Plant Cell, Tissue and Organ Culture 71: 213–222, 2002. © 2002 Kluwer Academic Publishers. Printed in the Netherlands. 213 Optimizing embryogenic callus production and plant regeneration from ‘Tifton 9’ bahiagrass seed explants for genetic manipulation Magali F. Grando 1 , Chandra I. Franklin 3 & Robert G. Shatters Jr. 1,2,∗ 1 Department of Agronomy, University of Florida, P.O. Box 110300 Gainesvillle, FL 32611-0300, USA; 2 USDA, ARS, USHRL, 2001 South Rock Rd. Fort Pierce, FL 34945, USA; 3 Department of Biology, Savannah State Uni- versity, P.O. Box 20600, Savannah, GA 31404, USA ( ∗ requests for offprints; Fax: +1-561-462-5986; E-mail: [email protected]) Received 10 July 2001; accepted in revised form 9 April 2002 Key words: forage grass, monocot, Paspalum notatum Flügge, somatic embryos, tissue culture Abstract Bahiagrass (Paspalum notatum Flügge) is a warm season forage grass widely cultivated in southeastern U.S. and South America. The cultivar Tifton 9 has several desirable characteristics such as high forage yield, more vigor at the seedling stage, etc.; but its forage quality is very low. As an initial step for future genetic manipulations to improve its forage characteristics, we have optimized in vitro culture conditions for plant regeneration. In this report, we describe an efficient method for embryogenic callus induction and plant regeneration from bahiagrass (cv. Tifton 9) seed explants, which are readily available and easy to manipulate, compared to other explant sources reported in the literature. Murashige and Skoog (MS) medium containing 30 µM dicamba and 5 µM 6-benzyladenine (BA) was optimal for callus induction and growth. Out of 9734 seeds cultured, 65.7% germinated and 21.4% produced embryogenic callus on this medium. Shoot formation was best when embryogenic calluses induced in this medium were trans- ferred to MS medium supplemented with 5 µM BA and 1 µM gibberellic acid with 1640 plantlets formed per gram fresh weight of callus tissue. When transferred to hormone-free SH medium, shoot systems produced well- developed root systems. The resulting plantlets grew normally produced viable seeds when transferred to soil in the greenhouse. Histochemical staining for GUS activity arising from transient expression of the introduced uidA (β -glucuronidase) gene indicated that bahiagrass embryogenic callus produced by this method is suitable for gene transfer via biolistic bombardment; and it can serve as a good target tissue for future genetic manipulations to improve the forage quality of bahiagrass (cv. Tifton 9). Abbreviations: MS-B – MS salts and B5 vitamins; BA – 6 benzyladenine; GA 3 – gibberellic acid; GUS – β -glucuronidase; NAA – naphthaleneacetic acid; 2,4-D – 2,4-dichlorophenoxyacetic acid; SH – Schenk and Hildebrandt medium Introduction Bahiagrass (Paspalum notatum Flügge) is a warm- season perennial forage widely cultivated in tropical and subtropical regions in the world. In the south- eastern USA, it is grown in 2.5 million hectares of improved pastures. The most common cultivar is Pensacola, a diploid out crossing population. The cul- tivar Tifton 9 was developed from a subpopulation of Pensacola cultivar using a recurrent restrictive phen- otype selection method, selecting for higher forage yield, and more vigor at seedling stage (Burton, 1989). Bahiagrass has many desirable pasture character- istics including tolerance to low soil fertility and poor drainage conditions, resistance to many diseases and pests, and persistence under low maintenance. Its nutritional value and digestibility, however; are low especially in mid-summer and early fall. The goal of

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Plant Cell, Tissue and Organ Culture 71: 213–222, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
213
Optimizing embryogenic callus production and plant regeneration from‘Tifton 9’ bahiagrass seed explants for genetic manipulation
Magali F. Grando1, Chandra I. Franklin3 & Robert G. Shatters Jr.1,2,∗1Department of Agronomy, University of Florida, P.O. Box 110300 Gainesvillle, FL 32611-0300, USA; 2USDA,ARS, USHRL, 2001 South Rock Rd. Fort Pierce, FL 34945, USA; 3Department of Biology, Savannah State Uni-versity, P.O. Box 20600, Savannah, GA 31404, USA (∗requests for offprints; Fax: +1-561-462-5986; E-mail:[email protected])
Received 10 July 2001; accepted in revised form 9 April 2002
Key words: forage grass, monocot, Paspalum notatum Flügge, somatic embryos, tissue culture
Abstract
Bahiagrass (Paspalum notatum Flügge) is a warm season forage grass widely cultivated in southeastern U.S. andSouth America. The cultivar Tifton 9 has several desirable characteristics such as high forage yield, more vigorat the seedling stage, etc.; but its forage quality is very low. As an initial step for future genetic manipulationsto improve its forage characteristics, we have optimized in vitro culture conditions for plant regeneration. In thisreport, we describe an efficient method for embryogenic callus induction and plant regeneration from bahiagrass(cv. Tifton 9) seed explants, which are readily available and easy to manipulate, compared to other explant sourcesreported in the literature.
Murashige and Skoog (MS) medium containing 30 µM dicamba and 5 µM 6-benzyladenine (BA) was optimalfor callus induction and growth. Out of 9734 seeds cultured, 65.7% germinated and 21.4% produced embryogeniccallus on this medium. Shoot formation was best when embryogenic calluses induced in this medium were trans-ferred to MS medium supplemented with 5 µM BA and 1 µM gibberellic acid with 1640 plantlets formed pergram fresh weight of callus tissue. When transferred to hormone-free SH medium, shoot systems produced well-developed root systems. The resulting plantlets grew normally produced viable seeds when transferred to soil inthe greenhouse. Histochemical staining for GUS activity arising from transient expression of the introduced uidA(β-glucuronidase) gene indicated that bahiagrass embryogenic callus produced by this method is suitable for genetransfer via biolistic bombardment; and it can serve as a good target tissue for future genetic manipulations toimprove the forage quality of bahiagrass (cv. Tifton 9).
Abbreviations: MS-B – MS salts and B5 vitamins; BA – 6 benzyladenine; GA3 – gibberellic acid; GUS –β-glucuronidase; NAA – naphthaleneacetic acid; 2,4-D – 2,4-dichlorophenoxyacetic acid; SH – Schenk andHildebrandt medium
Introduction
Bahiagrass (Paspalum notatum Flügge) is a warm-season perennial forage widely cultivated in tropicaland subtropical regions in the world. In the south-eastern USA, it is grown in 2.5 million hectaresof improved pastures. The most common cultivar isPensacola, a diploid out crossing population. The cul-tivar Tifton 9 was developed from a subpopulation of
Pensacola cultivar using a recurrent restrictive phen-otype selection method, selecting for higher forageyield, and more vigor at seedling stage (Burton, 1989).
Bahiagrass has many desirable pasture character-istics including tolerance to low soil fertility and poordrainage conditions, resistance to many diseases andpests, and persistence under low maintenance. Itsnutritional value and digestibility, however; are lowespecially in mid-summer and early fall. The goal of

214
our research is to use genetic engineering technologyto improve the forage quality of bahiagrass (cv. Tifton9) by modifying its protein and lignin content.
Establishment of a suitable tissue culture systemto produce regenerable target tissue is critical for theregeneration of transgenic grasses (Ritala et al., 1995).In gene transfer experiments with monocots, the em-bryogenic callus has been frequently used as the targettissue, because it can be manipulated to produce fertiletransformed plants. Transgenic maize (Genovesi et al.,1992; Wan at al., 1995), wheat (Vasil et al., 1992),oat (Somers et al., 1992; Cho et al., 1999), barley(Wan and Lemaux, 1994), rice (Sivamani et al., 1996),sugarcane (Bower and Birch, 1992; Gallo-Meagherand Irvine, 1996) and creeping bent grass (Zhong etal., 1993) plants have been obtained via particle bom-bardment mediated-DNA transfer using embryogeniccallus as the target tissue.
Explants such as immature embryos and younginflorescence tissues have been used to initiate em-bryogenic callus cultures capable of plant regenerationin numerous forage grass species including pearl mil-let (Pennisetum americanum) (Vasil and Vasil, 1981;Vasil, 1988), napier grass (Pennisetum purpureum)(Wang and Vasil, 1982), tall festuca (Fescua ar-undinacea), ryegrass (Lolium multiflorum), guineagrass (Panicum maximum) (see Ozias-Akins and Vasil,1988), bent grass (Agrostis palustris) (Zhong et al.,1993), buffalo grass (Cenchrus ciliaris) (Sankhla andSankhla, 1989), Kentucky bluegrass (Poa pratensis)(Van der Valk et al., 1989), dallisgrass (Paspalumdilatatum) (Bovo and Mroginski, 1986; Akashi andAdashi, 1992), and seashore paspalum (Paspalum va-ginatum) (Cardona and Duncan, 1997). Embryogeniccallus has also been produced from mature seeds ofKentucky bluegrass (Griffin and Dibble, 1995; Vander Valk et al., 1995), bent grass (Zhong et al., 1991),buffalo grass (Ross et al., 1995), caucasian bluestem(Bothriochloa caucasia) (Franklin et al., 1990) andoat (Cho et al., 1999). Shoot apical meristems isolatedfrom germinated seeds or seedlings were found to be ahighly regenerative explant source for callus inductionin barley (Weigel and Hughes, 1985; Vitanova et al.,1995), wheat (Wernicke and Milkovits, 1986; Varsh-ney et al., 1998), sorghum (Bhaskaran and Smith,1989; Gendy et al., 1996), and oat (Chen et al., 1995).
Plant regeneration has been reported from bahia-grass callus initiated from immature inflorescence(Bovo and Mroginski, 1986), mature and immatureembryos (Bovo and Mroginski, 1989), mature caryop-sis isolated from seeds by removing lemma and palea
(Marousky and West, 1990; Akashi et al., 1993), andleaf-stem sections (Shatters Jr. et al., 1994). Thusfar, there are no reports on regeneration of fertilebahiagrass plants from callus induced from germinat-ing mature seeds. Mature seed explants are preferredover other explants because they are readily availableyear-round, and the explant preparation prior to cul-ture initiation is less labor intensive. Bahiagrass seedscan be harvested, dried and stored at 4 ◦C for over ayear without compromising seed viability. Whereas,explants such as immature inflorescence or immatureembryos can be collected only during the reproductivephase of the plant, and they have to be used imme-diately after the harvest. Also, the use of mature fullseeds eliminates the laborious and time-consumingprocedures involved in isolation of immature embryosor isolation of mature caryopsis from each seed byremoving lemma and palea, which are very small inbahiagrass.
In this report, results from studies that led tothe development of an efficient tissue culture systemfrom bahiagrass (cv. Tifton 9) mature full-seed ex-plants are presented. In this system, high frequencyof embryogenic callus induction and fertile plant re-generation were observed. In addition, the potentialfor integrating this tissue culture system with particlebombardment mediated-DNA delivery for regenerat-ing transgenic bahiagrass is discussed.
Material and methods
Induction of embryogenic callus
Mature seeds of bahiagrass (cv. Tifton 9) were sur-face sterilized in 50% commercial bleach (Clorox�)solution with 2–3 drops of tween 20 (Sigma Chem-ical Co., St. Louis, MO) for 7 min followed by a70% ethanol treatment for 7 min, and three rinses insterile water. Surface-sterilized seeds were cultured inMS-B medium containing MS salts (Murashige andSkoog, 1962), B5 vitamins (Gamborg et al., 1968), 3%(w/v) sucrose, 0.8% (w/v) agar and various concentra-tions of auxins [3,6-dichloro-o-anisic acid (dicamba)or 2,4-dichlorophenoxyacetic acid (2,4-D)] along witha cytokinin 6-benzyladenine (BA). The pH was adjus-ted to 5.7 prior to the addition of agar. Each growthregulator treatment consisted of six replications withapproximately 100 seeds per replication. The cul-tures were maintained at 27 ◦C in the dark for callusinduction. The frequency of embryogenic callus in-

215
duction from germinated seeds was evaluated 6 weeksafter culture initiation. Embryogenic callus clumpswere then isolated from explants and subcultured ontofresh medium. Four weeks after the subculture, callusclumps were transferred to the regeneration medium.
Plant regeneration
To study the influence of growth regulators on the con-version of somatic embryos into plantlets two typesof experiments were performed. First, optimization ofcallus induction medium was performed using com-binations of callus induction and shoot developmentmedia. In this experiment, 30 callus clumps (approx-imately 5 mm in diameter) induced on each of the sixdifferent callus induction media were transferred tothree different shoot development media composed ofMS-B medium with or without BA (5 µM), or a com-bination of 13 µM BA and 5 µM naphthaleneaceticacid (NAA). Each treatment consisted of four replic-ates. The cultures were maintained at 27 ◦C with a16-h photoperiod, provided by cool white fluorescentlights. Four weeks after the transfer, the number ofvisible shoots formed per callus clump was determinedassuming each shoot system represented a regeneratedplantlet.
In the second experiment, shoot development re-sponse was compared using combinations of plantgrowth regulators in the shoot development media andsomatic embryos removed from the optimal callus in-duction medium (MS-B medium containing 30 µMdicamba and 5 µM BA). Thirty-six embryogenic cal-lus clumps (approximately 40 mg each) produced onMS-B+30 µM dicamba and 5 µM BA were evenlydivided among six shoot development media differingin BA and gibberellic acid (GA3) levels. Five weeksafter the transfer, the number of green shoots formedper gram (fresh weight) of callus initially used wasdetermined.
To promote the growth and development of rootsystems, approximately 30 plantlets (with shoot sys-tems 10 mm or longer, harvested from callus clumpscultured on shoot development medium) were trans-ferred to hormone-free SH medium (Schenk andHildebrandt, 1972) supplemented with B5 vitamins,3% (w/v) sucrose and 0.8% (w/v) agar. After 3 weeks,the quality of root systems produced by these plantletswas determined based on an arbitrary scale of one-to-three as described below. Grade 1, small primaryroots (≤2 cm) with no secondary roots; grade 2, smallprimary roots with secondary roots; and grade 3, well-
developed primary roots >2 cm with well-establishedsecondary roots.
Plantlets with well-established shoot and root sys-tems were transferred to the greenhouse, processedthrough the standard greenhouse acclimatization pro-cedure and grown to maturity. Seeds produced fromthese plants were collected and tested for viability bygerminating them on moist filter paper.
Analysis of variance (ANOVA) and the Duncanmultiple comparison test at the p<0.05 level of signi-ficance were used to analyze the data in this study.
Gene transfer
To test the use of embryogenic callus obtained frommature seed explants as a suitable target tissue forgenetic transformation of bahiagrass via particle bom-bardment mediated-DNA transfer, callus clumps werebombarded with 1 µm gold particles (Bio Rad Labor-atories, Hercules, CA) coated with the plasmid pAHC25 (Christensen et al., 1992; Christensen and Quail,1996), following the protocol described by Somers atal. (1992). The Biolistic� PDS 1000/He particle de-livery system (Bio Rad Laboratories, Hercules, CA)was used. The influence of particle velocity at heliumpressures 800, 1100, 1300 and 1500 psi; and the effectof bombarding the same target twice on the efficiencyof DNA transfer were tested. For each bombardment,3.5 µg of DNA was used and each treatment wasrepeated three times.
Efficiency of gene transfer was evaluated based onGUS (β-glucuronidase) activity encoded by the (uidA)reporter gene (contained in the plasmid pAHC 25)driven by the maize ubiquitin promoter. One day afterthe bombardment, callus samples were assayed forGUS activity following the standard GUS histochem-ical assay (Jefferson, 1987) using a modified buffersolution containing 25 mM NaPO4, pH 7.0, 0.25 mMEDTA, 1.25 mM potassium ferricyanide, 1.25 mMpotassium ferrocyanide, 0.25% Triton, and 1 mM5-bromo-4-chloro-3-indolyl glucuronic acid (X-gluc)(Labscientific Inc., Livingston, NJ) substrate.
Histology
For histological studies to determine the origin of cal-lus, explants were collected 2 weeks after culture ini-tiation. A standard histological procedure (Berlyn andMiksche, 1976) was followed for fixation (in FAA),paraffin embedding and sectioning. Sections were

216
Table 1. Frequency of total and embryogenic callus produced from mature bahiagrass seeds after 6 weeks of germination on MS basal mediasupplemented with 30 µM dicamba and 5 µM BA
Replication No. of seeds % of seeds % of seeds % of
used that germinated producing callus∗ seeds producing
embryogenic callus∗
1 1715 66.3 87.9 17.8
2 1466 65.9 91.2 22.8
3 1546 69.0 89.6 23.5
4 1449 65.1 90.5 27.8
5 1504 70.6 90.3 22.3
6 2054 59.4 77.9 16.1
Total 9734 65.7 87.5 21.4
∗Callus frequency from germinated seeds after 6 weeks of culture.
stained with toluidine blue as described previously(Franklin et al., 1991).
Results and discussion
Embryogenic callus induction
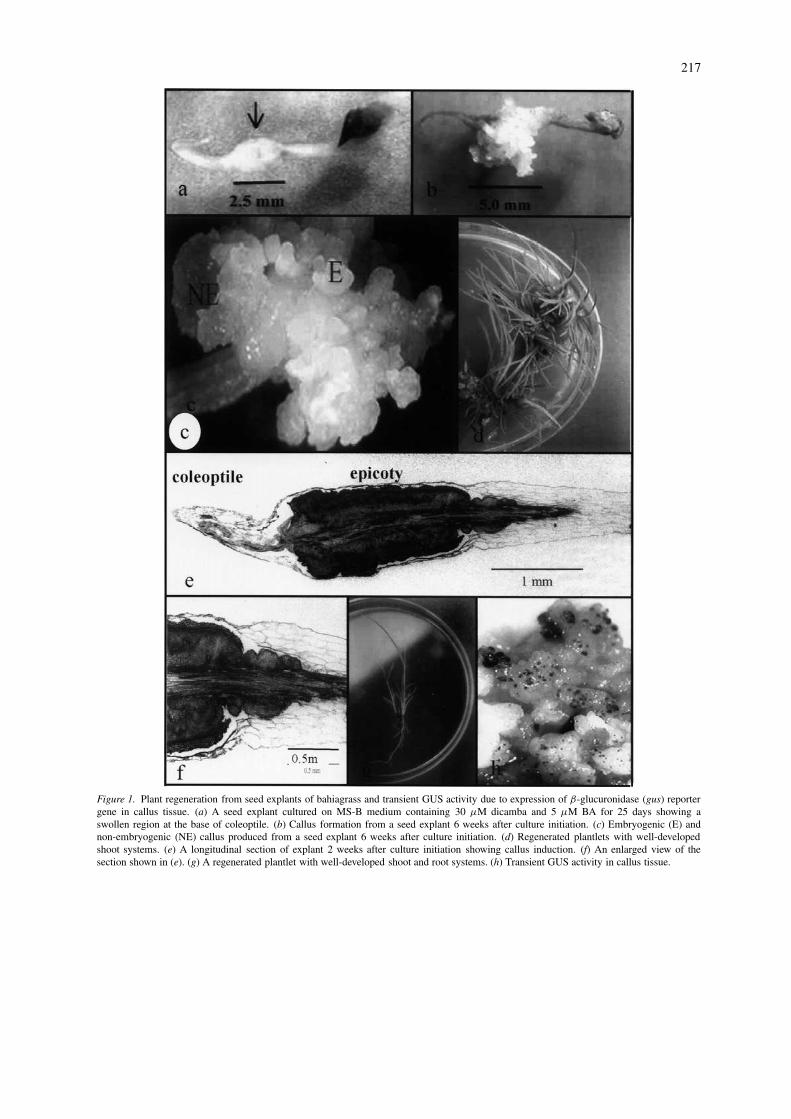
Bahiagrass seed explants germinated approximately 8days after placing on callus induction media. Two tothree weeks after culture initiation, the epicotyl regionat the base of the coleoptile showed swelling (Figure1a), and callus was produced from this swollen region6 weeks after culture initiation (Figure 1b). This ob-servation is consistent with results from histologicalstudies. Active proliferation of cells in the epicotylregion below the coleoptile was observed in the lon-gitudinal sections of 2-week-old explants (Figure 1eand f). Dense staining and relatively smaller size ofthese cells when compared to cells outside this regionsuggest active cell division and meristematic activityresulting in callus formation.
Six to eight weeks after the culture initiation, cal-lus produced by the seed explants consisted of bothembryogenic and non-embryogenic types (Figure 1c).The non-embryogenic callus was friable, soft andtranslucent. Whereas, the embryogenic callus wascompact, nodular and yellowish-white in color similarto the type I embryogenic callus described by Vasil(1988).
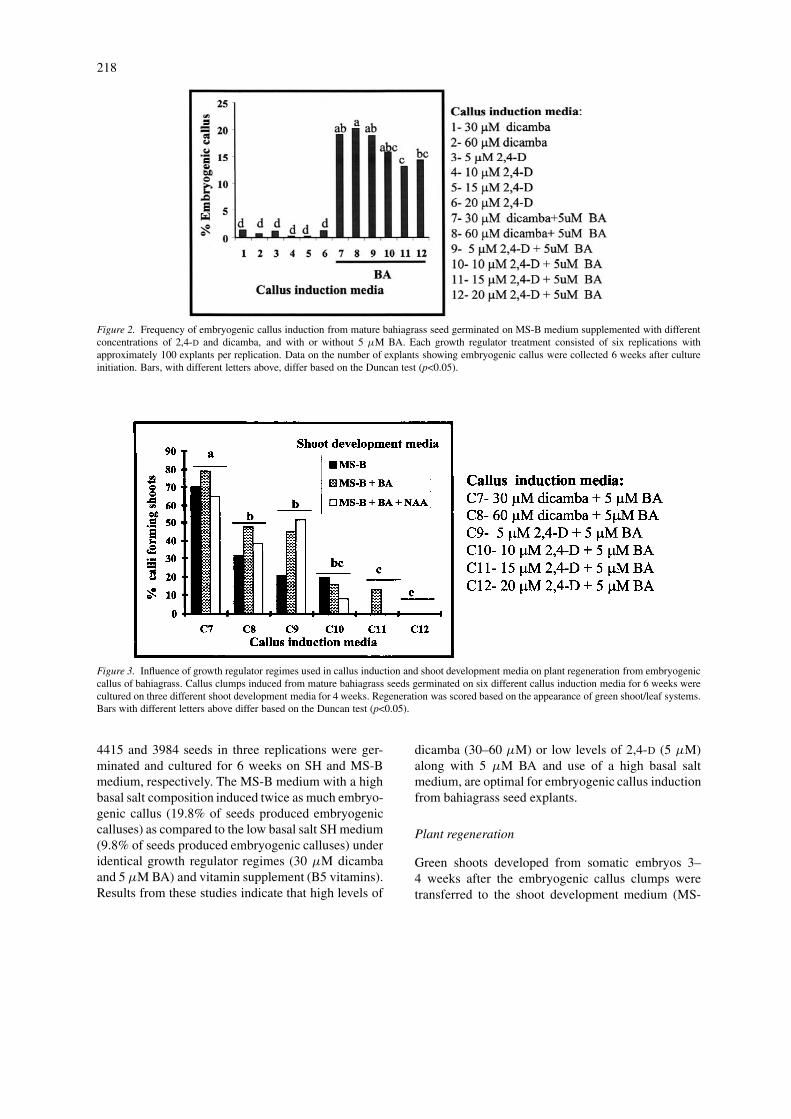
Significant (p<0.001) variation in the frequencyof embryogenic callus induction was observed amongdifferent growth regulator regimes tested (Figure 2).Data presented in Figure 2 clearly indicate that addi-tion of a cytokinin is essential for embryogenic callus
induction from bahiagrass seed explants. The highestfrequency of embryogenic callus production was ob-served with the addition of 5 µM BA in the presenceof either 30 or 60 µM dicamba (19.0 and 20.3%, re-spectively) or low levels, 5 and 10 µM of 2,4-D (18.9and 15.8%, respectively). A very low frequency (0.2–1.3%) of embryogenic callus was produced from seedscultured in the absence of BA. In another experimentdesigned to obtain a large amount of tissue for particlebombardment, approximately 9700 seeds were cul-tured on MS-B medium containing 30 µM dicambaand 5 µM BA. As shown in Table 1, 65.7% of theseseeds germinated, 87.5% of germinated seeds pro-duced callus and 21.4% of germinated seeds producedembryogenic callus. This frequency of embryogeniccallus production (21.4%) was higher than that ob-tained from mature bahiagrass caryopsis by Marouskyand West (1990) (11.6%) and Akashi et al. (1993)(15.2%).
Use of cytokinins in combination with auxins toinduce somatic embryogenesis in callus cultures hasbeen reported for a number of grasses including cer-eals (Bhaskaran and Smith, 1990). In our study, theaddition of BA to medium containing auxin increasedthe frequency of embryogenic callus formation ap-proximately 20-fold. This is consistent with workon sorghum shoot meristem cultures (Bhaskaran andSmith, 1989; Gendy et al., 1996) and corn (Zhonget al., 1992). The requirement of BA in somatic em-bryogenesis may be dependent on explant source, asobserved by Bhaskaran and Smith (1990).
Once the optimal growth regulator combinationwas chosen, the influence of medium salt compositionon somatic embryo production was tested. A total of
217
Figure 1. Plant regeneration from seed explants of bahiagrass and transient GUS activity due to expression of β-glucuronidase (gus) reportergene in callus tissue. (a) A seed explant cultured on MS-B medium containing 30 µM dicamba and 5 µM BA for 25 days showing aswollen region at the base of coleoptile. (b) Callus formation from a seed explant 6 weeks after culture initiation. (c) Embryogenic (E) andnon-embryogenic (NE) callus produced from a seed explant 6 weeks after culture initiation. (d) Regenerated plantlets with well-developedshoot systems. (e) A longitudinal section of explant 2 weeks after culture initiation showing callus induction. (f) An enlarged view of thesection shown in (e). (g) A regenerated plantlet with well-developed shoot and root systems. (h) Transient GUS activity in callus tissue.
218
Figure 2. Frequency of embryogenic callus induction from mature bahiagrass seed germinated on MS-B medium supplemented with differentconcentrations of 2,4-D and dicamba, and with or without 5 µM BA. Each growth regulator treatment consisted of six replications withapproximately 100 explants per replication. Data on the number of explants showing embryogenic callus were collected 6 weeks after cultureinitiation. Bars, with different letters above, differ based on the Duncan test (p<0.05).
Figure 3. Influence of growth regulator regimes used in callus induction and shoot development media on plant regeneration from embryogeniccallus of bahiagrass. Callus clumps induced from mature bahiagrass seeds germinated on six different callus induction media for 6 weeks werecultured on three different shoot development media for 4 weeks. Regeneration was scored based on the appearance of green shoot/leaf systems.Bars with different letters above differ based on the Duncan test (p<0.05).
4415 and 3984 seeds in three replications were ger-minated and cultured for 6 weeks on SH and MS-Bmedium, respectively. The MS-B medium with a highbasal salt composition induced twice as much embryo-genic callus (19.8% of seeds produced embryogeniccalluses) as compared to the low basal salt SH medium(9.8% of seeds produced embryogenic calluses) underidentical growth regulator regimes (30 µM dicambaand 5 µM BA) and vitamin supplement (B5 vitamins).Results from these studies indicate that high levels of
dicamba (30–60 µM) or low levels of 2,4-D (5 µM)along with 5 µM BA and use of a high basal saltmedium, are optimal for embryogenic callus inductionfrom bahiagrass seed explants.
Plant regeneration
Green shoots developed from somatic embryos 3–4 weeks after the embryogenic callus clumps weretransferred to the shoot development medium (MS-

219
Table 2. Root formation from shoots produced on six different shoot development media∗ and cultivated in SH basal medium with no growthregulators for 3 weeks
Shoot development No. shoots % of shoots Grade for root
medium inoculated producing roots (ns) growth∗∗
1-MS 25 86.0 2.00 bc
2-MS+5 µM BA 30 77.7 2.34 b
3-MS+10 µM BA 25 75.8 1.54 c
4-MS+5 µM BA+1 µM GA3 38 72.8 2.44 ab
5-MS+5 µM BA+2 µM GA3 40 85.7 2.84 a
6-MS+5 µM BA+3 µM GA3 27 84.5 2.39 b
∗Shoot systems were produced from embryogenic callus produced by germinating mature bahiagrass seeds on MS-B with 30 µM dicambaand 5 µM BA for 6 weeks. Shoot development was stimulated by transfer to the indicated medium for 6 weeks prior to transfer to SH basalmedium to stimulate root growth. Means followed by different letters differ by Duncan test (p<0.05).∗∗Grade 1 – Small roots (to 2 cm) with no ramification. Grade 2 – Small root with ramification. Grade 3 – Large roots with ramification.ns=differences in the values in this column were not significant.
B±growth regulators). Prolific differentiation of shootsystems (Figure 1d) was first observed followed by thedevelopment of root systems. Addition of growth reg-ulators such as 5 µM BA or 13 µM BA and 5 µMNAA to the medium did not significantly (two-wayANOVA p=0.222) influence the development of shootsystems (Figure 3), however; the effect of growthregulator levels in the callus induction medium onshoot development was significant (p<0.0001). Nosignificant interaction (p=0.864) was found betweenshoot development media and callus induction me-dium. Among various growth regulator regimes in thecallus induction media tested, a combination of 30µM dicamba and 5 µM BA produced callus with thebest shoot development potential (i.e. 71.2% of cal-luses producing shoots, Figure 3). Whereas, callusinduced on medium containing high levels of 2,4-D
(i.e. >10 µM) expressed lower shoot development po-tential. This result is consistent with an earlier report(Bovo and Mroginski, 1989) in which high concentra-tions of 2,4-D present during callus induction reducedplant regeneration from excised bahiagrass embryos.
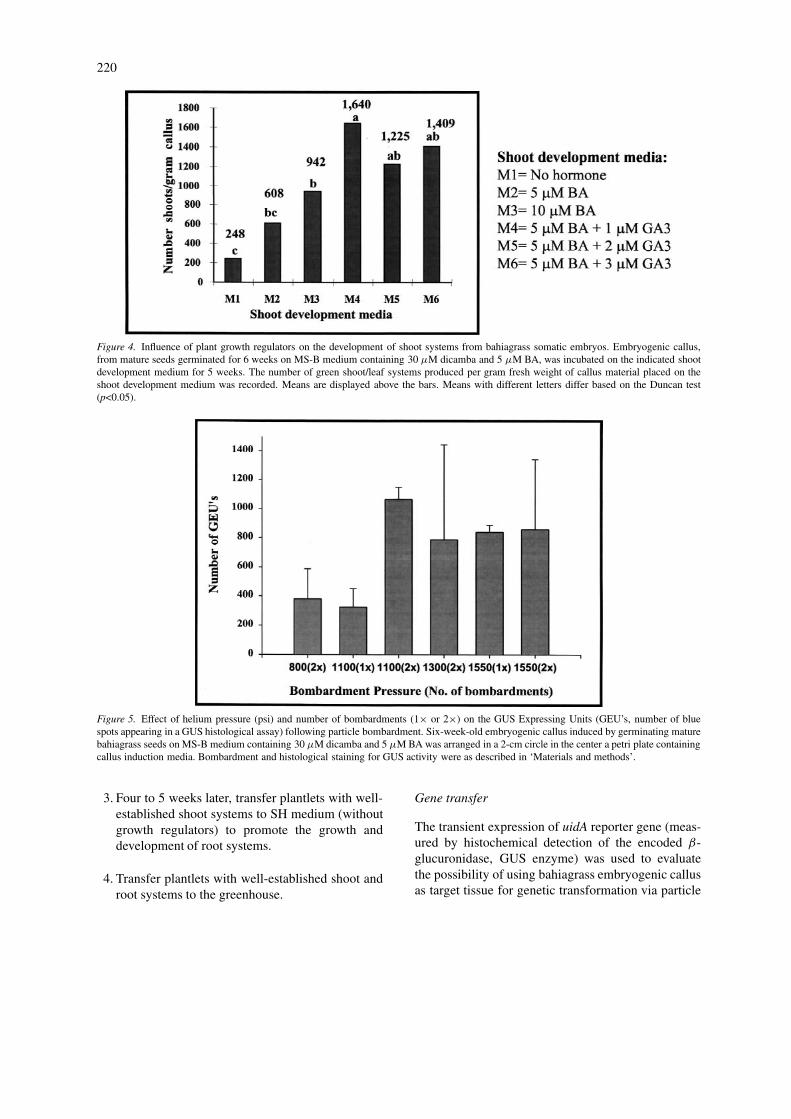
Using the optimal callus induction medium (MS-B with 30 µM dicamba and 5 µM BA), furtherexperiments were performed to optimize the growthregulator combination and level in the shoot devel-opment medium. Six-week-old embryogenic callusclumps produced on MS-B with 30 µM dicamba and5 µM BA were transferred to media culture containingdifferent combinations of BA and GA3. As shown inFigure 4, shoot development medium containing 5 µMBA and 1, 2 or 3 µM GA3 produced significantly more(p=0.0001) shoots per gram fresh weight of embryo-
genic callus than any other growth regulator regimestested.
Shoot systems ≥10 mm developed from this exper-iment were removed and transferred to hormone-freeSH medium to stimulate root development. Approxim-ately 80% of shoots produced on shoot developmentmedium developed root systems (Table 2). Growthregulator regimes in the shoot development mediumdid not significantly (p=0.653) influence the num-ber of shoots producing root systems. However, thequality of root systems was significantly (p<0.001) in-fluenced by the growth regulator regimes of the shootdevelopment medium. Plantlets cultivated on shoot de-velopment medium containing 5 µM BA and 1 or 2µM GA3 produced well-developed root systems con-sisting of primary roots >2 cm in length with profusesecondary roots. Plantlets, with well-established shootand root systems when transferred to the greenhouse,developed normally and produced viable seeds.
Optimal conditions for plant regeneration
Based on results presented above, it is concludedthat optimal conditions for plant regeneration frombahiagrass mature seed explants are:
1. Culture surface-sterilized seed explants on MS-Bmedium containing 30 µM dicamba and 5 µM BAfor 4 to 6 weeks for callus induction, which canbe subcultured on the same medium for two moreweeks to amplify tissue volume.
2. Transfer embryogenic callus clumps to shoot de-velopment medium (MS-B+5 µM BA+1 µM GA3to initiate plants development from somatic em-bryos.
220
Figure 4. Influence of plant growth regulators on the development of shoot systems from bahiagrass somatic embryos. Embryogenic callus,from mature seeds germinated for 6 weeks on MS-B medium containing 30 µM dicamba and 5 µM BA, was incubated on the indicated shootdevelopment medium for 5 weeks. The number of green shoot/leaf systems produced per gram fresh weight of callus material placed on theshoot development medium was recorded. Means are displayed above the bars. Means with different letters differ based on the Duncan test(p<0.05).
Figure 5. Effect of helium pressure (psi) and number of bombardments (1× or 2×) on the GUS Expressing Units (GEU’s, number of bluespots appearing in a GUS histological assay) following particle bombardment. Six-week-old embryogenic callus induced by germinating maturebahiagrass seeds on MS-B medium containing 30 µM dicamba and 5 µM BA was arranged in a 2-cm circle in the center a petri plate containingcallus induction media. Bombardment and histological staining for GUS activity were as described in ‘Materials and methods’.
3. Four to 5 weeks later, transfer plantlets with well-established shoot systems to SH medium (withoutgrowth regulators) to promote the growth anddevelopment of root systems.
4. Transfer plantlets with well-established shoot androot systems to the greenhouse.
Gene transfer
The transient expression of uidA reporter gene (meas-ured by histochemical detection of the encoded β-glucuronidase, GUS enzyme) was used to evaluatethe possibility of using bahiagrass embryogenic callusas target tissue for genetic transformation via particle

221
bombardment mediated-DNA transfer. The influenceof particle velocity (as determined by the helium pres-sure) and the effect of bombarding the same targettwice on GUS activity, as measured by the number ofGus Expressing Units (GEUs, number of blue spotsresulting from GUS histological assay, Figure 1h) ispresented in Figure 5. Maximum GEUs were detectedin callus bombarded twice with 1100 psi helium pres-sure, with an average of 1064 GEUs. This indicatesthe potential use of particle bombardment technologyto introduce and express foreign genes in this species.
The overall goal of this work was to develop an em-bryogenic callus induction and regeneration protocolto simplify the production of large quantities of bahia-grass (cv. Tifton 9) regenerable embryogenic callusfor use as the target tissue for gene transfer and re-generation of transgenic plants. We have successfullyachieved this goal, and have shown that the callus re-sponds to media and growth regulator supplements in amanner that would be expected based on the perform-ance of other gramineae species. This work should behelpful to researchers attempting to use biotechnolo-gical approaches to improve the forage characteristicsof bahiagrass and other related grass species.
Acknowledgements
The authors acknowledge Dr R. A. Wheeler, ARS-USDA, for technical support, Dr Rex Smith for criticalreview, and Dr S. H. West, Agronomy Dept. of Univer-sity of Florida, for donating the bahiagrass seeds usedin this experiment.
References
Akashi R & Adashi T (1992) Plant regeneration from suspensionculture-derived protoplast from apomitic dallisgrass (Paspalumdilatatum Poir.) Plant Sci. 82: 219–225
Akashi R, Hashimoto A & Adashi T (1993) Plant regeneration fromseed-derived embryogenic callus and cell suspension cultures ofbahiagrass (Paspalum notatum). Plant Sci. 90: 73–80
Berlyn GP & Miksche JP (1976) Botanical Microtechnique andCytochemistry (pp 30–65). Iowa State University Press, AmesIowa
Bhaskaran S & Smith RH (1989) Control of morphogenesis insorghum by 2,4-dichlorophenoxyacetic acid and cytokinins.Ann. Bot. 64: 217–224
Bhaskaran S & Smith RH (1990) Regeneration in cereal tissueculture: a review. Crop Sci. 30: 1328–1336
Bovo OA & Mroginski LA (1986) Tissue culture in Paspalum(Gramineae): plant regeneration from cultured inflorescences. J.Plant Physiol. 124: 481–492
Bovo OA & Mroginski LA (1989) Somatic embryogenesis and plantregeneration from cultured mature and immature embryos ofPaspalum notatum (Gramineae). Plant Sci. 65: 217–223
Bower R & Birch RG (1992) Transgenic sugarcane plants viamicroprojectile bombardment. Plant J. 2: 409–416
Burton GW (1989) Registration of ‘Tifton 9’ Pensacola bahiagrass.Crop Sci. 29: 1326
Cardona CA & Duncan RR (1997) Callus induction and highefficiency plant regeneration via somatic embryogenesis inPaspalum. Crop Sci. 37: 1297–1302
Chen Z, Zhuge Q & Sundqvist C (1995) Oat leaf base: tissue withan efficient regeneration capacity. Plant Cell Rep. 14: 354–358
Cho MJ, Jiang W & Lemaux PG (1999) High-frequency transform-ation of oat via microprojectile bombardment of seed-derivedhighly regenerative cultures. Plant Sci. 148: 9–17
Christensen AH, Sharrock RA & Quail PH (1992) Maize polyubi-quitin genes: structure, thermal perturbation of expression andtranscript splicing, and promoter activity following transfer toprotoplasm by electroporation. Plant Mol. Biol. 18: 675–689
Christensen AH & Quail PH (1996) Ubiquitin promoter-based vec-tors for high-level expression of selectable and/or screenablemarker genes in monocotyledonous plants. Transgenic Res. 5:213–218
Franklin CI, Trieu TN & Gonzales RA (1990) Plant regenerationthrough somatic embryogenesis in the forage grass Caucasianbluestem (Bothriochloa caucasica). Plant Cell Rep. 9: 443–446
Franklin CI, Trieu TN, Gonzales RA & Dixon RA (1991) Plantregeneration from seedling explants of green bean (Phaseolusvulgaris L.) via organogenesis. Plant Cell Tiss. Org. Cult. 24:199–206
Gallo-Meagher M & Irvine JE (1996) Herbicide resistant transgenicsugarcane plants containing the bar gene. Crop Sci. 36: 1367–1374
Gamborg OL, Miller RA & Ojima K (1968) Nutrient requirementsof suspension cultures of soybean root cells. Exp. Cell Res. 50:151–158
Gendy C, Séne M, Van Le B, Vidal J & Tran Thanh Van K (1996)Somatic embryogenesis and plant regeneration in Sorghumbicolor (l.) Moench. Plant Cell Rep. 15: 900–904
Genovesi D, Willetts N, Zachwieja S, Mann M, Spencer TM, FlickC & Gordon-Kamm WJ (1992) Transformation of an elite maizeinbred through microprojectile bombardment of regenerable em-bryogenic callus. In Vitro Cell Dev. Biol. 18: 189–200
Griffin JD & Dibble MS (1995) High-frequency plant regenerationfrom seed-derived callus cultures of Kentucky bluegrass (Poapratensis L.). Plant Cell Rep. 14: 721–724
Jefferson RA (1987) Assaying chimeric genes in plants: The GUSgene fusion system. Plant Mol. Biol. Rep. 5: 387–405
Marousky FJ & West SH (1990) Somatic embryogenesis andpant regeneration from cultured mature caryopses of bahiagrass(Paspalum notatum Flugge). Plant Cell Tiss. Org. Cult. 20:125–129
Murashige T & Skoog F (1962) A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol. Plant. 15:473–497
Ozias-Akins P & Vasil IK (1988) In vitro regeneration and geneticmanipulation of grasses. Physiol. Plant 73: 565–569
Ritala A, Aikasalo R, Aspegren K, Salmenkallio-Martila M, Aker-men S, Mannonen L, Kurtén U, Puupponen-Pimiäand R, TeeriTH & Kauppinen V (1995) Transgenic barley by particle bom-bardment. Inheritance of the transferred gene and characteristicof the transgenic barley plants. Euphytica 85: 81–88
Ross AH, Manners JM & Birch RG (1995) Embryogenic callusproduction, plant regeneration and transient gene expression fol-

222
lowing particle bombardment in pasture grass, Cenchrus ciliaris(Gramineae). Aust. J. Bot. 43: 193–199
Sankhla A & Sankhla N (1989) Tissue culture studies in desertplants: I. Cenchrus ciliaris cv. 75. Curr. Sci. 58: 872–874
Schenk RU & Hildebrandt AC (1972) Medium and techniques forinduction and growth of monocotyledonous plant cell cultures.Can. J. Bot. 50: 199–204
Shatters Jr. RS, Wheeler RA & West SH (1994) Somatic embryo-genesis and plant regeneration from callus culture of ‘Tifton 9’bahiagrass. Crop Sci. 34: 1378–1384
Sivamani E, Shen P, Opalka N, Beachy RN & Fauquet CM (1996)Selection of large quantities of embryogenic calli from indicarice seeds for production of fertile transgenic plants using thebiolistic method. Plant Cell Rep. 15: 322–327
Somers DA, Rines HW, Kaeppler HF & Bushnell WR (1992).Fertile, transgenic oat plants. Bio/Technology 10: 1589–1594
Van der Valk P, Ruis F, Tettelaar-Schrier AM & Van de Velde CM(1995) Optimizing plant regeneration from seed-derived callusculture of Kentucky bluegrass. The effect of benzyladenine. PlantCell Tiss. Org. Cult. 40: 101–103
Van der Valk P, Zaal MACM & Creemers-Molenaar J (1989) So-matic embryogenesis and plant regeneration in inflorescenceand seed derived callus culture of Poa pratensis L. (Kentuckybluegrass). Plant Cell Rep. 7: 644–647
Varshney A, Kant T & Kothari SL (1998) Plant regeneration fromcoleoptile tissue of wheat (Triticum aestivum L.). Biol. Plant. 40:137–141
Vasil IK (1988) Progress in the regeneration and genetic manipula-tion of cereal crops. Bio/Technology 6: 397–402
Vasil V, Castillo AM, Fromm ME & Vasil IK (1992) Herbi-cide resistant fertile transgenic wheat plants obtained by mi-croprojectile bombardment of regenerable embryogenic callus.Bio/Technology 10: 667–674
Vasil V & IK Vasil (1981) Somatic embryogenesis and plant regen-eration from tissue cultures of Pennisetum americanum and P.americanum×P. purpureum hybrid. Am. J. Bot. 68: 864–872
Vitanova Z, Vitanov V, Trifonova A, Savova D & Atanassov A(1995) Effect of 2,4-D precultivation on regeneration capacity ofcultivated barley. Plan Cell Rep. 14: 437–441
Wan Y & Lemaux PG (1994) Generation of large number of in-dependent transformed fertile barley plants. Plant Physiol. 104:37–48
Wan Y, Widholm JM & Lemaux PG (1995) Type I callus as a bom-bardment target for regeneration of fertile transgenic maize (Zeamays L.) Planta 196: 7–14
Wang D & Vasil IK (1982) Somatic embryogenesis and plant regen-eration from inflorescence segments of Pennisetum purpureumSchum. (Napier or elephant grass). Plant Sci. Lett. 25: 147–154
Weigel R & Hughes KH (1985) Long term regeneration by somaticembryogenesis in barley (Hordeum vulgaris L.) tissue culturesderived from apical meristem explants. Plant Cell Tiss. Org. Cult.5: 151–162
Wernicke W & Milkovits L (1986) The regeneration potential ofwheat shoot meristems in the presence and absence of 2,4-Dichlorophenoxyacetic acid. Protoplasma 131: 131–141
Zhong H, Srinivasan C & Sticklen MB (1991) Plant regeneration viasomatic embryogenesis in creeping bentgrass (Agrostis palustrisHuds.) Plant Cell Rep. 10: 453–456
Zhong H, Srinivasan C & Sticklen MB (1992) In vitro morpho-genesis in corn (Zea mays L.) I. Differentiation of multipleshoot clumps and somatic embryos from shoot tip. Planta 187:483–489
Zhong H, Boylard MG, Srinivasan C & Sticklen MB (1993) Trans-genic plants of turfgrass (Agrostis palustris L.) from micropro-jectile bombardment of embryogenic callus. Plant Cell Rep. 13:1–6
Related Documents











