Organic carbon sequestration in earthworm burrows Axel Don a, * , Bert Steinberg a , Ingo Scho ¨ ning a,1 , Karin Pritsch b , Monika Joschko c , Gerd Gleixner a , Ernst-Detlef Schulze a a Max-Planck-Institute for Biogeochemistry, Hans-Kno ¨ll Strasse 10, 07745 Jena, Germany b Helmholtz Zentrum Mu ¨nchen, German Research Center for Environmental Health (GmbH), Institute of Soil Ecology, Ingolstaedter Landstrasse 1, 85764 Neuherberg, Germany c Leibniz-Centre for Agricultural Landscape Research, Eberswalder Strasse 84, 15374 Muencheberg, Germany article info Article history: Received 16 August 2007 Received in revised form 12 March 2008 Accepted 19 March 2008 Available online 8 May 2008 Keywords: Anecic earthworms C turnover Earthworm burrows Lumbricus terrestris Soil organic carbon abstract Earthworms strongly affect soil organic carbon cycling. The aim of this study was to determine whether deep burrowing anecic earthworms enhance carbon storage in soils and decrease C turnover. Earthworm burrow linings were separated into thin cylindrical sections with different distances from the burrow wall to determine gradients from the burrow wall to the surrounding soil. Organic C, total N, radiocarbon ( 14 C) concentration, stable isotope values (d 13 C, d 15 N) and extracellular enzyme activities were measured in these samples. Anecic earthworms increased C stocks by 270 and 310 g m 2 accumulated in the vertical burrows. C-enrichment of the burrow linings was spatially highly variable within a distance of millimetres around the burrow walls. It was shown that C accumulation in burrows can be fast with C sequestration rates of about 22 g C m 2 yr 1 in the burrow linings, but accumulated C in the burrows may be mineralised fast with turnover times of only 3–5 years. Carbon stocks in earthworm burrows strongly depended on the earthworm activity which maintains continuous C input into the burrows. The enhanced extracellular enzyme activity of fresh casts was not persistent, but was 47% lower in inhabited burrows and 62% lower in abandoned burrows. Enzyme activities followed the C concentrations in the burrows and were not further suppressed due to earthworms. Radiocarbon concentrations and stable isotopes in the burrow linings showed an exponential gradient with the youngest and less degraded organic matter in the innermost part of the burrow wall. Carbon accumulation by anecic earthworm is restricted to distinct burrows with less influence to the surrounding soil. Contrary to the initial hy- pothesis, that organic C is stabilised due to earthworms, relaxation time experiments with nuclear magnetic resonance spectroscopy (NMR) did not reveal any enhanced adsorption of C on iron oxides with C stabilising effect. Our results suggest that earthworm activity does not substantially increase subsoil C stocks but burrows serve as fast ways for fresh C transport into deep soil horizons. Ó 2008 Elsevier Ltd. All rights reserved. 1. Introduction Soil carbon dynamics are influenced by an array of biotic and abiotic factors among which earthworms play a prominent role (Wolters, 2000). They consume up to 2 t litter ha 1 yr 1 which can be 100% of the annual litter fall, and simultaneously ingest 1200 t yr 1 of mineral soil (Raw, 1962). While processing mineral soil and detritus, less than 9% of the ingested organic carbon (C) is assimilated by the earthworms (Lee, 1985). Even in an extreme example where endogeic (horizontally burrowing) earthworms were newly introduced to North America causing rapid decline of the organic layer thickness, organic matter was only mixed into the mineral soil. Carbon loss of the entire system was small after 14 year of invasion of earthworms (Alban and Berry, 1994). Earthworms are present in almost all ecosystems around the globe with particularly high abundances in grasslands, where they increase productivity (Partsch et al., 2006). They have been cate- gorized into three functional groups, determined by their feeding behaviour and habitat (Bouche ´ , 1977): (1) Epigeic earthworms are litter feeders that live in the organic layer, (2) endogeic earthworms feed their way horizontally through the upper mineral soil and (3) anecic earthworms generally inhabit one single vertical burrow for their whole life. Anecic earthworms can build burrows down to 5 m depth at maximum, mostly reaching 1–2 m depth, and transport fresh organic detritus from the soil surface into the burrows while mixing with mineral soil (Lee, 1985). Carbon stored in the deeper mineral soil is assumed to be more protected from mineralisation * Corresponding author. Max-Planck-Institute for Biogeochemistry, Hans-Kno ¨ ll- Strasse 10, 07745 Jena, Germany. Tel.: þ49 3641 576184; fax: þ49 3641 577100. E-mail address: [email protected] (A. Don). 1 Present address: Friedrich Schiller University, Institute for Ecology, Dornburger Strasse 159, 07743 Jena, Germany. Contents lists available at ScienceDirect Soil Biology & Biochemistry journal homepage: www.elsevier.com/locate/soilbio 0038-0717/$ – see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.soilbio.2008.03.003 Soil Biology & Biochemistry 40 (2008) 1803–1812

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Soil Biology & Biochemistry 40 (2008) 1803–1812
Contents lists avai
Soil Biology & Biochemistry
journal homepage: www.elsevier .com/locate/soi lb io
Organic carbon sequestration in earthworm burrows
Axel Don a,*, Bert Steinberg a, Ingo Schoning a,1, Karin Pritsch b, Monika Joschko c, Gerd Gleixner a,Ernst-Detlef Schulze a
a Max-Planck-Institute for Biogeochemistry, Hans-Knoll Strasse 10, 07745 Jena, Germanyb Helmholtz Zentrum Munchen, German Research Center for Environmental Health (GmbH), Institute of Soil Ecology,Ingolstaedter Landstrasse 1, 85764 Neuherberg, Germanyc Leibniz-Centre for Agricultural Landscape Research, Eberswalder Strasse 84, 15374 Muencheberg, Germany
a r t i c l e i n f o
Article history:Received 16 August 2007Received in revised form 12 March 2008Accepted 19 March 2008Available online 8 May 2008
Keywords:Anecic earthwormsC turnoverEarthworm burrowsLumbricus terrestrisSoil organic carbon
* Corresponding author. Max-Planck-Institute for BStrasse 10, 07745 Jena, Germany. Tel.: þ49 3641 5761
E-mail address: [email protected] (A. Don).1 Present address: Friedrich Schiller University, Inst
Strasse 159, 07743 Jena, Germany.
0038-0717/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.soilbio.2008.03.003
a b s t r a c t
Earthworms strongly affect soil organic carbon cycling. The aim of this study was to determine whetherdeep burrowing anecic earthworms enhance carbon storage in soils and decrease C turnover. Earthwormburrow linings were separated into thin cylindrical sections with different distances from the burrowwall to determine gradients from the burrow wall to the surrounding soil. Organic C, total N, radiocarbon(14C) concentration, stable isotope values (d13C, d15N) and extracellular enzyme activities were measuredin these samples. Anecic earthworms increased C stocks by 270 and 310 g m�2 accumulated in thevertical burrows. C-enrichment of the burrow linings was spatially highly variable within a distance ofmillimetres around the burrow walls. It was shown that C accumulation in burrows can be fast with Csequestration rates of about 22 g C m�2 yr�1 in the burrow linings, but accumulated C in the burrows maybe mineralised fast with turnover times of only 3–5 years. Carbon stocks in earthworm burrows stronglydepended on the earthworm activity which maintains continuous C input into the burrows. Theenhanced extracellular enzyme activity of fresh casts was not persistent, but was 47% lower in inhabitedburrows and 62% lower in abandoned burrows. Enzyme activities followed the C concentrations in theburrows and were not further suppressed due to earthworms. Radiocarbon concentrations and stableisotopes in the burrow linings showed an exponential gradient with the youngest and less degradedorganic matter in the innermost part of the burrow wall. Carbon accumulation by anecic earthworm isrestricted to distinct burrows with less influence to the surrounding soil. Contrary to the initial hy-pothesis, that organic C is stabilised due to earthworms, relaxation time experiments with nuclearmagnetic resonance spectroscopy (NMR) did not reveal any enhanced adsorption of C on iron oxides withC stabilising effect. Our results suggest that earthworm activity does not substantially increase subsoil Cstocks but burrows serve as fast ways for fresh C transport into deep soil horizons.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Soil carbon dynamics are influenced by an array of biotic andabiotic factors among which earthworms play a prominent role(Wolters, 2000). They consume up to 2 t litter ha�1 yr�1 which canbe 100% of the annual litter fall, and simultaneously ingest1200 t yr�1 of mineral soil (Raw, 1962). While processing mineralsoil and detritus, less than 9% of the ingested organic carbon (C) isassimilated by the earthworms (Lee, 1985). Even in an extremeexample where endogeic (horizontally burrowing) earthwormswere newly introduced to North America causing rapid decline of
iogeochemistry, Hans-Knoll-84; fax: þ49 3641 577100.
itute for Ecology, Dornburger
All rights reserved.
the organic layer thickness, organic matter was only mixed into themineral soil. Carbon loss of the entire system was small after 14year of invasion of earthworms (Alban and Berry, 1994).
Earthworms are present in almost all ecosystems around theglobe with particularly high abundances in grasslands, where theyincrease productivity (Partsch et al., 2006). They have been cate-gorized into three functional groups, determined by their feedingbehaviour and habitat (Bouche, 1977): (1) Epigeic earthworms arelitter feeders that live in the organic layer, (2) endogeic earthwormsfeed their way horizontally through the upper mineral soil and (3)anecic earthworms generally inhabit one single vertical burrow fortheir whole life. Anecic earthworms can build burrows down to 5 mdepth at maximum, mostly reaching 1–2 m depth, and transportfresh organic detritus from the soil surface into the burrows whilemixing with mineral soil (Lee, 1985). Carbon stored in the deepermineral soil is assumed to be more protected from mineralisation

sample cylinder burrow
reference soil20 mm 0 - 0.75 mm
2.5-3.5 mm
1.5-2.5 mm
0.75-1.5 mm
Fig. 1. Sampling scheme of an earthworm burrow increment, illustrating the fourcylindrical samples taken around the burrow, which were between 0.75 and 1.0 mmthick and the reference sample which was taken for each burrow outside the drilo-sphere 20 mm from the burrow.
A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–18121804
than C in the organic layer or the A horizon (Paul et al., 1997). Themechanisms behind the stabilisation are still unclear. Low distur-bance frequencies, high amounts of minerals surfaces for physical Cprotection and a low nutritive value of organic matter in subsoilsmay explain high mean residual times. Moreover, anecic earth-worms can influence a horizontal soil volume of up to 1 cm thick-ness all along the burrows wall (Jegou et al., 2000). This zone iscalled the ‘‘drilosphere’’ (Lavelle, 1988) and C turnover may differhere to the surrounding soil.
On the one hand, earthworms enhance aggregation and areassumed to stimulate the formation of organo-mineral complexes.The formation of microaggregates within macroaggregates whichcontain protected occluded C seems to be enhanced by passagethrough the earthworm gut (Bossuyt et al., 2005). However, rapiddisintegration of aggregates may occur when polysaccharides andother organic gluing agents are mineralised and the protectedcarbon may again be released (Ge et al., 2001; Guggenberger et al.,1996). The passage of soil and detritus through the earthworm gutfacilitates the contact between C and mineral particles by grindingand mixing both components. Adsorption of C on mineral surfacesis considered to be an important stabilisation mechanism whichmay be enhanced by earthworm activity (von Lutzow et al., 2006).Bonds with iron and aluminium oxides are particularly strong andlead to decreased C turnover (Kaiser and Zech, 1999). Solid statenuclear magnetic resonance (NMR) spectroscopy was successfullyapplied to determine the formation of Fe–C bonds (Schoning et al.,2005).
On the other hand, several studies demonstrated a short termincrease of C turnover in earthworm casts due to priming: Earth-worm guts behave like bioreactors where microbial activity andbiomass are increased due to favourable conditions with readilyavailable C of mucus and water (Lavelle et al., 2004; Tiunov andScheu, 2000). Priming with mucus and water may persist for onlya short time. This effect can be demonstrated by measuring extra-cellular enzyme activities. Enzymes are excreted by microorgan-isms to hydrolyse and break down organic molecules to facilitatetheir ingestion. Enzymes are short lived and therefore, extracellularenzymes serve as indicators for the C turnover in soil. Lower Cturnover rates should be related to lower enzyme concentrationsper mass substrate. Whether enzyme activity and decompositionare suppressed in the long term, or if C stabilisation is enhanced byearthworm activity is largely unknown. Most existing earthwormstudies were performed with mesocosms for only short periods ofa few months maximum. Results from mesocosm studies cannot beused directly to scale up to larger scales. To our knowledge, there isno comprehensive field study that quantified the effect of anecicearthworm on C transport within the soil profile and C sequestra-tion in burrows over a longer time scale.
The following study was undertaken to answer the question asto whether anecic earthworms increase soil C stocks in the longterm and decrease C turnover in the drilosphere. The overall effectof anecic earthworms on C stocks and turnover in a temperategrassland system was quantified by analysing drilosphere micro-samples. Short term and long term effects caused by earthworms(Lumbricus terrestris) were investigated measuring enzyme activity,stable isotopes, NMR and 14C age of the burrow linings. Wehypothesised that persistent burrow structures provide space foradditional C in soils and that the C turnover in the burrows shouldbe decreased, due to physical stabilisation of organic C.
2. Material and methods
2.1. Site description
The study was carried out at two extensively managed grasslandsites located within Thuringia/Germany. The site ‘‘Jena’’ is situated
next to the River Saale in a floodplain with 587 mm average annualprecipitation and a mean annual temperature of 9.3 �C (11�37027E,50�57007N; 130 m.a.s.l.) (Roscher et al., 2004). The soil is a EutricFluvisol (WRB classification) developed from alluvial loam withgroundwater depth of approximately 2 m in summer. The site wasused as cropland between the early 1960s and 2000 and managedas grassland before and after this period. The sampling for thisstudy was conducted on grassland which was 4 years old. At the site‘‘Mehrstedt’’, Stagnic vertisols developed on clay rich limestonesediments, covered by loess (10�39022E, 51�16017N; 280 m.a.s.l.).The site was very dry (mean annual temperature 8.0 �C) due to thelimestone bedrock and low mean annual precipitation (551 mm). Itwas converted from cropland to grassland 23 years prior tosampling for this study (Scherer-Lorenzen et al., 2007).
2.2. Sampling
Horizontal cross-sections measuring 1�2 m2 at 10, 30, 60, 85and 110 cm depths were excavated on both sites. Mehrstedt addi-tionally had a 140 cm depth. Each site was prepared carefully andcleaned with a small broom and vacuum cleaner. These cross-sections were used to measure burrow numbers, diameter d (withdigital sliding calliper), and the location of burrows. Only burrowswith a diameter >3 mm were included in this study. The innersurface area of the burrows were calculated as pdl assuming a cy-lindrical burrow shape (l¼ length of the burrow). Burrows weresampled using 5 cm diameter core samplers to extract burrowincrements, 4 cm long (Fig. 1). Between 5 and 13 burrows weresampled at each sampling depth. Distances between sampledburrows were large enough (0.8–2.0 m) to insure the independenceof the samples.

A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–1812 1805
At the Jena site, we distinguished between inhabited andabandoned burrows: After excavating to a certain soil samplingdepth, we left the soil pit open and only loosely covered withthermoplastic foil for 20 h. Anecic earthworms closed the openingsof their burrows with faeces within a few hours to protect them.Closed burrows were sampled as ‘‘inhabited’’ and open burrowswere assumed to be ‘‘abandoned’’. The fresh casts on the burrowopenings were used to measure enzyme activity compared to thatin the burrow linings.
For enzyme analysis, earthworm burrows were sampled at30 cm depth at the Jena site. We sampled (1) inhabited, (2) longterm abandoned and (3) short term abandoned: inhabited burrowsfrom 30 cm soil depth were buried in place in horizontal arrange-ment to prevent any earthworm access for 11 days. Thus, with theexception of earthworm activity, the burrow conditions remainedunchanged. All three treatments were sampled with four replicates,stored at 2 �C and analysed within 14 days.
2.3. Sample preparation
The burrow linings (drilosphere) were divided into four sub-samples. Using a sequence of four drills with increasing diameters,we manually collected soil samples with increasing distances fromthe burrow wall (0.75, 1.5, 2.5, 3.5 mm) (Fig. 1). The two innersample cylinders were 0.75 mm thick; the outer two cylinders were1.0 mm thick. Additionally, the soil outside the drilosphere (ca.20 mm distance from burrow wall) was sampled (reference soil) tomeasure the background concentrations. Carbon concentrations inthese reference samples were similar to C concentrations ofindependent bulk samples for both sites (Don et al., 2007, Stein-beiss, S., personal communication). Only three different drilospherecylinders plus the reference soil were collected for enzymemeasurements. The inner sample cylinder was on average 1.1 mmthick; the other two were 1.25 mm.
2.4. Analysis
2.4.1. C, N analysisSamples were dried at 40 �C for 24 h and ground to powder.
Carbon concentrations of 250 mg ground subsamples weredetermined by dry combustion (VarioMax, Elementar) before (Ctot)and after (Cin) ignition at 450 �C for 16 h. Organic carbon (Corg)concentrations were calculated as Ctot� Cin. Total N was measuredsimultaneously with Ctot. Limit of determination of C was 0.12% andof N 0.05%. Throughout the following manuscript ‘‘C’’ is used asequivalent to ‘‘Corg’’.
Burrowing by anecic earthworms is mainly done by compres-sion of the surrounding soil with radial pressure of 72–93 kPa andaxial pressure of up to 65 kPa (Keudel and Schrader, 1999). Carbonwithin the compacted soil mass around the burrows and the soilmass void was accounted for in the following equation for the C-enrichment of the drilosphere [mass C] of each horizon k:
C-enrichment ¼ B1n
0@pr
12ðDh�1 � Dhþ1Þ
Xn
j¼1
X4
i¼1
�ci;j � cbulk;j
�
��
r2i;j � r2
i�1;j
�1A
where B is the number of burrows in the horizon k, r is the bulkdensity of horizon k [mass volume�1] (data from Don et al., 2007and Kreuziger, I., and Baade, J., personal communication), Dh is thedepth of the sampled horizon [length], c1.4 is the C concentrationof the cylindrical burrow sample [massC masssoil
�1 ], cbulk is theC concentration of the bulk soil outside the drilosphere
[massC masssoil�1 ], r1.4 is the radius of the cylindrical burrow sample
[length] and n is the number of samples j.The C-enrichment is a measure for additional C in a certain soil
depth due to the earthworm activity, taking into account thevarious C concentrations of the reference soil in different horizons.As burrow stability was high, we assumed that the soil was notdirectly influenced by earthworms if no burrow was visible(Hagedorn and Bundt, 2002).
Turnover times s for the upper 30 cm of the soil at the Jena sitewere calculated as (Paul and Clark, 1996):
s ¼ ðt � t0Þlnðct=ct0Þ
where t� t0 is the time span between C deposition by earthwormsand burrow sampling, ct0 is the organic C concentration in aban-doned burrows and ct0 is the organic C concentration in inhabitedburrows.
As a conservative estimation, we assumed a 2 years mean timeof burrows abundance, as all burrows had been destroyed byploughing 4 years before sampling. The earthworms mean life timewas assumed to be less than 1 year. Inhabited burrows wereassumed to represent C concentration of newly built burrows attime t.
2.4.2. Stable isotope and radiocarbon analysisd13C and d15N values were measured with continuous flow
isotope ratio mass spectrometry (IRMS). The isotope ratio massspectrometer (Deltaþ, Finnigan MAT) was combined with an ele-mental analyser (EA 1100, CE Instruments). The natural abundanceof 15N is reported as
d15N ¼�
Rsample=Rstandard � 1�
1000 ½&�
where R is the ratio of 15N/14N in samples and standards. Thestandard deviation of repeated measurements of laboratory stan-dards was 0.13&.
Radiocarbon concentration (14C) was determined with acceler-ator mass spectroscopy, 3 MV AMS (High Voltage EngineeringEurope) (Steinhof et al., 2004). Where necessary, samples weredecalcified with HCl and freeze dried to remove carbonate C.Carbondioxide evolved during dry combustion of 60–200 mgsample was reduced to graphite by heating a mixture of H2 and CO2
with iron powder at 650 �C. The graphite was pressed into targetsand measured with the AMS facility. All values were corrected forfractionation using d13C values. The mean analytical error was0.25 pMC. The 14C content was expressed in percent modern carbon(pMC).
2.4.3. Enzyme analysisThe activities of six different extracellular enzymes were
measured following the method of Pritsch et al. (2005) with thebelow detailed modifications. The measured enzymes are involvedin the turnover of plant components and soil organic matter such asstarch (a-glucosidase, a-gls) and cell wall compounds such ascellulose (b-glucosidase, b-gls; cellobiohydrolase, cel) or hemi-cellulose (xylosidase, xyl). Chitin (N-acetylglucosaminidase, Nag)originating from arthropod exoskeletons and fungal cell walls islikely to be enriched at the surface of burrows due to the coloni-sation of this nutrient rich habitat by fungi and arthropods. Bytaking up and transporting plant material and soil organic matter,these compounds are distributed and enriched by earthworms intheir casts and in the burrow linings and should therefore decreasetowards the bulk soil. Phosphatase (pho) activity was measured asan omnipresent, usually highly active enzyme in soil supplying cellswith phosphate that is required for all energy demanding processes.

0
20
40
60
80
100
120
140
0 100 200 300 400 500 600 700Number of burrows per m
2
So
il d
ep
th
[cm
]
Mehrstedt Jena
Fig. 2. Number of earthworm burrows >3 mm diameter at the sites Mehrstedt (opensymbols) and Jena (closed symbols).
A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–18121806
In brief, a mixture of 0.4 g of field-moist soil and 40 ml of distilledwater was shaken by hand for a few seconds, shaken with anextractor for 2�15 min and exposed to an ice-cooled ultrasonicbath with low energy input (<1 J) for 2� 3 min. The remainingmacroaggregates were carefully crushed with a spatula.
An aliquot of 50 ml of the suspension was transferred intoa microplate. The sample suspension was diluted with 50 ml ofdistilled water and 50 ml of methylumbelliferone (MU)-labelledsubstrates were added to start the incubation. All samples weremeasured in three replicates. Optimum substrate concentrationswere 400 mM for all except phosphatase (800 mM). All substrate-sample solutions were incubated for 60 min except MU-phosphate,which was incubated for just 20 min. Blank samples with distilledwater instead of sample suspension were added with six replicatesto test the substrates’ auto-fluorescence.
A calibration with all soil suspensions was done with six differentMU concentrations (0, 100, 200, 300, 400, 500 pmol MU in 50 mL)thus accounting for a possible quenching effect of the organic mol-ecules in the suspension. As there was no difference in quenchingbetween all three treatments(inhabited, short-term abandoned,long-term abandoned), we applied the same calibration curves forall treatments. Incubation was stopped with 2.5 M Tris (pH> 10).Microplates were centrifuged for 10 min and fluorescence emissionof MU was measured using a fluorescence spectrophotometer (CaryEclipse, Varian, Australia; excitation wave length: 360 nm, emission450 nm). The average emission rates of the samples without addedsubstrate were subtracted from the emission values of all samples.Calibration curves were used to calculate the enzyme activityexpressed as pmol MU mg�1 dry weight (DW) soil sample h�1.
2.4.4. NMR relaxation time experimentSolid state 13C NMR allow assessments of the chemical bonds in
soil organic matter by applying a cross-polarisation magic anglespinning. The obtained spectra are influenced by paramagneticmaterials such as iron. Close association between iron cations andorganic matter drastically reduce the spin-lattice relaxation time ofprotons in the rotating frame (T1rH). This feature is used by variablecontact time NMR experiments to assess soil organic carbon link-ages to iron oxides. A sample from each, the innermost part of anearthworm burrow and the surrounding soil outside the drilo-sphere (reference soil) from the Mehrstedt site (30 cm depth) wasanalysed in a Bruker DSX 200 spectrometer for 70 h each. Sampleswere spun in the magic angle at a frequency of 6.8 kHz. Untreatedsamples as well as samples which were previously treated with 10%hydrofluoric acid (HF) were used. Treatment with HF was applied toremove the paramagnetic iron from the soil and to concentrate thesoil organic carbon (Schoning et al., 2005). The total number ofscans with a constant recycle delay of 200 ms was 65,000 or 42,500for untreated samples and 14,000 for HF treated samples. NMRspectra were acquired using an array of 15 different contact timesvarying from 0.05 to 4.5 ms in untreated samples and using 24different contact times from 0.01 to 6 ms in HF treated samples. AllNMR spectra were subjected to Fourier transformation, phasingand baseline correction. The values of signal intensities of alkyl C(0–45 ppm), N-alkyl C (45–60 ppm) and O-alkyl C (60–110 ppm) atdifferent contact times were fitted with a two component approachto determine the spin relaxation time of protons (Abelmann et al.,2004). The fitting of a two component model with a fast and a slowdomain was applied using Mathcad 2000 Professional softwareaccording to the procedure described by Abelmann et al. (2004).
2.4.5. Species and biomass of earthwormsOn 26 May 2005, one soil block measuring 1 m� 1 m� 0.5 m
was excavated at both sites and hand sorted for earthworms. Inaddition, a mustard suspension was applied to the pit in order toexpel earthworms from the soil. The worms were subsequently
transported to the laboratory for further analysis. The fresh weightof earthworms was determined gravimetrically in a beaker withwater and species were determined according to Sims and Gerard(1985).
2.4.6. Statistical analysisVariates were characterised using classical descriptive statistics
(means and standard error of the mean). Throughout the paper,error bars and the error of mean values (�X) indicate the standarderror of the mean. The significance of different treatments wasaccessed using t-tests with a¼ 0.05. Statistical analysis was per-formed using Statistica 5.5 software.
3. Results
3.1. Burrow systems and the earthworm community
The number of earthworm burrows increased with increasingsoil depth up to a maximum at the 110 cm depth at the Mehrstedtsite and at 30 cm at the Jena site (Fig. 2). High burrow densities of>500 burrows per m2 were measured at both sites. Three hundredand forty four new burrows per m2 were constructed during a 4year period in the former ploughing horizon at the Jena site, ac-counting for almost 2/3 of the maximum burrow numbers at thissite. Taking into account all burrows >3 mm in diameter up to110 cm depth the soil surface exposed to the atmosphere increasedby a factor 8 at both sites, thus considerably increasing the surfacefor litter input. The mean burrow diameter was around 6 mm atboth sites and increased with increasing soil depth due to anincrease of larger burrows in the deeper soil horizons (Table 1).
The abundance of earthworms was low compared to the burrownumbers with 228 earthworms at the Jena site and 233 at theMehrstedt site. Anecic earthworms accounted for only 14% at theJena site (32 individuals) and 21% (49 individuals) of total earth-worms at the Mehrstedt site. Anecic earthworms made up 31% oftotal earthworm biomass in Jena (22 g m�2) and 57% in Mehrstedt(93 g m�2). This was due to their body size, which is considerablylarger than other earthworm groups. The only anecic earthworm

Table 1Earthworm burrow distribution for different size classes and sampling depths atJena and Mehrstedt sites [burrow number m�2] and mean burrow diameter [mm]
Diameter [mm] Jena Mehrstedt
10 cm 30 cm 60 cm 10 cm 30 cm 60 cm 85 cm 110 cm
3� x< 4 130 144 63 18 30 75 124 734� x< 5 91 134 69 29 44 58 95 1155� x< 6 62 75 70 19 31 76 101 786� x< 7 36 52 65 15 30 71 98 1137< x< 8 9 44 79 30 20 82 78 1088� x< 9 6 39 58 16 9 38 65 1009� x< 10 6 20 29 12 9 24 40 4010� x< 11 4 11 22 5 1 17 27 23�11 0 8 17 2 0 4 15 8
Mean burrowdiameter
4.5 5.3 6.5 5.9 5.4 6.0 5.9 6.3
A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–1812 1807
species at both sites was L. terrestris, which inhabited only 6–7% ofthe burrows at the horizon with maximum burrow density.
3.2. Increased C stocks due to earthworm burrows
At each depth, C concentrations were the highest in theinnermost part of the burrows and decreased exponentially withdistance from the burrow wall (Fig. 3). Total N concentrationsfollowed the same trend (data not presented). Down to 140 cmdepth, the drilosphere was C- and N-enriched compared to thesurrounding soil. However, the absolute and relative differencesbetween the drilosphere and the surrounding soil became smalleras soil depth increased.
At the Jena site, inhabited and abandoned burrows were sam-pled separately. We found a strong enhancing effect of earthwormpresence on the C concentrations in the burrows. To separate theeffect of the earthworms from the C dynamic in the soil profile, wecalculated the additional C in the drilosphere – called ‘‘C-enrichment’’ – by subtracting the background C concentration.Inhabited burrows showed significantly higher C-enrichmentcompared to abandoned burrows (Fig. 4). The C-enrichment inabandoned burrows was on average for all horizons only 33%compared to the enrichment in the inhabited burrows. The calcu-lated C-enrichment of the whole soil profile caused by anecic
10 cm depth30 cm depth60 cm depth85 cm depth110 cm depth140 cm depth
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
0.75 1.5 2.5 3.5 20Distance from burrow wall [mm]
Org
an
ic carb
on
co
ncen
tratio
n [%
]
Fig. 3. Organic C concentration [%] in the different layers around the earthwormburrows for different soil depths at the Mehrstedt site.
earthworms based on measured burrow volume and C concentra-tions in the drilosphere was 310 g C m�2 at the Mehrstedt site and270 g C m�2 at the Jena site, taking into account the different frac-tions of inhabited and abandoned burrows. The highest additionalC stock was detected in the subsoil between 30 and 60 cm depth(Fig. 5). Compared to the total C stocks of 11,200 g C m�2 for 0–100 cm depth at the Jena site and 8600 g m�2 at the Mehrstedt sitethe additional C of the drilosphere comprised 1.5% at the Jena siteand 2.8% at the Mehrstedt site (Steinbeiss, S., personal communi-cation; Don et al., 2007). At the Jena site, the drilosphere wassampled 4 years after the conversion of cropland to grassland. C-enrichment within the drilosphere of the former ploughing horizon(0–30 cm depth) took place during this period and was calculatedas 22 g C m�2 yr�1.
C turnover times (mean residence times) at 10 and 30 cm soildepths provided direct information on the C stability in earthwormburrows. In the innermost burrow wall, C turnover was about 5years at 10 cm depth and 3 years at 30 cm. Turnover times in-creased with increasing distance to the burrow wall with 8 and 6years (0.75–1.5 mm distance from burrow wall), 14 and 10 years(1.5–2.5 mm distance from burrow wall) and 10 and 35 years (2.5–3.5 mm distance from burrow wall) at 10 and 30 cm soil depths,respectively.
3.3. Origin and 14C age of carbon in the drilosphere
The horizontal gradients of C and N from the burrow wall to thebulk soil resembled those of vertical gradients within soil profiles.Along with the C and N gradients, we found d15N values increasedfrom the inner burrow wall to the surrounding reference soil(Fig. 6). Low stable isotope values indicated a low level of degra-dation of the organic matter within the drilosphere. Fresh detritushad a low stable d15N value of around 1.9& at the Mehrstedt siteand 3.8& at the Jena site. The d15N value difference betweenburrow wall and surrounding reference soil corresponded to aboutone trophic level of the food web (Minagawa and Wada, 1984). Thed13C values showed the same tendencies but had an offset of severalper mil depending on the carbonate content of the samples (notshown). In most horizons, the whole sampled drilosphere up to3.5 mm distance from the burrow wall was influenced by freshorganic matter.
Radiocarbon concentrations of the samples confirmed the C agegradient casts< burrows< surrounding soil (Table 2). This gradientshowed that the burrow walls were under continuous construction.Casts had younger 14C ages than burrows and the inner and outerpart of the burrows had different 14C ages too. The 14C concentra-tions in fresh casts at 60 cm on both sites were similar to recentatmospheric 14C concentration (102.7 and 101.5 pMC in casts,104.3 pMC in recent biomass). Thus, fresh detritus was transporteddirectly into deep soil horizons. High 14C ages at 60 cm depth in thesurrounding soil, especially at the site Mehrstedt with 79 pMC(>2000 yr turnover time), showed that there is little input of freshC into these horizons. The differences between 14C ages of burrowsand surrounding soil were less in the surface soil horizons. At 10 cmsoil depth, the input of fresh roots led to similar 14C ages in thesurrounding soil to that in the burrows.
3.4. Enzyme activity in burrows and casts
The activity of all measured extracellular enzymes wasenhanced in the drilosphere compared to bulk soil (Fig. 7). Thisdifference was significant (p< 0.05) for a- and b-glucosidase,cellobiohydrolase, xylosidase, and N-acetylglucosaminidase but notfor phosphatase in inhabited burrows but only for three out of sixenzymes in abandoned burrows. Similarly to the C and N gradients,enzyme activity gradients from the burrow wall to the surrounding

Distance from burrow wall [mm]
Org
an
ic carb
on
co
ncen
tratio
n [%
]
10 cm depth
0.0
1.5
3.0
4.5 30 cm depth
0.75 1.5 2.5 3.5 20 0.75 1.5 2.5 3.5 20
60 cm depth
0.0
1.5
3.0
4.5 85 cm depth
0.75 1.5 2.5 3.5 20 0.75 1.5 2.5 3.5 20
inhabited burrowsabandoned burrows
Fig. 4. Organic C concentrations [%] in the different layers around the earthworm burrow from inhabited and abandoned burrows at four different depths at the Jena site.
0
A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–18121808
soil were smaller in abandoned burrows compared to inhabitedburrows. Our results indicate a slight decrease in enzyme activityafter the earthworm burrow was recently abandoned: Within 11days after cessation of earthworm activity the enzyme activitydropped on average by 8% in the innermost ring of the earthwormburrow (Table 3). Only for xylosidase we found increased activity 11days after earthworm removal, however, this increase was notsignificant (p¼ 0.52). The earthworm casts derived from the samesoil horizon as the drilosphere samples, had up to seven timeshigher activity than the average drilosphere enzyme activity, 0–3.4 mm from the burrow wall (Table 3).
40
80
120
1600 20 40 60 80 100 120
C stock change due to earthworms [g m-2]
So
il d
ep
th
[cm
]
JenaMehrstedt
Fig. 5. C stock change [g C m�2] due to C-enrichment in anecic earthworm burrows fordifferent soil depth intervals at the Jena site (closed symbols) and the Mehrstedt site(open symbols).
3.5. Long term stabilisation due to adsorption on mineral surfaces –NMR relaxation time experiment
Adsorption on mineral surfaces were found to stabilise C in soils(Kaiser and Zech, 1999). Iron oxides in particular reveal largespecific surfaces and can interact with organic compounds. Here,a sample from the earthworm burrow was compared with thesurrounding soil (reference soil). No clear differences were visiblein the chemical composition of the 13C solid state NMR spectraacquired at a contact time of 1 ms. Both samples were dominatedby O-alkyl C with the main peak being at 71 ppm and alkyl C withthe main peak being at 27 ppm. The C recovery in HF treatedsamples was 81% for the burrow sample and 73% for the referencesoil sample. NMR relaxation experiments go beyond conventionalNMR spectra and provide an insight into Fe–organic matterinteractions (Schoning et al., 2005). Fe3þ impacts on 13C NMRspectra if it is in close vicinity to 13C–H groups by shortening thespin-lattice relaxation times (T1rH1 and T1rH2) of protons. Re-laxation times of protons surrounding the major C species did notshow any distinct difference between earthworm burrows ascompared to the reference soil (Table 4). Protons in the surroundingof alkyl C and N-alkyl C showed slightly shorter relaxation timesT1rH in the reference soil compared to the burrow. However, thesame tendency was found in the HF treated samples, which
indicates that the different relaxation times of burrow and refer-ence soil are not a result of different degrees of iron influence. Ingeneral, the proton relaxation times for alkyl, N-alkyl and O-alkyl inthe untreated sample from the burrows as well as in the referencesoil were in a similar range and were all >2 ms. There was no in-dication for a preferential interaction between iron oxides and anyspecific C species in the earthworm burrow. Our results showed

3
4
5
6
7
8
9
0.75 1.5 2.5 3.5 200.75 1.5 2.5 3.5 20
Distance from burrow wall [mm]
δ15N
valu
e [‰
]
110 cm depth85 cm depth30 cm depth10 cm depth
85 cm depth60 cm depth30 cm depth10 cm depth
Mehrstedt Jena
Fig. 6. d15N values in the different distances from the earthworm burrow wall for different soil depth at the sites Jena and Mehrstedt.
A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–1812 1809
that earthworms did not contribute to an increased adsorption ofany compound of soil organic C on iron oxides.
Table 214C concentrations [percentage modern carbon pMC %] of burrows and surroundingsoil samples from the sites Jena and Mehrstedt
Site Soil depth [cm] Sample pMC [%]
Mehrstedt 10 Inner burrow (0–1 mm) 102.0Mehrstedt 10 Surrounding soil 102.9Mehrstedt 30 Inner burrow (0–1 mm) 101.5Mehrstedt 30 Reference soil 94.4Mehrstedt 60 Cast 102.7Mehrstedt 60 Inner burrow (0–1 mm) 95.2Mehrstedt 60 Outer burrow (1–2 mm) 98.9Jena 60 Cast 101.5Jena 60 Inner burrow (0–1 mm) 100.8Jena 60 Outer burrow (1–2 mm) 96.5Jena 60 Reference soil 92.2
4. Discussion
4.1. The burrow system of anecic earthworms – persistent biogenicstructures
For temperate grasslands 100–800 burrows per m2 have beenreported by Lavelle (1988). Thus, the two sites in this study belongto the upper third of burrow rich sites with maximum burrownumbers of 527 and 660 (Table 1). Higher numbers of burrows inthe deep horizons compared to upper horizons indicate thatburrows had been destroyed at the surface (bioturbation, plough-ing) but persist in the depth (Fig 2). The loamy texture of both sitessupported the stability of the burrows in the subsoil. Ehlers (1975)found earthworm burrow numbers to increase with increasing soildepth from 2 to 60 cm. Particularly in tilled plots, as compared tono-tillage plots, most burrows were found below the ploughinghorizon.
Increasing burrow diameter with depth (Table 2), as found inthe present study, indicates high ages of the burrows compared tosurface soil burrows. While moving up and down inside theburrows, pressure is exerted onto the burrow walls which increasethe burrow diameter. Hagedorn and Bundt (2002) investigated theage of burrows and cracks with radionuclides and found burrows tobe persistent over decades. However, the age of burrows is difficultto determine because abandoned burrows can be reused by otherearthworms (Bastardie et al., 2005). Thus, a burrow can be datedyoung although it already exists for decades up to centuries.
Dry conditions during the summer in Mehrstedt, may haveforced the earthworms to build deeper burrows than those found inJena. Anecic earthworms use their burrows to escape unfavourableclimatic conditions. Anecic earthworms inhabit only one burrowper lifetime, if food scarcity does not force them to move (Edwardsand Bohlen, 1996). Thus, the high proportion of abandoned burrowssuggests that C storage in abandoned burrows plays a major role inthe total C balance of the drilosphere.
In addition to earthworm’s ability to transport C into the soildirectly, burrows as persistent structures may also facilitate other Ctransport processes. Roots use earthworm burrows to access deepsoil horizons, which are difficult to penetrate through the soil ma-trix (Springett and Gray, 1997). Carbon input in burrows with rootbiomass has never been quantified and was restricted in our studiessites to few burrows. Facilitation of plant roots access to soil waterand nutrients by earthworm burrows may increase C sequestration
along with increased productivity, thus increasing the long termsustainability of the ecosystem (Logsdon and Linden, 1992; Partschet al., 2006; Stockdill, 1982). However, missing earthworm freecontrol plots makes it difficult to quantify these long term effects.
C input with seepage water into the earthworm burrows may beanother C transport path. The numerous burrows with missing orblocked connections to the soil surface limit the water infiltration.Only under heavy rain conditions and water saturation of the soilcan the burrows high matrix potential be overcome to allow waterto flow from the soil into the burrow system. Additionally, hydro-phobic organic surfaces in the burrow suppress the waterinfiltration. During 12 summer storms, an average of only 3.9% ofrain was transported through the burrows, with 70% of the burrowsnever conducting any seepage water (Edwards et al., 1990). Thus, Ctransport in burrows with seepage water may play only a minorrole for C translocation compared to active C transport byearthworms.
4.2. Additional C storage within earthworm burrows
The high numbers of earthworm burrows led to a large increasein the soil surface through the burrow surfaces that can be regardedas micro-A horizons with similar features as the surface soil A ho-rizon. Carbon and N gradients in the micro-A horizons, as well asstable isotope values, were comparable to the A horizon of the soilprofiles (Figs. 3, 4 and 6). As in A horizons, there is a more or lesscontinuous C input in the micro-A horizons of the burrows. Theburrow diameter must be kept within a certain range to prevent theearthworm sliding into the burrow. Therefore, earthwormspermanently cast material onto their burrow wall to reduce theburrow diameter. The range of reported C-enrichments in the anecic

En
zym
e activity [p
mo
l M
U m
g-1 D
W so
il sam
ple h
-1]
Cellobiohydrolase (cel)
0
80
160
240
α-Glucosidase (α-gls)800
400
600
0
200
β-Glucosidase (β-gls)
Distance from burrow wall [mm]
0
80
160
240
1.0 2.2 3.4 20 1.0 2.2 3.4 20 1.0 2.2 3.4 20
Xylosidase (xyl)
1.0 2.2 3.4 20 1.0 2.2 3.4 20 1.0 2.2 3.4 20
N-acetylglucosaminidase (Nag)
2400
800
1600
0
Phosphatase (pho)
inhabited burrowsabandoned burrows
Fig. 7. Enzyme activity at 30 cm depth in inhabited (closed circles) and long term abandoned (open circles) anecic earthworm burrows in Jena.
A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–18121810
burrow linings compared to surrounding soil is between 5 and 500%(Hagedorn and Bundt, 2002; Jegou et al., 2000; Stehouwer et al.,1993). We showed that C-enrichment in the drilosphere is tempo-rarily and spatially highly variable within a distance of millimetres(Figs. 3 and 4). Therefore, sample thickness of the burrow linings iscrucial and has to be considered. From our results it is also evidentthat a distinction between inhabited and abandoned burrows isnecessary to assess the C sequestration potential due to, up to threetimes, higher C concentrations in inhabited compared to that foundin abandoned burrows (Fig. 4).
Once ploughing ended in Jena, many earthworm burrows werenewly constructed within 4 years. Anecic earthworms, in particular,are suppressed in cropland systems as a result of continuous tillagedisturbances (Wardle,1995). Conversion from cropland to grasslandled to a fast recovery in the earthworm population. While new
Table 3Extracellular enzyme activities at 30 cm depth in Jena [pmol MU mg�1 DW h�1] in fresh eain the inner burrow wall of inhabited burrows, in burrows abandoned for 11 days (short
Enzyme Enzyme activity [pmol MU mg�1 DW h�1]
Cast Drilosphere Inner burrowwall 0–1 mm
Inhabited Sa
a-gls 1352 290 780 5b-gls 374 71 134 1cel 229 44 74Nag 506 75 154 1pho 263 216 256 2xyl 797 158 230 2
n.s., no significant difference between inhabited and long term abandoned burrows. *Sign
burrows were being constructed in the young Jena grassland, Ctransport into the burrows was fast with a sequestration rate of22 g C m�2 yr�1 for 0–30 cm soil depth. Grasslands after conversionfrom cropland were estimated to sequester about 50 g C m�2 yr�1
(Potter et al., 1999; Soussana et al., 2004). Thus, earthworm activitycould account for 44% of this C accumulation. This is a conservativeestimate which does not include the deeper horizons where wefound the highest C-enrichment of the drilosphere. C-enrichment ofthe burrows may reach a steady state, which is determined by thesize of the earthworm population, the available substrate and themineralisation rate of the deposited C in the burrows. Compared tototal C stocks, the additional C in the anecic drilosphere was smallwith just 1.5% of the total C stock at the Jena site and 2.8% at theMehrstedt site. However, only continuous earthworm activity canmaintain this C-enrichment in the subsoil.
rthworm casts, in the drilosphere (average 0–3 mm distance from burrow wall) andterm abandoned) and in long term abandoned burrows
Enzyme activity/C
hort termbandoned
Long termabandoned
Inhabited Long termabandoned
27 262 54 44n.s.
21 61 313 191*71 38 30 28n.s.
36 66 62 48n.s.
42 226 103 164*71 135 92 99n.s.
ificant difference (p< 0.05) between inhabited and long term abandoned burrows.
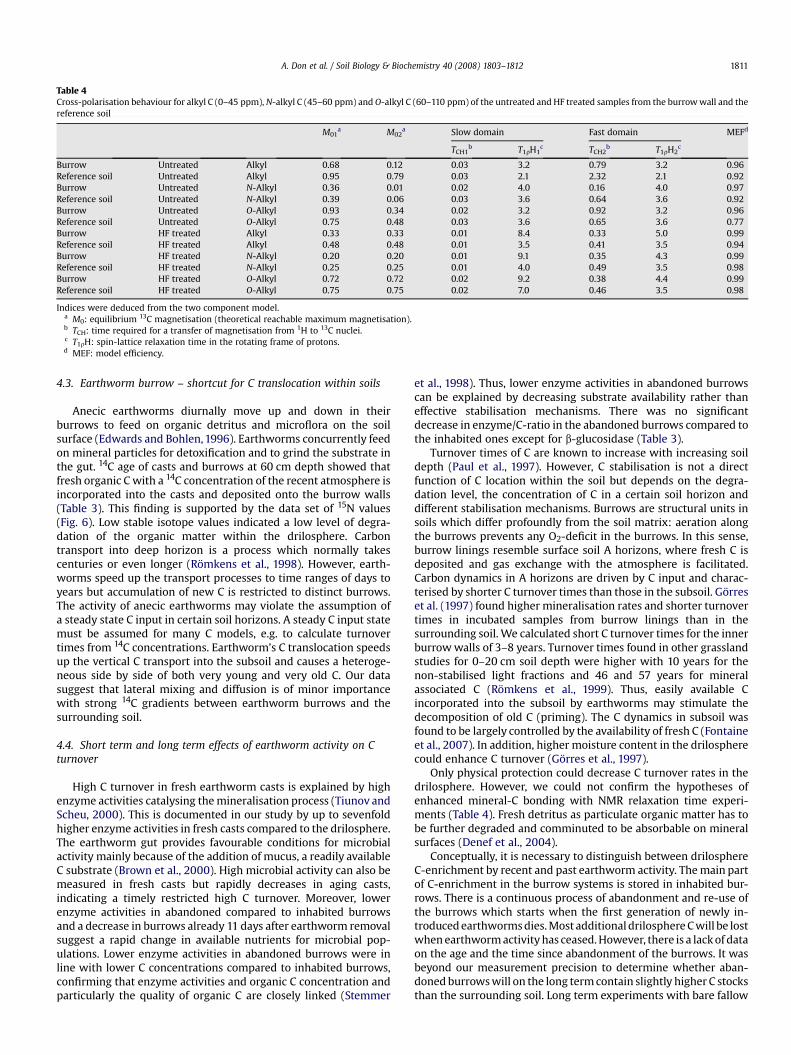
Table 4Cross-polarisation behaviour for alkyl C (0–45 ppm), N-alkyl C (45–60 ppm) and O-alkyl C (60–110 ppm) of the untreated and HF treated samples from the burrow wall and thereference soil
M01a M02
a Slow domain Fast domain MEFd
TCH1b T1rH1
c TCH2b T1rH2
c
Burrow Untreated Alkyl 0.68 0.12 0.03 3.2 0.79 3.2 0.96Reference soil Untreated Alkyl 0.95 0.79 0.03 2.1 2.32 2.1 0.92Burrow Untreated N-Alkyl 0.36 0.01 0.02 4.0 0.16 4.0 0.97Reference soil Untreated N-Alkyl 0.39 0.06 0.03 3.6 0.64 3.6 0.92Burrow Untreated O-Alkyl 0.93 0.34 0.02 3.2 0.92 3.2 0.96Reference soil Untreated O-Alkyl 0.75 0.48 0.03 3.6 0.65 3.6 0.77Burrow HF treated Alkyl 0.33 0.33 0.01 8.4 0.33 5.0 0.99Reference soil HF treated Alkyl 0.48 0.48 0.01 3.5 0.41 3.5 0.94Burrow HF treated N-Alkyl 0.20 0.20 0.01 9.1 0.35 4.3 0.99Reference soil HF treated N-Alkyl 0.25 0.25 0.01 4.0 0.49 3.5 0.98Burrow HF treated O-Alkyl 0.72 0.72 0.02 9.2 0.38 4.4 0.99Reference soil HF treated O-Alkyl 0.75 0.75 0.02 7.0 0.46 3.5 0.98
Indices were deduced from the two component model.a M0: equilibrium 13C magnetisation (theoretical reachable maximum magnetisation).b TCH: time required for a transfer of magnetisation from 1H to 13C nuclei.c T1rH: spin-lattice relaxation time in the rotating frame of protons.d MEF: model efficiency.
A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–1812 1811
4.3. Earthworm burrow – shortcut for C translocation within soils
Anecic earthworms diurnally move up and down in theirburrows to feed on organic detritus and microflora on the soilsurface (Edwards and Bohlen, 1996). Earthworms concurrently feedon mineral particles for detoxification and to grind the substrate inthe gut. 14C age of casts and burrows at 60 cm depth showed thatfresh organic C with a 14C concentration of the recent atmosphere isincorporated into the casts and deposited onto the burrow walls(Table 3). This finding is supported by the data set of 15N values(Fig. 6). Low stable isotope values indicated a low level of degra-dation of the organic matter within the drilosphere. Carbontransport into deep horizon is a process which normally takescenturies or even longer (Romkens et al., 1998). However, earth-worms speed up the transport processes to time ranges of days toyears but accumulation of new C is restricted to distinct burrows.The activity of anecic earthworms may violate the assumption ofa steady state C input in certain soil horizons. A steady C input statemust be assumed for many C models, e.g. to calculate turnovertimes from 14C concentrations. Earthworm’s C translocation speedsup the vertical C transport into the subsoil and causes a heteroge-neous side by side of both very young and very old C. Our datasuggest that lateral mixing and diffusion is of minor importancewith strong 14C gradients between earthworm burrows and thesurrounding soil.
4.4. Short term and long term effects of earthworm activity on Cturnover
High C turnover in fresh earthworm casts is explained by highenzyme activities catalysing the mineralisation process (Tiunov andScheu, 2000). This is documented in our study by up to sevenfoldhigher enzyme activities in fresh casts compared to the drilosphere.The earthworm gut provides favourable conditions for microbialactivity mainly because of the addition of mucus, a readily availableC substrate (Brown et al., 2000). High microbial activity can also bemeasured in fresh casts but rapidly decreases in aging casts,indicating a timely restricted high C turnover. Moreover, lowerenzyme activities in abandoned compared to inhabited burrowsand a decrease in burrows already 11 days after earthworm removalsuggest a rapid change in available nutrients for microbial pop-ulations. Lower enzyme activities in abandoned burrows were inline with lower C concentrations compared to inhabited burrows,confirming that enzyme activities and organic C concentration andparticularly the quality of organic C are closely linked (Stemmer
et al., 1998). Thus, lower enzyme activities in abandoned burrowscan be explained by decreasing substrate availability rather thaneffective stabilisation mechanisms. There was no significantdecrease in enzyme/C-ratio in the abandoned burrows compared tothe inhabited ones except for b-glucosidase (Table 3).
Turnover times of C are known to increase with increasing soildepth (Paul et al., 1997). However, C stabilisation is not a directfunction of C location within the soil but depends on the degra-dation level, the concentration of C in a certain soil horizon anddifferent stabilisation mechanisms. Burrows are structural units insoils which differ profoundly from the soil matrix: aeration alongthe burrows prevents any O2-deficit in the burrows. In this sense,burrow linings resemble surface soil A horizons, where fresh C isdeposited and gas exchange with the atmosphere is facilitated.Carbon dynamics in A horizons are driven by C input and charac-terised by shorter C turnover times than those in the subsoil. Gorreset al. (1997) found higher mineralisation rates and shorter turnovertimes in incubated samples from burrow linings than in thesurrounding soil. We calculated short C turnover times for the innerburrow walls of 3–8 years. Turnover times found in other grasslandstudies for 0–20 cm soil depth were higher with 10 years for thenon-stabilised light fractions and 46 and 57 years for mineralassociated C (Romkens et al., 1999). Thus, easily available Cincorporated into the subsoil by earthworms may stimulate thedecomposition of old C (priming). The C dynamics in subsoil wasfound to be largely controlled by the availability of fresh C (Fontaineet al., 2007). In addition, higher moisture content in the drilospherecould enhance C turnover (Gorres et al., 1997).
Only physical protection could decrease C turnover rates in thedrilosphere. However, we could not confirm the hypotheses ofenhanced mineral-C bonding with NMR relaxation time experi-ments (Table 4). Fresh detritus as particulate organic matter has tobe further degraded and comminuted to be absorbable on mineralsurfaces (Denef et al., 2004).
Conceptually, it is necessary to distinguish between drilosphereC-enrichment by recent and past earthworm activity. The main partof C-enrichment in the burrow systems is stored in inhabited bur-rows. There is a continuous process of abandonment and re-use ofthe burrows which starts when the first generation of newly in-troduced earthworms dies. Most additional drilosphere C will be lostwhen earthworm activity has ceased. However, there is a lack of dataon the age and the time since abandonment of the burrows. It wasbeyond our measurement precision to determine whether aban-doned burrows will on the long term contain slightly higher C stocksthan the surrounding soil. Long term experiments with bare fallow

A. Don et al. / Soil Biology & Biochemistry 40 (2008) 1803–18121812
showed that without C input from plants over many decades, soil Cstocks will decrease to a minimum C concentration which dependson the clay and silt content (Ruehlmann, 1999). We found no evi-dence that C-enrichment in the drilosphere would persist long afterthe burrows were abandoned. Thus, earthworm burrows are an in-put driven system where C concentrations may drop to the samebase level as determined in long term bare fallow experiments.
5. Conclusions
From our results, we conclude that the C distribution in soils ischanged by anecic earthworm’s activity with more C stored in thesubsoil where earthworms slightly increase the C stocks. The un-derlying mechanisms are based on enlarged surfaces throughwhich C enters the soil and can be stored (burrow linings). Up tonow, we found no indication to suggest that C in burrow linings hasa lower turnover rate and is stabilised on long term. The hypothesisthat C stabilisation is enhanced as a result of absorption on mineralsurfaces could not be confirmed. However, the translocation of Cfrom organic layer to the subsoil will decrease the C vulnerability tomineralisation. Carbon in the organic layer and the surface soil ismuch more prone to disturbances with rapid C loss than subsoil C.
Acknowledgements
We thank Heike Knicker for conducting the NMR relaxation timemeasurements and Ingrid Kogel-Knabner for her support with thisexperiment. We thank Kai Uwe Totsche for his help on analysing theNMR experiment. For help with the specification of the earthwormswe thank Doro Schneider. For fruitful discussions we thank StefanScheu, Martin J. Shipitalo, Paul F. Hendrix, Otto Daniel and PatrickLavelle. For helpful comments to the manuscript we thank AnnetteFreibauer, Kim Milferstedt and two anonymous reviewers.
References
Abelmann, K., Totsche, K.U., Knicker, H., Kogel-Knabner, I., 2004. CP dynamics ofheterogeneous organic material: characterization of molecular domains incoals. Solid State Nuclear Magnetic Resonance 25, 252–266.
Alban, D.H., Berry, E.C., 1994. Effects of earthworm invasion on morphology, carbon,and nitrogen of a forest soil. Applied Soil Ecology 1, 243–249.
Bastardie, F., Capowiez, Y., Renault, P., Cluzeau, D., 2005. A radio-labelled study ofearthworm behaviour in artificial soil cores in term of ecological types. Biologyand Fertility of Soils 41, 320–327.
Bossuyt, H., Six, J., Hendrix, P.F., 2005. Protection of soil carbon by microaggregateswithin earthworm casts. Soil Biology and Biochemistry 37, 251–258.
Bouche, M.B., 1977. Strategies lombriciennes. In: Lohm, U., Persson, T. (Eds.), SoilOrganisms as Components of Ecosystems, vol. 25, pp. 122–132. Stockholm.
Brown, G.G., Barois, I., Lavelle, P., 2000. Regulation of soil organic matter dynamicsand microbial activity in the drilosphere and the role of interactions with otheredaphic functional domains. European Journal of Soil Biology 36, 177–198.
Denef, K., Six, J., Merckx, R., Paustian, K., 2004. Carbon sequestration in micro-aggregates of no-tillage soils with different clay mineralogy. Soil Science Societyof America Journal 68, 1935–1944.
Don, A., Schumacher, J., Scherer-Lorenzen, M., Scholten, T., Schulze, E.-D., 2007.Spatial and vertical variation of soil carbon at two grassland sites – implicationsfor measuring soil carbon stocks. Geoderma 141, 272–282.
Edwards, C.A., Bohlen, P.J., 1996. Biology and Ecology of Earthworms, third ed.Springer, Berlin.
Edwards, W.M., Shipitalo, M.J., Owens, L.B., Norton, L.D., 1990. Effect of Lumbricusterrestris L. burrows on hydrology of continuous no-till corn fields. Geoderma46, 73–84.
Ehlers, W., 1975. Observations on earthworm channels and infiltration on tilled anduntilled loess soil. Soil Science 119, 242–249.
Fontaine, S., Barot, S., Barre, P., Bdioui, N., Mary, B., Rumpel, C., 2007. Stability oforganic carbon in deep soil layers controlled by fresh carbon supply. Nature450, 277–280.
Ge, F., Shuster, W.D., Edwards, C.A., Parmelee, R.W., Subler, S., 2001. Water stabilityof earthworm casts in manure- and inorganic-fertilizer amended agro-ecosystems influenced by age and depth. Pedobiologia 45, 12–26.
Gorres, J.H., Savin, M.C., Amador, J.A., 1997. Dynamics of carbon and nitrogenmineralization, microbial biomass, and nematode abundance within and out-side the burrow walls of anecic earthworms (Lumbricus terrestris). Soil Science162, 666–671.
Guggenberger, G., Thomas, R.J., Zech, W., 1996. Soil organic matter within earth-worm casts of an anecic–endogeic tropical pasture community, Colombia.Applied Soil Ecology 3, 263–274.
Hagedorn, F., Bundt, M., 2002. The age of preferential flow paths. Geoderma 108,119–132.
Jegou, D., Cluzeau, D., Hallaire, V., Balesdent, J., Trehen, P., 2000. Burrowing activityof the earthworms Lumbricus terrestris and Aporrectodea giardi and conse-quences on C transfers in soil. European Journal of Soil Biology 36, 27–34.
Kaiser, K., Zech, W., 1999. Release of natural organic matter sorbed to oxides anda subsoil. Soil Science Society of America Journal 63, 1157–1166.
Keudel, M., Schrader, S., 1999. Axial and radial pressure exerted by earthworms ofdifferent ecological groups. Biology and Fertility of Soils 29, 262–269.
Lavelle, P., 1988. Earthworm activities and the soil system. Biology and Fertility ofSoils 6, 237–251.
Lavelle, P., Charpentier, F., Villenave, C., Rossi, J., Derouard, L., Pashanasi, B., Andre, J.,Ponge, J., Bernier, N., 2004. Effects of earthworms on soil organic matter andnutrient dynamics at a landscape scale over decades. In: Edwards, C.A. (Ed.),Earthworm Ecology. CRC Press, Boca Raton, pp. 145–160. second ed.
Lee, K.E., 1985. Earthworms – Their Ecology and Relationship with Soils and LandUse. Academic Press, Sydney.
Logsdon, S.D., Linden, D.R., 1992. Interactions of earthworms with soil physicalconditions influencing plant-growth. Soil Science 154, 330–337.
von Lutzow, M., Kogel-Knabner, I., Ekschmitt, K., Matzner, E., Guggenberger, G.,Marschner, B., Flessa, H., 2006. Stabilization of organic matter in temperatesoils: mechanisms and their relevance under different soil conditions – a re-view. European Journal of Soil Science 57, 426–445.
Minagawa, M., Wada, E., 1984. Stepwise enrichment of N-15 along food-chains – further evidence and the relation between Delta-N-15 and animal age.Geochimica et Cosmochimica Acta 48, 1135–1140.
Partsch, S., Milcu, A., Scheu, S., 2006. Decomposers (Lumbricidae, Collembola) affectplant performance in model grasslands of different diversity. Ecology 87, 2548–2558.
Paul, E.A., Clark, F.E., 1996. Soil Microbiology and Biochemistry. Academic Press, SanDiego.
Paul, E.A., Follett, R.F., Leavitt, S.W., Halvorson, A., Peterson, G.A., Lyon, D.J., 1997.Radiocarbon dating for determination of soil organic matter pool sizes anddynamics. Soil Science Society of America Journal 61, 1058–1067.
Potter, K.N., Torbert, H.A., Johnson, H.B., Tischler, C.R., 1999. Carbon storage afterlong-term grass establishment on degraded soils. Soil Science 164, 718–725.
Pritsch, K., Luedemann, G., Matyssek, R., Hartmann, A., Schloter, M., Scherb, H.,Grams, T.E.E., 2005. Mycorrhizosphere responsiveness to atmospheric ozoneand inoculation with Phytophthora citricola in a phytotron experiment withspruce/beech mixed cultures. Plant Biology 7, 718–727.
Raw, F., 1962. Studies of earthworm populations in orchards. I. Leaf burial in appleorchards. Annals of Applied Biology 50, 389–404.
Romkens, P., Hassink, J., Van der Plicht, J., 1998. Soil organic C-14 dynamics: effectsof pasture installation on arable land. Radiocarbon 40, 1023–1031.
Romkens, P., van der Plicht, J., Hassink, J., 1999. Soil organic matter dynamics after theconversion of arable land to pasture. Biology and Fertility of Soils 28, 277–284.
Roscher, C., Schumacher, J., Baade, J., Wilcke, W., Gleixner, G., Weisser, W.W.,Schmid, B., Schulze, E.D., 2004. The role of biodiversity for element cycling andtrophic interactions: an experimental approach in a grassland community. Basicand Applied Ecology 5, 107–121.
Ruehlmann, J.,1999. A new approach to estimating the pool of stable organic matter insoil using data from long-term field experiments. Plant and Soil 213, 149–160.
Scherer-Lorenzen, M., Schulze, E.-D., Don, A., Schumacher, J., Weller, E., 2007.Exploring the functional significance of forest diversity: a new long-termexperiment with temperate tree species (BIOTREE). Perspectives in PlantEcology, Evolution and Systematics 9, 53–70.
Schoning, I., Knicker, H., Kogel-Knabner, I., 2005. Intimate association between O/N-alkyl carbon and iron oxides in clay fractions of forest soils. Organic Geo-chemistry 36, 1378–1390.
Sims, R.W., Gerard, B.M., 1985. Earthworms. Brill & Backhuys, London.Soussana, J.F., Loiseau, P., Vuichard, N., Ceschia, E., Balesdent, J., Chevallier, T.,
Arrouays, D., 2004. Carbon cycling and sequestration opportunities in tem-perate grasslands. Soil Use and Management 20, 219–230.
Springett, J., Gray, R., 1997. The interaction between plant roots and earthwormburrows in pasture. Soil Biology and Biochemistry 29, 621–625.
Stehouwer, R.C., Dick, W.A., Traina, S.J., 1993. Characteristics of earthworm burrowlining affecting atrazine sorption. Journal of Environmental Quality 22, 181–185.
Steinhof, A., Adamiec, G., Gleixner, G., van Klinken, G.J., Wagner, T., 2004. The newC-14 analysis laboratory in Jena, Germany. Radiocarbon 46, 51–58.
Stemmer, M., Gerzabek, M.H., Kandeler, E., 1998. Organic matter and enzymeactivity in particle-size fractions of soils obtained after low-energy sonication.Soil Biology and Biochemistry 30, 9–17.
Stockdill, S.M.J., 1982. Effects of introduced earthworms on the productivity of NewZealand pastures. Pedobiologia 24, 29–35.
Tiunov, A.V., Scheu, S., 2000. Microbial biomass, biovolume and respiration inLumbricus terrestris L. cast material of different age. Soil Biology andBiochemistry 32, 265–275.
Wardle, D.A., 1995. Impacts of disturbance on detritus food webs in agro-ecosys-tems of contrasting tillage and weed management practices. In: Begon, M.,Fitter, A.H. (Eds.), Advances in Ecological Research, vol. 26. Academic Press,London, pp. 105–185.
Wolters, V., 2000. Invertebrate control of soil organic matter stability. Biology andFertility of Soils 31, 1–19.
Related Documents











