Freshzvater Biology (1995) 33, 373-383 Nutrient enrichment effects on biofilm metabolism in a Mediterranean stream HELENA GUASCH, EUGENIA MARTf AND SERGI SABATER Departament d'Ecologia, Facultat de Biologia, Universitat de Barcelona, Diagonal 645, 08028, Barcelona, Spain SUMMARY 1. Biofilm biomass and metabolism were analysed in La Solana, a calcareous, undisturbed second-order stream. Measurements were carried out in two Mediterranean climatic extremes, summer and winter. Two on-site experimental channels were used to study changes following nutrient addition to one of them. 2. Alga! biomass (chlorophyll a), NDPP (net daily primary production), GDFP (gross daily primary production) and R (respiration) increased in the enriched channel, and these increases were greater in summer. 3. Photosynthetic capacity (Pmax'^*^') decreased during summer, possibly due to enhanced self-shading accompanying the increase in biomass. 4. In winter, Pmax*^*^' increased and reached values similar to those in the summer control channel. Because grazing was low, the higher values of Pmax*^**' were attributed to partial substitution of the cyanobacterial assemblage by a green-algae dominated assemblage with higher photosynthetic capacity as well as the low increase in biomass preventing any significant self-shading. Introduction The effect of nutrient limitation on primary producers has recently attached much attention in running waters. A growing body of evidence has shown that low concentrations of inorganic nutrients can limit primary production in rivers (e.g. Pringle, 1986), a situation analogous to that in phytoplankton commu- nities (Reynolds, 1984). A common procedure for studying nutrient limita- tion involves examining the effects of nutrient addition on primary producers. Some studies have investigated the response of the whole ecosystem (Peterson etal, 1983; Lock et ai, 1990), but this method is difficult to use in river ecosystems that drain populated areas. Other approaches include the use of artificial substrata, releasing nutrients directly into the river (Fairchild, Lowe & Richardson, 1985; Lowe, Golladay & Webster, 1986; Pringle, 1986), or alternatively artificial channels placed in situ (Hart & Robinson, 1990). While many previous studies have focused on changes in species composition and biomass of the biofilm, few have considered the resulting changes in metabolism (Lock © 1995 Blackwell Science Ltd et al., 1990). Here we apply an approach based on the earlier method of Odum (1956) in which biofilm metabolism is measured as the change occurring in oxygen production and consumption. On-site meas- urements of metabolism are appropriate because of the low degree of disturbance to the natural commun- ity. We used continuously flowing troughs adjacent to the stream to study the effect of nutrient enrichment on natural biofilm communities. This method enabled us to perform several measurements each day with a minimum of disturbance. This study was carried out in La Solana, an undis- turbed second-order stream, where biofilm is a thick calcareous crust in which cyanobacteria, algae and bacteria are interspersed to form a delicate microsys- tem that is extremely sensitive to pollution (Sabater, 1989). The impact of agricultural and residential devel- opment in the Mediterranean area is reflected, among other influences, in an increase in dissolved nutrients in the river water (Sabater et al., in press). High concentrations of nutrients entering the rivers cause 373

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Freshzvater Biology (1995) 33, 373-383
Nutrient enrichment effects on biofilm metabolism ina Mediterranean stream
HELENA GUASCH, EUGENIA MARTf AND SERGI SABATERDepartament d'Ecologia, Facultat de Biologia, Universitat de Barcelona, Diagonal 645, 08028, Barcelona, Spain
SUMMARY
1. Biofilm biomass and metabolism were analysed in La Solana, a calcareous,undisturbed second-order stream. Measurements were carried out in two Mediterraneanclimatic extremes, summer and winter. Two on-site experimental channels were used tostudy changes following nutrient addition to one of them.2. Alga! biomass (chlorophyll a), NDPP (net daily primary production), GDFP (grossdaily primary production) and R (respiration) increased in the enriched channel, andthese increases were greater in summer.3. Photosynthetic capacity (Pmax' * ') decreased during summer, possibly due toenhanced self-shading accompanying the increase in biomass.4. In winter, Pmax* * ' increased and reached values similar to those in the summercontrol channel. Because grazing was low, the higher values of Pmax* **' were attributedto partial substitution of the cyanobacterial assemblage by a green-algae dominatedassemblage with higher photosynthetic capacity as well as the low increase in biomasspreventing any significant self-shading.
Introduction
The effect of nutrient limitation on primary producershas recently attached much attention in runningwaters. A growing body of evidence has shown thatlow concentrations of inorganic nutrients can limitprimary production in rivers (e.g. Pringle, 1986), asituation analogous to that in phytoplankton commu-nities (Reynolds, 1984).
A common procedure for studying nutrient limita-tion involves examining the effects of nutrient additionon primary producers. Some studies have investigatedthe response of the whole ecosystem (Peterson etal,1983; Lock et ai, 1990), but this method is difficult touse in river ecosystems that drain populated areas.Other approaches include the use of artificial substrata,releasing nutrients directly into the river (Fairchild,Lowe & Richardson, 1985; Lowe, Golladay & Webster,1986; Pringle, 1986), or alternatively artificial channelsplaced in situ (Hart & Robinson, 1990). While manyprevious studies have focused on changes in speciescomposition and biomass of the biofilm, few haveconsidered the resulting changes in metabolism (Lock
© 1995 Blackwell Science Ltd
et al., 1990). Here we apply an approach based on theearlier method of Odum (1956) in which biofilmmetabolism is measured as the change occurring inoxygen production and consumption. On-site meas-urements of metabolism are appropriate because ofthe low degree of disturbance to the natural commun-ity. We used continuously flowing troughs adjacent tothe stream to study the effect of nutrient enrichmenton natural biofilm communities. This method enabledus to perform several measurements each day with aminimum of disturbance.
This study was carried out in La Solana, an undis-turbed second-order stream, where biofilm is a thickcalcareous crust in which cyanobacteria, algae andbacteria are interspersed to form a delicate microsys-tem that is extremely sensitive to pollution (Sabater,1989). The impact of agricultural and residential devel-opment in the Mediterranean area is reflected, amongother influences, in an increase in dissolved nutrientsin the river water (Sabater et al., in press). Highconcentrations of nutrients entering the rivers cause
373

374 H. Guasch, E. Marti and Sergi Sabater
an increase of chlorophyll a concentration (Sabater &Sabater, 1992) and induce changes in algal communitystructure (Sabater etal.. 1991). La Solana is a pristine,phosphorus-deficient stream (Marti, Sabater &Armengol, 1994) but, none the less, biomass andannual production are amongst the highest seen innon-polluted streams of a similar order feeding theTer system (Sabater et al., in press). Primary productionshows marked seasonal variation. The period fromlate spring to late summer is characterized by highproduction, which contrasts with low production dur-ing winter. During the latter season, low temperatureand light, as well as very low values of phosphate,are common (Marti et ai. 1994), suggesting that theseenvironmental factors may control primary production(Guasch & Sabater, 1994). To elucidate the effect ofnutrient limitation, we planned enrichment experi-ments in winter and summer, which represented theperiods with lowest and highest values of irradiance,water temperature and background nutrient concen-tration. We investigated the effect of nutrient enrich-ment on biofilm metabolism in La Solana, to evaluatehow calcareous streams behave under increased nutri-ent loading, a common situation in Mediterraneanstreams.
Study site
La Solana is located in the north-east of Spain, 90 kmnorth-west of Barcelona. The stream drains an undis-turbed, calcareous catchment. Vegetation in the basinis mainly composed of pines {Pinus sylvestris L.) andoaks {Quercus pubescens Willd.). Riparian vegetation isnot well developed because of the irregular discharge.Light reaches the river bed, but sunshine only arrivesdirectly from late winter onwards (Table 1). Dischargeranges between 1 and 95 I s~' Maximum flow occursduring heavy rains, especially in autumn and spring(99.9 l s ~ ' in June 1992). Water temperature variesbetween 0.2 and 19 "C according to season (Table 1).The stream has a high alkaliruty (between 3 and4meql"*) and conductivity (3(X)-450|iScm~*), butlow nutrient concentrations (0.1-17 jig l"^ of SRP, 29-300 ng I ^ of N-NO3 and 0-19 \ig T ' of N-NH4; Martiet al, 1994). The epilithic biofilm is characterized by a5-10 mm thick carbonate-encrusting community ofalgae and cyanobacteria (Sabater, 1989). Patches ofRivularia biasolettiana Menegh. coloruze the upper crustduring winter, and between late spring and summer
Table 1 Physical and chemical characteristics during winterand summer in the control and experiment channel. Themeans and standard errors (in parentheses) for each samplingday are indicated
Period Parameter
Winter 93 Water velocity (cm s"')Irradiance ()iE m^^s"')Temperature 'CN-NO3 (M-gl"')N-NH4 (|igl"^)
SRP (ugl""')DIN/SRP
Summer 93 Water velocity (cm s~')Irradiance (nE m~^s~')Temperature "CN-NO3 (^igl"')N-NH4 (fig I"" )SRP (ng l~')DIN/SRP
Control
1.3-2.7356 (94.8)
3.6 (1.4)206 (24)
6.8 (1.3)24 (12)
1.4 (0.2)427 (119)1.3-2.7908 (226)
18.7 (0.8)119 (13)22.4 (5.2)8.7 (0.7)46.7 (7.8)
Experiment
1.3-2.7
356 (94.8)3,6 (1.4)
216 (30)
13(5)37 (15)1.3-2.7908 (226)
18.7 (0.8)129 (22)30 (12)21(8)16.7 (8.2)
they are partially replaced by filamentous green algae(mainly Zygnema spp. ) and diatoms.
Materials and methods
The experimental design aimed to reproduce naturalconditions by using channels big enough to hold alarge number of intact colonized cobbles (35-40cobbles 15-30 cm diameter each, obtained from thestream bed), under a water velocity similar to that inthe stream during base flow conditions. However,replication between channels was not possible becausenatural discharge fluctuations would not have ensureda steady water flow for more than five channelssimultaneously.
The experimental troughs were placed on the streambed. They consisted of three PVC channels arrangedin parallel and mounted on a levelled steel frame. Thechannels were 8 m long and 20 cm wide. Water wascontinuously flushed to the channels through threepipes from a plastic container fed by a pond 50 mupstream. Equal flow was maintained in each channel,with velocities ranging between 1.3 and 2.7 cm s"^This was similar to stream velocities during the sum-mer. At these velocities the Reynolds number was 2100and 4300, respectively. Thus the water was maintainedclose to the boundary that separates laminar fromturbulent flow (i.e. Reynolds number of 1000).
Two channels (experimental and control) were filledwith colonized cobbles. Substrata were disposed into
© 1995 Blackwell Sdence Ltd, Freshwater Biology, 33, 373-383

the channels 15 days before starting the experiment.The third channel (UC) was filled with uncolonizedcobbles, and used to estimate dissolved oxygen dif-fusion into the channels. These cobbles and the PVCchannel had been acid-rinsed previously and werekept dry and covered with a black plastic sheetbetween sampling days to avoid colonization. Nutrientaddition was effected by a dosing system located atthe entry to the experimental channel (EC) whichallowed the slow dissolution of fertilizer tablets intothe water. This plant fertilizer (Gardena NPK) wascomposed (in percentage by weight) of 18% nitrogen(6.3% NO3 and 11.7% NH4), 8% phosphate (as PO4)and 13% K2O (manufacturer's values). The dosingsystem was refilled periodically to ensure a steadyinflow of nutrients, The second colonized channel(CC) was used as a control, with no enrichment.
Identical amounts of nutrients (one tablet of plantfertilizer per week) were added to the experimentalchannel in both the seasons studied. These additionsresulted in a slight increase in nitrate-N, and anobvious increase in ammonia-N with respect to back-ground concentrations in both seasons (Table 1).Because of the different background concentrations ofphosphate in winter and summer, the same nutrientaddition increased values about 10-fold in the winterand only 2-fold in summer. In winter the backgroundN : P ratio was about 400:1, which rose to 30:1following enrichment. In summer the N : P ratio fellfrom 47 : 1-17 :1 (Table 1). Results of nutrient uptakekinetics experiments, performed using short-timenutrient additions (E. Marti, unpublished data), wereevaluated to determine the concentration at whichuptake capacity of the natural community was satur-ated. In winter, natural community phosphate uptakewas saturated at concentrations above 10-15 | ig l"^The summer community was saturated at 25-30 Jig 1" ^Thus, despite different degrees of phosphate increasein the winter and summer experiments, fertilizationsaturated nutrient uptake capacity of the communityin both situations.
Enrichment experiments were carried out in winter1993 and summer 1993. Although both were plannedto last for 8 weeks, a sudden decrease in stream flowforced us to stop the summer experiment in thesixth week. Algal biomass as chlorophyll a, speciescomposition and metabolism (oxygen changes) weremeasured in both channels every 2 weeks. Knownareas of biofilm (4.25 cm^) were defined with a plastic
© 1995 Biackwell Science Ltd, Freshwater Biology, 33, 373-383
Enrichment effects in stream biofilms 375
frame and chosen at random along each channel andscraped for chlorophyll a analysis. Between three andsix samples were taken every 2 weeks during theexperiment. This was the maximum biomass that couldbe removed without significantly affecting metabolicactivity in the channels as a whole. However, moreintensive sampling was also carried out at the begin-ning and end of each experiment. In these cases,two-three whole cobbles were scraped completelyand aliquots were used for algal pigment analysis.Samples were kept at 4 °C, transported to the laborat-ory, and stored in a freezer at -20 "C until extraction.To ensure total chlorophyll extraction (Downes,Hrstich & Vincent, 1993), algal pigment analysisincluded passive extraction with 90% acetone (in thedark, at 4 "C for 12 h) as well as trituration with anhomogenizer. Chlorophyll a measurements includedcorrections for phaeopigments by measuring spectro-photometric absorbances at 665 nm before and afteracidifying the extracts up to 0.003 M HCl (Lorenzen,1967).
Algal biofilm samples for species composition studywere taken by scraping at random in the channels andpreserving the samples in 4% formalin. Algal materialwas observed under the microscope at 400X and iden-tified to species, Up to 300 cells were counted persample. Cells were always used as unit of measure-ment. Semiquantitative data on algal abundances wereused to calculate proportions of different algal groupsand cyanobacteria. Diversity was calculated with theShannon-Wiener index using logarithms to base 2.
Net primary production (NPP) and respiration (R)were measured in the channels using the open oxygenmethod (Odum, 1956; Edwards & Owen, 1962; Triskaet aL, 1983). For each colonized channel (CC and EC),NPP was estimated as the rate of change in dissolvedoxygen per unit area (Q). This was calculated asthe difference between oxygen concentrations at theoutflows of the colonized and uncolonized channels(O2C-O2U) measured simultaneously, and related tothe discharge (f), and the colonized area (S) as follows:
Q = (O2C-O2U) f S"' - NPP (1)
where O2C and O2U are in mgl" \ f in m- h ' and Sin m .
In the colonized channels, these variations integ-rated changes in oxygen concentration resulting fromgross primary production (GPP), respiration (R), anddiffusion (D) to or from the atmosphere. The change
376 H. Guasch, E. Marti and Sergi Sabater
100
100
Week
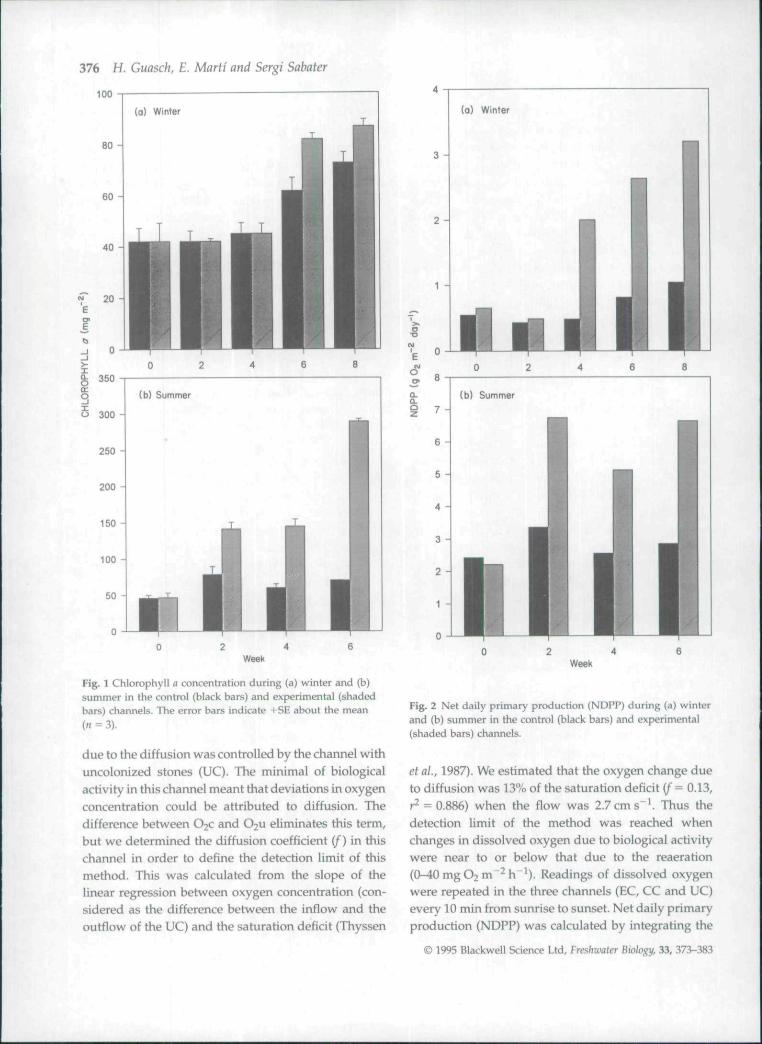
Fig. 1 Chlorophyll a concentration during (a) winter and (b)summer in the control (black bars) and experimental (shadedbars) channels. The error bars indicate +SE about the mean(" = 3).
due to the diffusion was controlled by the channel withuncolonized stones (UC). The minimal of biologicalactivity in this channel meant that deviations in oxygenconcentration could be attributed to diffusion. Thedifference between O2C and O2U eliminates this term,but we determined the diffusion coefficient if) in thischannel in order to define the detection limit of thismethod. This was calculated from the slope of thelinear regression between oxygen concentration (con-sidered as the difference between the inflow and theoutflow of the UC) and the saturation deficit (Thyssen
1 -
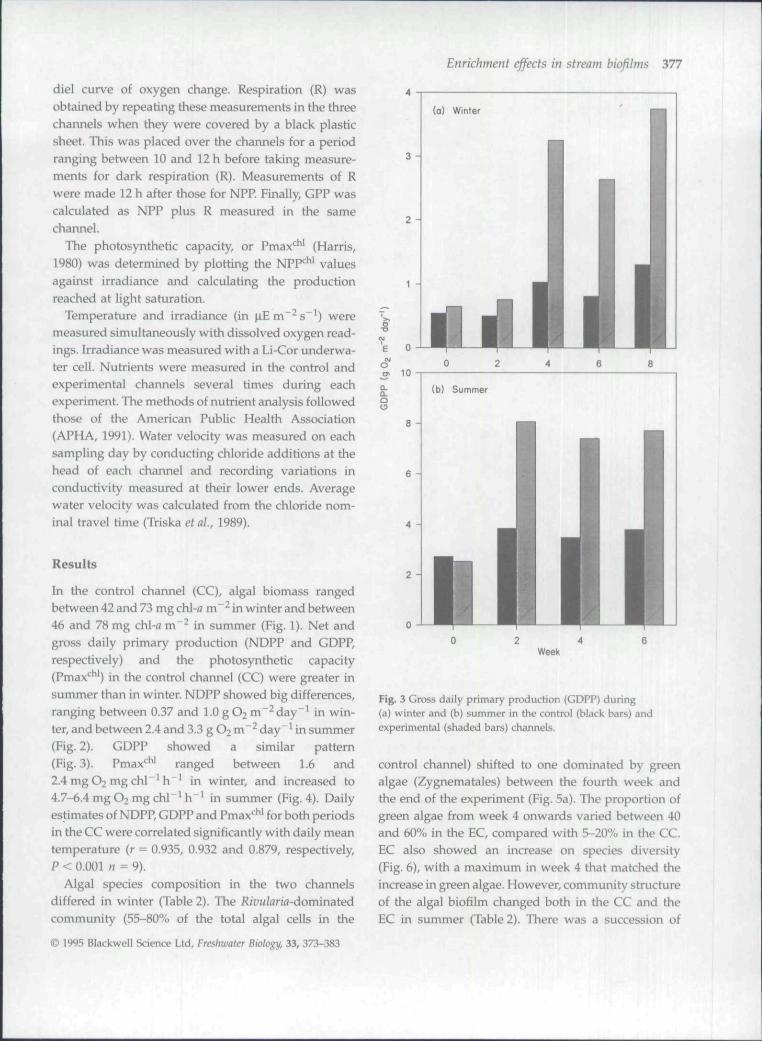
Fig. 2 Net daily primary production (NDPP) during (a) winterand (b) summer in the control (black bars) and experimental(shaded bars) channels.
et al.. 1987). We estimated that the oxygen change dueto diffusion was 13% of the saturation deficit if = 0.13,r - 0.886) when the flow was 2.7 cm s ' ^ Thus thedetection limit of the method was reached whenchanges in dissolved oxygen due to biological activitywere near to or below that due to the reaeration(0-40 mg O2 m"^ h"^). Readings of dissolved oxygenwere repeated in the three channels (EC, CC and UC)every 10 min from sunrise to sunset. Net daily primaryproduction (NDPP) was calculated by integrating the
© 1995 Blackwell Sdence Ltd, Freshwater Biology, 33, 373-383
Enrichment effects in stream biofilms 377
diel curve of oxygen change. Respiration (R) wasobtained by repeating these measurements in the threechannels when they were covered by a black plasticsheet. This was placed over the channels for a periodranging between 10 and 12 h before taking measure-ments for dark respiration (R). Measurements of Rwere made 12 h after those for NPP. Finally, GPP wascalculated as NPP plus R measured in the samechannel.
The photosynthetic capacity,, or Pmax*^ ' (Harris,1980) was determined by plotting the NPP * ' valuesagainst irradiance and calculating the productionreached at light saturation.
Temperature and irradiance (in ^iEm"^s"^) weremeasured simultaneously with dissolved oxygen read-ings. Irradiance was measured with a Li-Cor underwa-ter cell. Nutrients were measured in the control andexperimental channels several times during eachexperiment. The methods of nutrient analysis followedthose of the American Public Health Association(APHA, 1991). Water velocity was measured on eachsampling day by conducting chloride additions at thehead of each channel and recording variations inconductivity measured at their lower ends. Averagewater velocity was calculated from the chloride nom-inal travel time (Triska et aL, 1989).
Results
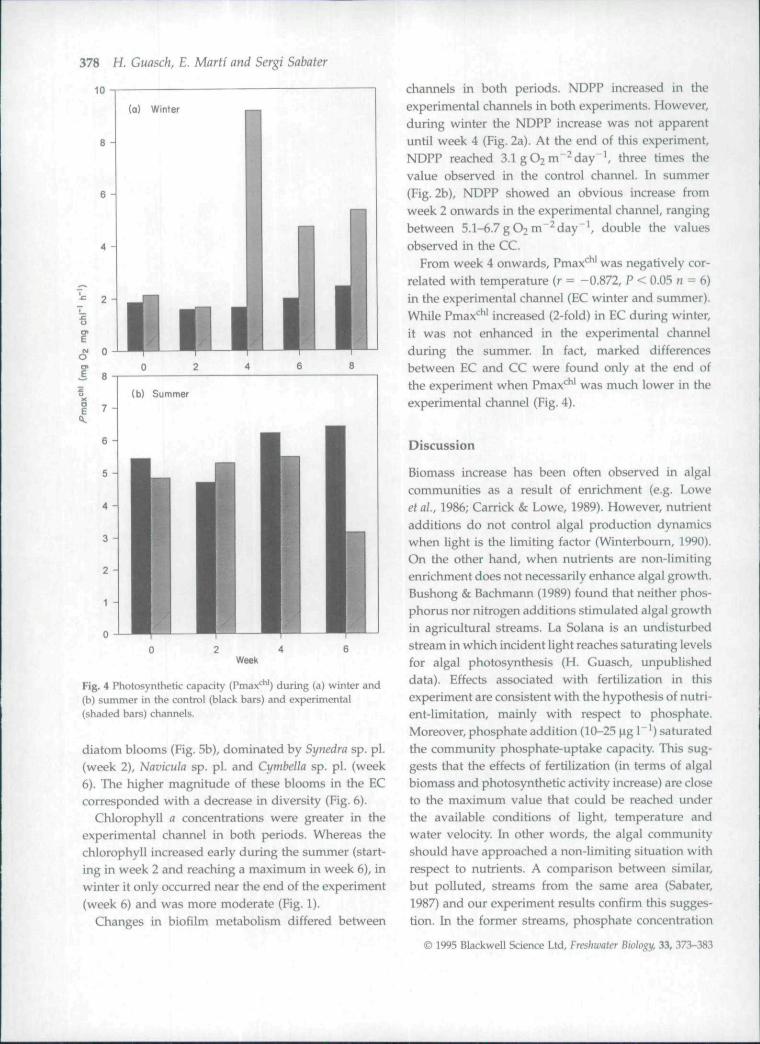
In the control channel (CC), algal biomass rangedbetween 42 and 73 mg chl-fl m" in winter and between46 and 78 mg chl-o m"^ in summer (Fig. 1). Net andgross daily primary production (NDPP and GDPP,respectively) and the photosynthetic capacity(Pmax* * ') in the control channel (CC) were greater insummer than in winter. NDPP showed big differences.
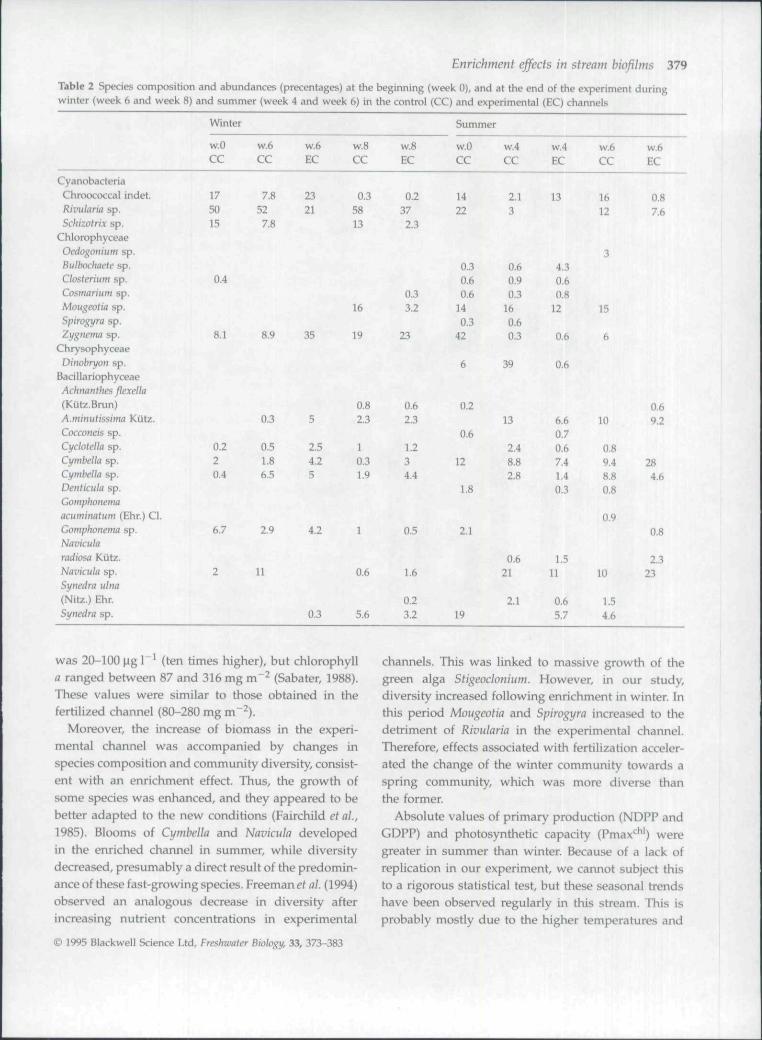
ranging between 0.37 and 1.0 gO2 m"^day"' in win-ter, and between 2.4 and 3.3 g O2 m~^ day"* in summer(Fig. 2). GDPP showed a similar pattern(Fig. 3). Pmax* ' ranged between 1.6 and2.4 mgO2mgchl"^ h~^ in winter, and increased to4.7-6.4 mgO2 mg chl"'h"^ in summer (Fig. 4). Dailyestimates of NDPP, GDPP and Pmax*^ ' for both periodsin the CC were correlated significantly with daily meantemperature (r = 0.935, 0.932 and 0.879, respectively,P<0 .001n = 9).
Algal species composition in the two channelsdiffered in winter (Table 2). The i?/uu/ana-dominatedcommunity (55-80% of the total algal cells in the
© 1995 Blackwell Science Ltd, Freshwater Biology. 33, 373-383
Week
Fig. 3 Gross daily primary production (GDPP) during(a) winter and (b) summer in the control (black bars) andexperimental (shaded bars) channels.
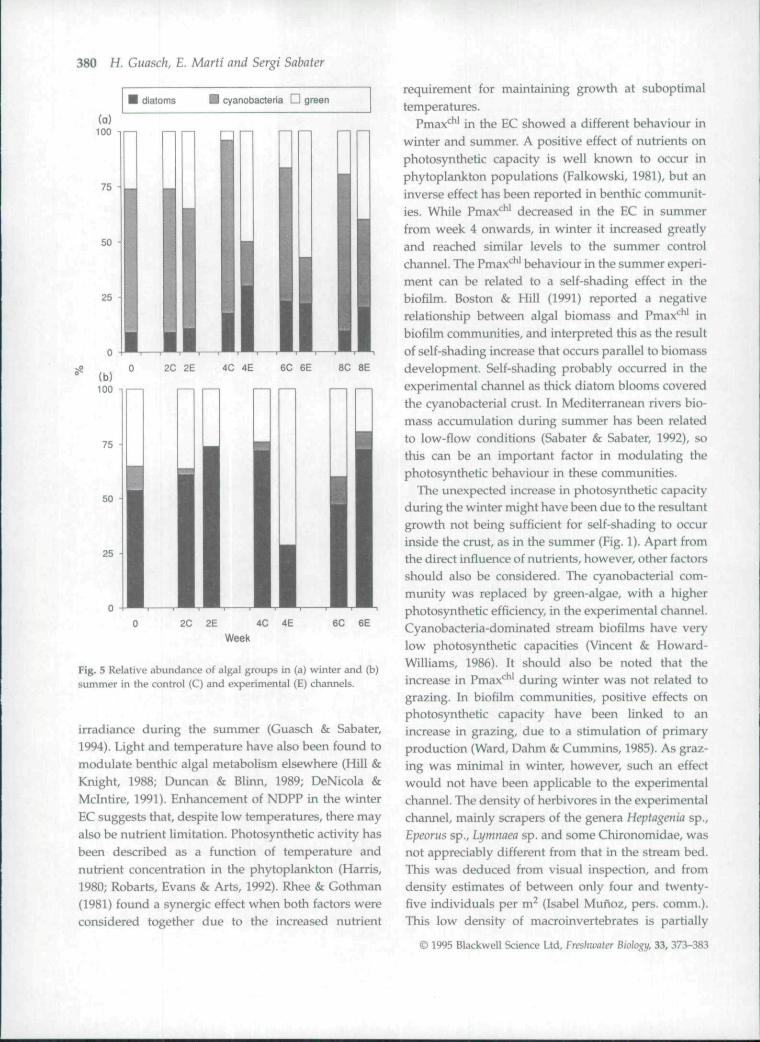
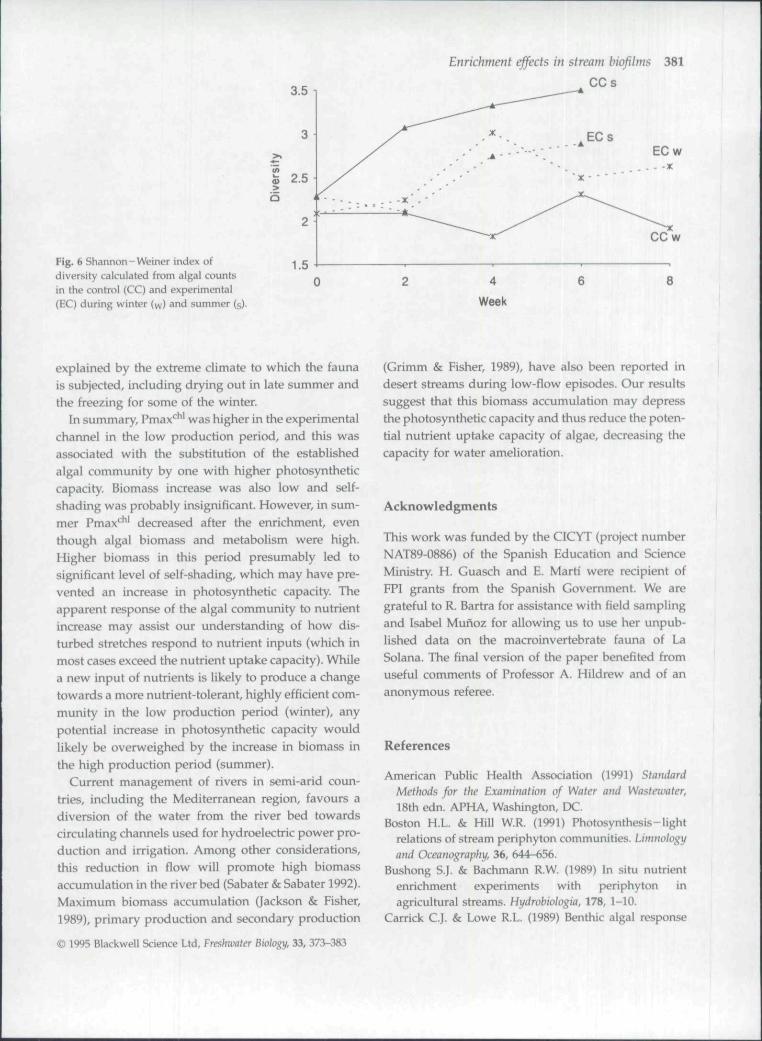
control channel) shifted to one dominated by greenalgae (Zygnema tales) between the fourth week andthe end of the experiment (Fig. 5a). The proportion ofgreen algae from week 4 onwards varied between 40and 60% in the EC, compared with 5-20% in the CC.EC also showed an increase on species diversity(Fig. 6), with a maximum in week 4 that matched theincrease in green algae. However, community structureof the algal biofilm changed both in the CC and theEC in summer (Table 2). There was a succession of
378 H. Guasch, E. Marti and Sergi Sabater
Week
Fig. 4 Photosynthetic capacity (Pmax** ) during (a) winter and(b) summer in the control (black bars) and experimental(shaded bars) channels.
diatom blooms (Fig. 5b), dominated by Synedra sp. pi.(week 2), Navicula sp. pi. and Cymbella sp. pi. (week6). The higher magnitude of these blooms in the ECcorresponded with a decrease in diversity (Fig. 6).
Chlorophyll a concentrations were greater in theexperimental channel in both periods. Whereas thechlorophyll increased early during the summer (start-ing in week 2 and reaching a maximum in week 6), inwinter it only occurred near the end of the experiment(week 6) and was more moderate (Fig. 1).
Changes in biofilm metabolism differed between
channels in both periods. NDPP increased in theexperimental channels in both experiments. However,during winter the NDPP increase was not apparentuntil week 4 (Fig. 2a). At the end of this experiment,NDPP reached 3.1 gO2 m^^day"', three times thevalue observed in the control channel. In summer(Fig. 2b), NDPP showed an obvious increase fromweek 2 onwards in the experimental channel, rangingbetween 5.1-6.7 gO2m~^ day" \ double the valuesobserved in the CC.
From week 4 onwards, Pmax' * ' was negatively cor-related with temperature (r = -0.872, P < 0.05 n = 6)in the experimental channel (EC winter and summer).While Pmax**' increased (2-fold) in EC during winter,it was not enhanced in the experimental channelduring the summer. In fact, marked differencesbetween EC and CC were found only at the end ofthe experiment when Pmax" ' was much lower in theexperimental channel (Fig. 4).
Discussion
Biomass increase has been often observed in algalcommunities as a result of enrichment (e.g. Loweet al, 1986; Carrick & Lowe, 1989). However, nutrientadditions do not control algal production dynamicswhen light is the limiting factor (Winterbourn, 1990).On the other hand, when nutrients are non-limitingenrichment does not necessarily enhance algal growth.Bushong & Bachmarui (1989) found that neither phos-phorus nor nitrogen additions stimulated algal growthin agricultural streams. La Solana is an undisturbedstream in which incident light reaches saturating levelsfor algal photosynthesis (H. Guasch, unpublisheddata). Effects associated with fertilization in thisexperiment are consistent with the hypothesis of nutri-ent-limitation, mainly with respect to phosphate.Moreover, phosphate addition (10-25 [J,g 1~') saturatedthe community phosphate-uptake capacity. This sug-gests that the effects of fertilization (in terms of algalbiomass and photosynthetic activity increase) are closeto the maximum value that could be reached underthe available conditions of light, temperature andwater velocity, ln other words, the algal communityshould have approached a non-limiting situation withrespect to nutrients. A comparison between similar,but polluted, streams from the same area (Sabater,1987) and our experiment results confirm this sugges-tion. In the former streams, phosphate concentration
© 1995 Blackwell Science Ltd, Freshwater Biology, 33, 373-383
Enrichment effects in stream biofilms 379
Table 2 Species composition and abundances (precentages) at the beginningwinter (week 6 and week 8) and summer (week 4 and week 6) in the control
Winter
w.OCC
w.6CC
w.6EC
w,8CC
w.8EC
(week(CC)
0),and
and at the end of the e)cperiment duringexperimental (EC) channels
Summer
w.OCC
w,4CC
w,4 w.6 w.6EC CC EC
CyanobacteriaChroococcal indet,Rivutaria sp.Schizotrix sp.
ChlorophyceaeOedogonium sp.Bulhochaete sp.Ctosterium sp,Cosmarium sp.Mougcotia sp.Spirogyra sp,Zygiiema sp.
ChrysophyceaeDiriobryon sp.
Bacillariophyceae" " ' • — " " ; flexeUa(Kutz.Bnjn)A.minutissima Kutz.Cocconeis sp.Cyclotelia sp.Cymbella sp.Cymbella sp.Denticula sp.
{Ehr.) Cl,Comphonema sp.
radiosa Kiitz,Navicula sp.Synedra ulna(Nitz.) Ehr.Synedra sp.
175015
0.4
8.1
7.8527.8
8.9
2321
35
0.35813
16
19
0.237
2.3
0.33.2
23
1422
0.30.60.6
140,3
42
2.13
0,60,90.3
160.60.3
13
4.30.60,8
12
0,6
1612
3
15
6
0.87.6
39 0.6
0.220,4
0.3
0.51,86,5
5
2.54,25
0.82.3
10.31.9
0.62.3
1.234.4
0.2
0,6
12
1.8
13
2.48,82.8
6,60.70,67.41.40.3
10
0.89.48.80.8
0.69.2
284.6
6.7 2.9 4,2 0.5 2,1
11 0.6 1.6
0.20.3 5.6 3.2 19
0.90.8
0.621
2.1
1.511
0.65.7
10
1.54,6
2,323
was 20-100 ig 1 ^ (ten fimes higher), but chlorophylla ranged between 87 and 316 mg m"^ (Sabater, 1988).These values were similar to those obtained in thefertilized channel (80-280 mg m"^).
Moreover, the increase of biomass in the experi-mental channel was accompanied by changes inspecies composifion and community diversity, consist-ent with an enrichment effect. Thus, the growth ofsome species was enhanced, and they appeared to bebetter adapted to the new condifions (Fairchild etal,1985). Blooms of Cymbella and Navicula developedin the enriched channel in summer, while diversitydecreased, presumably a direct result of the predomin-ance of these fast-growing species. Freeman et al. (1994)observed an analogous decrease in diversity afterincreasing nutrient concentrafions in experimental
© 1995 BlackweU Sdence Ltd, Freshwater Biology, 33, 373-383
channels. This was linked to massive growth of thegreen alga Stigeoclonium. However, in our study,diversity increased following enrichment in winter. Inthis period Mougeotia and Spirogyra increased to thedetriment of Rivularia in the experimental channel.Therefore, effects associated with fertilization acceler-ated the change of the winter community towards aspring community, which was more diverse thanthe former.
Absolute values of primary producfion (NDPP andGDPP) and photosynthetic capacity (Pmax" * ') weregreater in summer than winter. Because of a lack ofreplicafion in our experiment, we cannot subject thisto a rigorous stafistical test, but these seasonal trendshave been observed regularly in this stream. This isprobably mostly due to the higher temperatures and
380 H. Guasch, E. Marti and Sergi Sabater
(a)100 n
75 -
50 -
25
diatoms cyanobacteria D green
2C 2E 4C 4E 6C 6E 8C 8E
0 2C 2E 4C 4E 6C 6E
Week
Fig. 5 Relative abundance of algal groups in (a) winter and (b)summer in the control (C) and experimental (E) channels.
irradiance during the summer (Guasch & Sabater,1994). Light and temperature have also been found tomcxiulate benthic algal metabolism elsewhere (Hill &Knight, 1988; Duncan & Blinn, 1989; DeNicola &Mclntire, 1991). Enhancement of NDPP in the winterEC suggests that, despite low temperatures, there mayalso be nutrient limitation. Photosynthetic activity hasbeen described as a function of temperature andnutrient concentration in the phytoplankton (Harris,1980; Robarts, Evans & Arts, 1992). Rhee & Gothman(1981) found a synergic effect when both factors wereconsidered together due to the increased nutrient
requirement for maintaining growth at suboptimaltemperatures.
Pmax* * ' in the EC showed a different behaviour inwinter and summer. A positive effect of nutrients onphotosynthetic capacity is well known to occur inphytoplankton populations (Falkowski, 1981), but aninverse effect has been reported in benthic communit-ies. While Pmax*^ ' decreased in the EC in summerfrom week 4 onwards, in winter it increased greatlyand reached similar levels to the summer controlchannel. The Pmax*^ ' behaviour in the summer experi-ment can be related to a self-shading effect in thebiofilm. Boston & HiU (1991) reported a negativerelationship between algal biomass and Pmax' ' inbiofilm communities, and interpreted this as the resultof self-shading increase that occurs parallel to biomassdevelopment. Self-shading probably occurred in theexperimental channel as thick diatom blooms coveredthe cyanobacterial crust. In Mediterranean rivers bio-mass accumulation during summer has been relatedto low-flow conditions (Sabater & Sabater, 1992), sothis can be an important factor in modulating thephotosynthetic behaviour in these communities.
The unexpected increase in photosynthetic capacityduring the winter might have been due to the resultantgrowth not being sufficient for self-shading to occurinside the crust, as in the summer (Fig. 1). Apart fromthe direct influence of nutrients, however, other factorsshould also be considered. The cyanobacterial com-munity was replaced by green-algae, with a higherphotosynthetic efficiency, in the experimental channel.Cyanobacteria-dominated stream biofilms have verylow photosynthetic capacities (Vincent & Howard-WUliams, 1986). It should also be noted that theincrease in Pmax' * ' during winter was not related tograzing. In biofilm communities, positive effects onphotosynthetic capacity have been linked to anincrease in grazing, due to a stimulation of primaryproduction (Ward, Dahm & Cummins, 1985). As graz-ing was minimal in winter, however, such an effectwould not have been applicable to the experimentalchannel. The density of herbivores in the experimentalchannel, mainly scrapers of the genera Heptagenia sp.,Epeorus sp., Lymnaea sp. and some Chironomidae, wasnot appreciably different from that in the stream bed.This was deduced from visual inspection, and fromdensity estimates of between only four and twenty-five individuals per m^ (Isabel Mufioz, pers. comm.).This low density of macroinvertebrates is partially
© 1995 BlackweU Science Ltd, Freshwater Biology, 33, 373-383
3.5 1
3
ISfe 2.5
Fig. 6 Shannon - Weiner index ofdiversity calculated from algal countsin the control (CC) and experimental(EC) during winter (w) and summer (g
1.5
Enrichment effects in stream biofilms 381
CCs
CCw
4
Week
explained by the extreme climate to which the faunais subjected, including drying out in late summer andthe freezing for some of the winter.
In summary, Pmax*^ was higher in the experimentalchannel in the low production period, and this wasassociated with the substitution of the establishedalgal community by one with higher photosyntheticcapacity. Biomass increase was also low and self-shading was probably insignificant. However, in sum-mer Pmax' ' ' decreased after the enrichment, eventhough algal biomass and metabolism were high.Higher biomass in this period presumably led tosignificant level of self-shading, which may have pre-vented an increase in photosynthetic capacity. Theapparent response of the algal community to nutrientincrease may assist our understanding of how dis-turbed stretches respond to nutrient inputs (which inmost cases exceed the nutrient uptake capacity). Whilea new input of nutrients is likely to produce a changetowards a more nutrient-tolerant, highly efficient com-munity in the low production period (winter), anypotential increase in photosynthetic capacity wouldlikely be overweighed by the increase in biomass inthe high production period (summer).
Current management of rivers in semi-arid coun-tries, including the Mediterranean region, favours adiversion of the water from the river bed towardscirculating channels used for hydroelectric power pro-duction and irrigation. Among other considerations,this reduction in flow will promote high biomassaccumulation in the river bed (Sabater & Sabater 1992).Maximum biomass accumulation (Jackson & Fisher,1989), primary production and secondary production
© 1995 Blackwell Science Ltd, Freshwater Biology, 33, 373-383
(Grimm & Fisher, 1989), have also been reported indesert streams during low-flow episodes. Our resultssuggest that this biomass accumulation may depressthe photosynthetic capacity and thus reduce the poten-tial nutrient uptake capacity of algae, decreasing thecapacity for water amelioration.
Acknowledgments
This work was funded by the CICYT (project numberNAT89-0886) of the Spanish Education and ScienceMinistry. H. Guasch and E. Marti were recipient ofFPI grants from the Spanish Government. We aregrateful to R. Bartra for assistance with field samplingand Isabel Mufioz for allowing us to use her unpub-lished data on the macroinvertebrate fauna of LaSolana. The final version of the paper benefited fromuseful comments of Professor A. Hildrew and of ananonymous referee.
References
American Public Health Association (1991) StandardMethods for the Examination of Water and Wasteivater,18th edn. APHA, Wasliington, DC.
Boston H.L. & Hill W.R. (1991) Photosynthesis-lightrelations of stream periphyton communities. Limnologyand Oceanography, 36, 644-656.
Bushong S.J. & Bachmann R.W. (1989) In situ nutrientenrichment experiments with periphyton inagricultural streams. Hydrobiologia, 178, 1-10.
Carrick C.J. & Lowe R.L. (1989) Benthic algal response

382 H. Guasch, E. Marti and Sergi Sabater
to N and P enrichment along a pH gradient,Hydrobiologia, 179, 119-127.
DeNicola D.M. & Mclntire CD. (1991) Effects of hydraulicrefuge and Irradiance on Grazer-Periphytoninteractions in laboratory streams, journal of the NorthAmerican Benthological Society, 10, 251-262.
Downes M.T, Hrstich L. & Vincent W.R (1993) Extractionof chlorophyll and carotenoid pigments from antarticbenthic mats for analysis by HPLC. journal of AppliedPhycology, 5, 623-^28.
Duncan S.W. & Blinn D.W. (1989) Importance of physicalvariables on the seasonal dynamics of epilithic algaein a highly shaded canyon stream, journal of Phycology,25, 455-461.
Edwards R.W. & Owen M. (1962) The effect of plants onriver conditions. IV. The Oxygen Balance of a chalkstream, journal of Ecology, 50, 207-220.
Fairchild G.W., Lowe R.L. & Richardson W.B. (1985) Algalperiphyton growth on nutrient-diffusing substrates: anin situ bioassay. Ecology, 66, 465-472.
Falkowski PG. (1981) Light shade adaptations andassimilation numbers, journal of Plankton Research, 3,203-216.
Freeman C, Gresswell R., Guasch H., Hudson J., LockM.A., Reynolds B., Sabater S. & Sabater F (1994) Therole of drought in the impact of climatic change on themicrobiota of peatland streams. Freshwater Biology, 32,223-230.
Grimm N, & Fisher S.G. (1989) Stability of periphytonand macroinvertebrates to disturbance by flash floodsin a desert stream, journal of the North AmericanBenthological Society, 8, 293-307.
Guasch H, & Sabater S. (1994) Primary production ofepilithic communities in undisturbed mediterraneanstreams. Verhenlungen der Intemationale Vereinigung fiirtheoretische und Angewandte Limnologie, 25, 1761-1764.
Harris G.P. (1980) The measurement of phytopiankton innatural populations of phytopiankton. The Physiologicalecology of phytopiankton (ed. I. Morris), pp. 129-187,Blakwell Scientific Publications, Oxford.
Hart D.D. & Robinson CT. (1990) Resource limitaHon ina stream community: phosphorus enrichment effectson periphyton and grazers. Ecology, 71, 1494-1502.
Hawes I. (1993) Photosynthesis in thick cyanobacterialfilms: a comparison of annual and perennial antarcticmat communities. Hydrobiologia, 252, 203-209.
Hill W.R. & Knight A.W. (1988) Nutrient and lightlimitation of algae in two northern Califomia streams.journal ofPhycology, 24, 125-132.
Jackson J.K. & Fisher S.G. {1989) Secondary production,emergence, and export of Aquatic insects of SonoranDesert streams. Ecology, 67, 629-638.
Lock M.A, Ford TE., Hullar M.A.J., Kaufman M., Vestal
J.R., Volk G.S. & Ventullo R.M. (1990) Phosphoruslimitation in an Arctic river biofilm - A wholeecosystem experiment. Water Research, 24, 1545-1549.
Lorenzen CJ, (1967) Determination of chlorophyll andpheo-pigments: spectrophotometric equations.Limnology and Oceanography, 12, 343-346.
Lowe R.L., Goliaday S.W. & Webster J.R. (1986)Periphyton response to nutrient manipulation instreams draining clearcut and forested watersheds.journal of the North American Benthologicai Society, 5,221-229.
Marti E., Sabater F. & Armengol J. (1994) Day and nightnutrient uptake differences in a calcareous stream.Verhenlungen der Internationale Vereinigung fiirtheoretische und Angewandte Limnologie, 25, 1756-1760.
Marti E. & Sabater, F Seasonal phosphate and ammoniumuptake kinetics in natural epilithic community of amediterranean stream. Submitted to the journal of theNorth American Benthological Society.
Odum H.T. (1956) Primary production in flowing waters.Limnology and Oceanography, 1, 102-117.
Peterson B.J., Hobbie J.E., Corliss T.L., Friet K, (1983) Acontinuous-flow periphyton bioassay: tests of nutrientlimitation in a tundra stream. Limnology andOceanography, 28, 583-591.
Pringle C. (1986) Effects of water and substratum nutrientsupplies on lotic periphyton growth: an integralbioassay. Canadian Journal of Fisheries and AquaticSciences, 44, 619-629.
Reynolds C.S. (1984) Flie Ecology of FreshwaterPhytopiankton. Cambridge University Press,Cambridge.
Robarts R.D., Evans M.S. & Arts M.T. {1992) Light,nutrients and water temperature as determinants ofphytoptankton production in two saline prairie lakeswith high sulphate concentrations. Canadian journal ofFisheries and Aquatic Sciences, 49, 2281-2290,
Rhee G.Y. & Gothman I.J. (1981) The effect ofenvironmental factors on phytopiankton growth:Temperature and the interactions of temperature withnutrient limitation. Limnology & Oceanography, 26,635-648.
Sabater F. {1987) Estudi integrat del riu Ter i la seva conca:caracteristiques de I'aigua i els factors que les determinen.PhD thesis. University of Barcelona, Barcelona, Spain.
Sabater F, Guasch H., Marti E., Armengol J. & Sabater S.(in press) The Ter: a mediterranean river case-study inSpain. River and Stream Ecosystems. Ecosystems of theWorld, Vol. 22. {Eds C.E. Cushing, F Cummins &G.W, Minshall), pp. 371-390. Elsevier, Amsterdam.
Sabater S. (1988) Composition and seasonal variations ofthe chlorophyllic pigments in phytobenthos
© 1995 Blackweil Science Ltd, Freshwater Biology, 33, 373-383

assemblages of the Ter river along a nine-month period.Oecologia Aquatica, 9, 61-75.
Sabater S. (1989) Encrusting assemblages in aMediterranean river basin. Archiv fiir HydrobiologieErgebnisse der Limnologie, 114, 555-573.
Sabater S., Armengol J., Marti E., Sabater F. & Guasch H.(1991) Benthic diatom communities as descriptors ofdiscontinuities in the river Ter, Spain. Use of Algae forMonitoring Rivers (eds A. Whitton, E. Rott and G.Friedrich), pp. 157-163. Institut fiir Botanik, UniversitatInnsbruck.
Sabater S. & Sabater F (1992) Longitudinal changes ofbenthic algal biomass in a mediterranean river duringtwo high production periods. Archiv fUr HydrobiologieErgebnisse der Limnologie, 124, 475-487.
Triska F.J., Kennedy VC, Avanzino RJ. & Reilly B.N.(1983) Effect of simulated canopy cover on regulationof Nitrate uptake and primary production by naturalperiphyton assemblages. Dynamics of Lotic Ecosystems(eds T. D. Fontaine and S. M. Bartell), pp. 129-160.Ann Arbor Science Publishers, Michigan.
Triska F.J., Kennedy V.C., Avanzino R.J., Zellweger G.W.
Enrichment effects in stream biofilms 383
& Bencala K.E. (1989) Retention and transport ofnutrients in a third-order stream; channel processes.Ecology, 70, 1893-1905.
Thyssen N., Eriandsen M., Jeppesen E. & Ursin C. (1987)Reaeration of oxygen in shallow, macrophyte richstreams: Determination of the reaeration ratecoefficient. Internationale Reime der gesamtenHydrobiologie, 72, 405-429.
Vincent W.F & Howard-Williams C. (1986) Antarcticstream ecosystems: physiological ecology of blue-greenalgal epilithon. Freshwater Biology. 16, 219-233.
Ward A.K., Dahm CN. & Cummins K.W. (1985) Nostoc(Cyanophyta) productivity in Oregon streamecosystems: invertebrate influences and differencesbetween morphological types. Journal of Phycotogy, 21,223-247.
Winterbourn M.J. (1990) Interactions among nutrients,algae and invertebrates in a New Zealand mountainstream. Freshwater Biology. 23, 463-474.
(Manuscript accepted 10 November 1994)
© 1995 Blackwell Science Ltd, Freshwater Biology. 33, 373-383

Related Documents








![Session M1 Biofilm Reactors- Operational Overview and Applications for Nutrient Removal[1]](https://static.cupdf.com/doc/110x72/55cf92c2550346f57b994a4c/session-m1-biofilm-reactors-operational-overview-and-applications-for-nutrient.jpg)


