doi:10.1182/blood-2005-03-1204 Prepublished online August 16, 2005; 2005 106: 3907-3916 Mohammed, Ole N. Jensen, Giorgio Palestro, Giorgio Inghirami and Roberto Chiarle Carlotta Costa, Guido Tarone, Paola Defilippi, Emilio Hirsch, Elisabetta Boeri Erba, Shabaz Chiara Ambrogio, Claudia Voena, Andrea D. Manazza, Roberto Piva, Ludovica Riera, Laura Barberis, lymphoma kinase p130Cas mediates the transforming properties of the anaplastic http://bloodjournal.hematologylibrary.org/content/106/12/3907.full.html Updated information and services can be found at: (1930 articles) Signal Transduction (795 articles) Oncogenes and Tumor Suppressors (4217 articles) Neoplasia Articles on similar topics can be found in the following Blood collections http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: Copyright 2011 by The American Society of Hematology; all rights reserved. Washington DC 20036. by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.org From

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1182/blood-2005-03-1204Prepublished online August 16, 2005;2005 106: 3907-3916
Mohammed, Ole N. Jensen, Giorgio Palestro, Giorgio Inghirami and Roberto ChiarleCarlotta Costa, Guido Tarone, Paola Defilippi, Emilio Hirsch, Elisabetta Boeri Erba, Shabaz Chiara Ambrogio, Claudia Voena, Andrea D. Manazza, Roberto Piva, Ludovica Riera, Laura Barberis, lymphoma kinasep130Cas mediates the transforming properties of the anaplastic
http://bloodjournal.hematologylibrary.org/content/106/12/3907.full.htmlUpdated information and services can be found at:
(1930 articles)Signal Transduction � (795 articles)Oncogenes and Tumor Suppressors �
(4217 articles)Neoplasia �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Washington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

NEOPLASIA
p130Cas mediates the transforming properties of the anaplastic lymphoma kinaseChiara Ambrogio, Claudia Voena, Andrea D. Manazza, Roberto Piva, Ludovica Riera, Laura Barberis, Carlotta Costa,Guido Tarone, Paola Defilippi, Emilio Hirsch, Elisabetta Boeri Erba, Shabaz Mohammed, Ole N. Jensen,Giorgio Palestro, Giorgio Inghirami, and Roberto Chiarle
Translocations of the anaplastic lym-phoma kinase (ALK) gene have been de-scribed in anaplastic large-cell lympho-mas (ALCLs) and in stromal tumors. Themost frequent translocation, t(2;5), gener-ates the fusion protein nucleophosmin(NPM)–ALK with intrinsic tyrosine kinaseactivity. Along with transformation, NPM-ALK induces morphologic changes in fi-broblasts and lymphoid cells, suggestinga direct role of ALK in cell shaping. In thisstudy, we used a mass-spectrometry–based proteomic approach to search forproteins involved in cytoskeleton remod-
eling and identified p130Cas (p130 Crk-associated substrate) as a novel interac-tor of NPM-ALK. In 293 cells and infibroblasts as well as in human ALK-positive lymphoma cell lines, NPM-ALKwas able to bind p130Cas and to induceits phosphorylation. Both of the effectswere dependent on ALK kinase activityand on the adaptor protein growth factorreceptor–bound protein 2 (Grb2), sinceno binding or phosphorylation was foundwith the kinase-dead mutant NPM-ALKK210R or in the presence of a Grb2dominant-negative protein. Phosphoryla-
tion of p130Cas by NPM-ALK was partiallyindependent from Src (tyrosine kinasepp60c-src) kinase activity, as it was stilldetectable inSyf�/� cells.Finally,p130Cas�/�
(also known as Bcar1�/�) fibroblasts ex-pressing NPM-ALK showed impaired actinfilament depolymerization and were nolonger transformed compared with wild-type cells, indicating an essential role ofp130Cas activation in ALK-mediated trans-formation. (Blood. 2005;106:3907-3916)
© 2005 by The American Society of Hematology
Introduction
Nucleophosmin–anaplastic lymphoma kinase (NPM-ALK) is anoncogenic protein generated by the t(2;5)(p23;q35) translocation1
that characterizes most anaplastic large-cell lymphomas (ALCLs).Each of the 2 fusion-derived fractions that form NPM-ALK plays arole in the constitutive tyrosine kinase activity of the chimericprotein. NPM is a nucleolar phosphoprotein involved in the ribosomegenesis,2 in the centrosome duplication,3 and in the regulation ofthe basal level of p53.4 NPM promoter drives the constitutiveexpression of NPM-ALK in lymphoid cells; the oligomerizationdomain of NPM allows the dimerization5 and hence the autophos-phorylation of NPM-ALK. ALK is a 210-kDa tyrosine kinase (TK)receptor belonging to the insulin growth factor receptor superfam-ily, which is widely expressed in the nervous system duringembryogenesis but only focally in the adult brain.1,6,7 Although thephysiologic role of ALK receptor is still unknown, it seems to beinvolved in neuronal cell differentiation as suggested by the abilityto induce neurite outgrowth in vitro8 and by its fundamental role insynapse formation shown in Caenorhabditis elegans.9
The constitutive NPM-ALK activity transforms rat-1 fibro-blast10 and is responsible for the malignant transformation oflymphoid cells.11 Furthermore, NPM-ALK has the ability to induceB- and T-cell lymphomas in vivo.12,13 The signaling pathwaysdownstream from NPM-ALK follow a transduction pattern sharedwith most of deregulated tyrosine kinases, leading to increased
proliferation, resistance to apoptosis, and morphologic changes.The enhancement of cell growth in NPM-ALK–positive cellsresults from the activation of the rat sarcoma viral oncogenehomolog (Ras)/extracellular signal-related kinase (ERK) pathwaythrough the recruitment of the adaptors SHC (Src homology 2domain containing protein) and insulin receptor substrate 1 (IRS-1).14
In addition, NPM-ALK controls cellular mitogenity by binding tophospholipase C � (PLC-�).15
In parallel, the inhibition of apoptosis in the NPM-ALK signaltransduction derives from the activation of signal transducer andactivator of transcription 3 (Stat3),16 leading to an increasedexpression of the antiapoptotic factor Bcl-x(L) and from the stimulationof the phosphatidylinositol 3–kinase (PI3K)/Akt pathway,17 whichconfers resistance to apoptosis combined with a deregulatedproliferation rate due to the transcription factor forkhead boxO3a (FOXO3a).18
In contrast to these fairly well-characterized pathways, minimaldata are available regarding the effects of ALK tyrosine kinaseactivity on cell shape. Upon transduction in Hodgkin lymphomacell lines, NPM-ALK induces in these cells morphologic changesthat make them similar to ALCL cells.19 Moreover, in PC12 cells,ectopic expression of ALK induces a neurite outgrowth that hasbeen shown to depend on the mitogen-activated protein (MAP)kinase pathway.8 In C elegans, the degradation of ALK is required
From the Center for Experimental Research and Medical Studies (CERMS)and the Departments of Biomedical Sciences and Human Oncology and ofGenetics, Biology, and Biochemistry, University of Turin, Turin, Italy; and theDepartment of Biochemistry and Molecular Biology, University of SouthernDenmark, Odense, Denmark.
Submitted March 24, 2005; accepted July 18, 2005. Prepublished online asBlood First Edition Paper, August 16, 2005; DOI 10.1182/blood-2005-03-1204.
Supported by National Institutes of Health (NIH) R01-CA64033, Ministerodell’Universita e Ricerca Scientifica (MIUR), Compagnia di San Paolo, Torino
(Progetto Oncologia), Regione Piemonte (Ricerca Sanitaria Finalizzata andRicerca Scientifica), and Associazione Italiana per la Ricerca sul Cancro(AIRC).
Reprints: Roberto Chiarle, Department of Biomedical Sciences and HumanOncology, Via Santena 7 10126, Turin, Italy; e-mail: [email protected].
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. section 1734.
© 2005 by The American Society of Hematology
3907BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

for synapse stabilization,9 whereas in Drosophila, ALK is activatedby its ligand Jelly Belly to specify visceral mesoderm migrationand differentiation into muscle cells.20,21 Moreover, ALK ectopicexpression in Drosophila hemocytes promotes the formation oflamellocytes, specialized cells that participate in the encapsulationand killing of parasites, a phenotype that is coupled with theactivation of both the Janus kinase (JAK)/STAT and the Jun kinasepathways through Rho guanosine triphosphatase (GTPase) Rac1.22
In this paper we investigated molecules involved in cellmorphology that could also be relevant for the NPM-ALK–mediated transformation process. We started from a mass spectrom-etry–based screening of proteins interacting with NPM-ALK andinvolved in the cytoskeleton morphology. Through this analysis weidentified p130Cas (p130 Crk-associated substrate) and then dem-onstrated that NPM-ALK is able to bind and phosphorylatep130Cas. The p130Cas activation is mediated by the adaptorprotein growth factor receptor–bound protein 2 (Grb2) and ispartially independent from Src. Importantly, we show that p130Casactivation is required for NPM-ALK–mediated cell shape modifica-tions and actin filament depolymerization as well as for celltransformation, but not for cell migration, induced by NPM-ALK.
Taken together, these results suggest not only that p130Cas actsas one of the effectors of the cytoskeleton modifications induced byNPM-ALK but also that morphologic and growth cues are deeplyembedded in NPM-ALK–mediated oncogenic processes.
Materials and methods
Cell lines and culture
Human lymphoid cells TS, DHL, and Karpas (NPM-ALK–positive) andCEM, K562, and Namalwa (NPM-ALK–negative) were obtained fromNew York University and maintained in RPMI 1640 (BioWhittaker,Verviers, Belgium) containing 10% fetal calf serum, 2 mM glutamine(Eurobio Biotechnology, Les Ulis, France), 100 U/mL penicillin, and100 �g/mL streptomycin (Eurobio Biotechnology).
Human embryonal kidney cells 293T, 293GP, and 293 T-Rex Tet-On(Invitrogen, Carlsbad, CA) and murine fibroblasts MEF, NIH3T3, Syf �/�,Cas�/� (also known as Bcar1�/�), and MEF Tet-Off (Clontech, Palo Alto,CA) were maintained in Dulbecco modified Eagle medium (DMEM)supplemented with 10% fetal calf serum, 2 mM glutamine (EurobioBiotechnology), 100 U/mL penicillin, and 100 �g/mL streptomycin (Euro-bio Biotechnology). In MEF and 293 Tet systems, the working concentra-tion of tetracycline or doxycycline in the medium was 1 �g/mL.
Inducible ALK-shRNA interference TS cells were obtained by cotrans-duction with pLVTH vector containing the H1 promoter ALK-shRNAcassette and pLV-tTRKRAB vector,23 as described by Piva et al.53 Thesecells undergo NPM-ALK silencing when 1 �g/mL of doxycycline is addedto the medium for at least 72 hours.
Cell lysis, immunoprecipitation, and immunoblotting
Total cellular proteins were extracted with 20 mM Tris-HCl (pH 7.4), 150mM NaCl, 5 mM EDTA (ethylenediaminetetraacetic acid), 0.1% TritonX-100, 1 mM phenylmethylsulfonyl fluoride (PMSF), 10 mM NaF, 1 mMNa3VO4, and protease inhibitors (Roche, Mannheim, Germany). Celllysates were centrifuged at 10 000g (13 000 rpm) for 15 minutes and thesupernatants were collected and assayed for protein concentration using theBio-Rad protein assay method. Thirty micrograms of proteins was run onsodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE)under reducing conditions.
For immunoprecipitation experiments, 300 �g to 800 �g protein wasimmunoprecipitated with the appropriate antibody for 1 hour at 4°Cfollowed by 30 �L protein A–Sepharose beads (Amersham, Freiburg,
Germany). After SDS-PAGE, proteins were transferred to nitrocellulose,incubated with the specific antibody, and then detected with peroxidase-conjugated secondary antibodies and enhanced chemiluminescent reagent(Amersham).
Reagents and antibodies
PP2 Src-family kinase inhibitor was purchased from Calbiochem (Darm-stadt, Germany). The following antibodies were used: polyclonal anti-ALK(1:2000; Zymed, Basel, Switzerland), monoclonal anti-ALK (1:4000;Zymed), monoclonal anti-p130Cas (1:1000; Becton Dickinson, MountainView, CA), monoclonal anti–phospho-Tyr (PY20; 1:1000; TransductionLaboratories, Lexington, KY), polyclonal anti-Grb2 (1:2000; Santa CruzBiotechnology, Santa Cruz, CA), monoclonal anti-Crk (1:1000; Transduc-tion Laboratories), polyclonal anti-Src (1:1000; Santa Cruz Biotechnology),monoclonal antipaxillin (1:1000; Transduction Laboratories), monoclonalantiactin (1:1000; Santa Cruz Biotechnology), and monoclonal anti–focaladhesion kinase (anti-FAK; 1:1000; hybridome immunoglobulin G1 [IgG1]clone 4-4A, kindly gifted by Prof Silengo, University of Turin, Italy).Secondary antimouse or antirabbit peroxidase-conjugated antibodies werepurchased from Amersham.
Immunoprecipitation and mass spectrometry analysis
For anti-ALK immunoprecipitation, 293 T-Rex Tet-on cells (Invitrogen)transfected with a wild-type (wt) NPM-ALK and a kinase-dead mutantcontrol NPM-ALKK210R were grown to confluence and induced with1 �g/mL of tetracycline for 24 hours. Cells were lysed in 20 mM Tris (pH7.4), 150 mM NaCl, 5 mM EDTA, and 0.1% Triton X-100 in the presenceof protease and phosphatase inhibitors. Ten milligrams of cleared cell lysatewas mixed with 100 �g monoclonal anti-ALK coupled to agarose beads(Amersham) and rotated at 4°C for 2 hours. Precipitated immunocomplexeswere washed 3 times with lysis buffer, eluted, and boiled in Laemli bufferfor 5 minutes. Samples were resolved on SDS-PAGE and silver stained.Bands of interest were excised from the gel and subjected to in-gelreduction, alkylation, and digestion with trypsin (Promega, Madison, WI)and analyzed by matrix-assisted laser desorption ionization/time of flight(MALDI-TOF) mass spectrometry. Spectra were obtained using either aReflex III MALDI-TOF mass spectrometer or an Ultraflex TOF/TOF massspectrometer (Bruker Daltonics, Bremen, Germany), operating in thepositive ion delayed extraction reflector mode. Ions were generated byirradiation of analyte/matrix deposits by a nitrogen laser at 337 nm andanalyzed with an accelerating voltage of 20 kV. Each MALDI-TOFspectrum was generated by accumulating data corresponding to more than400 laser shots. Mass calibration in the range mass over electronic charge(m/z) 800 to 4000 was performed by using a bovine beta-lactoglobulintryptic peptide mixture. Postacquisition internal calibration was applied byusing theoretic masses of trypsin autodigestion peptides. Peptide massspectra were analyzed using Mascot Peptide Mass Fingerprinting software.24
Some samples were analyzed by liquid chromatography–tandem massspectrometry (LC-MS/MS) using a nanoflow–high-performance liquidchromatography (HPLC) system (Ultimate; LC Packings, Amsterdam, TheNetherlands) interfaced to electrospray quadrupole time-of-flight (Q-TOF)tandem mass spectrometers (QTOF Ultima or QTOF Micro; Waters/Micromass, Manchester, United Kingdom). Protein identification viapeptide MS/MS spectra was achieved by using Mascot software24 forsearching the National Center for Biotechnology Information (NCBI)nonredundant protein database.25
For antiphosphotyrosine immunoprecipitation, 293 T-Rex Tet-On cellswere grown in nonadherent conditions on Poly(2-hydroxyethylmethacry-late) (Poly-HEME; Sigma, St Louis, MO) coated plates, starved for12 hours, and induced with 1 �g/mL of tetracycline for 24 hours, pelleted,and stored at �80°C. Cells were lysed in 20 mM Tris (pH 7.4), 150 mMNaCl, 5 mM EDTA, and 1% Nonidet P-40 in the presence of protease andphosphatase inhibitors. Ten milligrams cleared cell lysate was mixed with50 �g agarose-conjugated monoclonal 4G10 (Upstate Biotechnology, LakePlacid, NY) and 10 �g monoclonal PY20 (Transduction Laboratories) andincubated rotating at 4°C overnight. Samples were processed as describedin “Cell lysis, immunoprecipitation, and immunoblotting.”
3908 AMBROGIO et al BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

Immunofluorescence staining
Cells were grown for 12 hours on glass coverslips pretreated withfibronectin (10 �g/mL phosphate-buffered saline [PBS]) at 37°C for 1 hourto facilitate cell adhesion. Samples were fixed in PBS containing 4%paraformaldehyde at room temperature for 10 minutes and permeabilizedwith PBS containing 0.3% Triton X-100 for 5 minutes. Coverslips wereincubated with PBS containing 3% bovine serum albumin (BSA) for 1 hourat room temperature and then stained with primary antibody for 1 hourfollowed by fluorescein isothiocyanate (FITC)– or tetramethyl rhodamineisothiocyanate (TRITC)–conjugated secondary antibody (1:200; Sigma) for1 hour at room temperature.
Primary polyclonal anti-ALK (Zymed) and monoclonal antipaxillin(Transduction Laboratories) antibodies were diluted 1:400 before use.Phycoerythrin (PE)–conjugated phalloidin (1:200 PBS; Sigma) was used tostain actin filaments. Nuclei were stained 10 minutes at room temperaturewith HOECHST (300 ng/mL; Sigma). Coverslips were mounted inantifading solution and viewed using a Leica TCS SP2 laser-scanningconfocal microscope driven by Leica Confocal Software; the images wereacquired at room temperature by means of a 63�/1.32 PL APO objective(Leica, Heidelberg, Germany). Brightfield images were acquired on a LeicaDM IRE2 microscope using a DC300F camera and were analyzed with IM50 software.
DNA constructs
Wild-type NPM-ALK or the kinase-dead mutant NPM-ALKK210R16 wascloned in the plasmid vector pcDNA5TO (Invitrogen) at HindIII/XhoI sitesand stably transfected into 293 T-Rex Tet-On cells using Effectene reagentsas described by manufacturer (Qiagen, Valencia, CA). Single cell–derivedclones were selected for NPM-ALK or NPM-ALKK210R expression levels.Wild-type NPM-ALK, digested to obtain a HindIII/XhoI-blunted fragment,was cloned in the plasmid vector pBIEGFP (Clontech) at MluI-blunted sitesand then stably transfected into MEF Tet-Off. Murine p130Cas was clonedin the plasmid vector pEGFP-C1 (Becton Dickinson) at EcoRI/BamHI sitesor digested to obtain an AseI/BamHI-blunted fragment and cloned in theplasmid vector Pallino26 at EcoRI/XhoI-blunted ends. Wild-type Grb2 or thedominant-negatives Grb2-P49L (mutated in the N-terminal SH3 domain)and Grb2-R86K (mutated in the SH2 domain) were cloned in the plasmidvector pRK5 (a kind gift from Dr A. Pellicer, New York University). Pallinovectors containing NPM-ALK or NPM-ALKK210R or ATIC-ALK (aminoimi-dazole-4-carboxamide ribonucleotide formyltransferase/inosine 5�-mono-phosphate [IMP] cyclohydrolase–ALK) were previously described.16 Full-length human ALK receptor was purchased from ATCC (Manassas, VA).
Retrovirus production and cell infection
NPM-ALK and p130Cas retroviruses were obtained by cotransfection ofPallino expression vector containing wild-type NPM-ALK or p130Cas withpMD2VSV-G plasmid into the 293GP packaging cell line (Invitrogen).Retroviruses released in culture medium were collected 24 hours aftertransfection. Five hundred microliters filtered (pore size 0.45 mm) superna-tants from GP cells was supplemented with 8 �g/mL polybrene (Sigma) andadded to 5 � 104 to 10 � 104 target cells. After 12 hours of incubation,1 mL complete medium was added and the cells were cultured for anadditional 2 days. The percentages of transduced cells were analyzed forGFP expression by fluorescence-activated cell sorter (FACS). Cells wereanalyzed for GFP content on a FACSCalibur flow cytometer (BectonDickinson). Calibration of the instrument was performed using Calibrationbeads (CALIBrite; Becton Dickinson). The CELLQuest software (BectonDickinson) was used for the data acquisition and analysis. For cell sorting,cells were suspended at the concentration of 10 � 106/mL in basic sortingbuffer (5 mM EDTA, 25 mM HEPES [N-2-hydroxyethylpiperazine-N�-2-ethanesulfonic acid; pH 7.0], 1% heat-inactivated fetal bovine serum[FBS]) and then sorted for GFP expression on a MoFlo High-Performancecell sorter (DAKO Cytomation, Glostrup, Denmark).
Soft-agar assay
For clonogenic assay, 5 � 105 cells/well were seeded in 0.35% agar on thetop of a base layer containing 0.7% agar into 6-well plates. After 3 to 5
weeks of cultivation, colonies were counted under a phase-contrastmicroscope. All of the experiments were done using triplicate plates forexperimental point.
Cell-cycle analysis
For DNA content determination, cells were fixed for 1 hour in 70% ethanolat 4°C. After washing, cells were treated with RNase (0.25 mg/mL) andstained with propidium iodide (50 �g/mL). The S-phase fraction wascalculated using the Modfit program from Becton Dickinson.
Chemotaxis assay
Cell migration was evaluated using 24-well, 5-�m pore size Transwellsystem plates (Costar, Cambridge, MA). Purified cells were washed once inRPMI 1640 (BioWhittaker) containing 0.1% fetal calf serum, 2 mMglutamine (Eurobio Biotechnology), 100 U/mL penicillin, and 100 �g/mLstreptomycin (Eurobio Biotechnology) and then adjusted to 1 � 106
cells/mL in the same medium. Cells (1 � 105 in 100 �L) were placed on thetop of the Transwell. Stromal-derived factor-1 � (SDF-1�; R&D Systems,Abingdon, United Kingdom), prepared at the indicated concentrations inthe same RPMI medium (600 �L total volume), was added to the bottom ofthe Transwell system. After 2 hours incubation at 37°C in a 5% CO2
atmosphere, the inserts of the transwell chambers were removed and thenumber of cells that had migrated into the lower well was counted bystaining with a DIF-Quick Kit (IMEB, San Marcos, CA).
Matrigel migration assay
Cells were seeded at a density of 1 � 105 cells onto Matrigel (BectonDickinson)–coated inserts (100 �g/insert) in 24-well, 8-�m pore sizeTranswell system plates (Costar). After a 48-hour incubation, the insertswere removed, the cells on the upper side of the inserts were removed with acotton swab, and the number of cells that migrated to the lower side of thefilters was counted under a phase-contrast microscope by staining withDIF-Quick Kit (IMEB).
Results
NPM-ALK expression modifies the cell cytoskeletonand the migration pattern
We selected different cell lines to test the effects of NPM-ALK oncell morphology, adhesion, and migration. We used 2 tetracycline-inducible cell lines: MEF Tet-Off fibroblasts were grown inDMEM medium containing tetracycline (1 �g/mL) and wereinduced to express NPM-ALK whenever tetracycline was depletedfrom the medium. The 293 T-Rex cells were induced to expressNPM-ALK when 1 �g/mL of tetracycline was added to themedium; NPM-ALK was detectable by Western blot (WB) as earlyas 6 hours after tetracycline induction. NIH3T3 fibroblasts wereretrovirally infected to stably express NPM-ALK. Upon NPM-ALK expression, MEF Tet-Off and NIH3T3 fibroblasts assumedthe typical transformed phenotype with spindle-shaped cell morphol-ogy (Figure 1A-B), whereas 293 T-Rex cells detached from theculture plate (Figure 1C-D).
In fibroblasts, the transformed morphology corresponded to adisruption of the actin cytoskeleton in immunofluorescence (Figure1E-F). Together with actin, we stained for the scaffolding proteinpaxillin as a marker of the cellular structural organization becauseit is found at the interface between the plasma membrane and theactin cytoskeleton.27,28 Noninfected NIH3T3 cells were character-ized by a strong paxillin signal at the focal adhesions, whereasNIH3T3 NPM-ALK cells showed few residual paxillin clusters atthe edge of the elongated cellular processes (Figure 1E-F greenarrows). Moreover, the actin cytoskeleton turned from a polarized
ROLE OF p130cas IN NPM-ALK–MEDIATED TRANSFORMATION 3909BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom
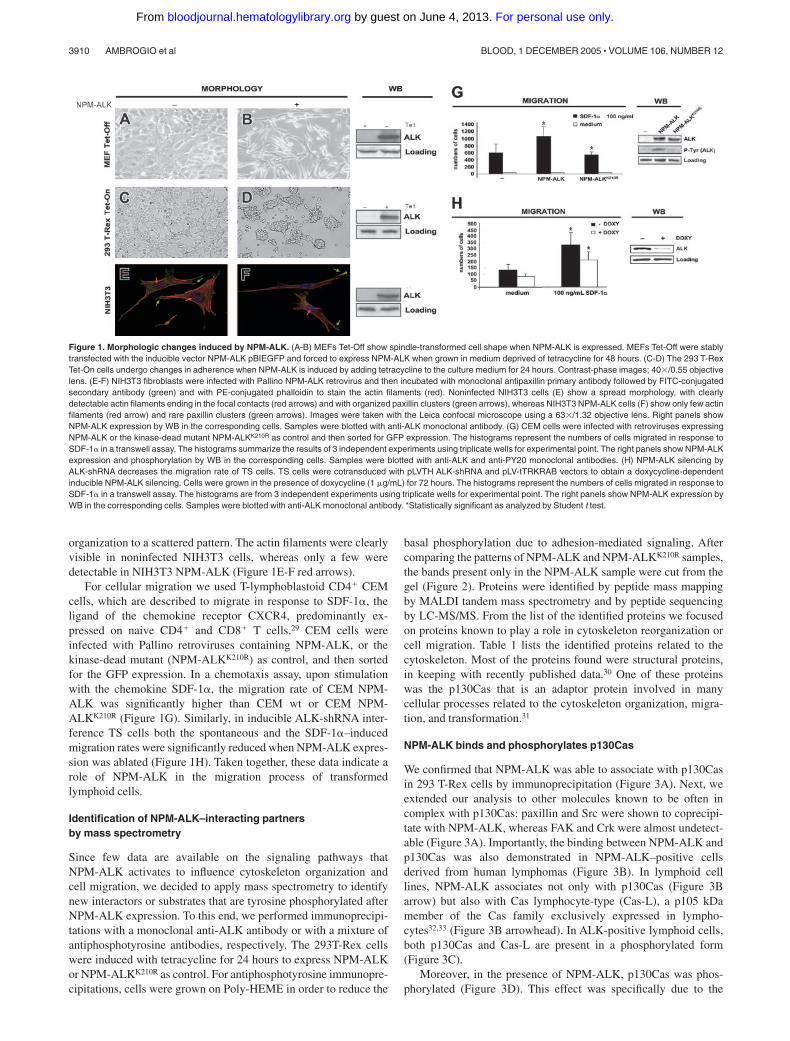
organization to a scattered pattern. The actin filaments were clearlyvisible in noninfected NIH3T3 cells, whereas only a few weredetectable in NIH3T3 NPM-ALK (Figure 1E-F red arrows).
For cellular migration we used T-lymphoblastoid CD4� CEMcells, which are described to migrate in response to SDF-1�, theligand of the chemokine receptor CXCR4, predominantly ex-pressed on naive CD4� and CD8� T cells.29 CEM cells wereinfected with Pallino retroviruses containing NPM-ALK, or thekinase-dead mutant (NPM-ALKK210R) as control, and then sortedfor the GFP expression. In a chemotaxis assay, upon stimulationwith the chemokine SDF-1�, the migration rate of CEM NPM-ALK was significantly higher than CEM wt or CEM NPM-ALKK210R (Figure 1G). Similarly, in inducible ALK-shRNA inter-ference TS cells both the spontaneous and the SDF-1�–inducedmigration rates were significantly reduced when NPM-ALK expres-sion was ablated (Figure 1H). Taken together, these data indicate arole of NPM-ALK in the migration process of transformedlymphoid cells.
Identification of NPM-ALK–interacting partnersby mass spectrometry
Since few data are available on the signaling pathways thatNPM-ALK activates to influence cytoskeleton organization andcell migration, we decided to apply mass spectrometry to identifynew interactors or substrates that are tyrosine phosphorylated afterNPM-ALK expression. To this end, we performed immunoprecipi-tations with a monoclonal anti-ALK antibody or with a mixture ofantiphosphotyrosine antibodies, respectively. The 293T-Rex cellswere induced with tetracycline for 24 hours to express NPM-ALKor NPM-ALKK210R as control. For antiphosphotyrosine immunopre-cipitations, cells were grown on Poly-HEME in order to reduce the
basal phosphorylation due to adhesion-mediated signaling. Aftercomparing the patterns of NPM-ALK and NPM-ALKK210R samples,the bands present only in the NPM-ALK sample were cut from thegel (Figure 2). Proteins were identified by peptide mass mappingby MALDI tandem mass spectrometry and by peptide sequencingby LC-MS/MS. From the list of the identified proteins we focusedon proteins known to play a role in cytoskeleton reorganization orcell migration. Table 1 lists the identified proteins related to thecytoskeleton. Most of the proteins found were structural proteins,in keeping with recently published data.30 One of these proteinswas the p130Cas that is an adaptor protein involved in manycellular processes related to the cytoskeleton organization, migra-tion, and transformation.31
NPM-ALK binds and phosphorylates p130Cas
We confirmed that NPM-ALK was able to associate with p130Casin 293 T-Rex cells by immunoprecipitation (Figure 3A). Next, weextended our analysis to other molecules known to be often incomplex with p130Cas: paxillin and Src were shown to coprecipi-tate with NPM-ALK, whereas FAK and Crk were almost undetect-able (Figure 3A). Importantly, the binding between NPM-ALK andp130Cas was also demonstrated in NPM-ALK–positive cellsderived from human lymphomas (Figure 3B). In lymphoid celllines, NPM-ALK associates not only with p130Cas (Figure 3Barrow) but also with Cas lymphocyte-type (Cas-L), a p105 kDamember of the Cas family exclusively expressed in lympho-cytes32,33 (Figure 3B arrowhead). In ALK-positive lymphoid cells,both p130Cas and Cas-L are present in a phosphorylated form(Figure 3C).
Moreover, in the presence of NPM-ALK, p130Cas was phos-phorylated (Figure 3D). This effect was specifically due to the
Figure 1. Morphologic changes induced by NPM-ALK. (A-B) MEFs Tet-Off show spindle-transformed cell shape when NPM-ALK is expressed. MEFs Tet-Off were stablytransfected with the inducible vector NPM-ALK pBIEGFP and forced to express NPM-ALK when grown in medium deprived of tetracycline for 48 hours. (C-D) The 293 T-RexTet-On cells undergo changes in adherence when NPM-ALK is induced by adding tetracycline to the culture medium for 24 hours. Contrast-phase images; 40�/0.55 objectivelens. (E-F) NIH3T3 fibroblasts were infected with Pallino NPM-ALK retrovirus and then incubated with monoclonal antipaxillin primary antibody followed by FITC-conjugatedsecondary antibody (green) and with PE-conjugated phalloidin to stain the actin filaments (red). Noninfected NIH3T3 cells (E) show a spread morphology, with clearlydetectable actin filaments ending in the focal contacts (red arrows) and with organized paxillin clusters (green arrows), whereas NIH3T3 NPM-ALK cells (F) show only few actinfilaments (red arrow) and rare paxillin clusters (green arrows). Images were taken with the Leica confocal microscope using a 63�/1.32 objective lens. Right panels showNPM-ALK expression by WB in the corresponding cells. Samples were blotted with anti-ALK monoclonal antibody. (G) CEM cells were infected with retroviruses expressingNPM-ALK or the kinase-dead mutant NPM-ALKK210R as control and then sorted for GFP expression. The histograms represent the numbers of cells migrated in response toSDF-1� in a transwell assay. The histograms summarize the results of 3 independent experiments using triplicate wells for experimental point. The right panels show NPM-ALKexpression and phosphorylation by WB in the corresponding cells. Samples were blotted with anti-ALK and anti-PY20 monoclonal antibodies. (H) NPM-ALK silencing byALK-shRNA decreases the migration rate of TS cells. TS cells were cotransduced with pLVTH ALK-shRNA and pLV-tTRKRAB vectors to obtain a doxycycline-dependentinducible NPM-ALK silencing. Cells were grown in the presence of doxycycline (1 �g/mL) for 72 hours. The histograms represent the numbers of cells migrated in response toSDF-1� in a transwell assay. The histograms are from 3 independent experiments using triplicate wells for experimental point. The right panels show NPM-ALK expression byWB in the corresponding cells. Samples were blotted with anti-ALK monoclonal antibody. *Statistically significant as analyzed by Student t test.
3910 AMBROGIO et al BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom
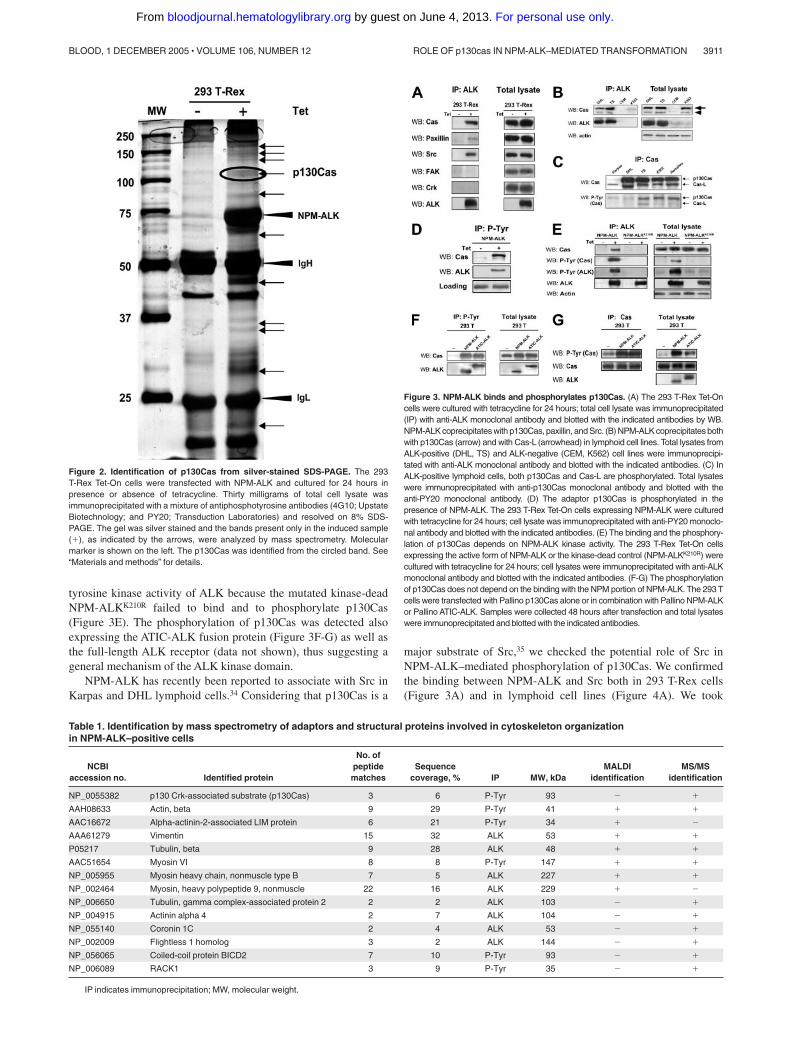
tyrosine kinase activity of ALK because the mutated kinase-deadNPM-ALKK210R failed to bind and to phosphorylate p130Cas(Figure 3E). The phosphorylation of p130Cas was detected alsoexpressing the ATIC-ALK fusion protein (Figure 3F-G) as well asthe full-length ALK receptor (data not shown), thus suggesting ageneral mechanism of the ALK kinase domain.
NPM-ALK has recently been reported to associate with Src inKarpas and DHL lymphoid cells.34 Considering that p130Cas is a
major substrate of Src,35 we checked the potential role of Src inNPM-ALK–mediated phosphorylation of p130Cas. We confirmedthe binding between NPM-ALK and Src both in 293 T-Rex cells(Figure 3A) and in lymphoid cell lines (Figure 4A). We took
Table 1. Identification by mass spectrometry of adaptors and structural proteins involved in cytoskeleton organizationin NPM-ALK–positive cells
NCBIaccession no. Identified protein
No. ofpeptidematches
Sequencecoverage, % IP MW, kDa
MALDIidentification
MS/MSidentification
NP_0055382 p130 Crk-associated substrate (p130Cas) 3 6 P-Tyr 93 � �
AAH08633 Actin, beta 9 29 P-Tyr 41 � �
AAC16672 Alpha-actinin-2-associated LIM protein 6 21 P-Tyr 34 � �
AAA61279 Vimentin 15 32 ALK 53 � �
P05217 Tubulin, beta 9 28 ALK 48 � �
AAC51654 Myosin VI 8 8 P-Tyr 147 � �
NP_005955 Myosin heavy chain, nonmuscle type B 7 5 ALK 227 � �
NP_002464 Myosin, heavy polypeptide 9, nonmuscle 22 16 ALK 229 � �
NP_006650 Tubulin, gamma complex-associated protein 2 2 2 ALK 103 � �
NP_004915 Actinin alpha 4 2 7 ALK 104 � �
NP_055140 Coronin 1C 2 4 ALK 53 � �
NP_002009 Flightless 1 homolog 3 2 ALK 144 � �
NP_056065 Coiled-coil protein BICD2 7 10 P-Tyr 93 � �
NP_006089 RACK1 3 9 P-Tyr 35 � �
IP indicates immunoprecipitation; MW, molecular weight.
Figure 2. Identification of p130Cas from silver-stained SDS-PAGE. The 293T-Rex Tet-On cells were transfected with NPM-ALK and cultured for 24 hours inpresence or absence of tetracycline. Thirty milligrams of total cell lysate wasimmunoprecipitated with a mixture of antiphosphotyrosine antibodies (4G10; UpstateBiotechnology; and PY20; Transduction Laboratories) and resolved on 8% SDS-PAGE. The gel was silver stained and the bands present only in the induced sample(�), as indicated by the arrows, were analyzed by mass spectrometry. Molecularmarker is shown on the left. The p130Cas was identified from the circled band. See“Materials and methods” for details.
Figure 3. NPM-ALK binds and phosphorylates p130Cas. (A) The 293 T-Rex Tet-Oncells were cultured with tetracycline for 24 hours; total cell lysate was immunoprecipitated(IP) with anti-ALK monoclonal antibody and blotted with the indicated antibodies by WB.NPM-ALK coprecipitates with p130Cas, paxillin, and Src. (B) NPM-ALK coprecipitates bothwith p130Cas (arrow) and with Cas-L (arrowhead) in lymphoid cell lines. Total lysates fromALK-positive (DHL, TS) and ALK-negative (CEM, K562) cell lines were immunoprecipi-tated with anti-ALK monoclonal antibody and blotted with the indicated antibodies. (C) InALK-positive lymphoid cells, both p130Cas and Cas-L are phosphorylated. Total lysateswere immunoprecipitated with anti-p130Cas monoclonal antibody and blotted with theanti-PY20 monoclonal antibody. (D) The adaptor p130Cas is phosphorylated in thepresence of NPM-ALK. The 293 T-Rex Tet-On cells expressing NPM-ALK were culturedwith tetracycline for 24 hours; cell lysate was immunoprecipitated with anti-PY20 monoclo-nal antibody and blotted with the indicated antibodies. (E) The binding and the phosphory-lation of p130Cas depends on NPM-ALK kinase activity. The 293 T-Rex Tet-On cellsexpressing the active form of NPM-ALK or the kinase-dead control (NPM-ALKK210R) werecultured with tetracycline for 24 hours; cell lysates were immunoprecipitated with anti-ALKmonoclonal antibody and blotted with the indicated antibodies. (F-G) The phosphorylationof p130Cas does not depend on the binding with the NPM portion of NPM-ALK. The 293 Tcells were transfected with Pallino p130Cas alone or in combination with Pallino NPM-ALKor Pallino ATIC-ALK. Samples were collected 48 hours after transfection and total lysateswere immunoprecipitated and blotted with the indicated antibodies.
ROLE OF p130cas IN NPM-ALK–MEDIATED TRANSFORMATION 3911BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

advantage of Src, Yes1, and Fyn triple knock-out fibroblasts toelucidate a potential role of Src kinases in NPM-ALK–mediatedp130Cas phosphorylation. NPM-ALK could induce p130Cas phos-phorylation even in the absence of Src, thus demonstrating thepresence of a Src-independent pathway (Figure 4B). In addition, nodifferences in NPM-ALK–dependent p130Cas phosphorylationwere found following treatment with the Src family inhibitor PP2both in MEF NIH3T3 cells and in lymphoid cell lines (Figures4C-D), thus confirming the results obtained in Src/Yes1/Fyn tripleknock-out fibroblasts.
NPM-ALK needs the adaptor Grb2 to bind p130Cas
Since p130Cas has no SH2 domains for a direct binding withphosphoproteins,31 we looked for an adaptor protein that couldmediate the association between NPM-ALK and p130Cas. Grb2has previously been shown to bind both NPM-ALK36 and p130Cas37
and we confirmed the association between NPM-ALK and Grb2 inALK-positive lymphoid cell lines (Figure 5A). Moreover, theR86K-mutated Grb2 has been reported to disrupt the associationbetween p130Cas and Grb2.37 By using 2 dominant-negative formsof Grb2, R86K and P49L, respectively mutated in the SH2 and inthe N-terminus SH3 domain, we were able to show that Grb2 wasinvolved in the binding of NPM-ALK to p130Cas. In fact the R86KGrb2 construct was able to disrupt the interaction between NPM-ALK and p130Cas, whereas the P49L mutation had no effect,indicating that the SH2 domain of Grb2 was fundamental for theassociation between NPM-ALK and p130Cas (Figure 5B). More-over, as a result of the releasing of NPM-ALK from the complex,p130Cas was no longer phosphorylated (Figure 5C).
p130Cas is required for NPM-ALK–mediated actin remodelingand cell transformation
Regarding the morphologic features, we asked if p130Cas couldinfluence NPM-ALK–induced cytoskeleton modifications. Weshowed that the expression of NPM-ALK in NIH3T3 MEF cellsled to spindle-shaped morphology and the outgrowth of elongatedprocesses (Figure 6A-B), but both of the effects were not evident inCas�/� MEFs (Figure 6C-D). In immunofluorescence, NIH3T3MEFs expressing NPM-ALK displayed a clear loss of the actinstructure (Figure 6F) in contrast to Cas�/� fibroblasts in which the
actin filaments were still detectable (Figure 6H), thus suggesting animportant role of p130Cas in NPM-ALK–mediated actin filamentremodeling.
Next, we investigated the functional role of p130Cas in someNPM-ALK–mediated biologic effects, such as cell transformationand migration. First, we tested the colony-forming ability ofNPM-ALK–expressing cells by soft-agar assay. We infected
Figure 4. The phosphorylation of p130Cas by NPM-ALK is independent from Src tyrosine kinase activity. (A) NPM-ALK coprecipitates with Src in lymphoid cells. Totallysates from ALK-positive (Karpas, DHL, TS) and ALK-negative (CEM, Namalwa) cell lines were immunoprecipitated with anti-ALK monoclonal antibody and blotted withanti-Src polyclonal antibody. (B) The NPM-ALK–mediated phosphorylation of p130Cas is Src independent. Syf triple knock-out fibroblasts were cotransfected with Pallinop130Cas and Pallino NPM-ALK or Pallino NPM-ALKK210R as a control. Samples were collected 48 hours after transfection and total lysates were immunoprecipitated withanti-PY20 monoclonal antibody and blotted with the indicated antibodies. (C) Src inhibition does not limit p130Cas phosphorylation. MEF NIH3T3 cells were cotransfected withPallino p130Cas and Pallino NPM-ALK or Pallino NPM-ALKK210R as a control. Forty-eight hours after transfection, samples were cultivated in the presence of 30 �M PP2 for 1hour and then collected. The adaptor p130Cas phosphorylation levels were detected by immunoprecipitation with anti-p130Cas monoclonal antibody followed by blotting withanti-PY20 monoclonal antibody. (D) ALK-positive (DHL and TS) and ALK-negative (K562) cells were cultivated in the presence of 30 �M PP2 for 1 hour and then collected. Theadaptor p130Cas phosphorylation levels were detected by immunoprecipitation with anti-p130Cas monoclonal antibody followed by blotting with anti-PY20 monoclonalantibody.
Figure 5. NPM-ALK binds p130Cas through the SH2 domain of Grb2. (A)NPM-ALK binds Grb2 in lymphoid cell lines. Total lysates were immunoprecipitatedwith anti-ALK monoclonal antibody and blotted with the indicated antibodies. (B-C)The 293 T cells were transfected with Pallino NPM-ALK, Pallino p130Cas, pRK5Grb2, and the dominant-negative Grb2 constructs pRK5 P49L (mutated in the SH3domain) or pRK5 R86K (mutated in the SH2 domain) as indicated. Samples werecollected 48 hours after transfection and total lysates were immunoprecipitated withanti-Grb2 polyclonal antibody and blotted with the indicated antibodies. Grb2 R86Kwas able to disrupt the binding of p130Cas to NPM-ALK (B) as well as p130Casphosphorylation (C).
3912 AMBROGIO et al BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

NIH3T3 MEFs and Cas�/� MEFs with retroviruses expressingNPM-ALK and NPM-ALKK210R as control (Figure 6I). NPM-ALKwas efficient in transforming NIH3T3 MEFs as expected but failedto transform Cas�/� fibroblasts, thus underlining the importance ofp130Cas in mediating the transforming properties of NPM-ALK(Figure 6K).
To exclude the possibility of an intrinsic proliferative defect ofCas�/� fibroblasts, we performed a cell-cycle analysis on noncon-fluent MEFs in logarithmic growth phase and found no prolifera-tive differences between Cas�/� cells and NIH3T3 MEFs (Figure
6J). In addition, we rescued Cas�/� fibroblasts by retroviralinfection with Pallino p130Cas. In soft-agar assay the ectopichyperexpression of p130Cas was sufficient to transform Cas�/�
fibroblasts (data not shown) in accordance to data previouslydescribed.38
Finally, we asked whether the overexpression of p130Cas couldincrease the cellular migration rate. For this purpose, we infectedboth Cas�/� and Cas�/� fibroblasts with Pallino NPM-ALKretrovirus (Figure 7A), sorted them for green fluorescent protein(GFP) content, and then performed a Matrigel migration assay. The
Figure 6. The adaptor p130Cas is required for NPM-ALK–mediated transformation and actin filamentsorganization. NIH3T3 MEFs and Cas�/� MEFs wereinfected with Pallino NPM-ALK or Pallino NPM-ALKK210R
retroviruses and sorted to obtain greater than 95%GFP-positive cells. (A-B) The expression of NPM-ALK inNIH3T3 MEFs led to spindle-shape morphology and tothe outgrowth of cellular processes. NPM-ALKK210R wasused as a control. (C-D) Cas�/� cells infected with PallinoNPM-ALK did not show evident morphologic differencesin comparison with the control NPM-ALKK210R. Phase-contrast images acquired with a 40�/0.55 objective lens.(E-H) Cas�/� fibroblasts expressing NPM-ALK retain theorganization of the actin cytoskeleton. Compared withNPM-ALK NIH3T3 MEFs that undergo an evident loss ofthe actin structure (F), NPM-ALK Cas�/� fibroblasts showa phenotype resembling Cas�/� infected with the controlNPM-ALKK210R (G-H). Cells were incubated with PE-conjugated phalloidin to stain the actin filaments (red).Images were taken with the Leica confocal microscopeequipped with a 63�/1.32 oil immersion objective lens.(I) Protein expression levels were analyzes by WB asindicated. (J) Cell-cycle analysis was performed by DNAcontent evaluation on cells in logarithmic growth phase.(K) NIH3T3 and Cas�/� MEFs, infected as indicated,were plated in soft agar and cultured for 3 weeks. Thehistograms represent the average numbers of coloniesfrom the indicated cells and constructs. Data are from 1 of3 independent experiments, each including triplicates forexperimental point.
Figure 7. The p130Cas is not required for NPM-ALK–mediated migration. (A) Cas�/� and Cas�/� rescuedfibroblasts were infected with Pallino NPM-ALK retrovirusand then sorted for GFP content. Protein expressionlevels were verified by WB as indicated. (B) Cells wereplated on a Matrigel-coated insert and the number ofmigrated cells was evaluated after 48 hours. The histo-grams represent the average number of migrated cellsfrom 3 independent experiments using triplicate wells forexperimental point. (C) CEM lymphoblastoid cells wereinfected with Pallino retroviruses containing NPM-ALK orNPM-ALKK210R together with retrovirus for p130Cas.Protein expression levels were verified by WB as indi-cated. (D) The histograms represent the average num-bers of migrated cells in response to increasing concen-trations of SDF-1� as indicated. Data are from 1 of 2independent experiments. Error bars indicate standarddeviation. *Statistically significant analysis as measuredby the Student t test.
ROLE OF p130cas IN NPM-ALK–MEDIATED TRANSFORMATION 3913BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

expression of NPM-ALK was sufficient to increase cell migrationas expected, although p130Cas expression did not provide anadditional effect on cell migration (Figure 7B). Nevertheless, wealso investigated the role of p130Cas in lymphoid cells migration;we overexpressed p130Cas together with either NPM-ALK orNPM-ALKK210R in CEM cells via retroviral infection, and GFP-positive cells were enriched by cell sorting. Despite the strongexpression levels of p130Cas and NPM-ALK obtained in CEMcells (Figure 7C) in a chemotaxis assay with SDF-1�, thesynergistic effect of SDF-1� and NPM-ALK on cell migration wasnot further increased by the overexpression of p130Cas, thussuggesting that p130Cas could not be a fundamental player in thisprocess (Figure 7D).
Discussion
Many canonic steps of malignant transformation in which thefusion protein NPM-ALK is directly involved have been fairly welldescribed and include increased mitogenity, protection from apopto-sis, and growth in the absence of adhesion or growth factors.10,39,40
The molecular mechanisms underlying these processes are similarto those employed by other tyrosine kinases and rely on theactivation of signaling molecules such as PI3K, PLC-�, Stat3, andMAPK.15-17,39,41
In line with these oncogenic properties, NPM-ALK is alsocapable of changing cell morphology in a similar fashion to otherwell-known oncogenes. In fibroblasts, NPM-ALK induces anelongated and birifrangent morphology and in lymphoid cells isable to increase the size of the cells as well as to drive anaplasticfeatures.19
In the present study we started from a proteomic approach toidentify molecules involved in these effects induced by NPM-ALK. By mass spectrometry on 293 T-Rex cells expressing atetracycline-inducible form of NPM-ALK, we identified somecytoskeleton-associated proteins that precipitate or are phosphory-lated as an effect of the tyrosine kinase activity of NPM-ALK.Interestingly, similar proteins, such as beta actin and tubulin, havealso been found to be phosphorylated in anaplastic lymphoma celllines (Karpas and DHL) with a recently described method ofimmunoaffinity profiling of tyrosine phosphorylation,30 thus render-ing our approach based on inducible 293 T-Rex cells a reliablemethod. Together with structural proteins, in the MALDI analysisof precipitated bands we found p130Cas, a 130-kilodalton (kDa)phosphotyrosine (pTyr)–scaffolding protein that normally associ-ates with 2 oncoproteins, pp60v-src (v-Src) and p47gag-crk(v-Crk).35,42
The fundamental role of p130Cas in actin cytoskeleton remodel-ing and cell migration has been suggested by several lines ofevidence including its phosphorylation following integrin engage-ment and its presence in the focal adhesions that form themolecular bridges between the extracellular matrix and the actincytoskeleton.43 In agreement with these functions, Cas�/� fibro-blasts show normal focal adhesion formation but have defects incell migration.38,44 In these events, the phosphorylation of p130Casis mediated by FAK, Src, and protein tyrosine kinase 2� (Pyk2) inan integrin-dependent fashion.45,46
Here we show that p130Cas precipitates with the oncogenictyrosine kinase NPM-ALK both in 293 T-Rex cells and inhuman-derived anaplastic lymphoma cells Karpas, DHL, and TS.In addition, p130Cas is phosphorylated when NPM-ALK isexpressed and this phosphorylation is strictly dependent on the
intrinsic tyrosine kinase activity of ALK since a kinase-deadmutant is no longer able to bind and phosphorylate p130Cas.Similarly to NPM-ALK, the ATIC-ALK fusion protein and thefull-length ALK receptor phosphorylate p130Cas, thus excludingan essential role of the NPM portion of the molecule and indicatinga general mechanism most likely shared by all ALK fusionproteins. Therefore, these findings add ALK to other tyrosinekinase receptors known to phosphorylate p130Cas such as EGF,fibroblast growth factor (FGF), insulin-like growth factor 1 (IGF-1), nerve growth factor (NGF), and platelet-derived growth factor(PDGF)31 and, more interestingly, are comparable to another fusionprotein involved in hematopoietic malignancies, break point clusterregion–Abelson (Bcr-Abl), which was demonstrated to bind andphosphorylate p130Cas in Bcr-Abl–positive cell lines and insamples obtained from chronic myeloid leukemia (CML) and acutelymphoblastic leukemia (ALL) patients.47,48
The phosphorylation of p130Cas at a molecular level has beenshown in vitro to be mediated by 4 different kinases (ie, Src, Fak,Pyk2, and Abl). Since it has recently been described that Src kinasecan bind to and be activated by NPM-ALK,34 we considered thepossibility that p130Cas phosphorylation by NPM-ALK requiredSrc as intermediate kinase. However, in triple knock-out Src, Yes1,and Fyn cells NPM-ALK was still able to phosphorylate p130Casat least to some extent and p130Cas phosphorylation was stilldetectable both in MEF NIH3T3 fibroblasts and in lymphoid celllines even after the treatment with the Src-family inhibitor PP2,thus indicating the existence of a Src-independent pathway inp130Cas activation. This pathway could pass through the adaptorprotein Grb2, a molecule previously known to bind both NPM-ALK36 and p130Cas.37 Indeed, we show in this paper that in 293T-Rex cells, an SH2 Grb2 mutant, acting as a dominant-negative,impaired both the NPM-ALK binding to and the phosphorylationof p130Cas, thus revealing Grb2 as a key regulator of thisinteraction.
The role of p130Cas in ALK-mediated cytoskeleton organiza-tion was evident by studying the polymerization of the actinfilaments. Deregulated ALK kinase activity forces cells to acquire atransformed phenotype with reduced adherence to the plate and aspindle, more birifrangent shape. In addition, ALK-transformedMEFs show long processes, in a way similar to the describedneurite outgrowth induced by ALK in PC12 cells,8 indicative of adirect role of ALK in cytoskeleton organization. Indeed, ALK-transformed MEFs showed a prominent actin depolymerizationsimilar to that induced by other oncogenic fusion proteins such asBcr-Abl.49 Interestingly, these effects were clearly impaired inCas�/� MEFs, indicating an essential role of p130Cas in ALK-mediated actin depolymerization. It still remains to be clarified ifp130Cas could be involved also in the cellular shaping ofNPM-ALK–expressing lymphoid cells.
Besides the functions on organizing the cell cytoskeleton,p130Cas is involved in broader oncogenic effects that include cellproliferation and apoptosis, indicating p130Cas as a major player inthe complex network that is connecting cell adhesion and migrationto cell growth and survival.31 Indeed, p130Cas overexpression issufficient to induce an anchorage-independent growth in cells,38,50
possibly through the activation of the c-Jun N-terminal kinase(JNK)/c-jun pathway,51 as well as protection from apoptosis.31 Inthis study we tested the role of p130Cas in NPM-ALK–mediatedtransformation, showing that Cas�/� fibroblasts can no longer betransformed by NPM-ALK. These effects on anchorage-indepen-dent growth could derive from a combination of cytoskeleton
3914 AMBROGIO et al BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

modifications and proliferative signals both provided by NPM-ALK in wt MEFs but impaired in the absence of p130Cas. Overallthe data are suggesting that NPM-ALK could control a Src-dependent and a Src-independent transformation pathway that bothuse p130Cas as pivotal adaptor. The Src-independent pathway mayrequire Grb2 as key adaptor molecule, as previously discussed. Inthis scenario, it is possible to further speculate that NPM-ALKactivates both Src and p130Cas, where phosphorylated p130Casmay further enhance Src activity in a sort of activating loop similarto what has already been described in other cancers.52
In this paper, we also describe a novel function of NPM-ALK(ie, its capability of increasing the migration rate of T lympho-cytes). CEM lymphoblastoid cells have previously been proven tomigrate in a gradient of SDF-1�, which is the ligand for CXCR4, achemokine receptor expressed in T cells.29 Upon retroviral expres-sion of NPM-ALK, CEM cells almost doubled their migration rate,indicating that NPM-ALK tyrosine kinase activity influences theactivation state of molecules involved in this process. However, in
contrast to transformation, we could not find a clear effect ofp130Cas overexpression in NPM-ALK–induced migration. Regard-ing lymphoid cells, this could be explained by considering that in Tlymphocytes a major role in beta1 integrin– and T-cell receptor(TCR)–mediated migration has been shown for Cas-L, a p105-kDaprotein that is a member of the Cas family selectively expressed inlymphocytes.32,33 Indeed, we showed that NPM-ALK is capable ofphosphorylating Cas-L in lymphocytes. It is therefore possible thatNPM-ALK in lymphocytes may promote migration through the activa-tion of different molecules than p130Cas. Further studies in lympho-cytes are needed to solve this issue.
In conclusion, in this study we show p130Cas as a noveldownstream interactor of NPM-ALK capable of modulating itstransforming properties. This role could involve not only thep130Cas effects on cytoskeleton remodeling but also its involve-ment in important proliferative and survival pathways. Furtherstudies are required to unravel this differential requirement of ALKfor p130Cas or its family members in different cell subtypes.
References
1. Morris SW, Kirstein MN, Valentine MB, et al. Fu-sion of a kinase gene, ALK, to a nucleolar proteingene, NPM, in non-Hodgkin’s lymphoma. Sci-ence. 1994;263:1281-1284.
2. Borer RA, Lehner CF, Eppenberger HM, Nigg EA.Major nucleolar proteins shuttle between nucleusand cytoplasm. Cell. 1989;56:379-390.
3. Okuda M. The role of nucleophosmin in centro-some duplication. Oncogene. 2002;21:6170-6174.
4. Kurki S, Peltonen K, Latonen L, et al. Nucleolarprotein NPM interacts with HDM2 and protectstumor suppressor protein p53 from HDM2-medi-ated degradation. Cancer Cell. 2004;5:465-475.
5. Chan PK, Chan FY. Nucleophosmin/B23 (NPM)oligomer is a major and stable entity in HeLacells. Biochim Biophys Acta. 1995;1262:37-42.
6. Morris SW, Naeve C, Mathew P, et al. ALK, thechromosome 2 gene locus altered by the t(2;5) innon-Hodgkin’s lymphoma, encodes a novel neu-ral receptor tyrosine kinase that is highly relatedto leukocyte tyrosine kinase (LTK). Oncogene.1997;14:2175-2188.
7. Iwahara T, Fujimoto J, Wen D, et al. Molecularcharacterization of ALK, a receptor tyrosine ki-nase expressed specifically in the nervous sys-tem. Oncogene. 1997;14:439-449.
8. Souttou B, Carvalho NB, Raulais D, Vigny M. Ac-tivation of anaplastic lymphoma kinase receptortyrosine kinase induces neuronal differentiationthrough the mitogen-activated protein kinasepathway. J Biol Chem. 2001;276:9526-9531.
9. Liao EH, Hung W, Abrams B, Zhen M. An SCF-like ubiquitin ligase complex that controls presyn-aptic differentiation. Nature. 2004;430:345-350.
10. Wellmann A, Doseeva V, Butscher W, et al. Theactivated anaplastic lymphoma kinase increasescellular proliferation and oncogene up-regulationin rat 1a fibroblasts. FASEB J. 1997;11:965-972.
11. Ma Z, Cools J, Marynen P, et al. Inv(2)(p23q35) inanaplastic large-cell lymphoma induces constitu-tive anaplastic lymphoma kinase (ALK) tyrosinekinase activation by fusion to ATIC, an enzymeinvolved in purine nucleotide biosynthesis. Blood.2000;95:2144-2149.
12. Kuefer MU, Look AT, Pulford K, et al. Retrovirus-mediated gene transfer of NPM-ALK causes lym-phoid malignancy in mice. Blood. 1997;90:2901-2910.
13. Chiarle R, Gong JZ, Guasparri I, et al. NPM-ALKtransgenic mice spontaneously develop T-celllymphomas and plasma cell tumors. Blood. 2003;101:1919-1927.
14. Fujimoto J, Shiota M, Iwahara T, et al. Character-
ization of the transforming activity of p80, a hy-perphosphorylated protein in a Ki-1 lymphomacell line with chromosomal translocation t(2;5).Proc Natl Acad Sci U S A. 1996;93:4181-4186.
15. Bai RY, Dieter P, Peschel C, Morris SW, DuysterJ. Nucleophosmin-anaplastic lymphoma kinase oflarge-cell anaplastic lymphoma is a constitutivelyactive tyrosine kinase that utilizes phospholipaseC-gamma to mediate its mitogenicity. Mol CellBiol. 1998;18:6951-6961.
16. Zamo A, Chiarle R, Piva R, et al. Anaplastic lym-phoma kinase (ALK) activates Stat3 and protectshaematopoietic cells from cell death. Oncogene.2002;21:1038-1047.
17. Bai RY, Ouyang T, Miething C, Morris SW, Pe-schel C, Duyster J. Nucleophosmin-anaplasticlymphoma kinase associated with anaplasticlarge-cell lymphoma activates the phosphatidyl-inositol 3-kinase/Akt antiapoptotic signalling path-way. Blood. 2000;96:4319-4327.
18. Gu TL, Tothova Z, Scheijen B, Griffin JD, GillilandDG, Sternberg DW. NPM-ALK fusion kinase ofanaplastic large-cell lymphoma regulates survivaland proliferative signalling through modulation ofFOXO3a. Blood. 2004;103:4622-4629.
19. Horie R, Watanabe M, Ishida T, et al. The NPM-ALK oncoprotein abrogates CD30 signalling andconstitutive NF-kappaB activation in anaplasticlarge cell lymphoma. Cancer Cell. 2004;5:353-364.
20. Lee HH, Norris A, Weiss JB, Frasch M. Jelly bellyprotein activates the receptor tyrosine kinase Alkto specify visceral muscle pioneers. Nature.2003;425:507-512.
21. Englund C, Loren CE, Grabbe C, et al. Jeb sig-nals through the Alk receptor tyrosine kinase todrive visceral muscle fusion. Nature. 2003;425:512-516.
22. Zettervall CJ, Anderl I, Williams MJ, et al. A di-rected screen for genes involved in Drosophilablood cell activation. Proc Natl Acad Sci U S A.2004;101:14192-14197.
23. Wiznerowicz M, Trono D. Conditional suppres-sion of cellular genes: lentivirus vector-mediateddrug-inducible RNA interference. J Virol. 2003;77:8957-8961.
24. Matrix Science. Mascot: Peptide Mass Finger-print. http://www.matrixscience.com/search_form_select.html. Accessed June 5, 2004.
25. National Center for Biotechnology Information.Nonredundant protein database. Available athttp://www.ncbi.nlm.nih.gov/BLAST. AccessedJune 10, 2004.
26. Grignani F, Kinsella T, Mencarelli A, et al. High-
efficiency gene transfer and selection of humanhaematopoietic progenitor cells with a hybridEBV/retroviral vector expressing the green fluo-rescence protein. Cancer Res. 1998;58:14-19.
27. Turner CE. Paxillin and focal adhesion signalling.Nat Cell Biol. 2000;2:E231-E236.
28. Turner CE. Paxillin interactions. J Cell Sci. 2000;113(Pt 23):4139-4140.
29. Rabin RL, Park MK, Liao F, Swofford R, StephanyD, Farber JM. Chemokine receptor responses onT cells are achieved through regulation of bothreceptor expression and signalling. J Immunol.1999;162:3840-3850.
30. Rush J, Moritz A, Lee KA, et al. Immunoaffinityprofiling of tyrosine phosphorylation in cancercells. Nat Biotechnol. 2005;23:94-101.
31. Bouton AH, Riggins RB, Bruce-Staskal PJ. Func-tions of the adapter protein Cas: signal conver-gence and the determination of cellular re-sponses. Oncogene. 2001;20:6448-6458.
32. Ohashi Y, Iwata S, Kamiguchi K, Morimoto C. Ty-rosine phosphorylation of Crk-associated sub-strate lymphocyte-type is a critical element inTCR- and beta 1 integrin-induced T lymphocytemigration. J Immunol. 1999;163:3727-3734.
33. van Seventer GA, Salmen HJ, Law SF, et al. Fo-cal adhesion kinase regulates beta1 integrin-de-pendent T cell migration through an HEF1 effec-tor pathway. Eur J Immunol. 2001;31:1417-1427.
34. Cussac D, Greenland C, Roche S, et al. Nucleo-phosmin-anaplastic lymphoma kinase of anaplas-tic large-cell lymphoma recruits, activates, anduses pp60c-src to mediate its mitogenicity. Blood.2004;103:1464-1471.
35. Matsuda M, Mayer BJ, Fukui Y, Hanafusa H.Binding of transforming protein, P47gag-crk, to abroad range of phosphotyrosine-containing pro-teins. Science. 1990;248:1537-1539.
36. Crockett DK, Lin Z, Elenitoba-Johnson KS, LimMS. Identification of NPM-ALK interacting pro-teins by tandem mass spectrometry. Oncogene.2004;23:2617-2629.
37. Hakak Y, Martin GS. Cas mediates transcriptionalactivation of the serum response element by Src.Mol Cell Biol. 1999;19:6953-6962.
38. Honda H, Nakamoto T, Sakai R, Hirai H.p130(Cas), an assembling molecule of actin fila-ments, promotes cell movement, cell migration,and cell spreading in fibroblasts. Biochem Bio-phys Res Commun. 1999;262:25-30.
39. Slupianek A, Nieborowska-Skorska M, Hoser G,et al. Role of phosphatidylinositol 3-kinase-Aktpathway in nucleophosmin/anaplastic lymphoma
ROLE OF p130cas IN NPM-ALK–MEDIATED TRANSFORMATION 3915BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

kinase-mediated lymphomagenesis. Cancer Res.2001;61:2194-2199.
40. Armstrong F, Duplantier MM, Trempat P, et al.Differential effects of X-ALK fusion proteins onproliferation, transformation, and invasion proper-ties of NIH3T3 cells. Oncogene. 2004;23:6071-6082.
41. Turner SD, Tooze R, Maclennan K, AlexanderDR. Vav-promoter regulated oncogenic fusionprotein NPM-ALK in transgenic mice causes B-cell lymphomas with hyperactive Jun kinase. On-cogene. 2003;22:7750-7761.
42. Reynolds AB, Roesel DJ, Kanner SB, ParsonsJT. Transformation-specific tyrosine phosphoryla-tion of a novel cellular protein in chicken cells ex-pressing oncogenic variants of the avian cellularsrc gene. Mol Cell Biol. 1989;9:629-638.
43. Harte MT, Hildebrand JD, Burnham MR, BoutonAH, Parsons JT. p130Cas, a substrate associatedwith v-Src and v-Crk, localizes to focal adhesionsand binds to focal adhesion kinase. J Biol Chem.1996;271:13649-13655.
44. Honda H, Oda H, Nakamoto T, et al. Cardiovas-
cular anomaly, impaired actin bundling andresistance to Src-induced transformation inmice lacking p130Cas. Nat Genet. 1998;19:361-365.
45. Schlaepfer DD, Hunter T. Focal adhesion ki-nase overexpression enhances ras-dependentintegrin signalling to ERK2/mitogen-activatedprotein kinase through interactions with andactivation of c-Src. J Biol Chem. 1997;272:13189-13195.
46. Schlaepfer DD, Broome MA, Hunter T. Fibronec-tin-stimulated signalling from a focal adhesionkinase-c-Src complex: involvement of the Grb2,p130cas, and Nck adaptor proteins. Mol Cell Biol.1997;17:1702-1713.
47. de Jong R, van Wijk A, Haataja L, Heisterkamp N,Groffen J. BCR/ABL-induced leukemogenesiscauses phosphorylation of Hef1 and its associa-tion with Crkl. J Biol Chem. 1997;272:32649-32655.
48. Salgia R, Pisick E, Sattler M, et al. p130CASforms a signalling complex with the adapter pro-tein CRKL in haematopoietic cells transformed by
the BCR/ABL oncogene. J Biol Chem. 1996;271:25198-25203.
49. Cheng K, Kurzrock R, Qiu X, et al. Reduced focal ad-hesion kinase and paxillin phosphorylation in BCR-ABL-transfected cells. Cancer. 2002;95:440-450.
50. Burnham MR, Bruce-Staskal PJ, Harte MT, et al.Regulation of c-SRC activity and function by theadapter protein CAS. Mol Cell Biol. 2000;20:5865-5878.
51. Oktay M, Wary KK, Dans M, Birge RB, Giancotti FG.Integrin-mediated activation of focal adhesion kinaseis required for signalling to Jun NH2-terminal kinaseand progression through the G1 phase of the cellcycle. J Cell Biol. 1999;145:1461-1469.
52. Burnham MR, Harte MT, Richardson A, ParsonsJT, Bouton AH. The identification of p130cas-binding proteins and their role in cellular transfor-mation. Oncogene. 1996;12:2467-2472.
53. Piva R, Chiarle R, Manzanna A, et al. Ablation ofoncogenic ALK is a viable therapeutic approachfor anaplastic large-cell lymphomas. Blood. 2005;in press.
3916 AMBROGIO et al BLOOD, 1 DECEMBER 2005 � VOLUME 106, NUMBER 12
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

Errata
Ambrogio et al. p130Cas mediates the transforming properties of the anaplastic lymphomakinase. Blood. 2005;106(12):3907-3916.
On page 3913 in the December 1, 2005, issue, there is an error inFigure 7A. A wrong panel was inserted corresponding to the WB:P-Tyr (Cas) lane. The correct Figure 7 is shown.
Nolte-’t Hoen et al. Activated T cells recruit exosomes secreted by dendritic cells via LFA-1.Blood. 2009;113(9):1977-1981.
On page 1979 in the February 26, 2009, issue, there is an error inFigure 2E. The unit of the molar concentration of MnCl2, indicated
at the right-hand side of the bar graph, should read “MnCl2 (mM)”(millimolar) and not “MnCl2 (�M)” (micromolar).
Figure 7. The p130Cas is not required for NPM-ALK–mediated migration. (A) Cas�/� and Cas�/� rescued fibroblasts were infected with Pallino NPM-ALK retrovirus andthen sorted for GFP content. Protein expression levels were verified by WB as indicated. (B) Cells were plated on a Matrigel-coated insert and the number of migrated cells wasevaluated after 48 hours. The histograms represent the average number of migrated cells from 3 independent experiments using triplicate wells for experimental point.(C) CEM lymphoblastoid cells were infected with Pallino retroviruses containing NPM-ALK or NPM-ALKK210R together with retrovirus for p130Cas. Protein expression levelswere verified by WB as indicated. (D) The histograms represent the average numbers of migrated cells in response to increasing concentrations of SDF-1� as indicated. Dataare from 1 of 2 independent experiments. Error bars indicate standard deviation. *Statistically significant analysis as measured by the Student t test.
2851BLOOD, 24 SEPTEMBER 2009 � VOLUME 114, NUMBER 13
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

Tai et al. CS1 promotes multiple myeloma cell adhesion, clonogenic growth, andtumorigenicity via c-maf–mediated interactions with bone marrow stromal cells. Blood.2009;113(18):4309-4318.
On page 4318 in the April 30, 2009, issue, a volume number wasomitted from one of the references. The correct Reference 30 isshown. The error was corrected in the online version, which nowdiffers from the print version.
30. Eychene A, Rocques N, Pouponnot C. A new MAFia in cancer. Nat Rev Cancer.2008;8:683-693.
2852 ERRATUM BLOOD, 24 SEPTEMBER 2009 � VOLUME 114, NUMBER 13
For personal use only. by guest on June 4, 2013. bloodjournal.hematologylibrary.orgFrom
Related Documents