Experimental Parasitology 93, 198–206 (1999) Article ID expr.1999.4450, available online at http://www.idealibrary.com on Onchocerca volvulus: Limited Heterogeneity in the Nuclear and Mitochondrial Genomes Elise M. Keddie,* ,1 Tarig Higazi,* Daniel Boakye,* Anthony Merriweather,* Michael C. Wooten,² and Thomas R. Unnasch* ,2 *Division of Geographic Medicine, University of Alabama at Birmingham, Birmingham, Alabama 35294, U.S.A.; and ² Department of Zoology and Wildlife Science, Auburn University, Auburn, Alabama 36849, U.S.A. Keddie, E. M., Higazi, T., Boakye, D., Merriweather, A., Wooten, INTRODUCTION M. C., and Unnasch, T. R. 1999. Onchocerca volvulus: Limited hetero- geneity in the nuclear and mitochondrial genomes. Experimental Para- sitology 93, 198–206. West African populations of Onchocerca volvu- lus endemic to the rain forest and savanna bioclimes of West Africa Onchocerca volvulus is the filarial parasite that causes differ in their ability to induce ocular disease in infected individuals. onchocerciasis, or river blindness. O. volvulus is endemic In recent years, both clinical- and animal-model-based studies have to most of sub-Saharan Africa and is also found in several implicated particular parasite antigens in the development of ocular foci in South and Central America. Infection with O. volvulus onchocerciasis. To test the hypothesis that the difference in pathogenic potential of blinding and nonblinding parasites might be reflected in can lead to various pathologies, including blindness (ocular qualitative differences in antigens that have been implicated in the onchocerciasis) or skin disease (onchodermatitis). The development of ocular onchocerciasis, we compared the sequences of mechanisms involved in the development of ocular oncho- two parasite antigens implicated in the development of ocular disease cerciasis remain incompletely understood. However, it is in blinding- and nonblinding-strain parasites. The results demonstrated clear that much of the ocular pathology is associated with a high level of homogeneity between the parasite strains in these genes. The study was extended to include additional nuclear genes encoding the host’s immune response to the parasite. This pathogenic antigens that are commonly recognized by individuals infected with immune response requires prior systemic exposure to para- O. volvulus and to the mitochondrial genome of the parasite. The site antigens, seems to be mediated through CD4 + T cells results demonstrate a high degree of homogeneity in both the nuclear (Ottesen 1995), and is regulated through the production of and the mitochondrial genomes among O. volvulus isolates collected specific cytokines, most importantly IL-4 and IL-12 from several different sites in Africa and in the Americas. This high degree of genetic homogeneity may reflect the passage of the parasite (Pearlman et al. 1997b, 1995). through a recent genetic bottleneck. q 1999 Academic Press Although the host’s immune response to the parasite is Index Descriptors and Abbreviations: Onchocerca volvulus; filaria; the mechanism through which ocular damage in onchocerci- nematode; mitochondrial genome; population genetics; IL-4, interleu- asis is accomplished, it is clear that parasite factors also play kin 4; IL-12; interleukin 12; OvCaR; Onchocerca volvulus calreticulin; a role in this process. For example, clinical and epidemiolog- OvPDI; Onchocerca volvulus protein disulfide isomerase; PCR; poly- merase chain reaction; nt; nucleotides; cDNA, copy DNA. ical studies have demonstrated that the severity of ocular onchocerciasis varies greatly between the rain forest and the savanna bioclimes in West Africa. In the savanna, a blinding form of onchocerciasis is found in which the prevalence of 1 Present Address: Department of Medical Pathology, University of O. volvulus-induced ocular disease is linearly related to the California at Davis, Davis, CA 95616, U.S.A. intensity of infection in the community (Remme et al. 1989). 2 To whom correspondence should be addressed. Fax: 1(205)933- 5671. E-mail:[email protected]. In contrast, in the rain forest, a nonblinding form of the 0014-4894/99 $30.00 198 Copyright q 1999 by Academic Press All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Experimental Parasitology 93, 198–206 (1999)
Article ID expr.1999.4450, available online at http://www.idealibrary.com on
Onchocerca volvulus: Limited Heterogeneity in the Nuclear andMitochondrial Genomes
Elise M. Keddie,*,1 Tarig Higazi,* Daniel Boakye,* Anthony Merriweather,*
am,Aub
Michael C. Wooten,† and Thomas R. Unnasch*,2
*Division of Geographic Medicine, University of Alabama at Birminghand †Department of Zoology and Wildlife Science, Auburn University,
Keddie, E. M., Higazi, T., Boakye, D., Merriweather, A., Wooten,M. C., and Unnasch, T. R. 1999. Onchocerca volvulus: Limited hetero-geneity in the nuclear and mitochondrial genomes. Experimental Para-sitology 93, 198–206. West African populations of Onchocerca volvu-lus endemic to the rain forest and savanna bioclimes of West Africadiffer in their ability to induce ocular disease in infected individuals.In recent years, both clinical- and animal-model-based studies haveimplicated particular parasite antigens in the development of ocularonchocerciasis. To test the hypothesis that the difference in pathogenicpotential of blinding and nonblinding parasites might be reflected inqualitative differences in antigens that have been implicated in thedevelopment of ocular onchocerciasis, we compared the sequences oftwo parasite antigens implicated in the development of ocular diseasein blinding- and nonblinding-strain parasites. The results demonstrateda high level of homogeneity between the parasite strains in these genes.The study was extended to include additional nuclear genes encodingantigens that are commonly recognized by individuals infected withO. volvulus and to the mitochondrial genome of the parasite. Theresults demonstrate a high degree of homogeneity in both the nuclearand the mitochondrial genomes among O. volvulus isolates collectedfrom several different sites in Africa and in the Americas. This highdegree of genetic homogeneity may reflect the passage of the parasite
through a recent genetic bottleneck. q 1999 Academic PressIndex Descriptors and Abbreviations: Onchocerca volvulus; filaria;nematode; mitochondrial genome; population genetics; IL-4, interleu-kin 4; IL-12; interleukin 12; OvCaR; Onchocerca volvulus calreticulin;OvPDI; Onchocerca volvulus protein disulfide isomerase; PCR; poly-merase chain reaction; nt; nucleotides; cDNA, copy DNA.
1Present Address: Department of Medical Pathology, University ofCalifornia at Davis, Davis, CA 95616, U.S.A.
2To whom correspondence should be addressed. Fax: 1(205)933-5671. E-mail:[email protected].
198
Birmingham, Alabama 35294, U.S.A.;urn, Alabama 36849, U.S.A.
INTRODUCTION
Onchocerca volvulus is the filarial parasite that causesonchocerciasis, or river blindness. O. volvulus is endemicto most of sub-Saharan Africa and is also found in severalfoci in South and Central America. Infection with O. volvuluscan lead to various pathologies, including blindness (ocularonchocerciasis) or skin disease (onchodermatitis). Themechanisms involved in the development of ocular oncho-cerciasis remain incompletely understood. However, it isclear that much of the ocular pathology is associated withthe host’s immune response to the parasite. This pathogenicimmune response requires prior systemic exposure to para-site antigens, seems to be mediated through CD4+ T cells(Ottesen 1995), and is regulated through the production ofspecific cytokines, most importantly IL-4 and IL-12(Pearlman et al. 1997b, 1995).
Although the host’s immune response to the parasite isthe mechanism through which ocular damage in onchocerci-asis is accomplished, it is clear that parasite factors also playa role in this process. For example, clinical and epidemiolog-ical studies have demonstrated that the severity of ocular
onchocerciasis varies greatly between the rain forest and thesavanna bioclimes in West Africa. In the savanna, a blindingform of onchocerciasis is found in which the prevalence ofO. volvulus-induced ocular disease is linearly related to theintensity of infection in the community (Remme et al. 1989).In contrast, in the rain forest, a nonblinding form of the0014-4894/99 $30.00Copyright q 1999 by Academic Press
All rights of reproduction in any form reserved.

gift from Dr. John Donelson (Donelson et al. 1988). cDNA sequenceswere obtained from the blinding and nonblinding strain libraries by
GENETIC HETEROGENEITY IN Onchocerca volvulus
disease is found. Here, although onchodermatitis is common,ocular onchocerciasis is rare, even in areas where the inten-sity of infection is extremely high (Dadzie et al. 1989a,b).Furthermore, data derived from isoenzyme studies and stud-ies of a repeated sequence family have suggested that geneticdifferences exist between O. volvulus populations endemicto the rain forest and savanna bioclimes (Cianchi et al. 1985;Flockhart et al. 1986; Zimmerman et al. 1994). These data,when taken together, have led to the hypothesis that distinctstrains of O. volvulus are endemic to the two bioclimesof West Africa which differ in their ability to induce ocu-lar disease.
In the past several years, evidence has accumulated thatsuggests that particular parasite antigens may play an im-portant role in the development of ocular onchocerciasis.For example, previous work has suggested a link betweenthe level of serum antibodies to the human autoantigen cal-reticulin and the development of anterior chamber oculardisease in patients infected with O. volvulus (Lux et al.1992). Further studies have identified a cross-reacting epi-tope present in human and O. volvulus calreticulin (OvCaR)located in the carboxyl terminal end of the molecule, sug-gesting a potential role for molecular mimicry in the develop-ment of ocular onchocerciasis (Rokeach et al. 1994). Simi-larly, studies employing a murine model of anterior chamberocular disease have demonstrated that certain cloned O.volvulus proteins are capable of inducing pathologies similarto those seen in ocular onchocerciasis when intracorneallyinjected into animals that have been presensitized with crudeadult worm antigen (Pearlman et al. 1994). The major patho-genic epitope of one such parasite protein, O. volvulus pro-tein disulfide isomerase (OvPDI), has been localized to a23-amino-acid peptide located in the carboxyl terminal por-tion of the molecule (Pearlman et al. 1997a).
We hypothesized that the difference in virulence of theblinding and nonblinding strains of O. volvulus might be
reflected in qualitative differences in those parasite antigens that have been implicated in the development of ocular on-chocerciasis. The initial goal of the study was to test thishypothesis. Based upon the initial findings, we expandedthe study to survey additional mitochondrial and nuclearsequences. The data presented suggest that a high degree of genetic homogeneity exists in O. volvulus collected fromseveral sites throughout Africa and in the Americas.MATERIALS AND METHODS
Adult O. volvulus were obtained from infected individuals by nod-ulectomy. Collections were carried out under the guidelines of the
199
World Health Organization. Following collection, the nodules werepreserved in isopropanol at ambient temperature. Collection data aresummarized in Table I.
DNA was prepared from the nodules following previously publishedprocedures (Unnasch and Meredith 1996). In brief, nodules were bi-sected in a tissue culture dish containing 150 mM NaCl, 10 mMTris–HCl, 1 mM EDTA (pH 8.0), and the included parasite materialwas removed. The solution containing the parasite material was broughtto a final concentration of 100 mg/ml proteinase K and 0.1% SDS.The mixture was incubated for 1 h at 558C, and the mixture wasextracted three times with a 1/1 mixture of phenol and chloroform,followed by a single extraction with chloroform alone. DNA wasprecipitated from the phenol-extracted samples by ethanol and resus-pended in 1 ml of 10 mM Tris–HCl (pH 8.0), 1 mM EDTA. Thesamples from West Africa were collected from villages in which theepidemiological pattern of disease had been characterized, as previouslydescribed (Dadzie et al. 1989a,b; Remme et al. 1989). The classificationbased upon the epidemiological disease pattern was subsequently con-firmed by amplification of the O-150 repeat family followed by hybrid-ization of the PCR products with a battery of strain-specific oligonucle-otide probes, as previously described (Unnasch and Meredith 1996).
Preparation of the cDNA library of adult parasites from the nonblind-ing strain has been described previously (Unnasch et al. 1988). ThecDNA library derived from the blinding strain of O. volvulus was a
PCR amplification employing primers specific for the 58 and 38 endsof the open reading frames of each of the four antigens included inthis study. The Accession Nos. of the DNA sequences for these antigensare as follows: OvCaR, M20565; OvPDI, U12440; OvRAL-2, U00693;and Ov33-3, X13313. Each PCR was carried out in a final volume of100 ml in a solution containing 10 mM Tris–HCl (pH 8.3), 50 mMKCl, 6 mM MgCl2, 0.01% gelatin, 200 mM each dNTP, 0.5 mM eachprimer, 1.25 units of Taq1 DNA polymerase (Applied Biosystems,
TABLE I
Collection Dates and Locations of O. volvulus Samples Used inThis Study
Code Datea Villagea Country Strainb
NB-1 1987 Bong Mine Liberia NonblindingNB-2 1990 Hemakono Cote d’Ivoire NonblindingNB-3 1990 Louga Cote d’Ivoire NonblindingNB-4 1990 Hemakono Cote d’Ivoire NonblindingNB-5 1990 Bonjeima Sierra Leone NonblindingB-1 1986 Manambougou Mali BlindingB-2 1990 Bielikpong Ghana BlindingB-3 1990 Bielikpong Ghana Blinding
B-4 1990 Bielikpong Ghana BlindingB-5 1990 Bielikpong Ghana BlindingNS-1 1996 Abu Hamad Sudan NASS-1 1996 Wau Sudan NAZ-1 NK NK Congo NAE-1 1995 Rio Cayapas Ecuador NAG-1 NK NK Guatemala NAa NK, not known.b NA, not applicable.

200
Norwalk, CT), and 10 ml of a plate lysate of the cDNA library (ca.107 plaque-forming units). Amplification conditions consisted of 40cycles of 1 min at 958C, 1 min at 508C, and 1 min at 728C. Theresulting PCR products were cloned into a T-tailed cloning vector(Invitrogen, San Diego, CA). The DNA sequence of positive cloneswas then determined, using Sequenase T4 DNA polymerase (Amers-ham/USB Cleveland, OH). The DNA sequence from at least threeindependent clones derived from each PCR product was determinedto ensure that any differences detected did not represent PCR artifacts.
Mitochondrial sequences were amplified by PCR employing primersderived from the published sequence of the O. volvulus mitochondrialgenome (Accession No. AF015193). In the case of the AT and tRNAtrp-ND6 intergeneic domains, primers flanking the noncoding domainswere developed. For fragments to be surveyed for restriction site poly-morphisms, the sequence of the complete mitochondrial genome wasfirst examined to identify regions that appeared to contain restrictionsites that would provide informative data on the degree of heterogeneityin the genome. From this analysis, it was decided to concentrate onseven regions of the genome that together contained putative recogni-tion sites for four different restriction enzymes, each recognizing a 4-bp site. The enzymes chosen included Alu1, Mse1, Rsa1, and TspE1.A series of primers were developed to amplify seven fragments, eachroughly 1 kb in length, from the mitochondrial genome. Amplificationswere carried out in a total volume of 100 ml in a solution containing60 mM Tris–HCl (pH 9.0), 15 mM (NH4)2SO4, 2 mM MgCl2, 200mM each of dATP, dGTP, dCTP, and TTP, 0.5 mM each primer, 2.5 unitsof Taq1 DNA polymerase, and 1 ml of the genomic DNA preparationsdescribed above. Amplification conditions consisted of 5 min at 958C,followed by 40 cycles of 1 min at 958C, 1 min at 508C, and 2 min at
728C. Following the PCR, the fragments containing the AT and the46-nt spacer domains were subjected to direct sequence analysis. Inmakeup of O. volvulus, a study was conducted to measure
Variation in Antigen cDNAs in the Blinding and Nonblinding
the case of the fragments to be subjected to PCR-RFLP, the fragmentswere digested with the four restriction enzymes in the buffers recom-mended by the manufacturer (New England Biolabs). The restrictionfragments were separated on a 10% polyacrylamide gel, and the frag-ments were visualized by staining with Syber Green (FMC Biopro-ducts), following the manufacturer’s instructions. Genetic distanceswere determined from pairwise comparisons of the RFLP data em-ploying the method of Nei and Miller (Nei and Miller 1990), usingthe RESTSITE computer program package (Miller 1991).
RESULTS
The initial goal of this study was to test the hypothesisthat qualitative differences in epitopes known to be associ-
ated with ocular pathology existed between the blinding andthe nonblinding strains of O. volvulus. Previous studies haddemonstrated that antibody recognition of an epitope en-coded in the carboxyl terminal portion of OvCaR was associ-ated with the development of sclerosing keratitis in individu-als infected with O. volvulus (Rokeach et al. 1994). Similarly,an epitope responsible for inducing ocular disease in a mousemodel of ocular onchocerciasis was localized to a 23-amino-acid stretch encoded in the carboxyl terminal portion ofKEDDIE ET AL
OvPDI (Pearlman et al. 1997a). Based upon these results,the sequences encoding these epitopes were amplified fromcDNA libraries derived from blinding- and nonblinding-strain O. volvulus, and the amplified product was subjectedto DNA sequence analysis, as described under Materialsand Methods. The results demonstrated that no sequencedifferences in the domains encoding the pathogenic epitopesexisted between the blinding- and the nonblinding-straincDNAs (data not shown). Based upon these results, thestudy was extended to include the coding sequences locatedoutside of the pathogenic domains of OvCaR and OvPDI,as well as to two additional O. volvulus cDNAs, OvRAL-2and Ov33-3. The latter two proteins were chosen becausethey encoded antigens that were commonly recognized byindividuals infected with O. volvulus and therefore mightbe expected to exhibit microsequence heterogeneity. Theresults of this study are summarized in Table II. Of a totalof 2994 nt analyzed, three sequence differences, includingtwo transversions and one transition, were found. None ofthe three sequence differences resulted in changes in thederived amino acid sequence of the encoded proteins (datanot shown).
The results summarized in Table II suggested that littlesequence heterogeneity in the coding sequences of com-monly recognized antigens existed between the blinding andthe nonblinding strains of O. volvulus. To determine if thisfinding reflected a generalized homogeneity in the genetic
the degree of heterogeneity in the mitochondrial genome ofO. volvulus. Recent studies have suggested that the mito-chondrial genome is useful for population-based studies ofgenetic diversity in nematodes (Blouin et al. 1998). Thecomplete DNA sequence of the mitochondrial genome ofO. volvulus has recently been reported (Keddie et al. 1998).The mitochondrial genome of O. volvulus is remarkablycompact, with just 17 intergenic regions (Keddie et al. 1998).
TABLE II
Strains of O. volvulus
Nucleotides Transitions/ GeneticcDNA analyzed transversions distancea
OvCar 1011 0/0 0OvPDI 843 1/1 0.0024Ov RAL-2 420 0/0 0Ov33-3 720 0/1 0.0014
a Determined using the method of Nei and Miller (1990).

GENETIC HETEROGENEITY IN Onchocerca volvulus
The largest of these exhibits characteristics suggesting thatit encodes the so-called D-loop or AT-rich region of themitochondrial genome. The AT domain is normally highlyvariable in sequence (Melton et al. 1997; Parsons et al.1997; Stoneking et al. 1991), suggesting that it might provide
a stringent test of the hypothesis that the genetic uniformityseen in the antigens was a general characteristic of O. volvu-lus. A 252-bp fragment derived from the AT domain wasFIG. 1. Comparison of the sequences of the D-loop domains fromdomain was amplified from O. volvulus and O. ochengi genomic DNAdescribed under Materials and Methods. “.”, Sequence identity with the
201
from two independent foci in the Americas, only two poly-morphisms were found. One of these, a C-T transition lo-cated in position 177, apparently represents an allelic poly-morphism seen within different O. volvulus populations. Thesecond polymorphism, a G-A transition at position 178, was
seen only in the parasite obtained from Ecuador. This highdegree of sequence homogeneity was not shared with otherspecies in the genus Onchocerca. When the sequence of thetherefore amplified from a number of different O. volvulus AT domain of O. ochengi a cattle parasite thought to be theclosest relative to O. volvulus was determined, it was foundindividuals and the DNA sequence of the resulting PCR
products was determined. As shown in Fig. 1, the degree of to be 20% divergent compared to the O. volvulus sequence(Fig. 1).variation in the AT domain of O. volvulus was found to be
extremely limited. In a total of 11 individuals collected from Two approaches were taken to determine if the homogene-ity seen in the putative AT domain extended to other areasdivergent parasite populations in East and West Africa and
12 individual O. volvulus and O. ochengi. A 252-bp portion of the ATand the DNA sequence of the amplified products was determined as
NB-1 sequence; “–”, a gap introduced to maximize sequence alignment.

202
of the O. volvulus mitochondrial genome. As mentionedabove, the mitochondrial genome of O. volvulus is extremelycompact, with very few intergenic regions. Apart from theputative AT domain, a single intergenic region greater than25 nt in length is found in the genome. This region, locatedbetween the tRNAtrp and ND6 genes, is 46 nt in size. TheDNA sequence of this 46-nt intergenic domain was deter-mined from seven individual parasites (four individuals fromthe blinding strain and three from the nonblinding strain).No polymorphisms were noted in this intergenic domainamong the seven individuals assayed (data not shown).
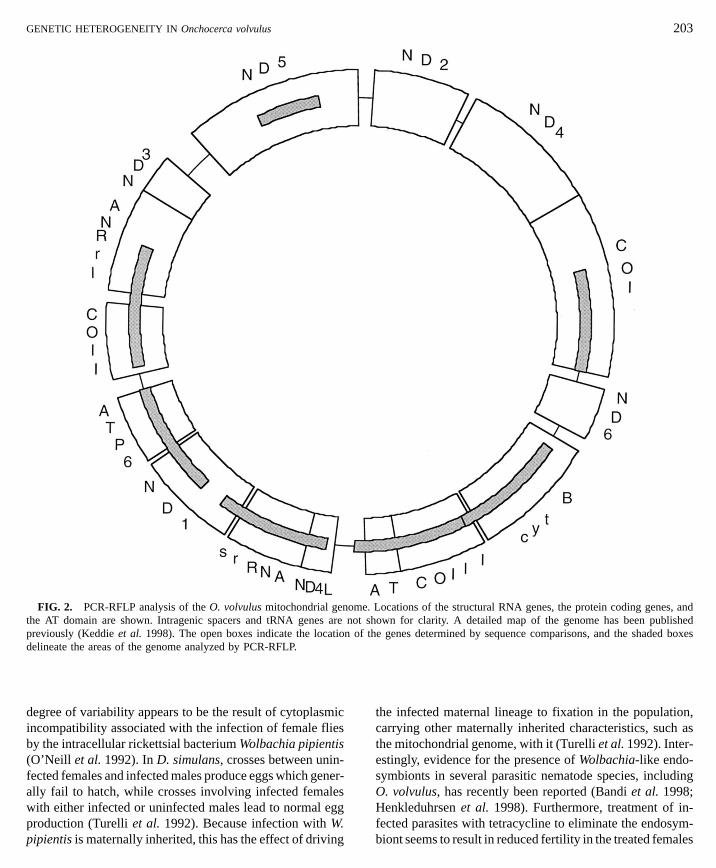
To determine if the homogeneity seen in the intragenicregions extended to the rest of the O. volvulus genome, atotal of seven fragments representing roughly 51% of theentire mitochondrial genome were amplified from 11 indi-vidual O. volvulus (Fig. 2). The resulting samples wereassayed for restriction-site polymorphisms as described un-der Materials and Methods. A total of 97 sites was assayed,
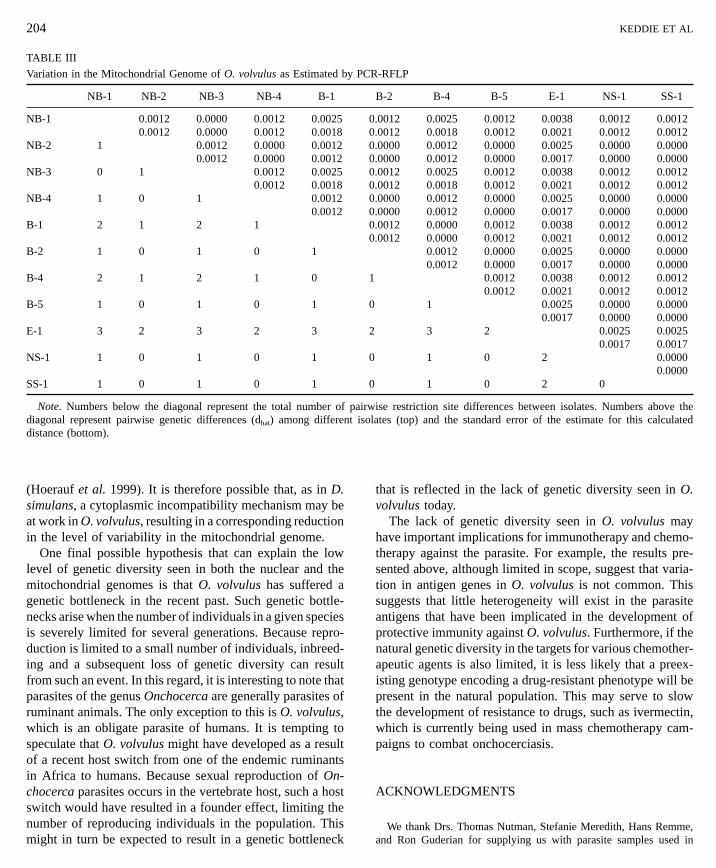
and the results are summarized in Table III. Only four restric-tion-site polymorphisms were found among the 11 individu-als examined. Two of these represented differences between the parasite collected in Ecuador and those collected in Af-rica, while the remaining two polymorphisms representedallelic polymorphisms seen in individuals from both theblinding and the nonblinding strains in Africa and in theindividual collected in Ecuador.DISCUSSION
The original goal of this project was to test the hypothesisthat qualitative differences between the blinding and thenonblinding strains of O. volvulus exist in epitopes that hadpreviously been associated with the development of ocularonchocerciasis. Previous studies have provided evidence forgenetic heterogeneity between different O. volvulus popula-tions. For example, blinding and nonblinding strains of theparasite can be distinguished on the basis of isoenzymepatterns (Cianchi et al. 1985; Flockhart et al. 1986) and by
the composition of a 150-bp repeated sequence family pres-ent in the O. volvulus nuclear genome (Zimmerman et al.1994). We found little or no sequence heterogeneity incDNAs encoding two proteins that have previously beenassociated with the development of ocular onchocerciasisor in two additional cDNAs encoding proteins that are com-monly recognized antigens in individuals infected with O.volvulus. This lack of genetic variation might be a result ofKEDDIE ET AL
functional constraints imposed upon these particular pro-teins. For example, both OvPDI and OvCaR represent evolu-tionarily conserved proteins, while Ov33-3 encodes a cys-teine protease inhibitor (Willenbucher et al. 1993). Thefunction of OvRAL-2 is not known. However, it representsa protein highly expressed in the hypodermis of adult andlarval parasites, suggesting that it might have an importantfunctional role as well (Bradley et al. 1993).
Whatever the reason for the high degree of homogeneityseen in these antigens, it is clear that the clinical and epidemi-ological differences in ocular onchocerciasis seen in theforest and savanna bioclimes of West Africa are not reflectedin qualitative differences in the parasite antigens examinedin this study. This may reflect the limited number of antigensanalyzed or it may be that the differences seen in the rainforest and savanna bioclimes are not due to qualitative differ-ences in the parasite antigens responsible for inducing oculardisease. It is possible that the differences in the ability toinduce ocular disease may result from quantitative differ-ences in the level of expression of the antigens responsiblefor ocular disease. Alternatively, genetic differences in thehost populations may play the deciding role in determiningif and when ocular onchocerciasis develops.
The results presented above suggest that the high levelof homogeneity seen in the antigen genes extends to themitochondrial genome of O. volvulus as well. In direct DNAsequence analysis of the AT domain, commonly one of themost variable portions of the mitochondrial genome, verylittle sequence divergence was seen, despite the fact thatthe parasites included in this study were collected frompopulations from two different continents. Furthermore, thehomogeneity seen in the AT domain reflected a low levelof sequence divergence in the mitochondrial genome as awhole. In a PCR-RFLP study covering over half of the totalmitochondrial genome, pairwise distances between individu-als collected from different locations ranged from 0 to0.0038. This level of divergence is roughly 10-fold less thanthat which is generally seen in other nematodes (Andersonet al. 1995; Blouin et al. 1998; Thomas and Wilson 1991).Indeed, in a recently published survey of variation in thenematode mitochondrial genome, it was concluded that mito-chondrial evolution in the nematode family was quite rapid,
suggesting that the mitochondrial genome of nematodesmight be a useful tool for population-based studies (Blouinet al. 1998). These results suggest that the level of genetichomogeneity exhibited in the O. volvulus mitochondrial ge-nome is not a characteristic of nematodes in general.Studies in Drosophila simulans populations have demon-strated that certain populations of this fly exhibit low levelsof variability in their mitochondrial genomes. This reduced
meot sof
FIG. 2. PCR-RFLP analysis of the O. volvulus mitochondrial genothe AT domain are shown. Intragenic spacers and tRNA genes are npreviously (Keddie et al. 1998). The open boxes indicate the locationdelineate the areas of the genome analyzed by PCR-RFLP.
degree of variability appears to be the result of cytoplasmic
incompatibility associated with the infection of female fliesby the intracellular rickettsial bacterium Wolbachia pipientis(O’Neill et al. 1992). In D. simulans, crosses between unin-fected females and infected males produce eggs which gener-ally fail to hatch, while crosses involving infected femaleswith either infected or uninfected males lead to normal eggproduction (Turelli et al. 1992). Because infection with W.pipientis is maternally inherited, this has the effect of driving. Locations of the structural RNA genes, the protein coding genes, andhown for clarity. A detailed map of the genome has been published
the genes determined by sequence comparisons, and the shaded boxes
the infected maternal lineage to fixation in the population,
GENETIC HETEROGENEITY IN Onchocerca volvulus 203
carrying other maternally inherited characteristics, such asthe mitochondrial genome, with it (Turelli et al. 1992). Inter-estingly, evidence for the presence of Wolbachia-like endo-symbionts in several parasitic nematode species, includingO. volvulus, has recently been reported (Bandi et al. 1998;Henkleduhrsen et al. 1998). Furthermore, treatment of in-fected parasites with tetracycline to eliminate the endosym-biont seems to result in reduced fertility in the treated females
0.0017 0.0000 0.0000E-1 3 2 3 2 3 2 3 2 0.0025 0.0025
airiso
protective immunity against O. volvulus. Furthermore, if the
NS-1 1 0 1 0 1
SS-1 1 0 1 0 1
Note. Numbers below the diagonal represent the total number of pdiagonal represent pairwise genetic differences (dhat) among differentdistance (bottom).
(Hoerauf et al. 1999). It is therefore possible that, as in D.simulans, a cytoplasmic incompatibility mechanism may beat work in O. volvulus, resulting in a corresponding reductionin the level of variability in the mitochondrial genome.
One final possible hypothesis that can explain the lowlevel of genetic diversity seen in both the nuclear and themitochondrial genomes is that O. volvulus has suffered agenetic bottleneck in the recent past. Such genetic bottle-necks arise when the number of individuals in a given speciesis severely limited for several generations. Because repro-duction is limited to a small number of individuals, inbreed-ing and a subsequent loss of genetic diversity can resultfrom such an event. In this regard, it is interesting to note thatparasites of the genus Onchocerca are generally parasites ofruminant animals. The only exception to this is O. volvulus,
which is an obligate parasite of humans. It is tempting tospeculate that O. volvulus might have developed as a resultof a recent host switch from one of the endemic ruminantsin Africa to humans. Because sexual reproduction of On-chocerca parasites occurs in the vertebrate host, such a hostswitch would have resulted in a founder effect, limiting thenumber of reproducing individuals in the population. Thismight in turn be expected to result in a genetic bottleneck0.0017 0.00170 1 0 2 0.0000
0.00000 1 0 2 0
wise restriction site differences between isolates. Numbers above thelates (top) and the standard error of the estimate for this calculated
that is reflected in the lack of genetic diversity seen in O.volvulus today.
The lack of genetic diversity seen in O. volvulus mayhave important implications for immunotherapy and chemo-therapy against the parasite. For example, the results pre-sented above, although limited in scope, suggest that varia-tion in antigen genes in O. volvulus is not common. Thissuggests that little heterogeneity will exist in the parasiteantigens that have been implicated in the development of
204 KEDDIE ET AL
TABLE III
Variation in the Mitochondrial Genome of O. volvulus as Estimated by PCR-RFLP
NB-1 NB-2 NB-3 NB-4 B-1 B-2 B-4 B-5 E-1 NS-1 SS-1
NB-1 0.0012 0.0000 0.0012 0.0025 0.0012 0.0025 0.0012 0.0038 0.0012 0.00120.0012 0.0000 0.0012 0.0018 0.0012 0.0018 0.0012 0.0021 0.0012 0.0012
NB-2 1 0.0012 0.0000 0.0012 0.0000 0.0012 0.0000 0.0025 0.0000 0.00000.0012 0.0000 0.0012 0.0000 0.0012 0.0000 0.0017 0.0000 0.0000
NB-3 0 1 0.0012 0.0025 0.0012 0.0025 0.0012 0.0038 0.0012 0.00120.0012 0.0018 0.0012 0.0018 0.0012 0.0021 0.0012 0.0012
NB-4 1 0 1 0.0012 0.0000 0.0012 0.0000 0.0025 0.0000 0.00000.0012 0.0000 0.0012 0.0000 0.0017 0.0000 0.0000
B-1 2 1 2 1 0.0012 0.0000 0.0012 0.0038 0.0012 0.00120.0012 0.0000 0.0012 0.0021 0.0012 0.0012
B-2 1 0 1 0 1 0.0012 0.0000 0.0025 0.0000 0.00000.0012 0.0000 0.0017 0.0000 0.0000
B-4 2 1 2 1 0 1 0.0012 0.0038 0.0012 0.00120.0012 0.0021 0.0012 0.0012
B-5 1 0 1 0 1 0 1 0.0025 0.0000 0.0000
natural genetic diversity in the targets for various chemother-apeutic agents is also limited, it is less likely that a preex-isting genotype encoding a drug-resistant phenotype will bepresent in the natural population. This may serve to slowthe development of resistance to drugs, such as ivermectin,
which is currently being used in mass chemotherapy cam-paigns to combat onchocerciasis.ACKNOWLEDGMENTS
We thank Drs. Thomas Nutman, Stefanie Meredith, Hans Remme,and Ron Guderian for supplying us with parasite samples used in

GENETIC HETEROGENEITY IN Onchocerca volvulus
this study. We also thank Mr. Mahmoud Rasheed, Mr. Rakonba, and
members of the Sudan National Onchocerciasis Task Force for provid-ing samples from Sudan. Appreciation is also extended to the Onchocer-ciasis Control Programme in West Africa and in particular to its director,Dr. Yankum Dadzie, for their support of this project. This investigationreceived financial support from the UNDP/World Bank/WHO SpecialProgramme for Research and Training in Tropical Diseases (TDR) andwas carried out with support of a training grant from TDR to T.H.REFERENCES
Anderson, T. J. C., Komuniecki, R., Komuniecki, P. R., and Jaenike,J. 1995. Are mitochondria inherited paternally in Ascaris? Interna-tional Journal for Parasitology 25, 1001–1004.
Bandi, C., Anderson, T. J. C., Genchi, C., and Blaxter, M. L. 1998.Phylogeny of Wolbachia in filarial nematodes. Proceedings of theRoyal Society of London—Series B: Biological Sciences 265, 2407–2413.
Blouin, M. S., Yowell, C. A., Courtney, C. H., and Dame, J. B. 1998.Substitution bias, rapid saturation, and the use of mtDNA for nema-tode systematics. Molecular Biology and Evolution 15, 1719–1727.
Bradley, J. E., Tuan, R. S., Shepley, K. J., Tree, T. I. M., Maizels, R.M., Helm, R., Gregory, W. F., and Unnasch, T. R. 1993. Onchocercavolvulus: Characterization of an immunodominant antigen presentin adult and larval parasites. Experimental Parasitology 77, 414–424.
Cianchi, R., Karam, M., Henry, M. C., Villani, F., Kumlien, S., andBullini, L. 1985. Preliminary data on the genetic differentiation ofOnchocerca volvulus in Africa (Nematoda: Filarioidea). Acta Tropica42, 341–351.
Dadzie, K. Y., Remme, J., Baker, R. H. A., Rolland, A., and Thylefors,B. 1989a. Ocular onchocerciasis and intensity of infection in thecommunity, III. West african rainforest foci of the vector Simuliumsanctipauli. Tropical Medicine and Parasitology 41, 376–382.
Dadzie, K. Y., Remme, J., Rolland, A., and Thylefors, B. 1989b. Ocularonchocerciasis and intensity of infection in the community, II. WestAfrican rainforest foci of the vector Simulium yahense. TropicalMedicine and Parasitology 40, 348–354.
Donelson, J. E., Duke, B. O. L., Moser, D., Zeng, W., Erondu, N.E., Lucius, R., Renz, A., Karam, M., and Zea-Flores, G. 1988.Construction of Onchocerca volvulus cDNA libraries and partialcharacterization of the cDNA for a major antigen. Molecular andBiochemical Parasitology 31, 241–250.
Flockhart, H. A., Cibulskis, R. E., Karam, M., and Albiez, E. J. 1986.Onchocerca volvulus: Enzyme polymorphism in relation to the dif-ferentiation of the forest and savannah strains of this parasite. Trans-actions of the Royal Society of Tropical Medicine and Hygiene80, 285–292.
Henkle-Duhrsen, K., Eckelt, V. H. O., Wildenburg, G., Blaxter, M.,and Walter, R. D. 1998. Gene structure, activity and localizationof a catalase from intracellular bacteria in Onchocerca volvulus.Molecular and Biochemical Parasitology 96, 69–81.
205
Hoerauf, A., Nissen-Pahle, K., Schmetz, C., Henkle-Duhrsen, K., Blax-ter, M. L., Buttner, D. W., Gallin, M. Y., Al-Qaoud, K. M., Lucius,R., and Fleischer, B. 1999. Tetracycline therapy targets intracellularbacteria in the filarial nematode Litomosoides sigmodontis and re-sults in filarial infertility. Journal of Clinical Investigation 103,11–17.
Keddie, E. M., Higazi, T., and Unnasch, T. R. 1998. The mitochondrialgenome of Onchocerca volvulus—Sequence, structure and phyloge-netic analysis. Molecular and Biochemical Parasitology 95,111–127.
Lux, F. A., McCauliffe, D. P., Buttner, D. W., Lucius, R., Capra, J.D., Sontheimer, R. D., and Lieu., T.-S. 1992. Serological crossreactivity between a human Ro/SS-A autoantigen (calreticulin) andthe lRAL-1 antigen of Onchocerca volvulus. Journal of ClinicalInvestigation 89, 1945–1951.
Melton, T., Ginther, C., Sensabaugh, G., Soodyall, H., and Stoneking,M. 1997. Extent of heterogeneity in mitochondrial DNA of sub-Saharan African populations. Journal of Forensic Sciences 42,582–92.
Miller, J. C. 1991. RESTSITE: A phylogenetic program that sorts rawrestriction data. Journal of Heredity 82, 262–263.
Nei, M., and Miller, J. C. 1990. A simple method for estimating averagenumber of nucleotide substitutions within and between populationsfrom restriction data. Genetics 125, 873–879.
O’Neill, S. L., Giordano, R., Colbert, A.M. E., Karr, T. L., and Robert-son, H. M. 1992. 16S rRNA phylogenetic analysis of the bacterialendosymbionts associated with cytoplasmic incompatibility in in-sects. Proceedings of the National Academy of Science USA 89,2699–2702.
Ottesen, E. A. 1995. Immune responsiveness and the pathogenesis ofhuman onchocerciasis. Journal of Infectious Diseases 171, 659–671.
Parsons, T. J., Muniec, D. S., Sullivan, K., Woodyatt, N., Alliston-Greiner, R., Wilson, M. R., Berry, D. L., Holland, K. A., Weedn,V. W., Gill, P., and Holland, M. M. 1997. A high observed substitutionrate in the human mitochondrial DNA control region. Nature Genet-ics 15, 363–368.
Pearlman, E., Diaconu, E., Hazlett, F. E., Merriweather, A., and Un-nasch, T. R. 1997a. Identification of an epitope of a recombinantOnchocerca volvulus protein that induces corneal pathology. Molecu-lar and Biochemical Parasitology 89, 123–135.
Pearlman, E., Kazura, J. W., Bardenstein, D. S., Unnasch, T. R., Perler,F., Diaconu, E., Hazlett, F. E., and Lass, J. H. 1994. Induction ofstromal keratitis by recombinant Onchocerca volvulus proteinsOv33, RAL2 and RAL6. In “Advances in Ocular Immunology” (R.B. Nussenblatt, S. M. Whitcup, R. R. Caspi, and I. Gery, Eds.), pp.289–292. Elsevier, Amsterdam.
Pearlman, E., Lass, J. H., Bardenstein, D. S., Hazlett, F. E., Albright,J. L., Diaconu, G., and Kazura, J. W. 1997b. IL-12 exacerbateshelminth-mediated corneal pathology by augmenting cell recruimentand chemokine expression. Journal of Immunology 158, 827–833.
Pearlman, E., Lass, J. H., Bardenstein, D. S., Kopf, M., Hazlett, F. E.,Diaconu, E., and Kazura, J. W. 1995. Interleukin 4 and T helper type2 cells are required for development of experimental onchocercalkeratitis (river blindness). Journal of Experimental Medicine 182,931–940.

206
Remme, J., Dadzie, K. Y., Rolland, A., and Thylefors, B. 1989. Ocularonchocerciasis and intensity of infection in the community I. WestAfrican savannah. Tropical Medicine and Parasitology 40, 340–347.
Rokeach, L. A., Zimmerman, P. A., and Unnasch, T. R. 1994. Epitopesof the Onchocerca volvulus RAL-1 antigen, a member of the calreti-culin family of proteins, recognized by sera from patients withonchocerciasis. Infection and Immunity 62, 3696–3704.
Stoneking, M., Hedgecock, D., Higuchi, R. G., Vigilant, L., and Erlich,H. A. 1991. Population variation of human mtDNA control regionsequences detected by enzymatic amplification and sequence-spe-cific oligonucleotide probes. American Journal of Human Genetics48, 370–382.
Thomas, W. K., and Wilson, A. C. 1991. Mode and tempo of molecularevolution in the nematode Caenorhabditis: Cytochrome oxidase IIand calmodulin sequences. Genetics 128, 269–279.
Turelli, M., Hoffmann, A. A., and McKechnie, S. W. 1992. Dynamicsof cytoplasmic incompatibility and mtDNA variation in natural Dro-sophila simulans populations. Genetics 132, 713–723.
KEDDIE ET AL
Unnasch, T. R., Gallin, M. Y., Soboslay, P. T., Erttmann, K. D., andGreene, B. M. 1988. Isolation and characterization of expressioncDNA clones encoding antigens of Onchocerca volvulus infectivelarvae. Journal of Clinical Investigation 82, 262–269.
Unnasch, T. R., and Meredith, S. E. O. 1996. The use of degenerateprimers in conjunction with strain and species oligonucleotides toclassify Onchocerca volvulus. In “Species Diagnostic Protocols:PCR and Other Nucleic Acid Methods” (J. P. Clapp, Ed.), pp. 293–303. Humana Press, Totowa, NJ.
Willenbucher, J., Hofle, W., and Lucius, R. 1993. The filarial antigensAv33/Ov33-3 show striking similarities to the major pepsin inhibitorfrom Ascaris suum. Molecular and Biochemical Parasitology 57,349–351.
Zimmerman, P. A., Katholi, C. R., Wooten, M. C., Lang-Unnasch, N.,and Unnasch, T. R. 1994. Recent evolutionary history of AmericanOnchocerca volvulus, based on analysis of a tandemly repeated DNAsequence family. Molecular Biology and Evolution 11, 384–392.
Received 4 March 1999; accepted with revision 4 August 1999
Related Documents











