Palaeoecological potential of the marine organic deposits of Posidonia oceanica: A case study in the NE Iberian Peninsula José A. López-Sáez a, ⁎, Lourdes López-Merino a , Miguel Á. Mateo b , Óscar Serrano b , Sebastián Pérez-Díaz a , Laura Serrano b a GI Arqueobiología, Instituto de Historia, CCHS, CSIC, Albasanz 26-28, 28037 Madrid, Spain b Centre d'Estudis Avançats de Blanes (CEAB), CSIC, Accés a la Cala St. Francesc 14, 17300 Blanes, Spain abstract article info Article history: Received 6 August 2008 Received in revised form 21 October 2008 Accepted 27 October 2008 Keywords: Posidonia oceanica Peat-like deposits Vegetation dynamics Human impact Holocene NE Iberian Peninsula A high-resolution sedimentary sequence sampled from the Portlligat Bay (NE Iberian Peninsula) has been studied in order to assess its palaeoecological potential. This sedimentary sequence is considered a very particular one, as it is a peat-like deposit derived from the accumulation of the belowground dead parts of the marine phanerogam Posidonia oceanica through the last 1600 years. This study represents the first attempt to explore the palaeoecological potential of such deposits from a palynological point of view, and has allowed the reconstruction of the vegetation dynamics and changes, human impact and the palaeoclimatic characteristics, demonstrating the value of these sedimentary records in palaeopalynological studies. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Pollen analyses on Holocene sedimentary records have been carried out in very different contexts, but they share two main characteristics: 1) the long-term accumulation of biogenic and/or minerogenic material; and 2) the preservation of pollen, spores, and other non-pollen microfossils. Peat-bogs and lakes provide the best examples of terrestrial palaeoenvironmental reconstruction from pollen analysis (Aaby and Berglund, 1986), although palaeosoils, caves, rockshelters and archaeological sites are also investigated (Carrión et al., 1999; Carrión, 2001; López-Sáez et al., 2003). In contrast, most of the studies carried out in oceanic environments come from the exploration of the sea bed (e.g. Tzedakis et al., 1997). In marine environments, the in situ accumulation of organic materials is fairly uncommon, but it has been described in swamps, sapropels and marine phanerogam meadows (e.g. Mateo et al., 1997; Rossignol-Strick, 1999; Rull, 2001). Palaeorecords of the only angios- perms that have invaded the submarine realm, seagrasses, are virtually lacking because, apparently, the appropriate preservation conditions have been very rarely met and because, with a few exceptions, seagrass pollen lacks exine, the outermost coat of the pollen grain. The endemic Mediterranean seagrass Posidonia oceanica is the most abundant marine phanerogam in the Mediterranean Sea, spreading over ca. 25,000–50,000 km 2 in waters of up to 45 m of depth (Bethoux and Copin-Montegut, 1986), and it shows the peculiarity of generating great amounts of organic litter from its roots, rhizomes and foliar sheaths, giving rise to a peat-bog-like biologic structure, known as ‘matte’ (Boudouresque et al., 1980; Mateo et al., 1997)(Fig. 1). The accumulation rate of these deposits, derived from the balance between the production, decomposition and erosion of the material, has been estimated as 0.1.–0.4 cm year - 1 , remaining as an organic sediment for millenia (Romero et al., 1994; Mateo et al., 1997, 2002). Thus, since the organic accumulation in the matte follows a coherent chronological pattern, interesting possibilities arise in the field of the ecology (Mateo et al., 2002; López-Sáez et al., 2006); given the fact that these sedimentary records are exclusive of Posidonia oceanica and that, up to date, they have not been exploited from a palaeopalynological point of view. Previous studies using radiocarbon techniques have revealed that these peats, which stratigraphy accurately reflect their formation chronology, constitute unique organic records comprising thousands of years (Boudouresque et al., 1980; Romero et al., 1994; Mateo et al., 1997, 2002). Damblon and Vanden Berghen (1993) carried out pollen analysis on a Posidonia oceanica supra-littoral deposit of only 15 cm located in the Djerba Islands in the south of Tunisia, in which they studied several specific samples dated ca. 4300–4200 BP, but not obtained from a single drill core. This study evaluates, for the first time, the palaeoenvironmental potential of a matte core of 482 cm underneath the meadow of Posi- donia oceanica, in the NE Iberian Peninsula. A pollen analysis (pollen grains, spores and non-pollen palynomorphs (NPPs)) of the uppermost Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224 ⁎ Corresponding author. Tel.: +34 916022477. E-mail address: [email protected] (J.A. López-Sáez). 0031-0182/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2008.10.020 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
Palaeoecological potential of the marine organic deposits of Posidonia oceanica:A case study in the NE Iberian Peninsula
José A. López-Sáez a,⁎, Lourdes López-Merino a, Miguel Á. Mateo b, Óscar Serrano b,Sebastián Pérez-Díaz a, Laura Serrano b
a GI Arqueobiología, Instituto de Historia, CCHS, CSIC, Albasanz 26-28, 28037 Madrid, Spainb Centre d'Estudis Avançats de Blanes (CEAB), CSIC, Accés a la Cala St. Francesc 14, 17300 Blanes, Spain
⁎ Corresponding author. Tel.: +34 916022477.E-mail address: [email protected] (J.A. López-Sáez).
0031-0182/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.palaeo.2008.10.020
a b s t r a c t
a r t i c l e i n f oArticle history:
A high-resolution sediment Received 6 August 2008Received in revised form 21 October 2008Accepted 27 October 2008Keywords:Posidonia oceanicaPeat-like depositsVegetation dynamicsHuman impactHoloceneNE Iberian Peninsula
ary sequence sampled from the Portlligat Bay (NE Iberian Peninsula) has beenstudied in order to assess its palaeoecological potential. This sedimentary sequence is considered a veryparticular one, as it is a peat-like deposit derived from the accumulation of the belowground dead parts ofthe marine phanerogam Posidonia oceanica through the last 1600 years. This study represents the firstattempt to explore the palaeoecological potential of such deposits from a palynological point of view, and hasallowed the reconstruction of the vegetation dynamics and changes, human impact and the palaeoclimaticcharacteristics, demonstrating the value of these sedimentary records in palaeopalynological studies.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Pollen analyses on Holocene sedimentary records have beencarried out in very different contexts, but they share two maincharacteristics: 1) the long-term accumulation of biogenic and/orminerogenic material; and 2) the preservation of pollen, spores, andother non-pollen microfossils. Peat-bogs and lakes provide the bestexamples of terrestrial palaeoenvironmental reconstruction frompollen analysis (Aaby and Berglund, 1986), although palaeosoils,caves, rockshelters and archaeological sites are also investigated(Carrión et al., 1999; Carrión, 2001; López-Sáez et al., 2003). Incontrast, most of the studies carried out in oceanic environmentscome from the exploration of the sea bed (e.g. Tzedakis et al., 1997).
In marine environments, the in situ accumulation of organicmaterials is fairly uncommon, but it has been described in swamps,sapropels and marine phanerogam meadows (e.g. Mateo et al., 1997;Rossignol-Strick, 1999; Rull, 2001). Palaeorecords of the only angios-perms that have invaded the submarine realm, seagrasses, are virtuallylacking because, apparently, the appropriate preservation conditionshave been very rarelymet and because, with a few exceptions, seagrasspollen lacks exine, the outermost coat of the pollen grain. The endemicMediterranean seagrass Posidonia oceanica is the most abundantmarine phanerogam in the Mediterranean Sea, spreading over ca.25,000–50,000 km2 in waters of up to 45 m of depth (Bethoux and
l rights reserved.
Copin-Montegut, 1986), and it shows the peculiarity of generatinggreat amounts of organic litter from its roots, rhizomes and foliarsheaths, giving rise to a peat-bog-like biologic structure, known as‘matte’ (Boudouresque et al., 1980; Mateo et al., 1997) (Fig. 1). Theaccumulation rate of these deposits, derived from the balance betweenthe production, decomposition and erosion of the material, has beenestimated as 0.1.–0.4 cm year−1, remaining as an organic sediment formillenia (Romero et al., 1994; Mateo et al., 1997, 2002). Thus, since theorganic accumulation in the matte follows a coherent chronologicalpattern, interesting possibilities arise in the field of the ecology (Mateoet al., 2002; López-Sáez et al., 2006); given the fact that thesesedimentary records are exclusive of Posidonia oceanica and that, up todate, they have not been exploited from a palaeopalynological point ofview.
Previous studies using radiocarbon techniques have revealed thatthese peats, which stratigraphy accurately reflect their formationchronology, constitute unique organic records comprising thousandsof years (Boudouresque et al., 1980; Romero et al., 1994; Mateo et al.,1997, 2002). Damblon and Vanden Berghen (1993) carried out pollenanalysis on a Posidonia oceanica supra-littoral deposit of only 15 cmlocated in the Djerba Islands in the south of Tunisia, in which theystudied several specific samples dated ca. 4300–4200 BP, but notobtained from a single drill core.
This study evaluates, for the first time, the palaeoenvironmentalpotential of a matte core of 482 cm underneath the meadow of Posi-donia oceanica, in the NE Iberian Peninsula. A pollen analysis (pollengrains, spores and non-pollenpalynomorphs (NPPs)) of the uppermost

Fig. 1. Top: A Posidonia oceanica matte as viewed in a reef barrier off the island ofFormentera (Balearic Archipelago, Spain). The matte wall in the image is 3 m high.Bottom: Detail of the core sampled in Portlligat Bay. The image shows a partial length ofthe uppermost 123 cm of the core, from which the pollen samples for this study wereisolated.
216 J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
123 cm is presented here. The aim of this work is to evaluate thepalaeoecologic potential of these marine deposits due to theunprecedented possibilities to perform detailed studies of vegetationand palaeoenvironmental history that the Posidonia peats offer.
Fig. 2. Situation map of Portlligat Bay in the NE Iberian Peninsula an
2. Description of the studied area
The Portlligat Bay (42°17′32″N; 3°17′28″E) is a small inlet locatedon the NE coast of the Iberian Peninsula, in the province of Girona,oriented to the NE towards the Mediterranean Sea (Fig. 2); and hasaverage minimum and maximum water depths of 2.2 m and 7.1 m,respectively. It is connected to the sea through a wide opening to theNW of ca. 213 m. The maximum length and width of the bay are1373 m and 252 m, respectively. About 68% of the bottom of the bay iscovered by living beds of Posidonia oceanica (ca. 9 ha) with shootdensities ranging from 100 to 900 shoots m−2.
Geologically, the Cape Creus and the Portlligat Bay are oldformations that originated ca. 400 million years ago along with thePyrenees. The substrate is of igneous and sedimentary origin, latertransformed through metamorphism and faulting, particularly duringthe Hercynian period. The bay receives episodic freshwater inputsfrom a typical Mediterranean temporary stream that flows into thebay from its NE shore. The annual precipitation ranges between 500and 800 mm and mainly occurs from October through December(average range for the period 2000–2006, as recorded by themeteorological station of Roses, Servei Meteorològic de Catalunya).
3. Materials and methods
3.1. Sediment retrieval and dating
The core (482 cm) was sampled in Portlligat Bay (Girona, NWMediterranean Sea; Fig.1) from a light floating drilling rig using a rotarydrilling pneumatic hammer and a 150 cm long, 8.8 cm diameter (inner)stainless steel corer. Sheath debris was dried at 70 °C to a constantweight and milled to a fine powder in a porcelain mortar prior toaccelerator mass spectrometry (AMS) 14C determination. Radiocarbon
d location of the palynologic drill of the Posidonia oceanica beds.
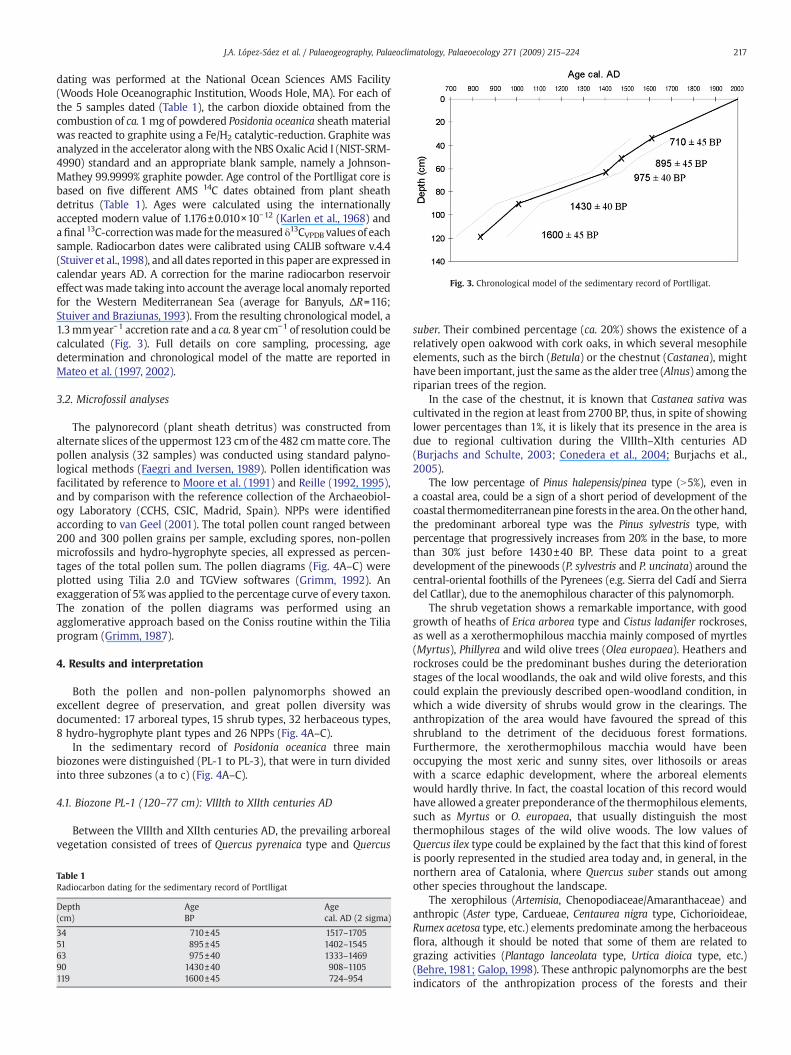
Fig. 3. Chronological model of the sedimentary record of Portlligat.
217J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
dating was performed at the National Ocean Sciences AMS Facility(Woods Hole Oceanographic Institution, Woods Hole, MA). For each ofthe 5 samples dated (Table 1), the carbon dioxide obtained from thecombustion of ca.1 mg of powdered Posidonia oceanica sheath materialwas reacted to graphite using a Fe/H2 catalytic-reduction. Graphite wasanalyzed in the accelerator alongwith the NBS Oxalic Acid I (NIST-SRM-4990) standard and an appropriate blank sample, namely a Johnson-Mathey 99.9999% graphite powder. Age control of the Portlligat core isbased on five different AMS 14C dates obtained from plant sheathdetritus (Table 1). Ages were calculated using the internationallyaccepted modern value of 1.176±0.010×10−12 (Karlen et al., 1968) andafinal 13C-correctionwasmade for themeasured δ13CVPDB values of eachsample. Radiocarbon dates were calibrated using CALIB software v.4.4(Stuiver et al.,1998), and all dates reported in this paper are expressed incalendar years AD. A correction for the marine radiocarbon reservoireffect wasmade taking into account the average local anomaly reportedfor the Western Mediterranean Sea (average for Banyuls, ΔR=116;Stuiver and Braziunas, 1993). From the resulting chronological model, a1.3mmyear−1 accretion rate and a ca. 8 year cm−1 of resolution could becalculated (Fig. 3). Full details on core sampling, processing, agedetermination and chronological model of the matte are reported inMateo et al. (1997, 2002).
3.2. Microfossil analyses
The palynorecord (plant sheath detritus) was constructed fromalternate slices of the uppermost 123 cm of the 482 cmmatte core. Thepollen analysis (32 samples) was conducted using standard palyno-logical methods (Faegri and Iversen, 1989). Pollen identification wasfacilitated by reference to Moore et al. (1991) and Reille (1992, 1995),and by comparison with the reference collection of the Archaeobiol-ogy Laboratory (CCHS, CSIC, Madrid, Spain). NPPs were identifiedaccording to van Geel (2001). The total pollen count ranged between200 and 300 pollen grains per sample, excluding spores, non-pollenmicrofossils and hydro-hygrophyte species, all expressed as percen-tages of the total pollen sum. The pollen diagrams (Fig. 4A–C) wereplotted using Tilia 2.0 and TGView softwares (Grimm, 1992). Anexaggeration of 5%was applied to the percentage curve of every taxon.The zonation of the pollen diagrams was performed using anagglomerative approach based on the Coniss routine within the Tiliaprogram (Grimm, 1987).
4. Results and interpretation
Both the pollen and non-pollen palynomorphs showed anexcellent degree of preservation, and great pollen diversity wasdocumented: 17 arboreal types, 15 shrub types, 32 herbaceous types,8 hydro-hygrophyte plant types and 26 NPPs (Fig. 4A–C).
In the sedimentary record of Posidonia oceanica three mainbiozones were distinguished (PL-1 to PL-3), that were in turn dividedinto three subzones (a to c) (Fig. 4A–C).
4.1. Biozone PL-1 (120–77 cm): VIIIth to XIIth centuries AD
Between the VIIIth and XIIth centuries AD, the prevailing arborealvegetation consisted of trees of Quercus pyrenaica type and Quercus
Table 1Radiocarbon dating for the sedimentary record of Portlligat
Depth(cm)
AgeBP
Agecal. AD (2 sigma)
34 710±45 1517–170551 895±45 1402–154563 975±40 1333–146990 1430±40 908–1105119 1600±45 724–954
suber. Their combined percentage (ca. 20%) shows the existence of arelatively open oakwood with cork oaks, in which several mesophileelements, such as the birch (Betula) or the chestnut (Castanea), mighthave been important, just the same as the alder tree (Alnus) among theriparian trees of the region.
In the case of the chestnut, it is known that Castanea sativa wascultivated in the region at least from 2700 BP, thus, in spite of showinglower percentages than 1%, it is likely that its presence in the area isdue to regional cultivation during the VIIIth–XIth centuries AD(Burjachs and Schulte, 2003; Conedera et al., 2004; Burjachs et al.,2005).
The low percentage of Pinus halepensis/pinea type (N5%), even ina coastal area, could be a sign of a short period of development of thecoastal thermomediterraneanpine forests in the area.On theotherhand,the predominant arboreal type was the Pinus sylvestris type, withpercentage that progressively increases from 20% in the base, to morethan 30% just before 1430±40 BP. These data point to a greatdevelopment of the pinewoods (P. sylvestris and P. uncinata) around thecentral-oriental foothills of the Pyrenees (e.g. Sierra del Cadí and Sierradel Catllar), due to the anemophilous character of this palynomorph.
The shrub vegetation shows a remarkable importance, with goodgrowth of heaths of Erica arborea type and Cistus ladanifer rockroses,as well as a xerothermophilous macchia mainly composed of myrtles(Myrtus), Phillyrea and wild olive trees (Olea europaea). Heathers androckroses could be the predominant bushes during the deteriorationstages of the local woodlands, the oak and wild olive forests, and thiscould explain the previously described open-woodland condition, inwhich a wide diversity of shrubs would grow in the clearings. Theanthropization of the area would have favoured the spread of thisshrubland to the detriment of the deciduous forest formations.Furthermore, the xerothermophilous macchia would have beenoccupying the most xeric and sunny sites, over lithosoils or areaswith a scarce edaphic development, where the arboreal elementswould hardly thrive. In fact, the coastal location of this record wouldhave allowed a greater preponderance of the thermophilous elements,such as Myrtus or O. europaea, that usually distinguish the mostthermophilous stages of the wild olive woods. The low values ofQuercus ilex type could be explained by the fact that this kind of forestis poorly represented in the studied area today and, in general, in thenorthern area of Catalonia, where Quercus suber stands out amongother species throughout the landscape.
The xerophilous (Artemisia, Chenopodiaceae/Amaranthaceae) andanthropic (Aster type, Cardueae, Centaurea nigra type, Cichorioideae,Rumex acetosa type, etc.) elements predominate among the herbaceousflora, although it should be noted that some of them are related tograzing activities (Plantago lanceolata type, Urtica dioica type, etc.)(Behre, 1981; Galop, 1998). These anthropic palynomorphs are the bestindicators of the anthropization process of the forests and their

Fig. 4. A. Palynological diagram (trees and shrubs) of Portlligat. B. Palynological diagram (herbs) of Portlligat. C. Palynological diagram (hydro-hygrophytes and NPPs) of Portlligat.
218 J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224

Fig. 4 (continued).
219J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
degradation to open woodlands. Furthermore, the presence of Sordaria(Type 55A) could confirm the impact of grazing, since it is a fungalascospore of coprophilous species (López-Sáez et al., 2000; van Geel,2001).
The percentage predominance of Foraminiferae (Type 700) amongthe NPPs, along with the presence of Tardigrada (Type 902) andAcritarche (Type 966), could indicate that in the chronological contextof the sedimentary record of Portlligat Bay, the prevailing conditionscould have been hyposaline (VanWaveren, 1989; Zamora et al., 2007).In the same way, the presence of Micrhystridium (Type 115) points tothe marine nature of the record (Bakker and van Smeerdijk, 1982). Onthe other hand, the documentation on Glomus cf. fasciculatum shouldbe correlated with erosive processes originated by the direct action ofthe ocean swell on the littoral area (Anderson et al., 1984).
A severe deforestation of the high mountain pine woods (PL-1b)occurred from the Xth and XIth centuries AD through to ca. 1150 AD(82 cm), along with the disappearance of the chestnut and the spreadof heliophilous elements, such as the birch. This process seems to havea negative influence on the oak woods, since the percentages of bothQuercus pyrenaica type and Quercus suber are slightly reduced. Thisprocess induced the spread of heathers (Erica arborea type), junipers(Juniperus type) and, to a certain extent, the pastures of anthropicorigin (Aster type, Cichorioideae, etc). Somehow, it seems that thisdeforestation event was centralized in the pinewoods of the Pyrenees,with minor importance in the littoral and prelittoral areas, althoughthey also seem to be under its influence.
Interestingly, some herbaceous palynomorphs of anthropozoo-genic origin, such as Chenopodiaceae/Amaranthaceae, Plantagolanceolata type or Urtica dioica type, or even Sordaria (Type 55A)among the NPPs, disappear during this PL-1b period, seemingly due toa deceleration in the rate of the local and regional migration of cattle.During this period, the percentages of Type 18, fungal ascospores (vanGeel, 1978), suffer a small initial increase, diminishing abruptly at theend of PL-1b, revealing the advent of a less humid phase (seeMartínez-Cortizas et al., 2005).
From the second half of the XIIth century AD the pine woods andthe deciduous oaks recover their percentages, the chestnut iscultivated again and, for the first time, the beech (Fagus) appears inthe pollen diagram, all of it in the final section of PL-1. Right at the endof this biozone, the percentage of Foraminiferae undergoes a sharpincrease over 100%, that could mean a return to the previoushyposaline conditions and environmental humidity present in mostof PL-1, except for the above mentioned period. The low percentage ofbeech suggests a regional character derived from several populationslocated in the eastern Pyrenees of the Iberian Peninsula.
4.2. Biozone PL-2 (77–45 cm): XIIIth to XVIth centuries AD
Between the XIIIth century AD and the first half of the XVIthcentury AD, the forests are of the same nature as during PL-1, that is,deciduous oakwoods (cork oaks and oak trees), though it seems thatthe oak tree is progressively losing its importance, while the cork tree(Quercus suber) remains constant. The percentage sum of both pollentypes continues to be ca. 20%, again showing a relatively openwoodland, favourable to mesophilous elements, such as the hazel(Corylus) or the birch (Betula), whereas the chestnut (Castanea sativa)would continue to have been cultivated. The most important featurewithin the herbaceous flora is the progressive increase in thepercentage of the high mountain pine woods in the Pyrenees (Pinussylvestris type) through PL-2, reaching a very notable percentage ca.1450–1530 AD (PL-2c) that could be suggesting the gradual reforesta-tion of the Catalan Pyrenees between the XVth andXVIth centuries AD.
Heathers (Erica arborea type) and rockroses (Cistus ladanifer) stillpredominate among the shrubby formations and their persistencecontinued during the substitution stages of the oakwoods. Thexerothermophilous macchia is now reduced only to Phillyrea andPistacia lentiscus, even though Olea europaea is present, but it ispossible that this palynomorph is pointing both to the presence ofwild olive tree, as an element of themacchia, or to the local cultivationof the olive tree.

220 J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
The herbaceous flora is mainly dominated by elements of nitrophi-lous and anthropic nature (Aster type, Cardueae, Cichorioideae) oranthropozoogenic nature (Chenopodiaceae/Amaranthaceae, Plantagolanceolata type, Urtica dioica type) (Behre, 1981; Galop, 1998), withhigher percentages than those occurring during PL-1. This could meanthat the anthropization of these territories through the XIIIth to XVIthcenturies AD reached a greater magnitude than during the previouscenturies, including a notable cattle pressure, as indicated by theimportance of Sordaria (Type 55A) (López-Sáez et al., 2000; van Geel,2001).
By this time, two different NPPs related to erosive processes,Pseudoschizaea circula and Glomus cf. fasciculatum, acquire someimportance, probably in direct relation with the erosion induced bythe ocean swelling on the local coast environment (Anderson et al.,1984) and with local erosive processes due to the anthropization ofthe forest (López-Sáez et al., 2000). The persistence of microforami-niferae (Type 700), Tardigrada (Type 902) and Acritarche (Type 966),with lower percentages, in general, than those occurring during PL-1would suggest the continuity of the hyposaline conditions in Portlligatthrough the XIIIth to XVIth centuries AD, while Micrhystridium's newappearance during PL-2b supports the evidence for the marine natureof the deposit.
The general dynamics proposed for PL-2 seem to be interruptedduring PL-2b (ca. 1380–1450 AD), since there is a reduction in themain arboreal elements, such as Pinus sylvestris type, Betula andQuercus pyrenaica type, or even a temporary disappearance of others,such as the alder and the chestnut. This decrease also affects theshrubby cover, since Cistus ladanifer, Juniperus type and Erica arboreatype all reduce their percentage values, especially the latter. Moreover,Type 18 also reduces its percentage, suggesting drier conditions(Martínez-Cortizas et al., 2005). Coinciding with this change in theclimate, these centuries also seem to be anthropized, since thepercentages of several nitrophilous and anthropic elements, such asCichorioideae, experience a spectacular rise. The economic modelsseem to correspond to amore intense cattle exploitation, since there isan increase in evidence for a greater grazing pressure, with maximumlevels of the anthropozoogenic palynomorphs, such as Plantagolanceolata type and Urtica dioica type (Galop, 1998), and NPPs, suchas Sordaria (Type 55A) and Riccia cf. sorocarpa (López-Sáez et al.,2000; Carrión and Navarro, 2002). The documentation of Chaetomium(Type 7A), fungal ascospores of carbonicolous species (van Geel, 1978;López-Sáez et al., 1998), during this period can be related to thedevelopment of fire, possibly induced to maintain the cattle pastures.In fact, it is probable that these fires and the human impact were thecause of a major level of nutrients in the sediments, indicating achange to mesotrophic conditions, like to indicate some NPPs such asZygnema type, Spirogyra and Zopfia (Bakker and van Smeerdijk, 1982;López-Sáez et al., 1998), or the percentage reduction of Foraminiferae,Tardigrada and Acritarche.
4.3. Biozone PL-3 (45–0 cm): XVIth to XXth centuries AD
From the second half of the XVIth century AD up to the present, thestructure of the forest undergoes very important changes, not only inthe environment immediately next to Portlligat, but also in the wholearea of the eastern Pyrenees.
In the middle of the XVIth century AD a whole series of signsindicating the development of arboricultural and agricultural activ-ities start to appear, in particular, the chestnut (Castanea) seems to becultivated more extensively than before and, at the same time, theculture of the walnut (Juglans), the grapevine (Vitis), the hemp(Cannabis/Humulus) and the cereals (Cerealia) begins; in addition, thepercentages of Olea europaea are increased during PL-3b and PL-3c,suggesting a greater intensity of its cultivation.
During the interval from ca. 1530 to 1700 AD (PL-3a), the abovementioned agricultural activities caused serious damage to the local
cork oak and oak forests, to such an extent that Quercus subercompletely disappears from the record and Quercus pyrenaica typereduces its percentages abruptly. At the same time, Quercus ilex typegains importance, but this fact must not be interpreted as a result ofthe development of its forests, but of a greater percentage representa-tion due to the disappearance of the other deciduous oaks (Riera-Moraand Esteban-Amat, 1994). Such an anthropic process would have leadto a greater development of the shrubby formations, specially thosecomposed of thermophilous elements such as Olea, Phillyrea andlentisc (Pistacia lentiscus), heathers and rockroses. The date of 710±45 BP (1517–1705 AD) would place this period between the middleXVIth and the early XVIIIth centuries AD. It is characterised by arelatively high anthropic impact on the landscape, as the prevalence ofnitrophilous elements and weeds, such as Aster type, Brassicaceae,Campanula type, Cardueae, Centaurea nigra type, Cichorioideae, Con-volvulus arvensis type, Corrigiola, Rumex acetosa type and Spergulaarvensis, demonstrates (Behre, 1981). Some grazing pressure is stillevident since anthropozoogenic elements, such as Chenopodiaceae/Amaranthaceae, Plantago lanceolata type and Urtica dioica type(Galop, 1998), and coprophilous NPPs, such as Sordaria (Type 55A)(López-Sáez et al., 1998), continue appearing in the record.
In the eastern Pyrenees a different situation is found, since most ofthe species, such as the fir (Abies) or the beech (Fagus), or even somemesophilous elements, like the birch (Betula), the hazel (Corylus) orthe lime tree (Tilia), experience a great increase. Undoubtedly, thesedata suggest little anthropic influence on the Pyrenees forests.Furthermore, the major element, Pinus sylvestris type, maintainsrelatively high percentages over 30%, except for two specific minimaduring PL-3b.
During PL-3b (1700–1840 AD) the forests at the Pyrenees weredisturbed, since fir disappears, beech is reduced, and high mountainpinewoods underwent substantial degradation. In parallel, and for thefirst time, the box (Buxus), a submediterranean element, and Ligistrumappear, as components of the degraded shrubby formations. Thechestnut and the walnut, the grapevine and the hemp, the cereals andthe olive tree, were all being cultivated. Percentages of the latter twoare sufficiently high to demonstrate the local existence of crops(López-Sáez et al., 2003). Even the forests of Quercus ilex type areaffected by this higher incidence of agriculture, disappearing from thepollen record, whereas the nitrophilous anthropic and the anthro-pozoogenic elements continue to be very abundant, in particularCichorioideae, Plantago lanceolata type and Sordaria (Type 55A).
The prevalence of agriculture during PL-3a and PL-3b has resultedin erosion of the uppermost level of the soil, distinguished by thepresence of Pseudoschizaea circula and Glomus cf. fasciculatum (López-Sáez et al., 2000; Burjachs and Schulte, 2003). This erosion can also berelated to more arid conditions, as indicated by the importanceacquired by xerophilous elements, such as Artemisia, Ephedradistachya type, Helianthemum type, Phillyrea or Pistacia lentiscus inthese two subzones. The decline of Type 18 in relation to its levelsduring PL-2 would be indicative of the same process (Martínez-Cortizas et al., 2005).
The last section of the pollen diagram (PL-3c), corresponding to thechronological interval between 1840 AD and the present, shows agradual recovery of the oak and cork oak woods, as well as the holmtree woods, due to the complete stoppage of the cultivation ofgrapevine, cereals, hemp and walnut, and to a reduction of thechestnut oak culture and, to a less extent, of the olive tree. Thisrecovery also involves the riparian forest, increasing the localimportance of the alder tree (Alnus) and heliophilous elements, suchas the birch. The anthropization pollen indicators are now less evident,moreover, most of them disappearing or having percentages notablyreduced. Nevertheless, the grazing pressure is still strong, with ahigher intensity than during PL-3b, since certain anthropozoogenicelements, such as Plantago lanceolata type, Urtica dioica type andRiccia cf. sorocarpa, can be identified (Carrión and Navarro, 2002).

221J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
From the point of view of the trophy of the sedimentationmedium,the prevalence of Foraminiferae during PL-3 would continue indicat-ing the presence of hyposaline conditions, mainly mesotrophic duringPL-3a and PL-3c, as a consequence of the contribution of a nutrientbudget to the waters, coming from agricultural activities and theassociated erosive processes. The identification of numerous NPPs(Type 128, Type 181, Mougeotia, Spirogyra, etc.) in both subzoneswould be the most evident sign of the mesotrophic conditions (López-Sáez et al., 2000; Carrión and Navarro, 2002). During PL-3b, whencereals and hemp cultures are at their peaks, the contribution ofnutrients is higher and therefore the conditions have tended towardseutrophy, allowing the documentation of a whole series of NPPs, suchas Types 174 and 425, related to these conditions as well as to a highdegree of landscape anthropization (van Geel et al., 1983).
5. Palaeoenvironmental and palaeoeconomical implications
The palaeopalynological study of the Posidonia oceanica beds atPortlligat shows a high level of diversity, both in pollen and NPPs,which demonstrated the great palaeoecological potential of this kindof submarine deposit. Its relative abundance in the Mediterranean Seaoffers the unique possibility, unknown and unpublished until thisstudy, of tackling similar investigations that could allow a compre-hensive and precise comparison among different coastal environ-ments and their surrounding palaeovegetation. This potentialbecomes of special relevance in those areas of the Mediterraneanshoreline where there is scarce or no previous documentation at all ofthe Holocene environmental evolution, e.g. the province of Girona.
The NE of the Iberian Peninsula is a relatively well-known territory,in relation to the history of its vegetation during the Holocene. Most ofthe available palaeopalynological sequences come from lacustrinerecords or archaeological sites (Burjachs et al., 1990; Riera-Mora andEsteban-Amat, 1994). However, our study is the first one undertakenfrom a marine organic deposit of Posidonia oceanica. It is completelycomparable to other regional terrestrial pollen records (Mallarachet al., 1985; Pérez-Obiol et al., 1991; Riera-Mora and Esteban-Amat,1994; Burjachs et al., 2005), and even to most of the existing recordson the western Mediterranean area (de Beaulieu et al., 2005),demonstrating the great palaeoecological potential of such sediments,in spite of its apparently complex taphonomy (Mateo et al., 1997).
The pollen sequence at Portlligat, an area affected by theMediterranean climate, suggests the presence of a relatively forestedlandscape through the last 1600 years, characterized by oak forma-tions (oaks, cork oaks and holm oaks) in the valleys and the areas nextto the coast, and pine woods, fir woods and beech woods at thePyrenees foothills. The importance of the shrubby formations ofheathers and rockroses in the whole sequence proves the anthro-pization of these territories in the above mentioned chronologicalframe, and this is confirmed by the abundance of nitrophilous andanthropozoogenic herbaceous elements. The equal amount of xer-othermophilous macchia typical bushes (Myrtus, Pistacia lentiscus,Phillyrea, Olea europaea) demonstrates the relationship between theforests of the area with the most thermophilous stage of the cork oakwoodland (Rivas-Martínez, 1987).
The abundance of some NPPs, such as microforaminiferae (Type700), Tardigrada (Type 902) and Acritarche (Type 966), from the baseof the deposit to approximately 45 cm depth (biozones PL-1 and PL-2)indicates the marine bay nature of the sedimentation environment,where hyposaline conditions predominate between the VIIIth centuryAD and the first half of the XVIth century AD. During PL-3, theprogressive reduction and even disappearance of the NPPs, except inthe last 15 cm of the deposit, can be related to a gradual and increasingcontribution of organic and/or inorganic compounds to the littoralareas during the periods of greater human impact.
In general, through the last 1600 years BP, the immediatesurroundings of Portlligat Bay underwent a relatively important
anthropization process, with a particular emphasis on cattle raising.The cultivation of chestnut and possibly olive tree is revealed from theVIIIth century AD on, almost without any interruption. However, thecultivation of walnut, grapevine, hemp and cereals only started fromabout 1530 AD. In contrast with our findings, the cultivation of hempstarted around 600–650 AD in other areas of Catalonia, although itwas not until ca. 1580 AD when it reached its highest diffusion (Rieraet al., 2004, 2006), in accordance with the information obtained fromthe Portlligat record.
The information collected from this study agrees with otherregional pollen records that also demonstrate how during the XVIthand XVIth centuries AD the coastal and Pyrenees areas experiencedmaximum forest exploitation, with high grazing pressure and a peakin agricultural expansion (Riera-Mora and Esteban-Amat, 1994;Pèlachs, 2004; Riera et al., 2004; Burjachs et al., 2005; Riera et al.,2006; Miras et al., 2007). Nevertheless, it is surprising that between1530 AD and 1840 AD the fir, beech and pine woods underwent someexpansion, even when these territories were suffering intensivegrazing activity. It is possible that the establishment of shelter areasfor some of these species, specially the pine and the fir, related to thedevelopment of coalmines, which appeared in the eastern Pyreneesaround these dates (Pèlachs, 2004; Miras et al., 2007), would havefavoured their survival and sustainable development. In the case ofthe beech, the increased anthropization of the Pyrenees could havepromoted its proliferation (Galop, 1998).
From 1850 AD, when the industrial period started, to the present,the anthropic pressure on the coastal environments has decreased,since hemp and cereals are not cultivated any more, allowing therecovery of the oak, cork oak and holm oak woods. Likewise, in thePyrenees the pine wood is recovering but the beechwood onlymaintains its condition, while the fir is greatly reduced.
Four different events are documented in the record of Portlligatdue to the great anthropic impact on the landscape and/orpalaeoenvironmental change.
The first one (82–90 cm), occurring ca. 1000–1100 AD (XIth–XIIthcenturies AD), is characterized by a great deforestation of the highmountain pine wood, the disappearance of the chestnut and a certainreduction of the oak and cork oak woods, while the shrubbyformations (heathers) and some heliophilous arboreal elements(birch) spread; all of which occurred during a decreasing period ofthe grazing pressure. These facts seem to be mediated by an intensedeforestation due to the anthropic impact in the pine woods of thePyrenees, in the NE Iberian Peninsula, since the medieval populationwas notably increased during this period (Bonnassie, 1988, 1990), andimportant social changes took place in the Catalan Pyrenees (Riu,2003). In the same period, there was a climate change towards morearid conditions and an increase in water salinity, since a decline ordisappearance of the NPPs, usually indicatives of hyposaline condi-tions, is observed. This was accompanied by a reduction of grazingpressure in the surrounding areas of Portlligat Bay. Possibly, all thesefacts were related with the beginning of the MWP (Fig. 5) that lastedfrom ca. 900 AD to 1400 AD (Riera et al., 2004; de Beaulieu et al.,2005). A similar event has been documented in the palaeoenviron-mental record of the Estanya Lake (Riera et al., 2004, 2006) between820 and 1075 AD, where the continuous recurrence of anthropic fireswould have induced a great deforestation similar to the one recordedat Portlligat, mostly mediated by a higher impact of cattle grazing andby a decrease in lake level due to the drier weather conditions. Thisevent documented in Portlligat has been illustrated in other pollenrecords in the province of Girona, demonstrating its correlation withthe deforestation of the Pyrenees woods in the NE of the IberianPeninsula from the XIth century AD (Mallarach et al., 1985; Pèlachs,2004).
At 70 cm depth (ca. 1300 AD) of the record something similaroccurs, though much less manifest than the previous one: there is apercentage reduction or even disappearance of most arboreal (alder
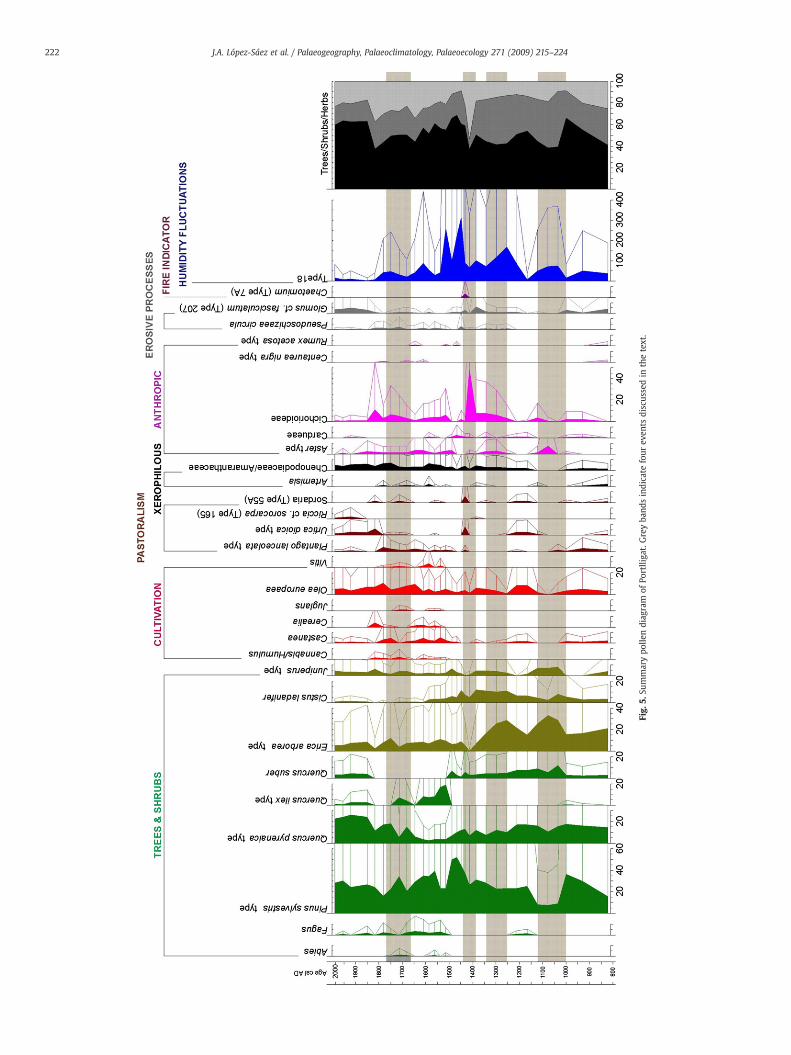
Fig.
5.Su
mmarypo
llendiag
ram
ofPo
rtlliga
t.Greyba
ndsindicate
four
even
tsdiscus
sedin
thetext.
222 J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224

223J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
tree, birch, chestnut, Pinus pinaster, Pinus sylvestris type) and shrubby(Cistus ladanifer, Erica arborea type) palynomorphs; there is also aslight increase in the anthropic herbs (Aster type, Cichorioideae);and a percentage decline in certain NPPs related to humid (Filicalesmonolete, Acari-Oribatei, Type 181, Type 182) or hyposaline (Type700) conditions. This short interval shows the end of local grazingactivity, since the indicative palynomorphs, such as Plantago lanceo-lata type, Urtica dioica type and Sordaria (Type 55A), disappear fromthe record. As awhole, these data suggest an important slowing downof the anthropic pressure on the landscape, particularly of grazing,probably due to a short episode of drier and colder conditions duringthe MWP.
The third notable event takes place ca. 1400 AD (XVth century AD,57–65 cm),when the forest cover (trees and shrubs) undergoes a declinethat seems to be mediated by the higher incidence of fire processes ofanthropic origin, due to an increase of cattle raising and, in general, to agreater anthropizationof the landscape in the studied area. These eventsinduced the change in the nutrient budget in the sediments, resulting inmesotrophic conditions during this period. Other pollen records in theNE Iberian Peninsula (Riera et al., 2004, 2006) suggest an increase oflacustrine levels between 1360 and 1580 AD, coincidental with the firstcold episode, with low solar activity, of the Little Ice Age (1400–1850AD), the so called SpörerMinimum(1400–1530AD, Fig. 5) (Despratet al., 2003; de Beaulieu et al., 2005).
In general, from a social point of view, the XIVth to XVIth centuriesAD entail a demographic and economic upheaval period in most ofEurope, due to the high incidence of diseases and epidemics, whichled to a lower human impact on the forests (Desprat et al., 2003). InCatalonia, the crisis that took place during the LateMiddle Ages beginsto be discerned from 1333 AD – the first ‘great year of famine’–,extending to 1347–1351 AD, along with the first surges of the BlackDeath, that lasted until the late XVth century AD, with deep economicand social crisis (Rodríguez-Puertolas, 1982; Riu, 2003; Feliú, 2004). Inthe pollen record of Portlligat these events are clearly noticeable. Inthat, except for the effects of the Spörer Minimum, the forests aremaintained through the XIIIth to XVIth centuries AD, specially in thePyrenees where the pine woods experienced an important expansionbetween the XVth and the XVIth centuries AD, facilitated by thedepopulation of the Catalan Pyrenees (Pèlachs, 2004). Such a criticalperiod triggered a depletion of the agricultural areas and a greatemigration process (Feliú, 2004), which, from a palaeoenvironmentalpoint of view, is demonstrated by a decreased anthropization of thelandscape.
A fourth outstanding event is documented between the 30 and23 cm depth of in record (ca.1650–1750 AD), at the transition betweenthe PL-3a and PL-3b biozones. In this interval, the chestnut culture isabruptly decreased and grapevines are also affected, while cerealcultivation is completely stopped; the percentages of several arborealelements (alder tree, birch, hazel, beech, pine, oak) are reduced oreven disappear, in contrast with the percentages of the indicators ofcolder andmore arid conditions (fir, holm oak), including some shrubs(Phillyrea). Among the herbaceous flora, the anthropization signs arescarcely evident, and only a minimum percentage of Foraminiferae,showing high saline conditions, could be detected. In summary, thisdata would point out towards a relatively short period with cold andarid conditions, during the Little Ice Age, in which the landscapeanthropization is suddenly diminished, thus involving the slowingdown of agricultural expansion, or even the abandonment of somecrops.
6. Conclusions
The highly organic marine litter generated by Posidonia oceanicagives rise to new research perspectives on palaeoenvironmentalstudies, especially on those of a palaeopalynological nature. Theassessment carried out in Portlligat Bay demonstrates the enormous
value of these formations, similar to the terrestrial peat-bogs, allowingthe reconstruction of palaeovegetation throughout the last 1600 years.This interest is increased due to the sensitivity of these deposits torecord several phases of human impact and palaeoenvironmentalchanges that are related to social crisis and different palaeoeconomicstrategies.
Acknowledgments
This work was supported by the PALEOMED and CARBOMEDprojects, respectively funded by the Spanish Ministry of Science andTechnology (McyT-BOS2002-02247) and the Spanish NationalResearch Council (CSIC) (PIF-CSIC 20050F0231). LLM has benefitedfrom a CSIC-ESF research contract (I3P predoctoral program). We aregrateful to Peter Kershaw, José Carrión and an anonymous reviewerfor their constructive comments. Peter Kershaw corrected the finalEnglish version of the manuscript.
References
Aaby, B., Berglund, B.E., 1986. Characterization of peat and lake deposits. In: Berglund, B.E.(Ed.), Handbook of Holocene Palaeoecology and Palaeohydrology. JohnWiley & Sons,Chichester, pp. 231–246.
Anderson, R.S., Homola, L., Davis, R.B., Jacobson Jr., G.L., 1984. Fossil remains of themycorrhizal fungal Glomus fasciculatum complex in postglacial lake sediments fromMaine. Canadian Journal of Botany 62, 2325–2328.
Bakker, M., van Smeerdijk, D.G., 1982. A palaeoecological study of a Late Holocenesection from “Het Ilperveld”, Western Netherlands. Review of Palaeobotany andPalynology 36, 95–164.
Behre, K.E., 1981. The interpretation of anthropogenic indicators in pollen diagrams.Pollen et Spores 23, 225–245.
Bethoux, J.P., Copin-Montegut, G., 1986. Biological fixation of atmospheric nitrogen inthe Mediterranean sea. Limnology and Oceanography 31, 1353–1358.
Bonnassie, P., 1988. Cataluña mil años atrás (siglos X–XI). Península Ediciones 62,Barcelona, 448 pp.
Bonnassie, P., 1990. La Catalogne au tournant de l'an mil: croissance et mutations d'unesociété. Albin Michel, Paris. 277 pp.
Boudouresque, C.F., Giraud, G., Thommeret, J., Thommeret, Y., 1980. First attempt atdating by 14C the undersea beds of dead Posidonia oceanica in the Bay of Port-Man(Port-Cros, Var, France). Travaux Scientifiques du Parc National de Port-Cros 6,239–242.
Burjachs, F., Schulte, L., 2003. El paisatge vegetal del Penedès entre la Prehistòria i elMónAntic. In: Guitart, J., Palet, J.M., Prevosti, M. (Eds.), Paisatges antics a laMediterrània ia la Cossetània oriental. Generalitat de Catalunya, Barcelona, pp. 249–254.
Burjachs, F., Esteban, A., Génova, R., Julià, R., Llasat, C., Montserrat, J., Parra, I., PérezObiol, R., Roure, J.M., Vilaplana, J.M., 1990. Vegetación y paleoclimatología en el N.E.peninsular durante el Tardi y Postglacial. In: Blanca, G., Díaz de, C., la Guardia,Fernández, M.C., Garrido, M., Rodríguez García, M.C., Romero García, A.T. (Eds.),Polen, esporas y sus aplicaciones. Universidad de Granada, Granada, pp. 465–470.
Burjachs, F., Bach, J., Buxó, R., Llàcer, P., MacGlade, J., Picazo, M., Piqué, R., Ros, M.T., 2005.El territori d'Emporion i les seves dades paleoambientals. Empúries 54, 25–32.
Carrión, J.S., 2001. Dialectic with climatic interpretations of Late-Quaternary vegetationhistory in Mediterranean Spain. Journal of Mediterranean Ecology 2, 145–156.
Carrión, J.S., Navarro, C., 2002. Cryptogam spores and other non-pollen microfossils assources of palaeoecological information: case-studies from Spain. Annales BotaniciFennici 39, 1–14.
Carrión, J.S., Munuera, M., Navarro, C., Burjachs, F., Dupré, M., Walker, M.J., 1999. Thepalaeoecological potential of pollen records in caves: the case of MediterraneanSpain. Quaternary Science Reviews 18, 1061–1073.
Conedera, M., Krebs, P., Tinner, W., Pradella, M., Torriani, D., 2004. The cultivation ofCastanea sativa (Mill.) in Europe, from its origin to its diffusion on a continentalscale. Vegetation History and Archaeobotany 13, 161–179.
Damblon, F., Vanden Berghen, C.V., 1993. Étude paléo-écologique (pollen et macro-restes) d'un dêpot tourbeux dans l'île de Djerba, Tunisie méridionale. Palynos-ciences 2, 157–172.
de Beaulieu, J.L., Miras, Y., Andrieu-Ponel, V., Guiter, F., 2005. Vegetation dynamics innorth-western Mediterranean regions: instability of the Mediterranean bioclimate.Plant Biosystems 139 (2), 114–126.
Desprat, S., Sánchez-Goñi, M.F., Loutre, M.F., 2003. Revealing climatic variability of thelast three millennia in northwestern Iberia using pollen influx data. Earth andPlanetary Science Letters 213, 63–78.
Faegri, K., Iversen, J., 1989. Textbook of Pollen Analysis. John Wiley & Sons, Chichester.328 pp.
Feliú, G., 2004. La crisis catalana de la Baja Edad Media: estado de la cuestión. Hispania64 (2), 435–466.
Galop, D., 1998. La forêt, l'homme et le troupeau dans les Pyrénées. 6000 ans d'histoirede l'environnement entre Garonne et Méditerranée. GEODE, Laboratoire d'EcologieTerrestre et FRAMESPA, Toulouse, 322 pp.

224 J.A. López-Sáez et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 271 (2009) 215–224
Grimm, E.C., 1987. Coniss: a Fortran 77 program for stratigraphically constrainedcluster analysis by the method of incremental sum of squares. Computers &Geosciences 13 (1), 13–35.
Grimm, E., 1992. Tilia, Version 2. USA Illinois State Museum, Research and CollectionCenter, Springfield. IL 62703.
Karlen, I., Olsson, I.U., Kållberg, P., Killiççi, S., 1968. Absolute determination of theactivity of two 14C dating standards. Arkiv Geofysik 4, 456–471.
López-Sáez, J.A., van Geel, B., Farbos-Texier, S., Diot, M.F., 1998. Remarques paléoéco-logiques à propos de quelques palynomorphes non-polliniques provenant desédiments quaternaires en France. Revue de Paléobiologie 17 (2), 445–459.
López-Sáez, J.A., van Geel, B., Martín Sánchez, M., 2000. Aplicación de los microfósilesno polínicos en Palinología Arqueológica. In: Oliveira-Jorge, V. (Ed.), Contributos dasCiências e das Technologias para a Arqueologia da Península Ibérica. Actas 3°Congresso de Arqueologia Peninsular, vol. IX, Vila-Real, Portugal, setembro de 1999.Adecap, Porto, pp. 11–20.
López-Sáez, J.A., López-García, P., Burjarchs, F., 2003. Arqueopalinología: Síntesis Crítica.Polen 12, 5–35.
López-Sáez, J.A., López-García, P., Pozuelo, R., Mateo, M.A., Renom, P., Garrido, D., 2006.Non-pollen palynomorphs from organic deposits of Posidonia oceanica: a new toolfor palaeoenvironmental studies in marine peat-like deposits. Palyno-Bulletin 2,38–40.
Mallarach, J.M., Pérez-Obiol, R., Roure, J.M.,1985. Aportaciones al conocimiento del climay la vegetación durante el Cuaternario reciente, en el N.E. de la Península Ibérica.Actas I Reunión del Cuaternario Ibérico. G.E.T.C.-G.T.P.E.Q, Lisboa, pp. 201–212.
Martínez Cortizas, A., Mighall, T., Pontevedra Pombal, X., Nóvoa Muñoz, J.C., PeiteadoVarela, E., Piñeiro Rebolo, R., 2005. Linking changes in atmospheric dust deposition,vegetation change and human activities in northwest Spain during the last5300 years. The Holocene 15 (5), 698–706.
Mateo, M.A., Romero, J., Pérez, M., Littler, M.M., Littler, D.S., 1997. Dynamics of millenaryorganic deposits resulting from the growth of theMediterranean seagrass Posidoniaoceanica. Estuarine, Coastal and Shelf Science 44, 103–110.
Mateo, M.A., Renom, P., Julià, R., Romero, J., Michener, R., 2002. An unexploredsedimentary record for the study of environmental change in Mediterraneancoastal environments: Posidonia oceanica (L.) Delile peats. The Study of Environ-mental Change Using Isotope Techniques. International Atomic Energy Agency C&SPapers Series 16P, Vienna, pp. 163–173.
Miras, Y., Ejarque, A., Riera, S., Palet, J.M., Orengo, H., Euba, I., 2007. Dynamique holocènede la végétation et occupation des Pyrénées andorranes depuis le Néolithiqueancien, d'après l'analyse pollinique de la tourbière de Bosc dels Estanyons (2180 m,Vall del Madriu, Andorre). Comptes Rendus Palévol 6, 291–300.
Moore, P.D., Webb, J.A., Collinson, M.E., 1991. Pollen Analysis, 2nd. ed. BlackwellScientific Publications, London. 216 pp.
Pèlachs, A., 2004. Deu mil anys de geohistòria ambiental al Pirineu central català. Ph.D.Thesis, Universitat Autònoma de Barcelona, Bellaterra, Spain, 581 pp.
Pérez-Obiol, R., Roure, J.M., Vilar, L., 1991. History of some plant communities of the Silszone (La Selva, Catalonia, Spain) based on pollen analysis. Scientia gerundensis 17,37–44.
Reille, M., 1992. Pollen et Spores d' Europe et d' Afrique du Nord. Laboratorie deBotanique Historique et Palynologie, Marseille. 543 pp.
Reille, M., 1995. Pollen et Spores d' Europe et d' Afrique du Nord. Supplement 1.Laboratorie de Botanique Historique et Palynologie, Marseille. 331 pp.
Riera, S., Wansard, G., Julià, R., 2004. 2000-year environmental history of a karstic lakein the Mediterranean Pre-Pyrenees: the Estanya lakes (Spain). Catena 55, 293–324.
Riera, S., López-Sáez, J.A., Julià, R., 2006. Lake responses to historical land use changes innorthern Spain: the contribution of non-pollen palynomorphs in a multiproxystudy. Review of Palaeobotany and Palynology 141, 127–137.
Riera-Mora, S., Esteban-Amat, A., 1994. Vegetation history and human activity duringthe last 6000 years on the central Catalan coast (northeastern Iberian Peninsula).Vegetation History and Archaeobotany 3, 7–23.
Riu, M., 2003. La vivienda rural en el Prepirineo y Pirineo catalán (siglos XI al XVI).Scripta Nova 7 (146), 1–5.
Rivas-Martínez, S., 1987. Memoria del MAPA de Series de Vegetación de España.Ministerio de Agricultura, Pesca y Alimentación, ICONA, Madrid, 268 pp.
Rodríguez-Puertolas, J., 1982. La crisis de la Baja Edad Media calatana y la poesía de laépoca. In: Bustos, E. (Ed.), Actas del Cuarto Congreso Internacional de Hispanistas.Universidad de Salamanca, Salamanca, pp. 487–498.
Romero, J., Pérez, M., Mateo, M.A., Sala, E., 1994. The below ground organs of theMediterranean seagrass Posidonia oceanica as a biogeochemical sink. AquaticBotany 47, 13–19.
Rossignol-Strick, M., 1999. The Holocene climatic optimum and pollen records ofsapropel 1 in the eastern Mediterranean, 9000–6000 BP. Quaternary ScienceReviews 18, 515–530.
Rull, V., 2001. Origen y evolución de los manglares del Caribe: evidencias palinológicas.In: Moreno, S. (Ed.), XIII Simposio de la Asociación de Palinólogos de LenguaEspañola (A.P.L.E.). Universidad Politécnica de Cartagena, Cartagena, pp. 333–349.
Stuiver, M., Braziunas, T., 1993. Modeling atmospheric 14C influences and 14C ages ofmarine samples to 10,000 BC. Radiocarbon 35, 137–189.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen, K.A., Kromer, B.,McCormac, G., van der Plicht, J., Spurk, M., 1998. INTCAL98 radiocarbon agecalibration 24000–0 cal BP. Radiocarbon 40 (3), 1041–1083.
Tzedakis, P.C., Andrieu, V., de Beaulieu, J.L., Crowhurst, S., Follieri, M., Hooghiemstra, H.,Magri, D., Reille, M., Sadori, L., Shackleton, N.J., Wijmstra, T.A., 1997. Comparison ofterrestrial and marine records of changing climate of the last 500,000 years. Earthand Planetary Science Letters 150, 171–176.
van Geel, B., 1978. A palaeoecological study of Holocene peat bog sections in Germanyand The Netherlands. Review of Palaeobotany and Palynology 25, 1–20.
van Geel, B., 2001. Non-pollen palynomorphs. In: Smol, J.P., Birks, H.J.B., Last, W.M.(Eds.), Tracking Environmental Change Using Lake Sediments. Terrestrial, Algal, andSiliceous Indicators, vol. 3. Kluwer Academic Publishers, Dordrecht, pp. 99–119.
van Geel, B., Hallewas, D.P., Pals, J.P., 1983. A Late Holocene deposit under theWestfrieseZeedijk near Enkhuizen (Prov. of Noord Holland, The Netherlands): palaeoecolo-gical and archaeological aspects. Review of Palaeobotany and Palynology 38,269–335.
Van Waveren, I.M., 1989. Pattern analysis of organic component abundances fromdeltaic and open marine deposits: palynofacies distribution (East Java, Indonesia).Netherlands Journal of Sea Research 23 (4), 441–447.
Zamora, N., Medeanic, S., Corrêa, I.C.S., 2007. Microforaminíferos como indicadorespaleoambientais: Estudos palinológicos da região sul da Costa Rica, AméricaCentral. Gravel 5, 75–87.
Related Documents







![Lack of Impact of Posidonia oceanica Leaf Nutrient ...Introduction In the Western Mediterranean, seagrass meadows are dominated by Posidonia oceanica (L.) Delile [1,2] where the fish](https://static.cupdf.com/doc/110x72/5e36a23aece3670988495fb0/lack-of-impact-of-posidonia-oceanica-leaf-nutrient-introduction-in-the-western.jpg)



