Oxidative damage–induced inflammation initiates age-related macular degeneration Joe G Hollyfield 1 , Vera L Bonilha 1 , Mary E Rayborn 1 , Xiaoping Yang 1 , Karen G Shadrach 1 , Liang Lu 2 , Rafael L Ufret 1 , Robert G Salomon 2 , and Victor L Perez 1 1 Cole Eye Institute (i31), Department of Ophthalmology, Cleveland Clinic Lerner College of Medicine, Cleveland, Ohio 44195, USA 2 Department of Chemistry, Case Western Reserve University, Cleveland, Ohio 44106, USA Abstract Oxidative damage and inflammation are postulated to be involved in age-related macular degeneration (AMD). However, the molecular signal(s) linking oxidation to inflammation in this late-onset disease is unknown. Here we describe AMD-like lesions in mice after immunization with mouse serum albumin adducted with carboxyethylpyrrole, a unique oxidation fragment of docosahexaenoic acid that has previously been found adducting proteins in drusen from AMD donor eye tissues 1 and in plasma samples 2 from individuals with AMD. Immunized mice develop antibodies to this hapten, fix complement component-3 in Bruch's membrane, accumulate drusen below the retinal pigment epithelium during aging, and develop lesions in the retinal pigment epithelium mimicking geographic atrophy, the blinding end-stage condition characteristic of the dry form of AMD. We hypothesize that these mice are sensitized to the generation of carboxyethylpyrrole adducts in the outer retina, where docosahexaenoic acid is abundant and conditions for oxidative damage are permissive. This new model provides a platform for dissecting the molecular pathology of oxidative damage in the outer retina and the immune response contributing to AMD. AMD is the most common cause of legal blindness in elderly individuals of industrialized countries 3,4 . Clinicians have long recognized that debris (termed drusen) below the retinal pigment epithetlium (RPE) in the macula is a risk factor for AMD. The presence of complement factor proteins in drusen in AMD eyes 1,5–7 and genetic variation in several complement factor genes in individuals with AMD 8–13 implicate inflammation as an important component in this disease. However, little is known about the signal(s) from the outer retina that initiates the immune system's involvement in AMD. As a potential initiating signal we evaluated carboxyethylpyrrole (CEP; Fig. 1a), an adduct that forms from an oxidation fragment of docosahexaenoic acid (DHA) 2 . DHA, the most oxidizable Correspondence should be addressed to J.G.H. ([email protected]) or V.L.P. ([email protected]). Note: Supplementary information is available on the Nature Medicine website. Author Contributions J.G.H. and V.L.P. designed and initiated the experiments. L.L. and R.G.S. prepared the CEP-MSA and CEP- BSA. R.L.U., V.L.P., K.G.S. and X.Y. immunized the mice. K.G.S. and X.Y. performed the protein chemistry and ELISA assays, as well as managing the day-to-day maintenance of the mice. M.E.R. and J.G.H. performed all of the histological and electron microscopic analysis. V.L.B. performed the confocal microscopy. J.G.H. analyzed all of the data and wrote the manuscript. R.G.S., R.L.U. and V.L.P. made critical comments and suggestions for revisions of the manuscript in response to the reviewers. Competing Interests Statement The authors declare competing financial interests: details accompany the full-text HTML version of the paper at www.nature.com/naturemedicine/. Published online at http://www.nature.com/naturemedicine Reprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions NIH Public Access Author Manuscript Nat Med. Author manuscript; available in PMC 2009 September 22. Published in final edited form as: Nat Med. 2008 February ; 14(2): 194–198. doi:10.1038/nm1709. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Oxidative damage–induced inflammation initiates age-relatedmacular degeneration
Joe G Hollyfield1, Vera L Bonilha1, Mary E Rayborn1, Xiaoping Yang1, Karen G Shadrach1,Liang Lu2, Rafael L Ufret1, Robert G Salomon2, and Victor L Perez11Cole Eye Institute (i31), Department of Ophthalmology, Cleveland Clinic Lerner College ofMedicine, Cleveland, Ohio 44195, USA2Department of Chemistry, Case Western Reserve University, Cleveland, Ohio 44106, USA
AbstractOxidative damage and inflammation are postulated to be involved in age-related maculardegeneration (AMD). However, the molecular signal(s) linking oxidation to inflammation in thislate-onset disease is unknown. Here we describe AMD-like lesions in mice after immunization withmouse serum albumin adducted with carboxyethylpyrrole, a unique oxidation fragment ofdocosahexaenoic acid that has previously been found adducting proteins in drusen from AMD donoreye tissues1 and in plasma samples2 from individuals with AMD. Immunized mice developantibodies to this hapten, fix complement component-3 in Bruch's membrane, accumulate drusenbelow the retinal pigment epithelium during aging, and develop lesions in the retinal pigmentepithelium mimicking geographic atrophy, the blinding end-stage condition characteristic of the dryform of AMD. We hypothesize that these mice are sensitized to the generation of carboxyethylpyrroleadducts in the outer retina, where docosahexaenoic acid is abundant and conditions for oxidativedamage are permissive. This new model provides a platform for dissecting the molecular pathologyof oxidative damage in the outer retina and the immune response contributing to AMD.
AMD is the most common cause of legal blindness in elderly individuals of industrializedcountries3,4. Clinicians have long recognized that debris (termed drusen) below the retinalpigment epithetlium (RPE) in the macula is a risk factor for AMD. The presence of complementfactor proteins in drusen in AMD eyes1,5–7 and genetic variation in several complement factorgenes in individuals with AMD8–13 implicate inflammation as an important component in thisdisease. However, little is known about the signal(s) from the outer retina that initiates theimmune system's involvement in AMD.
As a potential initiating signal we evaluated carboxyethylpyrrole (CEP; Fig. 1a), an adduct thatforms from an oxidation fragment of docosahexaenoic acid (DHA)2. DHA, the most oxidizable
Correspondence should be addressed to J.G.H. ([email protected]) or V.L.P. ([email protected]).Note: Supplementary information is available on the Nature Medicine website.Author Contributions J.G.H. and V.L.P. designed and initiated the experiments. L.L. and R.G.S. prepared the CEP-MSA and CEP-BSA. R.L.U., V.L.P., K.G.S. and X.Y. immunized the mice. K.G.S. and X.Y. performed the protein chemistry and ELISA assays, aswell as managing the day-to-day maintenance of the mice. M.E.R. and J.G.H. performed all of the histological and electron microscopicanalysis. V.L.B. performed the confocal microscopy. J.G.H. analyzed all of the data and wrote the manuscript. R.G.S., R.L.U. and V.L.P.made critical comments and suggestions for revisions of the manuscript in response to the reviewers.Competing Interests Statement The authors declare competing financial interests: details accompany the full-text HTML version ofthe paper at www.nature.com/naturemedicine/.Published online at http://www.nature.com/naturemedicineReprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions
NIH Public AccessAuthor ManuscriptNat Med. Author manuscript; available in PMC 2009 September 22.
Published in final edited form as:Nat Med. 2008 February ; 14(2): 194–198. doi:10.1038/nm1709.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of all long-chain polyunsaturated fatty acids, is abundant in the outer retina, where high oxygentension and light provide a permissive environment for oxidation14–16. CEP forms when thealdehyde group on a newly formed seven-carbon oxidation fragment of DHA covalentlyinteracts with an ε-lysyl amino group in a tissue protein1,2. AMD donor eyes contain moreCEP-modified proteins in the outer retina than are present in age-matched controls1, and CEP-adducted proteins are also more abundant in AMD plasma than in control samples2. CEPautoantibodies are also present in plasma and are more abundant in AMD than in controls2.This oxidation-generated hapten is noteworthy because of the long-recognized association ofAMD with oxidative damage17–21.
Because DHA is concentrated in RPE and photoreceptor cells14–16, we reasoned that thesetissues are a probable source of CEP adducts during aging. Because serum albumin is one ofthe major proteins modified with CEP in people with AMD2, we immunized mice with CEP-modified mouse serum albumin (CEP-MSA) in an attempt to raise the level of sensitivity toendogenously generated CEP (Fig. 1b). Our hypothesis was that immunized mice wouldgenerate a stronger immune response to CEP adducts, making the outer retina more vulnerableto immune-mediated damage.
We immunized mice with CEP-MSA or MSA controls in complete Freund's adjuvant (CFA)at day 0 and followed this with a challenge at day 10 in incomplete Freund's adjuvant (IFA).To generate a strong immunological response and confirm that CEP-MSA is immunogenic, agroup of ‘short-term’ experiments were performed in which animals received threeimmunizations during a 2–3–month period, with the final immunization given 10 d before themice were killed. To assess the effect of aging on the CEP-MSA immune response, a secondgroup of ‘long-term’ experiments were performed in which mice were treated with only thefirst immunization described above and maintained for 12–14 months before they were killed.(Mice used are listed in Supplementary Table 1 online.)
Titers of antibody to CEP in the short-term recovery mice were six to eight times higher in theCEP-MSA–immunized mice compared to naive and control mice immunized with MSA orCFA (Fig. 1c). CEP-specific antibody was not detected in Rag–deficient mice that lack matureT and B cells. Antibody titers were similar in the long-term recovery mice, with CEP-specificantibody titers higher in the CEP-MSA–immunized mice than in the control mice (Fig. 1d).These data show that CEP-MSA can induce an antibody-mediated immune response.
Histology of the CEP-MSA eyes revealed multiple RPE changes, including vesiculation andswelling of individual or multiple adjacent cells (Fig. 2a–c); cell lysis (Fig. 2b,c); pyknosis, asevidenced by intense toluidine blue staining (Fig. 2b); and the presence of monocytes in theinterphotoreceptor matrix (Fig. 2d,e). Some of the invading cells expressed the macrophagemarker F4/80 (Fig. 2e). RPE expanses with up to 14 contiguous swollen cells that appeared tobe undergoing lysis were common (Fig. 2f). In some eyes, the RPE was missing in focal areasand the overlying photoreceptors were greatly swollen (Fig. 2g and Supplementary Figs. 1-4).
The degree of pathology in the short-term tissues was evaluated in three or four sections pereye from the dorsal to the ventral retinal margin. Each sample was placed in one of the followingcategories based on the average number of lesions present per section: normal, minorpathology, moderate pathology or severe pathology. Two of the six naive mice studied had oneand three lysed RPE cells, respectively, and one of these also contained a single pyknotic RPEcell. Because these were naive mice that had not been immunized, these lesions were considerednormal background variability (Supplementary Fig. 1). When four or fewer lesions were found,the tissue was scored as normal. Samples in the minor pathology category contained five toten separate areas of RPE or outer retinal pathology in each section studied (SupplementaryFig. 2). Samples in the moderate pathology category contained 11–15 separate sites of RPE or
Hollyfield et al. Page 2
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

outer retinal pathology (Supplementary Fig. 3). Mice in the severe pathology category had 16or more sites of RPE pathology in each section. Four eyes in this group had lesions in the RPEthat were 20–70 μm long (Supplementary Fig. 4). We then plotted the degree of pathologyagainst the mean antibody titer of the mice in these groupings (Fig. 2h). These data show aclose relationship between the CEP-specific antibody titer and the severity of outer retinapathology.
C3d is a degradation product of C3b, a key complement protein required for the generation ofthe C3 and C5 convertases in the classical, lectin and alternate pathways22. We evaluated short-term tissues for C3d immunolocalization (Fig. 3a–h). C3d was observed in Bruch's membranebelow the RPE in the mice receiving CEP-MSA immunization (Fig. 3b). Some of the CEP-MSA tissues contained lysed RPE cells that could be labeled with an antibody to C3d (Fig.3c), suggesting that complement may be fundamentally involved in the lysis of these cells.Although the C3d immunofluorescence was somewhat patchy in eyes from CEP-MSA–immunized mice, it was not observed in control mice (Fig. 3d,e), except for minor fluorescenceoccasionally seen in the MSA–treated mice (Fig. 3d). CEP-MSA–immunized mice showed tentimes more immunofluorescence in Bruch's membrane than did control mice (Fig. 3f–h).
Examination of the fundus from the long-term protocol mice revealed patchy, reticular changesnot present in controls (Supplementary Fig. 5 online). Histology of 15 eyes from the long-termprotocol revealed an accumulation of sub-RPE deposits in the CEP-MSA-immunized mice(compare Fig. 4a to Fig. 4b and Fig. 4c to Fig. 4d). These deposits stained less intensely thandid the RPE cytoplasm, forming a near continuous band of variable thickness (2–4 μm) belowthe RPE (Supplementary Fig. 6 online). Choroidal neovascularization was not observed. Sub-RPE material did not develop in controls, although increased thickening of RPE basalinfoldings with aging was evident.
Sub-RPE deposits were measured with an image-processing algorithm that recognizes Bruch'smembrane and the basal infoldings and sub-RPE deposits in digitized images (Figs. 4c,d). Wemeasured the area occupied by basal infoldings in short-term recovery mice (Fig. 4e) and thearea of sub-RPE deposits plus basal infoldings in long-term recovery mice (Fig. 4f). Short-term recovery tissues had similar basal infolding dimensions (Fig. 4e). All eyes in the long-term protocol showed greater thickening below the RPE than was measured in the short-termtissues, except for the Rag-deficient mice. This probably reflects age-related changes.However, the CEP-MSA–immunized mice showed greater accumulation of basal deposits thanwith any other treatment (Fig. 4f).
In electron micrographs, the Bruch's membrane of long-term naive mice measured 0.54 ± 0.06μm (mean ± s.d.) thick, and RPE basal infoldings had a mean height of 1.19 ± 0.12 μm (Fig.4g). In contrast, Bruch's membrane in CEP-MSA tissues measured 1.71 ± 0.05 μm. Basalinfoldings were evident in CEP-MSA samples, but these profiles surrounded extracellularflocculent material that expanded this sub-RPE compartment to a thickness of 4.21 ± 1.14 μm(Fig. 4h). Similar sub-RPE debris is reported in mice subjected to dietary, lighting and/orgenetic manipulations23–26.
Our studies demonstrate that mice mount an antibody-mediated response to CEP-MSA. Theimmune system responds by depositing complement below the RPE, as evidenced by thelocalization of C3d in Bruch's membrane and the development of lytic changes in RPE cells.An intact immune system is required for this process, because CEP-MSA–immunized Rag-deficient mice, which are missing mature T cells and B cells27, showed none of the changesobserved in normal mice.
Although macrophages were found near some RPE lesions, it is unlikely that they initiate thepathology observed, as many lesions occurred in the absence of these cells. Macrophage
Hollyfield et al. Page 3
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

movement into this compartment may be due to the release of cytokines from lysed cells.Indeed, melanin-containing macrophages were observed, suggesting that there is debrisremoval after RPE lysis (Fig. 2d). Aging mice deficient in the macrophage chemokine Ccl2 orits receptor Ccr2 also show features similar to those in AMD23, suggesting a role formacrophages in maintaining the outer retina.
To our knowledge, this is the first study showing that immunization with a hapten generatedby oxidative damage to the DHA28 present in the drusen1 and plasma from AMD-affectedindividuals2 is sufficient to produce AMD-like lesions in mice. This model provides a newresource for understanding the early changes in the outer retina in AMD, as well as the diseaseprogression in mice with mutations or polymorphisms in complement pathway genes that arelinked to AMD in humans.
MethodsMice
We used C57BL/6 (Jackson Labs) and BALB/c (Taconic Labs) mice of both sexes at 2–3months of age. The Rag-deficient mice were on a C57BL/6 background. Protocols used herewere approved by the Institutional Animal Care and Use Committee at the Cleveland Clinic.
AntigensWe prepared the CEP-MSA and CEP-BSA from commercially available mouse serum albuminand bovine serum albumin (Sigma), which we converted to CEP-modified MSA and BSAfollowing published procedures28.
ImmunizationWe used standard mouse immunization protocols29. We anesthetized mice with ketamine-xylazine in PBS (80–90 mg/kg ketamine, 2–10 mg/ml xylazine). We used 200 μg of CEP-MSAin CFA or IFA (Difco Labs) for initial and all booster doses. We used the following controlmice for comparisons with CEP-MSA immunizations: mice immunized with non-adductedMSA (200 μg per inoculum), mice injected with CFA and subsequently boosted with IFAwithout any antigen, and age-matched naive mice. We also used Rag-deficient mice foridentical immunization protocols.
Carboxyethylpyrrole-specific antibody assayWe performed direct ELISA for the detection of antibody to CEP in 96-well plates coated withCEP-BSA (100 μl/well) at a 1:1,000 dilution in PBS and incubated at 37 °C for 1 h, using 1%BSA solution (Sigma) as a blank control. We used CEP-BSA as the coating agent(Supplementary Fig. 7 online). We washed the plate with PBS three times (300 μl/well) andblocked with 1% ovalbumin in PBS (Sigma) and then incubated at 37 °C for 1 h. We thenwashed the plate with 0.1% ovalbumin and 0.05% Tween-20 before loading the standards andmouse serum samples (at 1:10, 1:100 and 1:1,000 dilutions) and incubating them at 22 °C ona shaker for 1 h. We applied the alkaline phosphatase–conjugated secondary antibody to mouseIgG (Sigma) at a 1:2,000 dilution at 22 °C for 1 h, then rinsed and incubated the plate with Fastp-nitrophenyl phosphate tablets (Sigma). We measured absorbance (at 405 nm, using 655 nmas a reference) on a Spectra Max Plus (Molecular Devices) and defined the titer as the ratio ofserum binding to antigen versus serum binding to BSA. The antibody used on this western blotwas described previously2.
Hollyfield et al. Page 4
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fundus photographyWe examined the mouse fundus at various ages and post-immunization times. We anesthetizedand dilated the mice as described above. We used a handheld Kowa Genesis-D digital retinalcamera (Kowa Optimed) and a 90-mm intermediate lens.
HistologyWe prepared one eye from each mouse for histopathology and electron microscopy. We fixedthe eye in 2% glutaraldehyde and 2% formaldehyde (freshly prepared from paraformaldehyde)in 0.1 M cacodylate buffer, pH 7.2 at 4 °C overnight, then post-fixed it in 1% osmium tetroxide,dehydrated it in graded ethanol, and then in propylene oxide, and then transferred it to a plasticresin mixture containing Polybed 812 (Polysciences) and Araldite 502 (Polysciences) withpolymerizer. After polymerization, we cut five to ten 1-μm-thick sections, mounted them onmicroscope slides and stained them with toluidine blue.
Image analysisWe photographed retinal sections with a Zeiss Axiophot microscope equipped with aHamamatsu digital camera using a 63× oil-immersion lens. For quantitative analysis of Bruch'smembrane and sub-RPE boundaries, we took retinal images from three sections in the fundusof each of three eyes per treatment group, and we then batch-processed them with customizedmacros and algorithms generated for Image-Pro Plus 6.1 (Media Cybernetics).
ImmunocytochemistryWe froze one eye from each mouse in optimal cutting temperature medium forimmunocytochemistry. We prepared 7-μm cryosections through the center of each eye on acryostat HM 505E (Microm). For C3d localization, we used a polyclonal rabbit antibody tohuman C3d (Dako) followed by the secondary antibody (FITC-conjugated swine antibody torabbit immunoglobulin, Dako). We mounted the samples onto slides with Vectashield andexamined them for fluorescence. For F4/80 localization, we used rat antibody to mouse F4/80antigen (Serotec), followed by secondary antibody (Alexa Fluor 488–conjugated goat antibodyto rabbit IgG, Molecular Probes). We mounted the sections with Vectashield and evaluatedthem with a Zeiss Axiophot microscope equipped with epifluorescence illumination. Themounting medium contained either propidium iodide or DAPI for nuclear counterstaining. Weperformed the final imaging on a Leica TCS-SP (Exton) confocal microscope.
To compare fluorescence intensities in the C3d immunocytochemistry, we acquired a series of1.0-μm sections from the full retinal expanse. In the Leica confocal software, we saved thepinhole channel gain and offset as a macro and used it to collect all the images. We performedquantitative analysis using the stack profile function. We generated an area template with adimension of 4,512 μm2 (similar to that shown in Fig. 3f,g). We moved the template onto eachimage and positioned it so that fluorescence levels in equal areas along Bruch's membranecould be measured. We generated a graphic with the mean fluorescence intensity in each sectionof the stack. We defined ten areas from each section and measured sections from three or foureyes per treatment.
Electron microscopyWe thin-sectioned selected blocks on an RMC MT-XL ultramicrotome, placed the sections onnickel grids, stained them with uranyl acetate and lead citrate and viewed them with a Tecnai20 200-kV digital electron microscope equipped with a Gatan image filter. We evaluated atleast three tissue blocks from each treatment and captured over 50 images from each block ata 3,500× magnification, and we measured the dimensions of the indicated compartmentsdirectly from prints.
Hollyfield et al. Page 5
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Statistical analysisWe analyzed antibody titer and RPE basal deposit thickness data by ANOVA. Where theANOVA was significant, we made pairwise comparisons using Dunnett's test.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsSupported by the State of Ohio Biomedical Research and Technology Transfer Program, a Research Center Grantfrom the Foundation Fighting Blindness, and a Challenge Grant from Research to Prevent Blindness. The project wasalso supported by grants R56EY10240, R01EY014240, R24EY015638 (J.G.H.), R21EY017153 (V.L.B.),R01GM21249 (R.G.S.) and K08EY014912 (V.L.P.) from the US National Institutes of Health. The content is solelythe responsibility of the authors and does not necessarily represent the official views of the US National Eye Instituteof the National Institutes of Health. We thank J.W. Crabb for valuable discussions, B. Anand-Apte for criticalcomments on the manuscript, N.S. Peachey for help with the statistical comparisons, R.L. Fairchild for providing theRag-deficient mice, Y. Li for help with the histology, K. Sayanagi and T. Yakamoto for the fundus photography andA. Vasanji for developing the image-analysis algorithm used to define the areas of sub-RPE deposits.
References1. Crabb JW, et al. Drusen proteome analysis: an approach to the etiology of age-related macular
degeneration. Proc Natl Acad Sci USA 2002;99:14682–14687. [PubMed: 12391305]2. Gu X, et al. Carboxyethylpyrrole protein adducts and autoantibodies, biomarkers for age-related
macular degeneration. J Biol Chem 2003;278:42027–42035. [PubMed: 12923198]3. Javitt JC, Zhou Z, Maguire MG, Fine SL, Willke RJ. Incidence of exudative age-related macular
degeneration among elderly Americans. Ophthalmology 2003;110:1534–1539. [PubMed: 12917168]4. Augood CA, et al. Prevalence of age-related maculopathy in older Europeans: the European Eye Study
(EUREYE). Arch Ophthalmol 2006;124:529–535. [PubMed: 16606879]5. Hageman GS, et al. An integrated hypothesis that considers drusen as biomarkers of immune-mediated
processes at the RPE-Bruch's membrane interface in aging and age-related macular degeneration. ProgRetin Eye Res 2001;20:705–732. [PubMed: 11587915]
6. Anderson DH, Mullins RF, Hageman GS, Johnson LV. A role for local inflammation in the formationof drusen in the aging eye. Am J Ophthalmol 2002;134:411–431. [PubMed: 12208254]
7. Johnson LV, Ozaki S, Staples MK, Erickson PA, Anderson DH. A potential role for immune complexpathogenesis in drusen formation. Exp Eye Res 2000;70:441–449. [PubMed: 10865992]
8. Edwards AO, et al. Complement factor H polymorphism and age-related macular degeneration. Science2005;308:421–424. [PubMed: 15761121]
9. Hageman GS, et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH)predisposes individuals to age-related macular degeneration. Proc Natl Acad Sci USA 2005;102:7227–7232. [PubMed: 15870199]
10. Haines JL, et al. Complement factor H variant increases the risk of age-related macular degeneration.Science 2005;308:419–421. [PubMed: 15761120]
11. Klein RJ, et al. Complement factor H polymorphism in age-related macular degeneration. Science2005;308:385–389. [PubMed: 15761122]
12. Gold B, et al. Variation in factor B (BF) and complement component 2 (C2) genes is associated withage-related macular degeneration. Nat Genet 2006;38:458–462. [PubMed: 16518403]
13. Yates JR, et al. Complement C3 variant and the risk of age-related macular degeneration. N Engl JMed 2007;357:553–561. [PubMed: 17634448]
14. Anderson RE, Lissandrello PM, Maude MB, Matthes MT. Lipids of bovine retinal pigmentepithelium. Exp Eye Res 1976;23:149–157. [PubMed: 976364]
15. Anderson RE. Lipids of the ocular tissues. IV. A comparison of the phospholipids from the retina ofsix mammalian species. Exp Eye Res 1970;10:339–344. [PubMed: 4320824]
Hollyfield et al. Page 6
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

16. Fliesler SJ, Anderson RE. Chemistry and metabolism of lipids in the vertebrate retina. Prog LipidRes 1983;22:79–131. [PubMed: 6348799]
17. Seddon JM, Willett WC, Speizer FE, Hankinson SE. A prospective study of cigarette smoking andage-related macular degeneration in women. J Am Med Assoc 1996;276:1141–1146.
18. Klein R, Klein BE, Cruickshanks KJ. The prevalence of age-related maculopathy by geographicregion and ethnicity. Prog Retin Eye Res 1999;18:371–389. [PubMed: 10192518]
19. Snow KK, Seddon JM. Do age-related macular degeneration and cardiovascular disease sharecommon antecedents? Ophthalmic Epidemiol 1999;6:125–143. [PubMed: 10420212]
20. Christen WG, Glynn RJ, Manson JE, Ajani UA, Buring JE. A prospective study of cigarette smokingand risk of age-related macular degeneration in men. J Am Med Assoc 1996;276:1147–1151.
21. Solberg Y, Rosner M, Belkin M. The association between cigarette smoking and ocular diseases.Surv Ophthalmol 1998;42:535–547. [PubMed: 9635902]
22. Peakman M, Senaldi G, Vergan D. Review: assessment of complement activation in clinicalimmunology laboratories: time for reappraisal? J Clin Pathol 1989;42:1018–1025. [PubMed:2685048]
23. Ambati J, et al. An animal model of age-related macular degeneration in senescent Ccl-2– or Ccr-2–deficient mice. Nat Med 2003;9:1390–1397. [PubMed: 14566334]
24. Gottsch JD, Bynoe LA, Harlan JB, Rencs EV, Green WR. Light-induced deposits in Bruch'smembrane of protoporphyric mice. Arch Ophthalmol 1993;111:126–129. [PubMed: 8424710]
25. Cousins SW, et al. The role of aging, high-fat diet and blue light exposure in an experimental mousemodel for basal laminar deposit formation. Exp Eye Res 2002;75:543–553. [PubMed: 12457866]
26. Malek G, et al. Apolipoprotein E allele–dependent pathogenesis: a model for age-related retinaldegeneration. Proc Natl Acad Sci USA 2005;102:11900–11905. [PubMed: 16079201]
27. Mombaerts P, et al. RAG-1–deficient mice have no mature B and T lymphocytes. Cell 1992;68:869–877. [PubMed: 1547488]
28. Gu X, Sun M, Hazen S, Crabb JW, Salomon RG. Oxidatively truncated docosahexaenoatephosphoplipids: total synthesis, generation and peptide adduction chemistry. J Org Chem2003;68:3749–3761. [PubMed: 12737551]
29. Percopo CM, Hooks JJ, Shinohara T, Caspi R, Detrick B. Cytokine-mediated activation of a neuronalretinal resident cell provokes antigen presentation. J Immunol 1990;145:4101–4107. [PubMed:2147935]
Hollyfield et al. Page 7
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
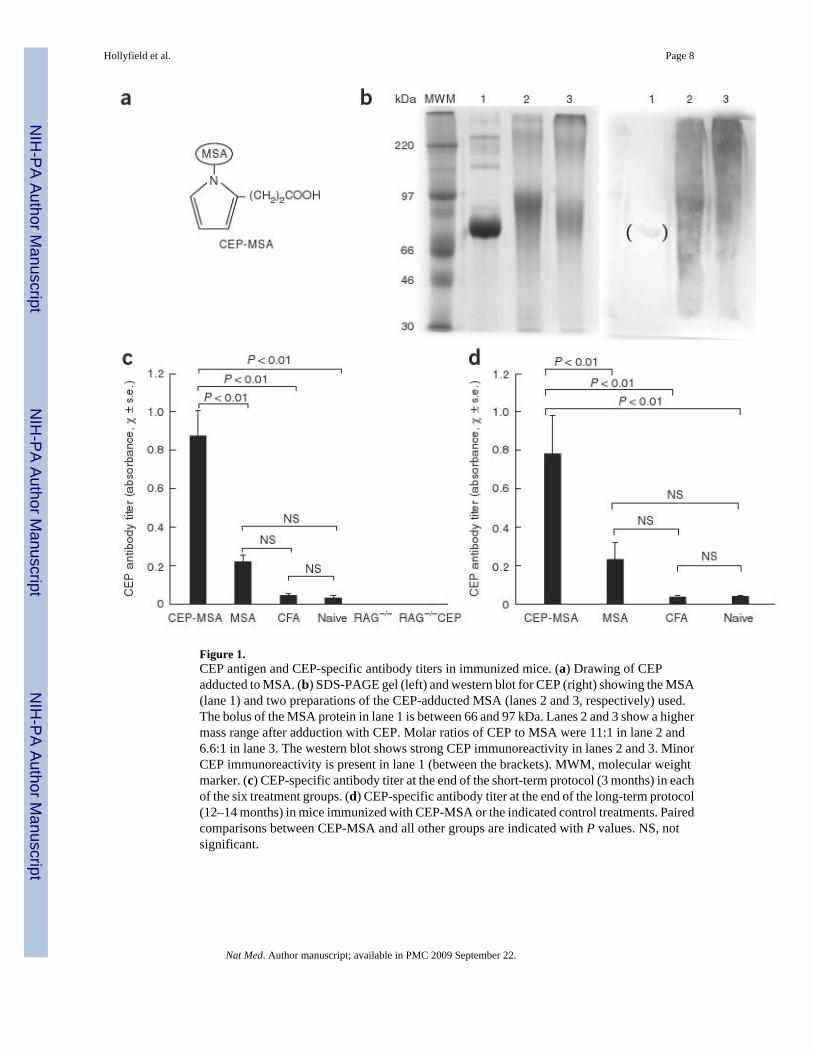
Figure 1.CEP antigen and CEP-specific antibody titers in immunized mice. (a) Drawing of CEPadducted to MSA. (b) SDS-PAGE gel (left) and western blot for CEP (right) showing the MSA(lane 1) and two preparations of the CEP-adducted MSA (lanes 2 and 3, respectively) used.The bolus of the MSA protein in lane 1 is between 66 and 97 kDa. Lanes 2 and 3 show a highermass range after adduction with CEP. Molar ratios of CEP to MSA were 11:1 in lane 2 and6.6:1 in lane 3. The western blot shows strong CEP immunoreactivity in lanes 2 and 3. MinorCEP immunoreactivity is present in lane 1 (between the brackets). MWM, molecular weightmarker. (c) CEP-specific antibody titer at the end of the short-term protocol (3 months) in eachof the six treatment groups. (d) CEP-specific antibody titer at the end of the long-term protocol(12–14 months) in mice immunized with CEP-MSA or the indicated control treatments. Pairedcomparisons between CEP-MSA and all other groups are indicated with P values. NS, notsignificant.
Hollyfield et al. Page 8
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
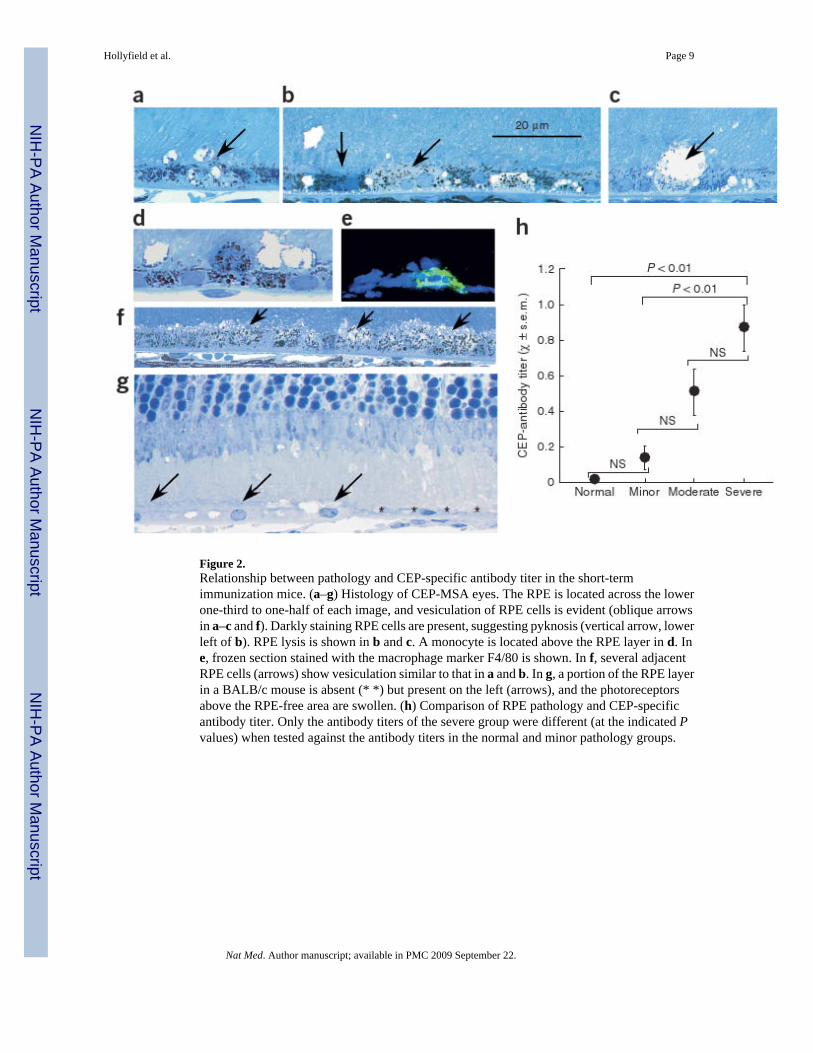
Figure 2.Relationship between pathology and CEP-specific antibody titer in the short-termimmunization mice. (a–g) Histology of CEP-MSA eyes. The RPE is located across the lowerone-third to one-half of each image, and vesiculation of RPE cells is evident (oblique arrowsin a–c and f). Darkly staining RPE cells are present, suggesting pyknosis (vertical arrow, lowerleft of b). RPE lysis is shown in b and c. A monocyte is located above the RPE layer in d. Ine, frozen section stained with the macrophage marker F4/80 is shown. In f, several adjacentRPE cells (arrows) show vesiculation similar to that in a and b. In g, a portion of the RPE layerin a BALB/c mouse is absent (* *) but present on the left (arrows), and the photoreceptorsabove the RPE-free area are swollen. (h) Comparison of RPE pathology and CEP-specificantibody titer. Only the antibody titers of the severe group were different (at the indicated Pvalues) when tested against the antibody titers in the normal and minor pathology groups.
Hollyfield et al. Page 9
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
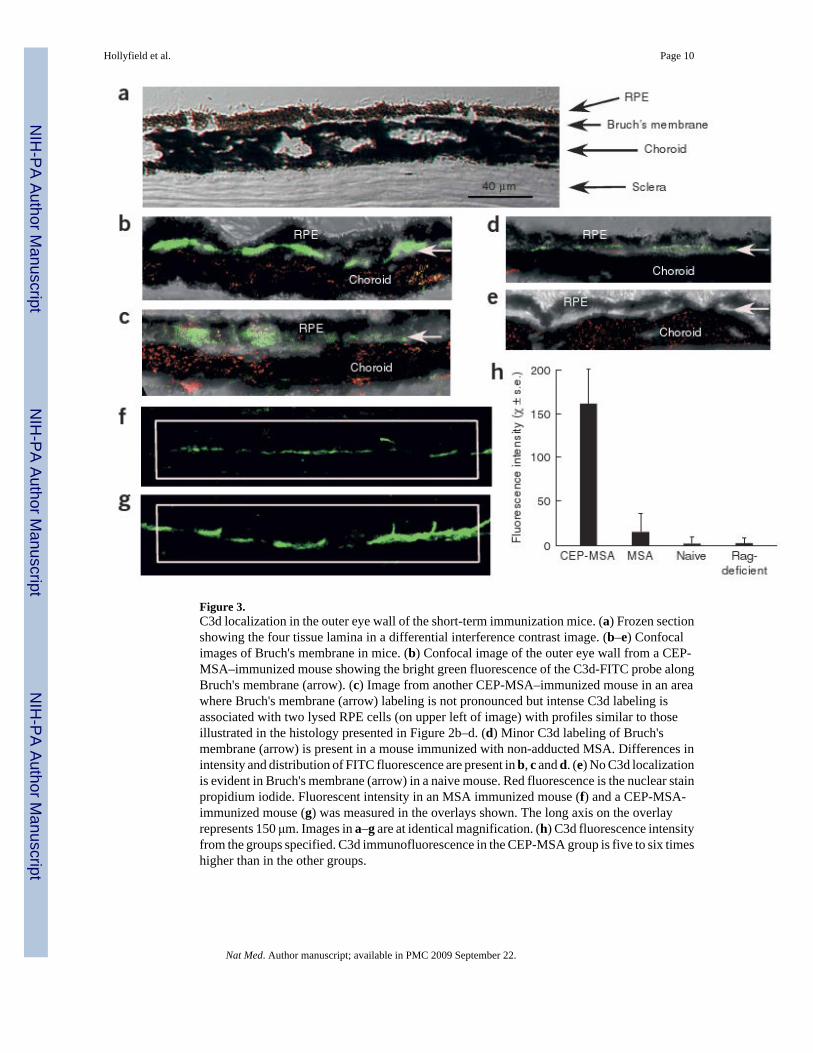
Figure 3.C3d localization in the outer eye wall of the short-term immunization mice. (a) Frozen sectionshowing the four tissue lamina in a differential interference contrast image. (b–e) Confocalimages of Bruch's membrane in mice. (b) Confocal image of the outer eye wall from a CEP-MSA–immunized mouse showing the bright green fluorescence of the C3d-FITC probe alongBruch's membrane (arrow). (c) Image from another CEP-MSA–immunized mouse in an areawhere Bruch's membrane (arrow) labeling is not pronounced but intense C3d labeling isassociated with two lysed RPE cells (on upper left of image) with profiles similar to thoseillustrated in the histology presented in Figure 2b–d. (d) Minor C3d labeling of Bruch'smembrane (arrow) is present in a mouse immunized with non-adducted MSA. Differences inintensity and distribution of FITC fluorescence are present in b, c and d. (e) No C3d localizationis evident in Bruch's membrane (arrow) in a naive mouse. Red fluorescence is the nuclear stainpropidium iodide. Fluorescent intensity in an MSA immunized mouse (f) and a CEP-MSA-immunized mouse (g) was measured in the overlays shown. The long axis on the overlayrepresents 150 μm. Images in a–g are at identical magnification. (h) C3d fluorescence intensityfrom the groups specified. C3d immunofluorescence in the CEP-MSA group is five to six timeshigher than in the other groups.
Hollyfield et al. Page 10
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
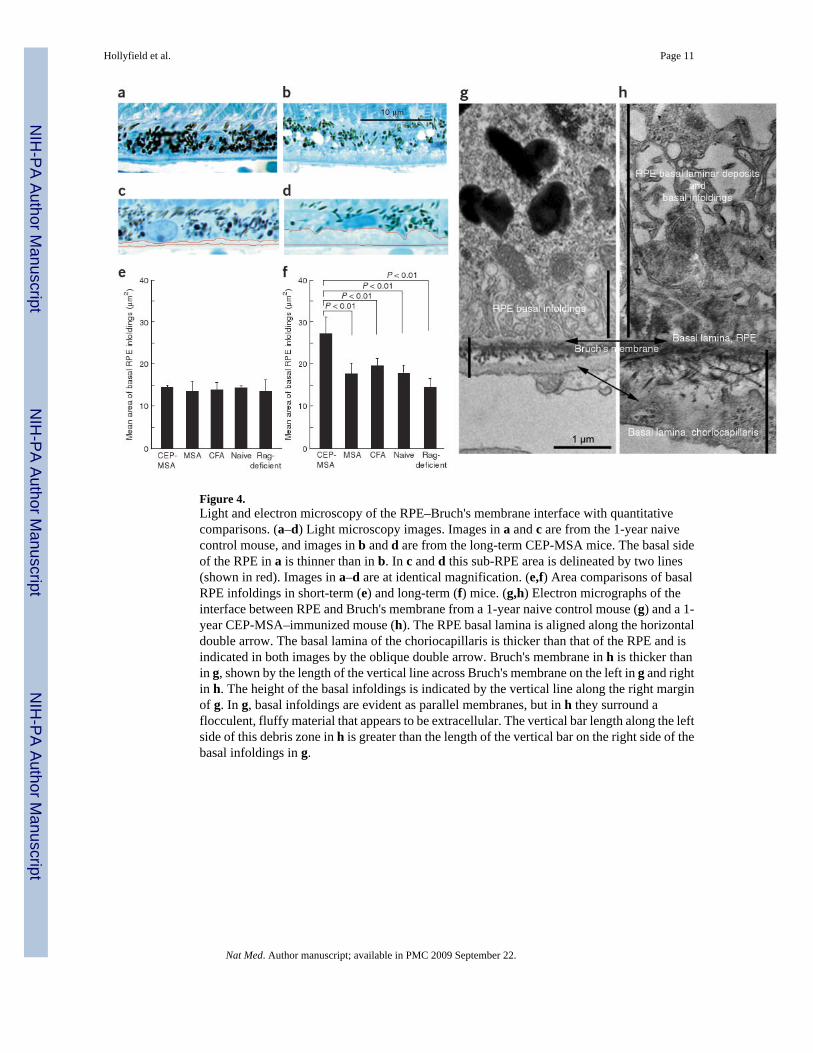
Figure 4.Light and electron microscopy of the RPE–Bruch's membrane interface with quantitativecomparisons. (a–d) Light microscopy images. Images in a and c are from the 1-year naivecontrol mouse, and images in b and d are from the long-term CEP-MSA mice. The basal sideof the RPE in a is thinner than in b. In c and d this sub-RPE area is delineated by two lines(shown in red). Images in a–d are at identical magnification. (e,f) Area comparisons of basalRPE infoldings in short-term (e) and long-term (f) mice. (g,h) Electron micrographs of theinterface between RPE and Bruch's membrane from a 1-year naive control mouse (g) and a 1-year CEP-MSA–immunized mouse (h). The RPE basal lamina is aligned along the horizontaldouble arrow. The basal lamina of the choriocapillaris is thicker than that of the RPE and isindicated in both images by the oblique double arrow. Bruch's membrane in h is thicker thanin g, shown by the length of the vertical line across Bruch's membrane on the left in g and rightin h. The height of the basal infoldings is indicated by the vertical line along the right marginof g. In g, basal infoldings are evident as parallel membranes, but in h they surround aflocculent, fluffy material that appears to be extracellular. The vertical bar length along the leftside of this debris zone in h is greater than the length of the vertical bar on the right side of thebasal infoldings in g.
Hollyfield et al. Page 11
Nat Med. Author manuscript; available in PMC 2009 September 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Related Documents











