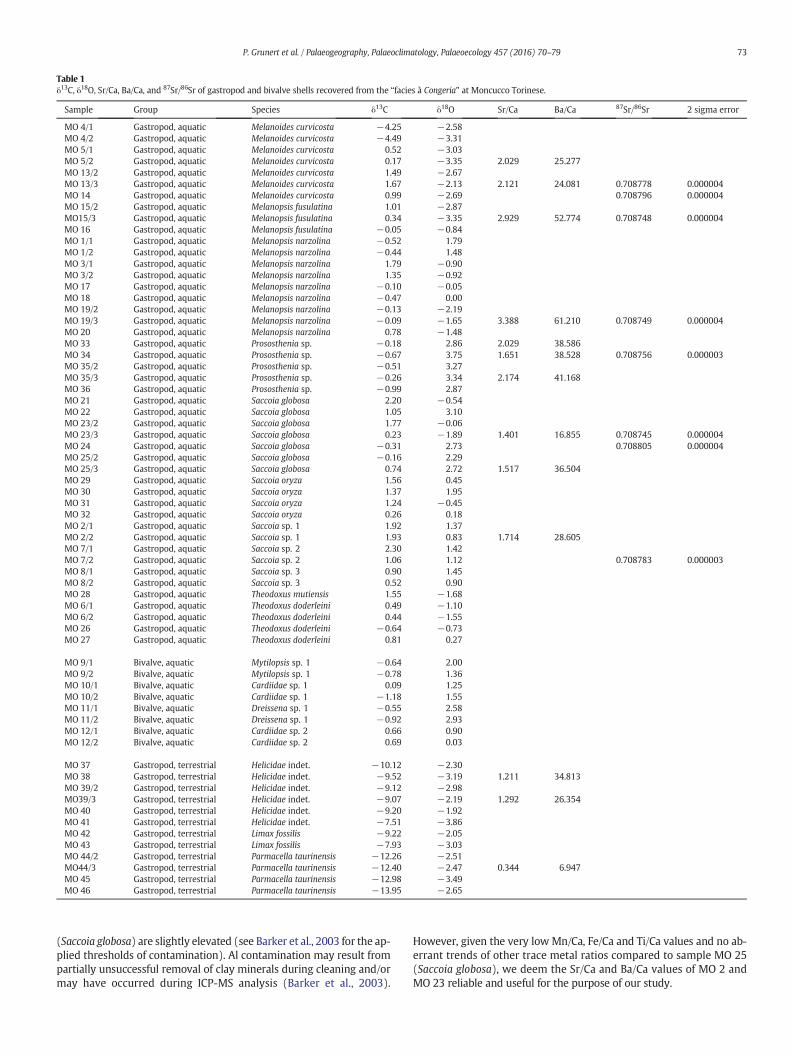
Table 1δ13C, δ18O, Sr/Ca, Ba/Ca, and 87Sr/86Sr of gastropod and bivalve shells recovered from the “facies à Congeria” at Moncucco Torinese.
Sample Group Species δ13C δ18O Sr/Ca Ba/Ca 87Sr/86Sr 2 sigma error
MO 4/1 Gastropod, aquatic Melanoides curvicosta −4.25 −2.58MO 4/2 Gastropod, aquatic Melanoides curvicosta −4.49 −3.31MO 5/1 Gastropod, aquatic Melanoides curvicosta 0.52 −3.03MO 5/2 Gastropod, aquatic Melanoides curvicosta 0.17 −3.35 2.029 25.277MO 13/2 Gastropod, aquatic Melanoides curvicosta 1.49 −2.67MO 13/3 Gastropod, aquatic Melanoides curvicosta 1.67 −2.13 2.121 24.081 0.708778 0.000004MO 14 Gastropod, aquatic Melanoides curvicosta 0.99 −2.69 0.708796 0.000004MO 15/2 Gastropod, aquatic Melanopsis fusulatina 1.01 −2.87MO15/3 Gastropod, aquatic Melanopsis fusulatina 0.34 −3.35 2.929 52.774 0.708748 0.000004MO 16 Gastropod, aquatic Melanopsis fusulatina −0.05 −0.84MO 1/1 Gastropod, aquatic Melanopsis narzolina −0.52 1.79MO 1/2 Gastropod, aquatic Melanopsis narzolina −0.44 1.48MO 3/1 Gastropod, aquatic Melanopsis narzolina 1.79 −0.90MO 3/2 Gastropod, aquatic Melanopsis narzolina 1.35 −0.92MO 17 Gastropod, aquatic Melanopsis narzolina −0.10 −0.05MO 18 Gastropod, aquatic Melanopsis narzolina −0.47 0.00MO 19/2 Gastropod, aquatic Melanopsis narzolina −0.13 −2.19MO 19/3 Gastropod, aquatic Melanopsis narzolina −0.09 −1.65 3.388 61.210 0.708749 0.000004MO 20 Gastropod, aquatic Melanopsis narzolina 0.78 −1.48MO 33 Gastropod, aquatic Prososthenia sp. −0.18 2.86 2.029 38.586MO 34 Gastropod, aquatic Prososthenia sp. −0.67 3.75 1.651 38.528 0.708756 0.000003MO 35/2 Gastropod, aquatic Prososthenia sp. −0.51 3.27MO 35/3 Gastropod, aquatic Prososthenia sp. −0.26 3.34 2.174 41.168MO 36 Gastropod, aquatic Prososthenia sp. −0.99 2.87MO 21 Gastropod, aquatic Saccoia globosa 2.20 −0.54MO 22 Gastropod, aquatic Saccoia globosa 1.05 3.10MO 23/2 Gastropod, aquatic Saccoia globosa 1.77 −0.06MO 23/3 Gastropod, aquatic Saccoia globosa 0.23 −1.89 1.401 16.855 0.708745 0.000004MO 24 Gastropod, aquatic Saccoia globosa −0.31 2.73 0.708805 0.000004MO 25/2 Gastropod, aquatic Saccoia globosa −0.16 2.29MO 25/3 Gastropod, aquatic Saccoia globosa 0.74 2.72 1.517 36.504MO 29 Gastropod, aquatic Saccoia oryza 1.56 0.45MO 30 Gastropod, aquatic Saccoia oryza 1.37 1.95MO 31 Gastropod, aquatic Saccoia oryza 1.24 −0.45MO 32 Gastropod, aquatic Saccoia oryza 0.26 0.18MO 2/1 Gastropod, aquatic Saccoia sp. 1 1.92 1.37MO 2/2 Gastropod, aquatic Saccoia sp. 1 1.93 0.83 1.714 28.605MO 7/1 Gastropod, aquatic Saccoia sp. 2 2.30 1.42MO 7/2 Gastropod, aquatic Saccoia sp. 2 1.06 1.12 0.708783 0.000003MO 8/1 Gastropod, aquatic Saccoia sp. 3 0.90 1.45MO 8/2 Gastropod, aquatic Saccoia sp. 3 0.52 0.90MO 28 Gastropod, aquatic Theodoxus mutiensis 1.55 −1.68MO 6/1 Gastropod, aquatic Theodoxus doderleini 0.49 −1.10MO 6/2 Gastropod, aquatic Theodoxus doderleini 0.44 −1.55MO 26 Gastropod, aquatic Theodoxus doderleini −0.64 −0.73MO 27 Gastropod, aquatic Theodoxus doderleini 0.81 0.27
MO 9/1 Bivalve, aquatic Mytilopsis sp. 1 −0.64 2.00MO 9/2 Bivalve, aquatic Mytilopsis sp. 1 −0.78 1.36MO 10/1 Bivalve, aquatic Cardiidae sp. 1 0.09 1.25MO 10/2 Bivalve, aquatic Cardiidae sp. 1 −1.18 1.55MO 11/1 Bivalve, aquatic Dreissena sp. 1 −0.55 2.58MO 11/2 Bivalve, aquatic Dreissena sp. 1 −0.92 2.93MO 12/1 Bivalve, aquatic Cardiidae sp. 2 0.66 0.90MO 12/2 Bivalve, aquatic Cardiidae sp. 2 0.69 0.03
MO 37 Gastropod, terrestrial Helicidae indet. −10.12 −2.30MO 38 Gastropod, terrestrial Helicidae indet. −9.52 −3.19 1.211 34.813MO 39/2 Gastropod, terrestrial Helicidae indet. −9.12 −2.98MO39/3 Gastropod, terrestrial Helicidae indet. −9.07 −2.19 1.292 26.354MO 40 Gastropod, terrestrial Helicidae indet. −9.20 −1.92MO 41 Gastropod, terrestrial Helicidae indet. −7.51 −3.86MO 42 Gastropod, terrestrial Limax fossilis −9.22 −2.05MO 43 Gastropod, terrestrial Limax fossilis −7.93 −3.03MO 44/2 Gastropod, terrestrial Parmacella taurinensis −12.26 −2.51MO44/3 Gastropod, terrestrial Parmacella taurinensis −12.40 −2.47 0.344 6.947MO 45 Gastropod, terrestrial Parmacella taurinensis −12.98 −3.49MO 46 Gastropod, terrestrial Parmacella taurinensis −13.95 −2.65
73P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
(Saccoia globosa) are slightly elevated (see Barker et al., 2003 for the ap-plied thresholds of contamination). Al contamination may result frompartially unsuccessful removal of clay minerals during cleaning and/ormay have occurred during ICP-MS analysis (Barker et al., 2003).
However, given the very low Mn/Ca, Fe/Ca and Ti/Ca values and no ab-errant trends of other trace metal ratios compared to sample MO 25(Saccoia globosa), we deem the Sr/Ca and Ba/Ca values of MO 2 andMO 23 reliable and useful for the purpose of our study.

Table 2Census counts and ecology of otolith taxa recovered from samples of the Moncucco Torinese section.
Bed
Family Taxon Ecology M2 M3 M3–4 M4 M4–5 M5 M7 + 25
Myctophidae Benthosema aff. suborbitale Marine mesopelagic 1 2 0 0 0 0 0Bolinichthys italicus Marine mesopelagic 0 1 0 0 0 0 0Diaphus befralai Marine mesopelagic 0 3 0 3 0 2 0Diaphus aff. pedemontanus Marine mesopelagic 0 3 0 4 0 2 0Diaphus aff. rubus Marine mesopelagic 0 0 0 1 0 0 0Diaphus splendidus Marine mesopelagic 0 6 0 9 0 0 0Diaphus taaningi Marine mesopelagic 0 1 0 2 1 0 0Hygophum aff. derthonensis Marine mesopelagic 0 0 0 3 1 1 1Myctophum coppa Marine mesopelagic 0 1 0 3 0 0 1
Gadidae Gadiculus labiatus Marine epi-mesopelagic; outer shelf and open ocean 0 1 0 0 0 0 0Moridae Physiculus sp. Marine epi-mesopelagic; outer shelf and open ocean 0 0 0 1 0 0 0Trachichthyidae Hoplostethus cf. mediterraneus Marine benthopelagic 0 0 0 0 1 0 0Bythitidae Grammonnus sp. Marine neritic 0 0 0 1 0 0 0Gobiidae Lesueurigobius sp. Marine neritic euryhaline 2 1 0 4 0 1 0Sciaenidae Argyrosomus sp. Marine neritic euryhaline 0 0 0 2 0 0 0
Sciaenidarum sp. nov. Estuarine resident 4 5 6 72 1 6 3“Trewasciaena” sp. Estuarine resident 1 1 2 9 0 4 0
74 P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
5. Discussion
5.1. Taphonomy
Macro- and microfossils of Moncucco Torinese form a highlyheterogeneous taphocoenosis (Angelone et al., 2010; Harzhauser et al.,2015). Terrestrial gastropods and vertebrates are consideredparautochthonous to the continental deposits of the “facies à Congeria”.Allochthonous components consist of (1) brackish mollusks reworkedby occasional fluvial flooding events from the directly underlying LagoMare layer (M1); and (2) marine microfossils reworked from thelower Messinian Marne di Sant'Agata Formation. The geochemical sep-aration of the terrestrial and brackish mollusks is in strong support ofthis interpretation, and the new results help to clarify the origin of theotoliths, unaddressed in previous publications.
The terrestrial gastropods, which reflect a heterogeneous landscapewith dry and rocky habitats as well as moist woody areas, provide anendmember for meteoric water (Harzhauser et al., 2015). This isreflected in the extremely depleted δ13C and low δ18O values and thevery low Sr/Ca of the helicids. The often erratic values of Parmacellasp. compared to the other terrestrial gastropods is most likely the resultof their different, two-layered shellmineralogy: no studies are available,but SEM images imply that a thick calcitic inner layer is followed by athin aragonitic outer layer. Substantially depleted Sr/Ca ratios and δ13Creflect the different mineralogy (Table 1; Lécuyer et al., 2012).
Partial reworking of theMarne di Sant'Agata Formation into the LagoMare deposits has been inferred from foraminiferal assemblages con-taining marker species such as Globorotalia miotumida (=Gr.conomiozea) and Gr. nicolae (Angelone et al., 2010; Lozar et al., 2010;Dela Pierre et al., 2011; Violanti et al., 2011). Sporadic occurrences of
Table 3δ13C and δ18O of otoliths recovered from the “facies à Congeria” at Moncucco Torinese.
Sample Taxon δ13C δ18O
OT-1A Diaphus spp. −2.43 1.75OT-1B Diaphus spp. −3.27 1.43OT-2A Diaphus spp. −3.55 2.54OT-2B Diaphus spp. −3.44 1.99OT-3A Diaphus spp. −2.45 2.82OT-3B Diaphus spp. −2.53 2.55OT-4 Hygophum spp. −1.53 2.79OT-5A Gadiculus labiatus −2.05 2.07OT-5B Gadiculus labiatus −3.84 2.65OT-6 Sciaenidarum spp. −5.35 −4.42OT-7 Hoplostethus cf. mediterraneus −0.10 1.90
planktic foraminifera (including Gr. miotumida) and bathyal benthic fo-raminifera have also been observed in sediment attached to one of theherein studied specimens of Saccoia globosa prior to cleaning. Microfos-sil preservation is generally poor (Violanti et al., 2011); herein observedforaminiferal shells are frequently broken, and have taken on a yellow-ish color. In contrast, mollusk shells and otoliths selected for the presentstudy are very well preserved, showing only minimal signs of mechan-ical damage and no indication for diagenesis (Fig. 3).
The absence of mollusks of theMarne di Sant'Agata Formation, suchas the frequent pectinid Amussiopecten, from the taphocoenosis pro-vides further evidence against re-working and may indicate size-sorting of the fossils according to their origin: While microfossils ofthe Marne di Sant'Agata Formation were easily remobilized, probablytransported over larger distances and redeposited throughout the “fa-cies à Congeria”, larger elements were only re-deposited during fluvialflooding events (Sturani, 1975; Violanti et al., 2011; Harzhauser et al.,2015).
A strong argument against reworking of the otoliths from theMarnedi Sant'Agata Formation comes from their 87Sr/86Sr values. Prior to theMSC, 87Sr/86Sr values of Mediterranean biogenic carbonates generallyfollow open ocean values until ~6.1 Ma, reflecting a well-establishedconnection to the Atlantic (Fig. 5; McArthur et al., 2001; Roveri et al.,2014a; Schildgen et al., 2014). Depleted 87Sr/86Sr values are only re-ported from a fewmarginal basins which became restricted at differenttimes due to local tectonic uplift (see Schildgen et al., 2014 for a detaileddiscussion). The TPB, however, remained well connected to the Medi-terranean prior to evaporite deposition, sustaining a diverse marinefauna (Sturani, 1975; Dela Pierre et al., 2011; Violanti et al., 2011). Ifthe otoliths originated from the hemipelagic Marne di Sant'Agata For-mation, their 87Sr/86Sr signature should reflect open ocean values and,consequently, have chronostratigraphic value. Cross-correlation withMcArthur et al. (2001) suggests individual ages between ~7.2 and~10 Ma for the otoliths (Table 4). Given the presence of Gr. miotumida(FCO-LO: 7.22–6.31 Ma; Krijgsman et al., 2004; Gradstein et al., 2012),Gr. nicolae (FO-LO: 6.83–6.72 Ma; Gradstein et al., 2012) and predomi-nantly sinistral coiling Neogloboquadrina acostaensis (acmes: 6.12 Ma,6.08 Ma; Lourens et al., 2004) in the reworked sediments, the87Sr/86Sr ages of the otoliths appear too old (within the 95% confidenceinterval) to be part of the reworked assemblage (Violanti et al., 2011).
5.2. Otoliths of marine fish in the Lago Mare deposits?
In contrast to the brackish mollusks, the otolith assemblage atMoncucco includes a diverse array of stenohaline open marine taxaalongside marginal marine taxa (Table 2). The herein analyzed

Fig. 3. Examples for gastropod shell preservation showing well-preserved crossed lamellar structure. Scale bar = 20 μm. (A) Melanoides curvicosta, sample MO 5; (B) Saccoia globosa,sample MO 25; (C) Prososthenia sp., sample MO 34.
75P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
Diaphus spp., Gadiculus labiatus and Hygophum aff. derthonensis areplanktivorous fish migrating between mesopelagic and epipelagicenvironments (e.g., Nafpaktitis et al., 1977; Cohen et al., 1990). TheMediterranean slimehead Hoplostethus cf. mediterraneus is abenthopelagic predatory fish that lives close to the muddy sea-floorbetween 100 and 1100 mwater depth (e.g., Maul, 1990). In contrast,the croaker Sciaenidarum sp. is considered a euryhaline neritic taxonthat thrives in estuarine environments (Carnevale et al., 2006a,2008b). This diversity of habitats is clearly expressed in the stable
Fig. 4. Stable isotope values (δ18O, δ13C) of mollusk shells and otoliths recovered from the“facies à Congeria” at Moncucco Torinese (Tables 1, 3). All values in‰ V-PDB.
isotopes (Fig. 4). Otolith δ13C is strongly determined bymetabolic ac-tivity, resulting in differences up to 4‰ between shallow (warm)and deep (cold) dwellers (Kalish, 1991; Thorrold et al., 1997; Linet al., 2012). Sciaenidarum sp. consequently shows severely depletedδ13C and δ18O, inhabiting the same oligohaline environment as themelanopsids (see below). Amongst oceanic taxa, δ13C of the deeperdwelling Hoplostethus cf. mediterraneus exceeds values of shallowerdwelling taxa. Similar patterns of δ13C and δ18O have been reportedfor mixed assemblages of brackish (Gobius, Sciaenidarum) and openmarine fish (Diaphus, Lophiiformes) in Lago Mare deposits from Tus-cany and Marche (Carnevale et al., 2006a, 2006b, 2008b; Caputoet al., 2009). The co-occurrence of taxa with such different ecologicalrequirements is related to predatory activity of fishes and other ver-tebrates, a taphonomic process that commonly contributes to the ac-cumulation of otoliths (Nolf, 1985). The digestive systems of largepredatory fishes foraging in the open sea (e.g., scombrids and caran-gids) very often contain numerous otoliths of myctophids and otherepipelagic and migratory mesopelagic fishes (Schäfer, 1966). Thesepredatory fishes commonly visit brackish biotopes such as estuariesattracted by the seasonal availability of food and thereby displace thepelagic otoliths. Unfortunately, their otoliths are extremely delicateand very rare in the fossil record, but their presence in Messinianbrackish paleobiotopes has been confirmed by skeletal remains(Carnevale et al., 2008a).
Otolith 87Sr/86Sr values clearly exceed typical values from LagoMaredeposits (Fig. 5; Roveri et al., 2014a; Flecker et al., 2015). The geologicalsetting of theMediterranean generally favors low 87Sr/86Sr river waters(e.g. Nile, Rhône), and depleted values during the late MSC have beenlinked to a humid climate and substantially increased runoff (Fleckeret al., 2002; Reuter et al., 2011; Topper et al., 2011). The elevated87Sr/86Sr values atMoncucco Torinese are thus best explained bymixingwith substantial amounts of Atlantic waters. At the same time otolithvalues fall slightly below late Messinian open ocean values (Δ-87Sr/86Sr = −11 ∗ 10−5 to −5.9 ∗ 10−5 for an average global value of0.709006 during the MSC; McArthur et al., 2001; Fig. 5) which maypoint to a rather sluggish exchange (Flecker et al., 2002). However,local effects cannot be discounted, as 87Sr/86Sr values in otoliths fromTuscany approachopen ocean values (Carnevale et al., 2008b). In a com-parably remote setting like the Adriatic subbasins, river input from theApennines might have had a large effect. Partly dissolution of Vena delGesso gypsum deposits of the Apennine foredeep might have furthercontributed to the low 87Sr/86Sr values (Müller and Mueller, 1991;Lugli et al., 2007). However, as the gypsumandotolith values aremerelyidentical, we consider its contribution minor. Mixing with Paratethyswaters cannot be entirely excluded; however, 87Sr/86Sr of Paratethyanwaters from this time interval is largely unknown, the location of a po-tential gateway is unclear, and Moncucco mollusks show no clear rela-tion to Paratethys faunas (Vasiliev et al., 2010; Flecker et al., 2015;Harzhauser et al., 2015).

Table 487Sr/86Sr ages inferred from the marine otolith taxa. A value of 0.000007 has been added to our data to correct for NBS 987 as used in McArthur et al. (2001).
Sample Taxon 87Sr/86Sr 2 sigma error NBS correction Age Min. Max. Error
OT-1 Diaphus spp. 0.708889 0.000004 0.708896 9.99 9.86 10.12 1.03OT-2 Diaphus spp. 0.708939 0.000004 0.708946 7.27 7.13 7.48 1.59OT-4 Hygophum spp. 0.708916 0.000004 0.708923 8.82 8.56 9.05 1.83OT-5 Gadiculus labiatus 0.708940 0.000005 0.708947 7.22 7.08 7.41 1.59
76 P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
5.3. Estuarine Lago Mare gastropods
The presence ofmarine fishes can only be explained by a connectionof the TPB to an open marine environment, most likely via the Adriaticbasin, which in turn implies a marginal marine habitat for themollusks.Mollusk 87Sr/86Sr values well below open ocean values (see Section 5.2)suggest substantial input of isotopically light river water into the TPB,characterizing the environment as an estuary. Based on the distributionof upper Messinian conglomerates, rivers most likely entered the basinfrom the west and the south (e.g., Ghibaudo et al., 1985) with catch-ment areas in Mesozoic carbonates and ophiolites (87Sr/86Sr =0.7070–0.7088; Faure et al., 1978; Irace et al., 2009; Marchina, 2015).Strontium isotopes further indicate oligo- to mesohaline conditions.87Sr/86Sr is most sensitive to changes in salinity b 5 psu while it ishard to distinguish an estuarine signal from the open marinesignal N 10 psu (Ingram and Sloan, 1992; Phillis et al., 2011). A compar-ison of the average offset between otolith (marine) andmollusk (brack-ish) 87Sr/86Sr of 0.000151, and the narrow range (Δ87Sr/86Srmax/min =0.00006) of the mollusk data to estuarine mixing models suggests thatsalinities at Moncucco most likely ranged between 5–10 psu (Philliset al., 2011).
Sr/Ca levels are typically depleted in freshwater systems comparedto marine systems, and paleo-Sr/Cawater can be estimated through thepartition coefficient (DSr) determined from recent mollusk carbonate(Rosenthal and Katz, 1989; Brown and Severin, 2009). As DSr varies be-tween taxa, we applied the values of Melanopsis spp. and Amnicolaintegra, the closest relatives to our taxa, for calculations (Treese et al.,1981; Rosenthal and Katz, 1989). Reconstructed Sr/Cawater for three
Fig. 5. Comparison of 87Sr/86Sr data for theMediterranean and the global ocean between 5and 7.5Ma. TheMessinian Salinity Crisis (MSC) and its different stages (including the LagoMare/LM) are clearly recognized by depleted values. Note the particularly wide spread ofdata during the Lago Mare stage. Circles refer to fossils (mollusks, ostracods, fish,foraminifera; dark grey: new data; light grey, white: data of previous studies). The rangeof data from evaporitic deposits are indicated for each MSC stage by shaded boxes.MO= mollusk samples of the present study; OT = otolith samples of the present study.The compilation of data is based on Roveri et al. (2014a) and Flecker et al. (2015).Maximum error for individual data-points of the compiled data-set is c. ± 0.000035(indicated at the bottom left corner; see Flecker et al., 2015).
species ranges from 3.3–7.9 mmol/mol (Table 5). These values exceedtypical values for lakes and rivers but fall below euhaline and hypersa-line values (Surge and Lohmann, 2002; Brown and Severin, 2009;Gillanders andMunro, 2012). However, they rangewithin typical valuesof estuaries, and their high variability is indicative of fluctuating oligo-to mesohaline conditions and mixing of different water masses(Gillanders andMunro, 2012). OnlyMelanopsis spp. exceed these values(9.4–10.9 mmol/mol) if calculated from the genus-specific DSr. The ele-vated Sr/Ca values might be explained by the episodic dissolution ofselenitic gypsum of the Vena de Gesso Formation, the local, temporaryinfluence of brines, or both (Rosenthal et al., 1989; Lugli et al., 2007).
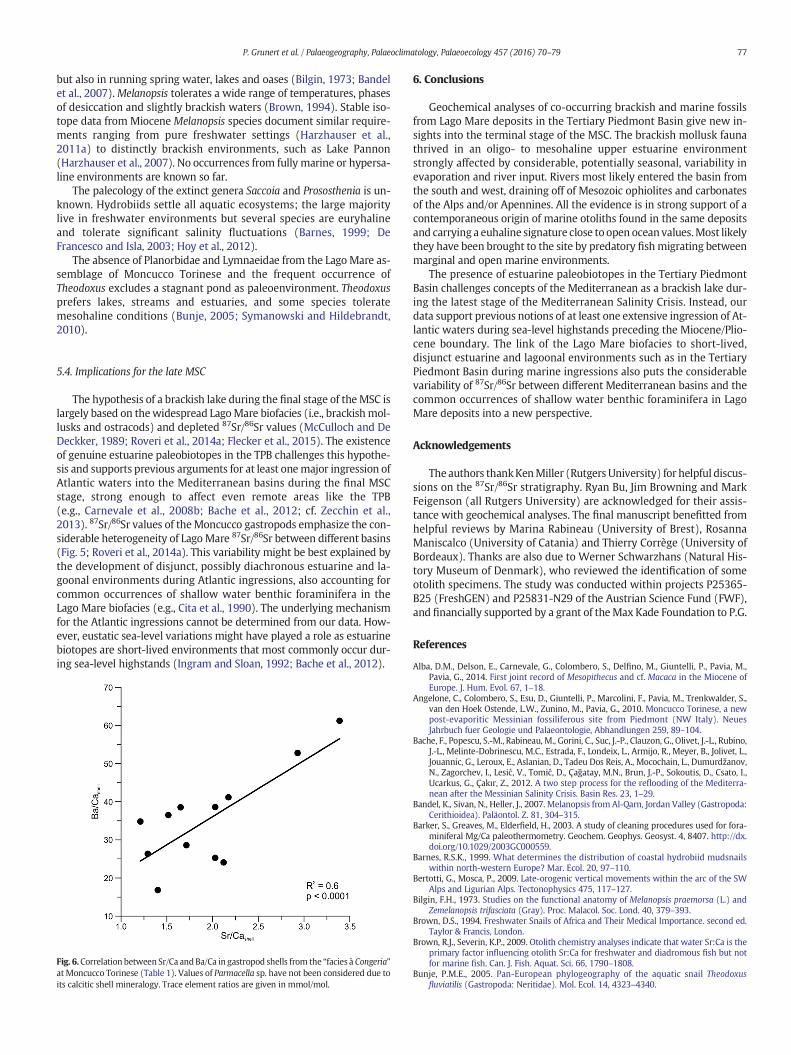
The positive correlation of Sr/Cashell and Ba/Cashell supports this in-terpretation (Fig. 6). Ba/Cawater shows ambivalent behaviorwith respectto salinity: while negatively correlated in mesohaline and euhaline wa-ters, positive correlations occur at salinities b 10 psu and N40 psu(Gillikin et al., 2006, 2008; Coffey et al., 1997; Gillanders and Munro,2012). Hypersaline conditions can be rejected for Moncucco from mol-lusk paleoecology and low, variable Sr/Ca values (Gillanders andMunro,2012; Harzhauser et al., 2015). In estuaries, dissolved Barium is re-moved at salinities b 10 psu, and the correlation to salinity and Sr/Cais positive (Coffey et al., 1997).
Fluctuations within this salinity range are not only indicated by var-iable Sr/Ca data but also by the stable isotope record. Due to their prox-imity to rivers, estuaries are more prominently affected by seasonalchanges in precipitation, runoff, and evaporation than other marginalmarine environments. Prominent seasonal δ18O variations in the orderof several permil also at salinities b 10 psu are well documented inMio-cene and recent estuaries (Surge and Lohmann, 2002; Harzhauser et al.,2011b; Mohan and Walther, 2015). We argue that the spread of δ18Odata in the time-averaged Moncucco taphocoenosis reflects suchchanges. To which extent each of these factors contributed to thedata-set cannot be deduced; however, the comparably small range ofδ13C and the non-existing/very weak negative correlation of δ18O andδ13C (R2 = 0.12, p = 0.63) suggests a prominent role of evaporation.δ18O variability up to 1.8‰within individual shells may also reflect sea-sonal changes; however, due to the small size of the analyzed gastro-pods, the extent of seasonal variation cannot be deducedwith certainty.
The paleoenvironmental conditions inferred from geochemistry arein agreement with the paleoecology of the analyzed taxa. A review ofthe paleobiology of the Lago Mare assemblage shows the prevalenceof euryhaline taxa (Harzhauser et al., 2015). Extant Melanopsis speciesin the circum-Mediterranean area live in freshwater streams and rivers
Table 5Calculated Sr/Ca of the original water. Sr/Ca 1 estimated from partition coefficient (DSr) ofMelanopsis spp. (DSr = 0.31; Rosenthal and Katz, 1989), Sr/Ca 2 estimated from Amnicolaintegra (DSr = 0.43; Treese et al., 1981).
Sample Taxon Shell Sr/Ca 1 Sr/Ca 2
MO5 Melanoides curvicosta 2.029 6.5 4.7MO13 Melanoides curvicosta 2.121 6.8 4.9MO15 Melanopsis fusulatina 2.929 9.4 6.8MO19 Melanopsis narzolina 3.388 10.9 7.9MO23 Saccoia globosa 1.401 4.5 3.3MO25 Saccoia globosa 1.517 4.9 3.5MO2 Saccoia sp. 1 1.714 5.5 4.0MO33 Prososthenia sp. 2.029 6.5 4.7MO34 Prososthenia sp. 1.651 5.3 3.8MO35 Prososthenia sp. 2.174 7.0 5.1
77P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
but also in running spring water, lakes and oases (Bilgin, 1973; Bandelet al., 2007).Melanopsis tolerates a wide range of temperatures, phasesof desiccation and slightly brackish waters (Brown, 1994). Stable iso-tope data from Miocene Melanopsis species document similar require-ments ranging from pure freshwater settings (Harzhauser et al.,2011a) to distinctly brackish environments, such as Lake Pannon(Harzhauser et al., 2007). No occurrences from fully marine or hypersa-line environments are known so far.
The palecology of the extinct genera Saccoia and Prososthenia is un-known. Hydrobiids settle all aquatic ecosystems; the large majoritylive in freshwater environments but several species are euryhalineand tolerate significant salinity fluctuations (Barnes, 1999; DeFrancesco and Isla, 2003; Hoy et al., 2012).
The absence of Planorbidae and Lymnaeidae from the Lago Mare as-semblage of Moncucco Torinese and the frequent occurrence ofTheodoxus excludes a stagnant pond as paleoenvironment. Theodoxusprefers lakes, streams and estuaries, and some species toleratemesohaline conditions (Bunje, 2005; Symanowski and Hildebrandt,2010).
5.4. Implications for the late MSC
The hypothesis of a brackish lake during the final stage of theMSC islargely based on thewidespread LagoMare biofacies (i.e., brackishmol-lusks and ostracods) and depleted 87Sr/86Sr values (McCulloch and DeDeckker, 1989; Roveri et al., 2014a; Flecker et al., 2015). The existenceof genuine estuarine paleobiotopes in the TPB challenges this hypothe-sis and supports previous arguments for at least onemajor ingression ofAtlantic waters into the Mediterranean basins during the final MSCstage, strong enough to affect even remote areas like the TPB(e.g., Carnevale et al., 2008b; Bache et al., 2012; cf. Zecchin et al.,2013). 87Sr/86Sr values of theMoncucco gastropods emphasize the con-siderable heterogeneity of LagoMare 87Sr/86Sr between different basins(Fig. 5; Roveri et al., 2014a). This variability might be best explained bythe development of disjunct, possibly diachronous estuarine and la-goonal environments during Atlantic ingressions, also accounting forcommon occurrences of shallow water benthic foraminifera in theLago Mare biofacies (e.g., Cita et al., 1990). The underlying mechanismfor the Atlantic ingressions cannot be determined from our data. How-ever, eustatic sea-level variations might have played a role as estuarinebiotopes are short-lived environments that most commonly occur dur-ing sea-level highstands (Ingram and Sloan, 1992; Bache et al., 2012).
Fig. 6.Correlation between Sr/Ca andBa/Ca in gastropod shells from the “facies à Congeria”at Moncucco Torinese (Table 1). Values of Parmacella sp. have not been considered due toits calcitic shell mineralogy. Trace element ratios are given in mmol/mol.
6. Conclusions
Geochemical analyses of co-occurring brackish and marine fossilsfrom Lago Mare deposits in the Tertiary Piedmont Basin give new in-sights into the terminal stage of the MSC. The brackish mollusk faunathrived in an oligo- to mesohaline upper estuarine environmentstrongly affected by considerable, potentially seasonal, variability inevaporation and river input. Rivers most likely entered the basin fromthe south and west, draining off of Mesozoic ophiolites and carbonatesof the Alps and/or Apennines. All the evidence is in strong support of acontemporaneous origin of marine otoliths found in the same depositsand carrying a euhaline signature close to open ocean values.Most likelythey have been brought to the site by predatory fishmigrating betweenmarginal and open marine environments.
The presence of estuarine paleobiotopes in the Tertiary PiedmontBasin challenges concepts of the Mediterranean as a brackish lake dur-ing the latest stage of the Mediterranean Salinity Crisis. Instead, ourdata support previous notions of at least one extensive ingression of At-lantic waters during sea-level highstands preceding the Miocene/Plio-cene boundary. The link of the Lago Mare biofacies to short-lived,disjunct estuarine and lagoonal environments such as in the TertiaryPiedmont Basin during marine ingressions also puts the considerablevariability of 87Sr/86Sr between different Mediterranean basins and thecommon occurrences of shallow water benthic foraminifera in LagoMare deposits into a new perspective.
Acknowledgements
The authors thankKenMiller (Rutgers University) for helpful discus-sions on the 87Sr/86Sr stratigraphy. Ryan Bu, Jim Browning and MarkFeigenson (all Rutgers University) are acknowledged for their assis-tance with geochemical analyses. The final manuscript benefitted fromhelpful reviews by Marina Rabineau (University of Brest), RosannaManiscalco (University of Catania) and Thierry Corrège (University ofBordeaux). Thanks are also due to Werner Schwarzhans (Natural His-tory Museum of Denmark), who reviewed the identification of someotolith specimens. The study was conducted within projects P25365-B25 (FreshGEN) and P25831-N29 of the Austrian Science Fund (FWF),and financially supported by a grant of theMax Kade Foundation to P.G.
References
Alba, D.M., Delson, E., Carnevale, G., Colombero, S., Delfino, M., Giuntelli, P., Pavia, M.,Pavia, G., 2014. First joint record of Mesopithecus and cf. Macaca in the Miocene ofEurope. J. Hum. Evol. 67, 1–18.
Angelone, C., Colombero, S., Esu, D., Giuntelli, P., Marcolini, F., Pavia, M., Trenkwalder, S.,van den Hoek Ostende, L.W., Zunino, M., Pavia, G., 2010. Moncucco Torinese, a newpost-evaporitic Messinian fossiliferous site from Piedmont (NW Italy). NeuesJahrbuch fuer Geologie und Palaeontologie, Abhandlungen 259, 89–104.
Bache, F., Popescu, S.-M., Rabineau, M., Gorini, C., Suc, J.-P., Clauzon, G., Olivet, J.-L., Rubino,J.-L., Melinte-Dobrinescu, M.C., Estrada, F., Londeix, L., Armijo, R., Meyer, B., Jolivet, L.,Jouannic, G., Leroux, E., Aslanian, D., Tadeu Dos Reis, A., Mocochain, L., Dumurdžanov,N., Zagorchev, I., Lesić, V., Tomić, D., Çağatay, M.N., Brun, J.-P., Sokoutis, D., Csato, I.,Ucarkus, G., Çakır, Z., 2012. A two step process for the reflooding of the Mediterra-nean after the Messinian Salinity Crisis. Basin Res. 23, 1–29.
Bandel, K., Sivan, N., Heller, J., 2007. Melanopsis from Al-Qarn, Jordan Valley (Gastropoda:Cerithioidea). Paläontol. Z. 81, 304–315.
Barker, S., Greaves, M., Elderfield, H., 2003. A study of cleaning procedures used for fora-miniferal Mg/Ca paleothermometry. Geochem. Geophys. Geosyst. 4, 8407. http://dx.doi.org/10.1029/2003GC000559.
Barnes, R.S.K., 1999. What determines the distribution of coastal hydrobiid mudsnailswithin north-western Europe? Mar. Ecol. 20, 97–110.
Bertotti, G., Mosca, P., 2009. Late-orogenic vertical movements within the arc of the SWAlps and Ligurian Alps. Tectonophysics 475, 117–127.
Bilgin, F.H., 1973. Studies on the functional anatomy of Melanopsis praemorsa (L.) andZemelanopsis trifasciata (Gray). Proc. Malacol. Soc. Lond. 40, 379–393.
Brown, D.S., 1994. Freshwater Snails of Africa and Their Medical Importance. second ed.Taylor & Francis, London.
Brown, R.J., Severin, K.P., 2009. Otolith chemistry analyses indicate that water Sr:Ca is theprimary factor influencing otolith Sr:Ca for freshwater and diadromous fish but notfor marine fish. Can. J. Fish. Aquat. Sci. 66, 1790–1808.
Bunje, P.M.E., 2005. Pan-European phylogeography of the aquatic snail Theodoxusfluviatilis (Gastropoda: Neritidae). Mol. Ecol. 14, 4323–4340.

78 P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
Butler, R.W.H., McCelland, E., Jones, R.E., 1999. Calibrating the duration and timing of theMessinian salinity crisis in the Mediterranean: linked tectonoclimatic signals inthrust-top basins of Sicily. J. Geol. Soc. 156, 827–835.
Caputo, D., Carnevale, G., Landini, W., 2009. Fish otoliths from the Messinian of Stradadegli Archi (Tuscany, Italy) – Taxonomy and palaeoecology. Ann. Naturhist. Mus.Wien 111A, 257–280.
Carnevale, G., Caputo, D., Landini, W., 2006a. Late Miocene fish otoliths from theColombacci Formation (Northern Apennines, Italy): implications for the Messinian“Lago-mare” event. Geol. J. 41, 537–555.
Carnevale, G., Landini, W., Sarti, G., 2006b. Mare versus Lago-mare: marine fishes and theMediterranean environment at the end of the Messinian Salinity Crisis. J. Geol. Soc.Lond. 163, 75–80.
Carnevale, G., Caputo, D., Landini, W., 2008a. A leerfish (Teleostei, Carangidae) from theMessinian evaporites of the Vena del Gesso basin (Romagna Apennines, Italy): paleo-geographical and paleoecological implications. Boll. Soc. Paleontol. Ital. 47, 169–176.
Carnevale, G., Longinelli, A., Caputo, D., Barbieri, M., Landini, W., 2008b. Did the Mediter-ranean marine reflooding precede the Mio-Pliocene boundary? Paleontological andgeochemical evidence from upper Messinian sequences of Tuscany, Italy.Palaeogeogr. Palaeoclimatol. Palaeoecol. 257, 81–105.
Cita, M.B., Santambrogio, S., Melillo, B., Rogate, F., 1990. Messinian paleoenvironments:new evidence from the Tyrrhenian sea (ODP Leg 107). In: Kastens, K.A., Mascle, J.,et al. (Eds.), Proceedings Ocean Drilling Program. Scientific Results. 107, pp. 211–227.
Coffey, M., Dehairs, F., Colette, O., Luther, G., Church, T., Jickells, T., 1997. The behaviour ofdissolved barium in estuaries. Estuar. Coast. Shelf Sci. 45, 113–121.
Cohen, D., Inada, T., Iwamoto, T., Scialabba, N., 1990. Gadiform fishes of the world (OrderGadiformes). An annotated and illustrated catalogue of cods, hakes, grenadiers andother gadiform fishes known to date. FAO Fish. Synop. 125 (10), 1–442.
Colombero, S., Pavia, G., Carnevale, G., 2014. Messinian rodents from Moncucco Torinese,NW Italy: palaeobiodiversity and biochronology. Geodiversitas 36, 5–59.
Colombero, S., Pavia, M., Carnevale, G., 2015. OldWorld porcupine (Rodentia, Hystricidae)remains from the late Messinian of Piedmont, NW Italy. Riv. Ital. Paleontol. Stratigr.121, 243–253.
De Francesco, C.G., Isla, F.I., 2003. Distribution and abundance of hydrobiid snails in amixed estuary and a coastal lagoon, Argentina. Estuaries 26, 790–797.
Dela Pierre, F., Bernardi, E., Cavagna, S., Clari, P., Gennari, R., Irace, A., Lozar, F., Lugli, S.,Manzi, V., Natalicchio, M., Roveri, M., Violanti, D., 2011. The record of the Messiniansalinity crisis in the Tertiary Piedmont Basin (NW Italy): the Alba section revisited.Palaeogeogr. Palaeoclimatol. Palaeoecol. 310, 238–255.
Faure, G., Assereto, R., Tremba, E.L., 1978. Strontium isotope composition of marine car-bonates of Middle Triassic to Early Jurassic age, Lombardic Alps, Italy. Sedimentology25, 523–543.
Flecker, R., de Villiers, S., Ellam, R.M., 2002. Modelling the effect of evaporation on thesalinity-87Sr/86Sr relationship in modern and ancient marginal-marine systems: theMediterranean Messinian Salinity Crisis. Earth Planet. Sci. Lett. 203, 221–233.
Flecker, R., Krijgsman, W., Capella, W., de Castro Martíns, C., Dmitrieva, E., Mayser, J.P.,Marzocchi, A., Modestu, S., Lozano Ochoa, D., Simon, D., Tulbure, M., van den Berg,B., van der Schee, M., de Lange, G., Ellam, R., Govers, R., Gutjahr, M., Hilgen, F.,Kouwenhoven, T., Lofi, J., Meijer, P., Sierro, F.J., Bachiri, N., Barhoun, N., Alami, A.C.,Chacon, B., Flores, J.A., Gregory, J., Howard, J., Lunt, D., Ochoa, M., Pancost, R.,Vincent, S., Yousfi, M.Z., 2015. Evolution of the Late Miocene Mediterranean-Atlantic gateways and their impact on regional and global environmental change.Earth-Sci. Rev. http://dx.doi.org/10.1016/j.earscirev.2015.08.007.
Ghibaudo, G., Clari, P., Perello, M., 1985. Litostratigrafia, sedimentologia ed evoluzionetettonico-sedimentaria dei depositi miocenici del margine sud-orientale del BacinoTerziario Ligure-Piemontese (Valli Borbera, Scrivia e Lemme). Boll. Soc. Geol. Ital.104, 349–397.
Gillanders, B.M., Munro, A.R., 2012. Hypersaline waters pose new challenges forreconstructing environmental histories of fish based on otolith chemistry. Limnol.Oceanogr. 57, 1136–1148.
Gillikin, D.P., Dehairs, F., Lorrain, A., Steenmans, D., Baeyens, W., Anré, L., 2006. Barium up-take into the shells of the common mussel (Mytilus edulis) and the potential for estu-arine paleo-chemistry reconstruction. Geochim. Cosmochim. Acta 70, 395–407.
Gillikin, D.P., Lorrain, A., Paulet, Y.-M., André, L., Dehairs, F., 2008. Synchronous bariumpeaks in high-resolution profiles of calcite and aragonite marine bivalve shells.Geo-Mar. Lett. 28, 351–358.
Gradstein, F., Ogg, J., Schmitz, M., Ogg, G., 2012. The Geologic Time Scale 2012. Elsevier,Amsterdam (1144 p).
Hart, S.R., Brooks, C., 1974. Clinopyroxene-matrix partitioning of K, Rb, Cs, Sr and Ba.Geochim. Cosmochim. Acta 38, 1799–1806.
Harzhauser, M., Latal, C., Piller, W.E., 2007. The stable isotope archive of Lake Pannon as amirror of Late Miocene climate change. Palaeogeogr. Palaeoclimatol. Palaeoecol. 249,335–350.
Harzhauser, M., Mandic, O., Latal, C., Kern, A., 2011a. Stable isotope composition of theMiocene Dinaride Lake System deduced from its endemic mollusc fauna.Hydrobiologia 682, 27–46.
Harzhauser, M., Piller, W.E., Müllegger, S., Grunert, P., 2011b. Changing seasonality pat-terns in Central Europe from Miocene Climate Optimum to Miocene Climate Transi-tion deduced from the Crassostrea isotope archive. Glob. Planet. Chang. 76 (1–2),77–84.
Harzhauser, M., Neubauer, T.A., Georgopoulou, E., Esu, D., D'Amico, C., Pavia, G., Giuntelli,P., Carnevale, G., 2015. Late Messinian continental and Lago-Mare gastropods fromthe Tertiary Piedmont Basin, NW Italy. Boll. Soc. Paleontol. Ital. 54, 1–53.
Hernández-Molina, F.J., Stow, D.A.V., Alvarez Zarikian, C.A., Acton, G., Bahr, A., Balestra, B.,Ducassou, E., Flood, R.D., Flores, J.-A., Furota, S., Grunert, P., Hodell, D., Jimenez-Espejo,F., Kim, J.K., Krissek, L., Kuroda, J., Li, B., Llave, E., Lofi, J., Lourens, L., Miller, M.,Nanayama, F., Nishida, N., Richter, C., Roque, A.C., Sanchez Goñi, M.F., Sierro, F.J.,
Singh, A.D., Sloss, C., Takashimizu, Y., Tzanova, A., Voelker, A., Williams, T., Xuan, C.,2014. Onset of Mediterranean outflow into the North Atlantic. Science 344,1244–1250.
Hoy, M., Boese, B.L., Taylor, L., Reusser, D., Rodriguez, R., 2012. Salinity adaptation of theinvasive New Zealand mud snail (Potamopyrgus antipodarum) in the ColumbiaRiver estuary (Pacific Northwest, USA): physiological and molecular studies. Aquat.Ecol. 46, 249–260.
Ingram, B.L., Sloan, D., 1992. Strontium isotopic composition of estuarine sediments aspaleosalinity-paleoclimate indicator. Science 255, 68–72.
Irace, A., Clemente, P., Nataliccho, M., Ossella, L., Trenkwalder, S., De Luca, D.A., Mosca, P.,Piana, F., Polino, R., Violanti, D., 2009. Geologia e idrostratigrafia profonda dellapianura padana occidentale. La Nuova Lito, Firenze (126 pages).
Irace, A., Clemente, P., Piana, F., De Luca, D.A., Polino, R., Violanti, D., Mosca, P.,Trenkwalder, S., Natalicchio, M., Ossella, L., Governa, M., Petricig, M., 2010.Hydrostratigraphy of the late Messinian-Quaternary basins in southern Piedmont(northwestern Italy). Memorie Descrittive della Carta Geologica d'Italia 90, 133–152.
Kalish, J.M., 1991. 13C and 18O isotopic disequilibria in fish otoliths: metabolic and kineticeffects. Mar. Ecol. Prog. Ser. 75, 191–203.
Krijgsman, W., Gaboardi, S., Hilgen, F.J., Iaccarino, S., de Kaenel, E., van der Laan, E., 2004.Revised astrochronology for the Ain el Beida section (Atlantic Morocco): no glacio-eustatic control for the onset of the Messinian Salinity Crisis. Stratigraphy 1, 87–101.
Lécuyer, C., Hutzler, A., Amiot, R., Daux, V., Grosheny, D., Otero, O., Martineau, F., Fourel, F.,Balter, V., Reynard, B., 2012. Carbon and oxygen isotope fractionations between ara-gonite and calcite of shells from modern molluscs. Chem. Geol. 332-333, 92–101.
Lin, H.-Y., Shiao, J.-C., Chen, Y.-G., Iizuka, Y., 2012. Ontogenetic vertical migration of gren-adiers revealed by otolith microstructures and stable isotopic composition. Deep-SeaRes. I 61, 123–130.
Lourens, L., Hilgen, F., Shackleton, N.J., Laskar, J., Wilson, D., 2004. The Neogene Period. In:Gradstein, F.M., Ogg, J.G., Smith, A.G. (Eds.), A Geologic Time Scale 2004. CambridgeUniversity Press, Cambridge, pp. 409–440.
Lozar, F., Violanti, D., Dela Pierre, F., Bernardi, E., Cavagna, S., Clari, P., Irace, A., Martinetto, E.,Trenkwalder, S., 2010. Calcareous nannofossils and foraminifers herald the MessinianSalinity Crisis: the Pollenzo section (Alba, Cuneo; NW Italy). Geobios 43, 21–32.
Lugli, S., Bassetti, M.A., Manzi, V., Barbieri, M., Longinelli, A., Roveri, M., 2007. TheMessinian “Vena del Gesso” evaporites revisited: characterization of isotopic compo-sition and organic matter. In: Schreiber, B.C., Lugli, S., Babel, M. (Eds.), EvaporitesThrough Space and Time. Geological Society, London, Special Publications Vol. 285,pp. 179–190.
Maino, M., Decarlis, A., Felletti, F., Seno, S., 2013. Tectono-sedimentary evolution of theTertiary Piedmont Basin (NW Italy) within the Oligo-Miocene central Mediterraneangeodynamics. Tectonics 32, 593–619.
Manzi, V., Gennari, R., Hilgen, F., Krijgsman, W., Lugli, S., Roveri, M., Sierro, F.J., 2013. Agerefinement of theMessinian salinity crisis onset in the Mediterranean. Terra Nova 25,315–322.
Marchina, C., 2015. Geochemical and Isotopic Investigation on the Po River Waters FromMonviso Sources to Its Delta: Natural and Anthropogenic Components (Ph.D. thesis,Università degli Studi di Ferrara. 117 pages).
Maul, G.E., 1990. Trachichthyidae. In: Quéro, J.C., Hureau, J.C., Karrer, C., Post, A., Saldanha,L. (Eds.), Check-list of the Fishes of the Eastern Tropical Atlantic (CLOFETA). UNESCO,Paris. vol. 2, pp. 620–622.
McArthur, J.M., Howarth, R.J., Bailey, T.R., 2001. Strontium isotope stratigraphy: LOWESSversion 3: best fit to the marine Sr-isotope curve for 0–509 Ma and accompanyinglook-up table for deriving numerical age. J. Geol. 109, 155–170.
McCulloch, M.T., De Deckker, P., 1989. Sr isotope constraints on the Mediterranean envi-ronment at the end of the Messinian salinity crisis. Nature 342, 62–65.
Mezger, E., 2012. HowDryWas theMessinian Salinity Crisis? AMolecular BiogeochemicalStudy of the Eraclea Minoa (Sicily) Section, Italy (Master thesis) University of Utrecht(34 pages).
Mohan, J.A., Walther, B.D., 2015. Spatiotemoral variation of trace elements and stable iso-topes in subtropical estuaries: II. Regional, local, and seasonal salinity-element rela-tionships. Estuar. Coasts 38, 769–781.
Müller, D.W., Mueller, P.A., 1991. Origin and age of the Mediterranean Messinian evapo-rites: implications from Sr isotopes. Earth Planet. Sci. Lett. 107, 1–12.
Nafpaktitis, B.G., Backus, R.H., Craddock, J.E., Haedrich, R.L., Robinson, B.H., Karnella, C.,1977. Family Myctophidae. In: Gibbs, R.H., Berry, F.H., Böhlke, J.E., Cohen, D.M.,Collette, B.B., Eschmeyer, W.N., Mead, G.W., Merriman, D., Pietsch, T.W. (Eds.), Fishesof the Western North Atlantic. Part Seven. Order Iniomi (Myctophyformes), SearsFoundation for Marine Research, Yale University, pp. 13–265.
Neubauer, T.A., Harzhauser, M., Kroh, A., Georgopoulou, E., Mandic, O., 2015. A gastropod-based biogeographic scheme for the European Neogene freshwater systems. EarthSci. Rev. 143, 98–116. http://dx.doi.org/10.1016/j.earscirev.2015.01.010.
Nolf, D., 1985. Otolithi Piscium. Handb. Paleoichthyology 10, 1–145.Orszag-Sperber, F., 2006. Changing perspectives in the concept of “LagoMare” inMediter-
ranean Late Miocene evolution. Sediment. Geol. 188-189, 259–277.Phillis, C.C., Ostrach, D.J., Ingram, B.L., Weber, P.K., 2011. Evaluating otolith Sr/Ca as a tool
for reconstructing estuarine habitat use. Can. J. Fish. Aquat. Sci. 68, 360–373.Reuter, M., Brachert, T.C., Grunert, P., 2011. Fossil round karren from Cyprus: a latest Mio-
cene record of a humid-temperate climate in the eastern Mediterranean. Facies 57,39–50.
Rosenthal, Y., Katz, A., 1989. The applicability of trace elements in freshwater shells forpaleogeochemical studies. Chem. Geol. 78, 65–76.
Rosenthal, Y., Katz, A., Tchernov, E., 1989. The reconstruction of quaternary freshwaterlakes from the chemical and isotopic composition of gastropod shells: the Dead SeaRift, Israel. Palaeogeogr. Palaeoclimatol. Palaeoecol. 74, 241–253.
Rossi, M., Mosca, P., Polino, R., Rogledi, S., Biffi, U., 2009. New outcrop and subsurface datain the Tertiary Piedmont Basin (NW-Italy): unconformity-bounded stratigraphic

79P. Grunert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 457 (2016) 70–79
units and their relationships with basin-modification phases. Riv. Ital. Paleontol.Stratigr. 115, 305–335.
Rouchy, J.M., Caruso, A., 2006. The Messinian salinity crisis in the Mediterranean basin: areassessment of the data and an integrated scenario. Sediment. Geol. 188-189, 35–67.
Roveri, M., Flecker, R., Krijgsman, W., Lofi, J., Lugli, S., Manzi, V., Sierro, F.J., Bertini, A.,Camerlenghi, A., De Lange, G., Gover, R., Hilgen, F.J., Hübscher, C., Meijer, P.T., Stoica,M., 2014a. The Messinian Salinity Crisis: past and future of a great challenge for ma-rine sciences. Mar. Geol. 352, 25–58.
Roveri, M., Manzi, V., Bergamasco, A., Falcieri, F.M., Gennari, R., Lugli, S., Schreiber, B.C.,2014b. Dense shelf water cascading and Messinian canyons: a new scenario for theMediterranean Salinity Crisis. Am. J. Sci. 314, 751–784.
Schäfer, W., 1966. Aktuopaläontologische Beobachtungen 6. Otolithen-Anreicherungen.Nat. Mus. 69, 439–444.
Schildgen, T.F., Cosentino, D., Frijia, G., Castorina, F., Dudas, F.Ö., Iadanza, A., Sampalmieri,G., Cipollari, P., Caruso, A., Bowring, S.A., Strecker, M.R., 2014. Sea level and climateforcing of the Sr isotope composition of late Miocene Mediterranean marine basins.Geochem. Geophys. Geosyst. http://dx.doi.org/10.1002/2014GC005332.
Sturani, C., 1975. Messinian facies in the Piedmont Basin. Mem. Soc. Geol. Ital. 16, 11–25.Surge, D.M., Lohmann, K.C., 2002. Temporal and spatial differences in salinity and water
chemistry in SW Florida estuaries: effects of human-impacted watersheds. Estuaries25, 393–408.
Symanowski, F., Hildebrandt, J.-P., 2010. Differences in osmotolerance in freshwater andbrackish water populations of Theodoxus fluviatilis (Gastropoda: Neritidae) are asso-ciated with differential protein expression. J. Comp. Physiol. B. 180, 337–346.
Thorrold, S.R., Campana, S.E., Jones, C.M., Swart, P.K., 1997. Factors determining δ13C andδ18O fractionation in aragonitic otoliths of marine fish. Geochim. Cosmochim. Acta 61,2909–2919.
Topper, R.P.M., Flecker, R., Meijer, P.T., Wortel, M.J.R., 2011. A box model of the Late Mio-cene Mediterranean Sea: implications from combined 87Sr/86Sr and salinity data.Paleoceanography 26, PA 3223.
Treese, T.N., Owen, R.M., Wilkinson, B.H., 1981. Sr/Ca andMg/Ca ratios in polygenetic car-bonate allochems from a Michigan marl lake. Geochim. Cosmochim. Acta 45,439–445.
Trenkwalder, S., Violanti, D., D'Atri, A., Lozar, F., Dela Pierre, F., Irace, A., 2008. The Mio-cene/Pliocene boundary in the Early Pliocene micropalaeontological record: newdata from the Tertiary Piedmont Basin (Moncucco quarry, Torino Hill, northwesternItaly). Boll. Soc. Paleontol. Ital. 47, 87–103.
Van der Schee, M., Sierro, F.J., Jiménez-Espejo, F.J., Hernández-Molina, F.J., Flecker, R., Flo-res, J.A., Acton, G., Gutjahr, M., Grunert, P., García-Gallardo, Á., Andersen, N., Evidenceof early bottom water current flow after the Messinian salinity crisis in the Gulf ofCadiz. Mar. Geol., http://dx.doi.org/10.1016/j.margeo.2016.04.005 (in press).
Vasiliev, I., Reichart, G.-J., Davies, G.R., Krijgsman, W., Stoica, M., 2010. Strontium isotoperatios of the Eastern Paratethys during the Mio-Pliocene transition: implications forinterbasinal connectivity. Earth Planet. Sci. Lett. 292, 123–131.
Violanti, D., Dela Pierre, F., Trenkwalder, S., Lozar, F., Clari, P., Irace, A., D'Atri, A., 2011. Bio-stratigraphic and palaeoenvironmental analyses of the Messinian/Zanclean boundaryand Zanclean succession in the Moncucco quarry (Piedmont, northwestern Italy).Bull. Soc. Geol. Fr. 182, 149–162.
Zecchin, M., Caffau, M., Di Stefano, A., Maniscalco, R., Lenaz, D., Civile, D., Muto, F., Critelli,S., 2013. The Messinian succession of the Crotone Basin (southern Italy) II: facies ar-chitecture and stratal surfaces across the Miocene-Pliocene boundary. Mar. Pet. Geol.48, 474–492.
Related Documents











