http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8 115 Ordovician nautiloid faunas of Central and Southern Thailand B. A. STAIT & C. F. BURRETT Geology Department, University of Tasmania, G.P.O. Box 252 C, Hobart, Tasmania 7001, Australia (Received 17 June 1983; accepted 19 August 1983) Abstract - Nautiloids from the shallow water Ordovician carbonates of Central and Southern Thailand can be grouped intofivebroad assemblages. 1. The Middle Ibexian fauna of indeterminate endocerids from Tarutao Island. 2. Hardmanoceras chrysanthimum (Kobayashi) of the Upper Ibexian age strata on Tarutao Island. 3. The Upper Ibexian Manchuroceras nakamense sp.nov. from Ron Phibum, Southern Thailand. 4. Wutinoceras sp., Chaohuceras ? sp. of Lower to Middle Whiterockian age strata from Satun Province. 5. Wutinoceras sp., Armenoceras chediforme Kobayashi and Georgina sp. of Lower-Middle Whiterockian age strata from Kanchanaburi, Central Thailand. All of these genera occur in Australia and China. 1. Introduction In the course of a general survey of the Ordovician of Thailand more than 70 nautiloid specimens were collected from several localities (Figs 1, 2). All specimens are from the carbonates of the Thung Song Formation (in southern Thailand) and Tha Manao Formation (in central Thailand), both of which conformably overlie the fossiliferous siliciclastics of the Tarutao Formation (on Tarutao Island) and its un- fossiliferous correlates elsewhere. These Upper Cam- brian and Ordovician formations occur in a narrow (150 km) north-south trending belt extending from the Shan States of Burma, through Thailand to the south of Kuala Lumpur in Malaysia. A recent, authoritative summary of the geology of western Thailand can be found in Bunopas (1982). Deformation of the Ordovician carbonates ranged from moderate to intense, and thermal metamorphism, based on the conodont colour alteration index of Epstein, Epstein & Harris (1977), ranged from 250-300 °C in southern Thailand to 400 °C in parts of central Thailand. This deformation and metamorphism has led to poor preservation of the conodonts and in most cases prohibits specific and sometimes the generic determinations of the nautiloids. 2. Previous work There has been very little published on the Ordovician nautiloids from Thailand. The vast majority of the palaeontological work was done by Kobayashi (1958, 1959, 1961) and Kobayashi & Hamada (1964), with the only other paper which includes descriptions and illustrations of Ordovician nautiloids being Ingavat, Muenlek & Udomratn (1975). Table 1 contains a list of the papers in which nautiloids are mentioned or described and the assignments made therein. Where a reassignment is considered possible Geol. Mag. 121 (2), 1984, pp. 115-124. Printed in Great Britain. t r \ J | " \ \ \j f ! M s v KANCHANABURI v PROVINCE \ 1 i / J 1 i 4 } \ i TARUTAO ISLAND ""~~ LANGKAWf ISLANDS y- t <> •l I < I \ f \ I AA pi ^ ^ r 1 \ . / ^ ^ y BANGKOK /* n r i ^v ? / ^ ^ ^| ORDOVICIAN LIMESTONE ' INTERNATIONAL BOttOEH 0 100 KM Q C? \ .THUNG SONG y /RON PHIBUU K c\ •- V / S A T U N PROVINCE - ^ ^ - \ \ r - v MALAYSIA ^/" S ~'' ~s \ \ s s \ -y U | J i >v •r Figure 1. Map indicating the distribution of Ordovician strata in Thailand and the collecting localities (A-D are shown in detail on Fig. 2). (Modified from Bunopas, 1982.) 8-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
115
Ordovician nautiloid faunas of Central andSouthern Thailand
B. A. STAIT & C. F. B U R R E T T
Geology Department, University of Tasmania, G.P.O. Box 252 C, Hobart, Tasmania 7001, Australia
(Received 17 June 1983; accepted 19 August 1983)
Abstract - Nautiloids from the shallow water Ordovician carbonates of Central and SouthernThailand can be grouped into five broad assemblages. 1. The Middle Ibexian fauna of indeterminateendocerids from Tarutao Island. 2. Hardmanoceras chrysanthimum (Kobayashi) of the Upper Ibexianage strata on Tarutao Island. 3. The Upper Ibexian Manchuroceras nakamense sp.nov. from RonPhibum, Southern Thailand. 4. Wutinoceras sp., Chaohuceras ? sp. of Lower to Middle Whiterockianage strata from Satun Province. 5. Wutinoceras sp., Armenoceras chediforme Kobayashi and Georginasp. of Lower-Middle Whiterockian age strata from Kanchanaburi, Central Thailand. All of thesegenera occur in Australia and China.
1. Introduction
In the course of a general survey of the Ordovician ofThailand more than 70 nautiloid specimens werecollected from several localities (Figs 1, 2). Allspecimens are from the carbonates of the Thung SongFormation (in southern Thailand) and Tha ManaoFormation (in central Thailand), both of whichconformably overlie the fossiliferous siliciclastics ofthe Tarutao Formation (on Tarutao Island) and its un-fossiliferous correlates elsewhere. These Upper Cam-brian and Ordovician formations occur in a narrow(150 km) north-south trending belt extending from theShan States of Burma, through Thailand to the southof Kuala Lumpur in Malaysia. A recent, authoritativesummary of the geology of western Thailand can befound in Bunopas (1982).
Deformation of the Ordovician carbonates rangedfrom moderate to intense, and thermal metamorphism,based on the conodont colour alteration index ofEpstein, Epstein & Harris (1977), ranged from250-300 °C in southern Thailand to 400 °C in parts ofcentral Thailand. This deformation and metamorphismhas led to poor preservation of the conodonts and inmost cases prohibits specific and sometimes thegeneric determinations of the nautiloids.
2. Previous work
There has been very little published on the Ordoviciannautiloids from Thailand. The vast majority of thepalaeontological work was done by Kobayashi(1958, 1959, 1961) and Kobayashi & Hamada (1964),with the only other paper which includes descriptionsand illustrations of Ordovician nautiloids beingIngavat, Muenlek & Udomratn (1975). Table 1contains a list of the papers in which nautiloids arementioned or described and the assignments madetherein. Where a reassignment is considered possible
Geol. Mag. 121 (2), 1984, pp. 115-124. Printed in Great Britain.
t r \J | " \
\ \j
f
!Ms v
KANCHANABURI vPROVINCE \
1
i/
J
1 i4} \i
TARUTAOISLAND ""~~
LANGKAWfISLANDS
y-
t
<>•lI
< I\f
\I
AA
pi
^ ^ r1
\ . / ^
^ y
BANGKOK /*
n ri ^v ?
/ ^ ^
^ | ORDOVICIAN LIMESTONE
' INTERNATIONAL BOttOEH
0 100 KM
Q
C?\ .THUNG SONG
y /RON PHIBUU
Kc\
•- V /SATUN PROVINCE
- ^ ^ - \ \
r - vMALAYSIA ^/"S~''
~s\\s
s\
-yU
|J
i> v
•r
Figure 1. Map indicating the distribution of Ordovicianstrata in Thailand and the collecting localities (A-D areshown in detail on Fig. 2). (Modified from Bunopas, 1982.)
8-2

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
116 B. A. STAIT& C. F. BURRETT
AO LA NGA
MALAKA CREEKl
Figure 2. Maps showing the precise nautiloid-collecting localities. A-D on the maps refer to areas on Fig. 1.

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
Ordovician nautiloids from Thailand
Table 1.
117
Brown et at. (1951)Kobayashi (1958)Kobayashi (1959)Kobayashi (1961)
Kobayashi & Hamada (1964)
Collins in Burton (1974)
Ingavat, Muenlek & Udomratn (1975)
Teraoka el al. (1982)
Actinoceras sp.Armenoceras chediformeArmenoceras chediformeArmenoceras cfr. chediforme(Armenoceras chediforme)
Ormoceras sp.(Wutinoceras sp.)
Endocerid siphuncleOrmocerid gen. et sp. indet.Armenoceras sp.A. coranicumSinoceras chinenseMichelinoceras sp.Endoceroid siphuncleSelkirkoceras aff. yokusenense(Armenoceras chediforme)
Eothinoceras sp.Orthoceras sp.Endoceroid
Southern ThailandSouthern ThailandSouthern ThailandNorthern Thailand
Northern Thailand
Southern ThailandSouthern ThailandSouthern ThailandSouthern ThailandSouthern ThailandSouthern ThailandSouthern ThailandCentral Thailand
Central ThailandCentral ThailandSouthern Thailand
or necessary it is given in parentheses. Review paperswhich only list previously described faunas are notincluded.
3. Age and correlations
International correlations and terminologies are basedon the chart of Ross et al. (1982).
The oldest nautiloids collected (Endoceratidae gen.et sp. indet.) are common within the two lowestmembers of the Thung Song Formation on TarutaoIsland (Fig. 2 A, 3). The underlying Tarutao Formationcontains a Tremadoc trilobite fauna (Stait, Burrett &Wongwanich, 1983). The lowest members of theThung Song Formation contain conodonts whichindicate a Middle Ibexian age (Teraoka et al. 1982)(Fig. 3).
Hardmanoceras is found in the penultimate memberof the Thung Song Formation on Tarutao Island (ofnodular, red and green calcareous shales) and is foundwith conodonts of the Prioniodus evae zone ofprobable Middle Arenig (Upper Ibexian-LowerWhiterockian) age. Hardmanoceras also occurs in theMiddle Arenig Emanuel Formation of WesternAustralia (Teichert & Glenister, 1954; McTavish,1973) and in the probably Middle Arenig of NorthChina (Chen, 1976).
Wutinoceras occurs with Chaohucerasl and stro-matoporoids in peritidal dolomicrites at Khao Nui inSatun Province (Fig. 2B, 3). Conodonts from thishorizon include Bergstroemognathus sp., Scolopodusrex and Eoneoprioniodus bilongatus. The last-namedspecies is found in the Joins Formation of Oklahoma(Mound, 1965) and the Ship Point Formation ofCanada (Barnes, 1977) and a Middle Whiterockianage is suggested (Ross et al. 1982). Wutinoceras andChaohuceras both occur in the Lower-Middle White-rockian of Tasmania (Stait, unpubl. Ph.D. thesis,Univ. Tasmania, 1981) and North China (Chen, 1976;Qi, 1980). Wutinoceras is also widespread in Lower-Middle Whiterockian strata of North America(Flower, 1957, 1968, 1976), Australia (Flower, 1968;
Stait, unpubl. Ph.D. thesis, Univ. Tasmania, 1981) andMalaysia (Stait & Burrett, 1982).
Dolomicrites in a quarry at Ron Phibum, nearThung Song (Fig. 2D) yield Manchuroceras and thegastropod Teiichispira. No specifically determinableconodonts have yet been found, but Manchuroceras istypical of the Upper Ibexian (formerly UpperCanadian) in North America (Hook & Flower, 1977)and correlates in China (Chen, 1976) and Australia(Teichert, 1947).
Nautiloid specimens from the Tha Manao Formationof Khao Tham in Kanchanaburi Province (Fig. 2 D)include Wutinoceras, Armenoceras and Georgina,which together suggest a Lower-Middle Whiterockianage.
4. Systematic palaeontology
Order ACTINOCERIDA Teichert, 1933Family ARMENOCERATIDAE Troedsson, 1926
Armenoceras Foerste, 1924Armenoceras chediforme Kobayashi, 1958
1958 Armenoceras chediforme Kobayashi; p. 229, pi.17, fig. 2a-b.1959 Armenoceras chediforme Kobayashi; Kobayashi,p. 401, pi. 27, figs 6-8.1961 Armenoceras cfr. chediforme Kobayashi; Koba-yashi, p. 82, pi. V, figs 1-3.
Materials. Six specimens from the Tha ManaoFormation of Kanchanaburi northern Thailand(locality D on Figs 1 and 2), TF1577-1582.
Remarks. As the Thai specimens have a siphunclemuch higher than long, recumbent septal necks, strongadnation of the connecting rings to both sides of thesepta and straight simple radial canals they are clearlyassignable to the broad concept of Armenoceras.Kobayashi (1958) described a species of Armenocerasfrom Thung Song Limestone at Ron Phibun nearThung Song, of which he remarked t h a t ' . . . this formis here recognized as an Armenoceras having a large,

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
118 B. A. STAIT& C. F. BURRETT
TARUTAOISLAND
SATUNPROVINCE
RONPHIBUM
KANCHANABURIPROVINCE
oc>
ozUJoc
o
sUJoc
A o n F i g . 1
I , I
i I I I Ii ^ r ^\\k I ^
/ B \ I >W\I . I I I/ // /
II II I/ /T i l l
B on Fig. C on F i g . 1 0 on F i g . 1
ION
i—
socou.
(Sao«/)(92=>XK
/
1|/1 ,/
111 ;/111|/
/1 1 1 1/
1 1 1 |/
1 1 1 1
1 1 1 1
1 1 1 1
1 1 1 1
I I I !
1 1 1 1
Mil
1 1 1 1/
/ /II II I
T l I I I
II II I
I . I I . I I
I I I T I/ /
1 1 1
KEY
LIMESTONE
DOLOMICRITE
STROMATOLITICLIMESTONE
NODULAR LIMESTONE
QUARTZ ARENITES
HARDMANOCERAS
ORTHOCONICNAUTILOIDS
Figure 3. Generalized stratigraphy and correlations of the Ordovician of southern and central Thailand.

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
Ordovician nautiloids from Thailand 119
Figure 4. Armenoceras chediforme Kobayashi. A. Naturallyweathered silicified siphuncle, TF1577, x 2. B. Enlargementof a siphuncular segment in TF1581 (note the radial canal),x 3. C. Thin section of four siphuncular segments, TF 1581,x 1.5. D. Naturally weathered silicified siphuncle, TF1578,x 2. E. Thin section of isolated siphuncle, TF1580, x 1.5.All specimens oriented with apex towards the bottom of thepage.
somewhat curved siphuncle near the center of theconch'. The type material of this species is in highlydeformed limestone, a fact even conceded byKobayashi (1959, p. 402), where he stated: 'Becausethe outline of the siphuncle and the curvature of thesepta are irregular in the type specimen [of A.chediforme] it is highly probable that its siphuncle wascurved by secondary deformation.'
The type material is only poorly preserved but it isconsidered better to retain this species of Armenocerasfrom Thailand and Malaysia rather than to try andcompare this inadequate material with other species,although when topotype material is available detailedcomparison may be possible. Although the material
in the present collections is only poorly preserved nofeature can be found that differentiates them from A.chediforme.
Ingavat, Muenlek and Udomratn (1975) describedSelkirkoceras aff. yokusenense from north of Kan-chanaburi at Amphoe Ban Rai. They assigned it toSelkirkoceras because of the apparent reduction ofwidth of the siphuncle with growth; however,examination of the drawing and photograph plusmaterial in the present collections suggest that it is anoblique section through a specimen of Armenoceras.
Family GEORGINIDAE Wade, 1977Georgina Wade, 1977
Georgina sp.Fig. 5D, E
Material. Two incomplete, isolated siphunclesTF 1570, TF 1571. Both specimens from Kanchanaburi(locality D on Figs 1, 2).
Description. Specimen TF1570 consists of five si-phuncular segments totalling 85 mm in length. Thefirst segment (adorally) is 28 mm wide while the fifthis 25 mm. The siphuncular segments are twice as wideas long, and are highly expanded between the septalforamina. The septal necks are cyrtochoanitic, broadlyopen, and slope apically from the venter to the dorsumat 70°. The engrafts are large and regularly spaced,12 mm long and 7.0 mm wide on the first segment.
Remarks. The presence of engrafts and the size andshape of the siphuncle are typical of the Georginidaeand in particular the genus Georgina. As only twoisolated incomplete siphuncles have been foundassignment to a species cannot be justified; however,comparison with the other known species is possible.Georgina has only previously been described from theLower to Middle Ordovician Coolibah and NoraFormations of Western Queensland (Wade, 1977). TheQueensland fauna contains five species of Georgina.Georgina sp. differs from G. taylori, G. andersonorum,G. beuteli and G. linda as the siphuncular segments aremuch longer relative to their height. G. dwyeri has asimilar length-to-height ratio for the siphuncularsegments as does Georgina sp. but the engrafts aremore numerous, smaller and narrower in G. dwyeri. Insummary, Georgina sp. has engrafts more typical of thespecies found in the upper Coolibah Formation(especially G. taylori) but the proportions of thesiphuncle are those of the younger species G. dwyeri(which occurs in the upper Nora Formation).
Family WUTINOCERATIDAE Shimizu & Obata, 1935Wutinoceras Shimizu & Obata, 1935
Wutinoceras sp.
Fig. 5A-C
1961 Ormoceras sp. Kobayashi, p. 83, pi. v, figs 4, 5.
Material. Two poorly preserved phragmocones;

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
120 B. A. STAIT& C. F. BURRETT
Figure 5. A-C. Wutinoceras sp. A, C oriented apex below;B oriented apex right. A. Naturally weathered phragmocone,lateral view, TF1561, x 1.8. B. Enlargement of the siphunclein TF 1561, x 3.5. C. Oblique opaque section of a partialphragmocone, TF1562, x 2. D-E. Georgina sp. D.Naturally weathered silicified siphuncle, venter right, apexbelow, TF1570, x 1.5. E. Oblique opaque section of thesiphuncle, apex left, TF 1570, x 2.
TF1561 from Kanchanaburi, Central Thailand (lo-cality D on Figs 1, 2) and TF1562 from SouthernThailand (locality B on Figs 1, 2).
Remarks. Only two specimens that could be includedin the genus Wutinoceras have been found in this study,which considering the abundance of specimens ofWutinoceras robustum in Northern Malaysia (Stait &Burrett, 1982) is surprising. Both of the specimens areweathered down to the siphuncle and the criticaldetails required for assignment to a species are notclearly preserved. However, the size and position of thesiphuncle and the few details of the interior of the
Figure 6. Manchuroceras nakamense sp.nov. A-E. A seriesof transverse sections of TF 1558, all x 2. A. Apicad facingcut. B. Orad facing cut 13 mm from A. C. Apicad facing cut.D. Apicad facing cut 12 mm from C. E. Orad facing cut10 mm from D. F. Sagittal thin section of TF 1558, x 2. G.Enlargement of ventral side of thin section in F, x 8.
siphuncle that can be observed are reminiscent ofWutinoceras robustum, to which they may be shown tobelong when better material becomes available.
Kobayashi (1961) described and illustrated Ormo-ceras sp. from northern Thailand. This specimen isindistinguishable from the specimens assigned toWutinoceras sp. in this study.
Order ENDOCERIDA Teichert, 1933Family MANCHUROCERATIDAE Kobayashi, 1935
Manchuroceras Ozaki, 1927Manchuroceras nakamense sp.nov.
Fig. 6A-G
Material. Three partially preserved endosiphocones,holotype TF1558, paratypes TF1559, TF1560 (T62,locality C on Figs 1, 2).
Description. The phragmocone is not preserved, onlyisolated endosiphuncles. The apical end of the

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
Ordovician nautiloids from Thailand 121
endosiphocone is not known, however the majority ispreserved. The deformation undergone by the speci-mens has resulted in an apparent increase in the rateof expansion of the endosiphuncle.
The endosiphuncle is circular in cross-section,slowly expanding, and the height increases from 16 to22 mm in 58 mm on TF1558. The endosiphoconeincreases in height from 3.5 to 12.0 mm in 58 mm andthe endosiphowedge decreases in height from 9.0 to7.0 mm in the same distance on TF1558. The septalnecks are not clearly preserved but a thin section ofTF1558 suggests that they were hemichoanitic orsubholochoanitic.
A triradiate endosiphoblade pattern is present withone vertical blade on the dorsal side of theendosiphocone. The other two blades are ventro-lateral, originating at the boundary between theendosiphowedge and the endocones proper.
Remarks. The generic taxobases of Manchurocerashave been discussed in detail since its first description(Kobayashi, 1935, 1977; Hook & Flower, 1977;Teichert, 1947; Teichert & Glenister, 1953). TheThailand specimens fit into the general concept ofManchuroceras as they have a triradiate endosipho-blade pattern, sub-holochoanitic septal necks, ventralelevation of the endocones and an endosiphotube onthe dorsal side of centre. However, the Thai materialis more longiconic and slender than is typical ofManchuroceras. M. asiaticum Balashov is longer andmore slender than M. nakamense, and as suggested byHook & Flower (1977, p. 42) may well be a new genus.M. qiugshuiheense Chen (1976) is also a slender formbut the endosiphotube is closer to the dorsal side ofthe endosiphuncle and the ventral elevation is higherthan in M. nakamense. Zou (1981) established threespecies of Manchuroceras, all relatively slendercompared to typical Manchuroceras, but they arepreserved as silicified isolated endosiphuncles, whichmakes comparison with the Thailand material virtuallyimpossible.
Genus Chaohuceras Qi, 1980Chaohuceras ? sp.
Fig. 7A-D
Material. Only one endosiphuncle is known, TF1572from T2 (locality B on Figs 1, 2).
Description. Only the endosiphuncle is known. It isstraight, circular in cross-section and slowly expanding,with the height increasing from 12 to 28 mm in 60 mmon TF1572. The endosiphoblade pattern changesfrom a pair of lateral blades at the apical end to atriradiate pattern at the adoral end of the specimen.The change in pattern is achieved when the right blade(looking adorally) splits into two which graduallyseparate, producing the triradiate pattern. Theconnecting rings and septal necks are unknown.
Remarks. Qi (1980) established the genus Chaohucerasfor an endocerid with a pair of lateral endosiphoblades.The holotype specimen only reaches a similar size tothe most apicad part of TF 1572. The endosiphobladepattern at the apical end of TF 1572 is similar to thatin the equivalent-sized transverse cuts in Chaohuceras,but more orad the pattern of the endosiphobladeschanges. There are two possible explanations for thischange; either Chaohuceras develops a triradiateendosiphoblade pattern with growth and the holotypeof Chaohuceras is the immature portion of thespecimen, or they are different genera and early stagesof TF1572 are a homeomorph of Chaohuceras. Untilmore material is available for study it is difficult todistinguish which of the two scenarios is correct. SoTF1572 is tentatively assigned to Chaohuceras,awaiting additional specimens.
ENDOCERATIDAE gen. et sp. indet.Fig. 8A-I
Material. Seven isolated endosiphuncles, TF1563-1569. Specimens collected from the ThungSong Formation, Tarutao Island (TF32, 33, 35;locality A on Figs 1, 2).
Description. Endosiphuncle straight, slowly expanding.The endosiphotube is crescentic, concave towardswhat is assumed to be the venter. The tube opens intoa dumb-bell-shaped endosiphocone with the larger'endosiphowedge' on the venter. The endosiphobladepattern is poorly preserved in most specimens butconsists of four blades which project from the lateralcorners of the endosiphocone. The blades are paired,with one pair to the dorsal side and the other at theventral side; both are at 45° to the lateral plane.
Figure 7. Chaohuceras ? sp. A-D. A series of transverse sections of TF 1572, x 2, all apicad facing cuts. B. 5 mm orad of A,C-B 13 mm, D-C 11 mm.

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
122 B. A. STAIT& C. F. BURRETT
Figure 8. Endocerida gen. et sp. indet. A-C. A series oftransverse sections of TF 1563, x 5. A. Apicad facing cut. B.Apicad facing cut 31 mm from A. C. Apicad facing cut25 mm from A. D. Sagittal thin section of endosiphunclebetween A and C, apex below, venter right, x 4. E-I. A seriesof transverse sections of TF 1564, x 4. E, F. Opposite cutsat the apicad end of specimen. E. Orad facing cut. F. Apicadfacing cut. G. Apicad facing cut 14 mm from F. H, I.Opposite cuts 8 mm from G.
Remarks. Assignment of isolated endosiphuncles to agenus is very difficult and in most cases inadvisable. Anumber of endocerid genera have been established onetched-out isolated endosiphuncles (Flower 1955,1956a, b); although most of these genera are valid itis difficult to compare material not preserved in thesame fashion. Therefore comparison with these generais at best tentative when material is preserved inlimestone and only a series of transverse and longi-tudinal sections are available. The material in the pre-sent collections not only lacks details of the septalnecks and connecting rings, but also recrystallizationmakes interpretation of the structures within theendosiphuncle itself difficult. Therefore although thereare similarities between the Thailand material and anumber of previously described genera (e.g. Allocoto-
ceras, Stenosiphon, Platysiphon and Retroclitendo-ceras), the detailed comparison required to enable adefinite assignment is not possible.
Order TARPHYCERIDA Flower in Flowerand Kummel, 1950
Family TROCHOLITIDAE Chapman, 1857Hardmanoceras Teichert & Glenister, 1952
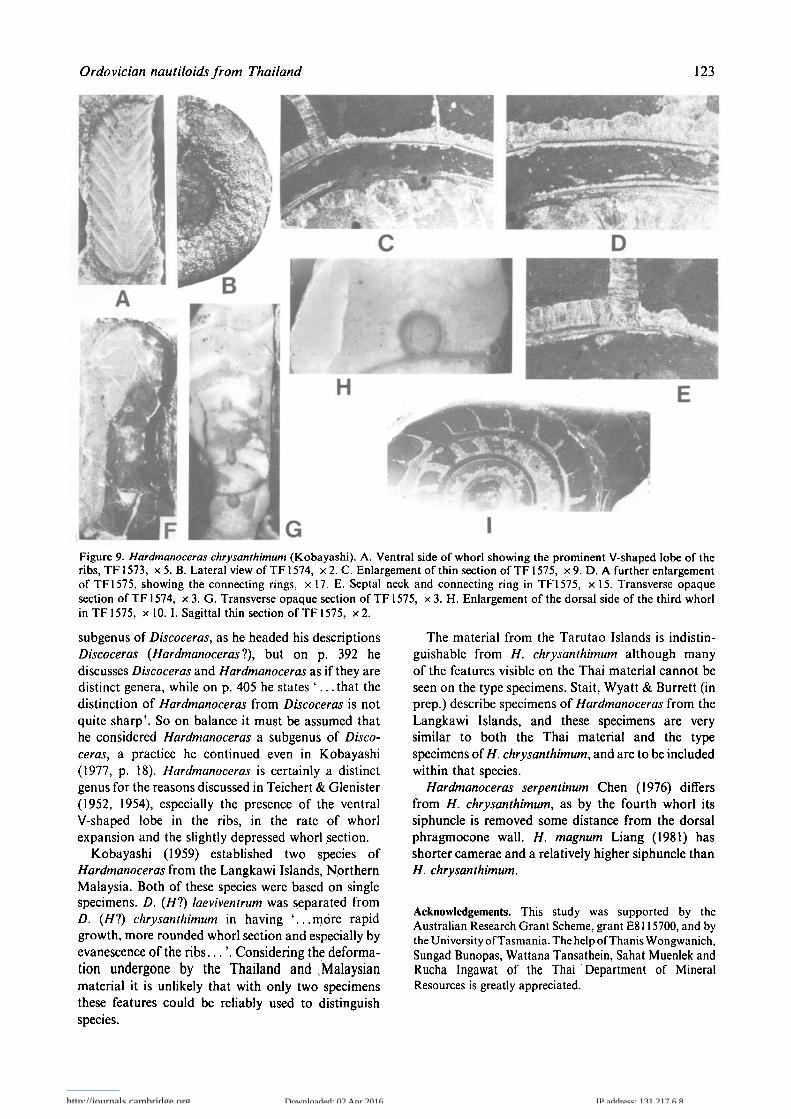
Hardmanoceras chrysanthimum (Kobayashi, 1959)
Fig. 9A-I1959 Discoceras {Hardmanoceras'}) chrysanthimumKobayashi, p. 404, pi. 26, fig. 1 a-b.1959 Discoceras (Hardmanoceras'?) laeviventrumKobayashi, p. 405, pi. 26, fig. 2a-b.
Material. Four specimens, TF 1573, TF 1574, TF 1575,TF 1576. Collected from the Thung Song Formationon the Tarutao Island (T55, locality A on Figs 1, 2).
Description. The conch is discoidal, 40 mm in diameter(TF1575). The best-preserved specimen (TF1575)consists of five slowly expanding whorls. TF 1575 hasslightly over half a whorl of the body chamberpreserved. All whorls are in contact and slightlyimpressed dorsally. The whorl section is broadlysub-rectangular and depressed, the flanks are gentlyand uniformly rounded. The height and width in mmof the whorls on TF 1574 from the inner to outer whorlare: 1.6?, 2.5; 2.1, 3.6; 3.3, 5.6; 5.6, 8.5 respectively.The corresponding measurements on TF 1575 are: 1.8,3.5; 2.3, 6.0; 3.2, 9.0; 5.4, 11.1; 6.3?,—. The diameterof the whorls on TF1575 are 3.6, 8.0, 14.6, 24.0,38.0? mm. The length of the camera on TF1575increases from 0.9 to 3.9 mm. The umbilicus has asmall perforation, 0.7 mm on TF1575. The suture isnot clear on any specimen, but there are probablydorsal and ventral lobes, while on the flanks it isstraight or possibly a slight saddle. There areprominent ribs across the flanks which become fainteras they bend back to form a deep V-shaped lobe acrossthe venter.
The siphuncle is approximately a quarter of theheight of the phragmocone, having a height of 0.7 mmat the third whorl of TF 1575. Siphuncle is in contactwith the dorsal wall of the phragmocone throughoutthe specimens. The septa are gently curved, septalnecks are orthochoanitic and one-seventh of the lengthof the camerae. The connecting rings are thick(0.1 mm) and do not expand between the septalforamina. The degree of recrystallization does notenable the distribution nor extent of cameral depositsto be reliably determined.
Remarks. There is some confusion in Kobayashi(1959) as to whether he considered Hardmanoceras tobe a separate genus or a subgenus of Discoceras.Kobayashi (1959) implies that Hardmanoceras was a
http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
Ordovician nautiloids from Thailand 123
IFigure 9. Hardmanoceras chrysanthimum (Kobayashi). A. Ventral side of whorl showing the prominent V-shaped lobe of theribs, TF1573, x 5. B. Lateral view of TF1574, x 2. C. Enlargement of thin section of TF1575, x 9. D. A further enlargementof TF1575, showing the connecting rings, x 17. E. Septal neck and connecting ring in TF1575, x 15. Transverse opaquesection of TF 1574, x 3. G. Transverse opaque section of TF 1575, x 3. H. Enlargement of the dorsal side of the third whorlin TF1575, x 10. I. Sagittal thin section of TF 1575, x2.
subgenus of Discoceras, as he headed his descriptionsDiscoceras {Hardmanoceras ?), but on p. 392 hediscusses Discoceras and Hardmanoceras as if they aredistinct genera, while on p. 405 he states ' . . . that thedistinction of Hardmanoceras from Discoceras is notquite sharp'. So on balance it must be assumed thathe considered Hardmanoceras a subgenus of Disco-ceras, a practice he continued even in Kobayashi(1977, p. 18). Hardmanoceras is certainly a distinctgenus for the reasons discussed in Teichert & Glenister(1952, 1954), especially the presence of the ventralV-shaped lobe in the ribs, in the rate of whorlexpansion and the slightly depressed whorl section.
Kobayashi (1959) established two species ofHardmanoceras from the Langkawi Islands, NorthernMalaysia. Both of these species were based on singlespecimens. D. (//?) laeviventrum was separated fromD. (//?) chrysanthimum in having ' . . . more rapidgrowth, more rounded whorl section and especially byevanescence of the r ibs . . . ' . Considering the deforma-tion undergone by the Thailand and ^Malaysianmaterial it is unlikely that with only two specimensthese features could be reliably used to distinguishspecies.
The material from the Tarutao Islands is indistin-guishable from H. chrysanthimum although manyof the features visible on the Thai material cannot beseen on the type specimens. Stait, Wyatt & Burrett (inprep.) describe specimens of Hardmanoceras from theLangkawi Islands, and these specimens are verysimilar to both the Thai material and the typespecimens of//, chrysanthimum, and are to be includedwithin that species.
Hardmanoceras serpentinum Chen (1976) differsfrom H. chrysanthimum, as by the fourth whorl itssiphuncle is removed some distance from the dorsalphragmocone wall. H. magnum Liang (1981) hasshorter camerae and a relatively higher siphuncle thanH. chrysanthimum.
Acknowledgements. This study was supported by theAustralian Research Grant Scheme, grant E8115700, and bythe University of Tasmania. The help of Thanis Wongwanich,Sungad Bunopas, Wattana Tansathein, Sahat Muenlek andRucha Ingawat of the Thai Department of MineralResources is greatly appreciated.

http://journals.cambridge.org Downloaded: 02 Apr 2016 IP address: 131.217.6.8
124 Ordovician nautiloids from Thailand
References
BARNES, C. R. 1977. Ordovician conodonts from the ShipPoint andv Bad Cache Rapids Formations, MelvillePeninsula, southeastern District of Mackenzie. Geo-logical Survey of Canada, Bulletin 269, 99-119.
BROWN, G. F., BURAVAS, S., CHARALJAVANAPHET, J.,
JALICHANDRA, N., JOHNSTON, W. D. JR., SRESTHAPUTRA,
V. & TAYLOR, G. C. 1951. Geologic Reconnaissance ofthe Mineral Deposits of Thailand. United StatesGeological Survey Bulletin no. 984, 183 pp.
BUNOPAS, S. 1982. Paleogeographic history of WesternThailand and adjacent parts of South-East Asia - aplate tectonics interpretation. Geological Survey ofThailand, Special Paper no. 5, 810 pp.
BURTON, C. K. 1974. The Satun Group (Nai Tak Formationand Thung Song Limestone) of Penninsular Thailand.Sains Malaysiana 3 (1), 15-34.
CHEN JUNYUAN. 1976. Advances in the Ordovicianstratigraphy of North China with a brief descrip-tion of nautiloid fossils. Ada PalaeontologicaSinica 15 (1), 55-74 (in Chinese with English summary).
EPSTEIN, A. G., EPSTEIN, J. B. & HARRIS, L. D. 1977.
Conodont color alteration - an index to organic meta-morphism. United States Geological Survey Profes-sional Paper 995, 27 pp.
FLOWER, R. H. 1955. New Chazyan Orthocones. Journal ofPaleontology 29 (5), 806-30.
FLOWER, R. H. 1956a. Cephalopods from the Canadian ofMaryland. Journal of Paleontology 30 (1), 75-96.
FLOWER, R. H. 19566. Some endoceroids from the El PasoLimestone. Journal of Paleontology 30 (1), 97-100.
FLOWER, R. H. 1957. Studies of the Actinoceratida. NewMexico Bureau of Mines and Mineral Resources,memoir no. 2, 100 pp.
FLOWER, R. H. 1968. The first great expansion of theactinoceroids. New Mexico Bureau of Mines andMineral Resources, memoir no. 19 (1), 1-16.
FLOWER, R. H. 1976. New American Wutinoceratidae withreview of actinoceroid occurrences in eastern hemi-sphere. New Mexico Bureau of Mines and MineralResources, memoir no. 28 (1), 5-12.
FOERSTE, A. F. 1924. Silurian cephalopods of northernMichigan. Universtiy of Michigan Museum of Paleonto-logy, contribution 2, 19-86.
HOOK, S. C. & FLOWER, R. H. 1977. Late Canadian (ZonesJ, K) cephalopod faunas from south-western UnitedStates. New Mexico Bureau of Mines and MineralResources, memoir no. 32, 9-102.
INGAVAT, R., MUENLEK, S. & UDOMRATN, C. 1975. On the
discoveries of some Permian fusulinids and Ordoviciancephalopods of Banrai, West Thailand. Journal of theGeological Society of Thailand 1 (1-2), 81-9.
KOBAYASHI, T. 1935. Restudy on Manchuroceras with a briefnote on the classification of endoceroids. Journal of theGeological Society of Japan 42 (506), 736-52.
KOBAYASHI, T. 1958. Some Ordovician fossils from theThailand-Malayan borderland. Japanese Journal ofGeology and Geography 29 (4), 223-31.
KOBAYASHI, T. 1959. On some Ordovician fossils fromnorthern Malaya and her adjacence. Journal of theFaculty of Science, University of Tokyo 11, 387-407.
KOBAYASHI, T. 1961. On the occurrence of Ordoviciannautiloids in North Thailand. Japanese Journal ofGeology and Geography 32(1), 79-84.
KOBAYASHI, T. 1977. Manchuroceras found in South Koreawith notes on the Manchuroceratidae and the Manchuro-ceras province. Transactions and Proceedings of thePalaeontological Society of Japan 105, 17-26.
KOBAYASHI, T. & HAMADA, T. 1964. On the Middle
Ordovician fossils from Satun, the Malaysian Frontierof Thailand. Geology and Palaeontology of SoutheastAsia 1, 269-78.
LIANG ZHONGFA. 1981. Ordovician cephalopods fromHunjiang region of Jilin and northern Neimongol.Ada Palaeontologica Sinica 20 (5), 393-9 (in Chinese
with English summary).MCTAVISH, R. A. 1973. Prioniodontacean conodonts from
the Emanuel Formation (Lower Ordovician) ofWestern Australia. Geologica el Palaeontologica 7,27-58.
MOUND, M. C. 1965. A conodont fauna from the JoinsFormation (Ordovician), Oklahoma. Tulane Studies inGeology 4 (1), 1-45.
OZAKI, K. 1927. On a new genus of Ordovician Cepholopodafrom Manchuria. Geological Society of Tokyo 34,45-50.
Qi DUNLUN. 1980. Ordovician cephalopods from Wuwei ofAnhui and their stratigraphical significance. AdaPalaeontologica Sinica 19 (4), 245-62 (in Chinese withEnglish summary).
Ross, R. J. JR. & 27 authors. 1982. The Ordovician systemin the United States. International Union of GeologicalSciences Publication, no. 12, 73 pp.
SHIMIZU, S. & OBATA, T. 1935. New genera of Gotlandianand Ordovician nautiloids. Shanghai Science InstituteJournal, section 2, 2, 1-10.
STAIT, B. A. & BURRETT, C. F. 1982. Wutinoceras (Nauti-loidea) from the Setul Limestone (Ordovician) ofMalaysia. Alcheringa 6, 193-6.
STAIT, B. A., BURRETT, C. F. & WONGWANICH, T. 1983.
Ordovician trilobites from the Tarutao Formation,Southern Thailand. Neues Jahrbuch fur Geologie undPalaontologie (in press).
STAIT, B. A., WYATT, D. & BURRETT, C. F. (in prep.).
Ordovician nautiloids from the Setal Limestone of theLangkawi Islands, Northern Malaysia.
TEICHERT, C. 1947. Early Ordovician cephalopods fromAdamsfield, Tasmania. Journal of Paleontology 21 (5),420-8.
TEICHERT, C. & GLENISTER, B. F. 1952. Fossil nautiloidfaunas from Australia. Journal of Paleontology 26 (5),730-52.
TEICHERT, C. & GLENISTER, B. F. 1953. Ordovician and
Silurian cephalopods from Tasmania, Australia. Bul-letins of American Paleontology 34, 66 pp.
TEICHERT, C. & GLENISTER, B. F. 1954. Early Ordoviciancephalopod fauna from Northwestern Australia. Bul-letins of American Paleontology 35 (150), 1-12.
TERAOKA, Y., SAWATA, H., YOSHIDA, T. & PUNGRASSAMI, T.
1982. Lower Paleozoic formations of the TarutaoIslands, southern Thailand - report of stratigraphicstudy team No. 1. Prince of Songkhla University,Geological Research Project Publication, no. 6, 54 pp.
WADE, M. 1977. Georginidae, new family of actinoceroidcephalopods, Middle Ordovician, Australia. Memoirsof the Queensland Museum 18 (1), 1-15.
Zou XIPING. 1981. Early Ordovician nautiloids fromQingshuihe, nei Monggol (Inner Mongolia) andPianguan, Shanxi Province. Ada Palaeontologica Sinica20 (4), 353-62 (in Chinese with English summary).
Related Documents











