REVIEW ARTICLE Overexpression of C 4 -cycle enzymes in transgenic C 3 plants: a biotechnological approach to improve C 3 -photosynthesis Rainer E. Ha ¨ usler 1,3 , Heinz-Josef Hirsch 2 , Fritz Kreuzaler 2 and Christoph Peterha ¨ nsel 2 1 Botanik II, Botanisches Institut der Universita ¨ t zu Ko ¨ ln, Gyrhofstrasse 15, D-50931 Cologne, Germany 2 Institut fu ¨ r Biologie I, RWTH, Worringer Weg 1, D-52074 Aachen, Germany Received 8 November 2001; Accepted 18 December 2001 Abstract The process of photorespiration diminishes the efficiency of CO 2 assimilation and yield of C 3 -crops such as wheat, rice, soybean or potato, which are important for feeding the growing world population. Photorespiration starts with the competitive inhibi- tion of CO 2 fixation by O 2 at the active site of ribulose- 1,5-bisphosphate carboxylase/oxygenase (Rubisco) and can result in a loss of up to 50% of the CO 2 fixed in ambient air. By contrast, C 4 plants, such as maize, sugar cane and Sorghum, possess a CO 2 concentrat- ing mechanism, by which atmospheric CO 2 is bound to C 4 -carbon compounds and shuttled from the mesophyll cells where the prefixation of bicarbonate occurs via phosphoenolpyruvate carboxylase (PEPC) into the gas-tight bundle-sheath cells, where the bound carbon is released again as CO 2 and enters the Calvin cycle. However, the anatomical division into mesophyll and bundle-sheaths cells (‘Kranz’- anatomy) appears not to be a prerequisite for the operation of a CO 2 concentrating mechanism. Sub- merged aquatic macrophytes, for instance, can induce aC 4 -like CO 2 concentrating mechanism in only one cell type when CO 2 becomes limiting. A single cell C 4 -mechanism has also been reported recently for a terrestrial chenopod. For over 10 years researchers in laboratories around the world have attempted to improve photosynthesis and crop yield by introducing a single cell C 4 -cycle in C 3 plants by a transgenic approach. In the meantime, there has been sub- stantial progress in overexpressing the key enzymes of the C 4 cycle in rice, potato, and tobacco. In this review there will be a focus on biochemical and physiological consequences of the overexpression of C 4 -cycle genes in C 3 plants. Bearing in mind that C 4 -cycle enzymes are also present in C 3 plants, the pitfalls encountered when C 3 metabolism is per- turbed by the overexpression of individual C 4 genes will also be discussed. Key words: Biotechnology, crop yield, transgenic C 3 plants. Introduction Photorespiration decreases the efficiency of CO 2 assimilation in C 3 plants Most of our crops, such as wheat, rice, soybean or potato are classified as C 3 plants as the first product of atmospheric CO 2 fixation is the 3-carbon compound 3-phosphoglycerate (3-PGA), which is produced in the Calvin cycle by Rubisco (the only enzyme capable of net carbon assimilation) in the chloroplast stroma. However, competition of O 2 with CO 2 at the active site of Rubisco (Chen and Spreitzer, 1992; Jordan and Ogren, 1984) results in a loss of up to 50% of the carbon fixed in a process known as photorespiration (Ogren, 1984). 3 To whom correspondence should be addressed. Fax: q49 221 470 5039. E-mail: [email protected] Abbreviations: C*, CO 2 compensation point in the absence of dark respiration in the light; CA, carbonic anhydrase; NAD(P)-MDH, NAD(P)-dependent malate dehydrogenase; NAD(P)-ME, NAD(P)-dependent malic enzyme; OAA, oxaloacetate; PEP, phosphoenolpyruvate; PEPC, phosphoenolpyruvate carboxylase; PEPCK, phosphoenolpyruvate carboxykinase; PEPS, phosphoenolpyruvate synthetase; 3-PGA, 3-phosphoglycerate; 2-PG, 2-phosphoglycollate; PFD, photon flux density; PPDK, pyruvate, orthophosphate dikinase; PPT, phosphoenolpyruvate/phosphate translocator; R d , dark respiration in the light; Rubisco, ribulose 1,5-bisphosphate carboxylase/oxygenase; TPT, triose phosphate/phosphate translocator. Journal of Experimental Botany, Vol. 53, No. 369, pp. 591–607, April 2002 ß Society for Experimental Biology 2002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REVIEW ARTICLE
Overexpression of C4-cycle enzymes in transgenic
C3 plants: a biotechnological approach to improve
C3-photosynthesis
Rainer E. Hausler1,3, Heinz-Josef Hirsch2, Fritz Kreuzaler2 and Christoph Peterhansel2
1 Botanik II, Botanisches Institut der Universitat zu Koln, Gyrhofstrasse 15, D-50931 Cologne, Germany2 Institut fur Biologie I, RWTH, Worringer Weg 1, D-52074 Aachen, Germany
Received 8 November 2001; Accepted 18 December 2001
Abstract
The process of photorespiration diminishes theefficiency of CO2 assimilation and yield of C3-cropssuch as wheat, rice, soybean or potato, which areimportant for feeding the growing world population.Photorespiration starts with the competitive inhibi-tion of CO2 fixation by O2 at the active site of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco)and can result in a loss of up to 50% of the CO2 fixedin ambient air. By contrast, C4 plants, such as maize,sugar cane and Sorghum, possess a CO2 concentrat-ing mechanism, by which atmospheric CO2 is boundto C4-carbon compounds and shuttled from themesophyll cells where the prefixation of bicarbonateoccurs via phosphoenolpyruvate carboxylase (PEPC)into the gas-tight bundle-sheath cells, where thebound carbon is released again as CO2 and entersthe Calvin cycle. However, the anatomical divisioninto mesophyll and bundle-sheaths cells (‘Kranz’-anatomy) appears not to be a prerequisite for theoperation of a CO2 concentrating mechanism. Sub-merged aquaticmacrophytes, for instance, can inducea C4-like CO2 concentrating mechanism in only onecell type when CO2 becomes limiting. A single cellC4-mechanism has also been reported recently for aterrestrial chenopod. For over 10 years researchersin laboratories around the world have attempted toimprove photosynthesis and crop yield by introducinga single cell C4-cycle in C3 plants by a transgenic
approach. In the meantime, there has been sub-stantial progress in overexpressing the key enzymesof the C4 cycle in rice, potato, and tobacco. In thisreview there will be a focus on biochemical andphysiological consequences of the overexpressionof C4-cycle genes in C3 plants. Bearing in mind thatC4-cycle enzymes are also present in C3 plants, thepitfalls encountered when C3 metabolism is per-turbed by the overexpression of individual C4 geneswill also be discussed.
Key words: Biotechnology, crop yield, transgenic C3 plants.
Introduction
Photorespiration decreases the efficiency ofCO2 assimilation in C3 plants
Most of our crops, such as wheat, rice, soybean orpotato are classified as C3 plants as the first product ofatmospheric CO2 fixation is the 3-carbon compound3-phosphoglycerate (3-PGA), which is produced in theCalvin cycle by Rubisco (the only enzyme capable of netcarbon assimilation) in the chloroplast stroma. However,competition of O2 with CO2 at the active site of Rubisco(Chen and Spreitzer, 1992; Jordan and Ogren, 1984)results in a loss of up to 50% of the carbon fixed ina process known as photorespiration (Ogren, 1984).
3 To whom correspondence should be addressed. Fax: q49221 470 5039. E-mail: [email protected]
Abbreviations: C*, CO2 compensation point in the absence of dark respiration in the light; CA, carbonic anhydrase; NAD(P)-MDH, NAD(P)-dependent
malate dehydrogenase; NAD(P)-ME, NAD(P)-dependent malic enzyme; OAA, oxaloacetate; PEP, phosphoenolpyruvate; PEPC, phosphoenolpyruvate
carboxylase; PEPCK, phosphoenolpyruvate carboxykinase; PEPS, phosphoenolpyruvate synthetase; 3-PGA, 3-phosphoglycerate; 2-PG,
2-phosphoglycollate; PFD, photon flux density; PPDK, pyruvate, orthophosphate dikinase; PPT, phosphoenolpyruvate/phosphate translocator; Rd,
dark respiration in the light; Rubisco, ribulose 1,5-bisphosphate carboxylase/oxygenase; TPT, triose phosphate/phosphate translocator.
Journal of Experimental Botany, Vol. 53, No. 369, pp. 591–607, April 2002
� Society for Experimental Biology 2002

Oxygenation of ribulose-1,5-bisphosphate (RubP) severelydiminishes the efficiency of CO2 assimilation in C3 plantsin ambient air and results in the formation of 3-PGA aswell as 2-phosphoglycollate (2-PG). The latter is meta-bolized in three compartments of the leaf cell, the chloro-plast, the peroxysomes and the mitochondria, involvingnumerous enzymatic reactions and transport processes(Fig. 1). The overall photorespiratory cycle is also linkedto amino acid metabolism in that glycine, serine, gluta-mate, and glutamine are metabolized at high rates (Keyset al., 1978). Both CO2 and ammonia are released at equalrates in the reaction catalysed by the mitochondrialglycine decarboxylase complex (Oliver, 1994). The lossof CO2 during photorespiration is reflected in a CO2
compensation point (C) of CO2 assimilation of between40–60 ml l�1 CO2 in the intercellular air space. At theCO2 compensation point, net CO2 assimilation equalsCO2 release through photorespiration and mitochondrialrespiration in the light. In high CO2 anduor low O2 theoxygenase activity of Rubisco is virtually absent, the fluxthrough the photorespiratory carbon cycle negligible andthe CO2 compensation point close to zero.
There are numerous reports on the improvement ofgrowth and crop yield of C3 plants in an atmosphere
containing elevated CO2 (Arp et al., 1998; Besford, 1990;Chen et al., 1997; Teramura et al., 1990). This ismainly based on a faster biomass production due to anincrease in CO2 assimilation rates and a suppression ofphotorespiration.
C4 plants have developed strategies to concentrateCO2 in the vicinity of Rubisco
During the evolution of higher plants, adaptationsto low water supply anduor hot environments havedeveloped independently several times (Edwards et al.,2001; Kellogg, 1999; Ku et al., 1996). Plants, which displayC4-metabolism, release CO2 at high rates in the vicinityof Rubisco and thereby increase the ratio of RubPcarboxylationuoxygenation substantially (Leegood, 1997,2002). This strategy prevents major losses of CO2 byphotorespiration and is accompanied by an increase inthe water and nitrogen use efficiency compared to C3
plants (Sage and Pearcy, 1987). Common C4 plants arecharacterized by the so-called ‘Kranz’-anatomy withmesophyll cells surrounded by relatively thin cell wallsand bundle-sheath cells surrounded by thick cell walls.This anatomical separation into different cell types is
Fig. 1. The photorespiratory carbon and nitrogen cycle typical for C3 plants in ambient air. Photorespiration starts with the oxygenase reaction ofRubisco (1). (2) Phosphoglycollate phosphatase, (3) glycollate oxidase, (4) glutamate : glyoxylate aminotransferase, (5) serine : glyoxylateaminotransferase, (6) glycine decarboxylase, (7a, b) NAD malate dehydrogenase, (8) hydroxypyruvate reductase, (9) glycerate kinase, (10) glutaminesynthetase, (11) glutamate synthase.
592 Hausler et al.

accompanied by a spatial separation of the prefixationof atmospheric CO2 in the mesophyll cells followed by therelease of CO2 and its refixation via the C3 (Calvin) cyclein the bundle-sheath cells. Moreover, in order to keepintercellular diffusion ways short, mesophyll cells are inclose proximity to the bundle-sheath (Dengler andNelson, 1999). The first step in the C4-cycle is thecarboxylation of PEP by phosphoenolpyruvate carb-oxylase (PEPC) in the cytosol of the mesophyll cells usingHCO�
3 as the inorganic carbon substrate (Cooper andWood, 1971; O’Leary, 1982). This yields the C4 dicar-boxylic acid oxaloacetate (OAA), which is either reducedto malate or transaminated to aspartate and is thentransported into the bundle-sheath cells, where CO2 isreleased at high rates by decarboxylating enzymes.Depending on the type of C4 plants, CO2 is releasedeither by chloroplastic NADP malic enzyme (ME),
mitochondrial NAD-ME, or cytosolic phosphoenol-pyruvate carboxykinase (PEPCK) (Hatch and Osmond,1976; Leegood, 2002) (Fig. 2). Since bundle-sheath cellshave low gas permeability, the CO2 concentration insolution is drastically increased, which causes a suppres-sion of the Rubisco oxygenase activity and, consequently,photorespiration (Leegood, 1997). Pyruvate released byNADP-ME or NAD-ME is transferred back into themesophyll chloroplasts where the inorganic carbonacceptor PEP is regenerated by pyruvate, Pi dikinase(PPDK). The reaction, catalysed by PPDK, consumes inaddition two ATP per CO2 assimilated. One ATP isdirectly required by PPDK, the second ATP is neededfor the conversion of the reaction product AMP intoADP catalysed by adenylate kinase (Fig. 2). The C4
strategy permits high rates of CO2 assimilation at arelatively small stomatal aperture and thereby increases
Fig. 2. CO2 concentrating mechanism in a NADP malic enzyme C4 plant, such as maize, sugar cane or Sorghum. CO2 is converted to HCO3�
by carbonic anhydrase (1) in the cytosol of the mesophyll cells and fixed by oxygen-insensitive PEPC (2). The oxaloacetate formed is imported intothe stroma of the mesophyll chloroplasts and reduced by NADP-MDH (3) using redox equivalents from non-cyclic electron transport. Malate isexported from the stroma in counter exchange with OAA catalysed by a malateuOAA transporter (4). In the mesophyll cell the concentration ofmalate is high, which allows diffusion along a concentration gradient to the bundle-sheath cells. Malate enters the bundle-sheath chloroplasts andis subjected to oxidative decarboxylation by NADP-ME (5). As bundle-sheath cells are gas tight, the high rate of oxidative malate decarboxylationresults in a steep increase in the CO2 concentration in the vicinity of Rubisco (6) and hence a suppression of the oxygenase activity andphotorespiration. As bundle-sheath chloroplasts of NADP-ME C4 plants lack photosystem II and hence the capacity for non-cyclic electron transport,NADPH formed by NADP-ME is utilized for the reduction of 50% of 3-PGA formed by Rubisco. The residual 3-PGA is exported by a C4-typeTPT (7a) and is transferred to the mesophyll chloroplasts, imported into the stroma via the TPT (7b) and reduced to triose phosphates. Pyruvate, theproduct of malate decarboxylation also diffuses into mesophyll cells, enters the chloroplasts via a pyruvate transporter (8) and the primary inorganiccarbon acceptor PEP is regenerated by PPDK (9). The PPi released by this reaction is cleaved by pyrophosphatase (10) and the AMP converted toADP by adenylate kinase (11). Hence, for the regeneration of PEP an additional two ATP are required. PEP is exported from the chloroplast via thePPT (12). For the sake of clarity not all cofactors are shown.
Biotechnology and C3 photosynthesis 593

the water use efficiency. Moreover, due to an almostcomplete suppression of RubP oxygenation the CO2
compensation point is close to zero or very low ascompared to C3 plants.
C4 plants lacking ‘Kranz’ anatomy
Since the discovery of the C4 cycle some 36 years ago thespatial separation into mesophyll cells and bundle sheathcells was thought to be a prerequisite for an efficient CO2-concentrating mechanism. It is therefore quite amazingthat the submerged aquatic plant Hydrilla verticillata wasidentified as being capable of inducing a C4-like meta-bolism (Fig. 3A), but lacking the ‘Kranz’-anatomy typicalfor C4 plants (Holaday and Bowes, 1980; Magnin et al.,1997; Salvucci and Bowes, 1981, 1983; Spencer et al.,1996). The switch from C3- to C4-like metabolism inH. verticillata is triggered by low CO2 concentrations(i.e. at high water temperatures) resulting in an increase
in PEPC, ME and PPDK activities, which causes a sub-stantial drop in the CO2 compensation point from around40 ml l�1 in the C3-state down to below 10 ml l�1 in theC4-induced state (Spencer et al., 1996; Reiskind et al.,1997) (Fig. 3A). Further evidence for a C4-like metab-olism without ‘Kranz’ anatomy emerged for otheraquatic species such as Egeria densa (Casati et al., 2000)and (probably) Elodea canadensis (de Groote andKennedy, 1977). Moreover, apart from the inducibleC4-photosynthesis these aquatic species exhibit apH-polarity of their leaf surfaces. The steady-state pHof the adaxial surface is acidic (pH 4.0), whereas theabaxial side of the leaf is alkaline (pH 10.0) (van Ginkelet al., 2001). An acidic environment allows higher CO2
concentrations at equilibrium despite a drop in the totalinorganic carbon in the surrounding water. This supple-mentary mechanism to increase the availability ofexternal CO2 is also apparent in the C4-induced state ofH. verticillata.
Fig. 3. (A) Induced C4 cycle in the submerged aquatic plant Hydrilla verticillata without ‘Kranz’ anatomy. (1) CA, (2) PEPC, (3) NAD-MDH,(4) NADP-MDH, (5) NADP-ME, (6) Rubisco, (7) PPDK, (8) PPT. (B) Genetically engineered single cell CO2 concentrating mechanism in trans-genic tobacco plants. The transformants contain all combinations of C4-cycle enzymes (PEPC, PEPCK, NADP-ME, PPDK or PEPS as well as thePPT). For the sake of clarity not all cofactors are shown.
594 Hausler et al.

Interestingly, single cell C4 photosynthesis appears notto be restricted to aquatic macrophytes. Recent determi-nations of d13C values (a measure for the activity ofPEPC compared to Rubisco, which discriminates 13CO2
in favour of 12CO2) combined with anatomical studiesrevealed that the terrestrial chenopod Borszczowia aralo-caspica (a halophyte with succulent leaves adapted to asemi-dry environment) is likely to carry out C4 meta-bolism in only one cell type (Freitag and Stichler, 2000).Moreover, the individual chlorenchyma cells of B. aralo-caspica carry two types of chloroplasts with a distinctenzyme equipment and capability to produce starch.Immunolocalization of enzyme proteins revealed thatRubisco (as well as starch) is localized in chloroplasts ofthe basal part of the chlorenchyma cells (i.e. closer tothe vascular bundles), whereas PPDK is localized in thechloroplasts of the distal parts of the cells (i.e. closer tothe intercellular air space). Like Rubisco, NAD malicenzyme, which appears to act as the decarboxylatingenzyme is also more abundant in the mitochondria ofthe basal parts of the chlorenchyma cells. PEPC is equallydistributed throughout the cytosol (Vozesenskaya et al.,2001).
Crassulacean acid metabolism (CAM) plants also exhibita single cell CO2-concentrating mechanism
A different strategy of a single cell CO2-concentratingmechanism is realized in Crassulacean acid metabolism(CAM) plants, which also show an adaptation to aminimized water supply. In CAM plants both theprefixation of HCO�
3 by PEPC and the release of CO2
by decarboxylating enzymes is temporally separated(Osmond, 1978; Cockburn et al., 1979; Spalding et al.,1979). HCO�
3 is prefixed in the dark by PEPC at lowtemperatures and open stomata and malic acid, whichaccumulates in the vacuole during the night is decarboxyl-ated during the next day, when stomata are closed, eitherby cytosolic NADP-ME in concert with mitochondrialNAD-ME or by cytosolic PEPCK. This mechanismefficiently reduces the loss of water and also results in ahigh CO2 environment in the mesophyll during the dayand thereby suppresses photorespiration.
Attempts to introduce a single cell CO2-concentratingmechanism into terrestrial crops
In laboratories around the world attempts to introducesingle cell C4-like features into terrestrial C3 plants bya transgenic approach are in progress (Matsuoka et al.,2001). It is believed that the introduction of an intra-cellular CO2 pump might improve the efficiency of C3
photosynthesis by a substantial suppression of photo-respiration. There has been progress in single, double andmultiple overexpressions of C4-cycle enzymes in C3 crops.In particular, very high expression levels could be achieved
in transgenic rice (Ku et al., 1999). However, physio-logical alterations caused by this approach have beenaddressed only in a few reports. As C4-cycle enzymesare also common in C3 plants, although with muchsmaller activities, the introduction of individual C4-cyclegenes might perturb not only primary metabolism, butin certain cases also secondary metabolism (Hausleret al., 2001).
Although a single cell C4-like system appears tooperate in aquatic macrophytes and also in a terrestrialchenopod adapted to semi-dry climates, it is still an openquestion whether a biotechnologically inserted CO2
pump in terrestrial C3 crops can efficiently increase thecarboxylationuoxygenation ratio of Rubisco. It is ques-tionable whether the additional CO2 released from theproducts of PEP carboxylation can be retained within themesophyll or whether it is just lost to the atmosphere,because of high diffusion rates of CO2 in the airspace ofthe leaves (Leegood, 2002). In order to match the require-ments of the metabolic environment of individual C3 cropspecies, future research might also focus more on genetic-ally engineered C4-cycle enzymes driven by more specificpromoters. In this review there will be a focus on bio-chemical and physiological consequences of the over-expression of C4-cycle genes in the C3 plants Solanumtuberosum and Nicotiana tabacum. The progress in thegenetic manipulation of rice plants, particularly theachievement of extremely high expression levels willbe dealt with in a second review.
The role of C4-cycle enzymes in C3 plants
Exceptionally, all enzymes and metabolite transportersinvolved in the C4 pathway also occur in C3 plants,although at much lower activities and different tissuespecificities. Before the question as to the feasibility of theapproach to establish a C4-like cycle in C3 plants istackled, one has to be aware of the physiologicalfunctions of the C3 forms of the individual enzymes andtransporters summarized in Table 1.
PEPC [EC 4.1.1.31]
As in C4 plants, PEPC in C3 plants is subject to complexregulation by metabolites and covalent modification byreversible phosphorylation (Andreo et al., 1987; Van Quyet al., 1991; Duff and Chollet, 1995; Zhang et al., 1995;Chollet et al., 1996). In certain types of heterotrophictissues from developing fruits and seeds of C3 plants, theCO2 respired is recaptured by PEPC. These tissues con-tain relatively high PEPC activities (reviewed by Latzkoand Kelly, 1983). In leaves of C3 plants one role ofPEPC is the supply of carbon skeletons for amino acidbiosynthesis following nitrate assimilation (Andrews,1986; Melzer and O’Leary, 1987) (Fig. 4). During nitrate
Biotechnology and C3 photosynthesis 595
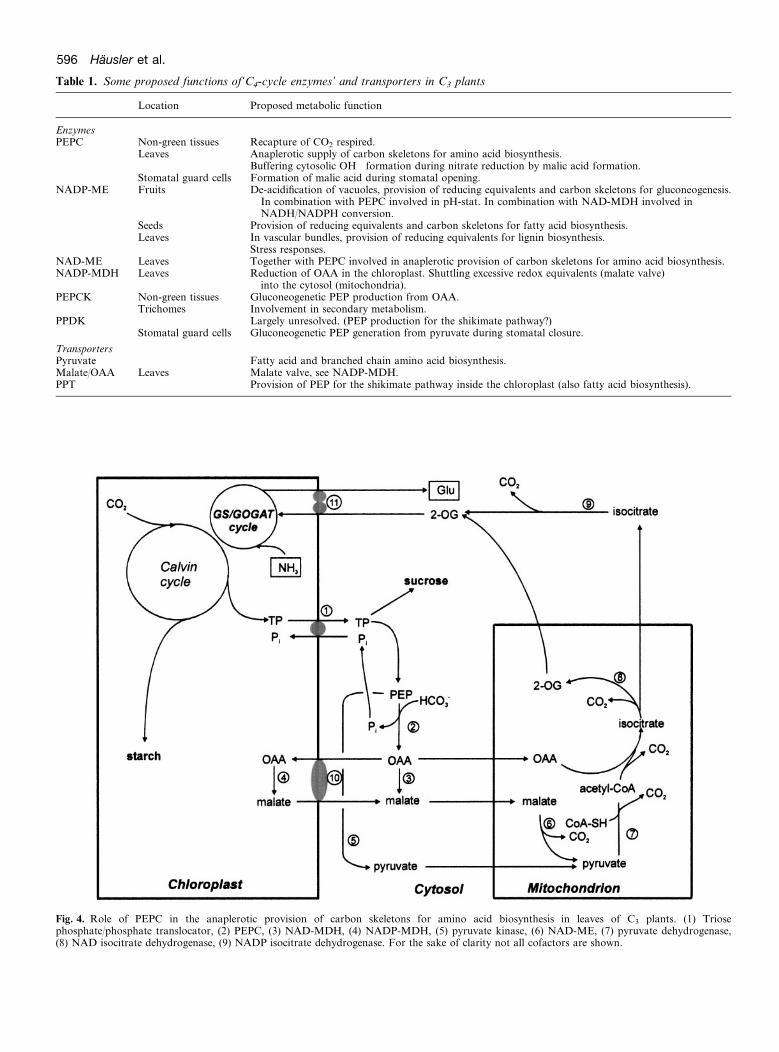
Table 1. Some proposed functions of‘C4-cycle enzymes’ and transporters in C3 plants
Location Proposed metabolic function
EnzymesPEPC Non-green tissues Recapture of CO2 respired.
Leaves Anaplerotic supply of carbon skeletons for amino acid biosynthesis.Buffering cytosolic OH� formation during nitrate reduction by malic acid formation.
Stomatal guard cells Formation of malic acid during stomatal opening.NADP-ME Fruits De-acidification of vacuoles, provision of reducing equivalents and carbon skeletons for gluconeogenesis.
In combination with PEPC involved in pH-stat. In combination with NAD-MDH involved inNADHuNADPH conversion.
Seeds Provision of reducing equivalents and carbon skeletons for fatty acid biosynthesis.Leaves In vascular bundles, provision of reducing equivalents for lignin biosynthesis.
Stress responses.NAD-ME Leaves Together with PEPC involved in anaplerotic provision of carbon skeletons for amino acid biosynthesis.NADP-MDH Leaves Reduction of OAA in the chloroplast. Shuttling excessive redox equivalents (malate valve)
into the cytosol (mitochondria).PEPCK Non-green tissues Gluconeogenetic PEP production from OAA.
Trichomes Involvement in secondary metabolism.PPDK Largely unresolved. (PEP production for the shikimate pathway?)
Stomatal guard cells Gluconeogenetic PEP generation from pyruvate during stomatal closure.
TransportersPyruvate Fatty acid and branched chain amino acid biosynthesis.MalateuOAA Leaves Malate valve, see NADP-MDH.PPT Provision of PEP for the shikimate pathway inside the chloroplast (also fatty acid biosynthesis).
Fig. 4. Role of PEPC in the anaplerotic provision of carbon skeletons for amino acid biosynthesis in leaves of C3 plants. (1) Triosephosphateuphosphate translocator, (2) PEPC, (3) NAD-MDH, (4) NADP-MDH, (5) pyruvate kinase, (6) NAD-ME, (7) pyruvate dehydrogenase,(8) NAD isocitrate dehydrogenase, (9) NADP isocitrate dehydrogenase. For the sake of clarity not all cofactors are shown.
596 Hausler et al.

reduction in the cytosol OH� ions are generated. Asboth processes, nitrate reduction and malate formation,are closely correlated the production of OAA via PEPCand the consecutive formation of malic acid may buffercytosolic pH (Martinoia and Rentsch, 1994). Moreover,PEPC also plays an important role in stomatal move-ment. An increased influx of potassium ions into theguard cells during stomatal opening and the concomitantefflux of protons via the plasma membrane-boundATP-dependent proton pump would result in a rapidalkalization of the cytosol (Raschke et al., 1988). Thisis counteracted by the synthesis of malic acid from neutralsugars or transitory starch via PEPC. For instance, asubstantial decrease in the stomatal aperture has beenreported for Vicia faba after the application of DCDP(3,3-dichloro-2-dihydroxyphosphinoyl-2-propeoate) aspecific inhibitor of PEPC (Asai et al., 2000).
NADP-ME [EC 1.1.1.40]
NADP-ME contributes to a huge variety of metabolicpathways in green and non-green tissues of C3 plants asreviewed previously (Edwards and Andreo, 1992). It isfound in leaves, etiolated tissues, seeds, roots, fruits, andtubers (potato) in the chloroplast as well as in the cytosol.In fruits, NADP-ME is involved in ripening and in thede-acidification of the vacuole as well as in the provisionof reducing equivalents and carbon skeletons for sucrosebiosynthesis via gluconeogenesis. This is comparable tothe situation in CAM plants in the light. Together withPEPC, NADP-ME might also serve as a pH-stat and incombination with NAD malate dehydrogenase (MDH)could be involved in the conversion of NADH to NADPH.In oil-storing tissues, such as seeds of rapeseed or wheatgerm, NADP-ME is engaged in the proliferation ofreducing power and carbon skeletons for fatty acidbiosynthesis.
Leaves of solanaceous species, such as potato andtobacco, contain substantial activities of cytosolicNADP-ME (Knee et al.,1996). There are indications thatthe cytosolic enzyme is associated with vascular bundles,particularly with developing xylem and internal phloem(Schaaf et al., 1995) suggesting that NADPH produced bythis reaction is required for lignin biosynthesis. Moreover,NADP-ME appears to be involved in stress responses(Casati et al., 1999). In leaf tissues, the expressionof NADP-ME is increased severely by woundingin combination with glutathionine treatment (Schaafet al., 1995).
NAD-ME [EC 1.1.1.39]
NAD-ME is localized in the mitochondria and in C3
plants is involved in the anaplerotic carbon supply foramino acid biosynthesis (Fig. 4). Malate formed by PEPCcan either be used in the citric acid cycle or it can be
decarboxylated to provide pyruvate and subsequentlyacetyl-CoA (Douce and Neuburger, 1989).
NADP-MDH [EC 1.1.1.82]
In leaves of C3 plants, NADP-MDH plays a central rolein shuttling excessive redox equivalents from the chloro-plast into the cytosol (Scheibe, 1987). This so-calledmalate valve operates at high light intensities, whenelectron transport rates and NADPH generation exceedthe demands for CO2 assimilation and thereby preventsoverreduction of the stroma anduor increased Mehler-peroxidase reaction (Polle, 1996). NADP-MDH issubjected to reversible activation by thioredoxin-m andthereby responds to the redox state of the stroma.
PEPCK [EC 4.1.1.39]
As reviewed recently (Leegood et al., 1999) plant PEPCKis a cytosolic enzyme and is basically involved in ATP-dependent gluconeogenetic PEP production from OAA.In C3 plants, high PEPCK activities were detected duringgermination of oil-storing seeds, during fruit ripening, inphloem-associated cells, and in developing seeds. PEPCKis also found in trichomes of tobacco and cucumberleaves and is believed to be involved in the productionof secondary metabolites. There also appears to be afunction of PEPCK in plant defence reactions.
PPDK [EC 2.7.9.1]
In C3 plants, low PPDK activities were reported fora number of different tissue types and, depending onthe species, it is found only in chloroplasts or in bothchloroplasts and the cytosol (Aoyagi and Bassham, 1984;Nomura et al., 2000). The function of PPDK in C3 plantsis less clear. However, for guard cells of Vicia faba it hasbeen proposed that chloroplastic PPDK in concert withcytosolic NADP-ME could play a role in gluconeogeneticPEP generation from pyruvate during stomatal closure(Schnabl, 1981).
Metabolite transporters
High fluxes of C4 photosynthesis are accompanied bytransport processes across membranes, particularly theinner chloroplast envelope. In the mesophyll cells,pyruvate enters the chloroplast via a pyruvate Hq
symporter as a substrate for PPDK (Flugge et al.,1985). Likewise, PEP generated within the chloroplastshas to get access to the cytosol via a PEPuphosphatetranslocator (PPT). Furthermore, there are significantlyhigh rates of OAAumalate exchange required for a fastproduction of malate inside the mesophyll chloroplasts.Moreover, in NADP-ME C4 plants, malate mustenter the bundle-sheath chloroplasts. Following oxidat-ive decarboxylation of malate by NADP-ME, pyruvate
Biotechnology and C3 photosynthesis 597

formed by this reaction has to be exported from thechloroplast.
Pyruvate imported into chloroplasts of C3 plantscan be used for fatty acid biosynthesis or the productionof branched chain amino acids. A rapid malateuOAAexchange across the inner envelope is required for themalate valve in C3 plants. However, the PPT is the onlymetabolite transporter also involved in C4 metabolism,which has been cloned from a variety of non-C4-tissuesand is found at low activities in the inner envelopemembrane of chloroplasts and non-green plastids ofC3 plants as well as in maize roots (Fischer et al., 1997).Most likely, the PPT provides plastids with PEP as a sub-strate for the shikimate pathway (Streatfield et al., 1999).Import of PEP (rather than export) is a prerequisite foraromatic amino acid biosynthesis, which is localizedentirely within this organelle and results in a huge varietyof secondary products downstream of phenylalanine(Schmid and Amrhein, 1995).
The need for an accelerated metabolite exchangeacross the chloroplast envelope as observed in C4 plantsought to be considered as well in attempts to introduceC4-like features into C3 plants. Therefore genes encodingthe relevant plastidial metabolite transporters, apart fromthe PPT, should be identified and isolated.
Progress to date in the physiological andbiochemical characterization of C3 plantsoverexpressing C4-cycle genes
As C4-cycle enzymes (and metabolite transporters) havedistinct functions in C3 plants (Table 1), an increase intheir activities by individual overexpression is likely toperturb metabolism or to trigger compensational changesin metabolic fluxes. Compensational changes in meta-bolism as a response to the overexpression of C4-cycleenzymes are interesting to study, as these changes maypoint to not yet unravelled regulatory interrelationshipsbetween different metabolic pathways. It is conceivablethat a fully operational single cell C4-cycle circumventssuch metabolic perturbations.
Overexpression of PEPC is the first step for establishinga C4 cycle in C3 plants
The efficient fixation of atmospheric CO2 by PEPC isa prerequisite for the insertion of a single cell C4 cycle inC3 plants. The first cDNAs of the C4-type PEPCsequences for cloning were obtained from maize (Izuiet al., 1986) and Flaveria trinervia (Poetsch et al., 1991). Asuccessful introduction of the maize PEPC into transgenictobacco plants under the control of its own promoter orthe mesophyll-specific promoter of the chlorophyll aubbinding protein gene (cab) was achieved (Hudspeth et al.,1992) and later on by using the constitutively expressing
cauliflower mosaic virus (CaMV) 35S promoter (Kogamiet al., 1994; Benfey and Chua, 1990). In these types oftransgenic plants PEPC activity was increased slightlymore than 2-fold. Apart from an increase in the malatecontents, there were no apparent effects on the rate ofCO2 assimilation. However, in contrast to the wild type,the quantum yield for CO2 assimilation appeared to beunaffected by increasing temperatures in one transgenicline (Kogami et al., 1994). The lack of decrease in thequantum yield with increasing temperature suggested arefixation of respired CO2 by PEPC in the transgenics.However, the CO2 compensation point measured in asealed chamber remained unaffected in transgenictobacco plants compared to the control plants(Hudspeth et al., 1992; Kogami et al., 1994).
In rice, high expression of the maize PEPC wasachieved recently using the complete maize PEPC geneincluding exons and introns and its own promoter fortransformation and resulted in a 110-fold increase inPEPC activity measured in vitro (Ku et al., 1999). Thereported decline in O2 inhibition of CO2 assimilationwould have been consistent with an attenuation of photo-respiration. However, a more detailed analysis of theseplants suggested that this effect was likely to be based onphosphate limitation of photosynthesis under conditionsthat promote high photosynthetic fluxes (Fukayama et al.,2001; Matsuoka et al., 2000). Still, it is hard to conceiveas to why phosphate limitation of photosynthesis shouldoccur with an increase in PEPC activitiy. Provided thatPEP is generated via glycolysis starting from triosephosphates exported from the chloroplast in the light,the phosphate stoichiometry would be balanced as foreach triose phosphate exported one phosphate is releasedfrom PEP by PEPC and could serve as a counter exchangesubstrate regardless of whether the flux into sucrose bio-synthesis was slowed down or not (Fig. 4). If phosphatelimitation was the cause for lower CO2 assimilation ratesunder optimum conditions, it is therefore more likelythat it occurs at the site of cytoplasmic glyceraldehyde-3-phosphate dehydrogenase, leading to the subsequentformation of ATP by phosphoglycerate kinase. Thisreaction sequence could (at least temporarily) depletecytoplasmic phosphate pools required for triose phos-phate counter exchange. It is also conceivable thatadditional OAA formed by PEPC is reduced to malatein the stroma and hence competes with reducing equi-valents needed for 3-PGA reduction. However, to theknowledge of the authors, neither of these alternativeexplanations has been addressed experimentally.
As potato was chosen as a model plant for one labor-atory study, cDNAs and genomic clones of the endo-genous C3-type PEPC were isolated (Merkelbach et al.,1993). However, it was then decided to use PEPC froma bacterial source rather than the endogenous enzymefrom potato or other plants. Bacterial enzymes lack, for
598 Hausler et al.

instance, the regulatory properties of plant enzymes, suchas the covalent modification by phosphorylation. Over-expression of two bacterial PEPC genes from Escherichiacoli and Corynebacterium glutamicum under the control ofthe CaMV 35S promoter resulted eventually in a 5-foldincrease in PEPC activity in potato leaves with the enzymefrom C. glutamicum (Gehlen et al., 1996). Depending onthe composition of the in vitro assay an increase of up to20-fold could be determined (Hausler et al., 2001). Similarto the report on the PEPC overexpressors of tobacco(Hudspeth et al., 1992; Kogami et al., 1994), the absoluterates of photosynthetic CO2 assimilation and electrontransport were not severely affected in potato plantsoverexpressing the bacterial PEPC or in plants with anantisense repression of the endogenous enzyme. Likewise,the CO2 compensation point (C), determined in a closedchamber was unaffected in the trangenics. However, darkCO2 release after illumination was considerably increasedin PEPC overexpressors and diminished in the antisenseplants (Gehlen et al., 1996). This suggested that PEPCaffects the rate of respiratory CO2 release. More recentlyit could be demonstrated that the CO2 compensationpoint (C*) independent of dark respiration in the light(Rd) (determined according to Brooks and Farquhar,1985), was appreciably diminished in PEPC overexpres-sors and slightly increased in potato plants with an anti-sense repression of the endogenous PEPCs (Fig. 5)(Hausler et al., 1999). Rd (the rate of dark respirationin the light) was increased in a range of PEPC over-expressors and slightly decreased in the antisense plants.As C* solely reflects the kinetic properties of Rubisco(Brooks and Farquhar, 1985), but not accounts forcarboxylation of PEP by O2-insensitive PEPC, the changesin C* were interpreted as changes in the CO2uO2 ratio inthe vicinity of Rubisco, which was most likely due toan increased release of CO2 from products of PEP carb-oxylation. In theory, even more CO2 can be deliberatedfrom PEP carboxylation products than is actually fixedvia PEPC in the form of HCO�
3 . If, for instance, malatewere completely oxidized in the citric acid cycle (Fig. 4),this would result in the release of four CO2 for eachcarboxylation of PEP on the expense of photoassimilates.However, all intermediate stoichiometry of HCO�
3 uptakeand CO2 release might occur as well. An enhanced CO2
release by decarboxylating enzymes would result in anefficient HCO�
3 uCO2 pump (provided that the activityof cytosolic carbonic anhydrase is absent or low), whichcould increase the CO2 concentration in the mesophyllcells as well as in the vicinity of Rubisco leading to aslight inhibition of photorespiration. These theoreticalconsiderations were reinforced by the observation thatoverexpression of PEPC induced the endogenous cyto-solic NADP-ME of potato plants by a factor of 4–6(Hausler et al., 2001). This induction was accompanied byincreased activities of a putative cytosolic pyruvate kinase
and a presumably cytosolic NADP isocitrate dehydro-genase (Chen, 1998) suggesting that an increased activityof cytosolic NADP-ME in potato leaves initiatesCO2 release from PEP carboxylation products (Hausleret al., 2001).
In another approach to increase PEPC activity, theendogenous potato enzyme was modified in a way thatthe phosphorylation site was mutated, which yielded an
Fig. 5. Typical dependencies of CO2 assimilation from the intercellularCO2 concentration (Ci ) required for the determination of C* and Rd
in control potato (A), a transformed line overexpressing PEPC fromC. glutamicum (B) and a transformed line with antisense repression ofthe endogenous PEPC (C). Gas exchange measurements were done at22 8C and a relative humidity of 30% at limited PFDs (11, 74, 100 mmolm�2 s�1) and a saturated PFD (320 mmol m�2 s�1). The intercept of theAuCi dependencies indicate C* (on the Ci-axis) and �Rd (on the A-axis).The error for the calculation of the intercept for the individual setsof measurements was below 5%. (This figure is taken from Hausleret al., 1999.)
Biotechnology and C3 photosynthesis 599

enzyme with a higher affinity for PEP and loweredsensitivity towards malate inhibition. A detailed reporton the analysis of these plants is in preparation byRademacher et al. Transgenic potato plants with a4–5-fold increase in the activity of the modified versionof potato PEPC also exhibited a pleiotropic increasein endogenous cytosolic NADP-ME (Hausler et al.,2001).
The sole increase in PEPC activity thereforeappears to perturb metabolic fluxes and can lead toa waste of photoassimilates (T Rademacher, personalcommunication).
Stomatal movement is affected in PEPC transgenicsof potato
PEPC from C. glutamicum was expressed under thecontrol of the CaMV 35S promoter. Hence the enzymeis constitutively expressed in most tissues and it wasvery likely to be also present in stomatal guard cells.Interestingly, stomatal opening was accelerated in PEPCoverexpressors and delayed in PEPC antisense plantscompared to the wild type (Gehlen et al., 1996). The aver-age half time for stomatal opening was 3 min, 9 min, and6 min for PEPC overexpressors, the antisense plants andthe wild type, respectively. Furthermore, oscillations instomatal conductance were also observed in PEPCoverexpressors. These side-effects, underlining the roleof PEPC during stomatal movement (see also Asai et al.,2000), deserve a more thorough investigation.
Pleiotropic effects on secondary metabolism
Overexpression of PEPC from bacterial and plant sourcesnot only affects primary metabolism, but also appearsto have an effect on the abundance of secondary plantproducts. The contents of UV protectants (flavonoids),for instance, were significantly smaller in potato plantsoverexpressing PEPC from C. glutamicum or the mutatedversion of the endogenous enzyme (Hausler et al., 2001)and were also lower in analogous transgenic tobaccoplants (Jun Li, unpublished data). This might be explainedin terms of a limitation on PEP import into the chloro-plast. PEPC might compete with the initial steps of theshikimate pathway for PEP, which serves as one of theprecursors, and thereby diminishes the flux throughthe shikimate pathway. This is an hypothesis which needsto be proven, particularly in comparison with plants over-expressing PPDK, which are capable of producingadditional PEP within the chloroplasts.
Overexpression of decarboxylating enzymes
The introduction of PEPC activity is only the first stepin establishing an intracellular CO2 pump in C3
plants. Depending on the desired mode of CO2 releaseeither PEPCK or NADP malic enzyme could serve as
decarboxylases within the chloroplasts. The simplestway to release CO2 would be to decarboxylate OAA byPEPCK (Fig. 3B). This would have the advantage ofgenerating PEP as the substrate for PEPC without anyfurther enzymatic steps, but would also require (i) aunidirectional import of OAA into the chloroplasts andan efficient export of PEP from the chloroplasts.
For the second alternative, the decarboxylation ofmalate by NADP-ME, either the unidirectional transportof OAA or of malate would be required as malate couldeither be formed from OAA by stromal NADP-MDH orby extraplastidial NAD-MDH isozymes. Furthermore, asNADP-ME produces pyruvate within the chloroplast, anadditional conversion into PEP by PPDK or analternative enzyme is necessary.
Overexpression of PEPCK
PEPCK is a cytosolic enzyme. It is therefore essential totarget it to the chloroplast by fusion to an appropriatechloroplast targeting peptide. A cytosolic overexpressionof PEPCK together with PEPC would result in a futilecycle. Recently, Susuki et al. have reported that an over-expression of PEPCK in rice can lead to a substantialactivity of 3 U mg�1 chlorophyll (Suzuki et al., 2000). ThePEPCK gene from Urochloa panicoides was fused tothe Rubisco small subunit (rbcS) transit sequence andexpressed under the control of the maize PEPC or PPDKpromoter. Elegant carbon isotope feeding experimentsrevealed that a higher flux of 14CO2 into C4 compoundswas presumably caused by increased cytosolic PEPlevels resulting in a higher flux through PEPC. Thesefindings suggest that the PPT allows a significant effluxof PEP from the chloroplast in rice plants. It also impliesthat OAA can enter the chloroplast without any knowncounter exchange substrate. Moreover, by feeding radio-labelled malate the same authors could show that CO2
released by the action of PEPCK enters the Calvin cycleand causes a 3-fold increase in the labelling of 3-PGA andsucrose compared to the controls. The rates of CO2
assimilation and the CO2 compensation point remainedunaffected. The lack of effect on photosynthesis is con-sistent with data obtained for transgenic tobacco plants(Hausler et al., 2001) overexpressing the PEPCK genefrom Sinorhizobium meliloti (Osteras et al., 1995) fused tothe rbcS stromal transit sequence. However, unlike inrice, PEPCK activity could only be detected reliablyin isolated chloroplasts of transgenic tobacco plants(Hausler et al., 2001). The introduction of the sameconstruct in potato plants was not successful.
Overexpression of chloroplastic NADP-ME
For the second approach, the increase in NADP-ME,the cDNA of chloroplastic NADP-ME from the C3
plant Flaveria pringlei (Lipka et al., 1994) was used to
600 Hausler et al.

transform potato and tobacco plants under the control ofthe double CaMV 35S pomoter (Lipka et al., 1999;Hausler et al., 2001). However, due to high activities ofendogenous cytosolic NADP-ME in solanaceous speciescombined with relatively poor expression levels, thepresence of the introduced chloroplastic enzyme couldonly be confirmed in chloroplast extracts (Lipka et al.,1999). It was increased 5-fold with reference to chloro-plast protein. Overexpression of the C3 chloroplastenzyme had no large effects on photosynthetic perform-ance of transgenic potato plants. Single NADP-MEoverexpressors of tobacco were also generated, but havenot been analysed on a physiological level.
High expression of the maize C4-type NADP-MEin rice under the control of either the cab or CaMV35S promoter have been reported independently by twoJapanese groups (Takeuchi et al., 2000; Tsuchida et al.,2001). The transgenic rice plants were severely comprom-ized in growth and exhibit photoinhibition and photo-damage combined with a decline in chlorophyll contentsprobably due to an excessive NADPH production withinthe stroma in the light. As an indicator for the stromalNADPHuNADP ratio the activation state of stromalNADP-MDH (i.e. the in vivo activity as a fraction of theactivity of the fully activated enzyme) was doubled in thetransgenic lines compared to the wild type (Tsuchidaet al., 2001). Moreover, in one approach the ultrastruc-ture of the chloroplasts appeared to be agranal, similar tobundle-sheath chloroplasts of NADP-ME C4 plants(Takeuchi et al., 2000). From attempts to determine C*,a perturbation of the stromal redox states was also pro-posed for potato plants overexpressing NADP-ME(Hausler et al., 2001). However, due to the relativelylow activity of the introduced NADP-ME, the transgenicpotato lines lacked any visible phenotype.
Overexpression of enzymes that regenerate PEPfrom pyruvate
If CO2 is released by overexpressed NADP-malic enzyme,the pyruvate formed has to be converted to PEP as thesubstrate for PEPC. This could be achieved by over-expressing PPDK or bacterial PEP synthetase (PEPS), anenzyme catalysing a similar reaction as PPDK but with-out PPi formation. As outlined earlier, PEP also serves asa substrate for the shikimate pathway, which is localizedentirely in the chloroplast. Chloroplasts and most non-green plastids lack the ability to produce PEP via glyco-lysis. Thus, PEP has to be imported from the cytosolby the PPT. It is therefore conceivable that theadditional formation of PEP by overexpressed PPDK orPEPS could stimulate the flux through the shikimatepathway. In order to prevent these unpredictable side-effects, PPDK may be overexpressed in the cytosol(Sheriff et al., 1998).
Overexpression of PPDK
A successful overexpression of PPDK in C3 plants hasbeen reported for tobacco (Sheriff et al., 1998), potato(Ishimaru et al., 1998) Arabidopsis (Ishimaru et al., 1997),and rice (Nomura et al., 2000; Fukuyama et al., 2001).Overexpression of the C4 maize PPDK gene in potato(Ishimaru et al., 1998) resulted in about a 5-fold increasein activity and caused a depletion in pyruvate contentand a slight elevation of PEP content. Malate content wasalso considerably increased, presumably because of higherfluxes through PEPC and subsequent malic acid forma-tion. For rice plants, a drastic increase in PPDK activitywas achieved when the maize PPDK gene, includingintrons and exons as well as its own promoter andterminator, was used for transformation. Despite the factthat approximately 35% of the total leaf protein consistedof introduced PPDK, the transformants lacked a visiblephenotype or changes in growth or fertility (Fukayamaet al., 2001). Transgenic potato and tobacco plantsoverexpressing the C4-type PPDK gene from Flaveriatrinervia under the control of the 35S promoter have alsobeen generated in the laboratory and await a thoroughanalysis.
Overexpression of PEPS
As plant PPDK is highly regulated (i.e. by reversiblephosphorylation), the PEPS gene from E. coli was isol-ated and characterized (Niersbach et al., 1992). Potatoplants were generated overexpressing PEPS targeted tothe chloroplasts by fusion to the rbcS transit sequence(Panstruga et al., 1997). There were no large effects onphotosynthetic CO2 assimilation and electron transport.However, transpiration rates were slightly higher com-pared to the wild type and stomatal closure was delayedappreciably indicating a perturbation in the metabolismof stomatal guard cells (Panstruga et al., 1997). Therewere some specific effects on the contents of certainamino acids in the transgenics. The leaf contents ofglutamate were lowered by 25% whereas aspartate andasparagine contents were increased by more than 30%compared to the wild type. Contents of branched chainamino acids deriving from pyruvate were also altered. Thereason for these changes has not been further elucidated(R Panstruga et al., unpublished information). PEPSoverexpressors were compromised in shoot fresh weight,final heights of the plants and tuber yield, and leavesturned slightly yellowish and appeared wilted whenthe plants were grown in a non-air-conditioned green-house during temporary hot periods in the summer(R Panstruga et al., unpublished observations). However,these effects were absent when plants were grown undera controlled temperature and light regime in a growthcabinet.
Biotechnology and C3 photosynthesis 601

Overexpression of the PPT
One prerequisite for an operational C4-cycle in C3 plantsis the efficient export of PEP formed in the chloroplastby PPDK or PEPCK targeted to the chloroplast. In C3
plants there is only low activity of a chloroplastic PPT(Fischer et al., 1997; however see also Suzuki et al., 2000).Tobacco transformants overexpressing the heterologousPPT from cauliflower buds under the control of theCaMV 35S promoter have been generated and character-ized in the laboratory of UI Flugge. In the transformants,transport rates for PEP were increased 10-fold comparedto the wild type (L Voll, P Nicolay, K Fischer, REHausler, UI Flugge, unpublished results).
Double transformants
The introduction of individual C4-cycle genes in C3 cropscan be regarded as a test for their functionality in thetransformed plant species. A combined overexpression ofC4-cycle genes could provide additional information onwhether or not these enzymes may work in concert. Asoutlined above, single overexpression of C4-cycle genesmight perturb metabolism, as all C4 enzymes also havedistinct functions in C3 plants. The analysis of doubleand multiple transformants might help to underline thefeasibility of the approach to establish a single cell C4
cycle in C3 crops.
Combined overexpression of PEPC and chloroplasticNADP-ME
In order to release CO2 directly at the site of Rubisco,potato double transformants with increased activitiesof PEPC and chloroplastic NADP-ME from Flaveriapringlei were generated (Lipka et al., 1999). NADP-MEactivity in isolated chloroplasts of the transformantswas increased about 5-fold compared to the wild type.These double transformants exhibited a less pronouncedtemperature-dependent decrease of CO2 assimilationcombined with a considerable decline in the electronrequirement for CO2 assimilation (i.e. the electron trans-port rate over apparent CO2 assimilation) at elevatedtemperatures. Moreover, O2 inhibition of CO2 assimila-tion was attenuated significantly both at 25 8C and 35 8C(Hausler et al., 2001). Lower rates of electron transportfor similar or even higher rates of apparent CO2 assimila-tion as well as an attenuation of O2 inhibition of CO2
assimilation may point at an appreciable decline in photo-respiration rates. However, determinations of C* and Rd
produced equivocal results for double PEPCuNADP-MEoverexpressors (Hausler et al., 2001). C* appeared to bedecreased from 45 ml l�1 in the wild type to about20 ml l�1 in the double transformants only at an inter-mediate photon flux density (PFD), but not at high lightor very low PFDs. This suggests that the NADPHproduced by chloroplastic NADP-ME might perturb
stromal redox potentials, particularly at high PFDs(compare Tsuchida et al., 2001).
From the potato system another interesting aspectemerged. The induction of the endogenous cytosolicNADP-ME as a result of PEPC overexpression wasattenuated in double transformants containing PEPCand chloroplastic NADP-ME in combination despite theidentical activities of overexpressed PEPC as comparedto the single transformants (Hausler et al., 2001). Thissuggests that a redirection of the metabolic flux into thechloroplast circumvents pleiotropic changes observed inthe single transformants and leads to the assumption thata fully operational C4 cycle could be decoupled from theresidual metabolism in C3 plants provided that all of itscomponents are expressed in a well-balanced way.
Combined overexpression of PEPC with NADP-ME orPEPCK in tobacco
A moderate overexpression of PEPC from C. glutamicumcombined with NADP-ME from F. pringlei or withPEPCK from S. meliloti in tobacco plants had nosubstantial effects on the photosynthetic performance ofthe plants (Hausler et al., 2001).
Combined overexpression of PEPC with PPDK inrice plants
Recently Ku et al. reported a 35% increase in thephotosynthetic capacity and a 22% increase in grain yieldin rice plants overexpressing PEPC and PPDK in com-bination (Ku et al., 2001). A detailed analysis of theseplants is awaited. It is conceivable that this observation islinked to an increased provision of PEP by PPDK for theshikimate pathway inside the chloroplast (Streatfield et al.,1999; Matsuoka et al., 2000), an assumption, whichrequires experimental support.
Multiple overexpression of C4-cycle enzymes
The introduction of a fully operational C4 cycle mightultimately give the clue as to whether or not C3 photo-synthesis can be improved. Multiple overexpressorsof potato have been obtained by stepwise transforma-tion with plasmids containing different resistance genes(kanamycin, hygromycin, sulphonylamide, and BASTA1
wbarx) for selection. Transgenic lines containing PEPCfrom C. glutamicum or the mutated endogenous PEPC,combined with NADP-ME, PPDK and the PPT havebeen generated and await a thorough analysis. Fortobacco, transgenic plants overexpressing single C4-cyclegenes were genetically crossed and a number of hetero-zygous lines were obtained that overexpress multiplegenes (PEPC, NADP-ME, PPDK, PEPS, PEPCK, as wellas the PPT wFig. 3Bx) in various combinations (Jun Li,unpublished results). The progenies of these plants are
602 Hausler et al.

being screened at the moment for better growth in anatmosphere containing a lowered CO2 concentration.
Species-dependent metabolic responses towardsoverexpression of C4-cycle genes
Recent data (Hausler et al., 2001) show that even closelyrelated species such as potato and tobacco respond dif-ferentially towards an overexpression of C4-cycle genes.For instance, the induction of cytosolic NADP-MEin PEPC overexpressors was less apparent in tobaccotransformants compared to transgenic potato plants.Moreover, the most pronounced attenuating effects onphotorespiration were observed with PEPCuNADP-MEdouble transformants of potato plants, but for singlePEPC overexpressors of tobacco plants. It is likely thatthese species-dependent differences in the acceptance forredirecting metabolism by introduced C4-cycle enzymesmight hinder a successful introduction of an operationalC4-cycle in all of the crop plants intended to be used inthis approach.
Future perspectives and possiblefield applications
There has been significant progress in the overexpressionof the key enzymes of C4-type biochemistry in transgenicC3 plants. Nevertheless, it is still uncertain whether thisapproach will be sufficient to suppress photorespiration.The following section will discuss possible alternativeanduor supplementary approaches to improve a C4-typephotosynthesis in C3 plants.
Is a C4-like CO2 pump in C3 plants realistic onthe basis of current experimental data?
So far only a limited number of transgenic plants con-taining a maximum of two C4-cycle enzymes have beeninvestigated physiologically. Most of the observed effectswere based on pleiotropic changes in metabolism. Theefficiency by which aquatic plants are capable of switch-ing from a C3-type to a C4-like metabolism in single cellswhen the availability of CO2 declines is encouraging inthe prospect that it might be possible to establish a similarsystem in terrestrial C3 crops by a transgenic approach.Moreover, the terrestrial chenopod B. aralocaspicaappears to contain a single cell CO2-concentratingmechanism, distributed between the cytosol, the mito-chondria and two types of chloroplasts in only one typeof chlorenchyma cells (Freitag and Stichler, 2000;Vozesenskaya et al., 2001). Although this is a very excit-ing discovery, it ought to be considered that the anatomyof this halophytic chenopod is adapted to semi-dryenvironments with succulent leaves, central vascularbundles, a hypodermis, large chlorenchyma cells, waterstoring cells, and, most relevantly, a small intercellular air
space. These features of leaf anatomy are particularlydifferent from those of the C3 crops used in this study’stransgenic approaches.
In an aquatic system, only diffusion of CO2 throughthe water and the cell wall resistance for CO2 have tobe considered, whereas in terrestrial C3 plants diffusionthrough the surrounding air, stomatal conductance forCO2, and diffusion through the air space within the leaveshave to be taken into account as well (Farquhar et al.,1980; Laisk and Loreto, 1996). In C4 plants the gas-tightbundle-sheath prevents losses of CO2 from the cells andallows high CO2 environment in the vicinity of Rubisco.It is still an open question, whether or not the CO2
released by the introduced HCO�3 uCO2 pump can be
efficiently retained within the leaf, or whether a good partof it diffuses out of the cells into the intercellular spaceand would then just result in a waste of energy (see alsoLeegood, 2002). It might, for instance, become necessaryto decrease the free air space in leaves of C3 crops anduorto minimize stomatal aperture.
What about carbonic anhydrase?
The initial step of C4 photosynthesis is the rapid con-version of CO2 into HCO�
3 catalysed by carbonicanhydrase (CA) to provide the substrate for carbon fixa-tion by PEPC (Hatch and Burnell, 1990). It is estimatedthat photosynthesis in C4 plants would be slowed downby a factor of 104 in the absence of cytoplasmic CA(Badger and Price, 1994). However, this aspect has notbeen addressed so far in biotechnological approaches totransfer C4-like features into C3 plants. Similar considera-tions as for CA might apply for pyrophosphatase andadenylate kinase activities, which are essential for themetabolism of the PPDK reaction products AMP andPPi in the chloroplast. However, both enzymes are highlyactive in C3 chloroplasts (Gould and Winget, 1973;Schlattner et al., 1996).
Engineered C4 enzymes
Most approaches so far have been aimed at the introduc-tion of the highest possible activities of the respectiveenzymes in transgenic C3 plants. This is reasonablebecause these enzymes are expressed to very high levels inleaves of C4 plants. However, a high expression of theintroduced enzyme is not necessarily linked with highin vivo activities. Modulation of enzyme activity bycovalent modification (i.e. PEPC, PPDK) and the avail-ability of co-factors and substrates (for review seeLeegood, 1997) ought to be considered as well. Forinstance, C4-specific PEPC is phosphorylated in the lightto reduce its sensitivity towards malate inhibition. It isquestionable whether the protein kinase required forphosphorylation is expressed in the correct temporaland spatial pattern in C3 plants. Furthermore, C4 PEPC
Biotechnology and C3 photosynthesis 603

has a low substrate affinity for PEP, which might beunfavourable in the C3 environment. C3 isoforms ofPEPC show higher affinities to their respective substrates,but are very sensitive to inhibition by malate (Svenssonet al., 1997). The catalytic properties of PEPC frompotato have been modified by genetic engineering(T Rademacher et al., unpublished data) in the way thatthe enzyme possesses both C3 (high affinity towards PEP)and C4 (reduced sensitivity towards product inhibition)features and is not subjected to covalent modification.It appears to be useful and perhaps even necessary toengineer more of the C4-cycle enzymes anduor therespective promoters to be better adapted to the specificrequirements of a C4-like cycle in a ‘C3 environment’.
Morphological adaptations
The introduction of a ‘true C4 cycle’ into C3 plants wouldultimately require alterations in the leaf anatomy. Thereare numerous deviations from the classical C3 pattern ofleaf anatomy associated with the C4 syndrome in bothmono- and dicotyledonous plants (Nelson and Langdale,1992). However, most of these features are not absolutelyessential to perform C4 photosynthesis (Dengler andNelson, 1999). For example, the suberin lamella aroundthe bundle-sheath cells and the agranal ultrastructure ofbundle-sheath chloroplasts observed in NADP-ME typeC4 plants like maize are absent in other C4 species. Event-ually, it will be crucial to separate primary carbon fixationand photosynthetic carbon reduction in two differenttissue types or in distinct parts of one cell (as realized inB. aralocaspica) which are in intimate contact, and tominimize the leakage of CO2 from the tissue or the part ofthe cell where Rubisco is active. Bundle-sheath cells withdistinct morphology compared to mesophyll cells are alsopresent in C3 plants. In Arabidopsis, reticulate mutantshave been isolated with disturbed chloroplast develop-ment in mesophyll cells, but intact chloroplasts in thebundle-sheath indicating that the different cell typesfollow a separated developmental programme (Kinsmanand Pyke, 1998). Differences in developmental pro-grammes of mesophyll and bundle-sheath in C3 plantsmight be utilized to target distinct components of the C4
cycle into the respective cell types. Moreover, the effici-ency of metabolite exchange between the different celltypes would require closer vein spacing inside the leaf.The signals determining the density of veins are notdefinitely identified, but there is significant variabilitywithin a single organism. For instance, maize foliar leavesshow a high vein density significant for C4 plants, whereasthe leaf sheath or the husk leaf exhibit distances betweenthe veins similar to C3 plants (Langdale et al., 1988).Refined mutant screens and comparative gene expressionanalyses will be necessary to identify the factorsresponsible for changes in leaf anatomy.
Promoters
One very successful strategy is the transfer of com-plete genes from maize into rice (Ku et al., 1999). Thisapproach leads to very high expression levels andresembles, at least in part, the spatial and light-dependentexpression patterns found in C4 plants. OverexpressedC4-cycle enzymes might perturb the metabolism of wholeplants if transcription is not restricted to the photo-synthetic tissues. The use of endogenous promoters fromC3 plants with properties similar to the C4 promoterswould be advantageous. Recently, Tsuchida et al. usedthe rice cab promoter in rice for the overexpression ofmaize NADP-ME, which led to a strong accumulation ofthe protein in chloroplasts (Tsuchida et al., 2001).Alternatives for leaf-specific and light-induced geneexpression include the use of other photosynthetic pro-moters like the lhcb promoter (Cerdan et al., 2000) or therbcS promoter (Kyozuka et al., 1993).
Furthermore, inducible promoters could be used (Gatzand Lenk, 1998; Zuo and Chua, 2000). A comparisonof the induced with the non-induced state of expressionwould aid physiological analysis under controlledconditions.
Field applications
C4 plants are more productive than C3 plants whenthey are grown under their respective optimum conditions(Brown, 1999). C4 plants exhibit higher water and nitro-gen use efficiencies compared to C3 plants, which resultsin an increased dry matter production (reviewed in Brown,1999). Concentrating CO2 at the site of Rubisco shouldallow engineered C3 plants to reduce stomatal conduct-ance under drought conditions without a dramatic declinein the rate of CO2 assimilation (Drake et al., 1997). Thiswould allow both the use of new areas for cropproduction required for feeding the growing worldpopulation and the reduction of inputs into the system(such as fertilizers) and thus conserving natural resources.Both factors are much more relevant to today’s necessitiesthan the mere increase in biomass production.
Acknowledgements
The authors would like to thank Dr Mitsue Miyoa and HiroshiFukayama for the kind provision of as yet unpublished data.
References
Andreo CS, Gonzales DH, Iglesias AA. 1987. Higher plantphosphoenolpyruvate carboxylase: structure and regulation.FEBS Letters 213, 1–8.
Andrews M. 1986. The partitioning of nitrate assimilationbetween root and shoot of higher plants. Plant, Cell andEnvironment 9, 511–519.
604 Hausler et al.

Aoyagi K, Bassham JA. 1984. Pyruvate orthophosphate dikinasemRNA organ specificity in wheat and maize plants. PlantPhysiology 76, 278–280.
Arp WJ, Van Mierlo JEM, Berendse FEM, Snijders W. 1998.Interaction between elevated CO2 concentration, nitrogen andwater: effects on growth and water use of six perennial plantspecies. Plant, Cell and Environment 21, 1–11.
Asai N, Nakajima N, Tamaoki M, Kamada H, Kondo N. 2000.Role of malate synthesis mediated by phosphoenolpyruvatecarboxylase in guard cells in the regulation of stomatalmovement. Plant Cell Physiology 41, 10–15.
Badger MR, Price GD. 1994. The role of carbonic anhydrase inphotosynthesis. Annual Review of Plant Physiology and PlantMolecular Biology 45, 369–392.
Benfey PN, Chua NH. 1990. The Cauliflower Mosaic Virus 35Spromoter combinatorial regulation of transcription in plants.Science 250, 959–966.
Besford RT. 1990. The greenhouse effects on the acclimationof tomato plants growing in high carbon dioxide relative tochanges in Calvin cycle enzymes. Journal of Plant Physiology136, 458–463.
Brooks A, Farquhar GD. 1985. Effect of temperature on theCO2uO2 specificity of ribulose-1,5-bisphosphate carboxylaseuoxygenase and the rate of respiration in the light. Planta 165,
397–406.Brown RH. 1999. Agronomic implications of C4 photosynthesis.
In: Sage RF, Monson RK, eds. C4 plant biology. AcademicPress, 473–507.
Casati P, Drincovich MF, Edwards GE, Andreo CS. 1999.Minireview. Malate metabolism in plant defence. Photosyn-thesis Research 61, 99–105.
Casati P, Lara MV, Andreo CS. 2000. Induction of a C4-likemechanism of CO2 fixation in Egeria densa, a submergedaquatic species. Plant Physiology 123, 1611–1621.
Cerdan PD, Staneloni RJ, Ortega J, Bunge MM,Rodriguez-Batiller MJ, Sanchez RA, Casal JJ. 2000.Sustained but not transient phytochrome A signaling targetsa region of an lhcb1*2 promoter not necessary for phyto-chrome B action. The Plant Cell 12, 1203–1211.
Chen K, Hu G, Kreutgen N, Lenz F. 1997. The effect ofCO2 concentration on strawberries. I. Plant growth analysis.Journal of Applied Botany 71, 168–172.
Chen R. 1998. Plant NADP-dependent isocitrate dehydro-genases are predominantly localized in the cytosol. Planta207, 280–285.
Chen Z, Spreitzer RJ. 1992. How various factors influencethe CO2uO2 specificity of ribulose-1,5 bisphosphatecarboxylaseuoxygenase. Photosynthesis Research 31, 157–164.
Chollet R, Vidal J, O’Leary MH. 1996. Phosphoenolpyruvatecarboxylase: a ubiquitous, highly regulated enzyme in plants.Annual Review of Plant Physiology and Plant MolecularBiology 47, 273–298.
Cockburn W, Ting IP, Sternberg LO. 1979. Relationshipsbetween stomatal behaviour and internal carbon dioxideconcentration in crassulacean acid metabolism plants. PlantPhysiology 63, 1029–1032.
Cooper TG, Wood HG. 1971. The carboxylation of phospho-enolpyruvate and pyruvate. II. The active species of‘CO2’ utilized by phosphoenolpyruvate carboxylase andpyruvate carboxylase. Journal of Biological Chemistry246, 5488–5490.
de Groote D, Kennedy RA. 1977. Photosynthesis in Elodeacanadensis Michx. Plant Physiology 59, 1133–1135.
Dengler NG, Nelson T. 1999. Leaf structure and development inC4 plants. In: Sage RF, Monson RK, eds. C4 plant biology.San Diego: Academic Press, 133–172.
Douce R, Neuburger M. 1989. The uniqueness of plantmitochondria. Annual Review of Plant Physiology and PlantMolecular Biology 40, 371–414.
Drake BG, Gonzalez-Meler MA, Long SP. 1997. More efficientplants: a consequence of rising atmospheric CO2? AnnualReview of Plant Physiology and Plant Molecular Biology48, 609–639.
Duff SMG, Chollet R. 1995. In vivo regulation of wheat-leafphosphoenolpyruvate carboxylase by reversible phosphoryla-tion. Plant Physiology 107, 775–782.
Edwards GE, Andreo CS. 1992. NADP-malic enzymes fromplants. Photochemistry 31, 1845–1857.
Edwards GE, Furbank RT, Hatch MD, Osmond CB. 2001. Whatdoes it take to be C4? Lessons from the evolution of C4
photosynthesis. Plant Physiology 125, 36–49.Farquhar GD, von Caemmerer S, Berry JA. 1980. A biochemical
model of photosynthetic CO2 assimilation in leaves of C3
species. Planta 149, 78–90.Fischer K, Kammerer B, Gutensohn M, Arbinger B, Weber A,
Hausler RE, Flugge UI. 1997. A new class of plastidicphosphate translocators: a putative link between primary andsecondary metabolism by the phosphoenolpyruvateuphosphate antiporter. The Plant Cell 9, 453–462.
Flugge UI, Stitt M, Heldt HW. 1985. Light-driven uptake ofpyruvate into mesophyll chloroplasts from maize. FEBSLetters 183, 335–339.
Freitag H, Stichler W. 2000. A remarkable new leaf type withunusual photosynthetic tissue in a central Asiatic genus ofChenopodiacea. Plant Biology 2, 154–160.
Fukayama H, Tsuchida H, Agarie S et al. 2001.Significant accumulation of C4-specific pyruvate, orthophos-phate dikinase in a C3 plant, rice. Plant Physiology127, 1136–1146.
Gatz C, Lenk I. 1998. Promoters that respond to chemicalinducers. Trends in Plant Science 3, 352–358.
Gehlen J, Panstruga R, Smets H, Merkelbach S, Kleines M,Porsch P, Fladung, Becker I, Rademacher T, Hausler RE,Hirsch H-J. 1996. Effects of altered phosphoenolpyruvatecarboxylase activities on transgenic C3 plant Solanumtuberosum. Plant Molecular Biology 32, 831–848.
Gould JM, Winget GD. 1973. A membrane-bound alkalineinorganic pyrophosphatase in isolated spinach chloroplasts.Archives of Biochemistry and Biophysics 154, 606–613.
Hatch MD, Burnell JN. 1990. Carbonic anhydrase activity inleaves and its role in the first step of C4 photosynthesis. PlantPhysiology 93, 825–828.
Hatch MD, Osmond CB. 1976. Compartmentation and trans-port in C4 photosynthesis. In: Stocking CR, Heber U, eds.Encyclopedia of plant physiology, New series, Vol. 3. Berlin:Springer Verlag, 144–184.
Hausler RE, Kleines M, Uhrig H, Hirsch H-J, Smets H. 1999.Overexpression of phosphoenolpyruvate carboxylase fromCorynebacterium glutamicum lowers the CO2 compensationpoint (C*) and enhances dark and light respiration intransgenic potato. Journal of Experimental Botany 336,
1231–1242.Hausler RE, Rademacher T, Li J, Lipka V, Fischer KL,
Schubert S, Kreuzaler F, Hirsch H-J. 2001. Single and doubleoverexpression of C4-cycle genes had differential effects on thepattern of endogenous enzymes, attenuation of photorespira-tion and on contents of UV protectants in transgenic potatoand tobacco plants. Journal of Experimental Botany 52,
1785–1803.Holaday AS, Bowes G. 1980. C4 acid metabolism and dark
CO2 fixation in a submerged aquatic macrophyte (Hydrillaverticillata). Plant Physiology 65, 331–335.
Biotechnology and C3 photosynthesis 605

Hudspeth RL, Grula JW, Dai Z, Edwards GE, Ku MSB. 1992.Expression of maize phosphoenolpyruvate carboxylase intransgenic tobacco. Plant Physiology 98, 458–464.
Ishimaru K, Ishikawa I, Matsuoka M, Ohsugi R. 1997. Analysisof a C4 maize pyruvate, orthophosphate dikinase expressed inC3 transgenic Arabidopsis plants. Plant Science 129, 57–64.
Ishimaru K, Okawa Y, Ishige T, Tobias DJ, Ohsugi R. 1998.Elevated pyruvate, orthophosphate dikinase (PPDK)activity alters carbon metabolism in C3 transgenic potatoeswith a C4 maize PPDK gene. Physiolgia Plantarum 103,
340–346.Izui K, Ishijima S, Yamagushi Y, Katagiri F, Murata T,
Shigesada K, Sugiyama T, Katsuki H. 1986. Cloning andsequence analysis of cDNA encoding active phosphoenol-pyruvate carboxylase of the C4-pathway of maize. NucleicAcid Research 14, 1615–1628.
Jordan DB, Ogren WL. 1984. The CO2uO2 specificity of ribulose1,5-bisphosphate carboxylaseuoxygenase. Dependence onribulose bisphosphate concentration, pH and temperature.Planta 161, 308–313.
Kellogg EA. 1999. Phylogenetic aspects of the evolution of C4
photosynthesis. In: Sage RF, Monson RK, eds. C4 plantbiology. London: Academic Press, 411–443.
Keys AJ, Bird IF, Cornelius MJ, Lea PJ, Wallsgrove RM,Miflin BJ. 1978. Photorespiratory nitrogen cycle. Nature 275,741–743.
Kinsman EA, Pyke KA. 1998. Bundle sheath cells and cell-specific plastid development in Arabidopsis leaves.Development 125, 1815–1822.
Knee M, Finger FL, Lagrimini M. 1996. Evidence for a cytosolicNADP-malic enzyme in tomato. Phytochemistry 42, 11–16.
Kogami H, Shono M, Koike T, Yanagisawa S, Izui K, Sentoku N,Tanifuji S, Uchimiya H, Toki S. 1994. Molecular andphysiological evaluation of transgenic tobacco plants expres-sing a maize phosphoenolpyruvate carboxylase gene under thecontrol of the cauliflower mosaic virus 35S promoter.Transgenic Research 3, 287–296.
Ku MSB, Agarie S, Nomura M, Fukayama H, Tsuchida H,Ono K, Hirose S, Toki S, Miyao M, Matsuoka M. 1999. High-level expression of maize phosphoenolpyruvate carboxylase intransgenic rice plants. Nature Biotechnology 17, 76–80.
Ku MSB, Cho DH, Li X, Jiao DM, Pinto M, Miyao M,Matsuoka M. 2001. Introduction of genes encoding C4
photosynthesis enzymes into rice plants: physiological con-sequences. Rice Biotechnology: Improving Yield, StressTolerance and Grain Quality 236, 100–116.
Ku MSB, Kano-Murakami Y, Matsuoka M. 1996. Evolution andexpression of C4 photosynthesis gene. Plant Physiology 111,
949–957.Kyozuka J, McElroy D, Hayakawa T, Xie Y, Wu R,
Shimamoto K. 1993. Light-regulated and cell-specific expres-sion of tomato rbcS-gusA and rice rbcS-gusA fusion genes intransgenic rice. Plant Physiology 102, 991–1000.
Laisk A, Loreto F. 1996. Determining photosynthetic para-meters from leaf CO2 exchange and chlorophyll fluorescence.Ribulose-1,5-bisphosphate carboxylaseuoxygenase specificityfactor, dark respiration in the light, excitation distributionbetween photosystems, alternative electron transport ratesand mesophyll diffusion resistance. Plant Physiology 110,
903–912.Langdale JA, Zelitch I, Miller E, Nelson T. 1988. Cell position
and light influence C4 versus C3 patterns of photosyntheticgene expression in maize. EMBO Journal 7, 3643–3651.
Latzko E, Kelly GJ. 1983. The many-faceted function ofphosphoenolpyruvate carboxylase in C3 plants. PhysiologiaVegetale 21, 805–815.
Leegood RC, Acheson RM, Tecsi LI, Walker RP. 1999. Themany-faceted function of phosphoenolpyruvate carboxy-kinase in plants. In: Kruger NJ, Hill SA, Ratcliffe RG, eds.Regulation of primary metabolism in plants. Dodrecht:Kluwer, 37–57.
Leegood RC. 1997. The regulation of C4 photosynthesis.Advances in Botanical Research 26, 251–316.
Leegood RC. 2002. C4 photosynthesis: principles of CO2 con-centration and prospects for its introduction into C3 plants.Journal of Experimental Botany 53, 581–590.
Lipka B, Steinmuller K, Rosche E, Borsch D, Westhoff P. 1994.The C3 plant Flaveria pringlei contains a plastidicNADP-malic enzyme which is orthologous to the C4 isoformof the C4 plant F. trinervia. Plant Molecular Biology 26,
1775–1783.Lipka V, Hausler RE, Rademacher T, Li J, Hirsch H-J,
Kreuzaler F. 1999. Solanum tuberosum double transgenicexpressing phosphoenolpyruvate carboxylase and NADP-malic enzyme display reduced electron requirement for CO2
fixation. Plant Science 144, 93–105.Magnin NC, Cooley BA, Reiskind JB, Bowes G. 1997.
Regulation and localization of key enzymes during theinduction of Kranz-less, C4-type photosynthesis in Hydrillaverticillata. Plant Physiology 115, 1681–1689.
Martinoia E, Rentsch D. 1994. Malate compartmentation—responses to a complex metabolism. Annual Review of PlantPhysiology and Plant Molecular Biology 45, 447–467.
Matsuoka M, Fukayama H, Tsuchida H, Nomura M, Agari S,Ku MSB, Miyao M. 2000. How to express some C4
photosynthesis genes at high levels in rice. In: Sheehy JE,Mitchell PL, Hardy B. eds. Redesigning rice photosynthesis toincrease yield. Proceedings of the workshop on The Questto Reduce Hunger: Redesigning rice photosynthesis, 30November to 3 December 1999, Los Banos, Phillipines.International Rice Research Institute and Amsterdam:Elsevier Science BV, 167–175.
Matsuoka M, Furbank RT, Fukayama H, Miyao M. 2001.Molecular engineering of C4 photosynthesis. Annual Review ofPlant Physiology and Plant Molecular Biology 52, 297–314.
Melzer E, O’Leary MH. 1987. Anaplerotic CO2 fixation byphosphoenolpyruvate carboxylase in C3 plants. PlantPhysiology 84, 58–60.
Merkelbach S, Gehlen J, Denecke M, Hirsch HJ, Kreuzaler F.1993. Cloning, sequence analysis and expression of a cDNAencoding active phosphoenolpyruvate carboxylase of theC3 plant Solanum tuberosum. Plant Molecular Biology 23,881–888.
Nelson T, Langdale JA. 1992. Developmental genetics of C4
photosynthesis. Annual Review of Plant Physiology and PlantMolecular Biology 43, 25–47.
Niersbach M, Kreuzaler F, Geerse RH, Postma PW, Hirsch HJ.1992. Cloning and nucleotide sequence of the Escherichia coliK-12 ppsA gene, encoding PEP synthase. Molecular Geneticsand Genomics 231, 332–336.
Nomura M, Sentoku N, Tajima S, Matsuoka M. 2000.Expression patterns of cytoplasmic pyruvate, orthophosphatedikinase of rice (C3) and maize (C4) in a C3 plant, rice.Australian Journal of Plant Physiology 27, 343–347.
O’Leary MH. 1982. Phosphoenolpyruvate carboxylase: anenzymologist’s view. Annual Review of Plant Physiology33, 297–315.
Ogren WL. 1984. Photorespiration: pathways, regulation andmodification. Annual Review of Plant Physiology 35, 415–442.
Oliver DJ. 1994. The glycine decarboxylase complex from plantmitochondria. Annual Review of Plant Physiology and PlantMolecular Biology 45, 323–337.
606 Hausler et al.

Osmond CB. 1978. Crassulacean acid metabolism, a curiosity incontext. Annual Review of Plant Physiology 29, 379–414.
Osteras M, Driscoll BT, Finan TM. 1995. Molecular andexpression analysis of the Rhizobium meliloti phosphoenolpyruvate carboxykinase ( pckA) gene. Journal of Bacteriology177, 1452–1460.
Panstruga R, Hippe-Sanwald S, Lee Y-K, Lataster M, Lipka V,Fischer R, Liao YC, Hausler RE, Kreuzaler F, Hirsch HJ.1997. Expression and chloroplast-targeting of active phopho-enolpyruvate synthetase from Escherichia coli in Solanumtuberosum. Plant Science 127, 191–205.
Poetsch W, Hermans J, Westhoff P. 1991. Multiple cDNAs ofphosphoenolpyruvate carboxylase in the C4 dicot Flaveriatrinervia. FEBS Letters 292, 133–136.
Polle A. 1996. Mehler reaction: friend or foe in photosynthesis?Botatica Acta 109, 84–89.
Raschke K, Hedrich R, Reckmann U, Schroeder JI. 1988.Exploring biophysical and biochemical components of theosmotic motor that drives stomatal movement. Botanica Acta101, 283–294.
Reiskind JB, Madsen TV, van Ginkel LC, Bowes G. 1997.Evidence that inducible C4-type photosynthesis is a chloro-plastic CO2-concentrating mechanism in Hydrilla, a sub-merged monocot. Plant, Cell and Environment 20, 211–220.
Sage RF, Pearcy RW. 1987. The nitrogen use efficiency of C3 andC4 plants. I. Leaf nitrogen, growth and biomass partitioningin Chenopodium album (L.) and Amaranthus retroflexus (L.).Plant Physiology 84, 954–958.
Salvucci ME, Bowes G. 1981. Induction of reduced photo-respiratory activity in submerged and amphibious aquaticmacrophytes. Plant Physiology 67, 335–340.
Salvucci ME, Bowes G. 1983. Two photosynthetic mechanismsmediating the low photorespiratory state in submergedaquatic angiosperm. Plant Physiology 73, 488–496.
Schaaf J, Walter MH, Hess D. 1995. Primary metabolism inplant defense. Regulation of bean malic enzyme genepromoter in transgenic tobacco by developmental andenvironmental cues. Plant Physiology 108, 949–960.
Scheibe R. 1987. NADPq-malate dehydrogenase in C3 plants:regulation and role of a light-activated enzyme. PhysiologiaPlantarum 71, 393–400.
Schlattner U, Wagner E, Greppin H, Bonzon M. 1996.Chloroplast adenylate kinase from tobacco: purification andpartial characterization. Phytochemistry 42, 589–594.
Schmid J, Amrhein N. 1995. Molecular organization of theshikimate pathway in higher plants. Phytochemistry 39,737–749.
Schnabl H. 1981. The compartmentation of carboxylating anddecarboxylating enzymes in guard cell protoplasts. Planta152, 307–313.
Sheriff A, Meyer H, Riedel E, Schmitt JM, Lapke C. 1998. Theinfluence of plant pyruvate, orthophosphate dikinase on a C3
plant with respect to the intracellular location of the enzyme.Plant Science 136, 43–57.
Spalding MH, Stumpf DK, Ku MSB, Burris RH, Edwards GE.1979. Crassulacean acid metabolism and diurnal variations ofinternal CO2 and O2 concentrations in Sedum praealtum DC.Australien Journal of Plant Physiology 6, 557–567.
Spencer WE, Wetzel RG, Teeri J. 1996. Photosyntheticphenotype plasticity and the role of phosphoenolpyruvatecarboxylase in Hydrilla verticillata. Plant Science 118, 1–9.
Streatfield SJ, Weber A, Kinsman EA, Hausler RE, Li J,Post-Beitenmiller D, Kaiser WM, Pyke KA, Flugge UI,Chory J. 1999. The phosphoenolpyruvateuphosphate trans-locator is required for phenolic metabolism, palisade celldevelopment and plastid-dependent nuclear gene expression.The Plant Cell 11, 1609–1621.
Suzuki S, Murai N, Burnell J, Arai M. 2000. Changes inphotosynthetic carbon flow in transgenic rice plants thatexpress C4-type phosphoenolpyruvate carboxykinase fromUrochloa panicoides. Plant Physiology 124, 163–172.
Svensson P, Blasing OE, Westhoff P. 1997. Evolution of theenzymatic characteristics of C4 phosphoenolpyruvate carb-oxylase—a comparison of the orthologous ppcA phospho-enolpyruvate carboxylases of Flaveria trinervia (C4) andFlaveria pringlei (C3). European Journal of Biochemistry 246,
452–460.Takeuchi Y, Akagi H, Kamasawa N, Osumi M, Honda H. 2000.
Aberrant chloroplasts in transgenic rice plants expressing ahigh level of maize NADP-dependent malic enzyme. Planta211, 265–274.
Teramura AH, Sullivan JH, Ziska LH. 1990. Interaction ofelevated UV-B radiation and carbon dioxide on productivityand photosynthetic characteristics in wheat, rice and soybean.Plant Physiology 94, 470–475.
Tsuchida H, Tamai T, Fukayama H, Agarie S, Nomura M,
Onodera H, Ono K, Nishizawa Y, Lee B-H, Hirose S, Toki S,
Ku MSB, Matsuoka M, Miyao M. 2001. High level expressionof C4-specific NADP-malic enzyme in leaves and impairmentof photoautotrophic growth in a C3 plant, rice. Molecularengineering of C4 photosynthesis. Plant Cell Physiology42, 138–145.
van Ginkel LC, Bowes G, Reiskind JB, Prins HBA. 2001. ACO2-flux mechanism operating via pH-polarity in Hydrillaverticillata leaves with C3 and C4 photosynthesis.Photosynthesis Research 68, 81–88.
Van Quy L, Lamaze T, Champigny M-L. 1991. Short-termeffects of nitrate on sucrose synthesis in wheat leaves. Planta185, 53–57.
Voznesenskaya EV, Franceschi VR, Kiirats O, Freitag H,Edwards GE. 2001. Kranz anatomy is not essential forterrestrial C4 plant photosynthesis. Nature 414, 543–546.
Zhang XQ, Li B, Chollet R. 1995. In vivo regulatory phos-phorylation of soybean nodule phosphoenolpyruvate carb-oxylase. Plant Physiology 108, 1561–1568.
Zuo J, Chua N-H. 2000. Chemical-inducible systems forregulated expression of plant genes. Current Opinion inBiotechnology 11, 146–151.
Biotechnology and C3 photosynthesis 607
Related Documents











