470 AAP Memoir 23 (2000) Saunders, J.B. & Perch-Neilsen, K. (eds), Plankton Stratigraphy, Cambridge University Press, Cambridge. Willis, P.A., 1995.Phylogeny ofAustralian crocodiles. Unpublished PhD thesis, University of New South Wales, Sydney. Willis, P.A., 1997. New crocodilians from the .late Miocene White Hunter Site, Riversleigh, northwestern Queensland. Memoirs of the Queensland Museum 41, 423-37. Wilson, G.J., 1967a. Some new species of Lower Tertiary dinoflagellates from McMurdo Sound, Antarctica. New Zealand Journal of Botany 5, 57- 83. Wilson, G.J., 1967b. Some new species of Wetzeliella Eisenack (Dinophyceae) from New Zealand Eocene and Paleocene strata. New Zealand Journal of Botany 5, 469-497. Wilson, G.J., 1984. New Zealand Late Jurassic to Eocene dinoflagellate biostratigraphy -A summary. Newsletters in Stratigraphy 13(2), 104-117. Wilson, G.J., 1988. Palaeocene and Eocene dinoflagellate cysts from Waipawa, Hawkes Bay, New Zealand. New Zealand Geological Survey Paleontological Bulletin 57, 1-96. Wilson, G.J. & Clowes, C.D., 1982.Arachnodinium, a new dinoflagellate genus from the Lower Tertiary of Antarctica. Palynology 6, 97-103. Woodburne, M.O. & Case, J.A., 1996. Dispersal, vicariance, and the late Cretaceous to early Teriary land mammal biogeography from South America to Australia. Journal of Mammalian Evolution 3, 121- 61. Woodbume, M.O., Macfadden, B.J., Case, J.A., Springer, M.S., Pledge, N.S., Power, J.D., Woodburne, J.M. & Springer, K.B., 1994. Land mammal biostratigraphy and magnetostratigraphy of the Etadunna Formation (late Miocene) of South Australia. Journal of Vertebrate Paleontology 13, 483-515. Woodburne, M:O., Tedford, R.H., Archer, M., Turnbull, W., Plane, M. & Lundelius, E.L., 1985. Biochronology of the continental mammal record of Australia and New Guinea. South Australian Department of Mines and Energy Special Publication 5, 347-65. Wrenn, J.H. & Beckmann, S.W., 1982. Macera], total organic carbon, and palynological analyses of Ross Ice Shelf Project Site J-9 cores. Science 216, 187- 189. Wrenn, J.H. & Hart, G .F., 1988. Paleogene dinoflagellate cyst biostratigraphy of Seymour Island, Antarctica. Geological Society of America Memoir 169, 321-447. Yassini, I. & Jones, B.G., 1995.Recent Foraminiferida and Ostracoda from estuarine and shelf 10 Palaeobiogeography of the Quaternary of Australasia P. KERSHAW 1 , P.G. QUILTY 2 , B. DAVID', S. VAN HUET 3 & A. McMINN 4 1 School of Geography and Environmental Science, Monash University, Vic. 3800 Australia 2 School of Earth Sciences, University of Tasmania, Box 252-79, Hobart Tas. 7050 Australia 3 Department of Earth Sciences, Monash University, Vic. 3800 Australia 4 Institute of Antarctic and Southern Ocean Studies, University of Tasmania, Box 252C, Hobart, Tas. 7001 Australia Wing, S.L., Sues, H.-D., Potts, R, DiMichele, W.A. & Behrensmeyer, A.K., 1992. Evolutionary paleoecology. 1-13 in Behrensmeyer, A.K., Damuth, J.D., DiMichele, W.A., Potts, R., Sues, H.-D. & Wmg, S.L., 1992. Terrestrial ecosystems through time: evolutionary paleoecology of terrestrialplants and animals . The Evolution of Terrestrial Ecosystems Consortium, University of Chicago Press, Chicago. Wood, G.R., 1986. Late Oligocene to Early Miocene palynomorphs from GSQ Sandy Cape 1-3R. Geological Survey of Queensland, Publication 387, 1-27. environments on the southeastern coast ofAustralia. University of Wollongong Press, Wollongong, 1-484. Yassini, I.,Jones, B.G. & Jones M.R., 1993. Ostracods from the Gulf of Carpentaria, northeastern Australia. Senkenbergiana lethaea 73(2), 375-406. Zachos, J.C., Quinn, T.M. & Salamy, K.A., 1996. High- resolution (1o4 years) deep-sea foraminiferal stable isotope records of the Eocene-Oligocene climate transition. Paleoceanography, 11, 251-266. Zachos, J.C., Stott, L.D. & Lohmann, K.D., 1994. Evolution of early Cenozoic marine temperatures. Paleoceanography 9, 353-387. THE QUATERNARY is characterised by dramatic changes in climate that relate to the most recent interval of glaciation on the globe. These changes appear to be triggered by small variations in the amount and distribution of solar radiation received by the earth as a result of predictable changes in the earth's orbit relative to the sun, known as Milankovitch cycles (Imbrie et al., 1984). Milankovitch cyclicity is believed to have occurred through the whole of geological time and has been demonstrated to have operated through at least the last 6 Ma (million years) (Shackleton et al., 1995). The significance of these cycles in the Quaternary is their great amplitude of change which results from the complex relationships between Milankovitch forcing, the distribution of land masses, the growth of mountains and high plateaux, the pattern of ocean circulation, ice sheet growth and decay and substantial changes in carbon dioxide and other 'Greenhouse' gases in the atmosphere (Williams et al., 1998). In the absence of significant changes in the position of, and relationship between, continental masses due to the limited time-span of the Quaternary, natural, terrestrial biogeographic patterns tend to be dominated by these climate changes. However, the movement northwards of Australia by about 100 km over this period may have had some influence on oceanic and atmospheric circulation and biotic migration patterns within the Australian and Asian regions, and episodic lowering of sea sea level up to 130m (Ferland et al., 1995) would have had an influence on both terrestrial and marine migration and isolation. During the last glaciation, both Tasmania and New Guinea were joined to the Australian mainland and ocean currents were deflected with the loss of Bass and Torres Straits. Throughout the world, the exposure of continental shelves resulted in the loss of shallow marine environments and no more significantly than in the maritime continent region between northern Australia and mainland Southeast Asia, which holds a central position in regional and perhaps global atmospheric and oceanic circulation patterns. Similar effects would have been felt in New Zealand as islands were joined during low sea level phases and the land area increased by 50%. Incontrast to the stable Australian mainland, the islands of New Guinea and New Zealand have also experienced marked physiographic changes with substantial tectonic activity and volcanism. KERSHAW, P., QUILTY, P.G., DAVID, B., VAN HUET, S. & McMINN, A., 2000: 12:20. Palaeobiogeography of the Quaternary of Australasia. Memoir of the Association of Australasian Palaeontologists 23, 471-516. ISSN 0810-8889

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

470 AAP Memoir 23 (2000)
Saunders, J.B. & Perch-Neilsen, K. (eds), Plankton Stratigraphy, Cambridge University Press, Cambridge.
Willis, P.A., 1995.Phylogeny ofAustralian crocodiles. Unpublished PhD thesis, University of New South Wales, Sydney.
Willis, P.A., 1997. New crocodilians from the .late Miocene White Hunter Site, Riversleigh, northwestern Queensland. Memoirs of the Queensland Museum 41, 423-37.
Wilson, G.J., 1967a. Some new species of Lower Tertiary dinoflagellates from McMurdo Sound, Antarctica. New Zealand Journal of Botany 5, 57- 83.
Wilson, G.J., 1967b. Some new species of Wetzeliella Eisenack (Dinophyceae) from New Zealand Eocene and Paleocene strata. New Zealand Journal of Botany 5, 469-497.
Wilson, G.J., 1984. New Zealand Late Jurassic to Eocene dinoflagellate biostratigraphy -A summary. Newsletters in Stratigraphy 13(2), 104-117.
Wilson, G.J., 1988. Palaeocene and Eocene dinoflagellate cysts from Waipawa, Hawkes Bay, New Zealand. New Zealand Geological Survey Paleontological Bulletin 57, 1-96.
Wilson, G.J. & Clowes, C.D., 1982.Arachnodinium, a new dinoflagellate genus from the Lower Tertiary of Antarctica. Palynology 6, 97-103.
Woodburne, M.O. & Case, J.A., 1996. Dispersal, vicariance, and the late Cretaceous to early Teriary land mammal biogeography from South America to Australia. Journal of Mammalian Evolution 3, 121- 61.
Woodbume, M.O., Macfadden, B.J., Case, J.A., Springer, M.S., Pledge, N.S., Power, J.D., Woodburne, J.M. & Springer, K.B., 1994. Land mammal biostratigraphy and magnetostratigraphy of the Etadunna Formation (late Miocene) of South Australia. Journal of Vertebrate Paleontology 13, 483-515.
Woodburne, M:O., Tedford, R.H., Archer, M., Turnbull, W., Plane, M. & Lundelius, E.L., 1985. Biochronology of the continental mammal record of Australia and New Guinea. South Australian Department of Mines and Energy Special Publication 5, 347-65.
Wrenn, J.H. & Beckmann, S.W., 1982. Macera], total organic carbon, and palynological analyses of Ross Ice Shelf Project Site J-9 cores. Science 216, 187- 189.
Wrenn, J.H. & Hart, G .F., 1988. Paleogene dinoflagellate cyst biostratigraphy of Seymour Island, Antarctica. Geological Society of America Memoir 169, 321-447.
Yassini, I. & Jones, B.G., 1995.Recent Foraminiferida and Ostracoda from estuarine and shelf
10
Palaeobiogeography of the Quaternary of Australasia
P. KERSHAW 1, P.G. QUILTY2, B. DAVID', S. VAN HUET3 & A. McMINN4
1 School of Geography and Environmental Science, Monash University, Vic. 3800 Australia 2 School of Earth Sciences, University of Tasmania, Box 252-79, Hobart Tas. 7050 Australia 3 Department of Earth Sciences, Monash University, Vic. 3800 Australia 4 Institute of Antarctic and Southern Ocean Studies, University of Tasmania, Box 252C, Hobart, Tas.
7001 Australia
Wing, S.L., Sues, H.-D., Potts, R, DiMichele, W.A. & Behrensmeyer, A.K., 1992. Evolutionary paleoecology. 1-13 in Behrensmeyer, A.K., Damuth, J.D., DiMichele, W.A., Potts, R., Sues, H.-D. & Wmg, S.L., 1992. Terrestrial ecosystems through time: evolutionary paleoecology of terrestrialplants and animals . The Evolution of Terrestrial Ecosystems Consortium, University of Chicago Press, Chicago.
Wood, G.R., 1986. Late Oligocene to Early Miocene palynomorphs from GSQ Sandy Cape 1-3R. Geological Survey of Queensland, Publication 387, 1-27.
environments on the southeastern coast ofAustralia. University of Wollongong Press, Wollongong, 1-484.
Yassini, I.,Jones, B.G. & Jones M.R., 1993. Ostracods from the Gulf of Carpentaria, northeastern Australia.
Senkenbergiana lethaea 73(2), 375-406. Zachos, J.C., Quinn, T.M. & Salamy, K.A., 1996. High-
resolution (1o4 years) deep-sea foraminiferal stable isotope records of the Eocene-Oligocene climate transition. Paleoceanography, 11, 251-266.
Zachos, J.C., Stott, L.D. & Lohmann, K.D., 1994. Evolution of early Cenozoic marine temperatures. Paleoceanography 9, 353-387.
THE QUATERNARY is characterised by dramatic changes in climate that relate to the most recent interval of glaciation on the globe. These changes appear to be triggered by small variations in the amount and distribution of solar radiation received by the earth as a result of predictable changes in the earth's orbit relative to the sun, known as Milankovitch cycles (Imbrie et al., 1984). Milankovitch cyclicity is believed to have occurred through the whole of geological time and has been demonstrated to have operated through at least the last 6 Ma (million years) (Shackleton et al., 1995). The significance of these cycles in the Quaternary is their great amplitude of change which results from the complex relationships between Milankovitch forcing, the distribution of land masses, the growth of mountains and high plateaux, the pattern of ocean circulation, ice sheet growth and decay and substantial changes in carbon dioxide and other 'Greenhouse' gases in the atmosphere (Williams et al., 1998).
In the absence of significant changes in the position of, and relationship between, continental masses due to the limited time-span of the Quaternary, natural, terrestrial biogeographic patterns tend to be dominated by these climate
changes. However, the movement northwards of Australia by about 100 km over this period may have had some influence on oceanic and atmospheric circulation and biotic migration patterns within the Australian and Asian regions, and episodic lowering of sea sea level up to 130m (Ferland et al., 1995) would have had an influence on both terrestrial and marine migration and isolation. During the last glaciation, both Tasmania and New Guinea were joined to the Australian mainland and ocean currents were deflected with the loss of Bass and Torres Straits. Throughout the world, the exposure of continental shelves resulted in the loss of shallow marine environments and no more significantly than in the maritime continent region between northern Australia and mainland Southeast Asia, which holds a central position in regional and perhaps global atmospheric and oceanic circulation patterns. Similar effects would have been felt in New Zealand as islands were joined during low sea level phases and the land area increased by 50%. Incontrast to the stable Australian mainland, the islands of New Guinea and New Zealand have also experienced marked physiographic changes with substantial tectonic activity and volcanism.
KERSHAW, P., QUILTY, P.G., DAVID, B., VAN HUET, S. & McMINN, A., 2000: 12:20. Palaeobiogeography of the Quaternary of Australasia. Memoir of the Association of Australasian Palaeontologists 23, 471-516. ISSN 0810-8889

472
The short timescale of the Quaternary is significant also for questions related to the evolution of organisms. Evolutionary rates of organisms as directly observed from stratigraphic ranges within the fossil record , indicate that evolutionary change is unlikely to have been a major factor in Quaternary biogeography at least at the species level. This conclusion is supported by an examination of the biological response of biota to Milankovich forcing (Bennett 1997). He determined that organisms effectively responded to dramatic climatic fluctuations by migrating and that there was no notable change in evolutionary rates during the Quaternary period. However, there is likely to have been a significant degree of evolutionary change at the subspecies level, morphologically invisible in many fossil taxa, and in species ecology. Furthermore, one aspect of evolution which is very evident in the Quaternary is extinction and here the role of people has to be taken into account.
The unique feature of the Quaternary is that it embraces the evolution of bipedal, toolmaking, fire-using hominids (Williams et al., 1998) who
· were capable of broad dispersal and also of transporting other components of the biota, as well as inflicting an unprecedented impact on native biotas. Although the bulk of human evolution is considered to have taken place on the African continent, there is strong evidence for the presence of Homo erectus in Southeast Asia by 1.8 Ma. (Swisher et al., 1994) and increasing evidence that individuals had crossed Wallace's line, and hence made sea crossings, to the Lesser Sunda Island of Flores by 800 -700 ka (Sondaar et al., 1994; Morwood et al., 1998). However, there is no indication that this migration continued into Australia and New Guinea, probably until between 60 and 50 ka, well after the emergence of Homo sapiens. The subsequent colonisation of most of the southwest Pacific occurred only after 4 ka (Enright & Gosden, 1992) and in New Zealand, after 1ka, and perhaps as late as 700 BP (Anderson & McGlone, 1992).
Palaeobiological study of the Quaternary has been dominated by a desire to document and understand patterns of climate change, and the impact of people, and consequently to assess the relative roles of climate and people on environmental change and the development of the present landscape . There has been a strong focus on .the more recent part of this period when human impact has been greatest and where the availability of Continuously accumulating sediments, readily datable by methods such as radiocarbon, has allowed the reconstruction of detailed records interpi-etable from present day analogues. This 'top down' approach has generally failed to penetrate
AAP Memoir 23 (2000)
far into the Quaternary because oflack of interest in older environments, the paucity of dating methods and/or dating precision, the discontinuous nature of the sediment record, and increasing uncertainty about the ecological or environmental meaning of recorded assemblages. On the other hand, the conventional 'bottom up' approach of geologically-oriented palaeontologists has generally faltered when it comes to the Quaternary because oflow turnover rates relative to the length of the period, and a lack of economic incentive. Only recently, with a developing global concern about future climates, has there been a general realisation of the significance of Quaternary patterns of environmental change as a whole.
This review is inevitably biased in its treatment of the Quaternary. We first provide some overview of the status of global stratigraphy, developed independently of Australasian evidence, to provide a context for discussion of regional data. This is followed by an examination of fossil groups which have been studied sufficiently in biostratigraphy or palaeoecology to provide useful biogeographic insights or groups which have the potential to do so. Evidence for humans is treated separately because of the distinct influence of this taxon group on biogeographic patterns of other groups of biota. Regional biogeographic analysis is focussed initially on the Last Glacial Maximum and Holocene 'Optimum' as the best known examples of the environmental extremes which characterised at least the latter part of the Quaternary, followed by a more generalised examination of the whole Quaternary with emphasis on terrestrial flora and on those few areas where stratigraphical and biological records are most substantial.
GLOBALQUATERNARYSTRATIGRAPHY
Debate over the subdivision of the Quaternary, or even whether the Quaternary has legitimate geological status at all (Berggren et al., 1995), is on-going (P.>ftridge, 1997a). However, there is some agreement that the period covered by the Quaternary includes an earlier Pleistocene epoch, which embraces the glacial and interglacial periods and can be subdivided into the Early, Middle and Late Pleistocene, and the Holocene, the latter defining the period from the end of the last glaciation. The beginning of the Holocene, defined by radiocarbon dating at 10 ka, marks the time when global temperatures achieved approximately those of today, and is generally easy to recognise in palaeorecords. The Late Pleistocene covers the last glacial cycle from the beginning of the Last Interglacial (c.132 ka BP) to the Pleistocene- Holocene boundary, whereas the base of the Middle Pleistocene marks the beginning of
AAP Memoir 23 (2000)
consistent high amplitude glacial/interglacial oscillations and corresponds with the last magnetic reversal from the Matuyama chron to the Brunhes chron. The base of the Early Pleistocene, and hence the Quaternary, is considered to mark the first evidence of cooling, as determined by marine fauna in the Quaternary section of Vrica, Italy, which is close to the top of the Olduvai palaeomagnetic event or subchron dated to 1.76Ma (Aguirre & Pasini, 1985).
In recent years, a detailed global stratigraphy for the Quaternary and later part of the Tertiary period has been constructed from continuous oxygen isotope records of foraminifera . As the lighter isotope of oxygen (160) is preferentially removed from the ocean when ice sheets expand during glaciations, general trends towards heavier 180 content reflect cooling conditions, while fluctuations indicate ice volume variation corresponding with, and controlling, sea level changes (see Figs. 5-7). Each internal phase is given a stage number (Shackleton & Opdyke, 1973) with odd-numbered stages indicating low ice volume and high sea level, ·equating generally with interglacial periods or interstadial events in more recent times, and even-numbered stages indicating high ice volume and low sea level, equating with glacials or stadials in the upper part of the record. Dating by radiocarbon and palaeomagnetism has demonstrated such a close correspondence between the isotope curves and predicted Milankovitch forcing (Berger & Loutre, 1991; Laskar, 1990) that the records can be calibrated against the astronomical timescale (Lourens et al., 1996; Shackleton et al., 1995). Thus, isotope curves can be used as an independent means of establishing ages.
The isotope record (Figs.5- 7) shows progressively increasing & 180 values, or decreasing temperatures and increasing variability over the last 3 Ma. A marked lowering of temperatures and a change from high frequency- low amplitude to lower frequency-high amplitude oscillations takes place around the Early to Middle Pleistocene boundary and marks a change in dominance of obliquity (tilting of the earth's axis relative to the sun) cycles, of about 41 ka duration, to eccentricity (the shape of the earth's orbit) cycles of about 100 ka duration. However, there is no sustained change around the formal Plio/ Pleistocene boundary, apparently negating the expectation that this boundary should mark the beginning of late Neogene climatic deterioration. The selection of this boundary was based on the first appearance of the cold water marine mollusc Arctica islandica and the benthic foraminifera Hyalinea balthica in uplifted, and therefore accessible, marine sediments in southern Italy.
473 However, the boundary is not supported by many other marker species and an alternative boundary is being actively considered by the Subcommission on Quaternary Stratigraphy of the international Commission on Stratigraphy (Partridge, l 997b).
The isotope record indicates a significant temperature lowering around 2.6 Ma, within a transition from a dominance of orbital precession (the season of the year when the earth is nearest to the sun) cycles which have a periodicity of about 23 ka, to obliquity cycles, and coincides with the major Gauss/Matuyama magnetic reversal. There is also evidence for the first major incursion of ice-rafted debris into the North Atlantic (Shackleton et al., 1984), the beginning of major loess deposition in China (Kukla et al., 1990), major changes in mammalian and vegetation communities in Europe (Sue, 1997), and the appearance of the earliest species, Homo rodolfensis , that is currently included in our own genus (Partridge et al., 1995) around this time. These factors;combined with the fact that there is a suitable stratotype marine section exposed in southern Italy showing the appearance of the first true cold indicator planktonic Foraminifera Neogloboquadrina atlantica, together with a variety of other biostratigraphic markers (Partridge, l 997b) makes it likely that a boundary change will happen in the near future. Consequently, it is apt to acknowledge both a major time of environmental change and anticipate a boundary change. As a result, the Quaternary will be regarded as having begun at 2.6 Ma but, to ·prevent total confusion, a distinction will be made in this chapter between the conventional Early Pleistocene (Early Pleistocene B, 1.76 to 0.78 Ma) and its temporal extension (Early Pleistocene A, 2.6 to 1.76 Ma) where appropriate .
NATUREAND APPLICATIONSOFFOSSIL EVIDENCE
The perception that the biotas of Australasia have been present for a long period of time, combined with their isolation from the rest of the world and the lack of massive regional disruptions such as those caused by the advances and retreats of ice sheets on northern hemisphere continents, has led to an emphasis on palaeoenvironmental reconstructions of individual sites or small areas rather than on broad palaeobiogeographic or evolutionary studies. However, the data gathered, although possibly not optimal for palaeo- geographic purposes, are providing a useful basis for examination of patterns of distribution in relation to reconstructions of environmental conditions.
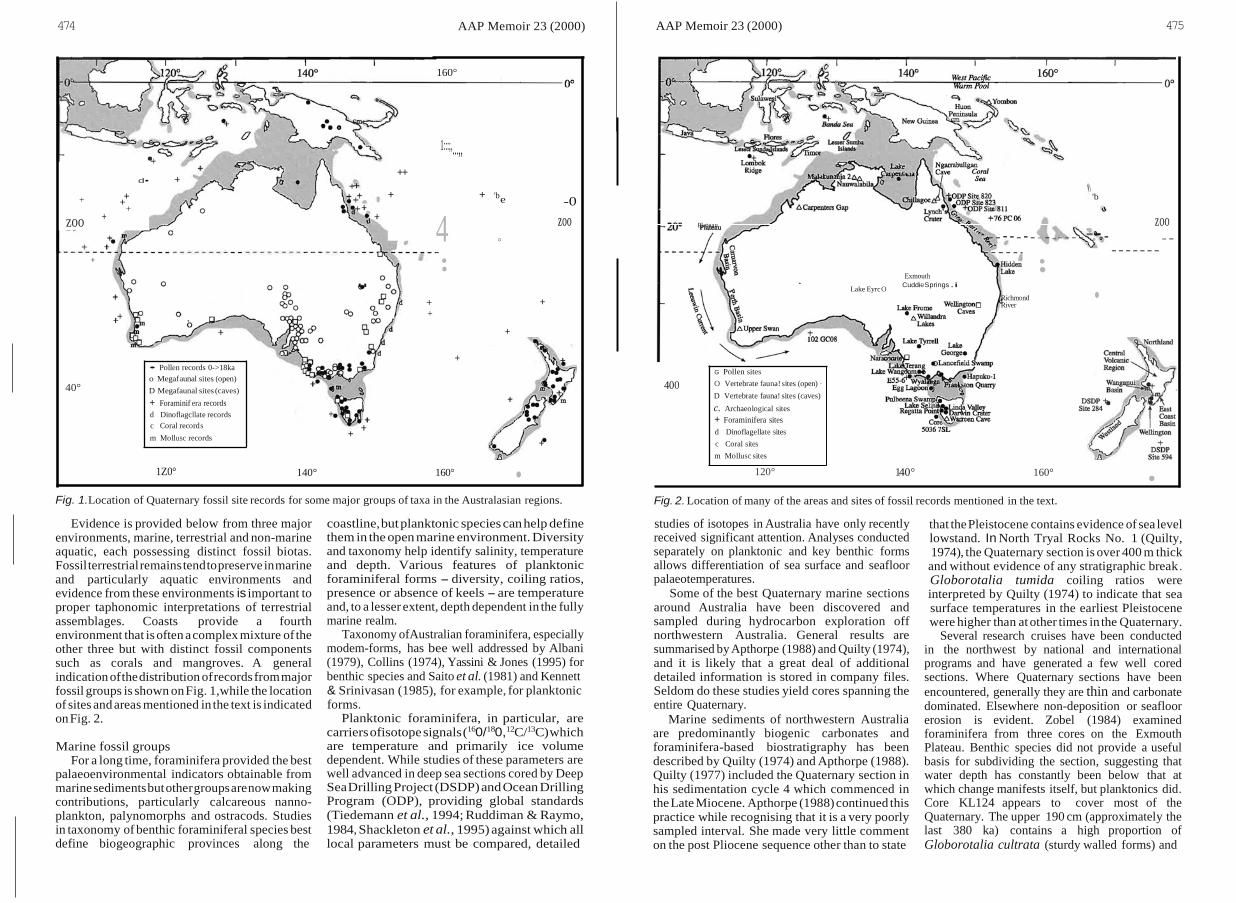
4
474 AAP Memoir 23 (2000) AAP Memoir 23 (2000) 475
160°
1:::;,...,, ++
d•
+ + + + 'b +
\\ 'b e -0
+ zoo +
zoo o
Plateau
- - - - - - - - - - - - - - -.- - zoo - -
+
.. •• + + +
+
Lake Eyrc O
Exmouth Cuddie Springs .i
••
Richmond River
- - - - - - -
•
40°
+ • Pollen records 0->18ka o Megaf aunal sites (open) D Megafaunal sites (caves) + Foraminif era records d Dinoflagcllate records c Coral record s m Mollusc records +
400
G Pollen sites O Vertebrate fauna! sites (open) · D Vertebrate fauna! sites (caves) c. Archaeological sites + Foraminifera sites d Dinoflagellate sites c Coral sites m Mollusc sites
1Z0° 140° 160° • 120° 140° 160° • Fig. 1.Location of Quaternary fossil site records for some major groups of taxa in the Australasian regions. Fig. 2. Location of many of the areas and sites of fossil records mentioned in the text.
Evidence is provided below from three major environments, marine, terrestrial and non-marine aquatic, each possessing distinct fossil biotas. Fossil terrestrial remains tend to preserve in marine and particularly aquatic environments and evidence from these environments is important to proper taphonomic interpretations of terrestrial assemblages. Coasts provide a fourth environment that is often a complex mixture of the other three but with distinct fossil components such as corals and mangroves. A general indication of the distribution of records from major fossil groups is shown on Fig. 1,while the location of sites and areas mentioned in the text is indicated on Fig. 2.
Marine fossil groups
For a long time, foraminifera provided the best palaeoenvironmental indicators obtainable from marine sediments but other groups are now making contributions, particularly calcareous nanno- plankton, palynomorphs and ostracods. Studies in taxonomy of benthic foraminiferal species best define biogeographic provinces along the
coastline, but planktonic species can help define them in the open marine environment. Diversity and taxonomy help identify salinity, temperature and depth. Various features of planktonic foraminiferal forms - diversity, coiling ratios, presence or absence of keels - are temperature and, to a lesser extent, depth dependent in the fully marine realm.
Taxonomy ofAustralian foraminifera, especially modem-forms, has bee well addressed by Albani (1979), Collins (1974), Yassini & Jones (1995) for benthic species and Saito et al. (1981) and Kennett & Srinivasan (1985), for example, for planktonic forms.
Planktonic foraminifera, in particular, are carriers ofisotope signals (160/180,12C/13C) which are temperature and primarily ice volume dependent. While studies of these parameters are well advanced in deep sea sections cored by Deep Sea Drilling Project (DSDP) and Ocean Drilling Program (ODP), providing global standards (Tiedemann et al., 1994; Ruddiman & Raymo, 1984, Shackleton et al., 1995) against which all local parameters must be compared, detailed
studies of isotopes in Australia have only recently received significant attention. Analyses conducted separately on planktonic and key benthic forms allows differentiation of sea surface and seafloor palaeotemperatures.
Some of the best Quaternary marine sections around Australia have been discovered and sampled during hydrocarbon exploration off northwestern Australia. General results are summarised by Apthorpe (1988) and Quilty (1974), and it is likely that a great deal of additional detailed information is stored in company files. Seldom do these studies yield cores spanning the entire Quaternary.
Marine sediments of northwestern Australia are predominantly biogenic carbonates and foraminifera-based biostratigraphy has been described by Quilty (1974) and Apthorpe (1988). Quilty (1977) included the Quaternary section in his sedimentation cycle 4 which commenced in the Late Miocene. Apthorpe (1988) continued this practice while recognising that it is a very poorly sampled interval. She made very little comment on the post Pliocene sequence other than to state
that the Pleistocene contains evidence of sea level lowstand. In North Tryal Rocks No. 1 (Quilty, 1974), the Quaternary section is over 400 m thick and without evidence of any stratigraphic break . Globorotalia tumida coiling ratios were interpreted by Quilty (1974) to indicate that sea surface temperatures in the earliest Pleistocene were higher than at other times in the Quaternary.
Several research cruises have been conducted in the northwest by national and international programs and have generated a few well cored sections. Where Quaternary sections have been encountered, generally they are thin and carbonate dominated. Elsewhere non-deposition or seafloor erosion is evident. Zobel (1984) examined foraminifera from three cores on the Exmouth Plateau. Benthic species did not provide a useful basis for subdividing the section, suggesting that water depth has constantly been below that at which change manifests itself, but planktonics did. Core KL124 appears to cover most of the Quaternary. The upper 190 cm (approximately the last 380 ka) contains a high proportion of Globorotalia cultrata (sturdy walled forms) and

476
other species indicating a subtropical-tropical fauna. From 190-420 cm (380-840 ka), the fauna is dominated by Globorotalia inflata, indicating somewhat cooler conditions in a 'transitional' fauna. Below that depth (to approximately 1.3ka), the subtropical-tropical fauna is present and this may coincide with Quilty's (1974) warmer interval.
Although few studies have been undertaken along the southern margin of Australia, relative to the size of the area, enough data are available to indicate that one of the major variables during the Quaternary (and earlier) has been the intensity of activity of the Leeuwin Current (Pearce & Walker, 1991). The effects are felt at least from southern Western Australia to western Victoria. The Leeuwin Current flows mainly in the upper few hundred metres, and not necessarily through
the full water column. Almond et al. (1993) and Li et al. (1997)
summarised the variation of foraminiferal faunas with standard Quaternary stratigraphic schemes. Almond et al. (1993) recognised five intervals (I to V) covering the last 125 ka in core 102GC08 taken in the Great Australian Bight. These do not coincide perfectly with the isotope stages of Shackleton & Opdyke (1973) but Interval III appears to be the same as Isotope Stage 5, Interval IV is Isotope Stage 6, and Interval V is the upper part at least oflsotope Stage 7. Interval II includes Isotope Stages 2 (lower part only) and 3 and 4. Interval I includes Isotope Stage 1 and the upper part of 2. Intervals I, III and V are the warmer water intervals characterised by high diversity (five planktonic species), dominated by Globigerinoides rober and Globorotalia inflata. During the alternating intervals, diversity is lower (four species) and dominance by any species is reduced .
There also are changes in the benthic species with lower infauna/higher epifauna during Intervals I, III and V. Benthic foraminiferal faunas respond to variations in global ice volume. High global ice volume (low sea level) leads to increased upwelling and nutrient supply and thus increased productivity (Arrhenius, 1952). Infauna! forms such as Uvigerina can be taken to imply times of higher productivity, and epifaunal faunas dominated by Cibicides, lower productivity. Almond et al. (1993) provide arguments for this interpretation.
This pattern, while having some scope for reinterpretation when longer cores are taken, suggests that during glacial intervals, the warm- water Leeuwin Current from the west was weaker, but was stronger during interglacials, such as at present. In consequence, warmer water faunas flourish across southern Australia in shallow water during interglacials.
AAP Memoir 23 (2000)
Wells & Okada (1996) used a more comprehensive array of indices on Verna core Vl 8-222, offshore from the Victoria-South Australia border. This core contains a record of Stage I-Stage 2 (last 14 ka), Stage 10 (350 ka) and Stage 12 (450 ka). Stage 1had a temperature maximum at about 8 ka BP with an annual range of 12-22°C, following a cooler Stage 2 (only the last 2 ka preserved). This coincides with high Leeuwin Current activity, low surface productivity and low upwelling. In contrast, Stages 10 and 12 were times oflower temperature (6-12°C) with higher productivity/higher upwelling and essentially no Leeuwin Current activity. The control on temperature may be oscillation of the Subtropical Convergence over the site. The results are broadly consistent with those of Almond et al. (1993).
There is an increasing emphasis on study of sections of the east coast of Australia. Samples normally are spot samples which lack a full stratigraphic context and, as pointed out by Chaproniere (1991), the foraminiferal biostratigraphy within the Quaternary depends on detailed knowledge of local events.
Nees (1997) studied a variety of features, such as infaunal/epifaunal ratios and abundance in benthic foraminiferal faunas from several cores in the Tasman Sea for the interval between Isotope Stage 6 and the present. Productivity increased during glacial stages, except on the Tasman Plateau, and surface productivity varied in relation to the position of the Tasman Front, which was considerably north of its present position during glacial Stage 6 and south of it in Substage 5e with higher productivity at the time over the Tasman Plateau.
In cores from the northern Tasman Sea, Martinez (1997) examined the population size of Globorotalia troncatulinoides as an index of productivity of Subantarctic Mode Water (SAMW) at the Antarctic Polar Frontal Zone over the last 150ka. Results showed that there has been a decrease j,M. abundance of SAMW generated over the recorded interval.
Chaproniere (1991) and Chaproniere & Pigram (1993) studied Quaternary Foraminifera from offshore Queensland and noted that there appears to have been little change through the Quaternary. Chaproniere (1991) recognised a sequence of events in the planktonic foraminiferal fauna but noted that there is essentially no evidence of cooling. He also included an oxygen isotope curve from Core 76PC06, which covers the interval of oxygen isotope stages 1-12, approximately the last 450 ka. He concluded that 'warm oligotrophic conditions similar to those today have been stable in the region since at least the Middle Pleistocene' (Chaproniere, 1991)
AAP Memoir 23 (2000)
Ocean Drilling Program Leg 133 conducted a drilling program off central and northeast Queensland and analyses of Foraminifera, magnetic properties, sediment composition, and many other parameters are documented in papers in McKenzie et al. (1993). In contrast with the view of Chaproniere (1991), these studies suggest major changes in the marine environment prior to and during the Quaternary in the Coral Sea region. Oxygen isotopic comparison of the planktonic Foraminifera Globigerinoides rober with that of the benthic Cibicidoides spp. in ODP Site 111 on the Queensland Plateau has revealed a sea surface temperature record through most of the last 10Ma (lsern et al., 1996).The age model was constructed from the biostratigraphic datums of Davies et al. (1991), Wei & Gartner, (1993) and Kroon et al. (1993) as well as comparison with the global isotope record. Major features include a substantial temperature increase around 2.5 Ma after a Late Pliocene low, and a further systematic increase, of around 4°C, superimposed on glacial/interglacial cyclicity, around 500-700 ka. This more recent increase is recorded in many other cores from the region and, from the most detailed isotope record, that of ODP Site 820 on the continental slope adjacerit to the Great Barrier Reef, the timing seems to be closer to 400-250 ka (Peerdemann et al., 1993). The youngest part of some sections shows the influence of the southward progression of the East Australia Current.
In contrast with accepted orthodoxy, higher concentrations of foraminifera and carbonate in sediments seem to coincide with glacial intervals, and there is considerable discussion over the cause of this apparent anomaly. Conversely, siliciclastic content is higher during warm intervals and this is expressed as higher magnetic susceptibility during intervals of high sea level. Substantial cooling is manifested during glacial intervals.
The Continental Shelf around Tasmania appears to be receiving little sediment at present but the Quaternary sediments known are dominantly carbonates and this gives hope of useful comparative sections which may provide standard sections for much of the Australian margin. Carbonate cores studied by Connell & Sikes (1997) cover the last 150 ka and these have patterns similar to those recorded from the Southern Ocean and Atlantic Ocean sub-Antarctic (Hodell, 1993; Howard & Prell, 1994). Carbonate content is at a minimum during glacial maxima, and there are grain size trends in the terrigenous content that relate to distance from sediment source.
In New Zealand, the application of foraminiferal studies to Quaternary palaeoenvironmental and stratigraphic reconstructions has had a much
477
longer history than in Australia due to the presence of thick, extensive marine sequences onshore accessible as exposures and to terrestrial drilling. This facilitated the development of a stratigraphic scheme for New Zealand which may have, to some extent, separated it scientifically from the rest of the world, but was of immense value in local terrestrial/marine correlation and in providing a timescale for understanding biogeographic change in the region. Marine sediments are particularly extensive in the Wanganui and East Coast Basins, in the southern part of the North Island which have been the focus of research . The deep water facies of the East Coast Basin have provided the best records of foraminifera, particularly for the Late Pliocene to Early Pleistocene, and these are described in Hornibrook (1981). These records have been correlated biostratigraphically with the classic marine sequence for the Late Cainozoic from DSDP Site 284 to the west of the South Island (Hornibrook, 1982). The oxygen isotope record from this m·arine core was important in demonstrating Late Miocene expansion of the Antarctic ice sheet and, more significantly for this review, the initiation of northern hemisphere glaciation around 2.5 Ma.
In addition to these regional syntheses of foraminiferal stratigraphy, a number of oxygen isotope records have been produced around Australasia covering the last or last few glacial cycles. Some of these have provided excellent chronologies for palynological records produced from the same cores. These records are considered under terrestrial fossil groups.
Closer to coasts, marine and estuarine molluscs become important relative to foraminifera in the establishment of biostratigraphies and palaeoenvironmental conditions. As they are more closely linked with adjacent coastlines, molluscs tend also to have more limited geographic distributions (with certain notable exceptions, particularly Anadara trapezia [Murray-Wallace et al., 2000]) although, due to their mobility frequently at the larval stage, many are common throughout the south Pacific and Indian Ocean regions. Their distributions mainly relate to water temperature, depth and salinity so they provide evidence of changes in the distribution of currents, water flow characteristics of rivers discharging into the sea, and sea levels. Their value is heightened by variation in the sediment types within which they are recorded and by the success achieved in dating middle to late Quaternary shells, particularly by amino acid racemisation (e.g. Murray-Wallace, 1995; Murray-Wallace et al., 1989).
Knowledge of the taxonomy and Quaternary palaeoecology of molluscs is substantial for New

478
Zealand (e.g. Beu & Maxwell, 1990) and for various parts of Australia, particularly the western seaboard of Western Australia and coastal South Australia (e.g. Kendrick et al., 1991, Ludbrook, 1984). Components of these regions, and especially New Zealand, have experienced uplift allowing the preservation of a variety of coastal landforms, especially those related to past sea level highs.
InNew Zealand, molluscs have played a key role, along with foraminifera and nannofossils (minute phytoplankton with calcareous tests) in the development of the Quaternary biostratigraphy. The Plio/Pleistocene boundary has been traditionally located at the time of extinction of typical late Tertiary, warm water molluscs such as Cucullaea, Olive/la and Polinices and their replacement by cold water arrivals, for example Zygochamys delicatula and Eucominia nassoides (Fleming, 1944) while subdivisions have relied heavily on the stratigraphic distribution of pectinid bivalves (Sectipecten, Phialopecten, Kaparachlamys, Zygochlamys and Pecten sensu stricto ) (Beu et al., 1987; Beu, 1995) and struthiolariid gastropods (Neef, 1970). The more parochial nature of molluscs relative to foraminifera and other marine microfossil groups has lessened their importance for broad regional correlation but they still remain critical to dating and facies identification in shallow marine sediments. Due to their diversity (c. 1000 species in Quaternary records), molluscs also provide much more data on environmental conditions.
The preservation of shell beds within the Camarvon and Perth Basins along the western coast of WesternAustralia provides important and, apart from vertebrate assemblages, generally the only glimpses from fossil evidence of Quaternary conditions within the western part of the continent (Kendrick et al., 1991). Mollusc assemblages from the Ascot Formation are considered to mark the Plio/Pleistocene boundary on the basis of correlation with the New Zealand sequence and the mix of extinct and extant species. Middle Pleistocene assemblages, attributed to stage 7 from field stratigraphic relationships with units of known age and direct dating by amino acid racemisation (Murray-Wallace & Kimber, 1989), contain a greater number of subtropical and tropical species than those of the earlier period, suggesting warmer sea surface temperatures. Itis considered by Kendrick et al. (1991) that these higher temperatures resulted from the development or intensification of the Leeuwin Current in the Middle Pleistocene. Assemblages from the Last Interglacial Maximum, dated by Uranium/Thorium (U/Th), suggest that river discharge, and hence climate, was less variable than
AAP Memoir 23 (2000)
today, and very similar to conditions in the region during the middle Holocene (Kendrick, 1977). A recent review of the historical biogeography of the estuarine mollusc Anadara trapezia suggests warmer sea surface temperatures during later interglacial maxima were widespread in the southern Australian and New Zealand regions, dating from isotope stage 11 in New Zealand and at least isotope stage 7 in southern Australia (Murray-Wallace et al., 2000).
At lower latitudes, corals are an important fossil group. The taxonomy and present day biogeography are relatively well known, especially for the Indo-Pacific region and they have a low rate of speciation, relative to most other faunal groups (Veron, 1995). A comparison of fossil assemblages from around the Plio/Pleistocene (B) boundary in southern Papua New Guinea with those living today in the region indicated similarity in growth environment and that the majority of species (65) showed no significant differences from extant species (Veron & Kelley, 1988). Only nine species showed taxonomically significant change while only two taxa are now unknown within the region. Scleractinian corals have well defined ecological ranges, good dispersal ability and can be identified to species level, although there is a great deal of morphological variability within species. These features make them useful contributors to the reconstruction of palaeo- environments during the Quaternary for near- shore environments. Their value is enhanced by the fact that many species contribute to the formation of reefs whose relative permanence in the landscape allows the reconstruction of environments over long periods of time.
Studies on coral assemblages through the last 95 ka in New Guinea have indicated little change in composition, suggesting either that sea surface temperatures have been relatively constant or that corals in the equatorial regions are relatively insensitive to temperature change (Pandolfi, 1996).;However, there have been major changes in the distribution and activity of reef growth off eastern Australia, including the Great Barrier Reef (McKenzie & Davies, 1993 ). The attenuation of coral species with increased latitude has indicated significant temperature changes in the southern parts of Australia. Off the coast of Western Australia, reef deposits, associated with the Last Interglacial and older periods have been recorded well south of their present position (Kendrick et al., 1991). Along the New South Wales coast, Last Interglacial reef deposits are found up to 700 km south of current reefs, suggesting that sea surface temperatures were around 2°C higher than today (Murray-Wallace & Belperio, 1991). Although many factors influence coral distributions and
AAP Memoir 23 (2000)
caution must be exercised in relating fossil occurrences simply to temperature levels (Veron, 1995), it is up.likely that temperature has not been a major factor in the creation of such large distributional changes.
The most extensively studied fossil corals are those forming reefs on the rising coastlines of New Guinea and neighbouring Indonesia. The record from the Huon Peninsula, dated by U/Th (Chappell, 1983, 1994) and by matching with the marine isotope stratigraphy (Chappell & Shackleton, 1986) provides one of the world's most detailed sea level records through about the last 300 ka. Oxygen isotope analysis of associated giant clams, has provided an independent sea surface temperature curve by extraction of the sea level component of the record from the isotope record (Aharon, 1983; Aharon & Chappell, 1986).
Dinoflagellates are common and often dominant elements in the phytoplankton of estuarine, nearshore and shelf environments, but mostly become less abundant with increasing distance from shore. Although dinoflagellates are common in both marine and freshwater environments, it is only in marine and estuarine environments that cysts are produced. It is these cysts that are used
in palaeontological studies. Modern dinoflagellate cysts show both a
latitudinal (McMinn, 1990) and a salinity gradient (McMinn, 199la, b) that can be used in Quaternary climate reconstruction, although they can also be used to investigate changes in circulation and frontal movements (Sun & McMinn, 1994; McMinn & Sun, 1994). Taxa characteristic of warm seas include Polysphaeridium zoharyi, Operculodinium israelianum and Tuberculo- dinium vancampoe. Taxa characteristic of cooler seas include Bitectatodinium tepikiense, Dalella chathamense and Pyxidiniopsis reticulata. Most Quaternary studies are based on Late Pleistocene and Holocene changes in southeastern Australia. These include Bulahdelah (McMinn, 1987), Botany Bay (McMinn, 1989), Richmond River (McMinn, 1992a), offshore southern N.S.W. (McMinn, 1992b), Lake Macquarie (McMinn, 199la) and offshore western Tasmania (McMinn & Wells, 1997). With the exception of the record from the Richmond River (McMinn, 1992a) which extends to Isotope Stage 7, none of the others examined material beyond Isotope Stage 5e.
In an examination of Pliocene to Holocene sequences from foundation test holes associated with the construction of production platforms in Bass Strait, McMinn (1992c) identified three biostratigraphic zones. The Achomosphaera ramulifera zone, based on the interval between the extinctions of Melitasphaeridium choanophorum and A. ramulifera, is Late Pliocene
479
to Early Pleistocene in age, while the Protoperidinium leonis zone, based on the interval between the extinction of A. ramulifera and the surface is Pleistocene to Holocene in age. Two subzones, based on the local extinction of Operculodinium israelianum, are recognised within the A. ramulifera zone. 0. israelianum is a common warm temperate and tropical species and so it is not possible to recognise this zone much further north.
Neogene and Quaternary sequences have been examined from offshore northwestern (McMinn, 1992d) and northeastern (McMinn, 1994a, b) Australia. The Quaternary samples from northwestern Australia contained evidence for reworking and so no attempt was made to determine first and last appearances, but the assemblages were typical of tropical environments. The dinoflagellate cysts in the study from the Great Barrier Reef, northeastern Australia, unfortunately showed little systematic variation coinciding with glacial/interglacial events. It is probable that the water temperature varied by too little in this location for changes in the dinoflagellate assemblages to be significant, a conclusion at variance with the interpretation of oxygen isotope records from the area.
Terrestrial fossil groups
By far the most important indicator of terrestrial palaeoenvironments is pollen which is preserved in almost all continuously waterlogged sediments including those of marine environments, in many lakes which experience drying particularly if there is a salt crust, and in:many cave deposits. All these environments have revealed useful measures of surrounding vegetation due to atmospheric and fluvial transport and mixing of pollen, with the size of the vegetation catchment recorded generally related to the size of the depositional basin.
Most onshore records are from more humid environments, particularly New Zealand, southeastern Australia and New Guinea Highlands where cool conditions combined with high precipitation are most conducive to the accumulation of peat deposits and contain sites formed by past glacial, periglacial and tectonic activity. The clusters of sites in northeast Queensland and in the drier western parts of Victoria reflect the availability of sites produced by volcanic activity. Other records largely within these areas are derived from depressions associated with high water tables, especially along river valleys and coasts. The majority of sequences provide evidence for the period since the Last Glacial Maximum and have allowed the construction of regional patterns of vegetation and

480
environmental, particularly climatic, change for this period. Regional summaries are provided in Dodson (1992), with a more detailed analysis of mainland southeastern Australia undertaken by Kershaw (1998).
Substantially longer continuous records from aquatic sites, covering at least the last glacial cycle, are confined to deep volcanic crater lakes such as Lake Terang (D'Costa & Kershaw, 1995) and Lake Wangoom (Edney et al., 1990, Harle et al., 1999) on the Western Plains of Victoria, Lake Leake in southeastern South Australia (Dodson, 1975) and Lynch's Crater and Strenekoff's Crater on the Atherton Tableland (Kershaw, 1994), the tectonic basins of Lake George near Canberra (Singh & Geissler, 1985; Kershaw et al., 1991) and Wellington, New Zealand (Mildenhall, 1994, 1995), the spring deposits of Pulbeena Swamp in northern Tasmania (Colhoun et al., 1982), the glaciated basin of Lake Selina in western Tasmania which survived the latest glaciation (Colhoun et al., 1999) and the Darwin Crater meteorite impact depression (Colhoun, 1988). More discontinuous long records have been constructed from the sites of Egg Lagoon on King Island (D'Costa et al., 1993) and Hidden Lake on Fraser Island (Longmore & Heijnis, 1999) inland of coastal dunes.
In the absence of direct, reliable, radiometric dating methods, timescales for these longer records have been estimated from pattern matching with the marine isotope record. However, such matching, even if generally reliable, inhibits investigation of lead and lag effects which are important for understanding the causes of climate change and vegetation responses. Analysis of pollen from marine sequences permits comparison of pollen and isotope records. Records produced from the Lombok Ridge (van der Kaars, 1991; Wang et al., 1999) and Banda Sea (van der Kaars et al., 2000) to the north of Australia, provide a regional picture of vegetation and flora in the northern Australia-New Guinea-Indonesian region over the last few glacial cycles; DSDP Site 594 off South Island, New Zealand (Reusser & van de Geer, 1994) and ODP Site 820 on the northeast Queensland continental slope (Moss & Kershaw, 2000) covering a similar period; and deep sea cores S036-7SL off western Tasmania (van de Geer et al.,1994 ) and E55-6 off the coast of western Victoria (Harle, 1997) covering much or all of the last glacial cycle. Despite the very different environments of deposition, all marine records show patterns of vegetation change similar to more localised records from terrestrial environments and, in the case of the three latter records, the aims of correlation with, and provision of a firmer timescale for, adjacent terrestrial records, were achieved.
AAP Memoir 23 (2000)
Only two records cover a large part of the Quaternary; that from Lake George and a coarse record through the last 1.4Ma from ODP Site 820 (Kershaw et al., 1993). However glimpses of Quaternary vegetation and flora, although poorly dated in the main, are provided from isolated polliniferous sequences in drillholes and exposures, particularly in the more dynamic landscapes of New Zealand (e.g. Mildenhall, 1978, 1983; Mildenhall & Suggate, 1981) and Tasmania (e.g. Augustinus & Macphail, 1997; Jordan et al., 1995) while Bussell (1988, 1990) and Bussell & Pillans (1992) provide a better dated and systematic representation of vegetation from the last few interglacials in the Wanganui region of North Island, New Zealand. Moar & Suggate (1996) piece together a picture of the last interglacial- glacial cycle from Westland on the South Island from about 20 isolated deposits. Summaries of pollen stratigraphy and vegetation are provided for the Late Pliocene (Macphail et al., 1995) and Early-Middle Pleistocene (Macphail et al., 1993) of Tasmania, and for at least parts of the Quaternary and the Tertiary/Quaternary transition in the general region by Mildenhall (1980), Kershaw (1988), Kershaw et al. (1994), Macphail (1997) and Nelson et al. (1988).
There are few continuous pollen records of any length published from semi-arid and arid environments within the region. The most substantial are from the salt lakes of Lake Frome (Singh, 1981; Singh & Luly, 1991) in South Australia and Lake Tyrrell (Luly, 1993) in northwest Victoria but both provide evidence only of the very late Quaternary. A more generalised record accompanies the important archaeological and vertebrate palaeontological site of Cuddie Springs in western New South Wales (Dodson et al., 1993). Dating reliability has been a problem but the recent development of the Accelerator Mass Spectrometry (AMS) radiocarbon dating technique, which can cope with the very small amounts .ilof carbon preserved in such environments, is encouraging greater activity in this area.
Also showing potential from these more arid areas is the study of pollen preserved in Leporillus (stick-nest rat) middens (Green et al., 1983; Nelson et al., 1990; Pearson & Dodson, 1993). These middens, composed of plant and other material cemented to rock crevices by urine, are similar to those formed by midden-building mammals in other semi-arid parts of the world, particularly the pack rats of the southeastern USA (Betancourt et al., 1990) and hyrax of southern Africa (Scott & Bousman, 1990), where methodologies for study have been established. Histories are pieced together from a number of
AAP Memoir 23 (2000)
middens dated by conventional or AMS radiocarbon dating. Comparison of recent spectra has demonstrated that, despite the very different setting of middens and potential for stick-nest rats to bias assemblages by incorporation of plant material, results are very similar to those derived from nearby spectra from soil surfaces (Pearson, 1997). Additional taphonomic studies have been conducted by Head et al. (1998), but full understanding of the processes of deposition, as well as the future value of these studies, is inhibited by the fact that both species of stick-nest rat have become extinct on the Australian mainland since the arrival of Europeans. The temporal scale of investigation is also likely to be restricted as the oldest middens found so far date back only to about 10 ka (McCarthy et al., 1996) although, importantly, spatial coverage is good over much of the centre of the continent lacking other suitable sites for palynological study.
The standard methodology in Quaternary palynology includes counting all recognisable pollen and fem spores and their percentage portrayal in diagrammatic form in relation to a determined pollen sum. Interpretation is facilitated by reference to the known ecology and distribution of parent plants and particularly by comparison with modern pollen samples from known vegetation and environments. This approach contrasts with that of pre-Quaternary palynology where the emphasis is placed on the recognition of stratigraphically important taxa. Although the Quaternary approach is useful in providing a good overview of vegetation and climate, it does have its limitations, particularly in Australia. In the first place, much of the continent is characterised, both in vegetation and pollen representation, by a few taxa including Eucalyptus, Casuarinaceae, Poaceae, Asteraceae, Chenopodiaceae, and Callitris which dominate interpretation. Unfortunately, these taxa are difficult to identify to more refmed taxonomic levels and this has led to very generalised reconstructions which, in some instances such as Lake Tyrrell, resulted in major debate over community representation and hence environmental interpretation (Luly, 1995; Sluiter & Parsons, 1995). Although refmed identification is a problem in palynology generally, Quaternary palynologists, with notable exceptions (eg. Moar, 1993; Boyd, 1992), have neglected pollen morphology and hence limited interpretation potential. In the second place, and leading on from the first, it is becoming increasingly clear that community composition is constantly changing and the use of surface samples for comparison purposes becomes increasingly difficult as records extend back in time. It has been established for one sequence in southeastern Australia, that
481
extends from the Last Glacial Maximum to present, that no satisfactory modem analogue exists within the general region until the mid- Holocene (Kershaw & Bulman, 1996).
One technique which could become an important complement to palynology is phytolith analysis. Itis widely adopted in archaeological and palaeoenvironmental studies in many parts of the world (Piperno, 1987) and its potential for Australia has been reviewed by Hart & Humphreys (1997). Phytoliths are composed of silica (opal) formed within and between plant cells, particularly those of grasses and other monocotyledons. They also are found in a range of dicotyledons and have been found in sediments dating back to the Eocene. They are of similar size to pollen and occur in similar abundance so that, theoretically, they can also be subject to similar quantitative analysis. However, most phytoliths are not diagnostic, with a number of shapes derived from the same plant and many taxa having similar structures, so that systematic variation in plant communities is difficult to determine. Their major values are in allowing more refined identification of specific groups of plants, and overseas they have been very successful in the important separation of cereals like rice and maize from other grasses, and also in providing evidence from soils and archaeological sites where pollen is not preserved. In the most substantial study in Australia to date, Bowdery (1996) has provided good evidence of some components of late Quaternary vegetation from archaeological sites in arid Australia.
Refined taxonomic identification of plants can be most usefully determined from the greater range of features preserved in macrofossils. Traditionally, analysis of the remains of leaves, fruit and seeds has focused on local aquatic plants and contributed to the documentation and understanding of hydroseral succession (Watts, 1978). However, the limited number of macrofossil studies undertaken in Australasia, often in association with pollen analysis, has been of greater importance to understanding of evolutionary and distributional patterns in selected terrestrial taxa as well as in refining palaeoclimatic estimates. Advantages of macrofossil remains over pollen, in addition to that of identification, lie in their structural expression of environmental conditions, particularly the size and shape of leaves and their degree of sclerophylly, as well as their limited dispersal. The most substantial and systematic studies on Quaternary macrofossils, which have included examination of dispersed cuticle, have been undertaken in Tasmania (Hill & Macphail, 1985; Jordan, 1995a, b, 1997; Jordan et al., 1991, 1995) and have revealed marked changes in the distribution and ecology of a number of rainforest

482
and sclerophyll taxa over the Quaternary period, including extinctions at the palynologically invisible level. One major conclusion is that caution must be exercised in applying present vegetation/climate patterns to the reconstruction of past climates as both plant/climate relationships and climate itself have been very different from those of today. In studies confined to the late Quaternary, however, the presence of macrofossils in the form of wood of Nothofagus in mainland southeastern Australia (McKenzie & Busby, 1992: McKenzie & Kershaw, 1997) and Agathis in North Island, New Zealand (Ogden et al., 1992), has been instrumental in documenting precise changes in the distribution of these taxa which have allowed detailed quantitative estimates of past climatic conditions. Inthe New Zealand case, an indication of the degree of climate change could also be gauged from a comparison of tree rings within modem and fossil wood.
Macrofossil examination is increasingly forming a component of archaeological studies (Beck et al., 1989) and although emphasis is usually placed on collected food and related materials, valuable data can be obtained on changes in plant distribution and climate. McConnell (1997), for example, provides a credible picture of climate change through the last 40 ka in the Kimberley region, an area which has proved essentially barren for palynological study. In addition, macroremains from stick-nest rat middens in arid areas can provide a useful complement to pollen as has been the case for packrats in the USA, although material is less abundant than in the USA (Head et al., 1998).
Another form of plant material, which comes in micro- and macro-fossil size ranges, is charcoal. This very resistant product of biomass burning is essential for understanding the biogeography of much of this part of the world where the biota has evolved in association with fire. Since the publication of Singh et al. (1981), which demonstrated the important link between vegetation change and fire, microscopic charcoal has been routinely counted in pollen analytical studies in Australia. This has been adopted by workers in New Zealand and New Guinea (see Kershaw et al., 1997; Ogden et al., 1998). It is clear that fire has been a constant feature of the whole Quaternary, and has generally increased in activity throughout this period, but, despite the important background research of Clark (1982, 1983, 1984), it has not proved possible as yet to quantify vegetation/fire relationships. Concerns over the accuracy of charcoal counting as a real measure of biomass burning has led recently to the chemical analysis of the elemental carbon component (e.g. Wang et al., 1999). The
AAP Memoir 23 (2000)
methodology, developed by Bird & Grocke (1997), also allows the isotopic composition of carbon to be analysed, providing a measure of the relative importance of taxa utilising the C3 and C4 photosynthetic pathways and hence an indication of the nature of the source vegetation. In the study of the tropical marine core by Wang et al. (1999), this measure approximates the amount of burning in forest relative to that in grassland.
A further recent development has been the utilisation of the fine structural detail preserved in macrofossil charcoal to allow identification of source plants (Hope, 1998). In northeast Queensland, the analysis and dating of soil charcoal has allowed a greater spatial assessment of late Quaternary burning than that recorded in the Lynch's Crater pollen record to be determined (Hopkins et al., 1993, 1996). The identification of most charcoal as Eucalyptus demonstrates that eucalypt woodland regionally replaced rainforest during the Last Glacial Maximum and has major applications to the debate over glacial rainforest refugia and the present distribution and floristic stability of rainforest communities. The ongoing examination of charcoal in archaeological sites to the west of the major areas of rainforest in northeast Queensland is providing a further spatial extension of the palaeoenvironmental record to an area where long pollen sequences are lacking (Nie Dolby, pers. comm.). The potential of this method to provide data on vegetation from arid environments has been demonstrated by the analysis of charcoal from an archaeological site in central Australia (Smith et al., 1995). Here, the development of vegetation similar to the present day was deduced from identification of mulga (Acacia aneura) charcoal, an identification that would not have been possible from pollen.
In contrast to the flora, fauna! groups have played relatively little part in the construction of Quaternary terrestrial sequences and envirents: consequently data with which to construct palaeogeographies is limited. This is in marked contrast to some other parts of the world, particularly much of the northern hemisphere, where large mammals have traditionally played a major role in biostratigraphy (Kurten, 1968; Kurten & Anderson, 1980) and where smaller vertebrates and invertebrates have demonstrated substantial palaeoecological value.
However, the unique nature of the vertebrate fauna of New Zealand, with its lack of mammals but array of endemic flightless birds, and Australia, and to a lesser extent New Guinea, with their preponderance of marsupials, has attracted a great deal of attention. As with other terrestrial groups, the lack of suitable sedimentary or preservational
AAP Memoir 23 (2000)
environments has inhibited the development of useful biostratigraphies or evolutionary trends, with the added impediment that macrofossil studies demand access to a large amount of material. Consequently records are restricted largely to erosional exposures within river systems, swamps, lakes, lunette and sand dunes and loess, and also to caves where the ratio of fossils to matrix is high and where assistance is often provided, for more recent time periods, by archaeologists. Archaeological middens also contribute a source of recent material. Much background on Quaternary faunas is presented, at length, in Vickers-Rich et al. (1991).
Throughout the region, there is little information on vertebrate faunas that can be dated to the earlier part of the Quaternary and the difficulty of placing sites in even a relative chronology suggests that there was little change in the fauna through most of the Quaternary period. Rich (1991) states that 'As a generality, with unfortunately all too few exceptions, it may be said that Australian terrestrial mammal sites fall into one of two age groups. Either they are younger than 35 ka and capable of being dated by the 14C-technique or they are regarded as Quaternary, beyond the range of 14C'. This conclusion is largely echoed for Quaternary avifauna where Baird (1991) notes only a few elements restricted to this period, while the Early and Middle Pleistocene are almost absent from the New Zealand terrestrial vertebrate record and the few bones from the Plio/Pleistocene are all attributable to taxa recorded at present or within the Holocene (Millener, 1991). However, there are a few sites in Australia which have the potential to provide long stratigraphic records. These include the Wellington Caves complex of New South Wales which may extend back into the Late Tertiary (Osborne, 1997), and Naracoorte Caves in South Australia where problems of dating are now being addressed by Thermal Ionisation Mass Spectrometry (TIMS) of speleothems (Ayliffe et al., 1997).
Late Quaternary sites are plentiful, a result of their accessibility, greater likelihood of preservation, archaeological interest and the fascination with late Quaternary megafaunal extinctions. InNew Zealand where there is a close association between extinction and the arrival of people which occurred in the very late Holocene, over 800 sites are known and, within midden sites, at least 28 of the 33 avian species including all 11 moa species known to have become extinct during the Polynesian period are recorded (Millener, 1991). From these data, recent, more substantial vertebrate records extending back to the very late Pleistocene (eg. Worthy & Holdaway, 1993, 1994,
483
1996;Worthy, 1998) and associated palynological research, it has been possible to reconstruct the ecology and distribution of the extinct avifauna, particularly the moas, an important prerequisite for the understanding of the cause(s) of extinction. The original assumption that most moas occupied grassland habitats has proved untenable in light of the substantial late Quaternary vegetation history of the islands (McGlone, 1988) which clearly shows that most of New Zealand was covered in forest until the arrival of Polynesians. The distribution of kill sites and examination of crop contents showing a high proportion of twigs, indicates that the majority of species inhabited open forest and scrubland with several confmed to lowland or upland forest, and one species to subalpine scrub and grassland (Anderson, 1989). There appears to be no pattern in the order of extinction between species and the chronology of sites shows a strong peak around 1300-1250 AD followed by a rapid decline. No sites are reliably dated to later than the 16th century (Anderson & McGlone, 1992) and it is possible that moas disappeared within a hundred years of the arrival of people (Holdaway & Jacomb, 2000).
InAustralia, there was mass extinction of an estimated 86% of existing 'megafauna' (Martin, 1984) including all 19 species greater than 100kg in mass (Flannery, 1990). Megafaunal extinctions were predominantly marsupials but included three reptiles and one flightless bird (Genyornis). The 'event' was accompanied by a lower but still significant proportion of extinction in a whole range of smaller terrestrial vertebrates and by dwarfmg in some extant species (Flannery, 1990; Murray, 1984). As the major extinction phase occurred much earlier in Australia than in New Zealand, within the Late Pleistocene, reconstruction of distribution patterns and habitats of extinct species is more difficult. However, from the examination of all sites containing extinct megafauna (Fig.2) combined with feeding habits determined from their dentition and assumed vegetation patterns, a substantial attempt has been made by Horton (1984), at least for the marsupials. He determined that there were three major groups of megafaunal species, one adapted to southern grasslands, a second to warmer and drier woodlands, and a third to cooler and wetter woodlands. On the basis of distribution patterns of surviving fauna, he allocated the megafauna to rainfall zones between 250 and 750 mm mean annual rainfall. He considered that their concentration here, and absence from both wetter forested and drier arid environments is consistent with biomass estimates and fauna!composition globally. An interesting feature of this reconstruction is that the only megafaunal species

484
of arid areas, the red kangaroo (Macropus rufa), did not become extinct. Horton credits this survival to the lack of stress on an arid-adapted species during the extinction period that he relates to a period of continental drying. He considers that the majority of taxa became extinct at or shortly after about 26 ka, the beginning of oxygen isotope stage 2, with the remainder succumbing to driest conditions around or after the Last Glacial Maximum between about 18 and 15 ka.
Horton's climatic model is but one of a number of hypotheses that have been forwarded in an attempt to explain megafaunal extinction and, partially due to the overwhelming evidence for extinction by humans in New Zealand, has not been widely accepted. However, it has been difficult to argue the case for a human explanation, due either to overkill (Flannery, 1990; 1994) or through habitat change (eg. Jones, 1973), when radiocarbon ages have persistently suggested that megafauna and people co-existed for at least 20 ka (see next section for timing of the arrival of people in Australia). Only recently, with the application of dating techniques additional to radiocarbon, has any confidence been generated in the timing of megafaunal extinction. The most convincing evidence for the timing of extinction comes from amino acid racemisation dating, supported by accelerator mass spectrometry (AMS) radiocarbon and TIMS CT-series analysis, of some 1200 egg shells of the giant flightless bird, Genyornis, from three arid to semi-arid regions in southern Australia (Miller et al. 1999). Genyornis was continuously present from more than 100 ka until 50 ka when it suddenly disappeared. Ina more humid part of southeastern Australia, electron spin resonance dating of extinct diprotodon teeth at Lancefield Swamp, a site critical to Horton's reconstruction, has put back the date of extinct megafauna from 26 ka to at least 40 ka (van Huet et al., 1998). Although these studies, on their own, do not provide an age for the extinction of all megafauna or allow determination of the exact mechanism of extinction, they suggest strongly that radiocarbon dating underestimated extinction dates and leave open the possibility that extinction coincided with the arrival of people.
.The bulk of palaeoenvironmental information is derived from the fauna that survived the extinction phase. A number of studies have provided data which complement or question reconstructions derived from pollen and geomorphic data and, in southwestern Australia where Climatic reconstructions are few and somewhat contradictory, faunal sequences have been important in establishing past environmental conditions (Merrilees, 1984). The value of bird remains has been demonstrated by Baird (1989)
AAP Memoir 23 (2000)
and Worthy & Holdaway (1993) in detecting the survival of palynologically invisible or uncertain location of small pockets of rainforest in southeastern Australia and Westland, New Zealand, respectively, during the last glacial period. As with plant fossils, it is frequently revealed that past assemblages, even during the late Quaternary, were very different from those of today, limiting their application to the reconstruction of past climates (Ride & Davis, 1997).
Faunal groups which have only recently begun to contribute to an understanding of Quaternary terrestrial environments are land snails and beetles. Land snails are commonly used in the northern hemisphere for providing details of past vegetation communities, and moisture and temperature estimates (eg. Goodfriend, 1992; Preece, 1990) but have been limited in their application in Australia because the detailed ecological and systematic background research necessary for full interpretation of fossil records has yet to be undertaken. However, several studies have been successfully completed. Two studies on land snails, from a limestone rock shelter at Chillagoe in north Queensland (David & Stanisic, 1990) and from a limestone cave in southwestern Tasmania (Porch & Allen, in prep.), provide local environmental records for archaeological sites in areas lacking suitable pollen sequences. A third record from Pulbeena Swamp in northern Tasmania (Porch, submitted) provides additional and supportive evidence of environmental changes deduced from the analysis of pollen (Colhoun et al., 1982) and ostracods (De Deckker, 1982d) at the site. Ithas also been established recently that, in northeast Queensland at least, there is systematic intraspecific variation in mollusc size in relation to rainfall which is providing a basis for reconstruction of past rainfall patterns from fossil sequences (Rowe et al., in press).
The interest in beetles, especially terrestrial beetles, lies in their sensitivity and rapid response to climateiJ>articularly temperature change (Elias, 1994). It has been demonstrated in the northern hemisphere that beetles provide a much more precise indicator of major glacial/interglacial temperature change than vegetation and organisms tethered to vegetation communities (Coope, 1986). Beetles are also considered to be reliable indicators of climate change over a long period of time in that they show remarkably slow rates of evolution, with few extinctions or new species recorded during the Quaternary period. The recent success achieved in an extension of study to southern South America, and the value of combining beetle and pollen analyses, (e.g. Ashworth et al., 1991) has encouraged the recent study of sites in southeastern Australia where
AAP Memoir 23 (2000)
pollen records have been constructed but where pollen has proved generally to be a less useful indicator of temperature than it has of rainfall (Porch & Elias, 2000).
Aquatic biota
Fossil studies have importance in Australian aquatic environments largely because of a general shortage of water and the sensitivity of water bodies to natural environmental variability and human impact. However, the frequent drying of lakes on all timescales has inhibited the development of research into many potential indicator fossils including rhizopods, rotifers, sponges, molluscs, insects and fishes, algae and cyanobacteria which have received attention in other parts of the world (De Deckker, 1982a). The majority of studies which have been undertaken have intentionally or fortuitously also involved pollen analysis, providing a basis for interpretation in relation to broader catchment areas.
The most widely studied animals are the small crustacean ostracods which occur in lakes throughout much of the Australian continent. These can be identified to species level from shell shapes and ornamentation, which are sensitive to physiochemical conditions in lakes (De Deckker, 1982a, 1988). They have been used to provide late Quaternary salinity and water level records for a range of sites, especially in southeastern Australia. These sites include volcanic crater lakes on the Western Plains of Victoria (De Deckker, 1982b; D'Costa et al., 1989; Edney et al., 1990), Lake George (De Deckker, 1982c), and Pulbeena Swamp (De Deckker, 1982d) where they have provided very valuable complements to other palaeoecological data, particularly pollen. Refinement of lake conditions, namely temperature, ionic composition and salinity has been achieved by use of the chemical composition of ostracod shells, especially the application of Sr/ Ca and Mg/Ca ratios (e.g. Chivas et al., 1986; De Deckker et al., 1988). Stable isotopes of carbon and oxygen, analysed from ostracod shells, have also helped reconstruct the origin of waters, their trophic status and salinity levels (Chivas et al., 1993).
The ecology and environmental controls on ostracod distributions in Australia, as well as their biogeography, is being further refined by multivariate analysis on several hundred modem assemblages from southeastern Australia (Lynda Taylor, pers. comm.) and a more general survey of the whole of the continent. Results indicate that a number of ostracod species have very specific requirements with respect to water chemistry and salinity. However, some fossil assemblages, notably in portions of the Lake George core, have
485
no modem analogues and it is thought that these may represent bicarbonate-rich waters which are rarely found on the continent today. There is a characteristic group of ostracods in the northern part of the continent which may provide useful information on the past incursion of summer rainfall conditions into southern Australia at times in the past. There is no evidence of evolution or extinction in Quaternary ostracods.
Challenging the supremacy of the ostracods in aquatic environmental reconstructions are diatoms which have been used in the study of Quaternary environments in North America and Europe for several decades and are now the most commonly used palaeolimnological indicators for the reconstruction of Late Pleistocene and Holocene lake histories in many parts of the world. Diatoms are sensitive to a range of water chemistry parameters including nutrient concentration, salinity, ionic composition and pH: community composition is also influenced by biotic factors such as grazing pressure and the relative proportion oflittoral to open water environments. Accordingly, diatom records have been used to provide insights into climate change, land use history, lake successional sequences, catchment processes and atmospheric pollution. Their global application is facilitated by the fact that identification can be achieved to species level and that most species are cosmopolitan and display consistent optima and ranges for key environmental parameters. Their abundance allows statistical treatment in a similar manner to pollen.
Despite these advantages, the study of diatoms in Australia has been retarded for a long period by the assertion of Joe Richardson (in De Deckker, 1982a) that the siliceous frustules of diatoms do not preserve well in sediments of a large number of aquatic environments, although an early study by Tudor (1973) did demonstrate the potential of the method. Within the last few years substantial advances have been made, largely by workers who took the time to learn methodology and taxonomy in foremost laboratories in the UK and France. A realistic appraisal of recent developments in Australia, acknowledging that there is still a problem with valve dissolution, has been undertaken by Reid et al. (1995).
Following Tudor's study and one by D'Costa et al. (1989) on volcanic lake sediments of the Western Plains of Victoria, Gell (1998) focussed on similar sites with reconstruction of past salinities soundly based on an extensive calibration set derived from over 150 surface samples from western Victoria (Gell, 1997). This approach has been adopted for factors influencing variation in billabongs and other river-related sites in the Murray-Darling Basin (Reid, 1997, Jenny Fluin,

486
pers. comm.) and changes in productivity within lakes and reservoirs within the eastern part of southeastern Australia (John Tibby, pers comm). The focus on recent climate change and human impact within the recent past has not provided any insights into evolutionary change among diatoms.
The study of sites primarily for pollen analysis has routinely included aquatic tax.a including some commonly occurring algae such as Botryococcus and Pediastrum. These have helped understand the nature of depositional sites and contributed to the reconstruction of past climates, but little attempt has been made to identify remains to a level that is useful in biogeographic or evolutionary studies. One notable exception is an early Holocene record for the monotypic species Brasenia schreberi, a floating leaved plant which was recorded in southern Victoria, well outside its present day range (Lloyd & Kershaw, 1997). This record suggests marked geographical response to higher temperatures during that
period. In contrast to overseas studies, few attempts
have been made to identify plant macro-remains to assist in aquatic community reconstruction though examination of dispersed cuticle at one site, Lynch's Crater, allowed the identification of swamp forest taxa present during oxygen isotope stage 5, and therefore a clearer separation of terrestrial and dryland vegetation (Bohte & Kershaw, 1999). One of the genera identified, Austromyrtus, demonstrates major range contraction as it has an older fossil record in Australia but no longer occurs on the continent.
ARCH AEOLOGICAL RECORD AND IDSTORY OF HUMAN SETTLEMENT
Although archaeology can be regarded as a distinct area of late Quaternary vertebrate palaeoecology and palaeobiogeography, people have had such an influence on faunal and floral biogeographic patterns that a separate section is considered warranted to emphasise the special position of human beings and to provide a basis for interpretation of late Quaternary changes in Australian biota. The influence of people stems from their time of arrival in Australasia, and particularly in Greater Australia. The timing of this arrival is still debated, in large part due to the practical limitations in radiocarbon dating. Where other dating methods, such as thermoluminescence (TL) or optically stimulated luminescence (OSL) have been applied, the antiquity of human colonisation as known from radiocarbon dating is extended by many thousands of years (see below). There is considerable debate in the literature over whether this is due to the use of different time clocks (ie radiocarbon or calender), or to the fact
AAP Memoir 23 (2000)
that the very earliest sites do not possess suitable carbon-bearing materials (such as charcoal or shell), necessary for radiocarbon dating (e.g. Malakunanja 2, Nauwalabila) (Allen, 1989; Allen & Holdaway, 1995; Chappell et al., 1996;Roberts et al., 1990, 1994). However, there are also other factors that contribute to a paucity of early archaeological sites, such as the difficulty of finding old, buried materials, many of which have been destroyed or deeply buried by subsequent geomorphological events. The very earliest sites in Greater Australia are likely to be located on the once-exposed continental shelf, having been inundated beneath tens of metres of water by rising sea levels since the terminal Pleistocene. Marine prehistoric archaeology is virtually non-existent in Australia, and the chances of ever finding such sites are very minimal.
The earliest undisputed evidence for people in Greater Australia comes from Carpenter's Gap rockshelter in the Kimberleys (Western Australia), radiocarbon dated to around 40 ka (O'Connor, 1995). Two radiocarbon determinations of 39,700±1000 and 39,220±870 BP, separated vertically by some 20 cm of sediments, were obtained on charcoal firmly associated with cultural materials (McConnell & O'Connor, 1997). The two ages are near the limit of reliable radiocarbon dating, and overlap within one standard deviation. This almost certainly indicates that at least the lowermost determination should be treated as a minimum age. How much older the deepest cultural deposits are is open to speculation.
Other early sites dated by radiocarbon means include Ngarrabullgan Cave in north Queensland (originally dated to >37,170 BP, but recently redated to c. 35,000 BP) (David, 1993), Upper Swan terrace in SW Western Australia (37,100+1600-1300 BP to 39,500+2300-1800 BP) (Pearce & Barbetti, 1981), the Willandra Lakes and lower Darling River system in SW New South ,;Wales (including Lake Mungo) (36,000±1100 and older) (e.g. Balme & Hope, 1990; Bowler et al., 1970; Bowler & Thorne, 1976), Warreen Cave in SW Tasmania (Allen, 1996), and Yombon in New Britain (Pavlides 1993; Pavlides & Gosdon, 1994). The fact that occupation in all of these early sites begins within a short span of time, has led Allen & Holdaway (1995) to suggest that this truly represents the timing of the earliest advent of people in Greater Australia. Furthermore, at Ngarrabullgan Cave early radiocarbon ages were paired with OSL determinations, indicating that radiocarbon and OSL/TL ages are unlikely to differ by more than 2.5 ka. The difference noted above between a 40
ka radiocarbon and an older, 60 ka TL/OSL year
AAP Memoir 23 (2000)
chronology (see below) is, therefore, almost certainly not simply due to an employment of two different time clocks (David et al., 1997).
The 37-40 ka limit for the antiquity of human presence in Australia, as informed by radiocarbon dating, is seriously challenged by Roberts et al. (e.g. 1990, 1994), who prefer to interpret the timing of the earliest radiocarbon ages as representing a methodological radiocarbon ceiling. They therefore employed TL in the late 1980s, and subsequently OSL, to date a number of sites previously suspected of being very old, but that could not be radiocarbon dated because of an absence of charcoal in the lowermost (i.e. earliest) deposits (Roberts et al., 1990, 1994; Roberts & Jones, 1994). At two sites from Kakadu, central north Australia -Nauwalabila and Malakunanja 2 - dates of 53 to 60 ka were obtained for sediments associated with cultural materials. At Malakunanja 2, the earliest cultural deposits were preceded by two metres of culturally sterile sediments. This was taken to indicate that people were not in Australia prior to about 60 ka by Roberts et al. (1990). This view is supported by a failure to find cultural materials in sediments (such as lunettes) dated to prior to 60 ka from other parts of Australia, despite many years of searching by both archaeologists and geomorphologists.
There have been other archaeological claims for the presence of people in Australia deeper in antiquity, but none of these has withstood the test of time. The most recent involve suggestions from thermoluminescence dating for human presence as far back as 116 ka and 176 ka (Fullagar et al., 1996)but recent OSL and AMS radiocarbon ages agree that the site is younger than 10 ka (Roberts et al., 1998).
It is commonly accepted by most, if not all, Australian archaeologists that the earliest peoples to reach Greater Australia came from Southeast Asia. The exact routes travelled are uncertain, but probably involved one of two possibilities:
1. A northern route from the Sunda shelf eastwards to what now are Sulawesi and Papua New Guinea or the Aru Islands, via smaller Indonesian islands.
2. A southern route, from Java (then a part of Sunda) through the what now are western Lesser Sunda Islands and Timor, southeastwards to Greater Australia or eastwards to the eastern Lesser Sunda Islands and then to Greater Australia (Birdsell 1977).
In each case, at least one sea crossing of more than 90 km was involved, making the first Australians the earliest known long-distance seafarers in the world. One implication is that co- operative behaviour (including language) of similar complexity to modern humans was
487
involved. This is due more to the requirements of planning and boat/raft construction than to negotiating the sea crossing itself. Some authors have argued that such behavioural implications would rule out early hominids such as Homo erectus (Davidson & Noble, 1992; but see Bednarik, 1997). This is consistent with the radiocarbon and TL/OSL chronologies, both of which lie firmly within the timeframe of Homo sapiens.
Considering the debate over the time of arrival, it is uncertain whether it took place during times of low sea levels, when sea crossings were shortest, or during times of rising seas, when population, territorial or resource shortages may have created social, political and/or economic pressures in the homelands. Resolution of this question depends on our ability to determine the timing of population movements and, consequently, of their arrival in Greater Australia. In either case, the first peoples were almost certainly Homo sapiens, and probably Homo sapiens sapiens, biologically akin to modern peoples (the earliest remains, including those of Lake Mungo, are fully modern) (e.g. Webb, 1989).
By 32 ka, the Huon Peninsula, New Britain, northern Australia, Tasmania, western New South Wales and southwestern Western Australia were all occupied. The only two major landscapes currently without occupational evidence by this early time are the arid core and the Southeastern Highlands. However, active sedimentation regimes in the former at least would render the discovery of such early sites very difficult; we do not, therefore, know . whether or not absence of evidence should be taken as evidence of absence.
By 22 Ma, the Last Glacial Maximum, all major Australian environments were occupied, including the arid zone, the Southeastern Highlands, and the Tasmanian high plateau (Cosgrove, 1995) although it is uncertain whether people actually lived in tropical rainforest until about 5 ka BP
(Horsfall & Hall, 1990). Most Australian archaeologists have
concentrated on researching early sites, either in search for 'the oldest', or so as to obtain long temporal sequences by which cultural trends and trajectories can be investigated. Despite this, it is only in some parts of Australia that Pleistocene sites are commonly found: the tropical north (the Kimberleys, Arnhem Land and Cape York Peninsula), the Western Australian sub-coastal zone, the dry lakes of western New South Wales, and southwestern Tasmania. Inthe tropical north, more than half of the 58 excavated and dated rockshelters have revealed Pleistocene cultural materials in each of the three regions (Lourandos & David, in press). Such high proportions of early

488 AAP Memoir 23 (2000) AAP Memoir 23 (2000) 489
sites are very unusual in the context of the rest of Australia, where Pleistocene rockshelters usually account for considerably less than l 0% of known sites. This northern distinction probably indicates demographic circumstances peculiar to northern Australia, including higher Pleistocene human populations in the warm, humid regions north of the tropic of Capricorn.
It was once thought that the first Australian settlers colonised the continent following the coast, where both water and marine resources familiar to seafaring peoples could be found (Bowdler, 1977). However, the numerous early archaeological sites subsequently found in inland regions implies that this is unlikely to have been the case (see also White & O'Connell, 1982). Indeed, Birdsell (e.g. 1957)had earlier argued that colonisation followed inland routes, via the once well-watered lakes and rivers of the arid and semi- arid core. However, chronological resolution is not presently fine enough to allow initial routes to be reconstructed with any confidence. This problem is heightened by uncertainty over the date when people first arrived on the continent.
There have been various attempts at modelling human responses to long-term environmental changes, but few have been systematically tested. Each model involves a contraction and expansion of populations and resource catchments with fluctuating fortunes, concerted with fluctuations
Australia's history. It was not until the mid to late Holocene, after the advent or invention of 1), systematic seed-based economies targetting the abundant r-selected seed grasses of much of arid and semi-arid Australia, and 2), extensive, risk- minimising kin networks capable of integrating vast population networks, that the dunefield deserts began to be permanently occupied. Until these mid to late Holocene innovations, barriers remained unoccupied, and corridors became abandoned during the Last Glacial Maximum when aridity was at its peak. Refuges alone remained occupied during the entire span of Australia's human history.
Hiscock (1988) and Lamb (1996) have also posited fluctuating settlement systems during the period covering the Last Glacial Maximum. Independently, their research at Lawn Hill (Hiscock, 1988) and Chillagoe (Lamb, 1996),both in Queensland, have revealed a tethering, so to speak, of human activity around sources of permanent water during periods of aridity, such as between 22 and 17 ka. This included a shift from the use and reduction of distant to local stone sources, and an increased incidence of site use, together interpreted as representing a reduction in the size of resource catchments. These tendencies are consistent with the settlement models presented above.
The most pronounced changes evident in the
20°
40°
:::: Playa lake G;;J Lake
-·· Mangrove swamp and coastline D Rainforest
Temperate forest D Savanna woodland
Tundra I steppe grassland D Xerophytic woodland and shrubland
160°
6 ka
t 'b
in palaeoclimates. Horton (1981) argued that the first Australians colonised the continent by following the better-watered regions, including coastlines, major river systems, and other areas
Australian archaeological record took place during the mid to late Holocene. It is then that off-shore islands begin to be systematically visited and used (Barker, 1995; O'Connor, 1992), the intensity of
120° 140° 160°
Fig. 3. Generalised reconstruction offeatures of the Australasian region at the height of the present (Holocene) interglacial, centred on 6 ka.
where water was available. Consequently, all parts of Australia except for the arid core were occupied early in the colonisation process. Subsequently, semi-arid areas became abandoned as aridity increased during the Last Glacial Maximum . In tandem with environmental fluctuations, people retreated to the coasts and major river systems, where surface water could be readily found. It is not until about 12 ka, when conditions again became suitable, that they returned to the hinterland. Veth (1989) proposed a settlement model for Australia's arid core based on general biogeographic principles , in many ways similar to Horton's. He argued that the Australian continent can be divided into three broad biogeographic types: refuges, where 'networks of permanent water sources capable of withstanding climatic extremes' can be found; corridors, where water sources have fluctuated significantly through the course of the Late Pleistocene and Holocene; and barriers, sandridge deserts where water is not commonly available. Given an absence of water sources, Veth suggested that dunefield deserts acted as barriers to human settlement for much of
occupation of individual sites increases markedly, intensities of regional land use increase, previously little used environments or resources become systematically exploited, often at a grand scale (e.g. eels in southwestern Victoria), large-scale fish traps first appear, grass seeds first begin to be systematically exploited, toxic plants begin to be processed, rendering them edible, and rock art becomes regymalised (see also McNiven, in press). Together, tnese factors imply both a significant population increase and a re-structuring of economic and politico-territorial networks. It is during this period that the arid core of Australia probably began to be permanently settled, a phenomenon in part made possible by the beginnings of systematic seed grinding. Lourandos (1983) has argued that this process of change involved an 'intensification' of both people-land and people-people interactions, and a possible increase in the structural complexity of relations between the two (see also David & Lourandos, 1998). It has also been argued that until the mid to late Holocene, long-term cultural trends as measured by site occupation broadly
mirrors environmental trends, as measured over the long term (i.e., occupational intensities increase with natural levels of bioproduction [Lourandos & David, in press]). After c. 5 ka, however, cultural trends diverge with occupational intensities increasing throughout the Holocene (but see Hiscock, 1986).David & Lourandos (1998) have recently argued that this divergence probably began during the early Holocene, when conditions were favourable to demographic expansion. This established a demographic momentum that continued well after the end of the early Holocene 'optimum'.
Intensification of occupation in Australia from about 5 ka is paralleled by an explosive human colonisation of the Southwest Pacific Islands from Melanesia (it is not known if these two processes were related), terminating in the eventual peopling of New Zealand (Enright & Gosden, 1992). The environmental impact on these islands was severe as a result of the introduction of new plants and animals, clearance of the native vegetation for agriculture, often using fire, and hunting of native fauna. Results included widespread species
extinction and accelerated soil erosion on hill slopes and deposition on adjacent lowlands (Enright & Gosden, 1992). Although these events are geologically recent, there is still a great deal of debate on actual times of arrival of people on individual islands and subsequent timing and specific causes of various impacts. Debate is nowhere more heated than in New Zealand where three occupation chronologies have been proposed; 'long (>1.5 ka), intermediate (c. 1 ka) and short (<0.6 ) (McGlone &Wilmshurst, 1999). Dates for settlement are based generally on radiocarbon and tephrochronological ages of first evidence for substantial and sustained burning and vegetation change from charcoal and pollen in sedimentary deposits, and on early archaeological sites. From an analysis of all sedimentary records, McGlone & Wilmshurst (1999) have concluded that variation in ages can be explained largely by various forms of contamination and that bogs provide the most reliable ages. These support the short chronology with most dates falling between 0.55 and 0.65 ka. However, support for at least ephemeral early

491 490 AAP Memoir 23 (2000) AAP Memoir 23 (2000)
D
contact, if not settlement as per the long chronology, has been boosted by AMS determinations on the bones of the introduced Pacific rat (Rattus exultans) which have provided dates as old as 1.8 ka (Holdaway, 1996), although, predictably, these early dates have been questioned (Anderson, 1996). Biogeographically, this debate is important for determination of pattern and rates of faunal extinction. It also has a major bearing on causes of extinction as rats could have had a marked impact on the biota perhaps before the major phases of human colonisation and establishment.
EXIREMESOFTHELASTGIACIALCYCLE
The detail available for the mid Holocene 'optimum' and the Last Glacial Maximum (LGM) provides some indication of the degree of environmental change which has characterised much of the Quaternary period as well as biogeographic responses to it. These periods provide, in many ways, recent analogues for the less well known earlier part of the Quaternary although they differ in that people were present and the megafauna had become extinct. Although the focus here is placed on the response of organisms, it .must be emphasised that estimates of environmental change are based on biological data, generally supplemented by complementary
14C years) and may have exceeded them by up to a metre in some places towards the end of the period. Overall, climatic controls over the region were similar to those of today with the exception of the El Nifto-Southem Oscillation (ENSO) whose activity is likely to have been lower until the very end of this period (McGlone et al., 1992).
Biogeographic patterns were not significantly different to those of today at the broad regional level. Rainforest had a marginally expanded distribution but essentially retained a similar fragmented and remnant distribution as it does today in Australia. In New Guinea and New Zealand, rainforest had expanded to cover most of these islands, replacing grassland and open forest in drier areas and grassland and subalpine vegetation at highest altitudes. In southeast Australia at least there was also a greater coverage of both wet and dry sclerophyll forest. Mangroves, which had flourished during the marine transgression, retained a much greater abundance than today although there is little evidence for a greater spatial distribution. There is little information on vegetation from the arid and semi- arid parts of Australia, but greater water availability in what are now ephemeral lake systems would have markedly altered aquatic assemblages. It is interesting to note though that the major playa lakes only filled partially during
20°
400
Present day coastline - Existing coastline •••• lOOmm isohyet D Lake '.:::; Dry lake • Glacier, ice cap - Periglacial activity A Active dunes
D Rainforest D Dry forest I woodland
Alpine I steppe grassland D
18 ka
• C5 20°
- - - - o - - - - - -- -,- - - - - - - - 0 0
0
information derived from the physical sciences. The mid Holocene 'optimum' occurred within
this interglacial period (eg. Magee et al., 1995). It is possible that even the small difference between
,.,.,.,,- Limit of Red Kangaroo (Arid meggiauna) · 120° 140° 160°
the range 7 to 5 ka (Fig. 3). For most of Australasia, palynological data suggest that conditions were wetter, perhaps by up to 25% and warmer, by about
this period and today has been augmented by the impact of Aboriginal people as intensity of occupation substantially increased throughout the
Fig. 4. Generalised reconstruction offeatures of the Australasian region at the Last Glacial Maximum, centred on 18 ka.
1°C, than present with reduced seasonal variation or inter-annual variability (Dodson & Harrison, 1993; Kershaw, 1995;McGlone et al., 1993). The period though does not necessarily incorporate either the highest temperatures or the most effective precipitation for all areas. In northeast Queensland, for example, bioclimatic analyses on pollen data indicate that highest values were achieved after 5 ka -although in concert with increased variability (Kershaw & Nix, 1988) - while similar analyses in southeastern Australia (Lloyd & Kershaw, 1997; Markgraf et al., 1991) suggest that maximum temperatures occurred earlier, around 9 ka. However, these estimates are derived from wetter areas where high and more continuous cloud cover may have effectively reduced incoming radiation during the 'optimum' period. Sea surface temperatures are also considered to have been slightly higher, and have been demonstrated to have been along the western and southern coasts of Australia with a more intense Leeuwen current (Kendrick, 1977; Murray- Wallace et al., 2000). Sea levels reached their present day levels by about 6.5-6 ka (uncorrected
Australian continent shortly after 5 ka. This event may have been triggered by the loss of environmental equability at this time or, as suggested by David & Lourandos (1998), environmental patchiness was created by a fragmentation of the political landscape after the mid Holocene, resulting in patch burning and use of accessible landscapes.
Conditionjl<luring the Last Glacial Maximum, centred on 20-18 ka (Fig. 4), were very different to those experienced by the first European colonists. In line with the rest of the world temperatures were cooler but, from available estimates, the degree of temperature lowering is uncertain. In southeastern Australia, New Zealand and the New Guinea Highlands, evidence from the extent of glaciers and position of the treeline suggest that mean annual temperatures were in the order of 6°C lower at higher altitudes and perhaps 4-6°C lower in the lowlands. The application of bioclimatic profiles of rainforest taxa to fossil assemblages in the Otway Ranges of Victoria suggest that temperatures were no more than 6°C lower(McKenzie & Kershaw, 1997),while mapping
of the extent of periglacial activity in southeastern Australia indicated a depression of up to 9°C (Barrows, 1995). A similar surprisingly extreme estimate has been proposed from amino-acid racemisation of egg shells preserved in the central part of Australia (Miller et al., 1999). These estimates of temperature lowering for terrestrial environments are generally larger than those predicted from global climatic modelling (Harrison & Dodson, 1993; M cGlone et al., 1993) and from sea surface temperatures based on distributional changes in foraminifera (Barrows et al., 1995; Wright et al., 1993), which do not exceed3-4°C. Greatest debate is over the extent of temperature lowering in the tropics. There is little evidence of temperature change from the terrestrial lowlands within the region but temperature lowering in pollen records in the Indonesian region immediately to the north suggest reductions in the order of 6°C, similar to those at higher tropical altitudes and to higher latitudes (van der Kaars, 1998). Sea surface temperature estimates vary from 0-2°C based on foraminiferal assemblages
(Barrows et al., 1996; Thunell et al. 1994) to about 5°C from tropical corals (Guilderson et al. 1994) lower than today.
During the Last Glacial Maximum, both absolute and effective precipitation was reduced. Estimates from biological and geomorphic data suggest a reduction ofup to 50% of those of today. These differences are much greater than those suggested from global model results (Wright et al., 1993). One major influence was the temperature lowering which reduced the ability of the atmosphere to take up moisture and which, in the tropics, caused a reduction in cyclogenesis. The situation was exacerbated by substantially reduced sea levels, down about 130 m, with the build up of ice in the northern hemisphere. Low sea levels resulted in greater continentality and moisture reduction within present terrestrial landscapes due to the loss of extensive continental shelves and coastal mountains. Evidence from corals, molluscs and marine plankton suggests that transport of warm water to higher latitudes along the western and southern Australian coasts was

( ';8 r Wi
=========== Winteraceae)
ar:!,d!fe:_ c!.a!!,w_;l (rsA:fa' J;J
I
=
492 AAP Memoir 23 (2000) AAP Memoir 23 (2000) 493
substantially lowered by the switching off of the Leeuwen current. Despite reduced temperatures, evaporation rates may have been high due to
considered most suitable for rainforest survival were covered in sclerophyll vegetation (Hopkins et al., 1993, 1996). The most likely pattern of
Gauss Matuyarna
i. [ Brunhes
higher winds associated with the compression of pressure belts towards the equator, although there is uncertainty as to whether the critical boundary between the westerly winds and subtropical high pressure belt across central Australia was located to the north or south of its present position (Bowler et al., 1976; Harrison & Dodson, 1993). Marine evidence suggests strongly that frontal
survival was as small patches, particularly along fire-protected and locally moist drainage lines and extending onto the exposed continental shelf. Similar patterns of rainforest survival have been suggested for New Zealand (McGlone et al., 1993) and southeastern Australia (Kershaw, 1995). In this latter area, some substance has been given to this hypothesis by the location of a cool
: 8 b e. C: o b o II I I ,' I I I I II I • I I • 0 0 )'!!'!L I _ l L_' _ --- LL Li
il' zones were north of their present positions during temperate rainforest 'refugium' in the Otway IMangapanian 1 IHaulawanl g Marahauan I Okehuan I Putikian
I [ I Nukumaraun I Castleclifian I Haweran
sand dune activity over much of the continent is
1999).
PLIOCENE
· PLEISTOCENE B PLEISTOCENE A
PLEISTOCENE gi.r.., j the last glacial period. The evidence for extensive consistent with drier, windier conditions.
There may have been some regional variation in the degree and even direction of change in effective precipitation. Little change is noted in records from highland New Guinea, but this may
Ranges of western Victoria (McKenzie & Kershaw, Mangrove pollen has strong representation in
coastal and marine records during the marine transgression, continuing to some degree through the Holocene period, but there is little evidence for mangroves at the Last Glacial Maximum.
H"' "' EARLY !( EARLY
:::ti z i;:c "T1-Cibicides molestus (F) . 11 11 11 11 Crqssostrea mg!!ns (Bl
$!! :5' Ph1a/opecten tnphook1 (B) Patro (B) "- Glycymeris (B) 5· ;! -------------------Eumarcia (B)
MIDDLE
simply be a function of the lack of sensitivity in these ever wet environments and certainly there
Reduced precipitation and temperatures would have had some influence, but the most important
15" - s· Bolivina fifei (f) . - $ RiticulOfenestra asanoi (mt.)
§ a Reticulojenestrp8 ,,::C, lia'>Jcunosa (N) Stylatractus universus R)
was regionally lower precipitation as indicated by pollen evidence from the ocean cores. Some lakes in the interior area of southwestern New South Wales appear to have supported higher water levels than present (Harrison & Dodson, 1993) which may have been due to a higher regional water table with excess moisture derived from snow melt in the southeastern highlands. It may also be that dates are unreliable as even in moister areas of southeastern Australia, there is often little
factor would have been the loss of shallow water habitats with the retreat of the sea to the continental margins. Survival would have been limited to narrow patches along the continental slope.
Rainforest vegetation and other mesic communities, particularly the eucalypt dominated wet and dry sclerophyll forests in southern and eastern Australia, were regionally replaced by more open eucalypt woodlands in Australia
Zygochlamys delicatula Globorotalia crassula Rotalia wan[;==ia nuncatulinoides (F)
Geophyrocapsa sinuosa (N) Globorotalia crassafonnis (dextraIJe )hyrocapsa omega (N). .
Pecten marwicki Pecten kupez ( Pe-ctena-otea-(B) ---
Pecten novaezealandiae (B) - Emiliania huxleyi (N) -
Not!wfagidites spinosus (Brasso¥XJra) Haloragacidites amolosus (Haloragis - Haloragaceae) Nothofagidites lonispina (Brassospora) Hamsipollenites annulatus (Zygogynum -
------Sapotaceoidaepollenites latJzam;i-tus (Sapotaceae) ======- Cupanieidites (Cupaneae - Sapmda.ceae).
sedimentary evidence for the actual height of the including some lowland parts of New Guinea, ========= Beaupreaidites elegansi orrna (Beauprea - Proteaceae)
last glacial period. Although conditions were generally dry, the Last Glacial Maximum was except for the extreme southeast of the continent, which, like most of New Zealand, was covered in :: Polycolpites reticulatus Reevsza Sterculiaceae)
Podosporites parvus ( icrocachrys -.Podocarp) fl:gfe!ifd1f;: ':k i 1f::' i1e-sJPhorbiaceae)
Motho gi & 1es matuu§rof'ondso1spsorraitsessweprourgaatus (MicrostroJoig,pJ!lf!'}, ff) iatus (Phymatosorus _ Polypodiaceae) wetter than the period succeeding it, from about 15,000 to 12,000 BP when rising temperatures on an essentially glacial landscape reduced effective precipitation to lowest levels within the last glacial cycle (Kershaw & Nanson, 1993).
One major regional response to glacial conditions was a reduction in the extent of mesic communities, particularly rainforest and mangroves. The only.recorded areas of substantial rainforest are in New Guinea and Northland, New Zealand. Both terrestrial and marine pollen records from the humid tropics area of northeastern Australia have demonstrated that rainforest survived but the regional study of soil charcoal has failed to locate its position. Even those areas
grassland steppe and low shrubland vegetation. It is considered that the almost total replacement of trees by steppe would require conditions more extreme than those accountable by reduced temperatures, rainfall and stronger winds, and McGlone (1988) postulates also the occurrence of frequent incursions of cold southerly winds and regular frosts for this period. There is little direct evidence o:Vuid land vegetation except that the cover must have been reduced to allow extensive mobilisation of sands over a large portion of Australia. The occurrence of red kangaroo bones though, outside of the present natural range of the species, indicates expansion of arid zone ecosystems (Horton, 1984).
========-_::_:-_:'._:-_:7_ _::7--:::7:---:7- ::-:-:---:--::-:--:-:;N;;o;;th,;o[_ Proteacidites minimus (Proteaceae spp.)
Cyathodesfasciculata Toronia toru (Persoonia----------M:cy:- o-porum--:la-ctu-m--(::M-;y-o-,porace-:a-e-;- - ---===
Proteaceae) Passijlora tetrandra (Passifloraceae ca_en_a-(Rosace_ae)_ -- Hedycarya (Monimiaceae)--
IIIWanganui Basin
·--Wellington •Central Volcanic Region •Northland
-Westland
---DSDP Site 594
Fig. 5. Pollen stratigraphic ranges for New Zealand over the last 3 Ma centred on the Wanganui Basin, in relation to the marine biostratigraphy for the Wanganui Basin and New Zealand biostratigraphic stages (Naish et al.,

1998), the timescale of Lourans et al (1996) and the composite benthonic oxygen isotope record of Shackleton et al. (1995). Pollen data from Bussell & Mildenhall (1990), Mildenhall (1978, 1980), Mildenhall & Suggate (1981) and Nelson et al. (1988). Detailed pollen records constructed for the Wanganui Basin (Bussell, 1986, 1998, 1990; Bussell & Pillans, 1992), Wellington (Mildenhall, 1983, 1994, 1995), Central Volcanic Region (McGlone & Topping, 1977, 1983), Northland (Newnham, 1999 and refs therein), Westland (Moar & Suggate, 1996), and DSDP Site 594 (Reusser & van de Geer, 1994).

494 AAP Memoir 23 (2000) 495 AAP Memoir 23 (2000)
-
:
_
Although it is possible to define the gross floristic and structural features of major communities, many assemblages were different
PATTERNS OF CHANGE THROUGH THE QUAlERNARY
Discussion is focused on three major areas
Gauss
Matuyama
0 I
Brunhes
to those of today. This is not unexpected as the range of climates was different to those at present. There is no identified modern analogue for the extensive grassland steppe vegetation (Kershaw, 1998) and components must be hidden in a variety of alpine, subalpine and cool woodland communities (Hope, 1994). One major component , a type of Asteraceae which produced a distinctive
where a combination of suitable sedimentary sequences and interest in analysis of a range of palaeoecological indicators has provided at least some picture of changing environments and biotic responses through much of the Quaternary period. Summaries of sequences from these areas are shown on Figures 5 to 7 in relation to the global marine oxygen isotope stratigraphy of
b b G: 0 b G; . t . I •. . .. I I I I d
if .,, type of pollen characterised by 'blunt spines', Shackleton et al. (1990, 1995) tuned to the !Mangapanian [ IHaulawanl o S Marahauan I Okehuan I Putildan I Haweran I Nukumanrun I Castleclifian I
Given the form name of Tubulifloridites
and the New Zealand biostratigraphy (Naish et
PLIOCENE
p.·
PLEISTOCENE B PLEISTOCENE A
I PLEISTOCENE 1 has been recorded only rarely in the Holocene . astronomical timescale of Lourens et al. (1996) ! EARLY H EARLY MIDDLE
pleistocenicus by Martin (1973), it may relate to several pollen taxa including Calomeria and
al., 1998). Time ranges only of first and last appearances of major biotic indicators are included
---- - - - - Araucariaceae
- - - - - - - Notlwfag us (Brassospora
) spp.
Cassinia arcuata (Macphail & Martin, 1991), but it is also possible that at least some component taxa have become extinct. In northeast Queensland, a rainforest genus, Dacrydium , appears to have become extinct within the Last Glacial Maximum (Kershaw, 1994). Within the fauna, Hope et al. (1978) record a more mixed faunal assemblage on Kangaroo Island, South Australia, than is present today including rodents from dense scrubs to open shrublands and pygmy possums now restricted to Tasmania.
In terms of general biogeographic responses to the environmental extremes, the majority of taxa, although responding in an individualistic manner, generally appear to have expanded and contracted within particular ranges rather than undertaking wholesale migration as is characteristic of organisms in glaciated parts of the world. In an examination of regional patterns of change in the Victorian region, rainforest expanded only marginally in the Holocene from retreats occupied during the glacial period (Kershaw, 1998). Greater expansion was probably inhibited by limited dispersal and by a sensitivity to fire in exposed situations. There is no indication that areas of rainforest separated by more than a few kilometres coalesced during the Holocene 'optimum' .
but the inferred time ranges of terrestrial floral sequences providing largely continuous and quantitative data are also discussed.
The most complete sequence is centred on the Wanganui Basin of New Zealand where a record of marine cyclicity dating back to 3.6 Ma has been identified and correlated with the astronomically- tuned marine isotope record, from palaeo- magnetism, oxygen isotope ratios, biostratigraphy, cyclostratigraphy and tephra-chronology (Naish et al., 1998). Evidence is provided of changes in foraminiferal, mollusc and pollen assemblages from this area and the broader region of the North Island (Fig. 5). Southeastern Australia has been the centre of Cainozoic pollen and plant macrofossil studies, but chronostratigraphical control is poor for the uneconomic Quaternary part of the record (Fig. 6). By contrast, a great deal of recent effort has been put into the establishment of the offshore stratigraphy of northeastern Australia, particularly to assist in the dating and understanding of environments associated with the development and history of coral reefs (McKenzie et al., 1993). The onshore and offshore records have also been tied together by palynological studies of isotopically and biostratigraphically dated offshore corey(Fig. 7).
- - - - - - - Dacrycarpus (Podocarpaceae) - - - - - - - Dacrydium cupressinum type (Podocarpaceae) - - - - - - - Cyatheocides annulatus (Cyatheaceae)
- Cassinia arcuata type (Asteraceac)
••..,,, Hapuka 1
Da crydium cupressinum type (Podocarpaceae) Araucaria (Araucariaceae)
Ascarina (Chloranthaceae) Dryandra type (Proteaceae)
--------- Beauprea (Proteaceae) --------- Notlwfagus (Brassospora ) spp.
Dacrycarpus (Podocarpaceae) Austromyrtus (m) (Mynaceae) Rubus (m) (Rosaceae)
Lauraceae (m) Allo casuarina sp. (m) (Casuarinaceac)
Banksia spp. (m) (Proteaceae) Austrobuxus (Euphorbiaceae) <ol J
Eucalyptus spathulata type (Myrtaceae) ::r Callitris I Actirwstrobos (m) (Cupressaceac) Q
--'---- ----------------- Symplocos (Symplocaceae) ('b ---------------------- Haloragodendron type (Haloragaceae) >-J
--- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Cunoniaceae(tricolpate) I» --------------------------- Quintinia (Escalloniaceae) "3'
Banksia kingii type (m) (Proteaceae) --------------------------- Gothanipollis perplexus i::
Astelia ----------------------- (Loranthaceae) !».
· I Linda Valley ·..:i • Regatta Point · -Lake Selina
-Pulbeena Swamp
·-Darwin Crater •Core S036 7SL
-Egg Lagoon
Microcochrys (Podocarpaceac)
--- Nothofagus (Fuscospora) spp. --- Dacrydium cupressinum type (Podocarpaceae) --- I.agarostrobos (Podocarpaccae)
--
-·---
--
--
--
--------- Araucaria (Araucariaceae)
---------- --------------------------------------------------------------------------------- ;g:i:;ti;;...<1tf:: r:c:r 0
Fig. 6.Pollen and plant macrofossil stratigraphic ranges for areas within southeastern Australia over the last 3 Ma in relation to New Zealand biostratigraphic stages (Naish et al., 1998), the timescale of Lourans et al (1996) and the composite benthonic oxygen isotope record of Shackleton et al. (1995). Pollen data for offshore Gippsland Basin from Macphail (1997) and Alan Partridge (pers. comm.), for onshore Tasmania from Macphail (1997)
Phylloc/adus (Podocarpaceae) [ ---------------------------- Gunnera (Gunneraceae) ;:;:
• •Lake George gJ I Frankston Quarry
Macphail et al. (1995) and Jordan et al. (1995), and for mainland southeastern Australia from Kershaw et al. (1991), McKenzie & Kershaw (2000), Merna McKenzie (pers comm.) and Barbara Wagstaff (pers. comm.). Plant macrofossil data from Hill & Macphail (1995), Jordan (1995a,b), Jordan et al. (1995) and Macphail et al. (1993). Detailed pollen records from Hapuka - 1(Macphail, 1997), Lake Selina (Colhoun et al., 1999), Pulbeena Swamp (Colhoun et al., 1982), Darwin Crater (Colhoun, 1988), Core S036 7SL (van de Geer et al. 1994), Egg Lagoon (D'Costa et al., 1993; D'Costa, 1997), Lake George (Kershaw et al., 1991; Singh & Geissler, 1985), Frankston Quarry (Pearson, 1996), Core E55-6 (Harle, 1997), Lake Wangoom (Harle et al., 1999), Lake Terang (D'Costa & Kershaw, 1995; D'Costa, 1989), Pejark Marsh (Barbara Wagstaff, pers. comm.) and Wyelangta (McKenzie & Kershaw, 2000).
•,:)
-Core E55-6
- Lake Wangoom
• • Lake Terang
Pejark Marsh
•Wyalangta
[ p;·

496 497 AAP Memoir 23 (2000) AAP Memoir 23 (2000)
I <g t:S ..a 0
: 8
r ;a
-
/ r
Gauss
;' 0
!·
Matuyama
Brunhes
til ::i z 0 0
-o-r &"_a o[
to 2.2 Ma is suggested, although this is likely to be an artefact of representation of discontinuous sequences and uncertain dating as well as geographical spread of sites rather than a series
temperature cannot be regarded as critical for these extinctions . However, any reduction in precipitation or increase in climatic variability would have been important for the survival of
b v. 0 b C:
I I I I I I I ..., ... I I
§Z§c. "! C>l.l
of abrupt events. Again there is little response at or around 1.8 Ma , with only Microstrobos disappearing slightly after this time.
rainforest generally within an area which is largely much drier than New Zealand. In the case of both southeastern Australia and
l :;<!
Marahauan I Okehuan I Putikian
c-"'g:O 'l
e:'z
t'1j Insoutheastern Australia, the only firmly dated pollen record which transcends the Plio- Pleistocene is that from Lake George which, from pollen evidence, indicates the abrupt decline of a
New Zealand, the scale of global climate change suggested from the marine isotope record appears insufficient to account for the extent and degree of correspondence of vegetation change \Mangapanian [ IHau!awaxil Nulrumaraun
2 §1 l I Castlcclifian l
Haweran 1il <> Cl.l S!.' a·! number of essentially cool temperate rainforest experienced around 2.5 Ma without an additional
g_ PLIOCENE PLEISTOCENE B e PLEISTOCENE A PLEIS1DCENE
EARLY EARLY MIDDLE taxa at some time within the latter part of the Gauss regional influence. One possible influence is !" il I &!: § E!'.c. 0
> o":";; filO 00
palaeomagnetic chrone, that ended about 2.6 Ma. There is little vegetation response around 1.8 Ma although one rainforest taxon -Podocarpaceae -is regionally present to about 2.2 Ma. Records from Tasmania and the offshore Gippsland Basin,
suggested by Bowler (1982) who, in an attempt to explain sedimentological sequences in southeastern Australia including that of Lake George, postulated that the steepening of the latitudinal gradient with the development of the
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Nothofagus (Brassosp ora) ------------------Dacrydium cupressinum type (Podocarpaceae) ------------------Dacrycarpus (Podocarpaceae) Dacrydium guillauminii (Podocarpaccae )
Austromy rtus (m) (Myrtaceae) --------------------------- Araucaria (Araucariaceae )
- - - - - - - - - - - - - - - .. - - Camptostemon (Euphorbiaceae)
ODP 823
------------ ODP Site 820 (General) - ODP Site 820 (Detailed)
- Lynch 's Crater
-Banda Sea
-Lombok Ridge - - - - Arau caria (Arancariace ae)
-----Fraser Island (Hidden Lake)
Fig. 7. Pollen stratigraphic ranges for northeastern Australia over the last 3 Ma in relation to New Zealand biostratigraphic stages (Naish et al., 1998), the timescale of Lourans et al (1996), the composite benthonic oxygen isotope record of Shackleton et al. (1995) and the planktonic isotope record of Peerdeman (1993). Pollen ranges from Bohte & Kershaw (1999), Kershaw et al. (1993), Moss & Kershaw (2000) and Martin & McMinn (1993). Pollen records from ODP Site 823 (Martin & McMinn, 1993), ODP Site 820 (Kershaw et al., 1993; Moss, 1999), Lynch's Crater, (Kershaw, 1986), Banda Sea (van der Kaars et al. (2000) and Lombok Ridge (Wang et
reviewed by Macphail (1997), show similar marked declines in rainforest or wet heath taxa around the Plio/Pleistocene boundary, and it is likely that in future, more continuous and better dated sequences will indicate some synchroneity of change about or slightly after 2.6 Ma.
Innortheastern Australia, there is a coarse but almost continuous record through the late Cainozoic and little evidence of floral turnover relating to either definition of the Plio-Pleistocene boundary. The only possible disappearance of a taxon at a palynologically visible level is Nothofagus subsection Brassospora but many other recognisable taxa which became extinct in southeastern Australia, and also New Zealand, survived in the northeast.
The most obvious explanation for the changes in New Zealand is a reduction in temperature indicated by marine indicators and suggested from pollen data by the loss of a number of taxa, including Sapotaceae, Cupaniae and Beauprea, which are now restricted to moist environments at lower latitudes. However, the loss also of taxa
Antarctic ice sheet resulted in the formation, expansion and movement northwards of the mid latitude high pressure belt. This contributed to continental drying in the Pliocene and to a switch from summer to winter rainfall in southeastern Australia around 2.5 Ma as this region, for the first time, came under the influence of the westerly wind belt. Such a change in seasonality have had a more dramatic impact on rainforest taxa than any reduction in temperature and rainfall and its influence is likely to have extended to New Zealand .
The lack of any substantial response in the flora of northeastern Australia at the Plio-Pleistocene boundary is consistent with the Bowler hypothesis in that nomajor change in atmospheric circulation is predicted for this part of the world and also with a reduced impact of any steepening of the temperature gradient in these tropical latitudes. The major oscillation in sea surface temperatures postulated for the Coral Sea in the Late Pliocene, culminating in a sharp rise in temperature around 2.5 Ma, appears not to have
al., 1999).
Plio-Pleistoc ene boundary There are few areas where it has been possible
to address the positioning and significance of the Plio-Pleistocene boundary. One of these is New Zealand where there has long been interest in relating the local biostratigraphic scheme to the global Quaternary stratigraphy. Evidence for cooling, noted as early as the 1940s, resulted in the base of the Nukwnaruan being correlated with the Plio-Pleistocene boundary (Fleming, 1953). Subsequent dating of this stage produced contradictory results and uncertainty about its global relationships. However, the weight of evidence, including that from Wanganui, indicates an age close to the Gauss/Matuyama boundary
,,, and adds weight to the proposal to move the Plio- Pleistocene boundary to around 2.6 Ma (Naish et al., 1998). A concentration oflast occurrences of typical Pliocene warm water bivalve molluscs and the first representation of a number of cool water taxa, including foraminifera, coccoliths and bivalves around 2.5 Ma and the lack of any real assemblage changes around 1.8 Ma, is a good illustration of the significance of the proposed boundary.
On land, changes in the pollen flora of North Island are even more dramatic, although extinctions far outweigh first appearances and include a variety of arboreal or shrub taxa, with rainforest affinities. A stepwise elimination from about 2.5
which are characteristic of cool temperate forests, such as Microcachrys and species of Nothofagus subsection Brassospora, suggest the influence of other factors of which increased climatic variability, particularly in the case of Nothofagus subsection Brassospora, presently restricted to montane environments in New Guinea and New Caledonia, is likely to have been important. Some increase in taxa of more open environments suggest also that somewhat drier conditions may have accompanied this variability.
In southeastern Australia, almost all identifiable taxa which became extinct have representatives in cool temperate rainforests or have extant relatives whose predicted temperature ranges should allow them to survive at these high latitudes (Kershaw, 1997). Consequently any decrease in
had any long term effect on the vegetation, although the period itself is notable for a lack of preserved fossil evidence in offshore cores analysed to date (Kershaw et al., in press).
Early Pleistocene I This interval covers at least two-thirds of the
Quaternary period, yet, because of a sparsity of 1 . evidence or at least dated evidence, is virtually ignored in any reconstruction of Quaternary environments or in its potential role in the development and understanding of present day environments and biogeography.
The substantial changes at the Plio-Pleistocene boundary in southeastern mainland Australia resulted in a landscape dominated by vegetation that has no recognisable modern analogue.

498
Asteraceae, including high values of the distinctive morphological type Tricolporites pleistocenicus, dominated the herbaceous component of the vegetation, while Casuarinaceae composed the major arboreal element, during the recorded period from Lake George (c. 2.5-1.5 Ma) (Kershaw et al., 1991), and the early part of the period in the Hapuku-1 core, offshore Gippsland Basin (Macphail, 1997). This combination continued as important through much of the Quaternary from Gippsland Basin evidence (Alan Partridge, pers. comm .). A dominance of Asteraceae is indicated around a TL age of about 1 Ma in a short sequence from a sand quarry bordering Port Phillip Bay (Pearson, 1996). In the western plains of Victoria, a 70 m core from the volcanic crater of Pejark Marsh provides a semi- continuous, fission track dated, pollen record from about 0.7 to 1 Ma (Wagstaff et al., in press). Poaceae shares dominance withAsteraceae in this record, probably because the heavy basaltic soils favoured grasses as they do today, while trees are relatively limited in their representation, with again Casuarinaceae as the most conspicuous component. A further problematic element is the frequent occurrence of Cyathea spores suggesting the representation of separate tree fem dominated communities.
The general impression from the vegetation is that, although climatic cyclicity is evident, it was not the biogeographic force that is seen in the later part of the Quaternary, with climate not achieving extremes of temperature or precipitation. From the modem pollen data set in southeastern Australia, the closest matches are with some semi-arid spectra where Casuarinaceae woodlands are most conspicuous and the dry conditions are conducive to proliferation of opportunistic Asteraceae. However, conditions appear to have been cooler than present as T. pleistocenicus values are much reduced during later interglacials. This reconstruction appears at odds with interpretations of the marine isotope record which suggests that conditions should have been as warm as today. Consequently it is likely that other factors were important in determining the vegetation cover of this period. One possibility is that major and predominantly arboreal components of the pre-existing vegetation were unable to adapt to the proposed switch from summer to winter rainfall and that cosmopolitan elements like the daisies filled this void. Major casualties were the Araucariaceae, indicative of drier rainforests which expanded over much of the eastern part of Australia as wetter rainforests declined during the Late Tertiary, and also the eucalypts and related Myrtaceae which, in the eastern part of the Murray Basin at least (Kershaw et al., 1994;
AAP Memoir 23 (2000)
Martin, 1991), appear to have formed a kind of wet sclerophyll forest in association with high levels of burning. Casuarinaceae maintained or increased its representation and it is not known whether this represents species survival from sclerophyll communities previously present or the evolution of new species in response to changed environmental conditions.
Visualising the landscape for this period in southeastern Australia is more difficult than for any other time during the Late Cainozoic. A mosaic of Casuarinaceae forest and more open vegetation with Asteraceae small trees and shrubs, and some eucalypts dominant above a heathy or grassy understorey may have existed with rainforest still prevalent in Tasmania and slightly more extensive and diverse than today in wetter parts of the mainland. The relative importance of these community types oscillated in relation to the global climate forcing. In addition, there is evidence of survival through much of the period, particularly from Pejark Marsh, of many more small patches of rainforest, particularly Araucariaceae, presumably in more protected environments on higher nutrient status soils. The recent discovery of wollemi pine growing in the Blue Mountains of New South Wales, a taxon not recorded palynologically since the Pliocene period, provides a demonstration of the ability of rainforest to survive in small patches through long periods of geological time (Jones et al., 1995).
The flora of New Zealand and southeastern Australia, particularly Tasmania, was more similar than today during the Early Pleistocene. Although there was some differential extinction at Plio- Pleistocene boundary, for instance Microcachrys in New Zealand and Dacrydium cupressinum type in Australia which could have been stochastic or a result of different levels of tectonic activity (Kershaw & McGlone, 1995), New Zealand had a mix of sclerophyll taxa including Acacia, Casuarinaceaepd probably Eucalyptus with the potential of developing a landscape similar to that of Australia (Kershaw, 1988).
In northeastern Australia, and probably along most of the eastern seaboard, the vegetation continued to be dominated by drier araucarian forest with wetter rainforest patches in coastal and montane areas probably little more extensive than at the time of arrival of European people (Kershaw et al., 1998). Itmight be inferred that the gradient of rainforest-related vegetation extended well inland with extensive areas of deciduous and semi-evergreen vine thickets, sensu Webb (1968), replacing araucarian forest, in semiarid environments with rainfall as low as about 600 mm per annum.
AAP Memoir 23 (2000)
There is virtually no evidence of conditions within the more arid centre of Australia, but a low resolution marine record from the similarly arid northwestern margin indicates a dominance of open vegetation including hummock grassland with an Acacia canopy and savanna grassland, probably with a Casuarinaceae dominated canopy (McMinn & Martin, 1992). Significant values of Gyrostemonaceae could suggest also a regional presence of rainforest-related thicket vegetation. These communities had been developing from the Late Miocene, presumably under increasing aridity, but there is no evidence of eucalypt woodlands .
Overall, the vegetation component of the landscape appears to have been relatively stable through this long Early Pleistocene period, allowing the evolution of a characteristic Quaternary fauna. Grasslands were extensive in northwestern Australia and possibly also in the centre of the Australian continent providing suitable environments for grazers. But with more extensive drier rainforest and thicket and Casuarinaceae woodlands and forests, and perhaps also woody Asteraceae communities, browsers would have been at an advantage over much of the continent.
Early-Middle Pleistocene transition
After a long period of relative stability, there is evidence of change around the Early-Middle Pleistocene boundary although the boundary itself is not clearly indicated in any available sequences. There are a group of extinctions, mainly bivalves , in the Wanganui sequence around 1.3 Ma, suggesting a further decrease in sea surface temperature. This coincides, approximately, with the beginning of the gradual temperature decline in the marine isotope record (Fig. 5) which continues to at least the Early-Middle Pleistocene boundary. Other extinctions and also first appearances of taxa, both nannnofossils and bivalves , occur through this period in the Wanganui sequence. A reduction in sea surface temperatures around the boundary is also suggested from northwestern Australia (Quilty, 1974; Zobel, 1984), but the evidence from northeastern Australia is different and somewhat contradictory with Chaproniere (1991) and Chaproniere & Pigram (1993) suggesting little change in planktonic foraminifera and no evidence of cooling (as is the case for the whole of the Quaternary), and oxygen isotope records indicating increasing sea surface temperatures (lsem et al., 1996; Peerdeman, 1997).
By contrast to the marine records, terrestrial palaeobotanical changes appear to be more closely related to the Early-Middle Pleistocene
499
boundary, although the degree of reliability is lower because of the discontinuous nature of the records . In New Zealand there are both last and first appearances, but little env ironmental information can be gleaned from them . In Tasmania, a range of taxa disappear although the list is augmented by species extinctions determined from macrofossils which are invisible in the pollen record. This input of macrofossil data is important in demonstrating that the plant landscape has probably been more dynamic than previously appreciated from a consideration of dominant pollen data. The mix of rainforest and sclerophyll taxa makes it difficult to determine the environmental causes of extinction, though it is probable from the loss of taxa such as Dacry carpus and Symplocos that a further increase in climatic variability was important and, from the latter taxon, that temperatures may also have decreased.
The generalised pollen record from ODP 820 (Kershaw et al., 1993) suggested that marked changes in components of north Queensland vegetation may have occurred around the Early- Middle Pleistocene transition ,perhaps relatable to the evident transition in the local and global isotope records from obliquity to eccentricity cycle dominance. Detailed examination of the pollen record between about 1 and 0.9 Ma (Russell Wild, pers. comm.) has confirmed the suggested increase in importance of Rhizophoraceae relative to Camptostemon, perhaps indicating a change from riverine to estuarine sources. One explanation is that the developmept of offshore reefs provided more sheltered estuarine environments for mangrove establishment and maintenance. However, there is no evidence for the existence of the Great Barrier Reef at this time (Feary et al. 1991). An alternative proposal is that estuarine mangrove development was facilitated by strong progradation at the end of glacial cycles as they increased in amplitude. The data certainly indicate a close relationship between mangrove peaks and 'glacial-interglacial' transitions as indicated in the isotope record from the core. The increase in amplitude of cycling may also have placed sufficient stress on the cool temperate conifers Dacrydium sp., Dacrycarpus and Dacrydium cupressinum to bring about their eventual demise but all are still hanging on to at least 900 ka.
Middle-Late Pleistocene
There is a substantial increase in the number of terrestrial palaeoecological records within this period although, only one, the discontinuous record from Lake George (Singh & Geissler, 1985), extends back to the Brunhes/Matuyama magnetic reversal. Other longer records include

500
Darwin Crater in Tasmania and Lake Terang in the western plains of Victoria which are both considered to extend to at least isotope stage 12. All records display marked fluctuations reflecting the high amplitude global climatic patterns. General correlation between marine and terrestrial records is possible and has been demonstrated for areas where comparisons between marine and terrestrial pollen records have been made, including northeastern and southeastern mainland Australia and New Zealand. Glacials were substantially drier, and in most cases demonstrably cooler than interglacials, with expansions of more open vegetation and reductions in lake levels. Geomorphic studies on riverine, lake and dune sediments from more arid parts of Australia (Kershaw & Nanson, 1993; Bowler et al., 1998; Magee et al., 1995; Wasson, 1989) and dust deposited in the Tasman Sea (Hesse, 1994) confirm that glacials were much drier than interglacials throughout the region. There is a suggestion also that precipitation levels were frequently higher within interglacials of the Middle-tate Pleistocene than had been achieved during much of the Early Pleistocene (Wagstaff et al., in press). In the western plains of Victoria, interglacial peaks in rainforest are generally more substantial than during the Early Pleistocene and suggest not only the attainment of higher rainfall but that the rainforest had the capacity to respond to climate change after existing for over a million years in small isolated patches. In the marine records, it is clear that glacial/interglacial alternations led to significant changes in oceanic conditions around Australia. Frontal zones moved north during times of expanded glaciation . The Leeuwin Current changed in importance while there were measurable changes in sea surface temperatures, current strength and upwelling .
Despite a general similarity between patterns of glacial and interglacial cycling within individual records, there are also notable differences that may have a variety of causes. The most obvious is the out-of-phase cyclicity of the orbital variables which produces global differences in seasonal and geographical solar insolation and hence precipitation. Superimposed on this variation is a proposed, although not fully explained, pattern of supercycles (Kukla & Cilek, 1996) whereby some glacials and succeeding interglacials appear climatically more extreme than others, at least in many terrestrial records. Supercycling has been used to explain substantial differences in vegetation response to individual climatic cycles in Lake Terang (Urban et al., 1996), and suggested for variation in Perched Lake (Longmore & Heijnis, 1998).A combination of glacial and supercycling may also explain
AAP Memoir 23 (2000)
features like the lack of response of Casuarinaceae to conditions during the penultimate interglacial when it is an important contributor to the vegetation of interglacial stages 7 and 1 at Lake Wangoom (Harle et al., 1999). Marked environmental differences in other records between the last and present interglacial are noted in other records, including the inland lakes of Lake Eyre (Croke et al., 1999) and Lake Woods (Bowler et al., 1998) where lake levels have been markedly lower during the Holocene than in Stage 5, and pollen records like Egg Lagoon on King Island (D'Costa, 1997), and Lynch's Crater, northeast Queensland, as well as Lake Wangoom where rainforest was more prevalent during substage 5e.
However, cyclical variation can not readily be used as an explanation for the degree of sustained environmental change in the late Quaternary which includes regional and continental level plant extinctions and the almost total loss of terrestrial megafauna. The most substantial landscape change is recorded in northeastern Australia where previously dominant moist araucarian forest was progressively replaced at study sites by open eucalypt vegetation. Perhaps the earliest evidence is from Fraser Island in southeastern Queensland where most of the replacement process, accompanied by a substantial and sustained increase in charcoal, is suggested, from preliminary and perhaps debatable uranium/thorium dates, to have taken place from before 350 ka to around 200 ka (Longmore & Heijnis, 1998). The last significant representation ofAraucaria though did occur within the last interglacial period and, like other records mentioned, failed, along with other rainforest taxa, to respond during the Holocene .
The ODP 820 record from northeastern Queensland provides a more detailed and continuous record through the last 250 ka. Here, Araucaria shows a clear two-tiered decline around 130 ka and 38 ka, the latter decline also recorded in tll# adjacent terrestrial record of Lynch's Crater. At both these sites the declines are accompanied by peaks in both eucalypts and charcoal with a general increase in both components through the record. The sites also demonstrate a regional extinction of the conifer Dacrydium guillauminii, now present only in New Caledonia, around 20 ka, and when combined with the loss of Austromyrtus from the Lynch's Crater record during the last glacial period, probably indicates a substantial reduction in the occurrence of peatland forest vegetation.
The earliest change in the ODP 820 record is a sustained increase in Poaceae around 175 ka, indicating a major expansion of grassland prior to substantial increases in eucalypts and charcoal.
AAP Memoir 23 (2000)
Similar increases in Poaceae are recorded in other marine cores, to the north of Australia, but in these, Poaceae expands relative to Eucalyptus . In the Lombok Ridge record Poaceae increases around 190ka and, like ODP 820, the event is not clearly related to any charcoal increase. However, charcoal does increase generally throughout the record with a major sustained rise at some time during the highly condensed or discontinuous record in the core of the period representing isotope stage 3. In the Banda Sea record, the increase in grasses relative to eucalypts occurs about 37 ka and is accompanied by a major rise in charcoal particles and also by a sustained decline in Dipterocarpaceae, a major element oflowland rainforest in Indonesia as well as New Guinea.
In southeastern Australia, the most substantial change occurs in the Lake George record with the general replacement of Casuarinaceae by Eucalyptus as the canopy dominant of sclerophyll forest or woodland during interglacial periods and an: increase in sclerophyllous shrubs. The change is also marked by a sustained increase in charcoal and, if the proposed timescale is accepted, occurred around 130 ka, the beginning of the last interglacial maximum. Broader regional vegetation change, especially during the last glacial cycle is suggested by a number oflast taxon occurrences. These include the temperate rainforest taxa Quintinia and perhaps Cunoniaceae (tricolpate) in Tasmania and the southern rainforest conifer Phyllocladus as well as the ancient herbaceous taxon Gunnera on the mainland both of which are now confined to Tasmania. These losses represent the last phase oflatitudinal sifting of once more homogeneous Australian rainforest although temperature is unlikely to have been the major cause. The last records of Phyllocladus and Gunnera are both from the Otway Ranges (McKenzie & Kershaw, 2000) and provide support for the suggestion that this area is biogeographically more similar to Tasmania than it is any other part of the Australian mainland. Data from the Otways tentatively suggest that Dacrycarpus may have survived here until the Late Pleistocene (Merna McKenzie, pers. comm.) but records of Nothofagus subsection Brassospora in the Late Pleistocene could well be the result of contamination from older sediments.
In contrast to Australia, there is little evidence for sustained vegetation change within New Zealand. Acacia, which appears to have been struggling since the Middle-Late Pleistocene boundary and eventually capitulates within the last glacial cycle, is the only recorded extinction. This is in contrast to the number of extinctions evident in the New Zealand marine record over the last 500 ka. However, the data sets are not easily
501
compared as the marine evidence is at species level and that of pollen at genus level. As indicated from Tasmanian macrofossil data, there may have been significant plant extinction at species level.
Since the publication of Singh et al. (1981), which incorporated the first long charcoal records from Australasian Quaternary sediments -from Lynch's Crater and Lake George -there has been a tendency for environmentalists to accept the proposition that vegetation change in association with increased burning of the landscape was caused by Aboriginal activities and hence the Australian landscape is a cultural phenomenon (Flannery, 1994). Concern expressed about the dating of the major vegetation change at Lake George around 130 ka (e.g. Wright, 1986) was lessened by a similar age assigned to the increase in burning and sustained vegetation change in the initial ODP 820 record. Archaeologists have rejected or remain very sceptical about an inference for the presence of people some 70 to 90 ka before any artefactual evidence or have even rejected the suggestion that People altered pre-existing burning
patterns (Horton, 1982). Any alternative explanation for sustained
vegetation change and increased burning had always faltered due to the lack of an obvious mechanism. The suggestion that recorded events were simply a continuation of a process of change to a more sclerophyllous vegetation, initiated in the central part of the continent by the drying trend in the late Tertiary (Kershaw, 1994), is shown here not to be viable. Most of the Quaternary experienced relatively stable conditions, despite the presence of fire-promoting eucalypt and grass vegetation with fire as a well-established environmental component. However, there is evidence from marine cores taken off the northern part of the continent of a regional increase in sea surface temperatures within the Middle Pleistocene. As mentioned, oxygen isotope records from the Coral Sea region suggest a systematic increase of about 4°C between about 500 and 250 ka (Isern et al., 1996; Peerdeman, 1993), estimates supported by sedimentary evidence for the development of the Great Barrier Reef within the last 500 ka (Feary et al., 1991). Off northwestern Australia, a similar increase in temperature, about 380 ka, was determined from foraminiferal assemblages (Zobel, 1984), while expansions of the bivalve Anadara trapezia (Murray-Wallace et al., 2000) during interglacials within the last 400 to 250 ka may indicate a regionally broader temperature increase. Isern et al. (1996) suggested that the temperature increase may have related to the development of a Coral Sea Warm Pool, a possible extension of the West Pacific Warm Pool. The cause of this temperature

502
increase has not been determined but could well relate to changing land-sea configurations and accompanying volcanic activity within the maritime continent, as Australia continued its movement northwards.
An increase in sea surface temperatures may have resulted in regionally increased precipitation which may help explain higher representation of rainforest relative to the Early Pleistocene in some pollen records, but could hardly explain the degree of instability which led to the expansion of sclerophyll vegetation and demise of fire-sensitive communities and taxa. However, if the evidence does relate to the development of the West Pacific Warm Pool, this would provide the Pacific temperature gradient required for the operation of El Nino-Southern Oscillation variability and, despite increased precipitation, the drought conditions necessary for increased fire activity and sclerophyll expansion.
A further question that needs to be addressed in consideration of the climate model is why do landscape changes appear to fall into discrete time periods? The change around 130 ka is clearly related to a major glacial-interglacial transition, but other sustained changes around 170 and 40 ka show no such correspondence with climatic events. One suggestion is that ENSO has not operated constantly or that its intensity has varied through time. Evidence from the Holocene of an increase in variability within the last 5 ka or so, which has been attributed to the latest onset of ENSO (McGlone et al., 1992; Shulmeister & Lees, 1995) provides support for this view. Consequently, change would be concentrated within those periods when ENSO was most active. Under these circumstances, any impact of people might simply have been an acceleration of a transformation that was already in progress. Of the times of change identified, the one around 40 ka is the most dramatic where it is recorded, despite its lack of correlation with a noted time of climate change. Considering the weight of archaeological evidence for continental colonisation by this time, a partial or complete human cause might be invoked.
Of major significance to the development of the present day landscape is the extinction of most of Australia's megafauna. Their loss not only transformed faunal biogeography and ecology but would have substantially impacted other components of the system, particularly the vegetation. The most widely publicised extinction scenario is that of Flannery (1990, 1994) who considered that overkill was the cause and that it was the loss of grazing and browsing pressure on the vegetation and consequent build up of fuel that led to increased burning and the expansion of fire-
AAP Memoir 23 (2000)
promoting sclerophyll vegetation. The high percentage of species becoming extinct is consistent with proportions on other land masses including New Zealand and the Americas where overkill provides the most convincing hypothesis (Martin, 1984). The acceptance of the overkill model has generally been prohibited by an apparent substantial overlap between the time of arrival of people and dated megafaunal remains. However, the establishment of a firm date for the demise of one megafaunal component, Genyornis, around 50 ka in the centre of Australia (Miller et al., l 999) combined with substantially older ages from the redating of critical sites from more coastal areas such as Lancefield Swamp (van Huet et al., 1998), and Spring Creek (Nick Porch, pers. comm.) and continuing uncertainty over the age of the Cuddie Springs megafauna, significantly reduce potential overlap time. As Flannery (1999) states 'there is currently no unequivocal evidence for the survival of any Australian megafauna after 40,000 years ago'. The reduction in the number of 'young' sites also makes the Horton model, whereby extinctions were caused primarily by a lowering of rainfall during the Last Glacial Maximum dating from about 25 ka, unlikely, although any intensification of ENSO around 40 ka may have contributed to the extinction process.
Habitat change due to Aboriginal burning is also unlikely to have been the predominant cause of extinction, as it is evident that both vegetation change and increased burning have been longer term and continuing processes, unlikely to have resulted in critical thresholds for megafaunal survival at the same time throughout the continent. However, it is possible that at least some megafauna were effected by these processes and that the extinction phase may have been initiated before the arrival of people (or the last colonisation phases), or that many species were vulnerable due to pre-existing environmental stress. There is also the question of the actual reason for an acceleration itll"rates of burning resulting from colonisation. The argument of Flannery, that burning increased as a result of fuel accumulation rather than direct Aboriginal burning , is not supported by evidence from the Banda Sea pollen record (van de Kaars et al., 1998). Here there was a sustained decline in the dipterocarp dominants of Indonesian rainforest around 37 ka in association with an increase in burning without any clear indication of megafaunal extinction. It suggests that people, colonising both parts of Indonesia and Australia around this time, were active users of fire.
Interestingly, while the Aboriginal past in Australia is now seen to have been dynamic rather than static, including changes in intensities of
AAP Memoir 23 (2000)
occupation of both geographical regions and individual sites, land use, artefact design, foraging strategies, diet breadths, resource extraction strategies, territorial networks, and the emergence of new technologies enabling the detoxification of poisonous plant foods (e.g. Cycas media) , the major periods of cultural change do not appear to have necessarily occurred in tandem with environmental or climatic alterations . Rather than occurring during the Last Glacial Maximum or during the early Holocene when climates were at their most extreme, by far the greatest degree of cultural change took place during the last Ska, and especially during the last 3 ka (although the influence of variable ENSO activity cannot be totally discounted). Partly because of this, most archaeologists today tend to view these changes as resulting from unfolding social processes rather than from external prime movers such as environmental or climatic changes. In this sense, the emergence of Homo sapiens during the Quaternary further challenges us to enquire into both the common links between, and the distinctive causes of, the various changes that have taken place during this, the most recent phase of earth's history. ·
CONCLUSIONS
Evidence for changing biogeographical patterns and their environmental causes during the Quaternary is provided by a large and increasing number of fossil groups and types. The most substantial information is available from marine and adjacent coastal environments while arid and semi-arid environments are poorly covered.
High temporal resolution and good spatial distribution of data for the last 20 ka has allowed good comparison between patterns of climate and biota during the Last Glacial Maximum and mid Holocene as largely representative of the extremes of glacial-interglacial cyclicity. Under regionally slightly wetter, warmer and less variable climates, forest ecosystems in the mid Holocene were slightly more extensive and, in Australia and New Guinea, the impact of people was probably significantly lower than today. During the Last Glacial Maximum, temperatures were probably at least 4°C and up to 8-9°C lower than today although substatial temperature lowering in the lowland tropics is debated . These lower temperatures combined with more regional influences, particularly substantial expansions in continental shelves, resulted in precipitation levels up to 50% lower than present. Forest systems were reduced to small isolated patches throughout most of the region although there is little evidence of regional migration. Only in the high latitudes of eastern Australia and New Zealand is there
503
evidence of the formation of community types that are not represented today, and these fall within the general category of grassland/steppe. The degree to which these two periods can be considered representative of extremes of glacial cycles over a longer period may be questioned in Australia and New Guinea, because of the presence of people and lack of megafauna. However, similarities in the nature and degree of change to New Zealand, that had yet to experience human settlement and megafaunal extinction, suggest that these influences on other components of the biota were not substantial in comparison with the impact of climate change.
Three areas, southeastern Australia, northeastern Australia and New Zealand (primarily the Wanganui Basin), contain the most substantial records of change through the Quaternary in general, and comparisons between them are made, largely on the pollen data. Records are generally oflow resolution and little information is provided on cyclicity, at least prior to the last 1 Ma. New Zealand and southeastern Australia show significant extinction of previously important vegetation components around the proposed Plio- Pleistocene boundary, some.2.5 Ma, reflecting the importance of global cooling perhaps augmented by the initiatiation of a winter rainfall regime at these latitudes. By contrast, northeastern Australia shows no clear response to the onset of the Quaternary, although a continuation of the late Cainozoic global drying trend is evident. Further floral extinctions and some sustained changes in taxon abundance are recorded around the Early- Middle Pleistocene transition ,probably in response to heightened precipitation and temperature contrasts between glacial and interglacial periods. Incontrast to New Zealand, and most of the globe, northeastern and southeastern Australia show marked trends towards more open vegetation with extinctions in some wetter elements and increased burning within the last few hundred thousand years . There is also evidence of an increase in sea surface temperatures off northeastern Queensland between 500 and 250 ka. It is hypothesised that this temperature increase is related to the development of the West Pacific Warm Pool and associated ENSO activity that created vegetation instability and its vulnerability to fire. It is likely that Aboriginal burning accelerated Late Quaternary vegetation changes but did not initiate them.
In relation to megafauna, this environmental reconstruction for Australia makes it unlikely that either climate or habitat change was the primary cause of Late Pleistocene extinction. Consequently, we consider that the most likely explanation is direct killing by people, a

504
conclusion supported by the evidence for the demise of megafauna in the late Holocene of New Zealand.
ACKNOWLEDGEMENTS
We thank Patrick De Deckker, Matt McGlone and Colin Murray-Wallace for their incisive comments and attempts to incorporate some consistency and balance into this review, Mike Macphail, Dallas Mildenhall and Alan Partridge, for valuable comments on Quaternary pollen stratigraphy, Nick Porch and Michael Reid for input on specific fossil groups, John Tibby for help with referencing and Gary Swinton for drafting the text figures. We apologise for the scant attention paid to some groups of biota. Inthe case of molluscs and corals, we thank Alan Beu and Charlie Veron respectively for their very valuable input and appreciate that heavy commitments prevented them making the contributions to this paper that they would have liked.
REFERENCES Aguirre, E. & Pasini, G., 1985. The Pliocene-Pleistocene
boundary. Episodes 8, 116-120. Aharon, P., 1983. 140,000-yr isotope climatic record
from raized coral reefs in New Guinea. Nature 304, 720-723.
Aharon, P. & Chappell, J., 1986. Oxygen isotopes, sea level changes and the temperature history of a coral reef environment in New Guinea over the last 105
years. Palaeogeography, Palaeoclimatology, Palaeoecology 56, 337-379.
Albani, A., 1979. Recent shallow water Foraminiferida from New South Wales. Australian Marine Sciences Association Handbook 3, 51 pp.
Allen, J., 1989. When did humans first colonise Australia? Search 20, 149-154.
Allen, J., 1996. Warreen Cave. 135-167 in Allen, J. (ed.), Report of the Southern Forests Archaeological Project 1: Site Descriptions, Stratigraphies and Chronologies. School of Archaeology, La Trobe University, Bundoora.
Allen, J. & Holdaway, S., 1995. The contamination of Pleistocene radiocarbon determinations in Australia. Antiquity 69, 101-112.
Almond, D., McGowran, B. & Li, Q.Y., 1993. Late Quaternary foraminiferal record from the Great Australian Bight and its environmental significance. Memoirs of the Association of Australasian Palaeontologists 15, 417-428.
Anderson, A.J., 1989. Prodigious Birds: Maas and Moa Hunting in Prehistoric New Zealand. Cambridge University Press, Cambridge . 238p.
Anderson, A.J., 1996. Was Rattus exulans in New Zealand 2000 years ago? AMS radiocarbon ages from Shag River Mouth. Archaeology in Oceania 31, 178-184.
AAP Memoir 23 (2000)
Anderson, A. & McGlone, M., 1992. Living on the edge - prehistoric land and people in New Zealand. 199- 241 in Dodson, J. (ed.) The Naive Lands: Prehistory and Environmental Change in Australia and the Southwest Pacific. Longman, Cheshire.
Apthorpe, M., 1988. Cainozoic depositional history of the North West Shelf. 55-84 in Purcell, P.G. & Purcell, R.R. (eds.) TheNorth West Shelf Australia. Proceedings of the Petroleum Exploration Society of Australia Symposium, Perth.
Arrhenius, G.O., 1952. Sediment cores from the east Pacific. Report of the Swedish Deep Sea Expedition, 1947-1948, 5, 1-228.
Ashworth, A.C., Markgraf, V. & Villagran, C., 1991. Late Quaternary climatic history of the Chilean Channels based on pollen and beetle analyses, with an analysis of the modem vegetation and pollen rain. Journal of Quaternary Science 6, 279-291.
Augustinus, P.C . & Macphail, M.K., 1997. Early Pleistocene stratigraphy and timing of the Bulgobac Glaciation, western Tasmania, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 128, 253-267.
Ayliffe, L.K., Moriarty, K.C., Marianelli, P.C & Wells, R.T., 1997. TIMS U/Th dating of secondary carbonates: theory and application to a Mid- Pleistocene fossil deposit. 11 in Programme and Abstracts: Conference on Australian Vertebrate Evolution, Palaeontology and Systematics, July, 1997, Western Australian Museum, Perth.
Baird, R.F., 1989. Fossil bird assemblages from Australian caves: precise indicators of late Quaternary environments. Palaeogeography, Palaeoclimatology, Palaeoecology 69, 241-244.
Baird, R.F., 1991.Avian fossils from the Quaternary of Australia. 809-870 in Vickers-Rich, P., Monaghan, J.M., Baird, R.F. & Rich, T.H. (eds.), Vertebrate Palaeontology of Australasia. Pioneer Design Studio, Lilydale.
Balme, J. & Hope, J., 1990. Radiocarbon dates from middens in the Darling River area of western New South Wales. Archaeology in Oceania 25, 85-101.
Barker, B. 19. The Sea People : maritime hunter- gatherers on the tropical coast. A late Holocene maritime specialisation in the Whitsunday Islands, central Queensland. Unpublished PhD thesis, University of Queensland, St Lucia.
Barrows, T.T., 1995. Three aspects of late Quaternary palaeoclimatic reconstruction in eastern Australia. Unpublished B.Sc.(Hons) thesis, Australian National University, Canberra.
Barrows, T.T., Ayress, M.A. & Hunt, G.R., 1996. A reconstruction of the Last Glacial Maximum sea- surface temperatures in the Australasian region. Quaternary Australasia 1411, 27-31.
Beck, W., Clarke, A. & Head, L. (eds.), 1989. Plants inAustralian Archaeology. Tempus 1. Anthropology Museum, University of Queensland, Brisbane. 213p.
AAP Memoir 23 (2000)
Bednarik, R., 1997. The origins of navigation and
language. The Artefact 20, 16-56. Bennett, K.D., 1997. Evolution and the Ecology: The
Pace of Life. Cambridge University Press, Cambridge, 241p .
Berger, A. & Loutre, M.F., 1991. Insolation values for the climate of the last 10 m.y. Quaternary Science Reviews JO, 297-317.
Berggren, W.A., Hilgen, F.J., Langereis, C.G., Kent, D.V., Obradovich, J.D., Raffi, I., Raymo, M.E. & Shackleton, N.J., 1995. Late Neogene chronology: new perspectives in high-resolution stratigraphy. Geological Society of America Bulletin 107, 1272- 1287.
Betancourt, J., Van Devender, T. & Martin, P., 1990. Packrat Middens: The Last 40,000 years of Biotic Change. The University of Arizona Press, Tucson . 419p .
Beu, A.G., 1995. Pliocene limestones and their scallops: Lithostratigraphy, pectinid biostratigraphy and palaeogeography of eastern North Island Late Neogene limestones. Institute of Geological and Nuclear Sciences Monograph 10, 243p.
Beu, A.G. & Maxwell, P.A., 1990. Cenozoic Mollusca of New Zealand . New Zealand Geological Survey Palaeontological Bulletin 58, 5l8p.
Beu, A.G., Edwards, A.R. & Pillans, B.J., 1987. A review ofNew Zealand Pleistocene stratigraphy, with emphasis on the marine rocks. 250-269 in ltiliara, M. & Kamei, T. (eds.) Proceedings of the First International Colloquium on Quaternary Stratigraphy of Asia and the Pacific Area , Osaka, Science Council of Japan, Osaka.
Bird, M.I. & Grocke, D.R., 1997. Determination of the abundance and carbon isotope composition of elemental carbon in sediments . Geochimica et Cosmochimica Acta 61, 3413-3423.
Birdsell, J. B. 1957. Some population problems involving Pleistocene Man. Cold Spring Harbor Symposia on Quantitative Biology 22, 41-69.
Birdsell, J. B. 1977. The recalibration of a paradigm for the first peopling of Australia.113-167 in Allen, J., Golson, J. & Jones, R. (eds.), Sunda and Sahu!. Academic Press, London .
Blom, W.M., 1988. Late Quaternary sediments and sea-levels in Bass Basin, southeastern Australia - a preliminary report. Search 19, 94-96.
Bohte, A. & Kershaw, A.P., 1999. Taphonomic influences on the interpretation of the palaeoecological record from Lynch's Crater, northeastern Australia. Quaternary International 57158, 49-59.
Bowdery, D., 1996. Phytolith Analysis Applied to Archaeological Sites in the Australian Arid Zone. Unpublished Ph.D. thesis, Australian National University, Canberra .
Bowdler, S., 1977. The coastal colonization of Australia. 205-246 in Allen, J., Golson, J. & Jones,
505
R. (eds.), Sunda and Sahu!, Academic Press , London.
Bowler, J .M., 1982. Aridity in the Tertiary and Quaternary of Australia. 35-45 in Barker, W.R. & Greenslade, P.J .M. (eds), Evolution of the Flora and Fauna of Arid Australia. Peacock Publications, Frewville.
Bowler, J.M., 1986. Spatial variability and hydrologic evolution of Australian lake basins: analogue for Pleistocene hydrologic change and evaporite formation. Palaeogeography, Palaeoclimatology, Palaeoecology 54, 21-41.
Bowler, J.M., Duller, G.A.T., Perret, N., Prescott, J.R. & Wyrwoll, K-H., 1998. Hydrologic changes in monsoonal climates of the last glacial cycle: stratigraphy and luminescence dating of Lake Woods, N.T., Australia. Palaeoclimates 3, 179-207.
Bowler, J., Jones, R., Allen, H. & Thome, A., 1970. Pleistocene human remains from Australia: a living site and cremation from Lake Mungo, western New South Wales. World Archaeology 2, 39-60.
Bowler, J. & Thome, A., 1976. Human remains from Lake Mungo: discovery and excavation of Lake Mungo 3. 127-138 in Kirk, R.L. & Thome, A. (eds.), The Origin of the Australians. Australian Institute of Aboriginal Studies, Canberra.
Boyd, W.E., 1992. A Pollen Flora of the Native Plants of South Australia and Southern Northern Territory, Australia. South Australian Geographical Papers 3, 212 pp.
Bussell, M.R., 1986. Palynological evidence for Upper Putikian (middle Pleistocene) interglacial and glacial climates at Rangitawa Stream, South Wanganui Basin, New Zealand. New Zealand Journal of Geology and Geophysics 29, 471-479,
Bussell, M.R., 1988. Mid and late Holocene pollen diagrams and Polynesian deforestation, Wanganui district, New Zealand. New Zealand Journal of Botany 26, 431-451. .
Bussell, M.R., 1990. Palynology of oxygen isotope 6 and substage Se from the cover beds of a marine terrace, Taranaki, New Zealand. Quaternary Research 34, 86-100.
Bussell, M.R. & Pillans, B., 1992. Vegetational and climatic history during oxygen isotope stage 9, Wanganui district, New Zealand, and correlation of the Fordell Ash. Journal of the Royal Society of New Zealand 22, 41-60.
Bussell, M.R . & Mildenhall, D.C., 1990. Extinct palynomorphs from middle and late Pleitocene terrestrial sediments, South Wanganui Basin, New Zealand. New Zealand Journal of Geology and Geophysics 33, 439-448.
Chappell, J.,1983. A revised sea-level record for the last 300,000 years from Papua New Guinea. Search 14, 99-101.
Chappell, J., 1994. Upper Quaternary Sea Levels, Coral Terraces, Oxygen Isotopes and Deep-sea

506
Temperatures, Journal of Geography (Japan) 103(7), 828-840.
Chappell, J., Head, J. & Magee, J., 1996. Beyond the radiocarbon limit in Australian archaeology and Quaternary research. Antiquity 70, 543-552.
Chappell, J. & Shackleton, N.J., 1986. Oxygen isotopes and sea level. Nature 324, 137-140.
Chaproniere, G.C.H., 1991. Pleistocene to Holocene planktic foraminiferal biostratigraphy of the Coral Sea, offshore Queensland, Australia. BMR Journal ofAustralian Geology and Geophysics I2, 195-221.
Chaproniere, G.C.H. & Pigram, C.J., 1993. Miocene to Pleistocene foraminiferal biostratigraphy of dredge samples from the Marion Plateau, offshore Queensland, Australia. A GSOJournal ofAustralian Geology and Geophysics I 4, 1-19.
Chivas, AR., De Deckker, P., & Shelley, J.M.G., 1986. Magnesium content ofnon-marine ostracod shells: a new palaeosalinometer and palaeothermometer. Palaeogeography, Palaeoclimatology, Palaeoecology 54, 43-61.
Chivas, AR., De Deckker, P., Cali, J.A., Chapman, A., Kiss, E. & Shelley, J.M.G., 1993. Coupled stable- isotope and trace-element measurements of lacustrine carbonates as.palaeoclimatic indicators. 113-121 in Swart, P.K., Lohmann, K.C., McKenzie, J. & Savin, S. (eds.) Climate Change in Continental Isotope Records, American Geophysical Union, Geophysical Monograph 78.
Clark, R.L., 1982. Point count estimation of charcoal in pollen preparations and thin sections of sediments . Pollen et Spores 24, 523-535.
Clark, R.L., 1983. Pollen and charcoal evidence for the effects of Aboriginal burning on the vegetation of Australia. Archaeology in Oceania I 8, 32-37.
Clark, R.L., 1984. Effects on charcoal of pollen preparation procedures. Pollen et Spores 26, 559- 576.
Colhoun, E.A., 1988. Cainozoic Vegetation of Tasmania. Special Paper, Department of Geography, University of Newcastle.
Colhoun, E.A., Pola, J.S., Barton, C.E. & Heijnis, H., 1999. Late Pleistocene vegetation and climate history of Lake Selina, western Tasmania. Quaternary International 57158, 5-23.
Colhoun, E.A., van de Geer, G., & Mook, W., 1982. Stratigraphy, pollen analysis and palaeoclimatic interpretation of Pulbeena Swamp, northwestern Tasmania. Quaternary Research I 8, 108-126.
Collins, AC., 1974. Port Phillip Survey 1957-63 Forarniniferida . Memoirs of the National Museum of Victoria 35, 1-62.
Collins, L.B., Wyrwoll, K-H & France, R.E., 1991. The Abrolhos carbonate platforms: geological evolution and Leeuwin Current activity. Journal of the Royal Society of Western Australia 74,47-57.
AAP Memoir 23 (2000)
Connell, R.D. & Sikes, E.L., 1997. Controls on Late Quaternary sedimentation of the South Tasman Rise. Australian Journal of Earth Sciences 44, 667-675.
Coope, G.R., 1986, Coleopteran analysis. 703-711 in Berglund, B.E. (ed.) Handbook of Holocene Palaeoecology and Palaeohydrology, John Wiley & Sons, Chichester.
Cosgrove, R., 1995. Late Pleistocene behavioural variation and time trends: the case from Tasmania. Archaeology in Oceania 30(3), 83-104.
Croke, J.C., Magee, J.W. & Wallensky, E.P., 1999. The role of the Australian monsoon I the western catchment of Lake Eyre, central Australia, during the last interglacial. Quaternary International 571 58, 71-80.
David, B., 1993. Nurrabullgin Cave: preliminary results from a pre-37,000 year old rockshelter, north Queensland. Archaeology in Oceania 28(1), 50-54.
David, B. & Lourandos, L., 1998. Rock art and socio- demography in northeastern Australian prehistory. World Archaeology 30(2), 193-219.
David, B., Roberts, R., Tuniz, C., Jones, R. & Head, J., 1997. New optical and radiocarbon dates from Ngarrabullgan cave, a Pleistocene archaeological site in Australia: implications for the comparability of time clocks and for the human colonisation of Australia. Antiquity 7I , 183-188.
Davidson, I. & Noble, W., 1992. Why the first colonisation of the Australian region is the earliest evidence of modem human behaviour. Archaeology in Oceania 27, 135-142.
Davies, P.J., McKenzie, J.A., Palmer-Julson, A. et al. 1991. Expanatory notes. Initial Reports , Ocean Drilling Program I33, 31-58.
D'Costa, D.M., 1989.Late Quaternary vegetation and environments from Lake Terang, western Victoria. Unpublished M.Sc. thesis, Monash Univeristy, Melbourne.
D'Costa, D.M., 1997. The reconstruction of Quaternary vegetation and climate on King Island, Bass Strait, Australia. Unpublished PhD thesis, Monash University, Melbourne.
D'Costa, D.M., Edney, P., Kershaw, AP.& De Deckker, P., 1989. La Quaternary palaeoecology of Tower Hill, Victoria. Journal of Biogeography I 6,461-482.
D'Costa, D.M., Grindrod, J. & Ogden, R., 1993. Preliminary environmental reconstructions from late Quaternary pollen and mollusc assemblages at Egg Lagoon, King Island, Bass Strait. Australian Journal
of Ecology 18, 351-366. D'Costa, D.M. & Kershaw, A.P., 1995. A Late
Pleistocene and Holocene pollen record from Lake Terang, Western Plains of Victoria, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 113, 57-67.
De Deckker, P., l 982a, Australian aquatic habitats and biota: their suitability for palaeolimnological investigations. Transactions of the Royal Society of South Australia I 06, 145-153.
AAP Memoir 23 (2000)
De Deckker, P.,-1982b, Holocene ostracods, other invertebrates and fish remains from cores of four maar lakes in southeastern Australia. Proceedings of the Royal Society of Victoria 94, 183-220.
De Deckker, P., l982c, Late Quaternary ostracods from Lake George, New South Wales .Alcheringa 6,305- 318.
De Deckker, P., 1982d, Non-marine ostracods from two Quaternary profiles at Pulbeena and Mowbray Swamps, Tasmania. Alcheringa 6, 249-274 .
De Deckker, P., Chivas, A.R., Shelley, M.G. & Torgersen, T., 1988. Ostracod shell chemistry: a new palaeoenvironmental indicator applied to a regressive/transgressive record from the Gulf of Carpentaria, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 66, 231-241.
Dodson, J.R., 1975. Vegetation history and water fluctuations at Lake Leake, southeastern Australia II. 50,000 B.P. to 10,000 B.P.Australian Journal of Botany 23, 815-831.
Dodson, J.R.(ed.), 1992. The Naive Lands: Prehistory and Environmental Change in Australia and the Southwest Pacific. Longman, Cheshire. 258p.
Dodson, J., Fullagar, R., Furby, J., Jones, R. & Prosser, I., 1993. Humans and megafauna in a late Pleistocene environment from Cuddie Springs, north-western New South Wales. Archaeology in Oceania 28, 94-99.
Edney, P.A., Kershaw, AP., De Deckker, P., 1990. A late Pleistocene and Holocene vegetation and environmental record from Lake Wangoom, Western Plains of Victoria, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 80, 325-343.
Elias, S.A., 1994, Quaternary Insects and Their Environments, Smithsonian Institution Press, Washington. 284p.
Emiliani, C., 1955. Pleistocene temperatures. Journal of Geology 63, 539-578.
Emight, N.J. & Gosden, 1992. Unstable archipelagos - south-west Pacific environment and prehistory since 30,000 B.P. 160-198 in Dodson, J. (ed.), The Naive Lands :Prehistory and Environmental Change in Australia and the Southwest Pacific. Longman, Cheshire.
Feary, D.A., Davies, P.J., Pigram, C.J. & Symonds, P.A., 1991. Climatic evolution and control on carbonate deposition in northeast Australia. Palaeogeography, Palaeoclimatology, Palaeoecology (Global and Planetary Change Section) 89, 341-361.
Ferland, M.A., Roy, P.S. & Murray-Wallace, C.V., 1995. Glacial lowstand deposits on the outer continental shelf of southeastern Australia. Quaternary Research 44, 294-299.
Flannery, T.F., 1990. Pleistocene megafaunal loss: implications of the aftershock for Australia's past and future. Archaeology in Oceania 25, 45-67.
Flannery, T., 1994. The Future Eaters. Reed, Chatswood. 423p.
507
Flannery, T.F., 1999. Debating extinction. Science 283, 182-183.
Fleming, C.A., 1944. Molluscan evidence of Pliocene climatic change in New Zealand. Transactions of the Royal Society of New Zealand 74,207-220.
Fleming, C.A., 1953. The geology of the Wanganui Subdivision. New Zealand Geological Survey Bulletin 52, 1-100.
Fullagar, R., Price, D. & Head, L., 1996. Early human occupation of northern Australia: Jinmium rockshelter. Antiquity 70, 751-773.
Gell, P.A., 1997, The development ofa diatom database for inferring lake salinity, Western Victoria, Australia: towards a quantitative approach for reconstructing past climates. Australian Journal of Botany 45, 389-423.
Gell, P.A., 1998, Quantitative reconstructions of the Holocene palaeosalinity of pairedcrater lakes based on a diatom transfer function Palaeoclimates 3, 83-96,
Goodfriend, G.A., 1992, The use of land snails in palaeoenvironmental reconstruction. Quaternary Science Reviews I l , 665-685.
Green, N., Caldwell, J., Hope, J. & Luly, J, 1983. Pollen from an 1800year old stick-nest rat (Leporillus spp.) midden from Gnalta, western New South Wales. Quaternary Australasia 1, 31-41.
Guilderson, T.P., Fairbanks, R.G. & Rubenstone, J.L., 1994. Tropical temperature variations since 20,000 years ago: modulating interhemispheric climate change . Science 263, 663-665.
Harle, K.J., 1997. Late Quaternary vegetation and climate change in southeastern Australia: palynological evidence from marine core E55-6. Palaeogeography , Palaeoclimatology, Palaeoecology 131, 465-483.
Harle, K.J., Kershaw, A.P. & Heijnis, H. (1999) The contributions of marine palynology and uranium/ thorium to the dating of the Lake Wangoom pollen record, western plains of Victoria, Australia. Quaternary International 57158, 25-34.
Harrison, S.P. & Dodson, J., 1993. Climates of Australia and New Guinea since 18,000 yr BP. 265-293 in Wright, H.E., Kutzbach, J.E., Webb, T., Ruddiman, W.E.F., Sreet-Perrott, F.A. & Bartlein, P.J. (eds.) Global Climates Since the Last Glacial Maximum. University of Minnesota Press, Minneapolis.
Hart, D.M. & Humphreys, G.S., 1997. Plant opal phytoliths: an Australian perspective. Quaternary Australasia 15(1), 17-25.
Head, L., McCarthy, L., Quade, J., Witter, D., Allen, V. & Lawson, E. 1998. Classification and radiocarbon dating of Leporillus nests in semi-arid Australia, and palaeoclimatic implications. Palaeoclimates : Data and Modelling 3, 161-177.
Hesse, P.P., 1994. The record of continental dust from Australia in Tasman sea sediments. Quaternary Science Reviews 13, 257-272.
Reusser, L.E. & van de Geer, G., 1994. Direct correlation of terrestrial and marine palaeoclimatic

508
records from four glacial-interglacial cycles - DSDP Site 594 Southwest Pacific. Quaternary Science Reviews 13, 273-282.
Hill, R.S. & Macphail, M.K., 1985. A fossil flora from rafted Pio-Pleistocene mudstones at Regatta Point, Tasmama. Australian Journal of Botany 33, 497- 517.
Hiscock, P., 1986. Technological change in the Hunter River valley and the interpretation of late Holocene change in Australia .Archaeology in Oceania 21(1), 40-50.
Hiscock, P., 1988. Prehistoric settlement patterns and artefact manufacture at Lawn Hill, northwest Queensland. Unpublished PhD thesis, University of Queensland, St. Lucia. ·
Hodell, D.A, 1993. Late Pleistocene paleoceanography of the South Atlantic sector of the Southern Ocean· Ocean Drilling Program Hole 704A'. Paleoceanography 8,47-68.
Holdaway, R.N., 1996.Arrival ofrats in New Zealand. Nature 384, 225-226.
Holdaway, R.N. & Jacomb, C., 2000. Rapid extinction of the moas (Aves: Dinornithiformes): model, test and inplications. Science 287, 2250-2254.
Hope, G.S., 1994. Quaternary vegetation, 386-389 in Hill, R.S. (ed.), The History of the Australian Veetaton : Cretaceous to Recent . Cambridge Umvers1ty Press, Cambridge.
Hope, G.S. (ed.), 1998. IdentifYing Wood-Charcoal Remains as Palaeo Evidence for Regions of Central and Northeast Australia. Research Papers in Archaeology and Natural History No. 28, 209p.
Hope, J.H., Lambert,R.J., Edmondson, L., Smith, M.J. & Van Tets, G.F., 1978. Late Pleistocene remains from Seton rock shelter, Kangaroo Island, South Australia. Journal of Biogeography 4, 363-385.
Hopkins, M.S., Ash, J., Graham, A.W., Head, J. & Hewett, R.K., 1993. Charcoal evidence of the spatial extent of the Eucalyptus woodland expansions and rainforest contractions in North Queensland during the late Pleistocene . Journal of Biogeography 20, 357-372.
Hopkins, M.S., Head, J., Ash, J.E., Hewett, R.K. and Graham, A.W., 1996. Evidence of a Holocene and continuing recent expansion of lowland rainforest in humid, tropical north Queensland. Journal of Biogeography 23, 737-745.
Hornibrook , N de B., 1981. Globorotalia (planktic Foraminiferida) in the Late Pliocene and Early Pleistocene of New Zealand. New Zealand Journal of Geology and Geophysics 24, 263-292.
Hornibrook, N de B., 1982. Globorotalia (Foraminiferida) from DSDP Leg 29, Site 284, Southwest Pacific. New Zealand Journal of Geology and Geophysics 25, 83-99.
Horsfall, N. & Hall, J., 1990. People and the rainforest: an archaeological perspective . 33-39 in Webb, L.J. & Kikkawa, J. {eds.), Australian Tropical
AAP Memoir 23 (2000)
Rainforests : Science-Values-Meaning . CSIRO, Melbourne.
Horton, D.R., 1981. Water and woodland: the peopling of Australia . Australian Institute of Aboriginal Studies Newsletter I6, 21-27.
Horton, D.R., 1982. The burning question: Aborigines, fire and Australian ecosystems. Mankind 13, 237- 251.
Horton, D.R., 1984. Red Kangaroos: last of the Australian megafauna. 639-680 in Martin, P.S. & Klein, R.G. (eds.) Quaternary Extinctions: A Prehistoric Revolution University of Arizona Press, Tucson.
Howard, W.R. & Prell, W.L., 1994. Late Quaternary carbonate production and preservation in the Southern Ocean: Implications for oceanic and atmospheric carbon cycling. Paleoceanography 9, 453-482.
lmbrie, J., Hays, J.D., Martinson, D.G., Mcintyre, A., Mix, A.C., Morley, J.J., Pisias, N.G., Prell, W.L. & Shackleton, N.L., 1984. The orbital theory of Pleistocene climate : support from a revised chronology of the marine 180 record. 269-305 in Berger, A. et al. (eds.), Milankovich and Climate D. Reidel, Norwell. '
Isern, A.R., McKenzie, J.A. & Feary, D.A., 1996. The role of sea-surface temperature as a control on carbonate platform development in the western Coral Sea. Palaeogeography, Palaeoclimatology, Palaeoecology 124, 247-272.
Jones, W.G., Hill, K.D. & Allen, J.M., 1995. Wollemia nobilis, a new living Australian genus and species in the Araucariaceae . Telopea 6, 173-176.
Jordan, G.J., 1995a. Early-Middle Pleistocene leaves of extinct and extant Proteaceae from western Tasmania, Australia. Botanical Journal of the Linnean Society 118, 19-35.
Jordan, G.J., 1995b. Extinct conifers and conifer diversity in the Early Pleistocene of western Tasmania. Review of Palaeobotany and Palynology 84, 375-387.
Jordan, G.J., 19_.97. Contrasts between the climatic ranges of tclssil and extant taxa: causes and consequences for palaeoclimatic estimates . Australian Journal of Botany 45, 465-474.
Jordan, G.J., Macphail, M.K., Barnes, R. & Hill, R.S., 1995. An Early to Middle Pleistocene flora of subalpine affinities in lowland western Tasmania. Australian Journal of Botany 43, 231-242.
Kendrick, G .W., 1977. Middle Holocene marine molluscs from near Gildford, Western Australia and evidence of climate change. Journal of the Royal Society of Western Australia 59, 97-104.
Kendrick, G.W., Wyrwoll, K-H. & Szabo, B.J. 1991. Plio-Pleistocene coastal events and history along the western margin of Australia. Quaternary Science Reviews JO, 419-439.
AAP Memoir 23 (2000)
Kennett, J.P. & Srinivasan, M.S., 1985. Neogene Planktonic Foraminifera: A Phylogenetic Atlas. Hutchinson Ross Publishing Company, Stroudsburg. 263p.
Kershaw, A.P., 1986. Climate change and Aboriginal burning through fue last two glacial-interglacial cycles from northeastern Queensland. Nature 322, 47-49.
Kershaw, A.P., 1988. Australasia . 237-306 in Huntley, B. & Webb, T. (eds.) Vegetation History. Kluwer Academic Publishers, New York.
Kershaw, A.P., 1994. Pleistocene vegetation of the humid tropics of northeastern Queensland, Australia . Palaeogeography, Palaeoclimatology, Palaeoecology 109, 399-412.
Kershaw, AP., 1995. Environmental Change in Greater Australia. Antiquity 69, 656-675.
Kershaw, A.P., 1997. A bioclimatic analysis of Early to Middle Miocene brown coal floras, Latrobe Valley, southeastern Australia. Australian Journal of Botany 45, 373-387.
Kershaw, A.P., 1998. Estimates of regional climatic variation within southeastern mainland Australia since the Last Glacial Maximum from pollen data. Palaeoclimates: Data and Modelling 3, 107-134.
Kershaw, A.P. & Bulman, D., 1996. A preliminary application of the analogue approach to the interpretation oflate Quaternary pollen spectra from southeastern Australia. Quaternary International 33, 61-71.
Kershaw,A.P. & McGlone, M.S., 1995. The Quaternary history of the southern conifers. 30-63 in Enright, N. & Hill, R.S. (eds.) The Ecology of the Southern Conifers. Melbourne University Press.
Kershaw, A.P. & Nanson, G.C., 1993. The last full glacial cycle in the Australian region. Global and Planetary Change 7, 1-9.
Kershaw, A.P. & Nix , H.A., 1988. Quantitative palaeoclimatic estimates from pollen spectra using bioclimatic profiles of extant taxa. Journal of Biogeography 15, 589-602.
Kershaw, A.P., Bush, M.B., Hope, G.S., Weiss, K-F., Goldammer, J.C. & Sanford, R., 1997. The contributions of humans to past biomass burning in the tropics. 413-442 in Clark, J.S., Cachier, H., Goldammer, J.G. & Stocks, B. (eds.) Sediment Records of Biomass Burning and Global Change. Springer, Berlin.
Kershaw, A.P., D'Costa, D.M., McEwen Mason, J.R.C., Wagstaff, B.E., 1991. Palynological evidence for Quaternary vegetation and environments of mainland southeastern Australia. Quaternary Science Reviews JO, 391-404.
Kershaw, A.P., McKenzie, G.M., McMinn, A., 1993. A Quaternary vegetation history of northeastern Queensland from pollen analysis of ODP site 820. Proceedings of the Ocean Drilling Program, Scientific Results 133, 107-114.
509
Kershaw, A.P., Martin, H.A. & McEwen Mason, J.R.C. (1994) The Neogene: a period of transition. 299- 327 in Hill, R.S. (ed.), TheHistory of theAustralian Vegetation: Cretaceous to Recent . Cambridge University Press, Cambridge.
Kershaw, A,P, Moss, P.T. & Wild, R., inpress. Patterns and causes of vegetation change in the Australian wet tropics over the last 10million years, in Moritz, C. & Bermingham, E. (eds.) Tropical Rainforests: Past and Future. Chicago University Press.
Kroon, D., 1993. Some planktonic forarniniferal datum levels during the last 10.4Ma, Leg 133.Proceedings of the Ocean Drilling Program, Scientific Results 133, 75-92.
Kukla, G. & Cilek, V., 1996. Plio-Pleistocene megacycles : record of climate and tectonics. Palaeogeography, Palaeoclimatology, Palaeoecology 120, 171-194.
Kukla, G., An, Z.S., Melice, J.L., Gavin, J. & Xiao,J.L., 1990. Magnetic susceptibility record of Chinese loess . Transactions of the Royal Society of Edinburgh , Earth Sciences 81, 263-288.
Kurten, B., 1968: Pleistocene mammals of Europe. Weidenfeld and Nicholson, London. 3l7p.
Kurten, B. & Anderson, E., 1980.Pleistocene Mammals of North America. Columbia University Press, London. 442p .
Lamb, L., 1996. Investigating changing stone technologies, site use and occupational intensities at Fern Cave, north Queensland . Australian Archaeology 42, 1-7.
Laskar, J., 1990. The chaotic motion of the solar system: a numerical estimate of the size of the chaotic zones. Icarus 88, 266-291.
Li, Q., Quilty, P.G., Moss, G. & McGowran, B., 1997. Southern Australian endemic and semi-endemic foraminifera: a preliminary report. Journal of Micropalaeontology 15, 169-185.
Lloyd, P.J. & Kershaw, A.P., 1997, Late Quaternary vegetation and early Holocene quantitative climatic estimates from Morwell Swamp, Latrobe Valley, south-eastern Australia. Australian Journal of Botany 45, 549-563.
Longmore, M .E. & Heijnis, H., 1999. Aridity in Australia: Pleistocene records of palaeoecological change from the Perched Lake sediments of Fraser Island, Queensland, Australia. Quaternary International 57158, 35-47.
Lorius, C., Jouzel, J., Ritz, C., Merlivat, L., Barkov, N.I., Korotkevich, Y.S. & Kotlyakov, V.M., 1985.A 150 000-year climatic record from Antarctic ice. Nature 316, 591-596.
Lourandos,H., 1983. Intensification: a late Pleistocene- Holocene archaeological sequence from southwestern Victoria. Archaeology in Oceania 18, 81-94.
Lourandos, H. & David, B., in press. Long-term archaeological and environmental trends: a

510
comparison from late Pleistocene-Holocene Australia, in Kershaw, A.P., Tapper, N.J., David, B., Bishop, P.M. & Penny, D. (eds.), The Environmental and Cultural History and Dynamics of the Australian-Southeast Asian Region. Catena Verlag. Reiskirchen, Germany.
Lourens, L.J., Antonarakou, A., Hilgren, F.J.,Van Hoof, A.AM., Vergnaud-Grazzini, C. & Zachariasse, W.J., 1996. Evaluation of the Plio-Pleistocene astronomical timescale, Paleoceanography 11, 391- 413.
Ludbrook, N.H., 1984. Quaternary molluscs of South Australia, Handbook, Vol. 9. Department of Mines and Energy, South Australia, Adelaide, 327 pp.
Luly, J., 1993. Holocene palaeoenvironments near Lake Tyrrell, semi-arid northwestern Victoria, Australia. Journal of Biogeography 20, 587-598.
Luly, J., 1995. Holocene palaeoenvironments at Lake Tyrrell - response to Sluiter and Parsons. Journal of Biogeography 22, 153-156.
Macphail, M.K., 1997. Late Neogene climates in Australia: fossil pollen- and spore-based estimates in retrospect and prospect. Australian Journal of Botany 45, 425-464.
Macphail, M.K. & Martin, H.A., 1991. 'Spineless' Asteraceae, (episode 2). Palynological and Palaeobotanical Association of Australasia Newsletter 23, 1-2.
Macphail, M.K., Colhoun, E.A. & Fitzsimons, S.J., 1995. Key periods in the evolution of the Cenozoic vegetation and flora in western Tasmania: the Late Pliocene.Australian Journal of Botany 43, 505-526.
Macphail, M.K., Jordan, G.J. & Hill, R.S., 1993. Key periods in the evolution of the Cenozoic vegetation and flora in western Tasmania: the Early-Middle Pleistocene. Australian Journal of Botany 41, 673- 707.
McCarthy, L. Head, L. & Quade, J., 1996. Holocene palaeoecology of the northern Flinders Ranges, South Australia, based on stick-nest rat (Leporillus spp.) middens: a preliminary overview. Palaeogeography, Palaeoclimatology, Palaeoecology 123,. 205-218.
McConnell, K., 1997. Palaeoethnobotanical remains of Carpentaria s Gap Site 1, The Kimberleys, Western Australia. Unpublished M.Sc. Thesis, Australian National University, Canberra.
McConnell, K. & O'Connor, S., 1997. 40,000 year record of food plants in the southern Kimberley Ranges, Western Australia. Australian Archaeology 45, 20-31.
McGlone, M.S., 1988. New Zealand, 557-599 in Huntley, B. & Webb, T. (eds.) Vegetation History. Kluwer Academic Publishers, Dordrecht.
McGlone, M.S. & Topping, W.W., 1977. Aranuian (post-glacial) pollen diagrams from the Tongariro region, North Island, New Zealand. New Zealand Journal of Botany 15, 749-760.
AAP Memoir 23 (2000)
McGlone, M.S. & Topping, W.W., 1983. Late Quaternary vegetation , Tongariro region , central North Island, New Zealand. New Zealand Journal of Botany 21, 53-76.
McGlone, M.S., Salinger, M.J. & Moar, N.T., 1993. Palaevegetation studies of New Zealand's climate since the Last Glacial Maximum. 294-317 in Wright, H.E., Kutzbach, J.E., Webb, T., Ruddiman, W.E.F., Sreet-Perrott, F.A. & Bartlein, P.J. (eds.) Global Climates since the Last Glacial Maximum . University of Minnesota Press, Minneapolis.
McGlone, M.S.;Kershaw, A.P. & Markgraf, V., 1992. El Nino/Southern Oscillation climatic variability in Australasian and South American palaeo- environmental records, 435-362 in Diaz, H.F. & Markgraf, V. (eds.) El Nino; Historical and Palaeoclimatic Aspects of the Southern Oscillation. Cambridge University Press, Cambridge.
McGlone, M.S. & Wilmshurst, J.M., 1999.Dating initial Maori environmental impact in New Zealand. Quaternary International 59, 5-16.
McKenzie, G.M . & Busby, J.R., 1992. A quantitative estimate of Holocene climate using a bioclimatic profile of Nothofagus cunninghamii (Hook.) Oerst. Journal of Biogeography 19, 531-540.
McKenzie, G.M. & Kershaw, A.P., 1997. A vegetation history and quantitative estimate of Holocene climate from Chapple Vale, in the Otway Region of Victoria, Australia. Australian Journal of Botany 45, 565-581.
McKenzie, G.M. & Kershaw, A.P., 2000. The last glacial cycle from Wyelangta, the Otway region of Victoria, Australia. Palaeogeography ,
Palaeoclimatology, Palaeoecology 155, 177-193. McKenzie, J.A. & Davies, P.J., 1993. Cenozoic
evolution of carbonate platforms on the northeastern margin: synthesis of Leg 133 drilling Results . Proceeedings of the Ocean Drilling Program , Scientific Results 133, 763-770.
McKenzie, J.A., Davies, P.J. & Palmer-Julson, A. (eds),1993. Proceeedings of the Ocean Drilling Program, Scientific Results 133, 826 pp.
McMinn, A., 1987. A Late Pleistocene dinoflagellate assembl from Bulahdelah, N.S.W. Proceedings of the Linnean Society of New South Wales 109, 175- 181.
McMinn, A., 1989. Late Pleistocene dinoflagellates from Botany Bay, N.S.W. Micropaleontology 35, 1-9.
McMinn, A., 1990. Distribution of <linoflagellate cysts in estuarine deposits from the east coast of Australia. Review of Palynology and Palaeobotany 65, 305- 309.
McMinn, A., 1991a. Quaternary dinoflagellate cyst distribution in Lake Macquarie. Quarterly Notes of the New South Wales Geological Survey 85, 1-6.
McMinn, A., 199lb. Recent dinoflagellate cysts from estuaries on the central coast of New South Wales, Australia. Micropalaeontology 37, 269-287.
AAP Memoir 23 (2000)
McMinn, A., 1992a. A late Pleistocene dinoflagellate and pollen record from the Richmond River Valley, northern N.S.W.; implications for the rate of sea level rise and climatic change. Quaternary Research 38, 347-358.
McMinn, A., 1992b. Recent and late Quaternary dinoflagellate cyst distribution on the continental shelf and slope of southeastern Australia. Palynology 16, 13-24.
McMinn, A ., 1992c. Pliocene through Holocene <linoflagellate cyst biostratigraphy of the Gippsland Basin, Australia. 147-161 in Head, M.J. & Wrenn, J.H. (eds.), Neogene and Quaternary Dinojlagellate Cysts and Acritarchs . American Association of Stratigraphic Palynologists Foundation, Dallas.
McMinn, A., 1992d. Neogene dinoflagellate distribution in the eastern Indian Ocean from Leg 123, Site 765. Proceedings of the Ocean Drilling Program, Scientific Results 123, 429-442.
McMinn, A., 1994a. Quaternary dinoflagellate distribution at Site 820, Great Barrier Reef, Australia . Proceedings of the Ocean Drilling Program , Scientific Results 133, 93-96.
McMinn, A., 1994b. Neogene dinoflagellate cyst biostratigraphy from Sites 815 and 823, Leg 133: Proceedings of the Ocean Drilling Program, Scientific Results 133, 97-106.
McMinn, A. & Martin, H.A., 1992. Late Cainozoic pollen history from Site 765, Eastern Indian Ocean. Proceedings of the Ocean Drilling Program, Scientific Results 123, 421-427.
McMinn, A. & Sun, X., 1994. Recent dinoflagellates from the Chatham Rise, east of New Zealand; a taxonomic study. Palynology 18, 41-53.
McMinn, A. & Wells, P., 1997. Use of dinoflagellate cysts to determine changing Quaternary sea surface temperature in southern Australia. Marine Micropaleontology 29, 407-422.
McNiven, I., 1999. Fissioning and Regionalisation :The social dimensions of changes in Aboriginal use of the Great Sandy Region, South East Queensland. 157-168 in Hall, J. & McNiven, l.J . (eds.), Australian Coastal Archaeology. Research School of Pacific and Asian Studies, Australian National University, Canberra.
Magee, J.M., Bowler, J.M., Miller, G.H. & Williams, D.L.G., 1995. Stratigraphy, sedimentology, chronology and palaeohydrology of Quaternary lacustrine deposits at Madigan Gulf, Lake Eyre, South Australia. Palaeogeography, Palaeoclimatologj; Palaeoecology 113, 3-42.
Markgraf, V., Bradbury, J.P. & Busby, J.R., 1986. Palaeoclimates in southwestern Tasmania during the last 13,000 years. Palaios 1, 368-380.
Martin, H.A., 1973. Palynology of some Tertiary and Pleistocene deposits, Lachlan River valley, New South Wales. Australian Journal of Botany Supplement 6, 1-57.
511
Martin, H.A., 1991. Tertiary stratigraphic palynology and palaeoclimate of the inland river systems in new South Wales. Special Publication of the Geological Society of Australia 18, 181-194.
Martin, H.A. & McMinn, A., 1993. Palynology of sites 815 and 823, The Neogene vegetation history of coastal vegetation of northeast Australia. Proceedings of the Ocean Drilling Program, Scientific Results 133, 115-125.
Martin, P.S., 1984. Prehistoric overkill: the global model. 354-403 in Martin,P.S. & Klein, R.G. (eds.) Quaternary Extinctions: A Prehistoric Revolution University of Arizona Press, Tucson.
Martinez, R .J .I., 1997. Decreasing influence of Subantarctic Mode Water north of the Tasman Front over the last 150 kyr. Palaeogeography, Palaeoclimatology, Palaeoecology 131, 355-364.
Merrilees, D., 1984. Comings and goings of late Quaternary mammals in extreme southwestern Australia. 629-638 in Martin, P.S. & Klein, R.G. (eds.) Quaternary Extinctions : A Prehistoric Revolution University of Arizona Press, Tucson.
Mildenhall, D.C., 1978. Palynology of the Waipipian and Hautewan Stages (Pliocene and Pleistocene), Wanganui, New Zealand. New Zealand Journal of Geology and Geophysics 21, 775-777.
Mildenhall, D.C., 1980. New Zealand Late Cretaceous and Cenozoic plant biogeography: a contribution. Palaeogeography, Palaeoclimatology, Palaeoecology 31, 197-233.
Mildenhall, D.C., 1983. Pollen diagram of a Middle Pleistocene section near Whilby, Pauatahanui, Porirua, New Zealand. New Zealand Geological Survey Report PAL 68, 13p.
Mildenhall, D.C.; 1994. Palynostratigraphy and palaeoenvironments of Wellington, New Zealand, during the last 80 ka, based on palynology of drill holes. New Zealand Journal of Geology and Geophysics 37, 421-436.
Mildenhall, D.C., 1995. Pleistocene palynology of the Petone and Seaview drillholes, Petone, Lower Hutt Valley, North Island, New Zealand. Journal of the Royal Society of New Zealand 25, 207-262.
Mildenhall, D.C. & Suggate,R.P., 1981.Palynology and age of the Tadmor Group (Late Miocene-Pliocene) and Porika Formation (Early Pleistocene), South Island, New Zealand New Zealand Journal of Geology and Geophysics 24, 515 -528.
Millener, P.R., 1991. The Quaternary avifauna of New Zealand. 1317-1344 in Vickers-Rich, P., Monaghan, J.M., Baird, R.F. & Rich, T.H. (eds.) Vertebrate Palaeontology of Australasia. Pioneer Design Studio, Lilydale.
Miller, G.H., Magee, J.W., Johnson, B.J., Fogel, M.L., Spooner, N.A., McCulloch and Ayliffe, L.K., 1999. Pleistocene extinction of Genyornis newtoni: human impact on Australian megafauna. Science 283, 205- 208.

512
Moar, N .T., 1993. Pollen Grains of New Zealand Dicotyledonous Plants. Whenua press , Lincoln. 200p.
Moar, N.T. & Suggate, R.P., 1996. Vegetation history from the K.aihinu (Last) Interglacial to the present, west coast, south Island, New Zealand . Quaternary Science Reviews 15, 532-547.
Morwood, M.J., O'Sullivan, P.B., Aziz, F. & Raza, A..1998. Fission-track ages of stone tools and fossils on the east Indonesian island of Flores.Nature 392, 173-176.
Moss, P.T., 1999.Late Quaternary environm ents of the humid tropics of northeastern Au s tralia . Unpublished PhD Thesis, Monash University, Melbourne.
Moss, P.T. & Kershaw,A.P., 2000. The last glacial cycle from the humid tropics of northeastern Australia: comparison of a terrestrial and a marine record. Palaeogeography, Palaeoclimatology, Pala eoecology 155, 155-176.
Murray, P., 1984. Extinction downunder : a bestiary of extinct Australian Late Pleistocene monotremes and marsupials , 600-628 in Martin, P.S. & Klein, R.G. (eds .) Quaternary Extinctions . A Prehistoric Revolution. ·University Arizona Press, Tucson.
Murray-Wallace , C.V., 1995. Aminostratigraphy of Quaternary coastal sequences in southern Australia -an overview. Quaternary International 26, 69-86.
Murray-Wallace, C.V.& Belperio,A.P., 1991. The Last Interglacial shoreline in Australia - a review. Quaternary Science Reviews 10, 441-461.
Murray-Wallace, C.V., Beu, A.G., Kendrick, G.W., Brown, L.J., Belperio, A.P. & Sherwood, J.E., 2000. Palaeoclimatic implications of the occurrence of the arcoid bivalve Andara trapezia (Deshayes) in the Quaternary of Australasia. Quaternary Science Reviews 19, 559-590.
Murray-Wallace , C.V. & Kimber, R.W.L., 1989. Quaternary marine arninostratigraphy - Perth Basin, Western Australia. Australian Journal of Earth Sciences 36, 553-568.
Murray-Wallace, C.V., Kimber, R.W.L ., Gostin, V.A. & Belperio, A.P., 1988. Amino acid racemisation dating of the 'older Pleistocene marine beds', Redcliffe, Northern Spencer Gulf, South Australia. Transactions of the Royal Society of South Australia 112, 51-55.
Naish , T., Abbott , S.T., Alloway, B.V., Beu, A.G ., Carter, R .M ., Edwards , A.R ., Joumeaux, T.D., Kamp, P.J.J., Pillans, B.J, Saul, G. & Woolfe, K.J., 1998. Astronomical calibration of a southern hemisphere Plio-Pleistocene reference section, Wanganui Basin, New Zealand. Quaternary Science Reviews 17, 695-710.
Naish, T., Pillans, B., and Kamp, P.J.J., 1997. Recurring global sea-level changes in shelf deposits near the G/ M polarity transition, Wanganui Basin, New Zealand. implications for redefining the Plio-Pleistocene boundary. Quaternary International 40, 61-72.
AAP Memoir 23 (2000)
Neef, G., 1970. Notes on the subgenus Pelicaria . New Zealand Journal of Geology and Geophysics 13, 436-476.
Nees, S., 1997. Late Quaternary palaeoceanography of the Tasman Sea: the benthic foraminiferal view. Palaeogeographu , Palaeoclimatology , Palaeoecology 131, 365-389.
Nelson, C.S., Mildenhall, D.C., Todd,A.J. & Pocknall, D.T., 1988. Subsurface stratigraphy, palaeoenvironments, palynology, and depositional history of the Late Neogene Tauranga Froup at Ohinewai, Lower Waikato Lowland , South Auckland , New Zealand . New Zealand Journal of Geology and Geophysics 31, 21-40.
Nelson, D., Webb, R. & Long, A., 1990. Analysis of stick-nest rat (Leporillus : Muridae) middens from central Australia. 428-434 in Betancourt , J., Van Devender, T. & Martin, P. (eds.) Packrat Middens : The Last 40,000 years of Biotic Change. The University of Arizona Press, Tucson.
Newnham, R ., 1999. Environmental change in Northland, New Zealand during the last glacial and Holocene. Quaternary International 57158, 61-70.
O'Connor , S., 1992. The timing and nature of prehistoric island use in northern Australia . Archa eology in Oceania 27, 49-60.
O'Connor, S., 1995. Carpenter's Gap rockshelter 1: 40,000 years of Aboriginal occupation in the Napier Ranges, Kimberley, WA. Australian Archaeology 40, 58-9.
Ogden, J., Basher, L. & McGlone, M., 1998.Fire, forest regeneration and links with early human habitation : evidence from New Zealand. Annals of Botany 81, 687-696.
Ogden, J., Wilson, A., Hendy, C., Hogg, A. & Newnham R.M., 1992. The late Quaternary history of kauri (Agathis australis) in New Zealand and its climatic significance.Journal of Biogeography 19,611-622.
Osborne, R.A.L., 1997. Rehabilitation of the Wellington Caves Phosphate Mine: implications for Cainozoic Stratigraphy. Proceedings of the Linnaeaociety of New South Wales 117, 175-180.
Pandolfi, J.M. , 1996. Limited membership in Pleistocene reef coral assemblages from the Huon Peninsula, Papua New Guinea: constancy during
global change. Palaeobiology 22, 152-176. Partridge, T.C., (Ed.) 1997a. The Plio-Pleistocene
Boundary. Special Issue, Quaternary International 40(1), 1-100.
Partridge, T.C., l 997b. Reassessment of the position of the Plio-Pleistocene boundary: is there a case for lowering it to the Gauss-Matuyama palaeomagnetic reversal? Quaternary International 40, 5-10.
Partridge, T.C., Wood, B.A. & deMenocal, P.B., 1995. The influence of global climate change and regional uplift on large-mammalian evolution in east and southern Africa . 331-355 in Vrba, E.S., Denton ,
AAP Memoir 23 (2000)
G.H ., Partridge , T.C. & Burckle , L.H . (eds .) Palaeoclimat e and Evolution with Emphasi s on Human Origins,Yale University Press, New Haven.
Pavlides , C., 1993. New archaeological research at Yombon, west New Britain, Papua New Guinea.
Archa eology in Oceania 28(1), 55-59. Pavlides , C. & Gosden, C., 1994. 35,000-year-old sites
inthe rainforests of West New Britain, Papua New Guinea. Antiquity 68, 604-610.
Pearce, R.H. & Barbetti, M., 1981. A 38,000-year-old archaeological site at Upper Swan, Western Australia. Archaeology in Oceania 16(3), 173-178.
Pearce , A.F. & Walker, D.I. (editors) 1991. The Leeuwin Current: an influence on the coastal climate and marine life of Western Australia .Journal of the
Royal Society of Western Australia 74, 1-140. Pearson . N., 1996. An Early Ple isto cene
pala eoecological record from Graham s Quarry, Franksto n, Australia. Unpublished BA (Hons.) thesis, Monash University, Melbourne.
Pearson, S., 1997. Stick-nest rat middens as a source of palaeoenvironmental data in central Australia. Unpublished PhD thesis, University of New South Wales.
Pearson , S. & Dodson, J., 1993. Stick-nest rat middens as sources of palaeoecological data in Australian deserts. Quaternary Research 39, 347-354.
Peerdeman , F.M., 1993. The Pleistocen e climatic and sea-level signature of the northeastern continental margin . Unpublished PhD Thesis, Australian National University, Canberra.
Peerdeman, F,M,, Davies, P.J. & Chivas, A.R., 1993. The .stable oxygen isotope signal in shallow-water, upper-slope sediments off the Great Barrier Reef (Hole 820A). Proceedings of the Ocean Drilling Program , Scientific Results 133, 163-173.
Piperno , D .R ., 1988. Phytolith Analysis: An Archa eological and Geological Perspectiv e. Academic Press, San Diego. 280p.
Porch , N ., submitted. Palaeoenvironmental interpretation of late Quaternary non-marine mollusc assemblages from Pulbeena Swamp, northwestern Tasmania. Alcheringa.
Porch, N. &Allen, J., in prep. The palaeoenvironmental significance of a 35,000 year record of landsnails and vertebrates from Warreen Cave, southwest Tasmania .
Porch , N . & Elias, S., 2000. Quaternary beetles : a review and issues for Australian studies.Australian Journal of Entomology 39, 1-9.
Preece, R.C., 1990. The molluscan fauna of the Middle Pleistocene interglacial deposits at Little Oakley, Essex, and its environmental and stratigraphic implications. Transactions of the Royal Society of London B328, 387-407.
Quilty, P.G., 1974. Tertiary stratigraphy of Western Australia . Jo urnal of the Geological Society of Australia 21, 301-318.
513
Quilty, P.G., 1977. Cenozoic sedimentation cycles in Western Australia. Geology 5, 336-340.
Reid, M. A., 1997. A diatom-based palaeoecological study of the Gou/burn River Billabongs , Southeastern Australia. Unpublished PhD Thesis, Monash University, Melbourne.
Reid, M.A., Tibby, J.C., Penny, D. & Gell, P.A., 1995. The use of diatoms to assess past and present water quality. Australian Journal of Ecology 20, 57-64.
Rich, T.H., 1991. Monotremes, placentals, and marsupials: their record in Australia and its biases. 894-1070 in Vickers-Rich, P., Monaghan , J.M., Baird , R.F & Rich , T.H . (eds .), Vertebrate Palaeontology of Australasia , Pioneer Design Studio, Lilydale, Victoria .
Ride, W.D.L. &Davis,A.C., 1997. Origins and setting: mammal Quaternary palaeontology in the eastern highlands of New South Wales. Proceedings of the Linnean Society of New South Wales 117, 197-222.
Roberts, R.G., Bird, M., Olley, J., Galbraith, R., Lawson, E., Laslett, G., Yoshida, H., Jones, R.,
Fullagar, R., Jacobsen, G. & Hua, Q., 1998. Optical and radiocarbon dating at Jinmium rock shelter in northern Australia. Nature 393, 358.
Roberts, R. G. & Jones, R., 1994. Luminescence dating of sediments: new light on the human colonisation of Australia. Australian Aboriginal Studies 199412,2-17.
Roberts, R. G., Jones, R. & Smith, M. A., 1990. Thermoluminescence dating of a 50,000 year-old human occupation site in northern Australia. Nature
345, 153-6. Roberts, R. G., Jones, R., Spooner, N. A., Head, J.,
Murray A. S. & Smith, M. A., 1994. The human colonisation of Australia: optical dates of 53,000 and 60,000 years bracket human arrival at Deaf Adder Gorge, Northern Territory. Quaternary Science Reviews 13, 575-83.
Rowe, C., David , B. & Stanisec, J., in press . The helicinid land snail Pleuropoma extincta (Odhner, 1917) as an environmental indicator in archaeology. Memoirs of the Queensland Museum.
Ruddiman, W.F. & Raymo, M.E., 1984. Northern hemisphere climate regimes during the last 3 Ma: possible tectonic connections. Philosophical Transactions of the Royal Society of London B318, 411-430.
Ruddiman, W.F., Raymo , M.E., Martinson, D.G., Clement, B.M. & Backman, J., 1989. Pleistocene evolution : Northern Hemisphere ice sheets and North Atlantic Ocean. Paleoceanography 4, 353- 412.
Saito, T., Thompson, P.R . & Breger, D., 1981. Systematic Index of Recent and Pleistocene Planktonic Foraminif era. University of Tokyo Press, Tokyo. 190p.
Scott, L. & Bousman, C., 1990. Palynological analysis of hyrax middens from southern Africa. Palaeogeograph y, Palaeoclimatology, Palaeoecology 79, 367-379.

514
Shackleton, N.J., 1995. New data on the evolution of Pliocene climatic variability. 242-248 in Vrba, E.S., Denton, G.H., Partridge, T.C. & Burckle, L.H. (eds.), Palaeoclimate and Evolution with Emphasis on Human Origins. Yale University Press, New Haven.
Shackleton, N.J., Backman, J. & Zimmerman, H.B., 1984. Oxygen isotope callibration of the onset of icerafting in DSDP Site 552A: history of glaciation in the North Atlantic region. Nature 307, 620-623,
Shackleton, N .J., Berger, A. & Peltier, W.R., 1990. An alternative astronomical calibration of the Lower Pleistocene timescale based on ODP Site 677. Transactions of the Royal Society of Edinburgh 81, 251-261.
Shackleton, N.J. & Opdyke, N.D., 1973. Oxygen isotope and palaeomagnetic stratigraphy of equatorial Pacific core V28-238: oxygen isotope temperatures and ice volumes on a 105 and 106 year scale. Quaternary Research 3, 39-59.
Shackleton, N.J., Crowhurst, S., Hagelberg, T., Pisias, N. & Schneider, D.A., 1995. A new late Neogene timescale: applications to leg 138 sites. Proceedings of Ocean Drilling Program, Scientific Results 138, 73-101.
Shulmeister, J. & Lees, B.G., 1995.Pollen evidence from tropical Australia for the onset of an ENSO dominated climate at c.4000 BP. The Holocene 5, 10-18.
Sikes, E.L. & Volkman, J.K., 1993. Calibration of alkenone unsaturation ratios (Uk '37) for paleotemperature estimation. Geochimica et Cosmochimica Acta 57, 1883-1889.
Sikes, E.L., Volkman, J.K., Robertson, L.G. & Pichon, J.-J., 1997. Alkenones and alkenes in surface water and sediments of the Southern Ocean: Implications for paleotemperature estimation in polar regions. Geochimica et Cosmochimica Acta 61, 1495-1505.
Singh, G., 1981 Late Quaternary pollen records and seasonal palaeoclimates of Lake Frome, South Australia. Hydrobiologia 82, 418-430.
Singh, G. & Luly, J., 1992. Changes in vegetation ad seasonal climate since the last full glacial at Lake Frome, South Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 84, 75-86.
Singh, G., Geissler, E.A., 1985. Late Cainozoic history of fire, lake levels and climate at Lake George, New South Wales, Australia. Philosophical Transactions of the Royal Society of London 311, 379-447.
Singh, G., Kershaw, A.P. & Clark, R., 1981, Quaternary vegetation and fire history in Australia. 23-54 in Gill, A.M., Groves, R.A. & Noble, LR. (eds.) Fire and the Australian Biota. Australian Academy of Science, Canberra.
Sluiter, I.R.K. & Parsons, R.F., 1995. On the Holocene palaeoenvironmental record from Lake Tyrrell, northwestern Victoria, Australia - a reply. Journal of Biogeography 22, 149-152.
AAP Memoir 23 (2000)
Smith, M.A., Yellen, L. & Pask, J., 1995. Vegetation history from archaeological charcoals in central Australia: the late Quaternary record from Puritjarra Rock Shelter. Vegetation History and Archaeobotany 4, 171-177.
Sondaar, P.Y., van den Burgh, G.D., Mubroto, B., Aziz, F., de Vos, J. &Batu, U.L., 1994.Middle Pleistocene faunal turnover and colonization of Flores (Indonesia) by Homo erectus. Comptes Rendues de la Academie des Sciences, Paris 319, Series II, 1255- 1262.
Sue, J.-P., Bertini, A., Leroy, S. & Suballyova, A.G. 1997. Towards the lowering of the Plio-Pleistocene boundary to the Gauss-Matuyama reversal. Quaternary International 40, 37-42.
Sun, X., and McMinn, A., 1994. Recent dinoflagellate cyst distribution associated with the Subtropical Convergence on the Chatham Rise, east of New Zealand. Marine Micropalaeontology 23, 345-356.
Swisher, C.C.W.J., Curtis, T., Jacob, A.G., Getty, A, Suprijo and Widiasmoro, 1994. The age of the earliest hominids in Indonesia. Science 263, 1118- 1121.
Tiedemann, R. Samthein, M. & Shackleton, N.J. 1994. Astronomic timescale for the Pliocene Atlantic d180 dust flux records of Ocean Drilling Program Site 659. Paleoceanography 9, 619-638.
Thunell, R., Anderson, D., Gellar, D. & Miao, Q., 1884. Sea surface temperature estimates for the tropical western Pacific, and their implications for the West Pacific Warm Pool. Quaternary Research 41, 255- 261.
Tudor, E.R., 1973. Hydrological interpretations of diatom assemblages in two Victorian crater lakes. Unpublished M. Sc. Thesis, University of Melbourne.
Urban, B., Kershaw, A.P., D'Costa, D.M. & Heijnis, H., 1996. The application of the concept of supercycles to the interpretation and correlation of terrestrial Quaternary records from Germany and Australia. Eos 77, 22.
van de Geer, G., Reusser, L.E., Lynch-Stieglitz, J. & Charles, Gl.D., 1994 Palaeoenvironments of Tasmania inferred from a 5-75 ka marine pollen record. Palynology 18, 33-40.
van de Graaff, W.J .E., Denman, P.D. & Hocking, R.M., 1977. Emerged Pleistocene marine terraces on Cape Range, Western Australia. Geological Survey of Western Australia, Annual Report (1975), 62-70.
van der Kaars, W.A., 1991. Palynology of eastern Indonesian marine piston-cores, A Late Quaternary vegetational and climatic record for Australasia. Palaeogeography , Palaeoclimatology, Palaeoecology 85, 239-302.
van der Kaars, S., 1998. Marine and terrestrial pollen records of the last glacial cycle from the Indonesian region: Bandung and Banda Sea. Palaeoclimates 3, 209-219.
AAP Memoir 23 (2000)
van der Kaars, S., Wang, X., Kershaw, A.P., Guichard, F. & Setiabudi, D.A. 2000. A late Quaternary palaeoecological record from the Banda Sea, Indonesia: patterns of vegetation, climate and biomass burning in Indonesia and northern Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 155, 135-153.
van Huet, S., Grun, R., Murray-Wallace, C.V., Redvers- Newton, N, and White, J.P., 1998. Age of the Lancefield megafauna: a reappraissal. Australian Archaeology 46, 5-11.
Veeh, H.H., Schwebel, D., van der Graaff, W.J.E .. & Denman, P.D ., 1979. Uranium-series ages of coralline terrace deposits in Western Australia. Journal of the Geological Society of Australia 26, 285-292.
Veron, J.E.N., 1995. Corals in space and time: the biogeography and evolution of the Scleractinia . University of New South Wales Press, Sydney. 321p.
Veron, J.E.N. & Kelley, R., 1988. Species stability in reef corals of Papua New Guinea and the Indo- Pacific. Memoir of the Association of Australasian
Palaeontologists 6, 1-69. Veth, P. 1989. Islands of the interior: a model for the
colonization of Australia's arid zone. Archaeology in Oceania 24, 81-92.
Vickers-Rich, P., Monaghan, J.M., Baird, R.F. & Rich, T.H. (eds.), 1991. Vertebrate Palaeontology in Australasia, Pioneer Design Studio, Lilydale.
Wagstaff, B.E., Kershaw, A.P., O'Sullivan, P., Harle, K.J. & Edwards, J., in press. An Early to Middle Pleistocene palynological record from the volcanic crater of Pejark Marsh, Western Plains of Victoria, southeastern Australia. Palaeogeography, Palaeoclimatology, Palaeoecology.
Wang, X., van der Kaars, S., Kershaw, P., Bird, M., and Jansen, F., 1999. A record of fire, vegetation and climate through the last three glacial cycles from Lombok Ridge core G6-4, eastern Indian Ocean, Indonesia . Palaeogeography, Palaeoclimatology, Palaeoecology 147, 241-256.
Wasson, R.J., 1989. Desert dune building, dust raising and palaeoclimate in the southern hemisphere during the last 280,000 years, pp. 123-137 in T.H. Donnelly and R.J. Wasson (eds.) CL/MANZ 3. CSIRO Division of Water Resources, Canberra.
Watts, W.A., 1978. Plant macrofossils and Quaternary palaeoecology. 53-68 in Walker, D. & Guppy, J.C. (eds.), Biology and Quaternary Environments. Australian Academy of Science, Canberra.
Webb., L.J., 1968. Environmental relationships of the structural types of Australian rainforest vegetation. Ecology 49, 296-311.
515
Webb, S., 1989. The Willandra Lakes Hominids . Department of Prehistory, Research School of Pacific Studies, Australian National University, Canberra. 194p.
Wei, W. & Gartner, S., 1993. Neogene calcareous nannofossils from Sites 811 and 819 through 825, offshore northeastern Australia. Proceedings of the Ocean Drilling Program, Scientific Results, 133, 19- 37.
Wells, P. & Okada, H., 1996. Holocene and Pleistocene glacial palaeoceanography off southeastern Australia, based on foraminifers and nannofossils in Verna cored hole V18-222. Australian Journal of Earth Sciences 43, 509-523.
White, J.P. & O'Connell, J.F., 1982. A Prehistory of Australia, New Guinea and Sahu/ .Academic Press, Sydney. 286p.
Williams, M.A.J., Dunkerley, D.L., De Deckker, P., Kershaw, A.P. & Chappell, J., 1998 Quaternary Environments. 2nd Edition. Edward Arnold, London. 329p.
Worthy, T.H., 1998. The Quaternary fossil faunas of Otago, South Island, New Zealand. Journal of the Royal Society of New Zealand 28, 421-521.
Worthy, T.H. & Holdaway, R.N., 1993. Quaternary fossil faunas from caves in the Punakaiki area, West Coast, South Island, New Zealand. Journal of the Royal Society of New Zealand 23, 147-254.
Worthy, T.H. & Holdaway, R.N., 1994. Quaternary fossil faunas from caves in Takaka Valley and on Takaka Hill, northwest Nelson, South Island, New Zealand. Journal of the Royal Society of New Zealand 24, 297-391.
Worthy, T.H. & Holdaway, R.N., 1996. Quaternary fossil faunas, overlapping taphonomies, and palaeofaunal reconstruction in North Canterbury, South Island, New Zealand . Journal of the Royal Society of New Zealand 26, 275-361 .
Wright, H.E. Jnr., Kutzbach, J.E., Webb T.I., Ruddiman, W.E.F, Street-Perrott, F.A. & Bartlein, P.J. (eds.), 1993. Global Climates Since the Last Glacial Maximum . University of Minnesota Press, Minneapolis. 569p.
Wright, R., 1986. How old is zone F at Lake George? Archaeology in Oceania 21, 138-139.
Yassini, I. & Jones, B.G., 1995. Foraminifera and Ostracoda from estuarine and shelf environments on the southeastern coast of Australia. University of Wollongong Press, Wollongong. 484p.
Zobel, B., 1984. Changes in Quaternary oceanographic bottom conditions as documented by foraminifera in sediment cores from the slope of the Exmouth Plateau off Western Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 48, 3-23.
-
I '
Related Documents











