Ontogeny of -Aminobutyric Acid-Immunoreactive Neuronal Populations in the Forebrain and Midbrain of the Sea Lamprey MIGUEL MELE ´ NDEZ-FERRO, 1 EMMA PE ´ REZ-COSTAS, 1 BEGON ˜ A VILLAR-CHEDA, 1 XESU ´ S MANOEL ABALO, 1 ROLANDO RODRI ´ GUEZ-MUN ˜ OZ, 2 MARI ´ A CELINA RODICIO, 1 AND RAMO ´ N ANADO ´ N 1 * 1 Department of Fundamental Biology, Faculty of Biology, University of Santiago de Compostela, 15782 Santiago de Compostela, Spain 2 Department of Biology of Organisms and Systems, Faculty of Biology, University of Oviedo, 33006 Oviedo, Spain ABSTRACT Although brain organization in lampreys is of great interest for understanding evolution in vertebrates, knowledge of early development is very scarce. Here, the development of the fore- brain and midbrain -aminobutyric acid (GABA)-ergic systems was studied in embryos, prolar- vae, and small larvae of the sea lamprey using an anti-GABA antibody. Ancillary immunochemi- cal markers, such as proliferating cell nuclear antigen (PCNA), calretinin, and serotonin, as well as general staining methods and semithin sections were used to characterize the territories containing GABA-immunoreactive (GABAir) neurons. Differentiation of GABAir neurons in the diencephalon begins in late embryos, whereas differentiation in the telencephalon and midbrain was delayed to posthatching stages. In lamprey prolarvae, the GABAir populations appear either as compact GABAir cell groups or as neurons interspersed among GABA-negative cells. In the telencephalon of prolarvae, a band of cerebrospinal fluid-contacting (CSF-c) GABAir neurons (septum) was separated from the major GABAir telencephalic band, the striatum (ganglionic eminence) primordium. The striatal primordium appears to give rise to most GABAir neurons observed in the olfactory bulb and striatum of early larval stages. GABAir populations in the dorsal telencephalon appear later, in 15–30-mm-long larvae. In the diencephalon, GABAir neurons appear in embryos, and the larval pattern of GABAir populations is recognizable in prolarvae. A small GABAir cluster consisting mainly of CSF-c neurons was observed in the caudal preoptic area, and a wide band of scattered CSF-c GABAir neurons extended from the preoptic region to the caudal infundibular recess. A mammillary GABAir population was also distinguished. Two compact GABAir clusters, one consisting of CSF-c neurons, were observed in the rostral (ventral) thalamus. In the caudal (dorsal) thalamus, a long band extended throughout the ventral tier. The nucleus of the medial longitudinal fascicle contained an early-appearing GABAir population. The paracommissural pretectum of prolarvae and larvae contained a large group of non-CSF-c GABAir neurons, although it was less compact than those of the thalamus, and a further group was found in the dorsal pretectum. In the midbrain of larvae, several groups of GABAir neurons were observed in the dorsal and ventral tegmentum and in the torus semicircularis. The development of GABAergic populations in the lamprey forebrain was similar Grant sponsor: Xunta de Galicia; Grant number: PGIDT99PXI20003B; Grant number: PGIDT99BIO20002; Grant sponsor: Spanish Science and Technology Ministry; Grant number: BMC2000-0283; Grant sponsor: Uni- versity of Santiago de Compostela. *Correspondence to: Ramo ´n Anado ´n, Department of Fundamental Biol- ogy, Faculty of Biology, University of Santiago de Compostela, 15782 Santiago de Compostela, Spain. E-mail: [email protected] Received 17 August 2001; Revised 19 December 2001; Accepted 18 Jan- uary 2002 DOI 10.1002/cne.10209 Published online the week of April 1, 2002 in Wiley InterScience (www. interscience.wiley.com). THE JOURNAL OF COMPARATIVE NEUROLOGY 446:360 –376 (2002) © 2002 WILEY-LISS, INC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ontogeny of �-AminobutyricAcid-Immunoreactive Neuronal
Populations in the Forebrain andMidbrain of the Sea Lamprey
MIGUEL MELENDEZ-FERRO,1 EMMA PEREZ-COSTAS,1 BEGONA VILLAR-CHEDA,1
XESUS MANOEL ABALO,1 ROLANDO RODRIGUEZ-MUNOZ,2
MARIA CELINA RODICIO,1AND RAMON ANADON1*
1Department of Fundamental Biology, Faculty of Biology, University of Santiago deCompostela, 15782 Santiago de Compostela, Spain
2Department of Biology of Organisms and Systems, Faculty of Biology, University ofOviedo, 33006 Oviedo, Spain
ABSTRACTAlthough brain organization in lampreys is of great interest for understanding evolution in
vertebrates, knowledge of early development is very scarce. Here, the development of the fore-brain and midbrain �-aminobutyric acid (GABA)-ergic systems was studied in embryos, prolar-vae, and small larvae of the sea lamprey using an anti-GABA antibody. Ancillary immunochemi-cal markers, such as proliferating cell nuclear antigen (PCNA), calretinin, and serotonin, as wellas general staining methods and semithin sections were used to characterize the territoriescontaining GABA-immunoreactive (GABAir) neurons. Differentiation of GABAir neurons in thediencephalon begins in late embryos, whereas differentiation in the telencephalon and midbrainwas delayed to posthatching stages. In lamprey prolarvae, the GABAir populations appear eitheras compact GABAir cell groups or as neurons interspersed among GABA-negative cells. In thetelencephalon of prolarvae, a band of cerebrospinal fluid-contacting (CSF-c) GABAir neurons(septum) was separated from the major GABAir telencephalic band, the striatum (ganglioniceminence) primordium. The striatal primordium appears to give rise to most GABAir neuronsobserved in the olfactory bulb and striatum of early larval stages. GABAir populations in thedorsal telencephalon appear later, in 15–30-mm-long larvae. In the diencephalon, GABAirneurons appear in embryos, and the larval pattern of GABAir populations is recognizable inprolarvae. A small GABAir cluster consisting mainly of CSF-c neurons was observed in thecaudal preoptic area, and a wide band of scattered CSF-c GABAir neurons extended from thepreoptic region to the caudal infundibular recess. A mammillary GABAir population was alsodistinguished. Two compact GABAir clusters, one consisting of CSF-c neurons, were observed inthe rostral (ventral) thalamus. In the caudal (dorsal) thalamus, a long band extended throughoutthe ventral tier. The nucleus of the medial longitudinal fascicle contained an early-appearingGABAir population. The paracommissural pretectum of prolarvae and larvae contained a largegroup of non-CSF-c GABAir neurons, although it was less compact than those of the thalamus,and a further group was found in the dorsal pretectum. In the midbrain of larvae, several groupsof GABAir neurons were observed in the dorsal and ventral tegmentum and in the torussemicircularis. The development of GABAergic populations in the lamprey forebrain was similar
Grant sponsor: Xunta de Galicia; Grant number: PGIDT99PXI20003B;Grant number: PGIDT99BIO20002; Grant sponsor: Spanish Science andTechnology Ministry; Grant number: BMC2000-0283; Grant sponsor: Uni-versity of Santiago de Compostela.
*Correspondence to: Ramon Anadon, Department of Fundamental Biol-ogy, Faculty of Biology, University of Santiago de Compostela, 15782Santiago de Compostela, Spain. E-mail: [email protected]
Received 17 August 2001; Revised 19 December 2001; Accepted 18 Jan-uary 2002
DOI 10.1002/cne.10209Published online the week of April 1, 2002 in Wiley InterScience (www.
interscience.wiley.com).
THE JOURNAL OF COMPARATIVE NEUROLOGY 446:360–376 (2002)
© 2002 WILEY-LISS, INC.

to that observed in teleosts and in mouse, suggesting that GABA is a very useful marker forunderstanding evolution of forebrain regions. The possible relation between early GABAergic cellgroups and the regions of the prosomeric map of the lamprey forebrain (Pombal and Puelles[1999] J. Comp. Neurol. 414:391–422) is discussed in view of these results and informationobtained with ancillary markers. J. Comp. Neurol. 446:360–376, 2002. © 2002 Wiley-Liss, Inc.
Indexing terms: development; prosomeres; GABAergic neurons; thalamus; ganglionic eminences;
Agnathans; Petromyzon marinus
�-Aminobutyric acid (GABA) is the main inhibitory neu-rotransmitter in the brain of vertebrates. It is produced byGABAergic neurons from glutamic acid by glutamic aciddecarboxylases (GADs). The few developmental studies onthis subject have found that GABA and GADs are amongthe first neurotransmitters and neurotransmitter-synthesizing enzymes produced by the vertebrate brain(Roberts et al., 1987; Aoki et al., 1989; Ekstrom and Ohlin,1995; Barale et al., 1996; Obata, 1997; Katarova et al.,2000), and this is also true for the retina (Cherubini et al.,1991; Messersmith and Redburn, 1993; Mitchell and Red-burn, 1996). Moreover, developmental studies in teleostsand mammals have found that GABA-immunoreactive(GABAir) or GAD gene-expressing neurons are distrib-uted according to segmental patterns (Ekstrom and Ohlin,1995; Katarova et al., 2000). Although these studies sug-gest that early GABAergic populations may be used incomparative studies for understanding brain organiza-tion, existing evidence is scant, and any generalization ofthis idea would require information based on studies inmore groups of vertebrates.
The presence of GABAergic neurons in some regions ofthe brain of large larvae and adult lampreys has alreadybeen noted (Rio et al., 1993; Pombal et al., 1997, 1999;Anadon et al., 1998a; Pombal and Puelles, 1999; Yanez etal., 1999; Melendez-Ferro et al., 2000, 2001). Knowledge ofthe early organization of the lamprey GABAergic systemsmay be valuable for understanding the evolution of thesesystems in vertebrate brains. However, there have been
no previous studies of the appearance of GABAergic struc-tures in the brain of lampreys, nor systematic studies offorebrain and midbrain GABAergic populations.
Lampreys belong to an ancient lineage of vertebratesthat flourished before the appearance of the gnathos-tomes. Here, we report the development of GABAergicsystems in the forebrain and midbrain of the sea lamprey.For defining GABAir populations during early larvalstages, we also made use of immunoreactivities to prolif-erating cell nuclear antigen (PCNA), serotonin, and cal-retinin (CR).
MATERIALS AND METHODS
Animals
Embryos (E10, E11, E13; n � 6), prolarvae (P0, P1, P2,P3, P4, P5, P6, P7, P10, P15, P23; n � 22), and larvae(9–40 mm in length; n � 28) of the sea lamprey (Petro-myzon marinus L.) were used. Embryos and prolarvaeoriginated from “in vitro”-fertilized eggs obtained fromsexually mature adult lampreys caught in the River Sella(northern Spain). Fertilized eggs were reared in the labo-ratory under appropriate conditions of darkness and tem-perature (16°C; Rodrıguez-Munoz et al., 2001). Larvallampreys were caught in the River Ulla (northwesternSpain) and kept in an aerated aquarium until processing.All experiments were conducted in accordance with Euro-pean Community guidelines on animal care and experi-mentation to minimize pain and discomfort.
Abbreviations
bu oral cavitycp pretectal nucleus of the posterior commissurecth caudal thalamic groupdi diencephalondp dorsal pretectal groupe eyeear otic vesiclefr fasciculus retroflexusha habenulaIII GABAir internuclear oculomotor neuronsir caudal infundibular recessist isthmuslip preoral liplp lateral palliumlpp lateral pallium primordiumlt lamina terminalism3 third midbrain Muller cellM5 Schober’s M5 nucleusma mammillary regionmc mitral cellsmch midbrain roof choroid plexusme mesencephalonmpp medial pallium primordiumnflm nucleus of the medial longitudinal fasciclenh naso-hypophysary canal
not notochordnpo “magnocellular” preoptic nucleusob olfactory bulbop olfactory placodep pineal organp1–5 prosomeres 1–5pc pretectal nucleus of the posterior commissureph pharynxphp1–3 pharyngeal pouches 1–3po posterior preoptic nucleuspoc postoptic commissurepp parapineal organrho rhombencephalonrpo preoptic recessrth rostral thalamic groupsc spinal cordsh subhippocampal lobesp septumstr striatumte telencephalontp von Kuppfer’s posterior tubercletu postoptic commissure-tuberal groupve oral velumVg trigeminal nerve ganglionzl zona limitans intrathalamica
361DEVELOPMENT OF GABA IN LAMPREY FOREBRAIN

Stages of embryos, prolarvae, and early larvae are indi-cated by age (e.g., E12 indicates 12 days after fertilization;P3 indicates 3 days posthatching, and so on; in our broodshatching occurred 11–13 days after fertilization). In addi-tion, to classify prolarvae, we used the stages defined forsea lamprey by Piavis (1971): hatching (P0–1), pigmenta-tion (P2–3), gill-cleft (P4–7), and burrowing (P8–23)stages. Larval stages (with the exception of larvae frombroods, in which age is known) are categorized simply bytotal body length.
GABA immunohistochemistry
The specimens were fixed after deep anesthesia with0.1% tricaine methane sulfonate (MS-222; Sigma, St.Louis, MO). Animals (late embryos, prolarvae, and 9–40mm larvae) were fixed in cold (4°C), freshly prepared 5%glutaraldehyde in 0.1 M phosphate buffer, pH 7.4 (PB),overnight. After rinsing in PB, samples were cryopro-tected with 30% sucrose in PB, embedded in Tissue Tek(Sakura, Torrance, CA), and frozen using liquid nitrogen-cooled isopentane. Transverse, sagittal, or horizontal sec-tions (10–12 �m thick) were cut on a cryostat, andmounted on gelatin-subbed slides. The sections were pro-cessed for immunocytochemistry as follows: Nonspecificbinding sites were blocked by incubation with 10% normalgoat serum (Vector Laboratories, Burlingame, CA). Thesections were then incubated in a humid chamber at roomtemperature with a polyclonal anti-GABA antibody (Af-finiti, Mamhead, United Kingdom), diluted 1:1,000, over-night. After rinsing in Tris-buffered saline (TBS; pH 7.4),the sections were incubated with a goat anti-rabbit immu-noglobulin (Sigma), diluted 1:100, for 1 hour, rinsed twicein TBS, and incubated in rabbit peroxidase-antiperoxidase (PAP; Sigma; diluted 1:400). The sectionswere developed using 0.6 mg/ml 3-3�-diaminobenzidine(DAB; Sigma) and 0.003% H2O2. All antibody dilutionswere made in TBS containing 0.2% Triton X-100 as deter-gent and 3% normal goat serum. After development, thesections were rinsed in TBS, dehydrated, and cover-slipped.
Other immunocytochemical markers
For PCNA, CR, and serotonin immunocytochemistry,some animals (late embryos, prolarvae, and 9–40 mmlarvae for PCNA; 30 mm larvae for CR; prolarvae and9–40 mm larvae for serotonin) were fixed in 4% parafor-maldehyde in PB (CR, serotonin) or Bouin’s fluid (PCNA).For PCNA or serotonin immunocytochemistry, the ani-mals were embedded in paraffin wax, cut in transversal orsagittal planes on a rotary microtome (8–10 �m thick-ness), and mounted on gelatin-subbed slides. Dewaxedsections were then immunostained by a procedure similarto that used for GABA, using either a polyclonal antisero-tonin antibody (Incstar, Stillwater, MN; diluted 1:1,000)or a mouse monoclonal anti-PCNA antibody (clone PC10;Sigma; diluted 1:4,000) overnight. After rinsing inphosphate-buffered saline (PBS; pH 7.4), sections wereincubated with goat anti-rabbit (Sigma) or anti-mouse(Dako, Glostrup, Denmark) immunoglobulin, diluted1:100 or 1:50, respectively, for 1 hour and developed withDAB and H2O2 as described above. For CR immunohisto-chemistry, larvae were cryoprotected, embedded, frozen,and cut on a cryostat in either transverse or sagittalplanes, as for GABA. Immunocytochemistry was per-formed as described above using a polyclonal anti-CR an-
tibody (Chemicon, Temecula, CA) diluted 1:1,000 over-night.
The specificity of the GABA, CR, serotonin, and PCNAantibodies has been tested by the suppliers. As a furthercontrol, some sections were processed without incubatingwith the primary antibody, or the primary antiserum wasreplaced by normal serum. In these sections, no immuno-staining was observed.
General stains and semithin sections
For topographic purposes, other prolarvae and smalllarvae were fixed in 1% paraformaldehyde and 1% glutar-aldehyde in PB, rinsed in PB, postfixed in 1% OsO4 in 0.1M cacodylate buffer (pH 7.4), dehydrated with acetone,and embedded in Spurr resin. Semithin sagittal sections(1 �m thick) were cut on an ultramicrotome and stainedwith a 1% toluidine blue-1% borax solution in water. Se-ries of hematoxylin-eosin-stained transverse and sagittalsections of prolarvae and larvae from our collection werealso at our disposal.
Photography and graphic reconstruction
Photomicrographs were taken with an Olympus DP-10color digital camera. The images were converted to a grayscale and adjusted for brightness and contrast using CorelPhoto-Paint 8. Photomontage and lettering were done us-ing Corel Draw 8.
To overcome mislocalization of GABAir structures ow-ing to the change of relative positions of territories ingrowing brains, we have made graphic reconstructions ofbrains using photomicrographs of complete series of trans-verse sections. The sections were drawn as equidistantparallel lines on brain profiles obtained from photomicro-graphs of sagittal sections of heads of lampreys of thesame age, and the GABAir regions represented on thebasis of distances to the top or bottom of each brain sec-tion. Reconstructions were also made for the ancillaryimmunohistochemical markers in 30-mm-long larvae fordefining their localization.
RESULTS
Landmarks in the forebrain and boundariesbetween major regions during lamprey
development
Knowledge of the early development of the lampreybrain is mainly based on the studies of von Kupffer (1906),Bergqvist (1932), and Kuratani et al. (1998). Here, wepresent a brief description of the main commissures andother landmarks visible in brains of prolarvae and earlylarvae useful for analysis of forebrain GABAergic popula-tions. A photograph of the head and a scheme of the brainof sea lamprey prolarvae are shown in Figure 1A,B.
In sagittal semithin sections of the brain of P7–17 pro-larvae (see Fig. 1C), the clearest limiting structure ob-served in the brain was a deep transverse groove and athin glial cell lamina separating the hindbrain and themidbrain, corresponding to the posterior intraencephalicsulcus of von Kupffer (1906). The third Muller cell (M3)was closely associated with this lamina. Caudally to thisgroove, the floor plate was rather thin and formed a char-acteristic midline glial septum. Rostrally to this groove,the floor plate contained a large commissural (“tegmen-tal”) region that extended over the notochord to a special-ized bulge of the ventral ventricular region (“posterior
362 M. MELENDEZ-FERRO ET AL.

Fig. 1. A: Photomicrograph of the head of a P7 prolarva showingthe location of some external landmarks 1–4, Pharyngeal pouches1–4. B: Schematic drawing of the brain and adjacent structures of aP10 prolarva. C: Schematic side view drawing of the fore- and mid-brain of a P7 prolarva showing the main landmarks used in thisstudy. Arrow, posterior commissure; star, pineal organ. Brokenstraight lines indicate the approximate planes of F–H; the dotted linesindicate the segmental limits. Rostral is to the left. D: Hematoxylin-eosin-stained sagittal section of the brain of a 18 mm larva showingthe different commissures and landmarks in the midline. The dottedlines indicate the segmental limits. Single arrow, posterior commis-sure; double arrow, habenular commissure; star, pineal organ. E: Sag-
ittal section of the brain of a P12 prolarva showing the appearance ofPCNA immunoreactivity in the ventricular zone of different regions.The arrowhead points to the PCNA-negative discontinuity in thep3–p2 boundary. Other symbols as in D. F–H: Semihorizontal sec-tions through the brain of a P12 prolarva showing clear differencesamong the ventricular zones of the different regions. The PCNA-negative discontinuities (arrowheads) between the striatum and theprimordium of the telencephalic hemispheres and between p3 and p2are indicated in F, that of the posterior tubercle in G, and that in thewalls of the caudal infundibular recess in H. For abbreviations, seelist. Scale bars � 100 �m in D, 50 �m in E–H.

tubercle” of von Kupffer, 1906). The median line exhibitedonly cycling cells, as indicated by their immunoreactivityto PCNA, which is in contrast with the PCNA-negativeventral midline glial septum of the hindbrain. Rostrally,the “posterior tubercle” was limited by the posterior in-fundibular recess that covered the tip of the notochord.The infundibular recess was covered with a dense PCNAirependymal layer that extended to a well-developed com-missural crest located transversely between the preopticrecess and the infundibulum. This commissural crest(“postoptic commissure” plate) was prominent at thesestages, despite the fact that the optic nerve and tract werepractically undeveloped. In the rostral part of the fore-brain, a deep sulcus (“anterior intraencephalic sulcus” ofvon Kupffer, 1906) separated the telencephalon from thediencephalon. The small roof region between the habenulaand the telencephalon consisted of a thin epithelial layerthat gave rise to the pineal and parapineal vesicles. Ahabenular commissure was present in the lamprey habe-nula that represents a large commissure for telencephalicaxons (Northcutt and Puzdrowsky, 1988; Yanez and Ana-don, 1994). A deep subhabenular sulcus separated thehabenula from thalamic-pretectal regions. Behind the ha-benula, the posterior commissure was prominent. Allthese landmarks were also visible in larvae (Fig. 1D).
PCNA immunoreactivity was of little help for distin-guishing between forebrain regions in embryos and earlyprolarvae. However, in the prosencephalon of later prolar-vae and small larvae, small PCNA-negative discontinui-ties in the ventricular zone and differences in the thick-ness and appearance of the PCNAir ventricular zonebetween different territories were observed, facilitatinganalysis of territories (Fig. 1E–H). Just below the habe-nula (Fig. 1E,F), a thin PCNA-negative ventricular bandseparated the dorsal and ventral thalamus primordia(p2–p3 interprosomeric boundary; see Pombal andPuelles, 1999). This band appears to extend toward aPCNA-negative area of the posterior tubercle (Fig. 1G). Inthe telencephalon, a PCNA-negative band separated thestriatum from the primordium of the telencephalic hemi-spheres (Fig. 1F). A thin PCNA-negative ventricular bandalso separated two unequal PCNAir ventricular zones inthe wall of the caudal infundibular recess, a continuouslateroventral oriented (tuberal) area and a little caudodor-sal (mammillary) area that is close to the tip of the noto-chord (Fig. 1H). This posterior tubercle discontinuity canbe followed toward the thalamus, but its end could not beassessed. No sharp limit between the pretectal region andthe midbrain could be traced with PCNA immunocyto-chemistry.
GABA distribution in the forebrain andmidbrain of late embryos and early
prolarvae
No GABAir neurons were observed in the forebrain ormidbrain of E10 embryos. A few GABAir neurons firstappeared in two locations of the basal diencephalon of E11embryos. The most rostral were cerebrospinal fluid-contacting (CSF-c) neurons, whereas, in the caudal dien-cephalon, the GABAir neurons occupy the outer part of theperiventricular mantle layer (Fig. 2A,B). In E13 embryosor P0 prolarvae (hatchlings), the number of GABAir neu-rons increased in the diencephalon relative to the numberof neurons in E11, forming two small groups, one in thepostoptic commissure-tuberal region and one in the pre-
tectal tegmentum (nucleus of the medial longitudinal fas-cicle; Fig. 2C,D). The rostral group consisted of CSF-cneurons. A few GABAir fibers were observed in the pri-mordial marginal layer of the diencephalon. In P1 prolar-vae, a few GABAir neurons also appeared in a band inter-mediate between the two early GABAir groups, which hadincreased their number of cells, and GABAir fibers crossedin the posterior commissure of P2 prolarvae.
In the diencephalon of P3 prolarvae, the number ofGABAir groups increased from two to six. In the postopticcommissure-tuberal region (hypothalamus), a large num-ber of CSF-c neurons extended in the wall, except in themost caudoventral zone of the infundibulum. Associatedwith the dorsocaudal part of this group, a rostral (ventral)thalamic GABAir group was distinguishable. In the cau-dal region, four GABAir groups of non-CSF-c neuronswere distinguished: caudal (dorsal) thalamus, nucleus ofthe posterior commissure, dorsal pretectal tegmentum,and nucleus of the medial longitudinal fascicle (Fig. 2E–H). Among these, the nucleus of the medial longitudinalfascicle exhibited a comparatively large number ofGABAir neurons. In the telencephalon, the first GABAircell group was observed in P4 prolarvae in the region ofthe terminal lamina, rostral to the preoptic recess. Itconsisted of scattered CSF-c neurons (Fig. 3A).
In P6–7 prolarvae, the GABAir groups of the postopticcommissure-tuberal region, rostral (ventral) thalamic,caudal (dorsal) thalamus, nucleus of the posterior commis-sure, dorsal pretectal tegmentum, and nucleus of the me-dial longitudinal fascicle were rather well separated (Fig.3B–G), and the marginal fiber layer between these groupsand the posterior commissure contained numerousGABAir fibers (Fig. 3B,E,F). The caudal end of the infun-dibular recess did not contain GABAir neurons (Fig. 3F).A graphic reconstruction of the GABAir cell groups ob-served in the forebrain of P7 prolarvae is shown in Fig-ure 4.
GABA distribution in late prolarvaeand larvae
In different forebrain and midbrain regions of late pro-larvae and early larvae, GABA immunoreactivity ap-peared either in compact areas of GABAir neurons or inwell-defined regions where GABAir neurons were inter-spersed among GABA-negative cells. In some areas (sep-tum, thalamic eminence, preoptic-tuberal, and mammill-ary regions and M5 nucleus of Schober), the GABAirneurons exhibited CSF-c dendrites (CSF-c neurons). TheCSF-c character was consistently conserved by many neu-rons of these regions from developing stages to adulthood(unpublished results). In most other regions, during earlystages of development, the GABAir neurons formed com-pact groups that were clearly different from GABA-negative adjacent regions. In some areas, such as thelateral telencephalic hemispheres and the pretectal nu-cleus of the posterior commissure, GABAir neurons didnot form compact clusters. A graphic reconstruction of theregions containing GABAir neurons in 18–30 mm larvaeis shown in Figure 7A.
Telencephalon. In early larvae, the number ofGABAir neurons in the region of the terminal laminaincreased considerably from the early prolarval stages,but these neurons conserved a close association with theterminal lamina during the larval period (Fig. 5C,E) andin adults (unpublished results). In view of its position inadults, this GABAir cell group may correspond to region
364 M. MELENDEZ-FERRO ET AL.

T4 (anterior commissure nucleus) of Schober (1964) and tothe paraterminal preoptic and neuroporic recess nuclei ofPombal and Puelles (1999).
At the transition to the larval period (P16), in additionto the paraterminal CSF-c group, the telencephalon exhib-ited a continuous cluster of non-CSF-c GABAir neurons inthe primordium of the striatum-olfactory bulbs. This clus-ter gave rise to the GABAir granule cells of the olfactorybulb of early larvae (P26–54), which exhibited numerous
GABAir processes in superficial areas surrounding smallgroups of GABA-negative neurons (very likely mitral cells;Fig. 5A,B). In the caudal region (striatum), the GABAirneurons formed a band close to the ventricular layer (Fig.5C,D). No migrated GABAir neurons could be observed inthe lateral pallium primordium of these early larvae (Fig.5C).
The most obvious change in the telencephalon of 15–30mm larvae with respect to early larvae was the appear-
Fig. 2. Sections of the forebrain of E11 embryos (A,B), hatchling(C,D), and P3 prolarvae (E–H) showing GABAir neurons. A,C: GABAirCSF-c neurons of the primordium of the postoptic commissure-tuberalregion. B,D: GABAir neuron of the nucleus of the medial longitudinalfascicle. E–G: Different sagittal sections of the same prolarva from lat-erally (E) to more medially (G), showing main GABAir groups.H: Oblique section passing through the plane indicated in E. Note the
CSF-c processes of neurons of the tuberal group. Dotted lines indicate theisthmomesencephalic and the telencephelic-diencephalic boundaries.Solid arrow, posterior commissure; arrowheads, CSF-c processes; openarrow, GABAir fibers; star, pineal organ. For abbreviations, see list.A–D: Semihorizontal sections; E–G: sagittal sections. Scale bars � 25 �min A,B,H, 10 �m in C,D, and 50 �m in E–G.
365DEVELOPMENT OF GABA IN LAMPREY FOREBRAIN
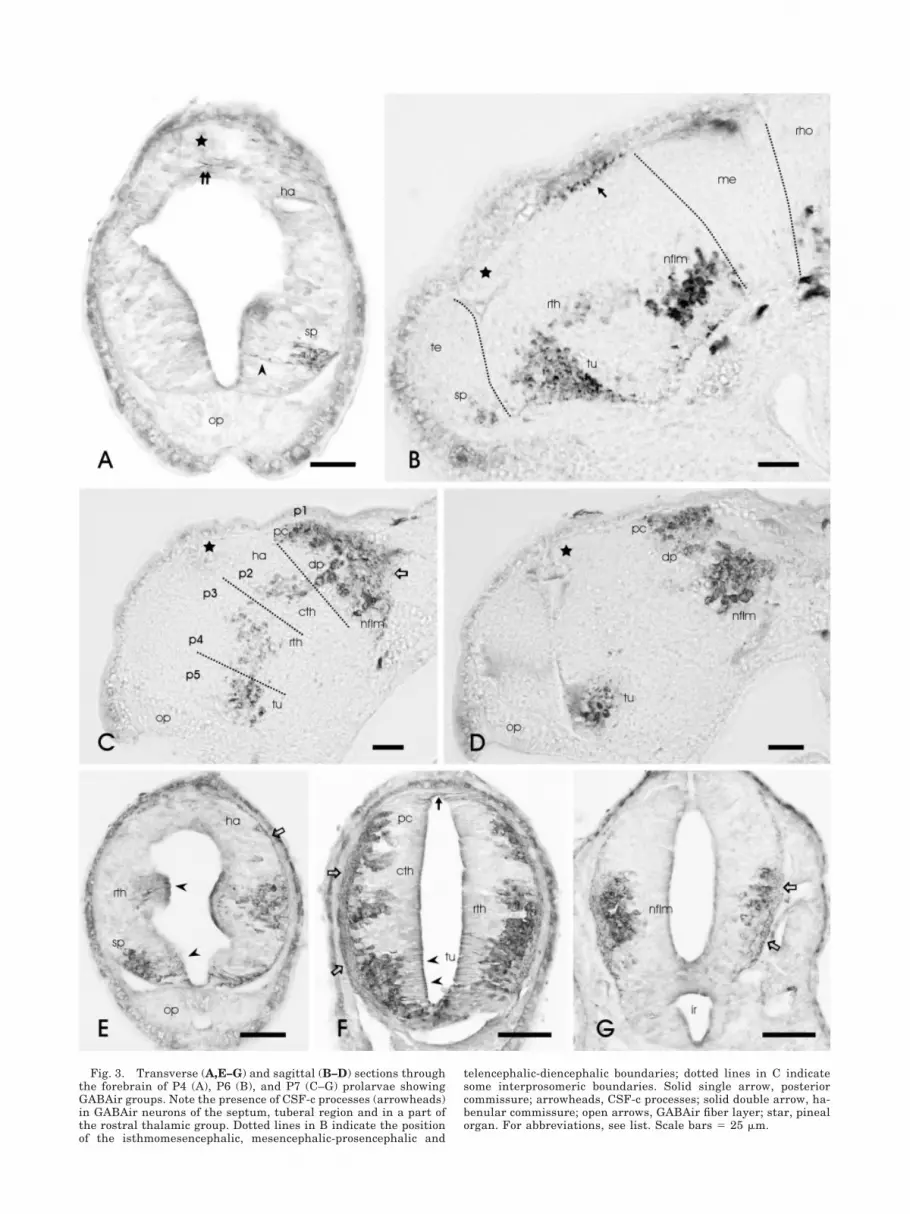
Fig. 3. Transverse (A,E–G) and sagittal (B–D) sections throughthe forebrain of P4 (A), P6 (B), and P7 (C–G) prolarvae showingGABAir groups. Note the presence of CSF-c processes (arrowheads)in GABAir neurons of the septum, tuberal region and in a part ofthe rostral thalamic group. Dotted lines in B indicate the positionof the isthmomesencephalic, mesencephalic-prosencephalic and
telencephalic-diencephalic boundaries; dotted lines in C indicatesome interprosomeric boundaries. Solid single arrow, posteriorcommissure; arrowheads, CSF-c processes; solid double arrow, ha-benular commissure; open arrows, GABAir fiber layer; star, pinealorgan. For abbreviations, see list. Scale bars � 25 �m.

ance of the telencephalic hemispheres, which now con-tained a scattered GABAir neuronal population migratedwithin a rather thickened cortical region (lateral pallium).These neurons were distributed among numerous GABA-negative cells in a cell and fibrous layer containing a largenumber of GABAir fibers (Fig. 5E,F). In 15 mm larvae, adeep intratelencephalic sulcus marked the origin of thetelencephalic lateral ventricles, which were well formed in30 mm larvae (Fig. 5E). The thickened periventricularregion of the telencephalic hemispheres of these larvaewas devoid of both GABAir perikarya and GABAir fibers.The olfactory bulb was by this time widely separated fromthe striatal GABAir cluster and contained numeroussmall GABAir neurons and a dense GABAir neuropil (Fig.5G). These small neurons contrast with the larger GABA-negative mitral cells. As the telencephalic hemispheresgrew, the striatum progressively became a smaller portionof the telencephalon, characterized by the presence of alayer of GABAir neurons rather close to the ventricle.During development, the region of the subhippocampallobe was characteristically devoid of GABAir perikarya(Fig. 5D,F), although it received GABAir fibers. In thelarval period studied (12–40 mm), the medial palliumprimordium was only a small GABA-negative region ros-tral to the habenula (Fig. 5F).
Diencephalon and dorsal pretectum. In the dien-cephalon of P10 prolarvae, a considerable number ofGABAir neurons could be observed in the hypothalamic,thalamic, and pretectal groups. The rostral thalamicGABAir group was better delimited from the large postop-tic commissure-tuberal GABAir group than in P4 prolar-vae and occupied a characteristic dorsorostral promi-nence, with only a small number of CSF-c neurons. Thecaudal thalamic group was clearly demarcated from thoseof the rostral thalamus and pretectum by bands free ofGABAir neurons. The posterior, habenular, and tegmen-tal commissures contained scarce GABAir fibers, whereasthe postoptic commissure was the most prominent. Themarginal region of the diencephalon exhibited a largenumber of GABAir processes, although such processeswere very scarce in the telencephalon at this stage.
In the diencephalon of early larvae (P26–54), the orga-nization of the GABAir groups was similar to that ob-served in later larval stages. From the caudal part of thepreoptic recess to the infundibular recess, an extensivearea with CSF-c GABAir neurons was observed (Fig. 6A–G). The appearance of this posterior preoptic-tuberal areawith GABA immunohistochemistry was rather homoge-neous, although in retrocommissural regions large non-CSF-c GABAir neurons could be observed in addition tosmaller bipolar CSF-c neurons (Fig. 6E,F). In the cau-dodorsal part of the mammillary recess, some CSF-cGABAir neurons could be observed forming a separategroup (Fig. 6H). The rostral thalamic GABAir groupshowed two parts, a ventral part with CSF-c neurons anda dorsal part with non-CSF-c neurons (Fig. 6A–C). Thegroup of CSF-c neurons became contiguous with the cau-dal preoptic-tuberal region (Fig. 6C). The caudal (dorsal)thalamic group formed an oblique band (Fig. 6A–F) thatcrossed the line of the fasciculus retroflexus. A wideGABA-negative region (“posterior tubercle”) caudally sep-arated the thalamohypothalamic GABAir groups fromthose of the pretectal-midbrain tegmentum (Fig. 6F–H).The pretectal nucleus of the posterior commissure formeda distinctive GABAir group (Fig. 6B–E). Unlike the tha-lamic GABAir bands, which contained closely groupedGABAir neurons, the GABAir neurons of the pretectalnucleus of the posterior commissure were intermingledwith GABA-negative cells.
Graphic reconstructions (Fig. 7A) and series of longitu-dinal sections allowed us to study the relationship be-tween the GABAir populations of the diencephalon andlandmarks in the ventricular surface and the fasciculusretroflexus in 30 mm larvae. The posterior preoptic-tuberal and rostral thalamus populations of CSF-cGABAir neurons become largely confluent. The rostral(dorsal) limits of these two populations were the GABA-negative magnocellular preoptic nucleus and the subhip-pocampal lobe, respectively. Moreover, the band of CSF-cGABAir neurons of the rostral (ventral) thalamus waspractically continuous through the dorsal tuberal regionwith those of the infundibular recess, lying rostroventrallyto Bergquist’s ventral diencephalic sulcus. The mammil-lary group of GABAir neurons is not a continuation withthis tuberal GABAir band. The caudal (dorsal) thalamicband of GABAir neurons showed a trajectory that ob-liquely crossed the fasciculus retroflexus (Fig. 7A,B), ex-tending far beyond it. In 30 mm larvae, this fascicle wasconspicuous and could be traced to the rostral part of thecommissural plate of the midbrain-diencephalon, just lat-eral to von Kupffer’s “posterior tubercle” (1906). Betweenthe habenula and this caudal thalamic GABAir band wasa wide GABA-negative dorsal thalamic region (Fig. 7A,B).This negative region had widened considerably with re-spect to that in early larvae (P26–54). A deep ventriculargroove caudal to the caudal thalamic GABAir band ap-peared as a double-flexured line, being practically verticalbetween the posterior tubercle GABA-negative region andthe ventral tegmentum. A GABA-negative transverse gapapparently marked the postulated p1–p2 interprosomericboundary (stars in Fig. 7B). No clear limit could be seenbetween the dorsal pretectal and the midbrain tegmentalgroups of GABAir neurons.
Pretectal tegmentum and midbrain. In the tegmen-tum of the pretectal region and in the midbrain of earlylarvae (P26–54), several groups of GABAir neurons can berecognized caudally to the negative “posterior tubercle.”
Fig. 4. Schematic drawing showing the distribution of GABAirgroups in the forebrain of a P7 prolarva. Same symbols as in Figure1A. For abbreviations, see list.
367DEVELOPMENT OF GABA IN LAMPREY FOREBRAIN

Ventrally in the tegmentum, a compact rostral group ofGABAir neurons was associated with larger GABA-negative cells that corresponds to the nucleus of the me-dial longitudinal fasciculus (Fig. 6H). A somewhat lesscompact group (dorsal pretectal nucleus) was located dor-sally to it (Fig. 6H), associated caudally with a few large
GABA-negative neurons, including the first Muller cell. Inthe ventral midbrain tegmentum, at least two GABAirgroups were present, a ventral group closely associatedwith the oculomotor nucleus and the third Muller cell(themselves GABA-negative) as well as a dorsal group(M5). Some GABAir neurons were also observed in the
Fig. 5. Transverse sections through the telencephalon of a P39(9–10 mm) larva (A–D) and a 30 mm larva (E,F) showing GABAirpopulations. A–D represent alternate sections of the telencephalon,from rostrally to caudally. Arrow in E points to the lateral telence-
phalic recess. H: Sagittal section through the telencephalon of a 30mm larva showing the GABAir neurons of the olfactory bulb. Notethat mitral cells are GABA-negative. For abbreviations, see list. Scalebars � 50 �m.
368 M. MELENDEZ-FERRO ET AL.

torus semicircularis (Fig. 6H). The limits between thedorsal groups of the pretectum and midbrain, and betweenthe ventral groups, were not clear.
In larger larvae (15–30 mm in length), rather scatteredGABAir neurons were observed in a midbrain longitudinalband corresponding to the torus semicircularis. The celllayers of the torus semicircularis were associated with aconspicuous GABAir neuropil extending from caudally tothe pretectal nucleus of the posterior commissure to the
isthmus. The dorsal tegmental nucleus of these larvaealso exhibited a band of CSF-c GABAir neurons, corre-sponding to the retinopetal M5 nucleus, which is known tobe GABAergic (see Rodicio et al., 1995; Anadon et al.,1998a). The optic tectum, located dorsally to the torus,lacked GABAir neurons in these stages, although in largelarvae it exhibited a GABAir tectal population (unpub-lished results). On the other hand, the midbrain GABAirpopulations were separated from the conspicuous isthmic
Fig. 6. A–H: Transverse sections through the diencephalon andpretectum of a P39 larva showing the distribution of the differentGABAir groups. Note in C the confluence between the rostral tha-lamic and preoptotuberal groups. Arrowheads point to groups withCSF-c neurons; thin arrows point to the separation between the
rostral and the caudal thalamic GABAir groups. Black star, “posteriortubercle” GABA-negative region; white star, dorsal portion of therostral thalamic group. For abbreviations, see list. Scale bars � 25�m.
369DEVELOPMENT OF GABA IN LAMPREY FOREBRAIN

GABAir populations by a band free of GABAir neurons.The distribution of these GABAir populations is shown inFigure 7A.
Distribution of ancillary immunochemicalmarkers during early development
Serotonin immunoreactivity was observed in the tha-lamic and infundibular regions of early larvae (Yanez,1992). In the thalamus of small larvae, a short row ofserotonin-ir neurons appeared in the thalamus, forming athin band parallel to the fasciculus retroflexus (Yanez,1992; present results). In the 30 mm larvae, this band waslocated approximately at the rostral limit of the caudal(dorsal) thalamic GABAir band (Fig. 8A). According toPombal and Puelles (1999), this band of non-CSF-cserotonin-ir neurons corresponds to the zona limitans in-trathalamica. Close examination of the forebrain of a 30mm larvae indicated that serotonin-ir infundibular neu-rons formed a CSF-c band that was coextensive with thedorsocaudal tuberal band containing GABAir CSF-c neu-rons (Fig. 8B,C). Unlike the GABAir population, thisserotonin-ir population does not extend to the rostral thal-amus. A second serotonin-ir group in the caudal infundib-ular (mammillary) recess was also coextensive with thecaudal hypothalamic group of GABAir neurons (Fig. 8D).
CR immunoreactivity has been used previously for iden-tification of thalamic territories in adult lamprey (Pombal
and Puelles, 1999). To clarify the relationship betweenGABA-containing populations and the thalamic domainsdefined by Pombal and Puelles, we examined CR distribu-tion in the diencephalon of 30 mm larvae. CRir neuronswere numerous in the forebrain and midbrain of theselarvae and in many regions formed coherent groups (Fig.8E,F,H–J). Moreover, the distributions of CRir neuronsand GABAir neurons in the thalamus appeared to becomplementary, the GABAir groups being CR-negativeand the CRir groups being GABA-negative (see Fig.8G,H).
The preoptic recess nucleus possessed GABAir neuronsbut lacked CRir cells (Fig. 8E). The subhippocampal lobeexhibited a number of CRir perikarya, although noGABAir cell bodies were observed (Fig. 8E). In the tha-lamic eminence (rostral ventral thalamus), a band of CRirneurons was observed just dorsal to the GABAir popula-tion. The tuberal region contained a rich CRir populationcoextensive with the region of scattered CSF-c GABAirneurons (Fig. 8F). In the habenula, a ventrocaudal CRirpopulation was observed. In the caudal (dorsal) thalamus,numerous CRir neurons formed a band just dorsal andparallel to that of GABAir neurons, crossing and extend-ing beyond the fasciculus retroflexus (Fig. 8F,H). In trans-verse sections through the pretectal commissural region, asmall group of CRir neurons appeared in a subventricularlocation, medial to the area containing GABAir neurons(Fig. 8H). A numerous CRir population extended through-out a GABA-negative territory to the posterior tubercle,joining the contralateral population and forming an un-even band below the floor of the mesencephalic ventricle(Fig. 8I,J).
DISCUSSION
In the present study, the temporal sequence and spatialdistribution of GABA immunoreactivity during develop-ment of the forebrain and midbrain is described for thefirst time for lampreys. The distribution of GABAir neu-rons observed in medium-sized (present results) and large(unpublished observations) larval lampreys suggests thatGABA immunoreactivity in early-differentiated GABAer-gic populations shows a consistent pattern throughout thelarval period. This indicates that immunoreactivity toGABA is useful for characterizing lamprey brain regions,as suggested by Ekstrom and Ohlin (1995) for a teleost.Moreover, the present results are in agreement with theidea that GABA is one of the first neurotransmitters toappear in the brain (Roberts et al., 1987; Aoki et al., 1989;Ekstrom and Ohlin, 1995). The early development ofGABAir neurons and the varied GABAergic populationsobserved in this study indicate that GABA may play arelevant role in brain development. Although the inhibi-tory functions of GABA have been well studied in thespinal cord of adult lamprey (see Christenson et al.,1991a,b), studies in other vertebrates also indicate thatGABA may have trophic functions and be excitatory dur-ing development (Madtes and Redburn, 1983; Cherubiniet al., 1991; LoTurco et al., 1995; Barker et al., 1998). Therich GABAir innervation observed in most parts of thedeveloping lamprey brain points to an early appearance ofinhibitory circuits in most forebrain and midbrain regions.
Our results with the anti-GABA antibody indicate thatonly cells with neuronal morphology and fibers were la-beled above background in the brain of the lamprey at thestages studied. Although the astroglia of the optic nerve of
Fig. 7. A: Schematic drawing showing the distribution of GABAircell groups in the fore- and midbrain of a 30 mm larva. B: Sagittalsection through the caudal thalamic GABAir group of a 30 mm larvashowing its relative position with respect to the right fasciculus ret-roflexus. The pretectal nucleus of the posterior commissure and thenucleus of the medial longitudinal fascicle are also shown in thissection. Note the presence of large GABA-negative neurons in thelatter nucleus. Stars indicate the GABA-negative cell band locatedbetween p2 and p1. For abbreviations, see list. Scale bar � 50 �m.
370 M. MELENDEZ-FERRO ET AL.

Fig. 8. Transverse sections through the brain of 30 mm larvaeshowing distribution of immunoreactivities to serotonin (A–D), cal-retinin (E,F,H–J), and GABA (G). Note the presence of serotonin-irneurons (thin solid arrows) in the zona limitans intrathalamica (A),dorsal tuberal region (B,C), and dorsal region of the mammillaryrecess (D). Arrowheads in B and C point to Bergquist’s ventral dien-
cephalic sulcus. Note also the numerous calretinin-ir neurons in thesubhippocampal lobe, dorsal thalamus, and posterior tubercle(E,F,H–J). In the pair in G and H, note the vicarious immunoreactiv-ity of thalamic nuclei to GABA and calretinin. Thick solid arrows,calretinin-ir band of the dorsal thalamus; open arrows, fasciculusretroflexus. For abbreviations, see list. Scale bars � 25 �m.

rat exhibit transient GABA immunoreactivity (Sakataniet al., 1992), GABAir cells were not observed in the opticnerve of prolarval or larval lampreys. The brain of larvallampreys, on the other hand, lacks blood vessels and,hence, GABA immunoreactive structures associated withbrain capillaries, as reported for mammals (Gragera et al.,1993; Benagiano et al., 2000).
Earliest prosencephalic GABAir populations
In many respects, the early development of the GABAirsystems of the lamprey brain was similar to that observedin a teleost (Gasterosteus aculeatus: Ekstrom and Ohlin,1995). As reported for Gasterosteus (Ekstrom et al., 1985;Ekstrom and Ohlin, 1995), GABA immunoreactivity ap-peared much earlier in lamprey than other early markers,such as serotonin (present results) or tyrosine hydroxylase(unpublished observations). Moreover, as in Gasterosteusand zebrafish embryos (Ekstrom and Ohlin, 1995; Martinet al., 1998; Doldan et al., 1999), GABA immunoreactivityappeared in discrete groups of neurons throughout thebrain. The presence of compact groups of neurons express-ing glutamic acid decarboxylase 65 (GAD65) and/or 67(GAD67) genes was also observed in the developing mousebrain by in situ hybridization (Katarova et al., 2000).Interestingly, the early pattern of GAD gene expressionobserved in the mouse prosencephalon is notably similarto that of prosencephalic GABAir neurons in lampreys andteleosts.
The two earliest GABAir groups of the lamprey prosen-cephalon were located in the rostral diencephalon andtegmental pretectum. The first group, which enlargedearly in development and became closely associated withthalamic populations, may correspond generically to the“nucleus of the tractus of the postoptic commissure” re-ported for teleost embryos (Ekstrom and Ohlin, 1995;Martin et al., 1998; Doldan et al., 1999). Alternatively, itmight correspond to the GABAir “optic tract neurons”observed in Xenopus embryos (Roberts et al., 1987). A cellgroup in the ventral thalamus of mouse also exhibitedGAD expression early in development (Katarova et al.,2000), but its possible correspondence with those observedin lamprey and teleosts is not clear. The pretectal tegmen-tal group appears to correspond to the nucleus of themedial longitudinal fasciculus, also appearing early in theteleost brain (Ekstrom and Ohlin, 1995; Martin et al.,1998; Doldan et al., 1999). An early GAD-expressing do-main was also reported for the pretectal tegmentum ofE10.5 mouse embryos (Katarova et al., 2000).
Appearance of GABAir telencephalicpopulations
The first GABAir telencephalic nucleus of lamprey con-sists of CSF-c neurons and may correspond in part to theGABAir “nucleus telencephali” of Gasterosteus (Ekstromand Ohlin, 1995). In lamprey, GABA immunoreactivity ofthis nucleus can be followed consistently throughout de-velopment to adulthood (unpublished observations). Thus,this nucleus probably corresponds in part to the T4 region(nucleus commissura anterioris) of Schober (1964) and theneuroporic recess-paraterminal preoptic nuclei of Pombaland Puelles (1999). A similar GABAir terminal brain pop-ulation is found in the telencephalon of Xenopus embryos(Roberts et al., 1987). By its topology, the earliest telen-cephalic GABAir nucleus of lamprey likely corresponds tothe GAD65-expressing neuronal populations of theseptum-diagonal band of developing mouse, which has
already appeared at the E10.5 stage (Katarova et al.,2000).
The second GABAir population appearing in the telen-cephalon of lamprey prolarvae is probably the source ofGABAir populations observed in both the striatum andthe olfactory bulbs of adults (Pombal et al., 1997;Melendez-Ferro et al., 2001). This compact nucleus occu-pies most of the telencephalic ventricular surface in earlylarvae, but, as development of the telencephalic hemi-spheres proceeds, its relative volume diminishes. In em-bryos of Gasterosteus, a distinction between two telence-phalic GABAir populations has not been made (Ekstromand Ohlin, 1995). Interestingly, the olfactory bulb-striatalGABAir population observed in lamprey prolarvae corre-sponds both topologically and by its early appearance tothe ganglionic eminences of mouse embryos, which ex-press both GAD genes early in development (E10.5 stage;Katarova et al., 2000).
Patterns of GABAir populations in thediencephalon
In late prolarvae, several GABAir groups were observedin the posterior preoptosupracommissural and tuberoin-fundibular regions, the ventral (rostral) and dorsal (cau-dal) thalamus, and the pretectum. These groups areroughly similar to those observed in the forebrain of 100hour embryos of Gasterosteus (Ekstrom and Ohlin, 1995).In mouse, corresponding populations expressing GADwere observed in the pretectum, ventral thalamus, tuberalarea, dorsal hypothalamus, and suprachiasmatic area. Inthe dorsal thalamus of mice, however, only a narrowstreak of cells (reticular thalamic neurons) expressedGAD (Katarova et al., 2000). This streak is reminiscent ofour caudal (dorsal) thalamic band and the p2 GABAirband of Gasterosteus (Ekstrom and Ohlin, 1995). In manyrespects, the diencephalic GABAergic populations of lam-prey, Gasterosteus, and mouse are strikingly similar.Moreover, the main region of the dorsal thalamus (medialand dorsal tiers) can be characterized by its strictlyGABA-negative character in both lamprey and mouse(present results; Katarova et al., 2000) and by its CRimmunoreactivity in both adult and larval lampreys (Pom-bal and Puelles, 1999; present results).
Appearance of GABAir fibers in forebraincommissures
The commissures observed in the prosencephalon andmidbrain (postoptic, habenular, posterior, and tegmentalcommissures) contain GABAir fibers in prolarval stages oflamprey (present results), and their presence is often themost obvious sign of the appearance of the commissures.GABA immunoreactivity is outstanding in the postopticand posterior commissures in early developmental stagesof teleosts (Ekstrom and Ohlin, 1995; Doldan et al., 1999)and Xenopus (Roberts et al., 1987), and GABA immunore-activity is also notable in the posterior commissure of ratand mouse embryos (Lauder et al., 1986; Katarova et al.,2000). The similar early appearance of GABA in commis-sures throughout vertebrate groups is in agreement withthe great resemblances observed in early GABAir popula-tions. However, none of these early forebrain commissuresappears to correspond to the main ventral GABAir com-missure of the brain of adult amphioxus (Anadon et al.,1998b).
372 M. MELENDEZ-FERRO ET AL.

GABAir populations and the segmentalorganization of the lamprey diencephalon
As reported for Gasterosteus (Ekstrom and Ohlin, 1995),the GABAir populations of the diencephalon and mesen-cephalon of larval lampreys form discrete bands or areas.For Gasterosteus, Ekstrom and Ohlin (1995) find goodagreement between the numbers and positions of thesebands and the transverse neuromeres hypothesized fromthe prosomeric model of organization of the mammalianforebrain (Puelles and Rubenstein, 1993), although theseauthors note that the exact boundaries of the prosence-phalic neuromeres cannot be predicted from their data.Their assignment of GABAir structures to prosomeres,however, shows important differences with respect to theprosomeric model of Puelles and Rubenstein (1993), suchas the inclusion of the whole telencephalon in p6. In themouse, good agreement was observed between GADmRNA expression domains and the different territoriesproposed in the neuromeric model (Katarova et al., 2000).The ontogenetic study of GABAergic systems may providevaluable information on the organization of forebrain ter-ritories (Ekstrom and Ohlin, 1995; Katarova et al., 2000;present results).
Brain incurvation in embryonic and early larval sealampreys is little more than a bent tube and thus mayfacilitate segmental analysis of GABAir populations. Thisis probably a result of the poor development of the visualsystem (eyes, optic nerves, optic tectum) during early lar-val stages (De Miguel and Anadon, 1987; De Miguel et al.,1990) and of the scarce outgrowth of the infundibularregion. However, a problem in identifying early GABAirstructures is that the prolarval and larval lamprey brainsshow very limited cytoarchitectural differentiation.
Several maps of diencephalic territories have beendrawn up for developing river lampreys (Bergqvist, 1932)and for large larvae and adults of river lamprey(Bergqvist, 1932; Heier, 1948; Schober, 1964; Pombal andPuelles, 1999). With regard to the general organization ofthe diencephalon, two of these maps follow roughly acolumnar (dorsoventral) scheme (Heier, 1948; Schober,1964), and two follow segmental models (Bergqvist, 1932;Pombal and Puelles, 1999; see Nieuwenhuys, 1998, for ahistorical review of these models). The two “columnar”maps are similar, the diencephalon being organized infour dorsoventral regions: epithalamus, dorsal and ven-tral thalamus, and hypothalamus, with further rostrocau-dal subdivisions in the hypothalamus and thalamus. Thetwo segmental maps divide the diencephalon into trans-verse segments, although the number of segments andlimits used were different.
The foundational study of Bergqvist (1932) on the ver-tebrate diencephalon included detailed mapping of thebrain territories of the river lamprey (Lampetra fluviati-lis) in development and in adulthood. He divided the thal-amus of larvae and adults into three transverse bands(caudal, medial, and frontal subregions). The ventral andmedial diencephalic sulci, and the fasciculus retroflexus,run within his frontal, medial, and caudal thalamic sub-regions, respectively. The ventral diencephalic region wassubdivided into the pars tuberculi posterioris (the ventralregion of his caudal and medial thalamic parts) and thehypothalamus (Bergqvist, 1932; Bergqvist and Kallen,1953). The hypothalamic region includes the partes inter-media and caudalis hypothalami, which correspond to theventral region of Bergqvist and colleagues’ pars dorsalis
telencephali and pars frontalis thalami and the pars ros-tralis hypothalami that corresponds to the ventral regionof the area ventralis telencephali and the preoptic region(Bergqvist, 1932; Bergqvist and Kallen, 1953).
More recently, the prosomeric model of forebrain orga-nization (Puelles and Rubenstein, 1993; Rubenstein et al.,1994; Puelles, 1995; Puelles et al., 1996; Milan andPuelles, 2000) has been applied to the adult lamprey onthe basis of cell distribution and several immunocyto-chemical markers (Pombal and Puelles, 1999). AlthoughPombal and colleagues also studied sea lampreys, thebasis of their study was the river lamprey. In their proso-meric map, the fasciculus retroflexus and transverse di-encephalic sulci were used as approximate limits of seg-ments, and as a result their p2 and p3 alar plate zones areshifted rostrally by about a half a segment with respect toBergqvist’s pars caudalis and nucleus medialis thalami.Moreover, Pombal and Puelles introduce a further seg-mental territory (p6) in Bergqvist’s hypothalamus.
The distribution of GABAir diencephalic populationsduring sea lamprey development does not show evidentsimilarities to the territories defined in “columnar” mapsof adult river lampreys but does show marked resem-blances with some territories postulated by the two seg-mental maps. Among these segmental maps, that elabo-rated by Pombal and Puelles (1999) has much greatertopological detail and uses the additional immunocyto-chemical markers employed in this study; in what follows,most comparisons will therefore be in reference to thismap.
In the preoptic region of prolarvae and larvae, GABAimmunocytochemistry allows us to distinguish between asickle-shaped negative band that roughly corresponds tothe magnocellular preoptic nucleus of Pombal and Puelles(1999) and a caudal GABAir portion extending over thecommissural optopostoptic plate from the bottom of theoptic recesses toward the infundibular region. The GABA-negative band may correspond in part to a band ofgonadotropin-releasing hormone cells observed in prolar-val and larval sea lampreys (Tobet et al., 1995, 1996).Neither Bergqvist (1932), nor Heier (1948), nor Schober(1964) distinguished between these regions. From itsstart, the GABAir posterior preoptic-supracommissuralband appears homogeneous, which does not support thesegmental division into p6 and p5 portions (epichiasmaticnucleus and paracommissural preoptic nucleus) indicatedby Pombal and Puelles (1999). The GABAergic characterof this caudal preoptic region was not reported by theseauthors.
For the rostral thalamus, we have distinguished twoadjacent GABAir populations, one ventrorostral, withCSF-c-type neurons, and the other dorsocaudal, withoutneurons of the CSF-c type. In view of their intermediateposition between the subhippocampal lobe, the dorsalthalamus, and the caudal preoptic region, these popula-tions probably correspond to the alar plate p3 and p4regions (ventral thalamus and “paraventricular nucleus”)of Pombal and Puelles (1999) and in part to zones of thefrontal and medial thalamic parts of Bergqvist (1932). Thepresence of two GABAir populations with different celltypes supports the distinction between p4 and p3 regionspostulated by Pombal and Puelles (1999). Although theseauthors considered that p4 was largely GABA-negative,our results indicate that this is not the case in developingsea lamprey.
373DEVELOPMENT OF GABA IN LAMPREY FOREBRAIN

A problem for interpretation of the GABAir domains inprolarvae and early larvae is the confluence of the GABAirpopulation of the caudal preoptic region with the alarp3–p4 GABAir populations and the GABAir tuberal pop-ulation, which extends from this confluence to the base ofthe caudal infundibular recess. All these populations formtogether a roughly Y-shaped figure. The caudal limit of thetuberal GABAir population is the ventral diencephalicsulcus of Bergqvist (1932), the same caudal limit as that ofthe serotoninergic tuberal population (present results).These four confluent GABAir territories form a figurereminiscent of the three-radial territory of expression ofthe dlx-2 gene and some class III POU genes (zp-12, zp-23)in the diencephalon of 32–36 hour postfertilization ze-brafish (Hauptmann and Gerster, 2000a,b). In earlymouse development, domains of GAD gene expression arealso temporally and spatially overlapped with those ofsome regulatory genes (Katarova et al., 2000).
The similarity of GABAir populations in the hypothala-mus sensu amplio of sea lamprey prolarvae and larvaemakes GABA expression relatively useless for detectingsegmental limits, such as those reported between theretrocommissural/tuberal (p6/p5) and tuberal/tuberomam-millary (p5/p4) regions (Pombal and Puelles, 1999). The re-ported distribution of substance P-immunoreactive neuronsalong a continuous ventral hypothalamic band extendingfrom the retrocommissural region to the tuberal region inlarval sea lampreys (Yanez, 1992) might also argue againstthe existence of the retrocommissural/tuberal (p6/p5) seg-mental boundary as proposed by Pombal and Puelles (1999)for adult lamprey. Studies of cell proliferation and geneexpression in zebrafish also point out the difficulty of subdi-viding the ventral hypothalamic region in terms of segments(Wullimann and Puelles, 1999; Hauptmann and Gerster,2000a). Whether the limits indicated by Pombal and Puelles(1999) for the tuberal region of lampreys correspond to seg-mental limits cannot be assessed from our data and requiresfurther investigation.
The dorsal region of the caudal infundibular recess ap-pears to have a different status on the basis of its GABAirand serotonin-ir populations, well separated from those ofthe tuberal region, and also in view of the presence of athin, PCNA-negative discontinuity (very likely differenti-ated glial cells) between the caudodorsal and the ventro-lateral PCNA-positive ventricular regions in prolarvae(present results). These results, together with the pres-ence of a characteristic non-CFS-c catecholaminergic pop-ulation (paratubercular nucleus) in this zone of sea lam-prey larvae (Yanez, 1992), are consistent with the ideathat the caudal mammillary region is well separated fromthe tuberal region sensu amplio and probably arises froma different prosomere (p3). In early larvae, an indepen-dent band of CSF-c serotonin-ir neurons courses from thepostchiasmatic region to the caudal tuber following acourse parallel to the ventral diencephalic sulcus ofBergqvist (Yanez, 1992; present results). This band corre-sponds to the hypothalamic periventricular organ of Pom-bal and Puelles (1999), which these authors also assign top3. In our opinion, this tuberal region arises from p4. Inadult lamprey brain, serotonin-ir populations of the mam-millary and tuberal regions were also considered as inde-pendent entities (Pierre et al., 1992).
A conspicuous caudal (dorsal) thalamic band of GABAirneurons follows a rather longitudinal course in thalamicregions, crossing the trajectory of the fasciculus ret-roflexus obliquely. This GABAir band is notably reminis-
cent of the GAD-expressing cell cord along the subtha-lamic sulcus of mouse (Katarova et al., 2000). Asomatostatin-ir population of the larval sea lamprey ex-tends in the same manner throughout the caudal thala-mus (Yanez, 1992; Yanez et al., 1992), and a thalamicband of calretinin-ir neurons is observed parallel but dor-sal to the GABAir band (present results). The extension ofthese three thalamic bands in larval sea lampreys clearlyexceeds the approximate caudal limit (fasciculus ret-roflexus) proposed by Pombal and Puelles (1999) for thedorsal thalamus (p2), although these bands can be settledwithin the caudal thalamic subregion of Bergqvist (1932).Our data on the sea lamprey suggest that the p2/p1boundary is far beyond the fasciculus retroflexus. In 30mm larvae, a GABA-negative band separates the caudalthalamic GABAir population from that of the rostral teg-mentum, and this negative band may be envisaged as theapproximate p2/p1 limit. It is interesting to note that inlamprey the relative position of the caudal thalamicGABAir band shifts ventrally as development of theGABA-negative dorsal thalamus domains proceed, indi-cating that the dorsal and medial tiers of the dorsal thal-amus have a comparatively late appearance, also indi-cated by the considerable thickness of the PCNAir layersobserved in the dorsal thalamus of early larvae (unpub-lished results). The dorsal thalamus of mouse, with theexception of the above-mentioned cord, is also GAD nega-tive (Katarova et al., 2000).
The early GABAir population observed in the tegmen-tum of p1 (nucleus of the medial longitudinal fascicle) hasbeen discussed above. The appearance of this early popu-lation is followed by those of GABAir populations in thedorsal pretectal tegmentum and the pretectal nucleus ofthe posterior commissure. Among these, the paracommis-sural population has the most characteristic appearance,being recognizable by the loose organization of its GABAirneurons from its beginning. Pombal and Puelles (1999)have divided their dorsal pretectal region into precommis-sural, commissural, and postcommissural tiers, using cy-toarchitectonic criteria. However, the homogeneous char-acteristics of the GABAir population throughout thedorsal pretectum do not support such a distinction, atleast in larvae. In zebrafish embryos, early regulatorygene expression is also homogeneous throughout this re-gion (Hauptmann and Gerster, 2000a,b). In later larvaeand adults of sea lamprey, the paracommissural pretectalregion is characterized by the abundance of GABAir neu-rons in the periventricular strata and the presence ofmigrated GABAir neurons scattered through intermedi-ate and superficial pretectal regions (unpublished obser-vations). This is a clear difference with respect to thedorsal tier of the dorsal thalamus of Pombal and Puelles(1999), which lacks GABAir neurons in the periventricu-lar strata of adults (unpublished observations). A similardistinction between GAD-negative dorsal thalamus andGAD-positive pretectal domains is found in mouse (Ka-tarova et al., 2000).
CONCLUSIONS
Study of the development of GABAir populations inlamprey reveals a striking resemblance between theseearly populations and those observed in teleosts and inmouse. The consistent pattern observed suggests a highdegree of conservation of the developmental pattern ofGABAergic populations throughout evolution. Accord-
374 M. MELENDEZ-FERRO ET AL.

ingly, knowledge of early GABAergic populations may bevery useful for understanding the general organization ofthe early domains of the forebrain of vertebrates and forcomparative studies of later prosencephalic territories.
ACKNOWLEDGMENTS
M.M.-F. received a predoctoral fellowship from the Uni-versity of Santiago de Compostela.
LITERATURE CITEDAnadon R, Melendez-Ferro M, Perez-Costas E, Pombal MA, Rodicio MC.
1998a. Centrifugal fibers are the only GABAergic structures of theretina of the larval sea lamprey: an immunocytochemical study. BrainRes 782:297–302.
Anadon R, Adrio F, Rodrıguez-Moldes I. 1998b. Distribution of GABAimmunoreactivity in the central and peripheral nervous system ofamphioxus (Branchiostoma lanceolatum Pallas). J Comp Neurol 401:293–307.
Aoki E, Semba R, Kashiwamata S. 1989. When does GABA-like immuno-reactivity appear in the rat cerebellar GABAergic neurons? Brain Res502:245–251.
Barale E, Fasolo A, Girardi E, Artero C, Franzoni MF. 1996. Immunohis-tochemical investigation of �-aminobutyric acid ontogeny and transientexpression in the central nervous system of Xenopus laevis tadpoles.J Comp Neurol 368:285–294.
Barker JL, Behar Y-X, Li Q-Y, Ma W, Maric D, Maric Y, Schaffner AE,Serafini R, Smith SV, Somogyi R, Vautrin JY, Wen X-L, Xian H. 1998.GABAergic cells and signals in CNS development. Perspect Dev Neu-robiol 5:305–322.
Benagiano V, Virgintino D, Rizzi A, Flace P, Troccoli V, Bormann J,Monaghan P, Robertson D, Roncali L, Ambrosi G. 2000. Glutamic aciddecarboxylase-positive neuronal cell bodies and terminals in the hu-man cerebellar cortex. Histochem J 32:557–564.
Bergqvist H. 1932. Zur Morphologie des Zwischenhirns bei niederen Wir-beltieren. Acta Zool 13:57–304.
Bergqvist H, Kallen B. 1953. Studies on the topography of migration areasin the vertebrate brain. Acta Anat 17:353–369.
Cherubini E, Gaiarsa JL, Ben-Ari Y. 1991. GABA: an excitatory transmit-ter in early postnatal life. Trends Neurosci 14:515–519.
Christenson J, Bongianni F, Grillner S, Hokfelt T. 1991a. PutativeGABAergic input to axons of spinal interneurons and primary sen-sory neurons in the lamprey spinal cord as shown by intracellularLucifer yellow and GABA immunohistochemistry. Brain Res 538:313–318.
Christenson J, Alford S, Grillner S, Hokfelt T. 1991b. Co-localized GABAand somatostatin use different ionic mechanisms to hyperpolarize tar-get neurons in the lamprey spinal cord. Neurosci Lett 134:93–97.
De Miguel E, Anadon R. 1987. Development of the retina and the optictectum of Petromyzon marinus L.: a light microscopic study. J Hirn-forsch 28:445–456.
De Miguel E, Rodicio MC, Anadon R. 1990. Organization of the optic nerveprojections of the larval lamprey. A HRP study. J Comp Neurol 302:529–542.
Doldan MJ, Prego B, Holmqvist BI, De Miguel E. 1999. Distribution ofGABA-immunolabeling in the early zebrafish (Danio rerio) brain. EurJ Morphol 37:126–129.
Ekstrom P, Ohlin L-M. 1995. Ontogeny of GABA-immunoreactive neuronsin the central nervous system in a teleost, Gasterosteus aculeatus L.J Chem Neuroanat 9:271–288.
Ekstrom P, Nyberg L, van Veen T. 1985. Ontogenetic development ofserotoninergic neurons in the brain of a teleost, the three-spinedstickleback. An immunohistochemical analysis. Dev Brain Res 17:209 –224.
Gragera RR, Muniz E, Martınez-Rodrıguez R. 1993. Electron microscopicimmunolocalization of GABA and glutamic acid decarboxylase (GAD)in cerebellar capillaries and their microenvironment. Cell Mol Biol39:809–817.
Hauptmann G, Gerster T. 2000a. Regulatory gene expression patternsreveal transverse and longitudinal subdivisions of the embryonic ze-brafish forebrain. Mech Dev 91:105–118.
Hauptmann G, Gerster T. 2000b. Combinatorial expression of zebrafishBrn-1- and Brn-2-related POU genes in the embryonic brain, proneph-ric primordium, and pharyngeal arches. Dev Dyn 218:345–358.
Heier P. 1948. Fundamental principles in the structure of the brain. Astudy of the brain of Petromyzon fluviatilis. Acta Anat 6(Suppl):1–213.
Katarova Z, Sekerkova G, Prodan S, Mugnaini E, Szabo G. 2000. Domain-restricted expression of two glutamic acid decarboxylase genes in midg-estation mouse embryos. J Comp Neurol 424:607–627.
Kuratani S, Horigome N, Ueki T, Aizawa S, Hirano S. 1998. Stereotypedaxonal bundle formation and neuromeric patterns in embryos of acyclostome, Lampetra japonica. J Comp Neurol 391:99–114.
Lauder JM, Han VKM, Henderson P, Verdoorn T, Towle AC. 1986. Prena-tal ontogeny of the GABAergic system in the rat brain: an immunocy-tochemical study. Neuroscience 19:465–493.
LoTurco JJ, Owens DF, Heath MJS, Davis MBE, Kriegstein AR. 1995.GABA and glutamate depolarize cortical progenitor cells and inhibitDNA synthesis. Neuron 15:1287–1298.
Madtes P, Redburn DA. 1983. GABA as a trophic factor during develop-ment. Life Sci 33:979–984.
Martin SC, Heinrich G, Sandell JH. 1998. Sequence and expression ofglutamic acid decarboxylase isoforms in the developing zebrafish.J Comp Neurol 396:253–266.
Melendez-Ferro M, Perez-Costas E, Gonzalez MJ, Pombal MA, Anadon R,Rodicio MC. 2000. GABA-immunoreactive internuclear neurons in theocular motor system of lampreys. Brain Res 855:150–157.
Melendez-Ferro M, Perez-Costas E, Rodrıguez-Munoz R, Gomez-LopezMP, Anadon R, Rodicio MC. 2001. GABA immunoreactivity in theolfactory bulbs of the sea lamprey Petromyzon marinus L. Brain Res893:253–260.
Messersmith EK, Redburn DA. 1993. The role of GABA during develop-ment of the outer retina in the rabbit. Neurochem Res 18:463–470.
Milan FJ, Puelles L. 2000. Patterns of calretinin, calbindin, and tyrosine-hydroxylase expression are consistent with the prosomeric map of thefrog diencephalon. J Comp Neurol 419:96–121.
Mitchell CK, Redburn DA. 1996. GABA and GABA-A receptors are maxi-mally expressed in association with cone synaptogenesis in neonatalrabbit retina. Dev Brain Res 95:63–71.
Nieuwenhuys R. 1998. Morphogenesis and general structure. In: Nieuwen-huys R, ten Donkelaar HJ, Nicholson C, editors. The central nervoussystem of vertebrates, vol 1. Berlin: Springer Verlag. p 159–228.
Northcutt RG, Puzdrowski RL. 1988. Projections of the olfactory bulb andnervus terminalis in the silver lamprey. Brain Behav Evol 32:96–107.
Obata K. 1997. Excitatory and trophic action of GABA and related sub-stances in newborn mice and organotypic cerebellar culture. Dev Neu-rosci 19:117–119.
Piavis GW. 1971. Embryology. In: Hardisty MW, Potter IC, editors. Thebiology of lampreys, vol 1. London: Academic Press. p 361–400.
Pierre J, Reperant J, Ward R, Vesselkin NP, Rio J-P, Miceli D, Kratskin I.1992. The serotoninergic system of the brain of the lamprey, Lampetrafluviatilis: an evolutionary perspective. J Chem Neuroanat 5:195–219.
Pombal MA, Puelles L. 1999. Prosomeric map of the lamprey forebrainbased on calretinin immunocytochemistry, Nissl stain, and ancillarymarkers. J Comp Neurol 414:391–422.
Pombal MA, El Manira A, Grillner S. 1997. Organization of the lampreystriatum—transmitters and projections. Brain Res 766:249–254.
Pombal MA, Yanez J, Marın O, Gonzalez A, Anadon R. 1999. Cholinergicand GABAergic neuronal elements in the pineal organ of lampreys, andtract-tracing observations of differential connections of pinealofugalneurons. Cell Tissue Res 295:215–223.
Puelles L. 1995. A segmental morphological paradigm for understandingvertebrate forebrains. Brain Behav Evol 46:319–337.
Puelles L, Rubenstein JLR. 1993. Expression patterns of homeobox andother putative regulatory genes in the embryonic mouse forebrainsuggest a neuromeric organization. TINS 16:472–479.
Puelles L, Milan FJ, Martınez de la Torre M. 1996. A segmental map ofarchitectonic subdivisions in the diencephalon of the frog Rana perezi:acetylcholinesterase-histochemical observations. Brain Behav Evol 47:279–310.
Rio JP, Vesselkin NP, Kirpitchnikova E, Kenigfest NB, Versaux-Botteri C,Reperant J. 1993. Presumptive GABAergic centrifugal input to thelamprey retina: a double-labeling study with axonal tracing and GABAimmunocytochemistry. Brain Res 600:9–19.
Roberts A, Dale N, Ottersen OP, Storm-Mathisen J. 1987. The earlydevelopment of neurons with GABA immunoreactivity in the CNS ofXenopus laevis embryos. J Comp Neurol 261:435–449.
Rodicio MC, Pombal MA, Anadon R. 1995. Early development and organi-zation of the retinopetal system in larval sea lamprey, Petromyzonmarinus L. An HRP study. Anat Embryol 192:517–526.
375DEVELOPMENT OF GABA IN LAMPREY FOREBRAIN

Rodrıguez-Munoz R, Nicieza AG, Brana F. 2001. Effects of temperature ondevelopmental performance, survival and growth of sea lamprey em-bryos. J Fish Biol 58:475–486.
Rubenstein JLR, Martınez S, Shimamura K, Puelles L. 1994. The embry-onic forebrain: the prosomeric model. Science 266:578–580.
Sakatani K, Black JA, Kocsis JD. 1992. Transient presence and functionalinteraction of endogenous GABA and GABAA receptors in developingrat optic nerve. Proc R Soc London [Biol] 247:155–161.
Schober W. 1964. Vergleichend-anatomische Untersuchungen am Gehirnder Larven und adulten Tiere von Lampetra fluviatilis und Lampetraplaneri. J Hirnforsch 7:107–209.
Tobet SA, Nozaki M, Youson JH, Sower SA. 1995. Distribution of lampreygonadotropin-releasing-hormone-III (GnRH-III) in brain of larval lam-preys (Petromyzon marinus). Cell Tissue Res 279:261–270.
Tobet SA, Chickering TW, Sower SA. 1996. Relationships of gonadotropin-releasing hormone (GnRH) neurons to the olfactory system in develop-ing lamprey (Petromyzon marinus). J Comp Neurol 376:97–111.
von Kupffer C. 1906. Die Morphogenie des Zentralnervensystems. In: Hertwig
O, editor. Handbuch der vergleichenden und experimentellen Entwick-lungslehre der Wirbeltiere, vol 2, part 3. Jena: Fischer. p 1–272.
Wullimann MF, Puelles L. 1999. Postembryonic neural proliferation in thezebrafish and its relationship to prosomeric domains. Anat Embryol329:329–348.
Yanez J. 1992. Estudio histoquımico e inmunohistoquımico sobre la orga-nizacion larvaria de algunos sistemas monoaminergicos y peptidergicosdel encefalo de lamprea de mar (Petromyzon marinus L.). PhD Thesis,University of Santiago de Compostela.
Yanez J, Anadon R. 1994. Afferent and efferent connections of the habe-nula in the larval sea lamprey (Petromyzon marinus L.). An experi-mental study. J Comp Neurol 345:148–160.
Yanez J, Rodrıguez-Moldes I, Anadon R. 1992. Organization ofsomatostatin-immunoreactive fibres in the brain of the larval lamprey(Petromyzon marinus). J Chem Neuroanat 5:511–520.
Yanez J, Pombal MA, Anadon R. 1999. Afferent and efferent connections ofthe parapineal organ in lamprey: a tract tracing and immunocytochem-ical study. J Comp Neurol 403:171–189.
376 M. MELENDEZ-FERRO ET AL.
Related Documents











