
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Overexpression of the FosB2 gene in thymocytes causes aberrant
development of T cells and thymic epithelial cells
Maria Luisa Carrozza1,3, Heinz Jacobs1, Dennis Acton1, Inder Verma2 and Anton Berns1
1Division of Molecular Genetics of the Netherlands Cancer Institute, Plesmanlaan 121, 1066 CX, Amsterdam,
The Netherlands; 2The Salk Institute for Biological Studies, Torry Pines Road, La Jolla, CA, USA
We have examined the role of the AP-1 transcription
factor on thymocyte maturation and thymus architecture
by overexpressing FosB2 in transgenic mice. FosB2 is a
naturally occurring splice variant of the FosB2 gene,
encoding a truncated protein which lacks two domains
necessary for transcriptional activation. The expression
of FosB2 in the thymocytes severely a�ected their
maturation and the structure of the whole thymus: the
phenotype developed slowly during the ®rst months of
life, resulting in a progressive expansion of the medulla
and concomitant reduction of the cortex. CD4+
thymocytes represented the major thymocyte population,
whereas the CD4+8+ thymocytes were virtually absent.
This phenotype appeared to be an intrinsic property of
bone marrow derived cells, as it could be reproduced in
bone marrow chimaeric mice. This pathology was very
reminiscent to that observed in mice overexpressing c-
Fos in thymic epithelium: also in that case the thymus
underwent with age a progressive expansion of the
epithelium and major changes in the ratio of thymocyte
subsets, but the phenotype appeared to be an intrinsic
property of the epithelial cells since it could not be
reproduced by transgenic bone marrow transplantation.
We speculate that both overexpression of FosB2 in
thymocytes and overexpression of c-Fos in thymic
epithelium results in aberrant signaling between thymo-
cytes and stroma, that ultimately alters the thymic
micromilieu, leading to this severe pathology.
Keywords: oncogene; thymus; development
Introduction
Generation of a diverse repertoire of T lymphocytes isa complex process which occurs in ± and depends on ±the specialized thymic microenvironment (Boyd andHugo, 1991; Zugic et al., 1991; Boyd et al., 1993).Bone-marrow derived progenitor cells enter the thymusat the cortico-medullary junction or subcapsular regionand di�erentiate during their migration through thecortex to the central medulla, where mature T cells exitinto the periphery. The thymocytes and the surround-ing stromal cells, which together create the specializedmicroenvironments of the di�erent thymic compart-ments, are bound by a symbiotic relationship: their
interaction is necessary both for thymocyte maturationand for correct development and maintenance of thestroma (Shores et al., 1991; Surh et al., 1992; Ritterand Boyd, 1993; van Ewijk et al., 1994; Bosma et al.,1983; Philpott et al., 1992; Zijlstra et al., 1990;Vukmanovich et al., 1992). A variety of moleculeshave been identi®ed, which are important for thesetwo-way cellular interactions: they include cell surfaceand extracellular matrix molecules, cytokines, recep-tors, the second messengers they activate, and c-Fos, aconstituent of the AP-1 transcription factor (Ritter &Boyd, 1993; Galy et al., 1992; Wolf & Cohen, 1993;Couture et al., 1992; RuÈ ther et al., 1988).AP-1 consists predominantly of Fos and Jun
proteins which dimerize through their leucine zipperdomains (for review see Ransone et al., 1990). Bothproteins are encoded by gene families, whose membersshow a high degree of homology at the gene andprotein level, are di�erently expressed during the cellcycle and exhibit crossregulatory activity in di�erentcell types and tissues (SchuÈ ttle et al., 1989; Yen et al.,1991). The various Fos and Jun heterodimers bind withsimilar a�nity in vitro to the target DNA sequence, theAP-1 binding site, also known as TRE (TPAresponsive element; Lee et al., 1987; Curran andFranza, 1988).In a variety of cell types in vivo and in vitro, AP-1
acts as a `main switch', controlling diverse cellularfunctions, such as proliferation (Greenberg and Zi�,1984), di�erentiation (MuÈ ller and Wagner, 1984;Morgan and Curran, 1991) and programmed celldeath (Smeyne et al., 1993). Some of the Fos andJun genes are expressed in the adult thymus (MuÈ ller,1986; Hirai et al., 1989). During T cell activation theycontrol gene expression and are found complexed withthe inducible nuclear factor NF-AT (Jain et al., 1993).Overexpression of c-Fos in the thymic epithelium of
transgenic mice results in profound alterations of thethymic structure and causes major shifts in the relativeproportions of the di�erent thymocyte subpopulations(RuÈ ther et al., 1988). Although changes in thymocytesubsets are also seen in fos-de®cient mice they arelikely the result of the poor health status of these mice(Okada et al., 1994).We wanted to study the role of AP-1 in thymocyte
maturation. Therefore, we have generated transgenicmice overexpressing FosB2 in the thymocytes. FosB2is a splice variant of FosB, ®rst detected in serumstimulated 3T3 mouse ®broblasts (Nakabeppu andNathans, 1991; Yen et al., 1991). It encodes a proteinof 237 amino acids, which has retained the DNA-binding basic region and the leucin-zipper dimeriza-tion domain of the intact FosB protein, but lacks theheptaproline motif (amino acid residues 257 to 263)
Correspondence: ML Carrozza3Present address: 1st Di Neuro®siologia Del Cnr, Via S. Zeno 51,Pisa, ItalyReceived 10 June 1996; revised 21 October 1996; accepted 29 October1996
Oncogene (1997) 14, 1083 ± 1091
1997 Stockton Press All rights reserved 0950 ± 9232/97 $12.00

which functions as a transactivation domain (Yen et
al., 1991; Wisdom et al., 1992) and the TBP (TATAbinding protein) binding motif, which is necessary forconferring full transcriptional activity (Metz et al.,1994).FosB2 is unable to activate transcription in F9 cells
(Nakabeppu and Nathans, 1991; Yen et al., 1991) andis a weak transcriptional activator in NIH3T3 and inHeLa cells (Mumberg et al., 1991; Dobrzanski et al.,1991). It is also unable to transform cells and acts in adominant negative fashion in transrepression andtransforming activity of the Fos, FosB and Junproteins (Nakabeppu and Nathans, 1991; Yen et al.,1991; Mumberg et al., 1991; Wisdom et al., 1992). Wereasoned that its overexpression might result in aphenotype which otherwise only might be achieved bythe inactivation of the multiple endogenous Fos genes.The expression of the FosB2 gene in the thymocytes
of transgenic mice dramatically a�ected the thymocytesand the thymic stroma. This phenotype was an intrinsicproperty of the haematopoietic cells as it could bereproduced in bone marrow chimeric mice.
Results
Generation of transgenic mice
Microinjection of the 4.8 kb SalI-ClaI fragment ofpTCDFosB, spanning the FosB2 cDNA and the T cellspeci®c promoter (TCRb) and enhancer (CD2) (Figure1), into fertilized oocytes resulted in ®ve founder mice,each with di�erent copy numbers of the construct. Twofemale founders, carrying high copy numbers, weremuch smaller than normal FVB mice, did not produceany o�spring, became sick and were sacri®ced. Theirovaries showed defective follicle development. Weestablished lines of the remaining three founders: line7785 with a low, line 7789 with an intermediate, andline 7787 with a high copy number (Figure 2a).Northern blot analysis of total RNA from di�erenttissues of the transgenic mice showed expression of thetransgene exclusively in thymus and spleen. Theexpression level was proportional to the copy number(Figure 2b), as was previously shown for CD2enhancer harbouring constructs (Greaves et al., 1989).
Phenotype of the FosB2 transgenic mice
Transgenic female mice of line 7787 and 7789 were ingeneral smaller than their non transgenic littermates,and were often not fertile or produced very smalllitters. Female o�spring of line 7785 were normal in
size. Male transgenic mice were slightly smaller thantheir normal littermates. The reduction in sizecorrelated with the expression level of the FosB2transgene. Both males and females did not show signof disease during the ®rst months of life, but becamesick at an age between 3 and 7 months. The health ofthe high copy number mice (line 7787 and 7789)deteriorated fast, whereas mice with low copy number(line 7785) remained in steady suboptimal healthconditions over longer periods. Sick animals werereported to su�er from anemia, from breathingproblems, and to have, in some cases, enlargedsubmaxillary glands. Whereas breathing problemscould relate to the vastly enlarged thymuses indiseased animals, and the anemia to the disturbancesin the haematopoietic organs, some of the otherpathological abnormalities are more di�cult to relateto the transgene. Sick mice were sacri®ced andhistological analysis was performed: the only grossabnormality found was a profoundly enlarged thymus.As at the time of sacri®ce the animals were at the endof the pathological process, we decided to follow thecourse of the disease by analysing the mice at di�erenttime points. Nontransgenic littermates were chosen asage-matched controls.
Histological analysis of the thymus
Thymuses of sick mice showed a dramatically alteredphenotype. The organ was signi®cantly enlarged ascompared to age-matched controls, maintaining a large
Figure 1 The FosB2 targeting construct. The 4.8 kb Sal l ±Cla l fragment was excised from the vector and microinjected intofertilized oocytes of the FVB strain. TCRb prom.=the promoter of the v gene segment of the b chain of the TCR. FosB2=the0.9 kb coding region of the FosB2 cDNA. pA=SV40 polyadenylation sequence. CD2 enhancer=enhancer sequence of the genecoding for the T cell speci®c, cell surface associated CD2 glycoprotein
1 2
1 2 3
1.4 kb
2.1 kb
0.7 kb
➝
➝
➝
a b
Figure 2 Southern and Northern blot analysis of DNA andRNA from line 7785 and line 7787. (a) Pst 1 digested tail DNA,hybridized to the murine FosB2 speci®c cNDA probe. Only theregion of the autoradiograph in which the transgenic fragmentsmigrate is shown. The 2.1 kb and the 0.7 kb bands are transgenespeci®c. lane 1: lane 7785 mouse; lane 2: line 7787 mouse. (b)Total RNA from thymus, hybridized to the FosB2 speci®c cDNAprobe. lane 1: line 7785 mouse; lane 2: line 7787 mouse; lane 3:control mouse. The size of the transgene mRNA is about 1.4 kb
Fos B2 expression affects thymus development
ML Carrozza et al
1084
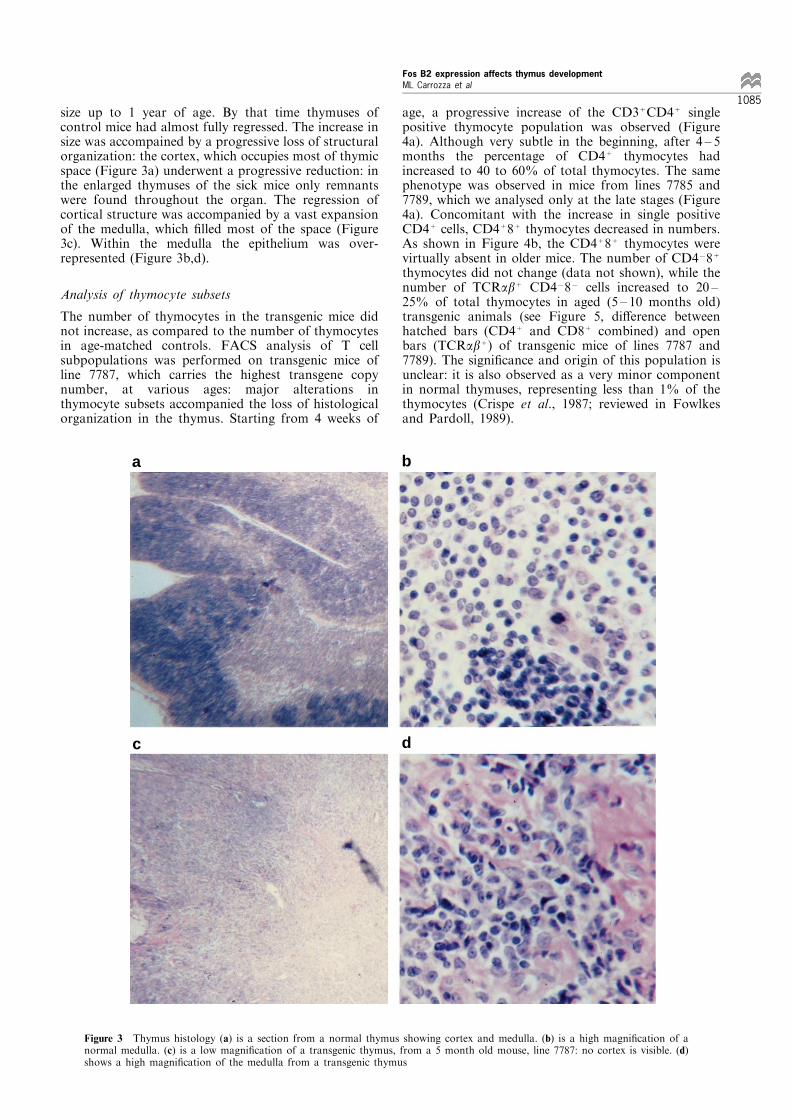
size up to 1 year of age. By that time thymuses ofcontrol mice had almost fully regressed. The increase insize was accompained by a progressive loss of structuralorganization: the cortex, which occupies most of thymicspace (Figure 3a) underwent a progressive reduction: inthe enlarged thymuses of the sick mice only remnantswere found throughout the organ. The regression ofcortical structure was accompanied by a vast expansionof the medulla, which ®lled most of the space (Figure3c). Within the medulla the epithelium was over-represented (Figure 3b,d).
Analysis of thymocyte subsets
The number of thymocytes in the transgenic mice didnot increase, as compared to the number of thymocytesin age-matched controls. FACS analysis of T cellsubpopulations was performed on transgenic mice ofline 7787, which carries the highest transgene copynumber, at various ages: major alterations inthymocyte subsets accompanied the loss of histologicalorganization in the thymus. Starting from 4 weeks of
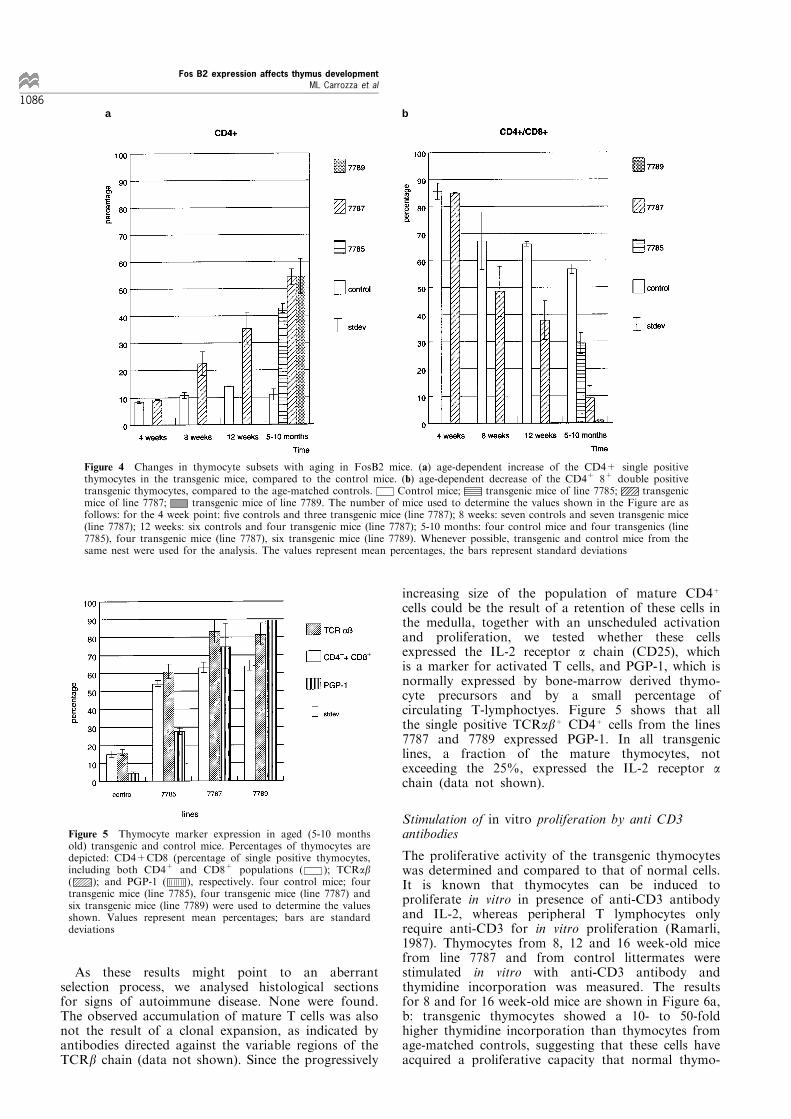
age, a progressive increase of the CD3+CD4+ singlepositive thymocyte population was observed (Figure4a). Although very subtle in the beginning, after 4 ± 5months the percentage of CD4+ thymocytes hadincreased to 40 to 60% of total thymocytes. The samephenotype was observed in mice from lines 7785 and7789, which we analysed only at the late stages (Figure4a). Concomitant with the increase in single positiveCD4+ cells, CD4+8+ thymocytes decreased in numbers.As shown in Figure 4b, the CD4+8+ thymocytes werevirtually absent in older mice. The number of CD478+
thymocytes did not change (data not shown), while thenumber of TCRab+ CD4787 cells increased to 20 ±25% of total thymocytes in aged (5 ± 10 months old)transgenic animals (see Figure 5, di�erence betweenhatched bars (CD4+ and CD8+ combined) and openbars (TCRab+) of transgenic mice of lines 7787 and7789). The signi®cance and origin of this population isunclear: it is also observed as a very minor componentin normal thymuses, representing less than 1% of thethymocytes (Crispe et al., 1987; reviewed in Fowlkesand Pardoll, 1989).
a b
dc
Figure 3 Thymus histology (a) is a section from a normal thymus showing cortex and medulla. (b) is a high magni®cation of anormal medulla. (c) is a low magni®cation of a transgenic thymus, from a 5 month old mouse, line 7787: no cortex is visible. (d)shows a high magni®cation of the medulla from a transgenic thymus
Fos B2 expression affects thymus development
ML Carrozza et al
1085
As these results might point to an aberrantselection process, we analysed histological sectionsfor signs of autoimmune disease. None were found.The observed accumulation of mature T cells was alsonot the result of a clonal expansion, as indicated byantibodies directed against the variable regions of theTCRb chain (data not shown). Since the progressively
increasing size of the population of mature CD4+
cells could be the result of a retention of these cells inthe medulla, together with an unscheduled activationand proliferation, we tested whether these cellsexpressed the IL-2 receptor a chain (CD25), whichis a marker for activated T cells, and PGP-1, which isnormally expressed by bone-marrow derived thymo-cyte precursors and by a small percentage ofcirculating T-lymphoctyes. Figure 5 shows that allthe single positive TCRab+ CD4+ cells from the lines7787 and 7789 expressed PGP-1. In all transgeniclines, a fraction of the mature thymocytes, notexceeding the 25%, expressed the IL-2 receptor a
chain (data not shown).
Stimulation of in vitro proliferation by anti CD3
antibodies
The proliferative activity of the transgenic thymocyteswas determined and compared to that of normal cells.It is known that thymocytes can be induced toproliferate in vitro in presence of anti-CD3 antibodyand IL-2, whereas peripheral T lymphocytes onlyrequire anti-CD3 for in vitro proliferation (Ramarli,1987). Thymocytes from 8, 12 and 16 week-old micefrom line 7787 and from control littermates werestimulated in vitro with anti-CD3 antibody andthymidine incorporation was measured. The resultsfor 8 and for 16 week-old mice are shown in Figure 6a,b: transgenic thymocytes showed a 10- to 50-foldhigher thymidine incorporation than thymocytes fromage-matched controls, suggesting that these cells haveacquired a proliferative capacity that normal thymo-
ba
Figure 4 Changes in thymocyte subsets with aging in FosB2 mice. (a) age-dependent increase of the CD4+ single positivethymocytes in the transgenic mice, compared to the control mice. (b) age-dependent decrease of the CD4+ 8+ double positivetransgenic thymocytes, compared to the age-matched controls. Control mice; transgenic mice of line 7785; transgenicmice of line 7787; transgenic mice of line 7789. The number of mice used to determine the values shown in the Figure are asfollows: for the 4 week point: ®ve controls and three transgenic mice (line 7787); 8 weeks: seven controls and seven transgenic mice(line 7787); 12 weeks: six controls and four transgenic mice (line 7787); 5-10 months: four control mice and four transgenics (line7785), four transgenic mice (line 7787), six transgenic mice (line 7789). Whenever possible, transgenic and control mice from thesame nest were used for the analysis. The values represent mean percentages, the bars represent standard deviations
Figure 5 Thymocyte marker expression in aged (5-10 monthsold) transgenic and control mice. Percentages of thymocytes aredepicted: CD4+CD8 (percentage of single positive thymocytes,including both CD4+ and CD8+ populations ( ); TCRab( ); and PGP-1 ( ), respectively. four control mice; fourtransgenic mice (line 7785), four transgenic mice (line 7787) andsix transgenic mice (line 7789) were used to determine the valuesshown. Values represent mean percentages; bars are standarddeviations
Fos B2 expression affects thymus development
ML Carrozza et al
1086
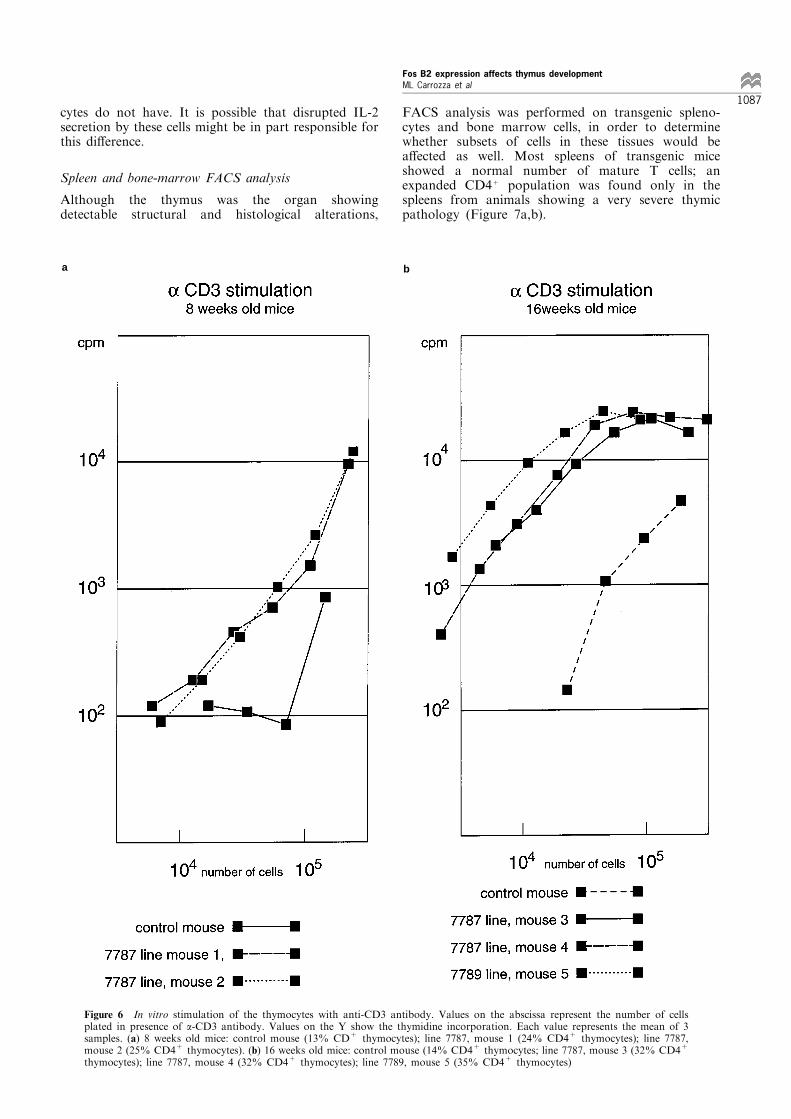
cytes do not have. It is possible that disrupted IL-2secretion by these cells might be in part responsible forthis di�erence.
Spleen and bone-marrow FACS analysis
Although the thymus was the organ showingdetectable structural and histological alterations,
FACS analysis was performed on transgenic spleno-cytes and bone marrow cells, in order to determinewhether subsets of cells in these tissues would bea�ected as well. Most spleens of transgenic miceshowed a normal number of mature T cells; anexpanded CD4+ population was found only in thespleens from animals showing a very severe thymicpathology (Figure 7a,b).
a b
Figure 6 In vitro stimulation of the thymocytes with anti-CD3 antibody. Values on the abscissa represent the number of cellsplated in presence of a-CD3 antibody. Values on the Y show the thymidine incorporation. Each value represents the mean of 3samples. (a) 8 weeks old mice: control mouse (13% CD+ thymocytes); line 7787, mouse 1 (24% CD4+ thymocytes); line 7787,mouse 2 (25% CD4+ thymocytes). (b) 16 weeks old mice: control mouse (14% CD4+ thymocytes; line 7787, mouse 3 (32% CD4+
thymocytes); line 7787, mouse 4 (32% CD4+ thymocytes); line 7789, mouse 5 (35% CD4+ thymocytes)
Fos B2 expression affects thymus development
ML Carrozza et al
1087
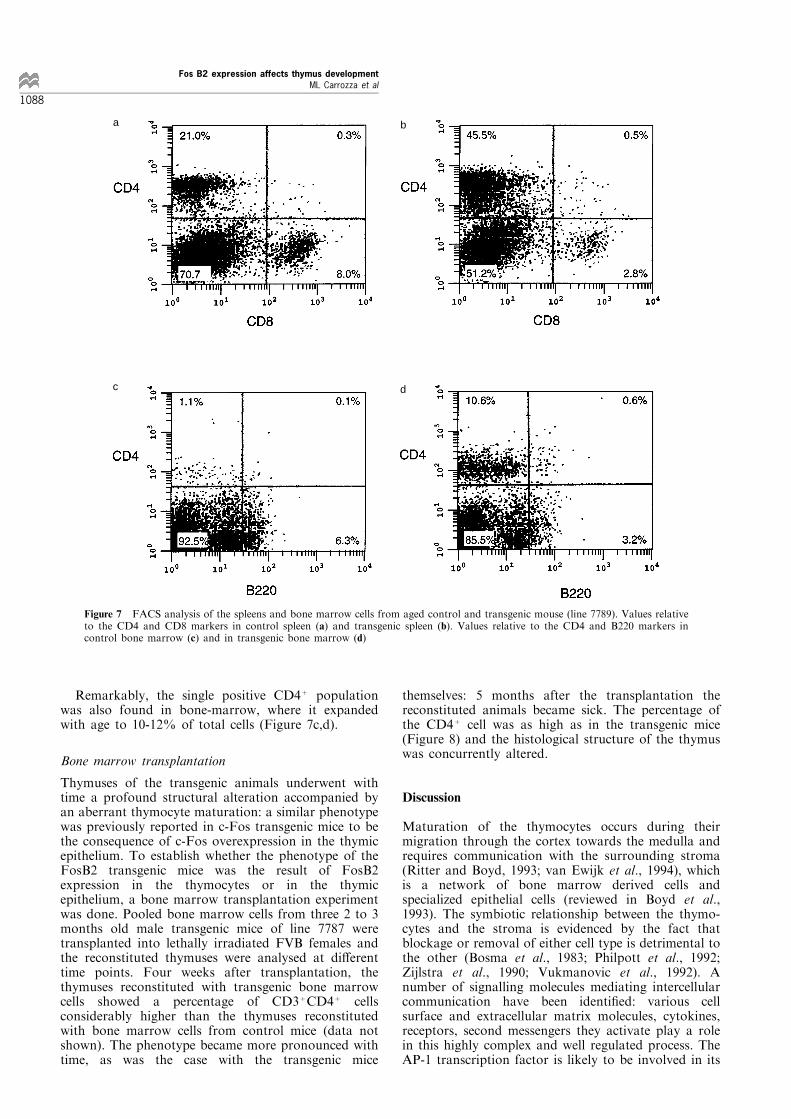
Remarkably, the single positive CD4+ populationwas also found in bone-marrow, where it expandedwith age to 10-12% of total cells (Figure 7c,d).
Bone marrow transplantation
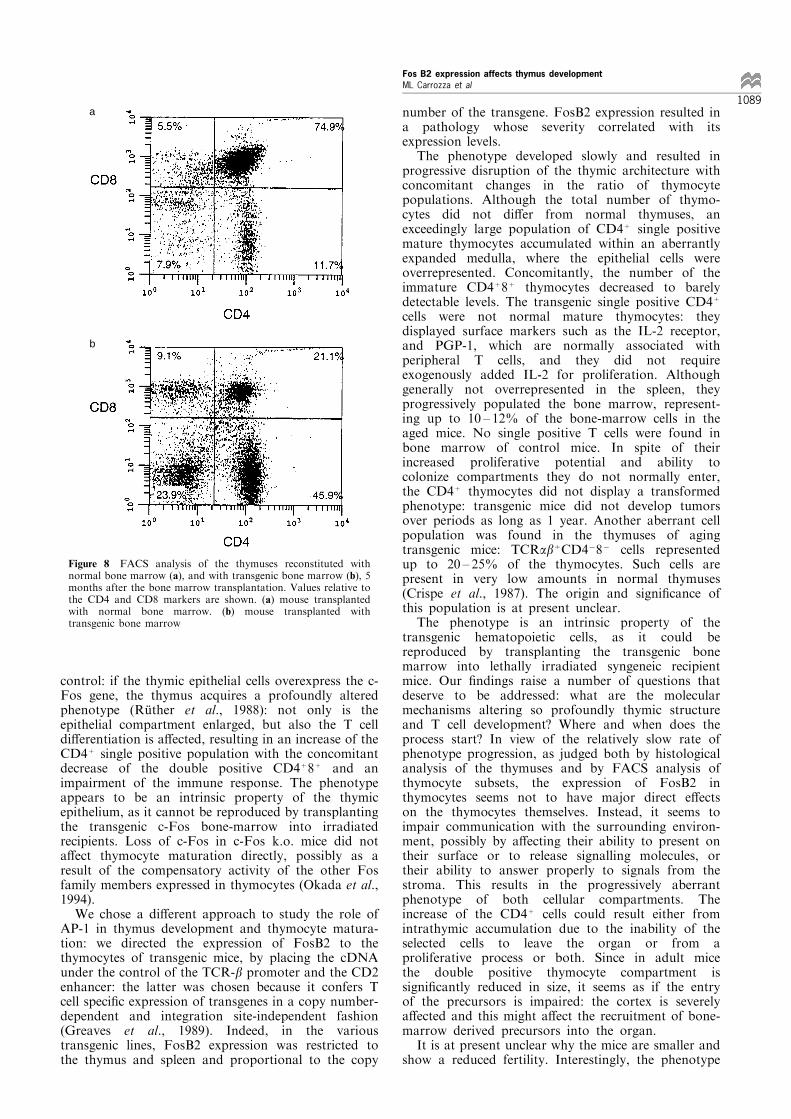
Thymuses of the transgenic animals underwent withtime a profound structural alteration accompanied byan aberrant thymocyte maturation: a similar phenotypewas previously reported in c-Fos transgenic mice to bethe consequence of c-Fos overexpression in the thymicepithelium. To establish whether the phenotype of theFosB2 transgenic mice was the result of FosB2expression in the thymocytes or in the thymicepithelium, a bone marrow transplantation experimentwas done. Pooled bone marrow cells from three 2 to 3months old male transgenic mice of line 7787 weretransplanted into lethally irradiated FVB females andthe reconstituted thymuses were analysed at di�erenttime points. Four weeks after transplantation, thethymuses reconstituted with transgenic bone marrowcells showed a percentage of CD3+CD4+ cellsconsiderably higher than the thymuses reconstitutedwith bone marrow cells from control mice (data notshown). The phenotype became more pronounced withtime, as was the case with the transgenic mice
themselves: 5 months after the transplantation thereconstituted animals became sick. The percentage ofthe CD4+ cell was as high as in the transgenic mice(Figure 8) and the histological structure of the thymuswas concurrently altered.
Discussion
Maturation of the thymocytes occurs during theirmigration through the cortex towards the medulla andrequires communication with the surrounding stroma(Ritter and Boyd, 1993; van Ewijk et al., 1994), whichis a network of bone marrow derived cells andspecialized epithelial cells (reviewed in Boyd et al.,1993). The symbiotic relationship between the thymo-cytes and the stroma is evidenced by the fact thatblockage or removal of either cell type is detrimental tothe other (Bosma et al., 1983; Philpott et al., 1992;Zijlstra et al., 1990; Vukmanovic et al., 1992). Anumber of signalling molecules mediating intercellularcommunication have been identi®ed: various cellsurface and extracellular matrix molecules, cytokines,receptors, second messengers they activate play a rolein this highly complex and well regulated process. TheAP-1 transcription factor is likely to be involved in its
a b
c d
Figure 7 FACS analysis of the spleens and bone marrow cells from aged control and transgenic mouse (line 7789). Values relativeto the CD4 and CD8 markers in control spleen (a) and transgenic spleen (b). Values relative to the CD4 and B220 markers incontrol bone marrow (c) and in transgenic bone marrow (d)
Fos B2 expression affects thymus development
ML Carrozza et al
1088
control: if the thymic epithelial cells overexpress the c-Fos gene, the thymus acquires a profoundly alteredphenotype (RuÈ ther et al., 1988): not only is theepithelial compartment enlarged, but also the T celldi�erentiation is a�ected, resulting in an increase of theCD4+ single positive population with the concomitantdecrease of the double positive CD4+8+ and animpairment of the immune response. The phenotypeappears to be an intrinsic property of the thymicepithelium, as it cannot be reproduced by transplantingthe transgenic c-Fos bone-marrow into irradiatedrecipients. Loss of c-Fos in c-Fos k.o. mice did nota�ect thymocyte maturation directly, possibly as aresult of the compensatory activity of the other Fosfamily members expressed in thymocytes (Okada et al.,1994).We chose a di�erent approach to study the role of
AP-1 in thymus development and thymocyte matura-tion: we directed the expression of FosB2 to thethymocytes of transgenic mice, by placing the cDNAunder the control of the TCR-b promoter and the CD2enhancer: the latter was chosen because it confers Tcell speci®c expression of transgenes in a copy number-dependent and integration site-independent fashion(Greaves et al., 1989). Indeed, in the varioustransgenic lines, FosB2 expression was restricted tothe thymus and spleen and proportional to the copy
number of the transgene. FosB2 expression resulted ina pathology whose severity correlated with itsexpression levels.The phenotype developed slowly and resulted in
progressive disruption of the thymic architecture withconcomitant changes in the ratio of thymocytepopulations. Although the total number of thymo-cytes did not di�er from normal thymuses, anexceedingly large population of CD4+ single positivemature thymocytes accumulated within an aberrantlyexpanded medulla, where the epithelial cells wereoverrepresented. Concomitantly, the number of theimmature CD4+8+ thymocytes decreased to barelydetectable levels. The transgenic single positive CD4+
cells were not normal mature thymocytes: theydisplayed surface markers such as the IL-2 receptor,and PGP-1, which are normally associated withperipheral T cells, and they did not requireexogenously added IL-2 for proliferation. Althoughgenerally not overrepresented in the spleen, theyprogressively populated the bone marrow, represent-ing up to 10 ± 12% of the bone-marrow cells in theaged mice. No single positive T cells were found inbone marrow of control mice. In spite of theirincreased proliferative potential and ability tocolonize compartments they do not normally enter,the CD4+ thymocytes did not display a transformedphenotype: transgenic mice did not develop tumorsover periods as long as 1 year. Another aberrant cellpopulation was found in the thymuses of agingtransgenic mice: TCRab+CD4787 cells representedup to 20 ± 25% of the thymocytes. Such cells arepresent in very low amounts in normal thymuses(Crispe et al., 1987). The origin and signi®cance ofthis population is at present unclear.The phenotype is an intrinsic property of the
transgenic hematopoietic cells, as it could bereproduced by transplanting the transgenic bonemarrow into lethally irradiated syngeneic recipientmice. Our ®ndings raise a number of questions thatdeserve to be addressed: what are the molecularmechanisms altering so profoundly thymic structureand T cell development? Where and when does theprocess start? In view of the relatively slow rate ofphenotype progression, as judged both by histologicalanalysis of the thymuses and by FACS analysis ofthymocyte subsets, the expression of FosB2 inthymocytes seems not to have major direct e�ectson the thymocytes themselves. Instead, it seems toimpair communication with the surrounding environ-ment, possibly by a�ecting their ability to present ontheir surface or to release signalling molecules, ortheir ability to answer properly to signals from thestroma. This results in the progressively aberrantphenotype of both cellular compartments. Theincrease of the CD4+ cells could result either fromintrathymic accumulation due to the inability of theselected cells to leave the organ or from aproliferative process or both. Since in adult micethe double positive thymocyte compartment issigni®cantly reduced in size, it seems as if the entryof the precursors is impaired: the cortex is severelya�ected and this might a�ect the recruitment of bone-marrow derived precursors into the organ.It is at present unclear why the mice are smaller and
show a reduced fertility. Interestingly, the phenotype
a
b
Figure 8 FACS analysis of the thymuses reconstituted withnormal bone marrow (a), and with transgenic bone marrow (b), 5months after the bone marrow transplantation. Values relative tothe CD4 and CD8 markers are shown. (a) mouse transplantedwith normal bone marrow. (b) mouse transplanted withtransgenic bone marrow
Fos B2 expression affects thymus development
ML Carrozza et al
1089

we observed is very similar to the one generated intransgenic mice overexpressing the c-Fos gene in theepithelial cells of the thymus (RuÈ ther et al., 1988),although in that case it appeared to be an intrinsicproperty of the epithelial cells, since it could not bereproduced by transgenic bone marrow transplanta-tion.These ®ndings deserve some discussion concerning
the mechanism(s) of action of the FosB2 transgene aswell as the relevance of its site of expression to thedevelopment of the phenotype.As to the ®rst issue, the Jun : FosB2 dimers
expressed in the transgenic thymocytes might eitherbe transcriptionally inactive, thereby depleting thethymocytes of the AP-1 function, or might weaklyactivate the transcription of di�erent sets of genes thanthe normal endogenous AP-1. Both mechanisms couldresult in unbalanced production of some relevantsignalling molecule(s).As to the second point, we observe that both the
overexpression of c-Fos in thymic epithelial cells and ofFosB2 in thymocytes alter the molecular communica-tion between stromal epithelium and developingthymocytes. It is likely that the action of bothtransgenes a�ects ultimately common targets, prob-ably the production of some factors involved in thetwo-way cellular communication.Independent from the mechanism by which FosB2
induces the observed phenotype, these mice might helpto gain access to factors that are of crucial importancefor the crosstalk between thymic epithelium anddeveloping thymocytes and thereby assist in unravel-ing the role of this interaction in selective outgrowth ordepletion of subsets of thymocytes.
Materials and methods
Construction of pTCDfosB2
A 1.5 kb Sal I-Hind III fragment corresponding to theTCRb gene promoter (Krimpenfort et al., 1988) wassubcloned into pACYC184: the resulting plasmid waslinearized with Hind III, the ends ®lled with Klenowpolymerase and Not I linkers added.
Following digestion of the plasmid with Cla I and Not I,the 2.1 kb Cla I-Not I fragment, spanning the CD2 enhancersequence (Greaves et al., 1989) and some unique restrictionsites adjacent to it in the construct pBluescript-CD2, wassubcloned next to the promoter sequence, the restriction sitesbeing located between the promoter and the enhancer. Theresulting plasmid was called pTCD.
A 0.9 kb BamHI-EcoRI fragment spanning the codingsequence of the murine FosB2 cDNA was treated withKlenow polymerase, ligated to Not I linkers and subclonedinto the unique Not I site of pTCD. The SV40 polyadenyla-tion sequence was inserted in the plasmid, 3' to the FosB2cDNA, as a 0.3 kb Xba I ± Sma I fragment. We used formicroinjection a 4.8 kb Sal I ±Cla I fragment encompassingthe TCRb promoter, the FosB2 cDNA with the polyadenyla-tion sequence and the CD2 enhancer.
Generation of the transgenic mice
The restriction fragment spanning the transgene and the T-cell speci®c control elements was microinjected intofertilized eggs of FVB mice. Two-cell stage embryos wereimplanted into pseudopregnant B6 fosters.
DNA and RNA analysis
Transgenic mice were identi®ed by Southern hybridizationon Pst I digested tail DNA. As a probe we used the 32P-labeled 0.9 kb BamH I ± EcoR I fragment corresponding tothe FosB2 cDNA, which detected two transgene speci®cfragments of 2.1 kb and 0.7 kb. Total RNA fromthymuses, spleens and other organs from transgenic andcontrol mice was separated on a 1.2% agarose gelcontaining 1.1% formaldehyde, transferred to nitrocellu-lose ®lters and hybridized under high stringency conditionsto the FosB2 speci®c probe. Expression of the endogenousFosB gene was not detected in adult mouse organs, asexpected (Nakabeppu and Nathans, 1991). The FosB probedoes also not detect c-Fos or Fra speci®c transcripts, dueto their limited similarity in DNA sequence (Zerial et al.,1989). Expression of the transgene resulted in a mRNA ofabout 1.4 kb.
Histological analysis
Tissues were isolated from mice and directly ®xed in EAF(4% v/v formol, 40% v/v/ ethanol, 0.43% wt/v NaCl and5% v/v acetic acid) for 24 h. Fixed and dehydrated tissueswere embedded in Histowax and cut at 5 mm sections andstained with hematoxylin and eosin stain.
Flow cytometry
Cells (36105) were incubated in 96-well plates for 30 ±45 min at 48C in 20 ml of PBS2+ (phosphate bu�ered salinewith 1% BSA and 0.1% sodium azide) and saturatingamounts of monoclonal antibody. Cell were washed twicewith PBS2= and incubated with streptavidin-phycoerythinfor biotinylated antibodies or mouse-anti-k-¯uorescein forunconjugated antibodies. The mAbs directed against CD3(145-2C11), CD4 (clone RM-4-5), CD8 (clone 53-6.7),TCRab (H57-597), Pgp-1 (CD44) (clone IM7), CD25(clone 7D4) and mouse-anti-rat k light chain (MRK-1)were purchased from Pharmingen (San Diego, CA). Flowcytometry was performed on a FACScan (Becton Dick-inson, Mountain View, CA), using Lysys II software.
In vitro proliferation assay
Thymocytes and splenocytes were isolated from 8, 12 and16 weeks old transgenic and control mice. Serially dilutedcell suspensions (56105 to 36103 cells) were incubated for3 days in microwells precoated with anti-CD3 antibody(145.2C11, 5 mg/ml in PBS, 50 ml/well); tritiated thymidinewas added (0.5 mCi in 25 ml Iscoves/well) and 5 h later thecells were harvested and processed to determine thethymidine incorporation.
Bone marrow transplantation
Two groups of ten 2 to 3 months old FVB female micewere lethally irradiated and reconstituted respectively with56104 and 56105 pooled bone marrow cells from three 2months old donor male mice from transgenic line 7787.Two parallel control groups were reconstituted with bonemarrow cells from normal FVB donor mice.
At regular intervals from 4 to 16 weeks after the transplanta-tion, the recipient mice were sacri®ced and the reconstitutedthymuses underwent histological and FACS analysis.
Acknowledgements
We would like to thank our colleagues from the animalfacility for their help in maintaining the mice, and M vander Valk for performing the pathology and evaluating thehistological samples. This work was in part supported bythe Dutch Cancer Society (DA).
Fos B2 expression affects thymus development
ML Carrozza et al
1090

References
Bosma GC, Custer RP and Bosma MJ. (1983). Nature, 301,527 ± 530.
Boyd RL and Hugo P. (1991). Immunol. Today, 12, 71 ± 78.Boyd RL, Tucek CL, Godfrey DI, Izon D, Wilson TJ,Davidson NJ, Bean AGD, Ladyman HM, Ritter AM andHugo P. (1993). Immunol. Today, 14, 445 ± 459.
Couture C, Amarante-Mendes B and Potworowski EF.(1992). Eur. J. Immunol., 22, 2579 ± 2585.
Crispe IN, Moore MW, Husmann LA, Smith L, Bevan MJand Shimonkevitz RP. (1987). Nature, 329, 336 ± 339.
Curran T and Franza Jr BR. (1988). Cell, 55, 395 ± 397.Dobrzansky P, Noguchi T, Kovary K, Rizzo CA, Lazo PSand Bravo R. (1991). Mol. Cell. Biol., 11, 5470 ± 5478.
Fowlkes BJ and Pardoll DM. (1989). Adv. in Immunol.,44,207 ± 263.
Galy AJ and Spits H. (1992). J. Immunology, 149, 775 ± 782.Greaves DR, Wilson FD, Lang G and Kioussis D. (1989).
Cell, 56, 979 ± 986.Greenberg ME and Zi� EB. (1984). Nature, 311, 433 ± 437.Hirai SI, Ryseck RP, Mechta F, Bravo R and Yaniv M.(1989). The EMBO J., 8, 1433 ± 1443.
Jain J, McCa�rey PG, Miner Z, Kerppola TK, Lambert JN,Verdine GL, Curran T and Rao A. (1993). Nature, 365,352 ± 355.
Krimpenfort P, De Jong R, Uematsu Y, Dembic Z, Ryser S,Von Boehmer H, Steinmetz M. and Berns A. (1988).EMBO J., 7, 745 ± 750.
Lee W, Mitchell P and Tjian R. (1987). Cell, 49, 741 ± 752.Metz R, Kouzarides T and Bravo R. (1994). The EMBO J.,
13, 3832 ± 3842.Morgan JI and Curran T. (1991). A. Rev. Neurosci., 14,421 ± 451.
MuÈ ller R and Wagner EF. (1984). Nature, 311, 438 ± 442.MuÈ ller R. (1986). Biochim, Biophys. Acta, 823, 207 ± 225.Mumberg D, Lucibello FC, Schuermann M and MuÈ ller R.(1991). Genes & Dev., 5, 1212 ± 1223.
Nakabeppu Y and Nathans D. (1991). Cell, 64, 751 ± 759.
Okada S, Wang ZQ, Grigoriadis AE, Wagner EF and vonRuÈ den T. (1994). Mol. Cell. Biol. 14, 382 ± 390.
Philpott KL, Viney JL, Kay G, Rastan S, Gardiner EM,Chae S, Hayday AC and Owen MJ. (1992). Science, 256,1448 ± 1452.
Ramarli D, Fox DA and Rheinherz EL. (1987). Proc. Natl.Acad. Sci., 84, 8598 ± 8602.
Ransone LJ and Verma IM. (1990). Ann. Rev. Cell. Biol., 6,539 ± 557.
Ritter MA and Boyd RL. (1993). Immunol. Today, 14, 462 ±469.
RuÈ ther U, MuÈ ller W, Sumida T, Tokuhisa T, Rajewsky Kand Wagner EF. (1988). Cell, 53, 847 ± 856.
SchuÈ ttle J, Viallet J, Nau M, Segal S, Fedorko J and MinnaJ. (1989). Cell, 59, 987 ± 997.
Shores EW, van Ewijk W and Singer A. (1991). Eur. J.Immunol., 21, 1657 ± 1661.
Smeyne RJ, Vendrell M, Hayward M, Baker SJ, Mia GG,Schilling K, Robertson LM, Curran T and Morgan JI.(1993). Nature, 363, 166 ± 169.
Surh CD, Ernst B and Sprent J. (1992). J. Exp. Med., 176,495 ± 505.
van Ewijk W, Shores EW and Singer A. (1994). Immunol.Today, 15, 214 ± 217.
Vukmanovic S, Grandea AG, Faas SJ, Knowles BB andBevan MJ. (1992). Nature, 359, 729 ± 732.
Wisdom R, Yen J, Rashid D and Verma IM. (1992). Genes &Dev., 6, 667 ± 675.
Wolf SS and Cohen A. (1993). Immunology, 77, 362 ± 368.Yen J, Wisdom RM, Tratner I and Verma I. (1991). Proc.
Natl. Acad. Sci., 88, 5077 ± 5081.Zerial M, Toschi L, Ryseck RP, Schuermann M, MuÈ ller andBravo R. (1989). EMBO J., 8, 805 ± 813.
Zijlstra M, Bix M, Simister ME, Loving JM, Raylet DH andJaenisch R. (1990). Nature, 344, 742 ± 746.
Zugic JN. (1991). Immunol. Today, 12, 65 ± 70.
Fos B2 expression affects thymus development
ML Carrozza et al
1091
Related Documents











