Palaeoenvironmental evolution of the ancient lagoon of Ostia Antica (Tiber delta, Italy) Cecile Vittori a, * , Ilaria Mazzini b , Ferr eol Salomon c , Jean-Philippe Goiran a , Simona Pannuzi d , Carlo Rosa e , Angelo Pellegrino d a Maison de l'Orient et de la M editerran ee, CNRS Unit e Mixte de Recherche 5133, 69365, Lyon Cedex 7, France b IGAG CNR Area della Ricerca di Roma1 e Montelibretti, Via Salaria Km 29,300, Monterotondo RM, Italy c Department of Archaeology, University of Southampton, Southampton, United Kingdom d MIBACT, Istituto Superiore per la Conservazione e il Restauro, Roma, Italy e Istituto Italiano di Paleontologia Umana, Roma, Italy article info Article history: Available online 23 August 2014 Keywords: Geoarchaeology Ostracodology Palaeoenvironment Ostia lagoon Tiber delta Abstract The ancient lagoon of Ostia played a key role in the development of the cultures of central Italy and especially that of Rome. The evolution of its environmental history and its partial transformation into a saline body of water during Antiquity remain as poorly known to archaeologists and historians as to geologists and geographers. A high chronostratigraphical resolution palaeoenvironmental study has been carried out on a sediment core. It is based on multi-proxy analysis and pays particular attention to os- tracods, which are very useful palaeoenvironmental indicators in marginal marine environments. Our study has unveiled five evolution phases of the landscapes in the countryside of Ostia: (1) between the 6th and the middle of the 2nd millennia BC a lagoon open to the sea was formed; (2) the lagoon was then subject to marine and fluvial influences,; (3) between the middle of the 2nd and the beginning of the 1st millennia BC, a marshland formed, indicating either a period of limited flooding or a shifting of the branch of the Tiber; (4) during the first part of the 1st millennium BC, the lagoon became again con- nected to the sea. In this area, its depth at the time of the founding of Ostia varied between 3.5 m and 4.5 m (below ancient sea level), which suggests that it could have served as a naturally sheltered place on the coast; (5) from the middle of the 1st millennia BC to the 19th century AD, the lagoon probably remained connected to the sea and progressively silted, until it disappeared in the 19th century. © 2014 Elsevier Ltd. All rights reserved. 1. Introduction Deltaic lagoons are coastal environments at the interface be- tween marine and continental dynamics. They record various coastlines process at different spatial and temporal scales (Bidet et al., 1982; Bird, 1994). Many palaeo-lagoons around the Medi- terranean coasts have been converted in salt works during histor- ical times to produce salt, an expensive commodity and necessity in the Mediterranean since the Egyptians (Delrue, 2011). The palaeo-lagoon of Ostia is one of the large lagoons that occupied the Tiber delta until programs to eliminate malaria in the 19th and 20th centuries led to land reclaim. The evolution of the lagoon environments is closely linked to the mobility of sandy barriers, sea-level variations and river and/or continental stream influences. Thus, the lagoons on the Tiber delta area can provide essential information concerning the palaeo-environments and the evolution of the dynamic of the Tiber delta. To these factors, modifications carried out to exploit and increase the salinity of the lagoon can be added as one of the first example of the impact of human industrial activity on the environment. Indeed, the Ostia palaeo-lagoon was located behind his eponymous ancient city which, according to Roman literary tradition was founded in the 7th century BC by King Ancus Marius, at the end of the salt war between the Latins of Rome and the Etruscan of the city of Veii. Thus, Titus Livius (Ab Urbe condita book I, 33, 6e9) wrote that the city was founded in order to make use of the salt works, located in the palaeo-lagoon of Ostia (Fig. 1). The ancient town of Ostia, intentionally founded near the ancient mouth of the Tiber (Tomassetti,1897), long played the role of port of supply for the city of Rome. Archaeology has brought to light its urban core, a fortress (castrum) made of large blocks of tuff, dated to the 4th century BC or beginning of the 3rd century BC (Calza et al., 1953; Zevi, 2001, * Corresponding author. Tel.: þ33 615 308 844. E-mail address: [email protected] (C. Vittori). Contents lists available at ScienceDirect Journal of Archaeological Science journal homepage: http://www.elsevier.com/locate/jas http://dx.doi.org/10.1016/j.jas.2014.06.017 0305-4403/© 2014 Elsevier Ltd. All rights reserved. Journal of Archaeological Science 54 (2015) 374e384

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Journal of Archaeological Science 54 (2015) 374e384
Contents lists avai
Journal of Archaeological Science
journal homepage: http: / /www.elsevier .com/locate/ jas
Palaeoenvironmental evolution of the ancient lagoon of Ostia Antica(Tiber delta, Italy)
Cecile Vittori a, *, Ilaria Mazzini b, Ferr�eol Salomon c, Jean-Philippe Goiran a,Simona Pannuzi d, Carlo Rosa e, Angelo Pellegrino d
a Maison de l'Orient et de la M�editerran�ee, CNRS Unit�e Mixte de Recherche 5133, 69365, Lyon Cedex 7, Franceb IGAG CNR Area della Ricerca di Roma1 e Montelibretti, Via Salaria Km 29,300, Monterotondo RM, Italyc Department of Archaeology, University of Southampton, Southampton, United Kingdomd MIBACT, Istituto Superiore per la Conservazione e il Restauro, Roma, Italye Istituto Italiano di Paleontologia Umana, Roma, Italy
a r t i c l e i n f o
Article history:Available online 23 August 2014
Keywords:GeoarchaeologyOstracodologyPalaeoenvironmentOstia lagoonTiber delta
* Corresponding author. Tel.: þ33 615 308 844.E-mail address: [email protected] (C
http://dx.doi.org/10.1016/j.jas.2014.06.0170305-4403/© 2014 Elsevier Ltd. All rights reserved.
A b s t r a c t
The ancient lagoon of Ostia played a key role in the development of the cultures of central Italy andespecially that of Rome. The evolution of its environmental history and its partial transformation into asaline body of water during Antiquity remain as poorly known to archaeologists and historians as togeologists and geographers. A high chronostratigraphical resolution palaeoenvironmental study has beencarried out on a sediment core. It is based on multi-proxy analysis and pays particular attention to os-tracods, which are very useful palaeoenvironmental indicators in marginal marine environments. Ourstudy has unveiled five evolution phases of the landscapes in the countryside of Ostia: (1) between the6th and the middle of the 2nd millennia BC a lagoon open to the sea was formed; (2) the lagoonwas thensubject to marine and fluvial influences,; (3) between the middle of the 2nd and the beginning of the 1stmillennia BC, a marshland formed, indicating either a period of limited flooding or a shifting of thebranch of the Tiber; (4) during the first part of the 1st millennium BC, the lagoon became again con-nected to the sea. In this area, its depth at the time of the founding of Ostia varied between 3.5 m and4.5 m (below ancient sea level), which suggests that it could have served as a naturally sheltered place onthe coast; (5) from the middle of the 1st millennia BC to the 19th century AD, the lagoon probablyremained connected to the sea and progressively silted, until it disappeared in the 19th century.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Deltaic lagoons are coastal environments at the interface be-tween marine and continental dynamics. They record variouscoastlines process at different spatial and temporal scales (Bidetet al., 1982; Bird, 1994). Many palaeo-lagoons around the Medi-terranean coasts have been converted in salt works during histor-ical times to produce salt, an expensive commodity and necessity inthe Mediterranean since the Egyptians (Delrue, 2011).
The palaeo-lagoon of Ostia is one of the large lagoons thatoccupied the Tiber delta until programs to eliminate malaria in the19th and 20th centuries led to land reclaim. The evolution of thelagoon environments is closely linked to the mobility of sandybarriers, sea-level variations and river and/or continental stream
. Vittori).
influences. Thus, the lagoons on the Tiber delta area can provideessential information concerning the palaeo-environments and theevolution of the dynamic of the Tiber delta. To these factors,modifications carried out to exploit and increase the salinity of thelagoon can be added as one of the first example of the impact ofhuman industrial activity on the environment. Indeed, the Ostiapalaeo-lagoon was located behind his eponymous ancient citywhich, according to Roman literary tradition was founded in the7th century BC by King Ancus Marius, at the end of the salt warbetween the Latins of Rome and the Etruscan of the city of Veii.Thus, Titus Livius (Ab Urbe condita book I, 33, 6e9) wrote that thecity was founded in order to make use of the salt works, located inthe palaeo-lagoon of Ostia (Fig. 1). The ancient town of Ostia,intentionally founded near the ancient mouth of the Tiber(Tomassetti, 1897), long played the role of port of supply for the cityof Rome. Archaeology has brought to light its urban core, a fortress(castrum) made of large blocks of tuff, dated to the 4th century BCor beginning of the 3rd century BC (Calza et al., 1953; Zevi, 2001,

Fig. 1. Location map of the Tiber delta, the studied area and position of the sediment cores.
C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384 375
2002). The close by salt marshes have been exploited since Antiq-uity and saltworks continued until 1895 (Segre in Dragone et al.,1967). The late Holocene evolution of the palaeo-lagoon of Ostiais thus closely linked to the salt marshes exploitation and to thehistory of the town of Ostia.
The lagoon of Maccarese, situated north of the Tiber, has beenthe subject of geomorphological (Belluomini et al., 1986; Bellottiet al., 1994, 2007), palaeo-environmental (Giraudi, 2002, 2004,2011) and, more recently, palynological studies (Di Rita et al.,2009). Instead, the lagoon of Ostia has only been the subject ofone publication (Bellotti et al., 2011), based on the study of somefifteen samples from three cores measuring between 3.5 and 4.5 mwhich contained sediment records dating from the second half ofthe 5th millennium BC onwards. The present study proposes totrace the environmental history of this still poorly known lagoonthrough high-resolution analysis of the sediments from a 20 mdeep borehole core. These data, integrated in a broader chro-nostratigraphy, contribute to the reconstruction of the natural dy-namics of the Tiber delta against human management actions.
2. Context of the study
2.1. Geographic and geomorphological context
The Tiber delta extends over 25 km (both submerged andemerged areas) from west to east and is bordered by Pleistoceneterraces, below which the interior deltaic plain extends (Fig. 1). Inthe past, two vast coastal lagoons, the palaeo-lagoon of Maccareseto the north and the palaeo-lagoon of Ostia to the south, occupiedthis space. They formed during the Holocene, built up by theaccumulation of sediments brought to the mouth of the Tiber riverdelta. Both lagoons were subjected to land improvement in the19th and 20th centuries. Since the 19th century, many studiesattempted to reconstruct the geomorphological history of the Tiberriver delta (Moro, 1871; Oberholtzer, 1875; Ponzi, 1875; Amenduni,1884; Bocci, 1892). But it was only in the second half of the 20thcentury that work carried out on many deep sediment cores
disclosed the stratigraphy of the deltaic plain (Dragone et al., 1967;Belluomini et al., 1986; Bellotti et al., 1989, 1994; 1995, 2007; 2011;Amorosi and Milli, 2001; Giraudi, 2002, 2004, 2011; Milli et al.,2013).
These authors, proposed the following evolution of the Tiberdelta area: (1) Between 15,000 and 8000 BP, a significant marinetransgression flooded the Pleistocene delta and the mouth of theTiber migrated eastward with the line of the coast (Bellotti et al.,2007). A protected delta was established behind a transgressivesandy barrier, which delimited a vast bay/lagoon (Milli et al., 2013).(2) About 8000e7000 BP, the increase in sea level began to slow(Lambeck et al., 2011). Then the Tiber prograded and cut the ancientbay/lagoon into two basins and thus isolated the lagoons of Mac-carese and Ostia (Bellotti et al. 2007; Milli et al., 2013). (3) Begin-ning in 6000 BP, the Tiber flowed into the sea, and three phases ofthe development of the delta, in relation to the mobility of itsmouth, followed each other: (a) between 6000 and 2700 BP, thedelta became established in the area where the harbour system ofPortus was built; (b) between 2700 and 1900 BP, after the abruptmigration of the Tiber, the delta developed more to the south(today Fiumara Grande); (c) from 1900 BP to the present time,following the construction of the channel of Fiumicino, the deltadeveloped from these two mouths.
2.2. Archaeological and historical context
In the territory of Ostia, on the left bank of the Tiber, theancient installations for salt production, consisted of two distinctbut connected basins, which the historical sources identify underthe names of “Stagno ostiense” and “Saline” (Fig. 2). The first re-fers to the southern part of the lagoon, which was linked to thesea by a canal (cf. Fossa papale). The second refers to the saltmarshes that make up the most internal zone, immediately southof the Tiber and north of the ancient Via Ostiense. The exact sizeof this vast zone has always been indicated differently on maps,from the Renaissance to the modern period (Frutaz, 1972). It waspartly defined thanks to recent archaeological excavations, which
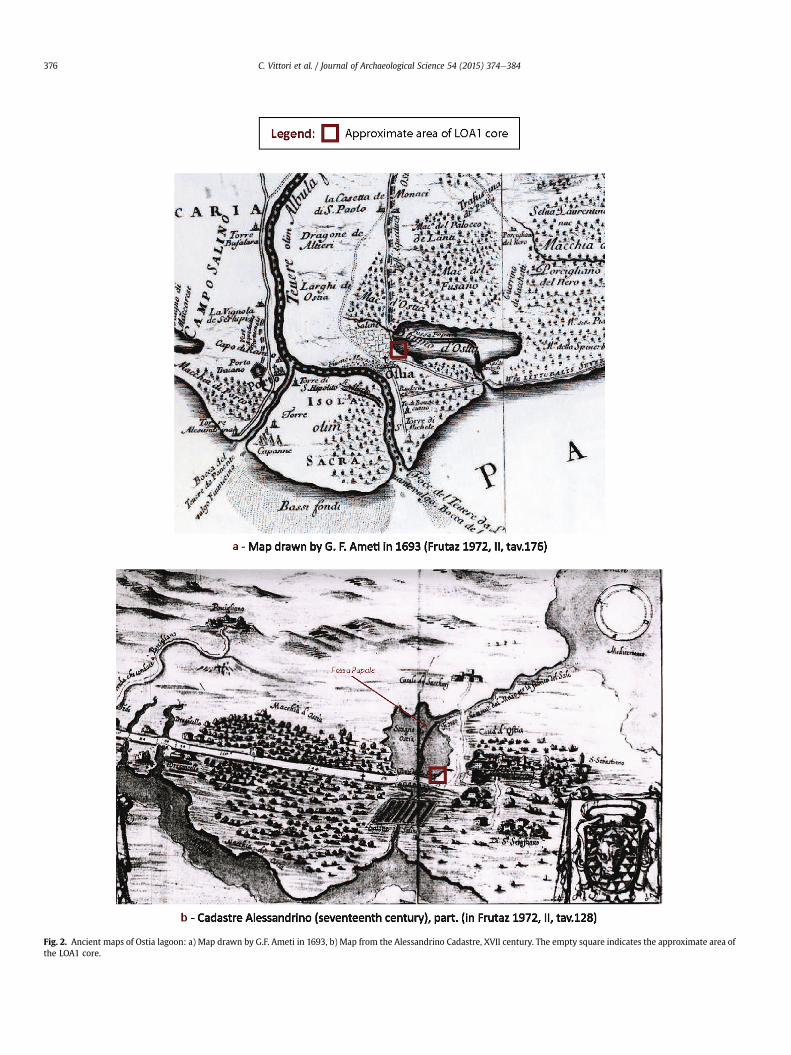
Fig. 2. Ancient maps of Ostia lagoon: a) Map drawn by G.F. Ameti in 1693, b) Map from the Alessandrino Cadastre, XVII century. The empty square indicates the approximate area ofthe LOA1 core.
C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384376
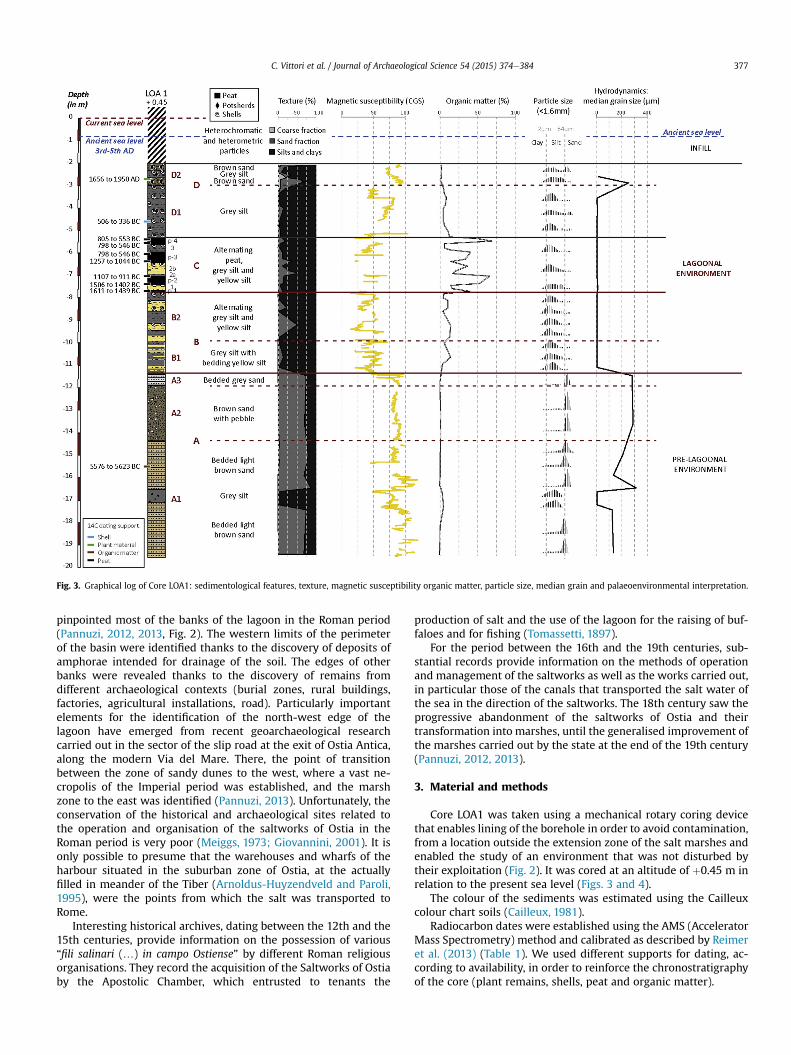
Fig. 3. Graphical log of Core LOA1: sedimentological features, texture, magnetic susceptibility organic matter, particle size, median grain and palaeoenvironmental interpretation.
C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384 377
pinpointed most of the banks of the lagoon in the Roman period(Pannuzi, 2012, 2013, Fig. 2). The western limits of the perimeterof the basin were identified thanks to the discovery of deposits ofamphorae intended for drainage of the soil. The edges of otherbanks were revealed thanks to the discovery of remains fromdifferent archaeological contexts (burial zones, rural buildings,factories, agricultural installations, road). Particularly importantelements for the identification of the north-west edge of thelagoon have emerged from recent geoarchaeological researchcarried out in the sector of the slip road at the exit of Ostia Antica,along the modern Via del Mare. There, the point of transitionbetween the zone of sandy dunes to the west, where a vast ne-cropolis of the Imperial period was established, and the marshzone to the east was identified (Pannuzi, 2013). Unfortunately, theconservation of the historical and archaeological sites related tothe operation and organisation of the saltworks of Ostia in theRoman period is very poor (Meiggs, 1973; Giovannini, 2001). It isonly possible to presume that the warehouses and wharfs of theharbour situated in the suburban zone of Ostia, at the actuallyfilled in meander of the Tiber (Arnoldus-Huyzendveld and Paroli,1995), were the points from which the salt was transported toRome.
Interesting historical archives, dating between the 12th and the15th centuries, provide information on the possession of various“fili salinari (…) in campo Ostiense” by different Roman religiousorganisations. They record the acquisition of the Saltworks of Ostiaby the Apostolic Chamber, which entrusted to tenants the
production of salt and the use of the lagoon for the raising of buf-faloes and for fishing (Tomassetti, 1897).
For the period between the 16th and the 19th centuries, sub-stantial records provide information on the methods of operationand management of the saltworks as well as the works carried out,in particular those of the canals that transported the salt water ofthe sea in the direction of the saltworks. The 18th century saw theprogressive abandonment of the saltworks of Ostia and theirtransformation into marshes, until the generalised improvement ofthe marshes carried out by the state at the end of the 19th century(Pannuzi, 2012, 2013).
3. Material and methods
Core LOA1 was taken using a mechanical rotary coring devicethat enables lining of the borehole in order to avoid contamination,from a location outside the extension zone of the salt marshes andenabled the study of an environment that was not disturbed bytheir exploitation (Fig. 2). It was cored at an altitude of þ0.45 m inrelation to the present sea level (Figs. 3 and 4).
The colour of the sediments was estimated using the Cailleuxcolour chart soils (Cailleux, 1981).
Radiocarbon dates were established using the AMS (AcceleratorMass Spectrometry) method and calibrated as described by Reimeret al. (2013) (Table 1). We used different supports for dating, ac-cording to availability, in order to reinforce the chronostratigraphyof the core (plant remains, shells, peat and organic matter).
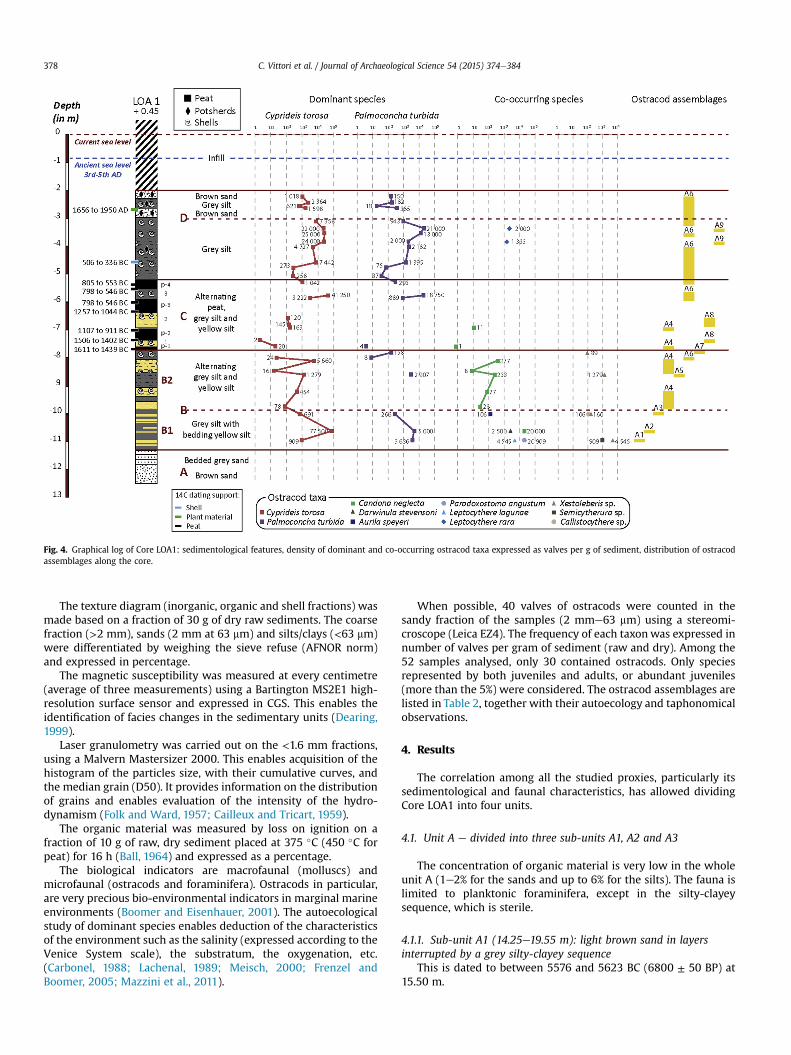
Fig. 4. Graphical log of Core LOA1: sedimentological features, density of dominant and co-occurring ostracod taxa expressed as valves per g of sediment, distribution of ostracodassemblages along the core.
C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384378
The texture diagram (inorganic, organic and shell fractions) wasmade based on a fraction of 30 g of dry raw sediments. The coarsefraction (>2 mm), sands (2 mm at 63 mm) and silts/clays (<63 mm)were differentiated by weighing the sieve refuse (AFNOR norm)and expressed in percentage.
The magnetic susceptibility was measured at every centimetre(average of three measurements) using a Bartington MS2E1 high-resolution surface sensor and expressed in CGS. This enables theidentification of facies changes in the sedimentary units (Dearing,1999).
Laser granulometry was carried out on the <1.6 mm fractions,using a Malvern Mastersizer 2000. This enables acquisition of thehistogram of the particles size, with their cumulative curves, andthe median grain (D50). It provides information on the distributionof grains and enables evaluation of the intensity of the hydro-dynamism (Folk and Ward, 1957; Cailleux and Tricart, 1959).
The organic material was measured by loss on ignition on afraction of 10 g of raw, dry sediment placed at 375 �C (450 �C forpeat) for 16 h (Ball, 1964) and expressed as a percentage.
The biological indicators are macrofaunal (molluscs) andmicrofaunal (ostracods and foraminifera). Ostracods in particular,are very precious bio-environmental indicators in marginal marineenvironments (Boomer and Eisenhauer, 2001). The autoecologicalstudy of dominant species enables deduction of the characteristicsof the environment such as the salinity (expressed according to theVenice System scale), the substratum, the oxygenation, etc.(Carbonel, 1988; Lachenal, 1989; Meisch, 2000; Frenzel andBoomer, 2005; Mazzini et al., 2011).
When possible, 40 valves of ostracods were counted in thesandy fraction of the samples (2 mme63 mm) using a stereomi-croscope (Leica EZ4). The frequency of each taxonwas expressed innumber of valves per gram of sediment (raw and dry). Among the52 samples analysed, only 30 contained ostracods. Only speciesrepresented by both juveniles and adults, or abundant juveniles(more than the 5%) were considered. The ostracod assemblages arelisted in Table 2, together with their autoecology and taphonomicalobservations.
4. Results
The correlation among all the studied proxies, particularly itssedimentological and faunal characteristics, has allowed dividingCore LOA1 into four units.
4.1. Unit A e divided into three sub-units A1, A2 and A3
The concentration of organic material is very low in the wholeunit A (1e2% for the sands and up to 6% for the silts). The fauna islimited to planktonic foraminifera, except in the silty-clayeysequence, which is sterile.
4.1.1. Sub-unit A1 (14.25e19.55 m): light brown sand in layersinterrupted by a grey silty-clayey sequence
This is dated to between 5576 and 5623 BC (6800 ± 50 BP) at15.50 m.
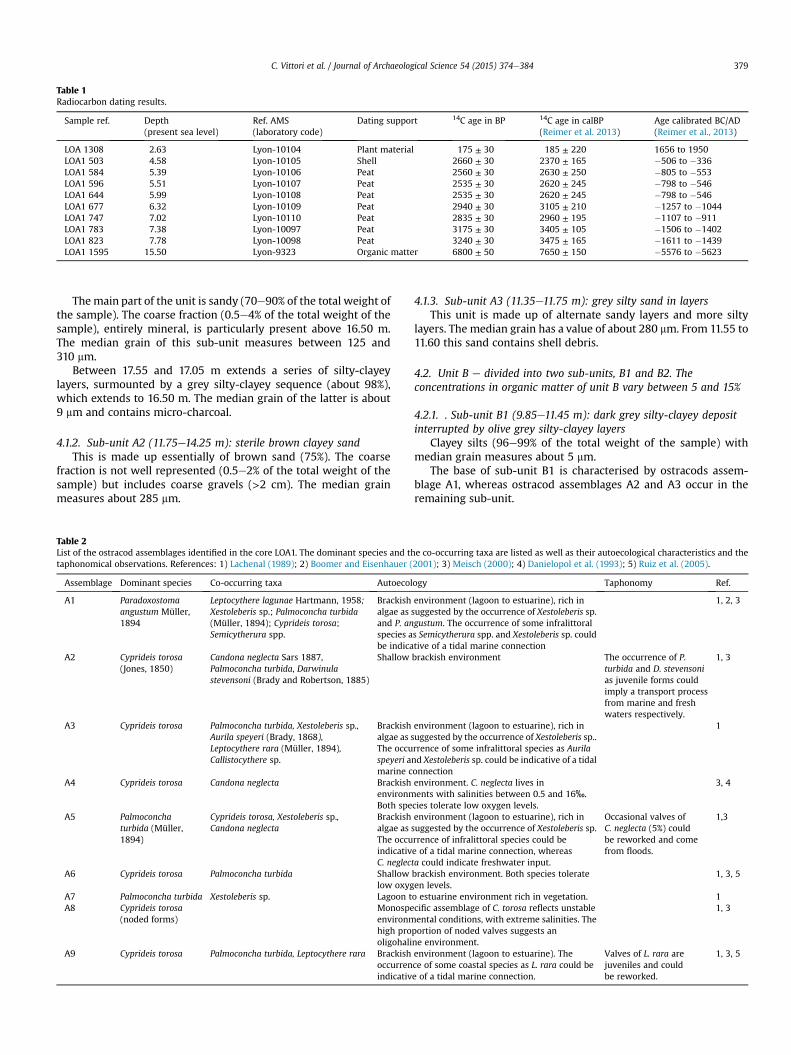
Table 1Radiocarbon dating results.
Sample ref. Depth(present sea level)
Ref. AMS(laboratory code)
Dating support 14C age in BP 14C age in calBP(Reimer et al. 2013)
Age calibrated BC/AD(Reimer et al., 2013)
LOA 1308 2.63 Lyon-10104 Plant material 175 ± 30 185 ± 220 1656 to 1950LOA1 503 4.58 Lyon-10105 Shell 2660 ± 30 2370 ± 165 �506 to �336LOA1 584 5.39 Lyon-10106 Peat 2560 ± 30 2630 ± 250 �805 to �553LOA1 596 5.51 Lyon-10107 Peat 2535 ± 30 2620 ± 245 �798 to �546LOA1 644 5.99 Lyon-10108 Peat 2535 ± 30 2620 ± 245 �798 to �546LOA1 677 6.32 Lyon-10109 Peat 2940 ± 30 3105 ± 210 �1257 to �1044LOA1 747 7.02 Lyon-10110 Peat 2835 ± 30 2960 ± 195 �1107 to �911LOA1 783 7.38 Lyon-10097 Peat 3175 ± 30 3405 ± 105 �1506 to �1402LOA1 823 7.78 Lyon-10098 Peat 3240 ± 30 3475 ± 165 �1611 to �1439LOA1 1595 15.50 Lyon-9323 Organic matter 6800 ± 50 7650 ± 150 �5576 to �5623
C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384 379
Themain part of the unit is sandy (70e90% of the total weight ofthe sample). The coarse fraction (0.5e4% of the total weight of thesample), entirely mineral, is particularly present above 16.50 m.The median grain of this sub-unit measures between 125 and310 mm.
Between 17.55 and 17.05 m extends a series of silty-clayeylayers, surmounted by a grey silty-clayey sequence (about 98%),which extends to 16.50 m. The median grain of the latter is about9 mm and contains micro-charcoal.
4.1.2. Sub-unit A2 (11.75e14.25 m): sterile brown clayey sandThis is made up essentially of brown sand (75%). The coarse
fraction is not well represented (0.5e2% of the total weight of thesample) but includes coarse gravels (>2 cm). The median grainmeasures about 285 mm.
Table 2List of the ostracod assemblages identified in the core LOA1. The dominant species and thtaphonomical observations. References: 1) Lachenal (1989); 2) Boomer and Eisenhauer (
Assemblage Dominant species Co-occurring taxa Autoecol
A1 Paradoxostomaangustum Müller,1894
Leptocythere lagunae Hartmann, 1958;Xestoleberis sp.; Palmoconcha turbida(Müller, 1894); Cyprideis torosa;Semicytherura spp.
Brackishalgae as sand P. anspecies abe indica
A2 Cyprideis torosa(Jones, 1850)
Candona neglecta Sars 1887,Palmoconcha turbida, Darwinulastevensoni (Brady and Robertson, 1885)
Shallow
A3 Cyprideis torosa Palmoconcha turbida, Xestoleberis sp.,Aurila speyeri (Brady, 1868),Leptocythere rara (Müller, 1894),Callistocythere sp.
Brackishalgae as sThe occuspeyeri amarine c
A4 Cyprideis torosa Candona neglecta BrackishenvironmBoth spe
A5 Palmoconchaturbida (Müller,1894)
Cyprideis torosa, Xestoleberis sp.,Candona neglecta
Brackishalgae as sThe occuindicativC. neglec
A6 Cyprideis torosa Palmoconcha turbida Shallowlow oxyg
A7 Palmoconcha turbida Xestoleberis sp. Lagoon tA8 Cyprideis torosa
(noded forms)Monospeenvironmhigh prooligohali
A9 Cyprideis torosa Palmoconcha turbida, Leptocythere rara Brackishoccurrenindicativ
4.1.3. Sub-unit A3 (11.35e11.75 m): grey silty sand in layersThis unit is made up of alternate sandy layers and more silty
layers. The median grain has a value of about 280 mm. From 11.55 to11.60 this sand contains shell debris.
4.2. Unit B e divided into two sub-units, B1 and B2. Theconcentrations in organic matter of unit B vary between 5 and 15%
4.2.1. . Sub-unit B1 (9.85e11.45 m): dark grey silty-clayey depositinterrupted by olive grey silty-clayey layers
Clayey silts (96e99% of the total weight of the sample) withmedian grain measures about 5 mm.
The base of sub-unit B1 is characterised by ostracods assem-blage A1, whereas ostracod assemblages A2 and A3 occur in theremaining sub-unit.
e co-occurring taxa are listed as well as their autoecological characteristics and the2001); 3) Meisch (2000); 4) Danielopol et al. (1993); 5) Ruiz et al. (2005).
ogy Taphonomy Ref.
environment (lagoon to estuarine), rich inuggested by the occurrence of Xestoleberis sp.gustum. The occurrence of some infralittorals Semicytherura spp. and Xestoleberis sp. couldtive of a tidal marine connection
1, 2, 3
brackish environment The occurrence of P.turbida and D. stevensonias juvenile forms couldimply a transport processfrom marine and freshwaters respectively.
1, 3
environment (lagoon to estuarine), rich inuggested by the occurrence of Xestoleberis sp..rrence of some infralittoral species as Auriland Xestoleberis sp. could be indicative of a tidalonnection
1
environment. C. neglecta lives inents with salinities between 0.5 and 16‰.cies tolerate low oxygen levels.
3, 4
environment (lagoon to estuarine), rich inuggested by the occurrence of Xestoleberis sp.rrence of infralittoral species could bee of a tidal marine connection, whereasta could indicate freshwater input.
Occasional valves ofC. neglecta (5%) couldbe reworked and comefrom floods.
1,3
brackish environment. Both species tolerateen levels.
1, 3, 5
o estuarine environment rich in vegetation. 1cific assemblage of C. torosa reflects unstableental conditions, with extreme salinities. The
portion of noded valves suggests anne environment.
1, 3
environment (lagoon to estuarine). Thece of some coastal species as L. rara could bee of a tidal marine connection.
Valves of L. rara arejuveniles and couldbe reworked.
1, 3, 5

C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384380
4.2.2. Sub-unit B2 (7.85e9.85 m): alternation of dark grey and ofolive grey silty-clayey deposits
This is made up of clayey silts (87e97% of the total weight of thesample), bearing more sand at the base (25e50% of the total weightof the sample). The median grain measures about 7 mm.
In the deposits that are grey in colour, ostracod assemblages A5,A6 and A7 represent the microfauna. The macrofauna is concen-trated between 9.85 and 10.05 m and between 7.80 and 8.05 m andis represented by the association of two marine brackish-watertaxa, Hydrobia ventrosa (Montagu, 1803) and Cerastoderma glau-cum (Brugui�ere, 1789) (Drake and Arias, 1995; Casagranda et al.,2005).
In the deposits that are olive grey in colour, the microfauna isrepresented by ostracod assemblage A4. The macrofauna isconcentrated between 9.70 and 9.85 m and between 8.05 and8.55 m and is represented by a freshwater taxon Bithynia tentacu-lata (Linnaeus, 1758) (Stryjak, 1994).
The base of unit B presents a significant micro-faunal density(570e8100 valves per gram of sediment), which tends to decreasetowards the top (2 valves per gram of sediment).
4.3. Unit C (5.30e7.85 m) e succession of peat layers interruptedby silty-clayey deposits
At its base, this unit is dated to between 1611 and 1439 BC(3240 ± 30 BP) and at its top between 805 and 553 BC(2560 ± 30 BP). It is also dated to between 798 and 546 BC(2535 ± 30 BP) at 5.51 and 5.99 m, to between 1257 and 1044 BC(2940 ± 30 BP) at 6.32 m, to between 1107 and 911 BC(2835 ± 30 BP) at 7.02 m and to between 1506 and 1402(3175 ± 30 BP) at 7.38 m (Fig. 3).
At the base of the unit, a 5 cm thick peat level (p-1) passes to anolive grey silty-clayey sequence (90e95% of the total weight of thesample) (Fig. 3). A 36 cm thick peat level (p-2) separates it by silty-clayey sequences (60e90% of the total weight of the sample) 2a and2b, which are respectively grey and olive grey in colour and becomesandy (60% of the total weight of the sample) at the top (Fig. 3). Athird peat level (p-3) extends from 6.21 to 5.97 m before beinginterrupted by the grey silty-clayey sequence 3 (about 90% of thetotal weight of the sample). Unit C is closed by the last peat level (p-4). The coarse fraction (less than 1%) of all these silty-clayey de-posits is made up of shell elements and the median grain variesbetween 5 and 15 mm.
Sequences 1 and 2 are richer in organic matter (16e21% and upto 38% for the sandy sequence) than the sequence 3 (7e14%). Thepeat units p-2 and p-4 are richer in organic matter (50e70%) thanpeat units p-1 and p-3 (about 35%).
The fauna of sequences 1 and 2 is represented by ostracod as-semblages A4 and A8, associated to freshwater gastropod(B. tentaculata). The microfauna of sequence 3 is characterised bythe ostracods assemblage A6, associated to two brackish-watermolluscs (H. ventrosa dominant and C. glaucum).
Overall, the microfaunal density tends to increase (about10e1100 valves per gram of sediment) throughout unit C.
4.4. Unit D e divided into two sub-units D1 and D2
4.4.1. Sub-unit D1 (3.10e5.30 m): dark grey mud containing shellDated to between 506 and 336 BC (2660 ± 30 BP) at 4.58 m.It is made up of very dark grey clayey silts (80e90% of the total
weight of the sample), which present traces of oxidation from 4.00to 4.80m and from 3.10 to 3.35m. Themedian grain varies between5 and 10 mm. The coarse fraction (0 and 3% of the total weight of thesample) is made up of shell elements, except around 5.05 and
4.05 m where mineral elements are also present. A pottery frag-ment was found at 4.22 m.
The concentrations of organic matter are greater in the oxidisedzones (12% as opposed to 6%).
The microfauna of sub-unit D1 is characterised by assemblagesA6 and A9 in the samples at 4.60 and 3.85 m. The macrofauna ismade up of two brackish-water molluscs (C. glaucum and H. ven-trosa). The microfaunal density tends to increase from the base tothe top of the unit (2500e15,000 valves per gram), in spite of adecrease in the oxidised sequences (5e70 valves per gram).Inversely, the macrofaunal density increases in the oxidisedsequences.
Moreover, a drop in magnetic susceptibility started at 3.55 m.
4.4.2. Sub-unit D2 (3.05 m to the fill), dark grey mud containingshell and sandy layers
Dated to between 1656 and 1950 AD (175 ± 30 BP) at 2.63 m.It is made up of dark grey sands (60e70% of the total weight of
the sample) interrupted by a silty-clayey stratum (between 50 and90% of silteclay) from 2.20 to 2.70 m. The coarse fraction, essen-tially mineral, represents between 5 and 20% of the total weight. Itis in general very rich in exogenous elements (23 pottery fragmentsand 12 examples of pozzolana). The median grain of the sandylayers is about 250 mm, as opposed to 15 mm for the silty-clayeylayers. The concentration in organic matter of this sub-unit isvery low (1e3%).
The sandy base of sub-unit D2 is made up of angular grainsassociated with planktonic foraminifera. The remaining sediment isalways characterised by ostracod assemblage A6 and some in-dividuals of brackish-water molluscs C. glaucum and H. ventrosa.Unit D2 finishes up with heterometric sediments and sterile het-erochromes, rich in fragments of faïence tiles.
5. Discussion
5.1. The pre-lagoon environment
The base of core LOA1 consists of marine bed sands, as suggestedby the presence of planktonic foraminifera.
These deposits, dated at their top to the 6th millennium BC,suggest an accumulation of transgressive sands mobilised by theeffect of a largemarine transgression that occurred between 15,000and 8000e7000 BP (Bellotti et al., 2007; Milli et al., 2013).
This sequence is interrupted by sterile silty-clayey depositscharacterised by a large quantity of micro-charcoal, which suggestscontributions of continental origin. This sequence could be relatedto the temporary slowing-down of the rise in sea level at thebeginning of the 7th millennium BC (Bard et al., 2010) which, byprovoking an inversion of dynamics, enabled the development of adelta behind the sandy barrier which was active until about6500 BC (Milli et al., 2013).
The seabed sands were interrupted by silty sands associatedwith few small pebbles (larger than 2 cm). These deposits, probablyof fluvial origin, could correspond to the load from the bottom ofthe Tiber mentioned by Milli et al. (2013). Thus they would havebeen deposited between the 5th and 3rdmillennia BC, in relation tothe progradation of the river (Bellotti et al., 2007; Milli et al., 2013).
Finally, the top of the pre-lagoon unit consists of marine sedi-ments, attested by the presence of planktonic foraminifera. Thesedeposits are characterised by an alternation of sandy and siltylayers. These suggest the progressive establishment of a protectedenvironment, probably related to the formation of sandy barriersfed by the mouth of the Tiber which flowed into the sea and iso-lated the palaeo-lagoon of Ostia around the 3rd millennium BC(Bellotti et al., 2007).

C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384 381
The absence of the ostracods in pre-lagoon environment couldbe due a too coarse sediment grain, a very high sedimentation rateor an environment too subject to waves. Barren sequences could belinked a post mortem dissolution process.
5.2. A lagoon subject to a double hydric influence up to the middleof the 2nd millennium BC
At the beginning, the lagoon environment remained relativelyopen to the sea (sub-unit B1), as attested by the ostracod assem-blages A1 and A3 (Tables 2 and 3). The occasional presence of thefreshwater species Candona neglecta and Darwinula stevensoni inunit B (assemblages A2, A4 and A5), which can tolerate oligohalineand occasionally mesohaline conditions reveals fluvial influxes(Table 3). The palaeo-lagoon of Ostiawas thus probably of estuarinetype, that is, the fluvial and coastal currents played a preponderantrole (Nichols and Allen, 1981), which suggests the maintenance ofthe Tiber to the north of the lagoon and a verymobile sandy barrier.
Over time, the palaeo-lagoon of Ostia tended to becomeconfined (sub-unit B2), attested by the decrease in the diversity ofostracod species (only 4 taxa: Cyprideis torosa, Palmoconcha turbida,Xestoleberis sp. and C. neglecta). This could be related to the thick-ening of the coastal bar by the sands of the progradation of theTiber which accelerated during the 2nd millennium BC (recordedbetween the river and the site of Portus, according to Salomon,2013). It is at this time characterised by the alternation of sedi-mentary deposits, sometimes deposited in a poly-euhaline(ostracod assemblages A5, A6 and A7) context dominated by
Table 3Salinity ranges of the 11 ostracod taxa as reported in the literature. The salinitytolerances are expressed following the Venice System (1959).
Ostracod taxa Salinity ranges Reference
Aurila speyeri (Brady, 1868) Prefers euhalineconditions; toleratesalinity rangeof 25e40‰
Lachenal, 1989
Callistocythere sp. UndefinedCandona neglecta Sars (1887) Prefers freshwater;
reported fromcoastal waters withsalinity range of0.5e16‰(record at 20‰)
Anadon et al.,1986; Meisch,2000
Cyprideis torosa (Jones, 1850) 0.4e150‰; greatestdevelopment atsalinities of 2e16.5‰;at about 5‰ nodedcarapaces dominatedsmooth carapaces
Meisch, 2000;Jahn et al., 1996
Darwinulastevensoni
(Brady andRobertson, 1885)
Prefers freshwater;tolerate an increaseof salinity up to 15‰
Anadon et al.,1986; Meisch,2000
Leptocytherelagunae
Hartmann (1958) Oligo-mesohalineexceptionallyalso polyhaline
Lachenal, 1989;Mazziniet al., 2011
Leptocythererara
(Müller, 1894) Brackish(meso-euhaline)
Lachenal, 1989
Palmoconchaturbida
(Müller, 1894) Marine euryhalinespecies that cansurvive also inmesohalinewaters; greatestdevelopment atsalinities of 35‰
Anadon et al.,1986; Lachenal,1989; Ruizet al., 2005
Paradoxostomaangustum
Müller (1894) Euhaline Lachenal, 1989
Semicytherura sp. Undefined; manyspecies euryhaline
Van Morkhoven,1963
Xestoleberis sp. Undefined
marine contributions, sometimes in an oligo-mesohaline (ostra-cods assemblage A4) context influenced by freshwater contribu-tions. The environmental variability of the lagoon during theseperiods is probably related to the mobility of the Tiber and/or asandy barrier. Indeed, the base of each of these deposits is char-acterised by and increase in the sandy fraction, which indicates agreater hydrodynamism during periods of hydrological changes inthe lagoon.
The oligo-mesohaline sequences indicate larger contributions infreshwater and fluvial sediments, which suggests a period of heavyflooding and/or the nearness of the branch of the Tiber that floodedthe north of the palaeo-lagoon of Ostia. The interruptions of theseby poly-euhaline sequences could have resulted from thedistancing of this branch and/or a rupture of the sandy barrier, dueto storms or floods. The sandy barrier would then have beenthickened by sediment deposits from the mouth of the Tiber andwould thus have progressively closed the lagoon against marineinfluence, favouring the return of an oligo-haline environment.
5.3. Peatification of the lagoon between the second half of the 2ndmillennium and the second half of the 1st millennium BC
The chronostratigraphy of core LOA1 reveals the establishmentof a series of peat formations starting in the first half of the 2ndmillennium BC (1611e1439 BC) up to the first half of the 1st mil-lennium BC (805e553 BC). This peat sequence is interruptedseveral times by silty-clayey sediments.
The cores of sediments from the palaeo-lagoon of Ostiadescribed in the literature also contain peat sequences: (1) acontinuous peat level in core S5 and (2) a discontinuous peat level,interrupted by fluvial sediments, in core S3 (Bellotti et al., 2011, p.1107e1110). They date respectively to between 1740 and 1530 BCand between 1880 and 1680 BC at their base and from 810 to 540 BCand from 910 to 820 BC at their top. These three contemporary peatsequences suggest a generalised peatification of the northern partof the palaeo-lagoon of Ostia.
Coastal peat is generally formed behind a sandy barrier thatlimits the incursions of salt water, in conditions of low oxygenationthat favour the preservation of organic deposits (Vella, 1999). Thepeatification of the palaeo-lagoon of Ostia is thus related to theisolation of the lagoon in relation to the sea and to the decrease offluvial incursions, which would have reduced the hydrodynamismwithin the lagoon. It could thus be due to (1) a phase of pro-gradation of the Tiber delta which favoured the enlargement of thesandy barrier, making it thicker and thus limiting the entry of saltwater into the lagoon, associated with (2) a period of low floodingand/or the progressive distancing of a branch of the Tiber whichflowed near the north side of the palaeo-lagoon of Ostia. Thedecrease of hydro-sedimentary contributions of fluvial origin atthat timewould have led to a decrease in the volume of water in thelagoon and thus accentuated the decrease of its exchanges with thesea (Bird, 1994), favouring the poor oxygenation of theenvironment.
At the base of unit C the inter-peat sequences are mainlydisposed in an oligo-mesohaline (assemblage A4) or even a clearlyoligohaline context, as indicated by the presence of the freshwatergastropod B. tentaculata (Stryjak, 1994) associated with ostracodsassemblage A8. The interruption of the peatification is related tolarge injections of water which ensured a better oxygenation of thelagoon, as well as to fluvial sediments, as suggested by more sandylayers and a high sedimentation rate (about 32.5 mm per yearcompared to only 20 mm per year for the poly-euhaline sequence)in these deposits. These phenomena can be explained by thealternation of periods of low flooding, corresponding to the se-quences of peat, and periods of strong flooding.

C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384382
The last inter-peat deposit is characteristic of a poly-euhalinelagoon in connection with the sea, indicated by the assemblagesA6 and A9. The interruption of these peat sequences could thushave been of (1) fluvial origin, related to one or several floodswhichcould have caused a break in the sandy barrier of the lagoon or of(2) marine origin, related to a storm which would have caused amarine incursion strong enough to have raised the salinity of thelagoon and thus stop the peatification.
According to Bellotti et al. (2011), the presence of pollen fromAlnus suggests that thewaters of the Tiber stopped the peatificationof the palaeo-lagoon of Ostia. However, the analysis of the ostracodsfrom the post-peat deposit in core LOA1 indicates the occurrence ofP. turbida, a marine brackish littoral species. These data tend toconfirm that the stoppage of the peatification was related not to afluvial incursion but to an increase in the salinity. Thus a brackishenvironment, tending to be under marine influence, would havebeen established, associated with fine sediments brought by theflooding of the river, which would also have brought the Alnuspollen revealed by Bellotti et al. (2011). The presence of the Tiber,which strongly influenced the hydrodynamism of this lagoon fromthe time of its formation, thus appears to have had much less in-fluence, which suggests that the river became distanced from thelagoon between the 2nd and the 1st millennia BC.
The coastal peat formations took shape in relation to the watertable and because of this were often slightly raised in relation to sealevel (Vella, 1999). However, the peat of the palaeo-lagoon of Ostiais situated at a depth of 3.5e4.5 m below the ancient biological sea-level (Goiran et al., 2010) in core LOA1, and at 2e3 m beneath theancient biological sea-level in cores S3 and S5 (Bellotti et al., 2011).These depths do not correspond to those that existed during theestablishment of these peat formations and indicate a possiblesubsidence of this sector with a compaction of the peat accentuatedby the underlying sediments.
5.4. A very confined euryhaline palaeo-lagoon beginning in the 9the mid 6th century BC
In the whole unit D, the continuous presence of the ostracodassemblages A6 and occasionally A9, indicates that after805e553 BC (2560 ± 30 BP) the palaeo-lagoon of Ostia remainedpoly-euhaline, maintaining a connectionwith the sea. However, thelow diversity of species (3 species of ostracods e C. torosa, P. turbidaand Leptocythere rara e, 2 species of molluscs e C. glaucum andH. ventrosae and 2 species of foraminifera e Ammonia beccarii(Linn�e, 1758) and punctual presence of Haynesina germanica(Ehrenberg, 1840) associated with a high faunal density suggests aconfined lagoon environment, as described by Guelorget andPerthuisot (1983).
This hypothesis is supported by the fact that the ostracods, aswell as the bivalve C. glaucum and the gastropod H. ventrosa, canwithstand all hypoxic conditions (Ruiz et al., 2005; Jahn et al., 1996;Gamenick et al., 1996; Boyden, 1972). Such an assemblage couldindicate the existence of thermohaline stratification, related to alow hydrodynamism associated with a high solar radiance, at leastseasonally. This thermohaline stratification would have preventedwater circulation and resulted in a hypoxia of the lagoon floor.
Beginning in the 9th century BC, the palaeo-lagoon of Ostia thusbecame a very stable environment in relation with little fluvialinfluence, as indicated by the poly-euhaline salinity and the finesedimentation observed in core LOA1. At that time, the lagoon wasfed by freshwater only through runoff, the rise of the groundwatertable and occasional river water during floods. The continuousenlargement of the sandy barrier by the sediments of the pro-gradation of the Tiber, caused the course of the river to becomemore andmore distant from the lagoon. Its connectionwith the sea
was probably no longer ensured by a simple opening in the sandybarrier but probably, from this period forward, by a true connectioncanal comparable to that visible on old maps (Fig. 2).
After the beginning of the 5th century/middle of the 4th centuryBC, the lagoonwas still connected to the sea and its depth varied, atleast locally, between 3.5 and 4.5 m in depth (in reference to theancient biological sea-level at the 3rde5th centuries AD accordingto Goiran et al., 2010). Thus, after the foundation of Ostia in thesecond half of the 4th century BC, it would have potentially servedas a natural coastal shelter, although this is not mentioned in thetexts.
5.5. Maintenance and infilling of the palaeo-lagoon of Ostia
The presence of rare coarse mineral elements associate to a fewjuvenile valves of L. rara in sub-unit D1 could be due to contact ofthe lagoon and marine waters or transported by storms. The first isdated to between 506 and 336 BC (2660 ± 30 BP) and is thus moreor less contemporary with the construction of the Castrum. Itspresence suggests that the distance between the Ostia lagoon andthe sea was at least one kilometre (Bellotti et al., 2011). Thesemarine inputs into the lagoon could be linked to an anthropicregulation of the connection between the lagoon and the sea,related to human activity in these salt marshes. Indeed, the con-necting canal between the palaeo-lagoon of Ostia and the sea couldhave been maintained by dredging, as was the case from 16th to19th century AD, notably at the end of the 16th century at thedemand of Pope Clement VIII (Fea, 1831). The necessity for cleaningthe canal, which often silted up, is probably related to the phase ofrapid progradation which occurred between the 16th and the 20thcenturies (Bersani and Moretti, 2008). The continuous existence ofthe connection between the lagoon and the sea during Antiquity, inspite of rapid progradation (Bicket et al., 2009), indicates the pos-sibility of maintenance by humans.
From the 17the20th centuries onward, human presence is moreand more visible in the sediments, with the introduction of manyexogenous elements (terra cotta, mortar, pozzolana, etc.). It couldbe linked to human activities typical of the area such as, forexample, fishing and salt exploitation. The last lagoon sedimentsare characterised by a faunal assemblage similar (C. torosa domi-nant, P. turbida and punctual presence of L. rara, as well as themolluscs C. glaucum and H. ventrosa) to that which has persistedsince Antiquity. It indicates the existence of a continuing connec-tion with the sea, which could be linked to the occurrence of thesalinity wedge in the groundwater (Pepe et al., 2013), and nothingshows that there was a decrease in the amounts of water. The sandydeposits of pozzolana, which came to fill the lagoon are angular andcontain taxa that are typically marine (planktonic foraminifera)associated with a terrestrial gastropod, which appears to indicatethat this last deposit is not natural.
6. Conclusion
The new stratigraphic sequence from core LOA1 has revealedthat the flooding of the palaeo-lagoon of Ostia by the waters of theTiber was recurrent throughout the stratigraphy, since the begin-ning of the formation of the lagoon (post 6th millennium BC) to the9th century BC (Fig. 3). After a lagoon open to the sea was formed itevolved into a lagoon under the double influence of the sea and theriver. Because of this, it differed from the palaeo-lagoon of Mac-carese, which was mainly open to marine influences up to the 9thcentury (Bellotti et al., 1994, 1995; 2007).
Over time, the fluvial influence became more and more presentin the palaeo-lagoon of Ostia, in contrast to the palaeo-lagoon ofMaccarese, which became isolated from the sea and was

C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384 383
transformed into an area of peat around the beginning of the 4thmillennium BC (Giraudi, 2004; Bellotti et al., 2007; Di Rita et al.,2009). These different situations tend to confirm the presence ofa branch of the Tiber situated near the northern part of the palaeo-lagoon of Ostia. This branch, by maintaining a sufficient level ofwater in the lagoon, participated in the control of hydric exchangesbetween the lagoon and the sea.
During the second half of the 2nd millennium BC, the marineand fluvial contributions to the lagoon diminished, which led to theformation of peat. The bank of the lagoon was then progressivelytaken over by the sands of progradation of the delta, and thepalaeo-lagoon of Ostia became more and more isolated from thesea. Between the 9th and the 6th centuries BC, both the palaeo-lagoons of Maccarese (Giraudi, 2002, 2004; Di Rita et al., 2009)and Ostia were re-opened. The re-opening could be anthropic inorigin (Bellotti et al., 2011) as highlighted by an increase in Che-nopodiaceae, a sign of increased salinity (Di Rita et al., 2009;Bellotti et al., 2011).
The new data from core LOA1 indicate that the peatification ofthe lagoon of Ostia was interrupted by a marine incursion and thatthe Tiber could have become distanced from the lagoon betweenthe 2nd and 1st millennia BC.
Beginning in the 9th century BC, the preservation of theconnection between the sea and the palaeo-lagoon of Ostia, in spiteof the progressive distancing of the shore, suggests that thelagoonal environment was partly regulated by human intervention,such as the maintenance of the connecting canal between the seaand the lagoon, necessary for successful human activity related tothe salt marshes.
Acknowledgements
Ecole Française de Rome, Soprintendenza Speciale per i BeniArcheologici di Roma e Sede di Ostia, Maison de l'Orient et de laM�editerran�ee (UMR 5133), Intelligence des mondes urbains (LabExIMU), Universit�a la Sapienza & Roma 3, CNRS, Universit�e de Lyon,Institut Universitaire de France, J. Cavero (topographer), OMEAAplatform, ARTEMIS-SHS program and radiocarbon laboratory ofLyon.
References
Amenduni, G., 1884. Sulle opere di bonificazione della plaga litoranea dell'AgroRomano che comprende le paludi e gli stagni di Ostia, Porto, Maccarese e delleterre vallive di Stracciacappa, Baccano, Pantano e Lago dei Tartari. Relazione delprogetto generale 15/7.
Amorosi, A., Milli, S., 2001. Late Quaternary depositional architecture of Po andTevere river deltas (Italy) and worldwide comparison with coeval deltaic suc-cessions. Sediment. Geol. 144 (3e4), 357e375.
Anadon, P., De Deckker, P., Julia, R., 1986. The Pleistocene lake deposits of the NEBaza Basin (Spain): salinity variations and ostracod succession. Hydrobiologia143, 199e208.
Arnoldus-Huyzendveld, A., Paroli, L., 1995. Alcune considerazioni sullo sviluppostorico dell'ansa dell Tevere presso Ostia e sul porto-canale. Archeologia Laziale,pp. 383e392.
Ball, D.F., 1964. Loss-on-ignition as an estimate of organic matter and organic car-bon in non-calcareous soils. J. Soil Sci. 15 (1), 84e92.
Bard, E., Hamelin, B., Delanghe-Sabatier, D., 2010. Deglacial meltwater pulse 1B andYounger Dryas sea levels revisited with boreholes at Tahiti. Science 327,1235e1237.
Bellotti, P., Calderoni, G., Carboni, M.G., Bella, L.D., Tortora, P., Valeri, P.,Zernitskaya, V., 2007. Late Quaternary landscape evolution of the Tiber riverdelta plain (Central Italy): new evidence from pollen data, biostratigraphy and14C dating. Z. Geomorphol. 51, 505e534.
Bellotti, P., Carboni, M.G., Milli, S., Tortora, P., Valeri, P., 1989. La Piana deltizia delTevere: analisi di facies ed ipotesi evolutiva dall'ultimo low stand glacialeall'attuale. G. Geol. 51, 7e11.
Bellotti, P., Chiocci, F.L., Milli, S., Tortora, P., Valeri, P., 1994. Sequence stratigraphyand depositional setting of the Tiber delta: integration of high-resolutionseismics, well logs, and archeological data. J. Sediment. Res. Section Stratigr.Glob. Stud. 64, 416e432.
Bellotti, P., Milli, S., Tortora, P., Valeri, P., 1995. Physical stratigraphy and sedimen-tology of the Late Pleistocene-Holocene Tiber Delta depositional sequence.Sedimentology 42, 617e634.
Bellotti, P., Calderoni, G., Rita, F.D., D'Orefice, M., D'Amico, C., Esu, D., Magri, D.,Martinez, M.P., Tortora, P., Valeri, P., 2011. The Tiber river Delta Plain (CentralItaly): coastal evolution and implications for the ancient Ostia roman settle-ment. Holocene 21, 1105e1116.
Belluomini, G., Iuzzolini, P., Manfra, L., Mortari, R., Zalaffi, M., 1986. Evoluzionerecente del delta del Tevere. Geol. Romana 25, 213e234.
Bersani, P., Moretti, D., 2008. Evoluzione storica della linea di costa in prossimit�adella foce del Tevere. L'Acqua, 77e88.
Bicket, A.R., Rendell, H.M., Claridge, A., Rose, P., Andrews, J., Brown, F.S.J., 2009.A multiscale geoarchaeological approach from the Laurentine shore (Cas-telporziano, Lazio, Italy). G�eomorphologie 4, 257e270.
Bidet, J.-G., Carruesco, C., Klingebiel, A., 1982. L'approche g�eologique des environ-nements lagunaires. Centre International pour la Formation et les EchangesG�eologiques, Paris.
Bird, E.C.F., 1994. Physical setting and geomorphology of coastal lagoons. CoastalLagoon Processes. Elsevier Science Publishers, Amsterdam, pp. 9e39.
Bocci, D., 1892. Il Delta Tiberino. Tipografia del Genio civile, Roma.Boomer, I., Eisenhauer, G., 2001. Ostracods as palaeoenvironmental indicators. In:
Holmes, J., Horne, D. (Eds.), The Ostracoda. Applications in Quaternary Research.AGU Books, pp. 135e149.
Boyden, C.-R., 1972. The behaviour, survival and respiration of the cockles Cera-stoderma Edule and C. Glaucum. Air J. Mar. Biol. Assoc. U. K. 52, 661e680.
Cailleux, A., Tricart, J., 1959. Initiation �a l'�etude des sables et des galets. Centre dedocumentation universitaire, Paris.
Cailleux, A., 1981. Code des couleurs des sols. Boub�ee, Paris.Calza, G., Becatti, G., Gismondi, I., 1953. Scavi di Ostia 1, Topografia generale. Libreria
dello Stato, Roma.Carbonel, P., 1988. Ostracods and the transition between fresh and saline waters. In:
De Deccker, P., Colin, J.-P., Peyrpouquet, J.-P. (Eds.), Ostracoda in the Earth Sci-ences, pp. 157e173.
Casagranda, C., Boudouresque, C.F., Francour, P., 2005. Abundance, populationstructure and production of Hydrobia ventrosa (Gastropoda: Prosobranchia) in aMediterranean brackish lagoon, Lake Ichkeul, Tunis. Arch. Hydrobiol. 164 (3),411e428.
Danielopol, D.L., Handl, M., Yin, Y., 1993. Benthic ostracods in the pre-Alpine deepLake Mondsee: notes on the origin and distribution. In: McKenzie, K.G.,Jones, P.J. (Eds.), Ostracoda in the Earth and Life Sciences, pp. 465e480.
Dearing, J.A., 1999. Environmental Magnetic Susceptibility, Using the BartingtonMS2 System M, second ed. Chi Publishing, England.
Delrue, V., 2011. Salt tanning, dyeing and cleaning in ancient Egypt. In: Alexianu, M.,Weller, O., Curca, R.G. (Eds.), “Archaeology and Anthropology of Salt: aDiachronic Approach” Proceedings of the International Colloquim, BritishArchaeological Reports International Series 2198. Archaeopress, Oxford,pp. 163e168.
Di Rita, F., Celant, A., Magri, D., 2009. Holocene environmental instability in thewetland north of the Tiber delta (Rome, Italy): seaelakeeman interaction.J. Paleolimnol. 44 (1), 51e67.
Dragone, F., Maino, A., Malatesta, A., Segre, A.G., 1967. Note Illustrative Della C.G.I.Alla Scala 1:100.000.
Drake, P., Arias, A.M., 1995. Distribution and production of three Hydrobia species(Gastropoda: Hydrobiidae) in a shallow coastal lagoon in the bay of C�adiz,Spain. J. Molluscan Stud. 61, 185e196.
Folk, R.L., Ward, W.C., 1957. Brazos river bar [Texas]; a study in the significance ofgrain size parameters. J. Sediment. Res. 27 (1), 3e26. Foglio 149 (Cerveteri).Servizio Geologico d'Italia.
Fea, C., 1831. Storia delle Saline d'Ostia introdotte da Anco Marcio quarto re di Romadopo la fondazione di quella, Roma.
Frenzel, P., Boomer, I., 2005. The use of ostracods from marginal marine, brackishwaters as bioindicators of modern and Quaternary environmental change.Palaeogeogr. Palaeoclimatol. Palaeoecol. 225 (1e4), 68e92.
Frutaz, A.P., 1972. Le carte del Lazio. Roma.Gamenick, I., Jahn, A., Vopel, K., Giere, O., 1996. Hypoxa and sulphide as structuring
factors in a macrozoobenthic community on the Baltic Sea shore: colonisationstudies and tolerance experiments. Mar. Ecol. Prog. Ser. 144, 3e85.
Giovannini, A., 2001. Les salines d'Ostie, in Ostia. Porte et porte de la Rome antique(Catalogo della Mostra di Ginevra). Ginevra, pp. 373e384.
Giraudi, C., 2002. Evoluzione ambientale tardo-olocenica nell'area comprente il stoeneolitico di Maccarese (Fiumicino). In: Manfredini, A. (Ed.), Le Dune, Il Lago, IlMare. Una communit�a di villaggio dell'Et�a del Rame a Maccarese, Istituto ital-iano di preistoria e protostoria, Firenze, pp. 25e35.
Giraudi, C., 2004. Evoluzione tardo-olocenica del delta del Tevere. Il Quaternario-Italian J. Quat. Sci. 17, 477e482.
Giraudi, C., 2011. The sediments of the ‘Stagno di Maccarese’ marsh (Tiber riverdelta, central Italy): a late-Holocene record of natural and human-inducedenvironmental changes. Holocene 21 (8), 1233e1243.
Goiran, J.P., Tronch�ere, H., Salomon, F., Carbonel, P., Djerbi, H., Ognard, C., 2010.Palaeoenvironmental reconstruction of the ancient harbors of Rome: Claudiusand Trajan's marine harbors on the Tiber delta. Quat. Int., 3e13.
Guelorget, J.P., Perthuisot, O., 1983. Le Domaine paralique: expressions g�eologiques,biologiques et �economiques du confinement. Presses de l'�Ecole NormaleSup�erieure, Paris.

C. Vittori et al. / Journal of Archaeological Science 54 (2015) 374e384384
Jahn, A., Gamenick, I., Theede, H., 1996. Physiological adaptations of Cyprideis tor-osa(Crustacea, Ostracoda) to hydrogen sulphide. Mar. Ecol. Prog. Ser. 142,215e223.
Lachenal, A.-M., 1989. �Ecologie des ostracodes du domaine m�editerran�een : appli-cation au golfe de Gab�es (Tunisie orientale). Les variations du niveau marindepuis 30 000 ans, 239 pp.
Lambeck, K., Antonioli, F., Anzidei, M., Ferranti, L., Leoni, G., Scicchitano, G.,Silenzi, S., 2011. Sea level change along the Italian coast during the Holoceneand projections for the future. Quat. Int. 232 (1e2), 250e257.
Mazzini, I., Faranda, C., Giardini, M., Giraudi, C., Sadori, L., 2011. Late Holocenepalaeoenvironmental evolution of the ancient harbour of Portus (Latium,Central Italy). J. Palaeolimnol 46, 243e256.
Meiggs, R., 1973. Roman Ostia. Oxford, 16e20 and 268e269.Meisch, C., 2000. Freshwater ostracoda of western and Central Europe. In:
Schwoerbel, J., Zwick, P. (Eds.), Süsswasserfauna von Mitteleuropa. SpektrumAkademischer Verlag 8/3, Heidelberg, pp. 1e522.
Milli, S., D'Ambrogi, C., Bellotti, P., Calderoni, G., Carboni, M.G., Celant, A., Bella, L., DiRita, F., Di Frezza, V., Magri, D., Pichezzi, R.M., Ricci, V., 2013. The transition fromwave-dominated estuary to wave-dominated delta: the Late Quaternary strat-igraphic architecture of Tiber river deltaic succession (Italy). Sediment. Geol.284e285, 159e180.
Moro, G., 1871. Lo stagno di Ostia, Monogrfia geologica ed idraulica. Tipografiaeditrice, Firenze.
Nichols, M., Allen, G.P., 1981. Sedimentary Process in Coastal Lagoons, CoastalLagoon Research Present and Future. UNESCO Technical papers in marine sci-ences 33, pp. 27e80.
Oberholtzer, F., 1875. Le foci del Tevere. GB Paravia, Roma.Pannuzi, S., 2013. La laguna di Ostia: produzione del sale e trasformazione del
paesaggio dall'et�a antica all'et�a moderna. MEFRM (M�elanges de l'�Ecole fran-çaise de Roma- Moyen Age) [Online], 125e2, 2013. URL: http://mefrm.revies.org/1507 http://mefrm.revies.org/1507.
Pannuzi, S., 2012. Le saline e lo stagno di Ostia: utilizzo e trasformazione del sub-urbio orientale ostiense dall'et�a antica all'et�a moderna (XVI-XVIII secolo). VICongresso Nazionale di Archeologia Medievale (L'Aquila 2012), Firenze,pp. 321e326.
Pepe, C., Giardini, M., Giraudi, C., Masi, A., Mazzini, I., Sadori, L., 2013. Plant land-scape and environmental changes recorded in marginal marine environments:the ancient Roman harbour of Portus (Rome, Italy). Quat. Int. 303, 73e81.
Ponzi, G., 1875. Il delta del Tevere: storia naturale del Tevere. Boll. della Soc. Geogr.It 12, 1e22.
Reimer, P.J., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., Bronk Ramsey, C.,Grootes, P.M., Guilderson, T.P., Haflidason, H., Hajdas, I., Hatte, C., Heaton, T.J.,Hoffmann, D.L., Hogg, A.G., Hughen, K.A., Kaiser, K.F., Kromer, B., Manning, S.W.,Niu, M., Reimer, R.W., Richards, D.A., Scott, E.M., Southon, J.R., Staff, R.A.,Turney, C.S.M., Van der Plicht, J., 2013. IntCal13 and Marine13 radiocarbon AgeCalibration curves 0-50,000 Years cal BP. Radiocarbon 55 (4), 1869e1887.
Ruiz, F., Abad, M., Bodergat, A.M., Carbonel, P., Rodríguez-L�azaro, J., Yasuhara, M.,2005. Marine and brackish-water ostracods as sentinels of anthropogenic im-pacts. Earth-Science Rev. 72 (1e2), 89e111.
Salomon, F., 2013. G�eoarch�eologie du Delta du Tibre : �evolution g�eomorphologiqueholoc�ene et contraintes hydros�edimentaires dans le syst�eme Ostie e Portus.Th�ese de doctorat en g�eographie physique/g�eoarch�eologie. Universit�e Lyon 2.
Stryjak, J.F., 1994. Recherches sur la biologie de Bithynia tentaculata LINNE 1758.Effets de l'�echauffement r�egulier du milieu aquatique sur le cycle biologique dece Mollusque Gast�eropode Prosobranche (Th�ese de doctorat). Universit�e deMetz.
Tomassetti, G., 1897. Della campagna romana, Via Ostiense e Laurentina. ASRSP, XX,pp. 45e94.
Van Morkhoven, F.P., 1963. Post-Palaeozoic Ostracoda: Their Morphology, Taxon-omy and Economic Use. Elsevier Publishing Company, Amsterdam and NewYork.
Vella, C., 1999. Perception et �evaluation de la mobilit�e du littoral Holoc�ene sur lamarge orientale du delta du Rhone. Th�ese de g�eomorphologie. Universit�e Aix-Marseille I.
Venice System, 1959. Symposium on the classification of brackish waters, Venice,April 8e14, 1958. Arch. Ocean. Limnol. (Suppl. 11), 1e248.
Zevi, F., 2001. Les d�ebuts d'Ostie. In: Descoeudres, J.P. (Ed.), Ostia, port et porte de laRome antique. GEORG, Geneve, pp. 3e9.
Zevi, F., 2002. Origini di Ostia. In: Bruun, C., Zevi, A.G. (Eds.), Ostia e Portus nelle lororelazioni con Roma, pp. 11e32.
Related Documents











