In Silico Biology 10 (2010) 207–221 207 DOI 10.3233/ISB-2010-0437 IOS Press Overlapping Genes May Control Reprogramming of Mouse Somatic Cells into Induced Pluripotent Stem Cells (iPSCs) and Breast Cancer Stem Cells Ettore Mosca a,1,∗ , Cinzia Cocola a,1 , Davood Sabour b,1 , Paride Pelucchi a,1 , Giovanni Bertalot c , Orazio Palumbo d , Massimo Carella d , Martin G¨ otte e , Hans R. Sch ¨ oler b , Rolland Reinbold a,b,∗ , Ileana Zucchi a and Luciano Milanesi a,∗ a ITB-CNR, Segrate Milan, Italy b Max Planck Institute, M¨ unster, Germany c Azienda Ospedaliera Desenzano d/G, Manerbio (BS) Italy d IRCCS Casa Sollievo della Sofferenza, Laboratorio di Genetica Medica, San Giovanni Rotondo, (FG) Italy e University of M¨ unster Medical Center, Gynecology and Obstetrics, M¨ unster, Germany ABSTRACT: Recent findings suggest the possibility that tumors originate from cancer cells with stem cell properties. The cancer stem cell (CSC) hypothesis provides an explanation for why existing cancer therapies often fail in eradicating highly malignant tumors and end with tumor recurrence. Although normal stem cells and CSCs both share the capacity for self-renewal and multi-lineage differentiation, suggesting that CSC may be derived from normal SCs, the cellular origin of transformation of CSCs is debatable. Research suggests that the tightly controlled balance of self-renewal and differentiation that characterizes normal stem cell function is dis-regulated in cancer. Additionally, recent evidence has linked an embryonic stem cell (ESC)-like gene signature with poorly differentiated high-grade tumors, suggesting that regulatory pathways controlling pluripotency may in part contribute to the somatic CSC phenotype. Here, we introduce expression profile bioinformatic analyses of mouse breast cells with CSC properties, mouse embryonic stem (mES) and induced pluripotent stem (iPS) cells with an emphasis on how study of pluripotent stem cells may contribute to the identification of genes and pathways that facilitate events associated with oncogenesis. Global gene expression analysis from CSCs and induced pluripotent stem cell lines represent an ideal model to study cancer initiation and progression and provide insight into the origin cancer stem cells. Additionally, insight into the genetic and epigenomic mechanisms regulating the balance between self-renewal and differentiation of somatic stem cells and cancer may help to determine whether different strategies used to generate iPSCs are potentially safe for therapeutic use. INTRODUCTION Recent studies have linked an embryonic stem cell (ESC)-like gene signature with poorly differentiated and highly aggressive tumors (Ben-Porath et al., 2008). This linkage suggests that the molecular profiles of pluripotent stem cells, cancer cells and tissue specific cancer stem cells (CSCs) might be closely 1 Authors contributed equally to this work. ∗ Corresponding author: Ettore Mosca, ITB-CNR, Via Cervi 93, Segrate Milan, Italy. E-mails: [email protected], [email protected], [email protected]. 1386-6338/10/$27.50 2010 – IOS Press and the authors. All rights reserved

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

In Silico Biology 10 (2010) 207–221 207DOI 10.3233/ISB-2010-0437IOS Press
Overlapping Genes May ControlReprogramming of Mouse Somatic Cellsinto Induced Pluripotent Stem Cells (iPSCs)and Breast Cancer Stem Cells
Ettore Moscaa,1,∗, Cinzia Cocolaa,1, Davood Sabourb,1, Paride Pelucchia,1, Giovanni Bertalotc,Orazio Palumbod, Massimo Carellad, Martin Gottee, Hans R. Scholerb, Rolland Reinbolda,b,∗,Ileana Zucchia and Luciano Milanesia,∗
aITB-CNR, Segrate Milan, ItalybMax Planck Institute, Munster, GermanycAzienda Ospedaliera Desenzano d/G, Manerbio (BS) ItalydIRCCS Casa Sollievo della Sofferenza, Laboratorio di Genetica Medica, San Giovanni Rotondo, (FG) ItalyeUniversity of Munster Medical Center, Gynecology and Obstetrics, Munster, Germany
ABSTRACT: Recent findings suggest the possibility that tumors originate from cancer cells with stem cell properties. Thecancer stem cell (CSC) hypothesis provides an explanation for why existing cancer therapies often fail in eradicating highlymalignant tumors and end with tumor recurrence. Although normal stem cells and CSCs both share the capacity for self-renewaland multi-lineage differentiation, suggesting that CSC may be derived from normal SCs, the cellular origin of transformation ofCSCs is debatable. Research suggests that the tightly controlled balance of self-renewal and differentiation that characterizesnormal stem cell function is dis-regulated in cancer. Additionally, recent evidence has linked an embryonic stem cell (ESC)-likegene signature with poorly differentiated high-grade tumors, suggesting that regulatory pathways controlling pluripotency mayin part contribute to the somatic CSC phenotype. Here, we introduce expression profile bioinformatic analyses of mouse breastcells with CSC properties, mouse embryonic stem (mES) and induced pluripotent stem (iPS) cells with an emphasis on howstudy of pluripotent stem cells may contribute to the identification of genes and pathways that facilitate events associated withoncogenesis. Global gene expression analysis from CSCs and induced pluripotent stem cell lines represent an ideal modelto study cancer initiation and progression and provide insight into the origin cancer stem cells. Additionally, insight into thegenetic and epigenomic mechanisms regulating the balance between self-renewal and differentiation of somatic stem cells andcancer may help to determine whether different strategies used to generate iPSCs are potentially safe for therapeutic use.
INTRODUCTION
Recent studies have linked an embryonic stem cell (ESC)-like gene signature with poorly differentiatedand highly aggressive tumors (Ben-Porath et al., 2008). This linkage suggests that the molecular profilesof pluripotent stem cells, cancer cells and tissue specific cancer stem cells (CSCs) might be closely
1Authors contributed equally to this work.∗Corresponding author: Ettore Mosca, ITB-CNR, Via Cervi 93, Segrate Milan, Italy. E-mails: [email protected],
[email protected], [email protected].
1386-6338/10/$27.50 2010 – IOS Press and the authors. All rights reserved

208 E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells
related. The CSC hypothesis does not specifically address the cellular mechanisms of transformation orthe origin of CSCs, but it is based on the concept that a small population of cells within a tumor displaystem-cell-like properties with tumor initiation and propagation capacity (Kalirai et al., 2006). Morerecently, various studies suggest that the processes that lead to the production of induced pluripotentstem cells (iPSCs) by reprogramming of non-pluripotent cells and transformation of somatic cells tomalignant cells overlap and that CSCs might initially arise through a reprogramming-like mechanismsimilar to somatic cell reprogramming to derive iPSCs (Utikal et al., 2009). For instance, self-renewal,the process by which stem cells divide to make more stem cells in order to perpetuate the stem cell poolthroughout life, requires cell cycle control. Self-renewal most likely involves networks that balancesproto-oncogenes (promoting self-renewal), gate-keeping tumor suppressors (limiting self-renewal), andcare-taking tumor suppressors (maintaining genomic integrity) (He et al., 2009). Reduced stem cellfunction and tissue regenerative capacity during aging may be caused by changes in stem cell self-renewal and tumor suppression programs. Therefore, cancer may arise from mutations or throughaberrant modulation of gene expression through epigenomic mechanisms that inappropriately activateself-renewal and decrease tumor suppression programs.
Induced pluripotent stem cells while generated by forced expression of a defined and limited setof transcription factors results in extensive but not well characterized changes at the chromosomaland epigenomic level, changes that may contribute to malignant cellular transformation of the iPSCsthemselves or more lineage restricted derivative cells such as tissue specific stem cells (Takahashi et al.,2006). Recently it has become apparent that significant obstacles have to be overcome before iPSCscan be considered for clinical trial studies, such as reliably determining and predicting whether specificpatient produced iPSCs have tumorigenic potential. As a research tool, the iPSC technology can alsoprovide opportunities to study cancer transformation and the etiology of cancer and the cellular originof CSCs. The detailed mechanisms for reprogramming of iPSCs and malignant transformation areunknown and are topics of intense research. The promise of patient-specific therapy will initially requirethe establishment of iPSC banks for the identification and screening of malignant genetic and epigenomicmodifications that are inadvertently introduced into patient cells used as starting material.
We propose that comparisons of gene expression profiles between normal pluripotent cells (such asESCs), tissue specific CSCs and iPSC lines may allow for detailed evaluation of the potential safety ofthe new iPSC lines and for the identification of potential cancer inducing and repressing genes. Similarsafety issues were raised previously for the therapeutic use of human ES cells since human ES cellsgrown in long-term culture have been shown to have genetic instability (Lefort et al., 2008). Whileearly iPSCs production relied on genetic modification of target cells, even with newer technologiesthat promise cellular reprogramming without genetic modification, presumably iPSCs will also displaygenetic instability due to transient epigenomicmodifications introduced during their derivation, culturing,expansion and differentiation. Safety of patient-derived iPSCs will depend on whether derivative cellsfrom reprogrammed cells will develop into tumors, even after many years when the patients have receivedthe reprogrammed cells.
We introduce a bioinformatics approach that may provide a prognostic tool to help determine thetumorgenic potential of iPSC and of iPS derivative cells. Although this approach utilizesmousemammarycells with CSC properties as an example of proof of principle, candidate cancer genes identified by thisstrategy using CSCs from human adult tissue may eventually help to better evaluate the safety of patientderived iPSCs. The strategy utilizes expression data derived from purified mouse pluripotent (mES)cells, mouse iPSCs and purified mouse breast cells with CSC properties. This strategy also represent ageneral approach that may suggest therapeutic strategies that can allow for selective destruction of tissuespecific CSCs.

E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells 209
RESULTS
Spontaneously transformed NMuMG cells exhibit tumor initiation potential and have the capacity togenerate complex 3D structures reminiscent of the mammary architecture
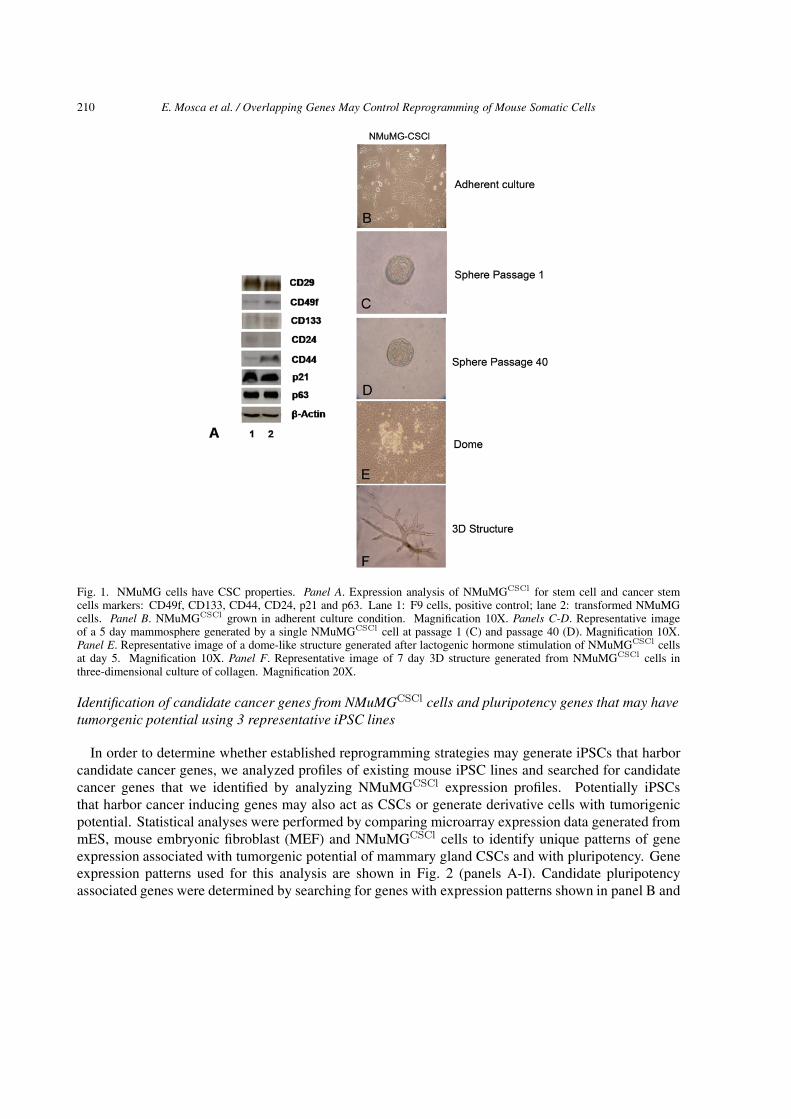
Combined knowledge of mutated genes and of the genes whose expression is altered in cellular trans-formation and reprogramming is necessary to predict whether iPSCs have tumorigenic potential and todevelop safe strategies for iPSCs production. To determine if iPSCs expressing potential cancer inducinggenes may result in tumor formation when used to generate adult SCs, spontaneously transformed normalmouse mammary epithelial NMuMG cells were used as a model. Spontaneous transformation may beinitiated by mutations, genetic instability, epigenomic alterations and aberrant changes associated withcellular defense mechanisms such as defective DNA repair programs (Gerson et al., 2006). To identifygenes that may contribute to mammary gland cellular transformation and to investigate whether existingpublished mouse iPSCs harbor candidate cancer genes, we used a cell line with tumorgenic potentialthat we established in our laboratory through the isolation of spontaneously transformed malignant cellsgenerated in culture from normal non-malignant mouse mammary epithelial NMuMG cells (Owens etal., 1974, Soriano et al., 1995). While normal NMuMG cells have the capacity to reconstitute normalmammary gland after mouse fat pad clearing (Soriano et al., 1998; Kamalati et al., 1999) transformedNMuMG-derivative cells when transplanted into nude mice have the ability to develop tumors (Cocolaand Zucchi, unpublished results). We determined that transformed NMuMG cells have the capacity toinitiate heterogeneous tumors that recapitulate morphologically the 3D architecture of the mammary tree(not shown) and generate all of the cell types of the mammary gland in vivo and in vitro (not shown).Transformed NMuMG cells, like rat LA7CSCs (Zucchi et al., 2007, Zucchi et al., 2008) are character-ized by expression of CD29 and CD49f, which mark a mammary stem cell–enriched population in mice(Al-Hajj et al., 2003, O’Brien 2007, Collins et al. 2005, Li et al., 2007, Stingl et al., 2006, Shackletonet al., 2006), CD44 and CD133, both markers for breast, colon, prostate and pancreatic CSCs and brainstem cells (Ricci-Vitiani et al., 2007, Richardson et al., 2004, Hermann et al., 2007, Lee et al., 2005),p21/WAF1, a gene involved in the maintenance of the quiescence of mammary, hematopoietic and brainstem cells (Woodward et al., 2005, Cheng et al., 2000, Kippin et al., 2005), p63 and by low or noexpression of CD24 (Fig. 1, Panels A and B).
Previously we have demonstrated that extensive capacity to regenerate spheres in non-adherent culturecondition is a property of self-renewing CSCs (Zucchi et al., 2007), therefore to determine whethertransformed NMuMG-cells have extensive capacity for self-renewal, cells were cultured in non-adherentconditions (Dontu et al., 2003). Mammospheres were observed 5–7 days after seeding of the cells (panelC) and when mechanically and enzymatically dissociated into a single-cell suspension, mammosphereswere serially regenerated for more than 40 passages (panel D). To assess whether these cells at the singlecell level can recapitulate alveolar-like structures, lactogenic hormones were used to induce dome-likestructure formation (panel E, Zucchi et al., 2002). Since 3D collagene culture systems provide thephysiological signals necessary for mammary morphogenesis in vitro and enable mammary cells to alsorecapitulate the spatial orientation and architecture of the mammary tree occurring in vivo (Zucchi et al.,2007), collagen was used to induce the cells to generate branched duct-like structures (panel F). Theseassays and the ability of the transformed NMuMG cells to generate heterogenous tumors demonstratedthat these clonal-derivative cells retained CSC properties and we therefore designated them NMuMGCSCl
cells (CSCl for Cancer Stem Cell like).
210 E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells
Fig. 1. NMuMG cells have CSC properties. Panel A. Expression analysis of NMuMGCSCl for stem cell and cancer stemcells markers: CD49f, CD133, CD44, CD24, p21 and p63. Lane 1: F9 cells, positive control; lane 2: transformed NMuMGcells. Panel B. NMuMGCSCl grown in adherent culture condition. Magnification 10X. Panels C-D. Representative imageof a 5 day mammosphere generated by a single NMuMGCSCl cell at passage 1 (C) and passage 40 (D). Magnification 10X.Panel E. Representative image of a dome-like structure generated after lactogenic hormone stimulation of NMuMGCSCl cellsat day 5. Magnification 10X. Panel F. Representative image of 7 day 3D structure generated from NMuMGCSCl cells inthree-dimensional culture of collagen. Magnification 20X.
Identification of candidate cancer genes from NMuMGCSCl cells and pluripotency genes that may havetumorgenic potential using 3 representative iPSC lines
In order to determine whether established reprogramming strategies may generate iPSCs that harborcandidate cancer genes, we analyzed profiles of existing mouse iPSC lines and searched for candidatecancer genes that we identified by analyzing NMuMGCSCl expression profiles. Potentially iPSCsthat harbor cancer inducing genes may also act as CSCs or generate derivative cells with tumorigenicpotential. Statistical analyses were performed by comparing microarray expression data generated frommES, mouse embryonic fibroblast (MEF) and NMuMGCSCl cells to identify unique patterns of geneexpression associated with tumorgenic potential of mammary gland CSCs and with pluripotency. Geneexpression patterns used for this analysis are shown in Fig. 2 (panels A-I). Candidate pluripotencyassociated genes were determined by searching for genes with expression patterns shown in panel B and

E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells 211
Fig. 2. Representation of genes differentially expressed or in common between NMuMGCSCl, MEF, mES cells and establishediPSC lines. Panels A-I. Representative gene expression profiles between mESCs, MEFs and NMuMGCSCl cells obtained frommicroarray expression data used to identify candidate cancer genes from NMuMGCSCl cells, pluripotency associated genesfrom mESCs and pluripotency associated genes that may contribute to the tumorgenic potential of iPSCs; vertical axis showsnormalized intensity values obtained from microarray expression data. Panel A. Gene expression patterns representative of genesdifferentially expressed only between MEF and mESC. Panel B. Genes uniquely associated with mESCs and not differentiallyexpressed in MEF and NMuMGCSCl. Panel C. Genes differentially expressed only between mESC and NMuMGCSCl. PanelD. Genes uniquely associated with MEF and not differentially expressed in mESC and NMuMGCSCl Panel H. Genes uniquelyassociated NMuMGCSCl and not differentially expressed in mESCs and MEF. Panel I. Genes differentially expressed onlybetween MEF and NMuMGCSCl . Panels E-G. Expression pattern representative of all variations of genes differentiallyexpressed when comparing mESCs, MEF and NMuMGCSCl . Panel J. Venn diagram showing the distribution of the genesdifferentially expressed between mESCs, MEF and NMuMGCSCl cells. Each set contains genes differentially expressed in thecomparison between the cell types reported above it. The total numbers of genes differentially expressed in each subset of theVenn diagram are shown. Roman numerals I-VII for each subset refers to the representative gene expression profiles shownin Panels A-I. Subset VI are candidate cancer genes, subset II are candidate pluripotency genes and subsets III are candidategenes that are likely to have a role in cancer and/or pluripotency of iPSCs. Panel K. Overlap between candidate cancer genesidentified in NMuMGCSCl cells and genes that are differentially expressed between mESC and representative iPSCs. Thethree gene sets composed of 13, 18 and 45 genes, contain candidate cancer genes (from subset VI) that are also differentiallyexpressed between mESCs and iPSCs. Panel L. Overlap between candidate cancer genes identified in NMuMGCSCl cells andgenes that are differentially expressed between mESC and that may provide tumorgenic potential to iPSCs. The three gene setscomposed of 2, 11 and 16 genes contain candidate cancer genes (from subset III) that are also differentially expressed betweenmESC and iPSCs. For panel K and L, 134, 166 and 634 genes are differentially expressed between mESC and iPS reportedby Kang et al. (2009), Cho et al. (2010) and Chen et al. (2009); gene symbols reported in the figure refer to genes belongingto the intersections; the p values assess the significance of the observed overlaps and were calculated using the appropriatehypergeometric tests.

212 E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells
A
Fig. 3. Functional grouping of the GO biological processes enriched (FDR < 0.05) in subsets III (panel A) and VI (panel B).Term grouping is based on the similarity of their associated genes. The size of the nodes reflects the statistical significance ofthe terms. The group leading term is the most significant term within the group. We obtained these networks using ClueGO(Bindea et al., 2009) a plugin for Cytoscape (Shannon et al., 2003).
displaying a significant (FDR < 0.05) expression variation between NMuMGCSCl compared to mESCsand between mESCs compared to MEF, but having similar expression levels in NMuMGCSCl and MEF.Candidate cancer genes were determined by searching for genes with expression patterns shown in panelH and displaying a significant (FDR < 0.05) expression variation between NMuMGCSCl comparedto MEF and between NMuMGCSCl cells compared to mESCs but have a similar expression level inmESCs and MEF cells. Panel H also includes genes that may not have a role in cancer but may havea role in normal mammary gland SC maintenance and/or function. Therefore these genes need to beexperimentally verified as cancer inducing or repressing genes.

E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells 213
B
Fig. 3. continued.
A total number of 1129 genes were identified as candidate pluripotency genes (Fig. 2 Panel J, subsetII), 567 genes as candidate cancer genes from NMuMGCSCl (subset VI), and 370 genes as candidatepluripotency genes that when aberrantly expressed may contribute to the tumorigenic potential of theNMuMGCSCl (subset III); details regarding genes belonging to subsets VI and III are reported inSupplementary Tables 1-2. Using MEF cells in the analysis of the expression profiles allowed genesrelated to epithelial and/or fibroblast cell specific function and maintenance (shown in Fig. 2 subset IV)to be distinguished from cancer and pluripotency.
We analyzed the GO biological processes and KEGG pathways regulated by the candidate cancer genes(identified in subset VI) and candidate pluripotency genes that when aberrantly expressed may contributeto tumorigenesis (subset III) (Figs 3 and 4, Panels A and B). The functional grouping shown in Panel Aprovides support that the identified candidate cancer genes may inappropriately regulate self-renewal anddecrease tumor suppression programs such as the negative regulation of apoptosis and immune system
214 E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells
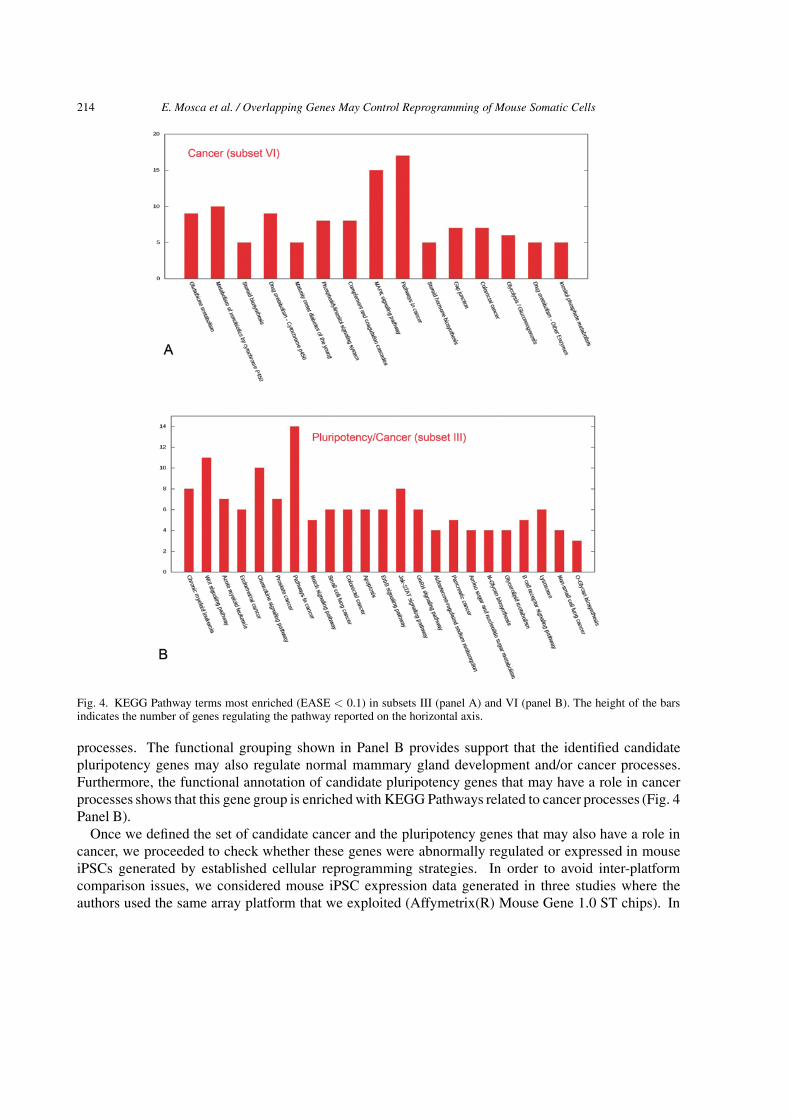
Fig. 4. KEGG Pathway terms most enriched (EASE < 0.1) in subsets III (panel A) and VI (panel B). The height of the barsindicates the number of genes regulating the pathway reported on the horizontal axis.
processes. The functional grouping shown in Panel B provides support that the identified candidatepluripotency genes may also regulate normal mammary gland development and/or cancer processes.Furthermore, the functional annotation of candidate pluripotency genes that may have a role in cancerprocesses shows that this gene group is enriched with KEGG Pathways related to cancer processes (Fig. 4Panel B).
Once we defined the set of candidate cancer and the pluripotency genes that may also have a role incancer, we proceeded to check whether these genes were abnormally regulated or expressed in mouseiPSCs generated by established cellular reprogramming strategies. In order to avoid inter-platformcomparison issues, we considered mouse iPSC expression data generated in three studies where theauthors used the same array platform that we exploited (Affymetrix(R) Mouse Gene 1.0 ST chips). In

E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells 215
Table 1Number of candidate cancer genes that show differential expression between iPSCs-ESCs inexpression data generated by Kang et al. (2009), Cho et al. (2010) and Chen et al. (2009). Thecolumns labelledwith the symbol “p” contain the p values that assess the degree of significanceof the observed intersection; “#” indicates the number of genes in the intersection; romannumbers refer to subsets of Fig. 2; “total” is the total number of genes in our dataset ofexpression data; “∩” indicates the intersection between two sets
iPSCs-ESCs VI III total iPSCs-ESCs ∩ VI IPSCs-ESCs ∩ III# p # p
Kang 134 567 370 19138 13 1.77E-004 2 0.735Chen 634 567 370 19138 45 6.10E-008 16 0.169Cho 166 567 370 19138 18 2.23E-006 11 3.97E-004
particular, we analyzed the “Kang” (Kang et al., 2009) and the “Chen” (Chen et al., 2009) datasets, andfound respectively, 155 and 713 genes differentially expressedbetween their reprogrammed “pluripotent”cells (iPSCs) and “normal” pluripotent cells (mESCs). Moreover, we considered the list of 188 genesdifferentially expressed between iPSCs and mESCs provided by Cho et al. (2010). The overlap of thecomparison among these three gene lists and the set of candidate cancer genes is depicted in Fig. 2(Panels J, K and L) and summarized in Table 1.
What is observed from panels J, K and L is that while the three labs generated iPSCs independently,several genes (namely Hmgn5, Jun, Ly6a, Crmp1, Ccnd1, Serpinb9b, Phlda1) with tumorgenic potentialwere found as common differentially expressed genes between iPSCs and mESCs, suggesting that dif-ferent reprogramming strategies may share a common mechanism leading to aberrant levels of specificgenes compared to mESCs. In particular, the labs of Kang and Cho both used protein based repro-gramming without genetic manipulation of the starting cells to produce iPSCs in both cases, genes Junand Hmgn5 identified with NMuMGCSCl cells as having tumorgenic potential, are aberrantly expressedin their respective iPS cells. This suggests that “safe” reprogramming strategies may share a commonmechanism leading to aberrant levels of these genes compared to mESCs.
The exact role of the genes identified in our study in cellular reprogramming to generate iPSCs andcancer is currently not known. Nuclear protein NSBP1 (HMGN5) is a recently discovered member ofthe HMGN nucleosome binding protein family. The interaction of NSBP1(HMGN5) with nucleosomesin euchromatin counteracts linker histone-mediated chromatin compaction and modulates transcription(Rochman et al., 2009). The proto-oncogene JUN is a putative transforming gene that is a componentof AP1 (activating protein 1) that regulates the expression and function of cell cycle regulators suchas Cyclin D1, p53, p21cip1/waf1, p19 (ARF) and p16 and has been recently described as having a rolein breast cancer invasion (Jiao et al., 2010). In addition Ly6a, which encodes for SCA1 protein, is amember of the Ly6 multigene family of type V glycophosphatidylinositol anchored cell surface proteins.Ly6a is expressed on various hematopoietic, mammary gland and mesenchymal progenitor/stem cellsand is a candidate marker for tissue specific CSCs (such as prostate and mammary gland) (Woodwardet al., 2005). The Crmp1 (collapsin response mediator protein 1) gene encodes a member of a familyof cytosolic phosphoproteins and is predominantly expressed in the nervous system (McIntyre et al.,2010). Crmp1 behaves as a tumor suppressor and negative regulator of invasion suppression and loss ofCrmp1 contributes to increased invasive phenotype of glioblastoma tumors and lymph node metastasisin lung cancer patients (Pan et al., 2010). PHLDA1 is a putative epithelial stem cell marker in thehuman small and large intestine and contributes to migration and proliferation in colon cancer cells(Sakthianandeswaren et al., 2011).

216 E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells
DISCUSSION
Pluripotent cells were initially derived from the inner cell mass of pre-implantation embryos and aretermed embryonic stem cells (mESCs) (for review see, Boiani et al., 2005). Undifferentiated pluripotentstem cells have the potential for expansion with the retention of normal karyotype in vivo and the abilityto generate cells of all three germ layers that can then further differentiate into many specific somaticstem cell lineages. Therefore pluripotent cells have almost unlimited potential in therapeutic regenerativemedicine. The recent discovery that non-pluripotent cells such as fibroblast cells can be “reprogrammed”by transient expression of a small number of genes into induced pluripotent stem cells (iPSCs) makeshuman iPSCs an ideal cell source for cell therapy given that patient derived iPSCs are genetically identicalto all cells of the patient. Therefore immune response and tissue rejection are not an issue when iPSCs areused for cell regeneration and tissue replacement therapies. However, the full extent of the relationshipof iPSCs to bona fide ESCs has not yet been fully determined and potential safety in their therapeutic usehas not been sufficiently evaluated. In fact, even embryo-derived ES cells have been tested in only a fewsettings, and their effectiveness and safety are currently not well established. Since an extensive numberof methods for producing iPSCs have been proposed and described, the issue into how to identify geneswith malignant potential that are inadvertently introduced to the starting patient cells have not been befully evaluated.
Recently, several studies have shown that the acquisition of immortality is a crucial and rate-limitingstep towards the establishment of a pluripotent state in somatic cells and emphasizes the similaritiesbetween induced pluripotency and tumorigenesis (Utikal et al., 2009). Notably we observed that manygenes regulating mammary gland oncogenesis were also present in normal pluripotent cells (Fig. 3 PanelB) suggesting that certain genes have both a role in pluripotency and breast development and cancer.
CONCLUSIONS
Our study was aimed at identifying candidate pluripotency genes that may be involved in the cancerprocess when aberrantly expressed in breast SCs. Our results suggest that reprogramming even withoutgenetic manipulation (such as protein based reprogramming) of the starting cells may result in theproduction of iPSCs that harbor malignant genes or aberrant gene expression patterns that contribute tocancer development of iPSCs and lineage restricted cells derived from iPSCs. Many of the genes weidentified are known to have a role in cancer initiation and/or progression (such as Jun) or are markersfor cancer cells and CSCs. Further studies will be required to confirm and identify the etiology of thetumorigenic potential of these genes and how the aberrant cancer gene expression patterns were inducedinto the iPSCs. Since the different iPSC lines we analyzed have similar genes with aberrant expression(eg, Jun and Hmgn5), this suggests that there maybe a common mechanism related to the process ofhow malignant potential is induced into the iPSCs regardless of the starting cell type and reprogrammingstrategies utilized. This for instance maybe due to the acquisition of immortality which is required forthe establishment of a pluripotent state or general genomic instability associated with culturing of iPSCsrather than due to a specific derivation strategy. Additionally, our analyses provides evidence of theexistence of pathways in common in reprogramming and cancer transformation and suggest that iPSCassays can provide a new approach for investigating how the normal cell state may be subverted in cancer.

E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells 217
METHODS
NMuMGCSCl cell line
The spontaneously transformed cell was isolated from a non tumorigenic NMuMG pool of cells. TheNMuMG cells exhibit cancer stem cell properties of cell self-renewal and differentiation in vitro.
Adherent and non-Adherent culture conditions
Adherent cells were cultured in DMEM, 10% fetal bovine serum (FBS, Gibco Laboratories), supple-mented with 10 μg/ml insulin (Sigma). Differentiation was induced in NMuMGCSCl post confluentculture using prolactin [3 μg/ml (Sigma)] for 96 hrs. Cells were harvested with 0.05% trypsin containing0.53 mM EDTA (Invitrogen).
Non-adherent cells were cultured in the DMEM, 5% FBS containing B27 (Invitrogen), 5 μg/ml insulin,20 ng/ml epidermal growth factor (EGF, Sigma), 20 ng/ml basic fibroblast growth factor (bFGF) and4 μg/ml heparin (Sigma) in low attachment plates (Sterilin, Stone Staffs) at clonogenic density (1000cells/ml). The capacity for sphere regeneration was tested by dissociating spheres with 0.05% trypsincontaining 0.53 mM EDTA and re-plating the cells at clonogenic density.
3D culture conditions
Monolayer cultures were detached with trypsin, cells were mixed with rat tail-derived collagen (pre-pared according to Soriano et al., 1995) and plated at a concentration of 1000 cells/well in 24-well plates.After collagen solidification, 500 μl of medium was added to each well and half of medium was changedevery 2–3 days.
Western Blot analysis
Cells were lysed on ice as described (Zucchi et al., 1998). Gel electrophoresis was performed usinga 12% of SDS denaturing poly-acrylamide gel. After transfer to PVDF membrane (GE-Biotechnology)the membranes were incubated with primary antibodies (all from Santa Cruz Biotechnology): goatpolyclonal anti-α6-Integrin (sc-6597), rabbit polyclonal anti-β1-Integrin (sc-8978), goat polyclonal anti-CD133 (sc-23797), rabbit polyclonal anti-CD44 (sc-7946),mouse monoclonal anti-p21 (sc-6246),mousemonoclonal anti-p63 (sc-8431) and goat polyclonal β-Actin (sc-1651), according to the manufacturers’recommendations. Secondary antibodies were all from Santa Cruz Biotechnology (rabbit anti-goat,sc-2769; goat anti-mouse, sc-2005; goat anti-rabbit, sc-2004). F9 mouse embryonic carcinoma cellswere used as positive controls.
mRNA isolation, hybridization, and microarray analysis
RNA isolation was performed using TRIzol Reagent (InvitrogenTM Life Technologies, Carlsbad, CA)according with manufacturers’ instructions and analyzed using 2100 Bioanalyzer (Agilent Technologies,Santa Clara, CA) to determine the quantity and the integrity of the sample.
The Affymetrix (Santa Clara, CA) Mouse Gene 1.0 ST Array whole transcript-based array interrogates28,853 well annotated genes. Staining, hybridization, washing and scanning of the array were performedfollowing manufacturers’ protocols. Three biological replicates were run independently, providingtriplicates for each condition. Gene expression data were stored as .cel format file that could beinterpreted by array analysis software. This dataset is designated as “Zucchi”.

218 E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells
Gene expression analysis of iPSC lines
Expression data generated with the platform Affymetrix(R) Mouse Gene 1.0 ST chips from the GeneExpression Omnibus database were analyzed (Barret et al., 2009). Six samples of mESCs (3 replica),and mouse iPSCs (3 replica) available in GEO under accession number GSE17004 were analysed fromKang (Kang et al., 2009), Chen (Chen et al., 2009) GEO accession GSE19604; and from (Cho et al.,2010) GEO accession GSE13770.
MICROARRAY DATA PRE-PROCESSING AND FILTERING
Zucchi, Kang and Chen datasets separately pre-processed in order to avoid the loss of biological signifi-cance which may occur as a consequence of global normalisation of data from different sources. Datasetswere pre-processed using the Robust Multichip Average strategy (background adjustment, quantile nor-malisation and probe summarisation) implemented in the R package aroma.affymetrix (Bengtsson et al.,2008). To filter out extremely low expression values, a filtering criteria was implemented establishinga background expression cutoff ε equal to the median of the intronic probe set signals: 50 for Zucchiand Kang datasets, 16 for Chen dataset. This filtering procedure discarded genes with a mean intensityvalue lower than ε in all the three cell types. Genes with mean intensity values higher then ε in at leastone cell type were retained but converted such that all the intensities lower than ε were corrected toε. The filtering procedure determined an enrichment of 141% (Zucchi dataset), 146% (Kang dataset)and 138% (Chen dataset). We calculated the enrichment as 100nc’ / (n’c), where (n, c) and (n’, c’) arerespectively, the total number of probe sets and the number of control probe sets before and after thefiltering procedure. Of the total 28,853 genes, 19,138 (Zucchi), 18,383 (Kang) and 20,325 (Chen) geneswere retained for further analyses.
Assessment of differential expression
Gene differential expression was assessed using LIMMA (LinearModels forMicroarray Data) (Irizarryet al., 2003). We used FDR < 0.05 criteria to select genes showing a significant expression changebetween a pair of conditions.
Functional annotation and grouping
The Functional Annotation tool provided by the DAVID Bioinformatics Resources was used forannotation and grouping of genes (Huang et al., 2009). We compared the distribution of KEGG(Kanehisa et al., 2002) pathways in our gene lists with the background distribution specific for theAffymetrix(R) Mouse Gene 1.0 ST chip and selected the annotation terms enriched with an EASE score< 0.1.
The networks representing the relations among the Gene Ontology annotations were generated withClueGO (Bindea et al., 2009), a plugin for Cytoscape (Shannon et al.,2003). ClueGO creates functionallygrouped annotation networks that show the relationships among the terms based on the similarity of thegenes associated to the annotations. The size of the nodes depends on the statistical significance ofthe terms. Each group has a specific colour and the group leading term is the most significant termwithin a group. We considered the Gene Ontology biological process annotation (excluding those withevidence code equal to IEA = inferred from electronic annotation), the enrichment (one-sided test) andthe Benjamini-Hochberg p value correction.

E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells 219
Significance of the number of overlapping genes
The significance of the observed overlap between two lists of geneswas calculated using the appropriatehypergeometric tests.
ACKNOWLEDGMENTS
We thank Loredana Ansalone and John Hatton for extraordinary administrative secretarial supportand the staff of the Department of Pathology of Leno BS for their excellent technical support. IZgrant support: Net2Drug Grant 037590, the Italy-U.S.A.-Project Grant N.527 B-B7, MIUR-FIRBGrant RBIP0695BB 003, Telethon Grant GG004247 and Fondazione CARIPLO Nobel and CARIPLO-Progetti-Internazionali. This work has been also supported by the MIUR FIRB ITALBIONET(RBPR05ZK2Z), BIOPOPGEN and the Flagship INTEROMICS projects.
SUPPLEMENTARY TABLES
Table 1. List of candidate cancer genes (see Fig. 2 subset VI); available at the URL ftp://fileshare.itb.cnr.it/Public/isb437/ST1.xls.
Table 2. List of candidate pluripotency genes that, when aberrantly expressed, may contribute tothe tumorgenic potential of the NMuMGCSCl cells (see Fig. 2, subset III); available at the URL ftp://fileshare.itb.cnr.it/Public/isb437/ST2.xls.
REFERENCES
• Al-Hajj, M., Wicha, M.S., Benito-Hernandez, A., Morrison, S.J., Clarke, M.F. (2003). Prospective identification oftumorigenic breast cancer cells. Proc Natl Acad Sci USA. 100. 3983-3988.
• Barrett, T, Troup, D.B., Wilhite, S.E., Ledoux, P., Rudnev, D., Evangelista, C., Kim, I.F., Soboleva, A., Tomashevsky,M., Marshall, K.A., Phillippy, K. H., Sherman, P.M., Muertter, R.N. and Edgar, R. (2009). NCBI GEO: archive forhigh-throughput functional genomic data Nucl. Acids Res. 37(suppl 1): D885-D890.
• Bengtsson, H., Simpson, K., Bullard, J., Hansen, K. (2008). aroma.affymetrix: A generic framework in R for analyzingsmall to very large Affymetrix data sets in bounded memory, Tech Report #745, Department of Statistics, University ofCalifornia, Berkeley.
• Ben-Porath, I., Thomson, M.W., Carey, V.J., Ge, R., Bell, G.W., Regev, A., Weinberg, R.A. (2008). An embryonic stemcell-like gene expression signature in poorly differentiated aggressive human tumors. Nat Genet. May. 40(5), 499-507.
• Boiani, M., Scholer, H.R. (2005). Regulatory networks in embryo-derived pluripotent stem cells., Nat Rev Mol Cell Biol.6(11), 872-84.
• Carpenter MK, Couture LA. (2010 Jul). Regulatory considerations for the development of autologous induced pluripotentstem cell therapies. Regen Med. 5(4), 569-579.
• Chen, J., Pei, D., He, W. (2009). Expression profile of early stage druing induced pluripotency regulated by BMP/FBS,GEO dataset GSE19604.
• Cheng, T., Rodrigues, N., Shen, H., Yang, Y., Dombkowski, D., Sykes, M., Scadden, D.T. (2000). Hematopoietic stemcell quiescence maintained by p21cip1/waf1. Science. 287, 1804-1808.
• Cho, H.J., Lee, C.S., Kwon, Y.W., Paek, J.S., Lee, S.H., Hur, J., Lee, E.J., Roh, T.Y., Chu, I.S., Leem, S.H., Kim, Y,.Kang, H.J., Park, Y.B., Kim, H.S. (2010). Induction of pluripotent stem cells from adult somatic cells by protein-basedreprogramming without genetic manipulation., Blood. 22;116(3):386-95.
• Collins, A.T., Berry, P.A., Hyde, C., Stower, M.J., Maitland, N.J. (2005). Prospective identification of tumorigenic prostatecancer stem cells. Cancer Res. 65, 10946-10951.
• Dontu, G., Abdallah, W.M., Foley, J.M., Jackson, K.W., Clarke, M.F., Kawamura, M.J., Wicha, M.S. (2003). Genes Dev.17, 1253-1270.

220 E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells
• Gerson, S.L., Reese, J., Kenyon, J. (2006). DNA repair in stem cell maintenance and conversion to cancer stem cells.Ernst Schering Found Symp Proc. 5, 231-44.
• He, S., Nakada, D., Morrison, S.J. (2007). Mechanisms of stem cell self-renewal. Annu Rev Cell Dev Biol. 2009. 25,377-406.
• Hess, P.G., (2009). Risk of tumorigenesis in first-in-human trials of embryonic stem cell neural derivatives: Ethics in theface of long-term uncertainty. Account Res. 16(4), 175-198.
• Huang, D.W., Sherman, B.T., Lempicki, R.A. (2009). Systematic and integrative analysis of large gene lists using DAVIDBioinformatics Resources. Nature Protoc. 4(1), 44-57.
• Kalirai, H., Clarke, R.B. (2006). Human breast epithelial stem cells and their regulation. J Pathol. 208, 7-16.• Kamalati, T., Niranjan, B., Yant, J., Buluwela, L. (1999). HGF/SF in mammary epithelial growth and morphogenesis: in
vitro and in vivo models. J Mammary Gland Biol Neoplasia. 4(1), 69-77.• Kang, L., Wang, J., Zhang, Y., Kou, Z., Gao, S. (2009). iPS Cells Can Support Full-Term Development of Tetraploid
Blastocyst-Complemented Embryos, Cell Stem Cell, 5 (2), 135-138.• Kippin, T.E., Martens, D.J., van der Kooy, D. (2005). p21 loss compromises the relative quiescence of forebrain stem cell
proliferation leading to exhaustion of their proliferation capacity. Genes and Development. 19, 756-67.• Jiao, X., Katiyar, S., Willmarth, N.E., Liu, M., Ma, X., Flomenberg, N., Lisanti, M.P., Pestell R.G. (2010). c-Jun induces
mammary epithelial cellular invasion and breast cancer stem cell expansion. J Biol Chem. 285(11), 8218-8226.• Irizarry, R.A., Bolstad, B.M., Collin, F., Cope, L.M., Hobbs, B., Speed, T.P. (2003). Summaries of Affymetrix GeneChip
probe level data. Nucleic Acids Research. 31(4), e15• Lefort, N., Feyeux, M., Bas, C., Feraud, O., Bennaceur-Griscelli, A., Tachdjian, G., Peschanski, M., Perrier, A.L. (2008).
Human embryonic stem cells reveal recurrent genomic instability at 20q11.21 Nat Biotechnol. 26(12):1364-6.• Li, C., Heidt, D.G., Dalerba, P., Burant, C.F., Zhang, L., Adsay, V., Wicha, M., Clarke, M.F., Simeone, D.M. (2007)
Identification of pancreatic cancer stem cells. Cancer Res. 67, 1030-1037.• Liras A, (2010). Future research and therapeutic applications of human stem cells: general, regulatory, and bioethical
aspects. J Transl Med. 10(8), 131.• McIntyre, J.C., Titlow, W.B., McClintock, T.S., (2010). Axon growth and guidance genes identify nascent, immature, and
mature olfactory sensory neurons. J. Neurosci Res. 15; 88(15), 3243-56.• Mukherjee, J., DeSouza, L.V., Micallef, J., Karim, Z., Croul, S., Siu, K.W., Guha, A. (2009). Loss of collapsin response
mediator Protein1, as detected by iTRAQ analysis, promotes invasion of human gliomas expressing mutant EGFRvIII.Cancer Res. 69(22), 8545-54.
• O’Brien, C.A., Pollett, A., Gallinger, S., Dick, J.E. (2007). A human colon cancer cell capable of initiating tumour growthin immunodeficient mice. Nature 445, 106-110.
• Owens, R.B., Smith, H.S., Hackett, A.J. (1974). Epithelial cell cultures from normal glandular tissue of mice. J NatlCancer Inst. 53(1), 261-9.
• Pan, S.H., Chao, Y.C., Chen, H.Y., Hung, P.F., Lin, P.Y., Lin, C.W., Chang, Y.L., Wu, C.T., Lee, Y.C., Yang, S.C., Hong,T.M., Yang, P.C. (2010). Long form collapsin response mediator protein-1 (LCRMP-1) expression is associated withclinical outcome and lymph node metastasis in non-small cell lung cancer patients. Lung Cancer. 67(1), 93-100.
• Ricci-Vitiani, L., Lombardi, D.G., Pilozzi, E., Biffoni, M., Todaro, M., Peschle, C., De Maria, R. (2007). Identificationand expansion of human colon-cancer initiating cells. Nature. 445, 111-115.
• Rochman, M., Postnikov, Y., Correll, S., Malicet, C., Wincovitch, S., Karpova, T.S., McNally, J.G., Wu, X., Bubunenko,N.A., Grigoryev, S., Bustin, M. (2009). The interaction of NSBP1/HMGN5 with nucleosomes in euchromatin counteractslinker histone-mediated chromatin compaction and modulates transcription., Mol Cell. 35(5), 642-56.
• Sakthianandeswaren A, Christie M, D’Andreti C, Tsui C, Jorissen RN, Li S, Fleming NI, Gibbs P, Lipton L, Malaterre J,Ramsay RG, Phesse TJ, Ernst M, Jeffery RE, Poulsom R, Leedham SJ, Segditsas S, Tomlinson IP, Bernhard OK, SimpsonRJ, Walker F, Faux MC, Church N, Catimel B, Flanagan DJ, Vincan E, Sieber OM. (2011). PHLDA1 expression marksthe putative epithelial stem cells and contributes to intestinal tumorigenesis. Cancer Res. 71, 3709-19.
• Shackleton, M., Vaillant, F., Simpson, K.J., Stingl, J., Smyth, G.K., Asselin-Labat, M.L., Wu, L., Lindeman, G.J., Visvader,J.E. (2006). Generation of a functional mammary gland from a single stem cell. Nature 439, 84-88.
• Soriano, J.V., Pepper, M.S., Nakamura, T., Orci L., Montesano, R. (1995). Hepatocyte growth factor stimulates extensivedevelopment of branching duct-like structures by cloned mammary gland epithelial cells. J Cell Sci. 108, 413-430.
• Soriano, J.V., Pepper, M.S., Orci, L., Montesano, R. (1998). Roles of hepatocyte growth factor/scatter factor andtransforming growth factor-beta1 in mammary gland ductal morphogenesis. J Mammary Gland Biol Neoplasia. 3(2),133-50.
• Stingl, J., Eirew, P., Ricketson, I., Shackleton, M., Vaillant, F., Choi, D., Li, H.I., Eaves, C.J. (2006). Purification andunique properties of mammary epithelial stem cells. Nature 439, 993-997.
• Takahashi, K., Yamanaka, S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblastcultures by defined factors. Cell. 126, 663-676.

E. Mosca et al. / Overlapping Genes May Control Reprogramming of Mouse Somatic Cells 221
• Utikal, J., Polo, J.M., Stadtfeld, M., Maherali, N., Kulalert, W., Walsh, R.M., Khalil, A., Rheinwald, J.G., Hochedlinger,K. (2009). Immortalization eliminates a roadblock during cellular reprogramming into iPS cells. Nature. 460(7259),1145-8.
• Woodward, W.A., Chen, M.S., Behbod, F., Rosen, J.M. (2005). On mammary stem cells. J Cell Sci. 118, 3585-3594.• Zucchi, I., Montagna, C., Susani, L., Vezzoni, P., Dulbecco, R. (1998). The rat gene homologous to the human gene 9-27
is involved in the development of the mammary gland. Proc Natl Acad Sci USA. 95, 1079-1084.• Zucchi, I., Astigiano, S., Bertalot, G., Sanzone, S., Cocola, C., Pelucchi, P., Bertoli, G., Stehling, M., Barbieri, O.,
Albertini, A., Scholer, H.R., Neel, B.G., Reinbold, R.A. and Dulbecco, R. (2008). Distinct populations of tumor-initiatingcells derived from a tumor generated by rat mammary cancer stem cells. Proc Natl Acad Sci USA. 105, 6940-16945.
• Zucchi, I., Bini, L., Albani, D., Valaperta, R., Liberatori, S., Raggiaschi, R., Montagna, C., Susani, L., Barbieri, O.,Pallini, V., Vezzoni, P., Dulbecco, R. (2002). Dome formation in cell cultures as expression of an early stage of lactogenicdifferentiation of the mammary gland. Proc Natl Acad Sci USA. 99(13), 8660-5. Epub 2002 Jun 19.
• Zucchi, I., Sanzone, S., Astigiano, S., Pelucchi, P., Scotti, M., Valsecchi, V., Barbieri, O., Bertoli, G., Albertini, A.,Reinbold R.A. and Dulbecco R. (2007). The properties of a mammary gland cancer stem cell. Proc Natl Acad Sci USA(2007). 104, 10476-10481.
Related Documents
![STEM CELLS EMBRYONIC STEM CELLS/INDUCED PLURIPOTENT STEM CELLS Stem Cells.pdf · germ cell production [2]. Human embryonic stem cells (hESCs) offer the means to further understand](https://static.cupdf.com/doc/110x72/6014b11f8ab8967916363675/stem-cells-embryonic-stem-cellsinduced-pluripotent-stem-cells-stem-cellspdf.jpg)










