MOLECULAR AND CELLULAR BIOLOGY, Jan. 1990, p. 217-222 Vol. 10, No. 1 0270-7306/90/010217-06$02.00/0 Copyright ©D 1990, American Society for Microbiology Overexpression of the STE4 Gene Leads to Mating Response in Haploid Saccharomyces cerevisiaett MALCOLM WHITEWAY,* LINDA HOUGAN, AND DAVID Y. THOMAS Genetic Engineering Section, National Research Council, Biotechnology Research Institute, 6100 Royalmount Avenue, Montreal, Quebec, Canada H4P 2R2 Received 10 July 1989/Accepted 6 October 1989 The STE4 gene of Saccharomyces cerevisiae encodes the I8 subunit of the yeast pheromone receptor-coupled G protein. Overexpression of the STE4 protein led to cell cycle arrest of haploid cells. This arrest was like the arrest mediated by mating pheromones in that it led to similar morphological changes in the arrested cells. The arrest occurred in haploid cells of either mating type but not in MATa/MATTa diploids, and it was suppressed by defects in genes such as STE12 that are needed for pheromone response. Overexpression of the STE4 gene product also suppressed the sterility of cells defective in the mating pheromone receptors encoded by the STE2 and STE3 genes. Cell cycle arrest mediated by STE4 overexpression was prevented in cells that either were overexpressing the SCGI gene product (the a subunit of the G protein) or lacked the STE18 gene product (the -y subunit of the G protein). This finding suggests that in yeast cells, the i subunit is the limiting component of the active ,(y element and that a proper balance in the levels of the G-protein subunits is critical to a normal mating pheromone response. The Saccharomyces cerevisiae mating pheromone re- sponse pathway has a remarkable similarity to G-protein- mediated signal transduction systems. Such systems were initially described in mammalian cells (see references 4, 8, 24, and 29 for recent reviews), where they were shown to consist of tripartite (a, 1, and -y subunits) guanine nucle- otide-binding proteins (G proteins) coupled to cell surface receptor proteins. In S. cerevisiae, the STE2 and STE3 genes encode the mating pheromone receptor proteins, which have seven potential membrane-spanning regions and thus are similar to mammalian G-protein-linked receptors (3, 9, 23). The GPAI (SCGI), STE4, and STE18 genes encode, respec- tively, the a, 1, and -y subunits of a G protein that is functionally coupled to these receptors (6, 22, 31). A current model for G-protein function in mammalian cells is that an interaction between the agonist and the receptor leads to guanine nucleotide exchange on the a subunit and the activated a subunit dissociates from the f3y subunit. The dissociated a subunit may then interact with an intracellular effector protein, such as adenylyl cyclase or cyclic GMP phosphodiesterase, and modulate its activity (8, 24, 29). Another model for some systems is that the free 13y subunit plays a role in effector modulation. This latter idea has been controversial (16, 33), but current evidence suggests that Py acts at least in the modulation of some phospholipase A2-mediated pathways (12, 13). Subsequent hydrolysis of the GTP on the a subunit allows reconstitution of the inactive atpy heterotrimer and turns the signaling pathway off (24). In yeast cells, the genetic evidence suggests that the 13y element acts positively in the mating pheromone response pathway. Disruption of either STE4 (I) or STE18 (-y) elimi- nates the ability of cells to respond to pheromones (33), whereas loss of the GPAJ (SCGI) protein (a) causes a constitutive mating response and a haploid-specific cell cycle * Corresponding author. t National Research Council Canada publication no. 30671. t This article is dedicated to the memory of Allen P. James, yeast geneticist. arrest (6, 11, 22). Both ste4 and stel8 mutations suppress this cell cycle arrest (33). Although there is currently no bio- chemical data on the mechanism of G-protein action in yeast cells, the structural similarities in the yeast and mammalian subunits and the functional distinctions between STE4 and STE18 on one hand and GPAI (SCGI) on the other make it likely the yeast mating response pathway functions similarly to the mammalian systems. Biochemical assessments of the roles of the various G- protein subunits in mammalian systems have treated the 1 and y proteins mainly as a single unit because they remain tightly associated throughout purification (24). In yeast cells, it was impossible to clone the STE4 gene from 2,um-based libraries (17), whereas the STE18 gene was readily obtained from such libraries (L. Hougan, unpublished data). This failure to isolate high-copy-number plasmids containing the STE4 gene suggests that high levels of the STE4 gene product may be detrimental to the cell, whereas high levels of the STE18 product are tolerated. Here, we investigate the phenotypic consequences of regulated overexpression of the STE4 gene by placing the STE4 coding sequence under the control of the GAL] promoter. Use of the GAL] promoter allows for facile manipulation of STE4 expression, since the promoter is efficiently induced when cells are grown on galactose but tightly repressed when cells are grown on glucose. Thus, cells can be grown in the absence of a potentially detrimental gene product and then induced to allow analysis of the results of the expression of this gene product. MATERIALS AND METHODS Strains. See Table 1 for a description of the yeast strains used. Plasmids. Plasmids pC3, which is a YEp13 plasmid con- taining the SCGI gene (6), and pJK6, which is a centromere plasmid containing the MFaJ gene under GAL] control (32; J. Kurjan, personal communication), were gifts of Janet Kurjan. Plasmid pSUL16 is YIp5 containing a LEU2 disrup- tion of STE12 (7) and was obtained from Stan Fields. Plasmids pSL376, which is pBR322 with a LEU2 disruption 217 on April 16, 2016 by guest http://mcb.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MOLECULAR AND CELLULAR BIOLOGY, Jan. 1990, p. 217-222 Vol. 10, No. 10270-7306/90/010217-06$02.00/0Copyright ©D 1990, American Society for Microbiology
Overexpression of the STE4 Gene Leads to Mating Response inHaploid Saccharomyces cerevisiaett
MALCOLM WHITEWAY,* LINDA HOUGAN, AND DAVID Y. THOMASGenetic Engineering Section, National Research Council, Biotechnology Research Institute, 6100 Royalmount Avenue,
Montreal, Quebec, Canada H4P 2R2
Received 10 July 1989/Accepted 6 October 1989
The STE4 gene of Saccharomyces cerevisiae encodes the I8 subunit of the yeast pheromone receptor-coupledG protein. Overexpression of the STE4 protein led to cell cycle arrest of haploid cells. This arrest was like thearrest mediated by mating pheromones in that it led to similar morphological changes in the arrested cells. Thearrest occurred in haploid cells of either mating type but not in MATa/MATTa diploids, and it was suppressedby defects in genes such as STE12 that are needed for pheromone response. Overexpression of the STE4 geneproduct also suppressed the sterility of cells defective in the mating pheromone receptors encoded by the STE2and STE3 genes. Cell cycle arrest mediated by STE4 overexpression was prevented in cells that either wereoverexpressing the SCGI gene product (the a subunit of the G protein) or lacked the STE18 gene product (the-y subunit of the G protein). This finding suggests that in yeast cells, the i subunit is the limiting componentof the active ,(y element and that a proper balance in the levels of the G-protein subunits is critical to a normalmating pheromone response.
The Saccharomyces cerevisiae mating pheromone re-sponse pathway has a remarkable similarity to G-protein-mediated signal transduction systems. Such systems wereinitially described in mammalian cells (see references 4, 8,24, and 29 for recent reviews), where they were shown toconsist of tripartite (a, 1, and -y subunits) guanine nucle-otide-binding proteins (G proteins) coupled to cell surfacereceptor proteins. In S. cerevisiae, the STE2 and STE3 genesencode the mating pheromone receptor proteins, which haveseven potential membrane-spanning regions and thus aresimilar to mammalian G-protein-linked receptors (3, 9, 23).The GPAI (SCGI), STE4, and STE18 genes encode, respec-tively, the a, 1, and -y subunits of a G protein that isfunctionally coupled to these receptors (6, 22, 31).A current model for G-protein function in mammalian cells
is that an interaction between the agonist and the receptorleads to guanine nucleotide exchange on the a subunit andthe activated a subunit dissociates from the f3y subunit. Thedissociated a subunit may then interact with an intracellulareffector protein, such as adenylyl cyclase or cyclic GMPphosphodiesterase, and modulate its activity (8, 24, 29).Another model for some systems is that the free 13y subunitplays a role in effector modulation. This latter idea has beencontroversial (16, 33), but current evidence suggests that Pyacts at least in the modulation of some phospholipaseA2-mediated pathways (12, 13). Subsequent hydrolysis ofthe GTP on the a subunit allows reconstitution of theinactive atpy heterotrimer and turns the signaling pathwayoff (24).
In yeast cells, the genetic evidence suggests that the 13yelement acts positively in the mating pheromone responsepathway. Disruption of either STE4 (I) or STE18 (-y) elimi-nates the ability of cells to respond to pheromones (33),whereas loss of the GPAJ (SCGI) protein (a) causes aconstitutive mating response and a haploid-specific cell cycle
* Corresponding author.t National Research Council Canada publication no. 30671.t This article is dedicated to the memory of Allen P. James, yeast
geneticist.
arrest (6, 11, 22). Both ste4 and stel8 mutations suppress thiscell cycle arrest (33). Although there is currently no bio-chemical data on the mechanism of G-protein action in yeastcells, the structural similarities in the yeast and mammaliansubunits and the functional distinctions between STE4 andSTE18 on one hand and GPAI (SCGI) on the other make itlikely the yeast mating response pathway functions similarlyto the mammalian systems.
Biochemical assessments of the roles of the various G-protein subunits in mammalian systems have treated the 1and y proteins mainly as a single unit because they remaintightly associated throughout purification (24). In yeast cells,it was impossible to clone the STE4 gene from 2,um-basedlibraries (17), whereas the STE18 gene was readily obtainedfrom such libraries (L. Hougan, unpublished data). Thisfailure to isolate high-copy-number plasmids containing theSTE4 gene suggests that high levels of the STE4 geneproduct may be detrimental to the cell, whereas high levelsof the STE18 product are tolerated.
Here, we investigate the phenotypic consequences ofregulated overexpression of the STE4 gene by placing theSTE4 coding sequence under the control of the GAL]promoter. Use of the GAL] promoter allows for facilemanipulation of STE4 expression, since the promoter isefficiently induced when cells are grown on galactose buttightly repressed when cells are grown on glucose. Thus,cells can be grown in the absence of a potentially detrimentalgene product and then induced to allow analysis of theresults of the expression of this gene product.
MATERIALS AND METHODSStrains. See Table 1 for a description of the yeast strains
used.Plasmids. Plasmids pC3, which is a YEp13 plasmid con-
taining the SCGI gene (6), and pJK6, which is a centromereplasmid containing the MFaJ gene under GAL] control (32;J. Kurjan, personal communication), were gifts of JanetKurjan. Plasmid pSUL16 is YIp5 containing a LEU2 disrup-tion of STE12 (7) and was obtained from Stan Fields.Plasmids pSL376, which is pBR322 with a LEU2 disruption
217
on April 16, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

218 WHITEWAY ET AL.
TABLE 1. Yeast strains
Strain Markers Source
Sc225 MATa adel leu2-3,2-112 ura3-52 J. HopperSc252 MATa adel leu2-3,2-112 ura3-52 J. HopperL1-3B MATa ura3 leu2-3,2-112, his3-11,3-15 This workL1-14D MATa ura3 leu2-3,2-112, his3-11,3-15 This workDL41 MMa adel ura3-52 leu2-3.2-112 + This work
MATa + ura3 leu2-3,2-112 his3-11,3-15
of STE3 (9), and pAB506, which is pBR322 containing aLEU2 disruption of STE2 (14), were obtained from KarenClark and Cathy Jackson, respectively. Plasmid YCp5O is acentromere-containing plasmid carrying the URA3 select-able marker (25). Plasmid M65pl is pTZ18R containing aLEU2 disruption of STE18 (31). Plasmid pL19 is a cen-tromere plasmid with STE4 under the control of the GAL]promoter and was constructed as follows. Plasmid M88p5,which consists of a 4.0-kilobase EcoRI fragment containingthe STE4 gene inserted into the EcoRI site of plasmidpTZ18R (20), was modified by oligonucleotide-directed mu-tagenesis. This placed an EcoRI and a BamHI site at the 5'end of the gene and a SalI site at the 3' end (Fig. 1). TheBamHI-to-Sall fragment was excised from this modifiedplasmid (M88L18) and cloned into the BamHI-to-SalI regionof plasmid pJK6 (Kurjan, personal communication). Thisreplaces the MFal sequences of pJK6 with STE4 andgenerates pLl9, a centromere plasmid carrying the STE4gene linked to the GALl promoter. Plasmid pL18-31 is aYEp213 plasmid containing the STE18 gene (L. Hougan,unpublished data). Recombinant DNA manipulations wereperformed by using standard protocols (18).Media and genetic methods. The media used have been
described previously (28, 32), and standard procedures wereused for strain constructions (28).
Transformation. Cells were transformed by recombinantplasmids or by DNA fragments by the lithium cation proto-col (10). Disruptions of STE2, STE3, STE12, and STE18 bythe LEU2 gene were constructed by one-step gene replace-ment (26). The STE2 disruption was created using a BamHIfragment containing a LEU2 replacement of STE2 obtainedfrom plasmid pAB506. The STE3 disruption fragment was aHindIll fragment from plasmid pSL376, the STE12 disrup-tion fragment was a SacI-to-SphI fragment from pSUL16,and the STE18 disruption used a PstI-to-HindIII fragmentcontaining a LEU2 replacement of STE18 derived fromM65pl.
Patch matings. Matings of strains containing plasmid pLl9were performed by growing the strains as patches on uracil-lacking (-Ura) glucose plates and then replica plating themto mating tester lawns on either yeast extract-peptone-galactose (YEPGAL) (for GALl promoter induction) oryeast extract-peptone-dextrose (YEPD) medium. Theseplates were incubated overnight at 30°C and then replica
STE45'GGTA CAC ATT ACG ATG GCA GCA CAT
IGAATTC T GGATCC
EcoRI BamHl
plated to minimal plates to detect the formation of pro-totrophic diploids.
Plasmid stability. Plasmid stability was measured by grow-ing strains overnight in YEPD or YEPGAL medium and thenspreading suitable dilutions on YEPD plates. After 2 days ofgrowth, the colonies were replica plated to drop-out mediumto detect colonies that retained the selectable marker char-acteristic of the plasmid under study. This procedure detectsthe loss of plasmids producing toxic proteins, since cellscontaining these plasmids will be at a growth disadvantage.RNA analysis. Total RNA was isolated from galactose-
induced and uninduced strains as described previously (19).Induced strains were grown in the presence of galactose asthe sole carbon source for 6 h; uninduced strains were grownfor an identical time in glucose-containing medium. TheRNA was denatured by boiling and run on a 1% agarose gelin TBE buffer, followed by direct hybridization with anend-labeled oligonucleotide probe to the dried gel (19).
Oligonucleotide-directed mutagenesis. Single-stranded DNAwas isolated from strain CJ236 (15) transformed withpTZ18R (20)-derived plasmids after infection with the helperbacteriophage M13KO7 as described elsewhere (30). Intro-duction of the 5' EcoRI and BamHI sites was achieved byannealing the oligonucleotide 5'-GGTACACATTACGiAATTCTFGGATCCATGGCAGCACAT-3' to plasmid M88p5and extending it with T4 DNA polymerase as describedpreviously (15). This generated plasmid M88L17, which wasthen mutagenized with oligonucleotide 5'-CTTCGAATTGGAGTCGACATTACTGTGAGC-3' to introduce a 3' Sall siteand create plasmid M88L18. Oligonucleotides were synthe-sized on an Applied Biosystems 380A DNA synthesizer andpurified as described previously (27). Correctly mutatedplasmids were identified by screening for the presence of thecreated restriction sites.
RESULTS
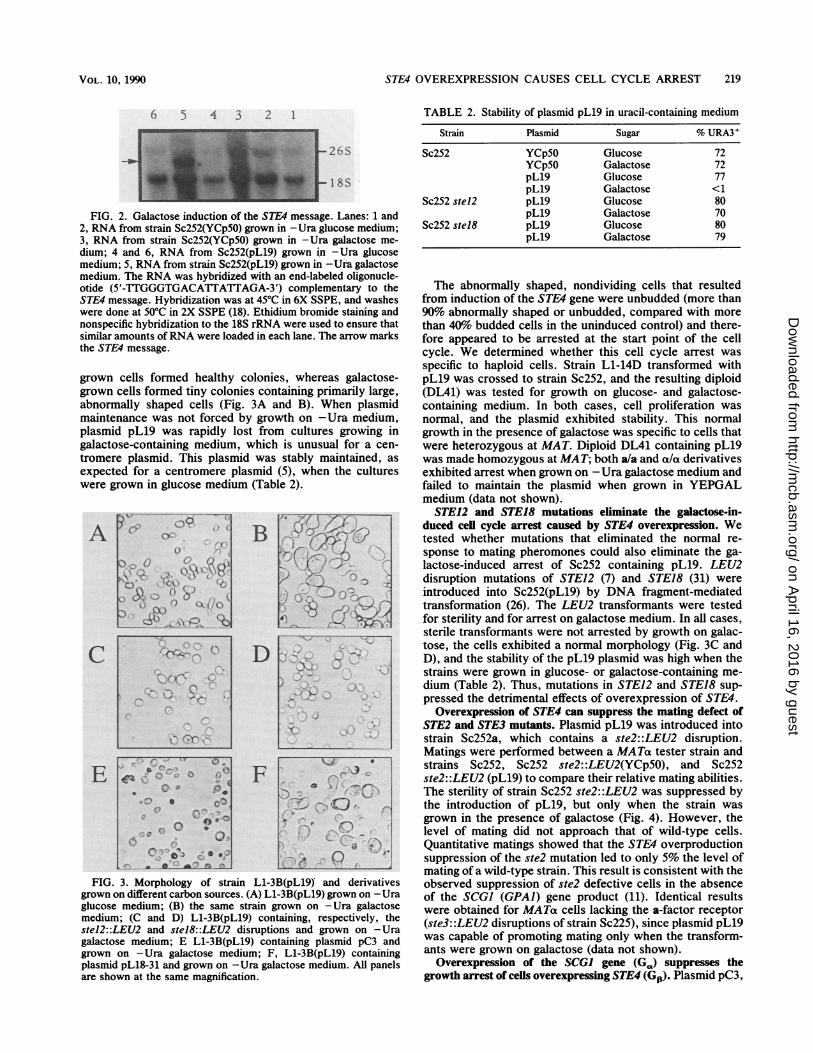
Construction of a galactose-inducible STE4 gene. We con-structed a derivative of the STE4 gene that replaced thenormal promoter and 5' regulatory sequences of this genewith the promoter of the GAL] gene. Northern (RNA)analysis was used to investigate the expression levels andregulation of the GAL] STE4-containing plasmid, designatedpL19. Total RNA was isolated from strain Sc252 containingeither pLl9 or YCp50 grown in medium containing eitherglucose or galactose. Growth in the presence of galactoseinduced a high level of the ST4-specific message only fromcells containing pL19 (Fig. 2).
Expression of STE4 under GAL control leads to haploid-specific cell cycle arrest. Plasmid pL19 was transformed intoa variety of ura3- GAL' strains, including Sc252, L1-3B,and L1-14D, and the phenotype of the resulting transfor-mants was checked after growth on medium containingeither glucose or galactose. When maintenance of the plas-mid was selected by growth on -Ura medium, glucose-
......TAG CTT CGA ATT GGA AAT ACT GTG AGCI
GTC GACSall
FIG. 1. Position of the newly created restriction sites in the modified STE4 gene. Oligonucleotides were used to direct the insertion ofEcoRl and BamHI sites at the 5' end of the STE4 gene and a Sall site downstream of the termination codon. The STE4 ATG and TAG codonsare shown in boldface.
MOL. CELL. BIOL.
on April 16, 2016 by guest
http://mcb.asm
.org/D
ownloaded from
STE4 OVEREXPRESSION CAUSES CELL CYCLE ARREST 219
6 5 4 3 2 1
26 S
18S
FIG. 2. Galactose induction of the STE4 message. Lanes: 1 and2, RNA from strain Sc252(YCp5O) grown in -Ura glucose medium;3, RNA from strain Sc252(YCp5O) grown in -Ura galactose me-
dium; 4 and 6, RNA from Sc252(pL19) grown in -Ura glucosemedium; 5, RNA from strain Sc252(pL19) grown in -Ura galactosemedium. The RNA was hybridized with an end-labeled oligonucle-otide (5'-TTGGGTGACATTATTAGA-3') complementary to theSTE4 message. Hybridization was at 45°C in 6X SSPE, and washeswere done at 50°C in 2X SSPE (18). Ethidium bromide staining andnonspecific hybridization to the 18S rRNA were used to ensure thatsimilar amounts of RNA were loaded in each lane. The arrow marksthe STE4 message.
grown cells formed healthy colonies, whereas galactose-grown cells formed tiny colonies containing primarily large,abnormally shaped cells (Fig. 3A and B). When plasmidmaintenance was not forced by growth on -Ura medium,plasmid pLl9 was rapidly lost from cultures growing ingalactose-containing medium, which is unusual for a cen-tromere plasmid. This plasmid was stably maintained, asexpected for a centromere plasmid (5), when the cultureswere grown in glucose medium (Table 2).
A
C
A) Cl\6(2(
D) -!g\:
B
D
F
FIG. 3. Morphology of strain L1-3B(pLl9) and derivativesgrown on different carbon sources. (A) L1-3B(pL19) grown on -Uraglucose medium; (B) the same strain grown on -Ura galactosemedium; (C and D) L1-3B(pL19) containing, respectively, thestel2::LEU2 and stel8::LEU2 disruptions and grown on -Uragalactose medium; E L1-3B(pLl9) containing plasmid pC3 andgrown on -Ura galactose medium; F, L1-3B(pLl9) containingplasmid pL18-31 and grown on -Ura galactose medium. All panelsare shown at the same magnification.
TABLE 2. Stability of plasmid pL19 in uracil-containing medium
Strain Plasmid Sugar % URA3+
Sc252 YCp50 Glucose 72YCp50 Galactose 72pL19 Glucose 77pL19 Galactose <1
Sc252 stel2 pL19 Glucose 80pL19 Galactose 70
Sc252 stel8 pL19 Glucose 80pL19 Galactose 79
The abnormally shaped, nondividing cells that resultedfrom induction of the STE4 gene were unbudded (more than90% abnormally shaped or unbudded, compared with morethan 40o budded cells in the uninduced control) and there-fore appeared to be arrested at the start point of the cellcycle. We determined whether this cell cycle arrest wasspecific to haploid cells. Strain L1-14D transformed withpLl9 was crossed to strain Sc252, and the resulting diploid(DL41) was tested for growth on glucose- and galactose-containing medium. In both cases, cell proliferation was
normal, and the plasmid exhibited stability. This normalgrowth in the presence of galactose was specific to cells thatwere heterozygous at MAT. Diploid DL41 containing pLl9was made homozygous at MAT; both a/a and ot/a derivativesexhibited arrest when grown on -Ura galactose medium andfailed to maintain the plasmid when grown in YEPGALmedium (data not shown).STE12 and STE18 mutations eliminate the galactose-in-
duced cell cycle arrest caused by STE4 overexpression. Wetested whether mutations that eliminated the normal re-sponse to mating pheromones could also eliminate the ga-lactose-induced arrest of Sc252 containing pLl9. LEU2disruption mutations of STE12 (7) and STE18 (31) were
introduced into Sc252(pLl9) by DNA fragment-mediatedtransformation (26). The LEU2 transformants were testedfor sterility and for arrest on galactose medium. In all cases,sterile transformants were not arrested by growth on galac-tose, the cells exhibited a normal morphology (Fig. 3C andD), and the stability of the pL19 plasmid was high when thestrains were grown in glucose- or galactose-containing me-dium (Table 2). Thus, mutations in STE12 and STE18 sup-pressed the detrimental effects of overexpression of STE4.
Overexpression of STE4 can suppress the mating defect ofSTE2 and STE3 mutants. Plasmid pL19 was introduced intostrain Sc252a, which contains a ste2::LEU2 disruption.Matings were performed between a MATca tester strain andstrains Sc252, Sc252 ste2: :LEU2(YCp5O), and Sc252ste2::LEU2 (pL19) to compare their relative mating abilities.The sterility of strain Sc252 ste2::LEU2 was suppressed bythe introduction of pL19, but only when the strain was
grown in the presence of galactose (Fig. 4). However, thelevel of mating did not approach that of wild-type cells.Quantitative matings showed that the STE4 overproductionsuppression of the ste2 mutation led to only 5% the level ofmating of a wild-type strain. This result is consistent with theobserved suppression of ste2 defective cells in the absenceof the SCGI (GPAI) gene product (11). Identical resultswere obtained for MATca cells lacking the a-factor receptor(ste3: :LEU2 disruptions of strain Sc225), since plasmid pL19was capable of promoting mating only when the transform-ants were grown on galactose (data not shown).
Overexpression of the SCGI gene (G.,) suppresses thegrowth arrest of cells overexpressing STE4 (G"). Plasmid pC3,
(7I
.- _
.- -. Ct l
'._.._ ._.._ _ .-__ _ ._.__
3
3I
0>f0A.,,
VOL. 10, 1990
on April 16, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

220 WHITEWAY ET AL.
Sc252[YCp50]
ste2::14EU2IYCIp501
ste2::L-EU2[pl.191
FIG. 4. Effect of the overproduction of STE4 on the mating of a ste2 mutant. Wild type (Sc252) and Sc252 ste2::LEU2 strains transformedwith either YCp5O or pL19 were grown on -Ura glucose medium to maintain the plasmid. They were then replicated to a MATa tester lawnon either YEPGAL (GAL) or YEPD (GLU) medium to allow mating, followed by replication to minimal medium to select prototrophicdiploids.
which contains the SCGI gene on the high-copy-number2,um-based vector YEp13 (6), was transformed into strainL1-3B containing the galactose-inducible STE4 plasmidpL19. In the absence of pC3, this strain arrested as morpho-logically abnormal cells when grown on galactose medium.When cultures were grown in YEPGAL medium, plasmidpL19 was rapidly lost. However, when the strain alsocontained plasmid pC3, the galactose-inducible arrest wassuppressed, and the cultures contained primarily morpho-logically normal cells (Fig. 3E). When these doubly trans-formed cells were grown on YEPGAL medium, both theURA3 marker defining pL19 and the LEU2 marker definingpC3 were maintained such that no colonies were formedfrom cells that contained only plasmid pL19. In contrast,when these transformants were grown on YEPD medium,the plasmids were independently lost (Table 3).
Increased expression of the STE18 (G,,) gene does notsuppress the growth arrest of cells overproducing STE4 (Ga).Plasmid pL18-31, which contains the STE18 gene cloned onthe high-copy-number 2,um-based plasmid YEp13 (2), wasintroduced into strain L1-3B(pL19). In contrast to the resultsobtained by overexpression of the a subunit, the increasedexpression of STE18 did not suppress the growth arrest ofthe strain when it was grown on galactose (Fig. 3F) and didnot stabilize plasmid pL19 during growth in YEPGAL me-dium (Table 3). In addition, pL18-31 by itself had no effecton the pheromone responsiveness of haploid yeast strains.Thus, the presumed overexpression of the yeast -y-subunitgene had no phenotypic consequences either in wild-typecells or in cells overexpressing the P-subunit gene, althoughthe actual levels of expression are unknown.
TABLE 3. Plasmid stabilities in strain L1-3B(pL19)
Plasmid Sugar % URA3+ % LEU2+
pL18-31 Glucose 64 21Galactose <1 4
pC3 Glucose 56 38Galactose 41a 53
a All URA3+ colonies were also LEU2+.
DISCUSSION
G proteins, whether in mammalian or yeast cells, appearto modulate effector pathways after the a subunit is disso-ciated from the Py element. In mammalian cells, this effectormodulation can be regulated either by the GTP-bound a
subunit (29) or by the Py element (12) and is shut off by
reassociation of the a^y heterotrimer after hydrolysis ofGTP to GDP (24). In yeast cells, the mating responsepathway is apparently activated by the f3y subunit of themating pheromone receptor-coupled G protein (31).
Previous work has shown that yeast strains defective inthe GPAI (SCGI) gene, which encodes the a subunit of theyeast mating pheromone response pathway G protein, con-stitutively express this mating response. As a consequence,mutations in the GPAI (SCGI) gene cause haploid-specificcell cycle arrest (6, 11, 22). This haploid-specific arrestpresumably results because the defective or missing a sub-unit is not capable of complexing with the active 13y subunitto shut off the pheromone response pathway. A dominantmutation within the STE4 gene, the yeast , subunit, can alsocause haploid-specific arrest. This mutation may affect thesite of a-P interaction and thus interfere with the formationof the transductionally inactive aI3y heterotrimer (1).
This work establishes that high levels of the wild-typeSTE4 gene product created an equivalent constitutive matingresponse phenotype. Galactose-induced overexpression ofthe P subunit caused haploid-specific cell cycle arrest andsuppressed the sterility of yeast cells with defects in thepheromone receptor proteins. This finding suggests that theSTE4 gene product is normally a limiting component in theresponse pathway in the absence of pheromone; in haploidwild-type cells, increased expression of the G,3 subunitalone, in the absence of any pheromone, leads to an activa-tion of the pheromone response pathway.One interpretation of this observation is that the STE4
protein is capable of directly modulating the effector of themating response pathway, so free p is capable of activatingthe pheromone response signal. However, overexpression ofthe STE4 gene product does not lead to cell cycle arrest incells that lack the STE18 gene. Thus, the y subunit of theyeast G protein has an essential role in the mating response
- URA GAL L.U
MOL. CELL. BIOL.
on April 16, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

STE4 OVEREXPRESSION CAUSES CELL CYCLE ARREST 221
pathway that is not eliminated by overproduction of the fisubunit. These observations can be explained if the -yelement is the effector modulator and if -y is normally inexcess in the cell, so that the level of functional P3y isprimarily a reflection of the level of the 1B subunit. Alterna-tively, P and fy may be at stoichiometric levels, but if free 1is capable of complexing with a, overexpression of 1B wouldfree some Py effector modulator from a repression. In eithercase, the results provide strong evidence for an essential rolefor both 1 and y in the activation of the response pathway.Previous biochemical manipulations of purified G-proteinsubunits have always used combined f3y proteins becausethe 13 and -y subunits remain tightly associated during isola-tion (24). Our genetic manipulations have allowed an exam-ination of the effect of excess 13 in the presence and absenceof y and support the implicit assumption of these previousstudies that the 1 and -y subunits form a functional unit.The phenotypic consequences of overproduction of the
STE4 gene product are consistent with the suggestion thatthe 13y element acts as the positive element in the pheromoneresponse pathway and that this element is antagonized bythe a subunit. Although there is no biochemical evidence asto the actual levels of GDP or GTP bound to the a subunit,all manipulations that potentially either decrease the level ofthe aGDP subunit (6, 11, 22) or increase the level of either the13y element (this study) or the aGTP subunit (21) lead to aconstitutive mating response or enhance pheromone respon-siveness, respectively. Conversely, potential increases inthe level of the aGDP subunit, either in wild-type or sst2mutant cells (6) or in cells overexpressing 13y (this study),reduce pheromone responsiveness or suppress the constitu-tive response signal.We would anticipate a sensitivity to the imbalance of
G-protein subunit levels shown in this study to be charac-teristic of all G-protein systems because efficient activation-deactivation cycles should depend on the presence of the aand 13y components at similar concentrations. It may beinformative to see the consequences of similar manipulationsin mammalian cells, although the variety of independent GQ,subunits will be a complicating factor. For example, theconstruction of cell lines or transgenic animals containingG-protein subunits under the control of strong regulatablepromoters may shed some light on the varied roles of Gproteins in higher organisms by activating or deactivating theresponse pathways under the control of a particular Gprotein.
ACKNOWLEDGMENTS
We thank J. Kurjan, S. Fields, K. Clark, C. Jackson, and J.Hopper for their generous gifts of plasmids and strains and T.Vernet and F. Laliberte for advice and assistance with the site-directed mutagenesis. We also thank Carl Juby for the synthesis ofoligonucleotides and M. Dennis for insightful comments on themanuscript.
LITERATURE CITED
1. Blinder, D., S. Bouvier, and D. D. Jenness. 1989. Constitutivemutants in the yeast pheromone response: ordered function ofthe gene products. Cell 56:479-486.
2. Broach, J. R., J. N. Strathern, and J. B. Hicks. 1979. Transfor-mation in yeast: development of a hybrid cloning vector andisolation of the CAN] gene. Gene 8:121-133.
3. Burkholder, A. C., and L. H. Hartwell. 1985. The yeast a-factorreceptor: structural properties deduced from the sequence ofthe STE2 gene. Nucleic Acids Res. 13:8463-8475.
4. Casey, P. J., and A. G. Gilman. 1988. G protein involvement in
receptor-effector coupling. J. Biol. Chem. 263:2577-2580.5. Clarke, L., and J. Carbon. 1980. Isolation of a yeast centromere
and construction of functional small circular chromosomes.Nature (London) 187:504-509.
6. Dietzel, C., and J. Kurjan. 1987. The yeast SCGI gene: a G,,-likeprotein implicated in the a- and a-factor response pathway. Cell50:1001-1010.
7. Fields, S., and I. Herskowitz. 1987. Regulation by the mating-type locus of STE12, a gene required for cell-type-specificexpression. Mol. Cell. Biol. 7:3818-3821.
8. Gilman, A. 1987. G proteins: transducers of receptor-generatedsignals. Annu. Rev. Biochem. 56:615-649.
9. Hagen, D. C., G. McCaffrey, and G. F. Sprague, Jr. 1986.Evidence the yeast STE3 gene encodes a receptor for thepeptide pheromone a-factor: gene sequence and implications forthe structure of the presumed receptor. Proc. Natl. Acad. Sci.USA 83:1418-1422.
10. Ito, H., Y. Fukuda, K. Murata, and A. Kimura. 1983. Transfor-mation of intact yeast cells treated with alkali cations. J.Bacteriol. 153:163-168.
11. Jahng, K. -Y., J. Ferguson, and S. I. Reed. 1988. Mutations in agene encoding the a subunit of a Saccharomyces cerevisiae Gprotein indicate a role in mating pheromone signaling. Mol. Cell.Biol. 8:2484-2493.
12. Jelsema, C. L., and J. Axelrod. 1987. Stimulation of phospholi-pase A2 activity in bovine rod outer segments by the 13y subunitsof transducin and its inhibition by the a subunit. Proc. Natl.Acad. Sci. USA. 84:3623-3627.
13. Kim, D., D. L. Lewis, L. Graziadei, E. J. Neer, D. Bar-Sagi, andD. E. Clapham. 1989. G protein 13y-subunits activate the cardiacmuscarinic K'-channel via phospholipase A2. Nature (London)337:557-560.
14. Konopka, J. B., D. D. Jenness, and L. H. Hartwell. 1988. The Cterminus of the S. cerevisiae a pheromone receptor mediates anadaptive response to pheromone. Cell 54:609-620.
15. Kunkle, T. A., J. D. Roberts, and R. A. Zakour. 1987. Rapid andefficient site-specific mutagenesis without phenotypic selection.Methods Enzymol. 154:367-382.
16. Logothetis, D. E., Y. Kurachi, J. Galper, E. J. Neer, and D. E.Clapham. 1987. The 03y subunits of GTP-binding proteins acti-vate the muscarinic K' channel in heart. Nature (London)325:321-326.
17. MacKay, V. L. 1983. Cloning of yeast STE genes in 2,u vectors.Methods Enzymol. 101:325-343.
18. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
19. McIntosh, E. M., R. W. Ord, and R. K. Storms. 1988. Tran-scriptional regulation of the cell cycle-dependent thymidylatesynthase gene of Saccharomyces cerevisiae. Mol. Cell. Biol.8:4616-4624.
20. Mead, D. A., E. Szczesna-Skorpa, and B. Kemper. 1986. Single-stranded DNA "blue" T7 promoter plasmids: a versatile tan-dem promoter system for cloning and protein engineering.Protein Eng. 1:67-74.
21. Miyajina, I., K.-I. Arai, and K. Matsumoto. 1989. GPA1vav-50mutation in the mating-factor signaling pathway in Saccharomy-ces cerevisiae. Mol. Cell. Biol. 9:2289-2297.
22. Miyajima, I., M. Nakafuku, N. Nakayama, C. Brenner, A.Miyajima, K. Kaibuchi, K. Arai, Y. Kaziro, and K. Matsumoto.1987. GPAI, a haploid essential gene, encodes a yeast homologof mammalian G protein which may be involved in mating factorsignal transduction. Cell 50:1011-1019.
23. Nakayama, N., A. Miyajima, and K. Arai. 1985. Nucleotidesequences of STE2 and STE3, cell type specific sterile genesfrom Saccharomyces cerevisiae. EMBO J. 4:2643-2648.
24. Neer, E. J., and D. E. Clapham. 1988. Roles of G proteinsubunits in transmembrane signaling. Nature (London) 333:129-134.
25. Rose, M. D., P. Novick, J. H. Thomas, D. Botstein, and G. R.
VOL. 10, 1990
on April 16, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

222 WHITEWAY ET AL. MOL. CELL. BIOL.
Fink. 1987. A Saccharomyces cerevisiae genomic plasmid bankbased on a centromere containing shuttle vector. Gene 60:237-243.
26. Rothstein, R. 1983. One step gene disruption in yeast. MethodsEnzymol. 101:202-211.
27. Sanchez-Pescador, R., and M. S. Urdea. 1984. Use of unpurifiedsynthetic deoxynucleotide primers for rapid dideoxynucleotidechain termination sequencing. DNA 3:339-343.
28. Sherman, F., G. R. Fink, and J. B. Hicks. 1982. Methods inyeast genetics. Cold Harbor Laboratory, Cold Spring Harbor,N.Y.
29. Stryer, L., and H. R. Bourne. 1986. G proteins: a family of signaltransducers. Annu. Rev. Cell Biol. 2:391-419.
30. Vernet, T., D. Dignard, and D. Y. Thomas. 1987. A family of
yeast expression vectors containing the phage fl origin ofreplication. Gene 52:225-233.
31. Whiteway, M. S., L. Hougan, D. Dignard, D. Y. Thomas, L. Bell,G. C. Saari, F. J. Grant, P. O'Hara, and V. L. MacKay. 1989.The STE4 and STE18 genes of yeast encode potential ,B and ysubunits of the mating factor receptor-coupled G protein. Cell56:467-477.
32. Whiteway, M. S., L. Hougan, and D. Y. Thomas. 1988. Expres-sion of MFal in MATa cells supersensitive to a-factor leads toself-arrest. Mol. Gen. Genet. 214:85-88.
33. Yatani, A., H. Hamm, J. Codina, M. R. Mazzoni, L. Birn-baumer, and A. M. Brown. 1988. A monoclonal antibody to thea subunit of Gk blocks muscarinic activation of atrial K+channels. Science 241:828-831.
on April 16, 2016 by guest
http://mcb.asm
.org/D
ownloaded from
Related Documents











