Handedness and Asymmetry of Hand Representation in Human Motor Cortex J. VOLKMANN, A. SCHNITZLER, O. W. WITTE, AND H.-J. FREUND Department of Neurology, Heinrich-Heine University, D-40225 Duesseldorf, Germany Volkmann, J., A. Schnitzler, O. W. Witte, and H.-J. Freund. to detect consistent anatomic asymmetries in these studies Handedness and asymmetry of hand representation in human motor may be due to the lack of historical information about hand- cortex. J. Neurophysiol. 79: 2149–2154, 1998. The cortical repre- edness in postmortem studies and the inaccuracy of delineat- sentation of five simple hand and finger movements in the human ing hand function in these structures based on anatomic motor cortex was determined in left- and right-handed people with landmarks. whole-head magnetoencephalography. Different movements were Functional neuroimaging techniques now exist to measure found to be represented by spatially segregated dipolar sources in the cortical representation of skilled movements noninva- primary motor cortex. The spatial arrangement of neuronal sources sively with high spatial accuracy. Here we report in vivo for digit and wrist movements was nonsomatotopic and varied measurements of hand movement representation in left- and greatly between subjects. As an estimator of hand area size in primary motor cortex, we determined the smallest cuboid volume right-handed subjects using whole-head magnetoencepha- enclosing the five dipole sources within the left and right hemi- lography (MEG). We assessed possible functional asym- sphere of each subject. Interhemispheric comparison revealed a metries of primary motor cortex in the context of individual significant increase of this volume in primary motor cortex opposite hand preference and performance. to the preferred hand. This asymmetry was due to a greater spatial segregation of neuronal dipole generators subserving different hand and finger actions in the dominant hemisphere. Mean Euclidean METHODS distances between dipole sources for different movements were Movement-related neuromagnetic fields were recorded with the 10.7 { 3.5 mm in the dominant and 9.4 { 3.5 mm in the nondomi- 122-channel Neuromag MEG system in 10 healthy, male subjects nant hemisphere ( mean { SD; P Å 0.01, two-tailed t-test). The ( 5 right-handed subjects, age: 34.0 { 6.1 yr ; 5 left-handed subjects, expansion of hand representation in primary motor cortex could age: 28.2 { 2.5 yr) for five simple finger and hand movements: not simply be attributed to a greater number of pyramidal cells flexion of the distal phalanx of the thumb, index finger abduction, devoted to each particular movement as inferred from current index finger extension, little finger abduction, and wrist flexion. source amplitudes. The degree of hemispheric asymmetry of hand All subjects showed consistent hand preference in every-day activi- area size in the primary motor cortex was correlated highly with ties as assessed by a handedness questionnaire (Witelson 1989) the asymmetry of hand performance in a standardized handedness and compatible asymmetry of hand performance in a standardized test ( r Å00.76, P õ 0.01). These results demonstrate for the first hand dominance test (Ja ¨ncke 1996). Surface electromyograms time a biological correlate of handedness in human motor cortex. (EMG) of the musculi extensor indicis (ei), interosseus dorsalis The expansion of hand motor cortex in the dominant hemisphere 1 ( id1 ) , flexor pollicis longus ( fpl ) , abductor digiti minimi ( adm) , may provide extra space for the cortical encoding of a greater and flexor carpi ulnaris (fcu), vertical electrooculogram (EOG), motor skill repertoire of the preferred hand. and neuromagnetic activity (band-pass 0.01–100 Hz, sample rate 512 Hz) were recorded simultaneously. The subjects were seated inside a magnetically shielded room in a nonmagnetic chair with INTRODUCTION both forearms supported by an armrest. For index abduction, index flexion, thumb flexion, and little finger abduction both hands were Hand preference is the most prominent behavioral indica- resting on the volar side with all fingers outstretched on the arm tor for hemispheric specialization in humans. About 90% support. For wrist flexions, both hands were supinated and lying of humans are right-handed and therefore left-hemisphere on the dorsal side with all fingers passively flexed and the forearm dominant for manual skills. The neurobiological correlate fully supported. These positions ensured a minimal muscular effort of handedness, however, is still a matter of debate. Some for maintaining hand posture throughout a trial. Subjects were investigators have attributed handedness to a possible hemi- instructed to perform brisk isolated movements from complete re- laxation approximately once every 4 s, alternating between hands. spheric asymmetry of cortical association areas controlling No pacing signal was provided. Subjects were free in choosing the the cognitive-motor requirements of skilled movements starting hand. Feedback about performance based on surface EMG (Haaland and Harington 1996). Others have argued that was given during a short initial training period. By visual inspec- hand preference should be reflected by functional or struc- tion, the examiner moreover controlled for involuntary cocontrac- tural asymmetries of the primary motor cortex and descend- tions of other limb parts and corrected motor performance during ing pathways (Amunts et al. 1996; Nathan et al. 1990; White the training period if necessary. Each subject participated in three et al. 1997). But conclusive evidence has not been found recording sessions on different days during a 3-wk period in which for either view. Recent studies have revealed conflicting the sequence of the five movement conditions was randomized. results about structural hemispheric differences at the level During each recording session, a total of 100 – 120 movements was of primary motor cortex and pyramidal tract (Amunts et al. collected for each hand and condition. We made sure that each subject kept the hand posture constant for the individual move- 1996; Nathan et al. 1990; White et al. 1997). The failure 2149 0022-3077/98 $5.00 Copyright q 1998 The American Physiological Society J800-7 / 9k27$$ap07 03-18-98 20:08:34 neupa LP-Neurophys

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Handedness and Asymmetry of Hand Representation in Human MotorCortex
J. VOLKMANN, A. SCHNITZLER, O. W. WITTE, AND H.-J. FREUNDDepartment of Neurology, Heinrich-Heine University, D-40225 Duesseldorf, Germany
Volkmann, J., A. Schnitzler, O. W. Witte, and H.-J. Freund. to detect consistent anatomic asymmetries in these studiesHandedness and asymmetry of hand representation in human motor may be due to the lack of historical information about hand-cortex. J. Neurophysiol. 79: 2149–2154, 1998. The cortical repre- edness in postmortem studies and the inaccuracy of delineat-sentation of five simple hand and finger movements in the human ing hand function in these structures based on anatomicmotor cortex was determined in left- and right-handed people with landmarks.whole-head magnetoencephalography. Different movements were
Functional neuroimaging techniques now exist to measurefound to be represented by spatially segregated dipolar sources inthe cortical representation of skilled movements noninva-primary motor cortex. The spatial arrangement of neuronal sourcessively with high spatial accuracy. Here we report in vivofor digit and wrist movements was nonsomatotopic and variedmeasurements of hand movement representation in left- andgreatly between subjects. As an estimator of hand area size in
primary motor cortex, we determined the smallest cuboid volume right-handed subjects using whole-head magnetoencepha-enclosing the five dipole sources within the left and right hemi- lography (MEG). We assessed possible functional asym-sphere of each subject. Interhemispheric comparison revealed a metries of primary motor cortex in the context of individualsignificant increase of this volume in primary motor cortex opposite hand preference and performance.to the preferred hand. This asymmetry was due to a greater spatialsegregation of neuronal dipole generators subserving different handand finger actions in the dominant hemisphere. Mean Euclidean M E T H O D Sdistances between dipole sources for different movements were
Movement-related neuromagnetic fields were recorded with the10.7 { 3.5 mm in the dominant and 9.4 { 3.5 mm in the nondomi-122-channel Neuromag MEG system in 10 healthy, male subjectsnant hemisphere (mean { SD; P Å 0.01, two-tailed t-test) . The(5 right-handed subjects, age: 34.0{ 6.1 yr; 5 left-handed subjects,expansion of hand representation in primary motor cortex couldage: 28.2 { 2.5 yr) for five simple finger and hand movements:not simply be attributed to a greater number of pyramidal cellsflexion of the distal phalanx of the thumb, index finger abduction,devoted to each particular movement as inferred from currentindex finger extension, little finger abduction, and wrist flexion.source amplitudes. The degree of hemispheric asymmetry of handAll subjects showed consistent hand preference in every-day activi-area size in the primary motor cortex was correlated highly withties as assessed by a handedness questionnaire (Witelson 1989)the asymmetry of hand performance in a standardized handednessand compatible asymmetry of hand performance in a standardizedtest (r Å 00.76, P õ 0.01). These results demonstrate for the firsthand dominance test (Jancke 1996). Surface electromyogramstime a biological correlate of handedness in human motor cortex.(EMG) of the musculi extensor indicis (ei) , interosseus dorsalisThe expansion of hand motor cortex in the dominant hemisphere1 (id1), flexor pollicis longus (fpl) , abductor digiti minimi (adm),may provide extra space for the cortical encoding of a greaterand flexor carpi ulnaris (fcu) , vertical electrooculogram (EOG),motor skill repertoire of the preferred hand.and neuromagnetic activity (band-pass 0.01–100 Hz, sample rate512 Hz) were recorded simultaneously. The subjects were seatedinside a magnetically shielded room in a nonmagnetic chair withI N T R O D U C T I O Nboth forearms supported by an armrest. For index abduction, indexflexion, thumb flexion, and little finger abduction both hands wereHand preference is the most prominent behavioral indica-resting on the volar side with all fingers outstretched on the armtor for hemispheric specialization in humans. About 90% support. For wrist flexions, both hands were supinated and lying
of humans are right-handed and therefore left-hemisphere on the dorsal side with all fingers passively flexed and the forearmdominant for manual skills. The neurobiological correlate fully supported. These positions ensured a minimal muscular effortof handedness, however, is still a matter of debate. Some for maintaining hand posture throughout a trial. Subjects wereinvestigators have attributed handedness to a possible hemi- instructed to perform brisk isolated movements from complete re-
laxation approximately once every 4 s, alternating between hands.spheric asymmetry of cortical association areas controllingNo pacing signal was provided. Subjects were free in choosing thethe cognitive-motor requirements of skilled movementsstarting hand. Feedback about performance based on surface EMG(Haaland and Harington 1996) . Others have argued thatwas given during a short initial training period. By visual inspec-hand preference should be reflected by functional or struc-tion, the examiner moreover controlled for involuntary cocontrac-tural asymmetries of the primary motor cortex and descend-tions of other limb parts and corrected motor performance duringing pathways (Amunts et al. 1996; Nathan et al. 1990; White the training period if necessary. Each subject participated in three
et al. 1997). But conclusive evidence has not been found recording sessions on different days during a 3-wk period in whichfor either view. Recent studies have revealed conflicting the sequence of the five movement conditions was randomized.results about structural hemispheric differences at the level During each recording session, a total of 100–120 movements wasof primary motor cortex and pyramidal tract (Amunts et al. collected for each hand and condition. We made sure that each
subject kept the hand posture constant for the individual move-1996; Nathan et al. 1990; White et al. 1997) . The failure
21490022-3077/98 $5.00 Copyright q 1998 The American Physiological Society
J800-7/ 9k27$$ap07 03-18-98 20:08:34 neupa LP-Neurophys
J. VOLKMANN, A. SCHNITZLER, O. W. WITTE, AND H.-J. FREUND2150
For source localization, a spherical head model was fitted to theindividual magnetic resonance image (MRI; Siemens-Magnetom1.5 Tesla, T1–3D-Flash sequence) after alignment of the MRI andMEG coordinate system based on fiducial point markers. Softwaretools provided by Neuromag were used for the segmentation of MRimages and computation of three-dimensional surface renderings.
To quantify hemispheric asymmetries in the spatial extent ofhand representation in primary motor cortex, we computed thesmallest cuboid volume containing all contralateral dipole sourcesof the investigated movements in either hemisphere. Each side ofthe cuboid was defined by the maximal Euclidean distance betweendipole sources within each spatial direction of the MEG coordinatesystem. A hemispheric asymmetry index (R L/R / L) then wascalculated for each subject expressing the proportional differenceof hand area volume in the left (L) and right hemisphere (R).Negative values indicate left and positive values indicate righthemisphere dominance.
In a second analytic approach, we also investigated the func-tional activation of ipsilateral motor cortex to determine the degreeof hemispheric asymmetry of motor output activity during unilat-eral hand movements. The contribution of ipsilateral motor cortexactivity to each movement was investigated using the followingapproach: after calculating the optimal source locations for the
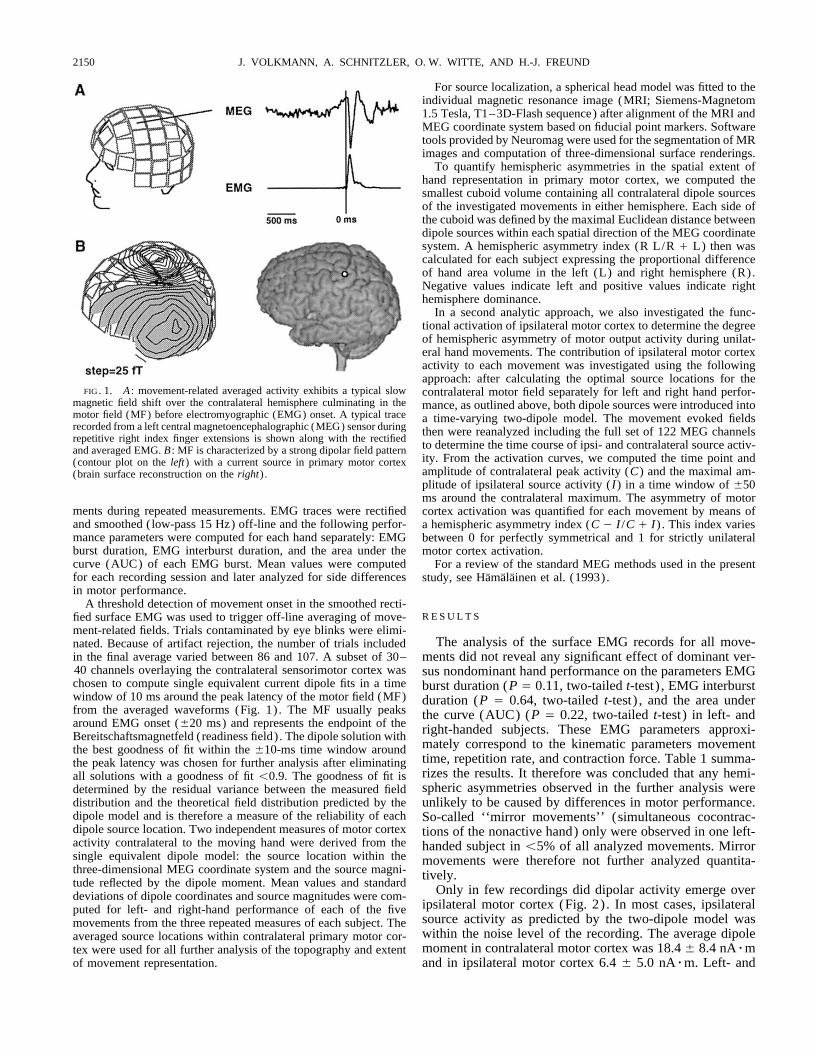
FIG. 1. A : movement-related averaged activity exhibits a typical slow contralateral motor field separately for left and right hand perfor-magnetic field shift over the contralateral hemisphere culminating in the mance, as outlined above, both dipole sources were introduced intomotor field (MF) before electromyographic (EMG) onset. A typical trace a time-varying two-dipole model. The movement evoked fieldsrecorded from a left central magnetoencephalographic (MEG) sensor during then were reanalyzed including the full set of 122 MEG channelsrepetitive right index finger extensions is shown along with the rectified
to determine the time course of ipsi- and contralateral source activ-and averaged EMG. B : MF is characterized by a strong dipolar field patternity. From the activation curves, we computed the time point and(contour plot on the left) with a current source in primary motor cortexamplitude of contralateral peak activity (C) and the maximal am-(brain surface reconstruction on the right) .plitude of ipsilateral source activity (I) in a time window of {50ms around the contralateral maximum. The asymmetry of motor
ments during repeated measurements. EMG traces were rectified cortex activation was quantified for each movement by means ofand smoothed (low-pass 15 Hz) off-line and the following perfor- a hemispheric asymmetry index (C 0 I /C / I) . This index variesmance parameters were computed for each hand separately: EMG between 0 for perfectly symmetrical and 1 for strictly unilateralburst duration, EMG interburst duration, and the area under the motor cortex activation.curve (AUC) of each EMG burst. Mean values were computed For a review of the standard MEG methods used in the presentfor each recording session and later analyzed for side differences study, see Hamalainen et al. (1993).in motor performance.
A threshold detection of movement onset in the smoothed recti-R E S U L T Sfied surface EMG was used to trigger off-line averaging of move-
ment-related fields. Trials contaminated by eye blinks were elimi-The analysis of the surface EMG records for all move-nated. Because of artifact rejection, the number of trials included
in the final average varied between 86 and 107. A subset of 30– ments did not reveal any significant effect of dominant ver-40 channels overlaying the contralateral sensorimotor cortex was sus nondominant hand performance on the parameters EMGchosen to compute single equivalent current dipole fits in a time burst duration (P Å 0.11, two-tailed t-test) , EMG interburstwindow of 10 ms around the peak latency of the motor field (MF) duration (P Å 0.64, two-tailed t-test) , and the area underfrom the averaged waveforms (Fig. 1) . The MF usually peaks the curve (AUC) (P Å 0.22, two-tailed t-test) in left- andaround EMG onset ({20 ms) and represents the endpoint of the right-handed subjects. These EMG parameters approxi-Bereitschaftsmagnetfeld (readiness field) . The dipole solution with
mately correspond to the kinematic parameters movementthe best goodness of fit within the {10-ms time window aroundtime, repetition rate, and contraction force. Table 1 summa-the peak latency was chosen for further analysis after eliminatingrizes the results. It therefore was concluded that any hemi-all solutions with a goodness of fit õ0.9. The goodness of fit isspheric asymmetries observed in the further analysis weredetermined by the residual variance between the measured field
distribution and the theoretical field distribution predicted by the unlikely to be caused by differences in motor performance.dipole model and is therefore a measure of the reliability of each So-called ‘‘mirror movements’’ (simultaneous cocontrac-dipole source location. Two independent measures of motor cortex tions of the nonactive hand) only were observed in one left-activity contralateral to the moving hand were derived from the handed subject in õ5% of all analyzed movements. Mirrorsingle equivalent dipole model: the source location within the movements were therefore not further analyzed quantita-three-dimensional MEG coordinate system and the source magni- tively.tude reflected by the dipole moment. Mean values and standard Only in few recordings did dipolar activity emerge overdeviations of dipole coordinates and source magnitudes were com-
ipsilateral motor cortex (Fig. 2) . In most cases, ipsilateralputed for left- and right-hand performance of each of the fivesource activity as predicted by the two-dipole model wasmovements from the three repeated measures of each subject. Thewithin the noise level of the recording. The average dipoleaveraged source locations within contralateral primary motor cor-moment in contralateral motor cortex was 18.4 { 8.4 nArmtex were used for all further analysis of the topography and extent
of movement representation. and in ipsilateral motor cortex 6.4 { 5.0 nArm. Left- and
J800-7/ 9k27$$ap07 03-18-98 20:08:34 neupa LP-Neurophys

HANDEDNESS AND MOTOR CORTEX ASYMMETRY 2151
TABLE 1. Summary of motor parameters
Movement Typeand Muscle Burst Duration, s Interburst Duration, s Area Under the Curve
Dominant hand adm 0.61 { 0.27 4.36 { 1.26 0.83 { 0.40ei 0.36 { 0.14 4.73 { 1.40 0.34 { 0.20fcu 0.42 { 0.14 4.96 { 1.83 0.52 { 0.22fpl 0.32 { 0.17 4.88 { 1.52 0.32 { 0.16idl 0.76 { 0.96 4.46 { 1.46 1.10 { 0.51
Nondominant hand adm 0.57 { 0.23 4.60 { 1.22 0.82 { 0.31ei 0.32 { 0.13 4.71 { 1.11 0.26 { 0.14fcu 0.42 { 0.20 4.94 { 1.56 0.56 { 0.30fpl 0.28 { 0.14 5.13 { 1.61 0.31 { 0.14idl 0.52 { 0.27 4.66 { 1.36 0.88 { 0.40
This table summarizes the motor performance parameters obtained from surface electromyographic analysis separated by movement type and muscle:adm (m. abductor digiti minimi), little finger abduction; ei (m. extensor indicis), index extension; fcu (m. flexor carpi ulnaris), wrist flexion; fpl (m.flexor pollicis longus), thumb flexion; and idl (m. interosseus dorsalis 1), index abduction. Values are means { SD.
right handed subjects did not differ significantly in the hemi- vidual high resolution magnetic resonance scans of the brain.Motor output maps had a low intraindividual variability inspheric asymmetry of motor cortical output during unilateral
movements as determined by the hemispheric asymmetry repeated measurements with an SEõ 1.5 mm in each spatialdirection. Figure 3 shows typical examples of hand motorindex of the dipole moment (P Å 0.73; two-tailed t-test) .
Pooled data of both groups, however, revealed a statistical representation in a right-handed (RH) and a left-handed(LH) subject. These cases illustrate the three characteristictendency for a stronger activation of ipsilateral motor cortex
during movements of the nonpreferred hand (mean asymme- features of motor output organization in M1 that were ob-served in all subjects. 1) Different digit and wrist movementstry index 0.49 { 0.24) than during movements of the pre-
ferred hand (mean asymmetry index 0.54 { 0.26; P Å 0.09; were represented by spatially segregated neuronal sources inprimary motor cortex. Across subjects 87% of all interdipoletwo-tailed t-test) .
For each subject, we reconstructed the topographic organi- distances in the nondominant hemisphere and 89% in thedominant hemisphere were significant (95% lower confi-zation of hand and finger movement representation in pri-
mary motor cortex by projecting equivalent current dipole dence limit of the Euclidean interdipole distance ú0 in re-peated measurements) . 2) The spatial arrangement of neu-sources of contralateral motor output activity into the indi-
FIG. 2. Examples of the time course of ipsilateral andcontralateral motor cortex activation during the abduction ofthe right little finger (adm) in 2 right-handed subjects. A :ipsilateral motor cortex activity is apparent in this subject bya weaker dipolar field pattern over the ipsilateral hemisphere.Ipsilateral field pattern appears slightly asymmetric due tothe interaction with coinciding activation of the other hemi-sphere. Time course of the ipsilateral dipole moment isroughly parallel to the contralateral source. A gray bar under-lying the activation curves indicates the time window of {50ms around the contralateral source maximum, which we usedto determine the maximal ipsilateral source activity. B : inthe majority of recordings, however, no dipolar field patternemerged over the ispilateral hemisphere as demonstrated inthis case. Goodness of fit of the 2-dipole model is displayedas a function of time in the lowest chart.
J800-7/ 9k27$$ap07 03-18-98 20:08:34 neupa LP-Neurophys
J. VOLKMANN, A. SCHNITZLER, O. W. WITTE, AND H.-J. FREUND2152
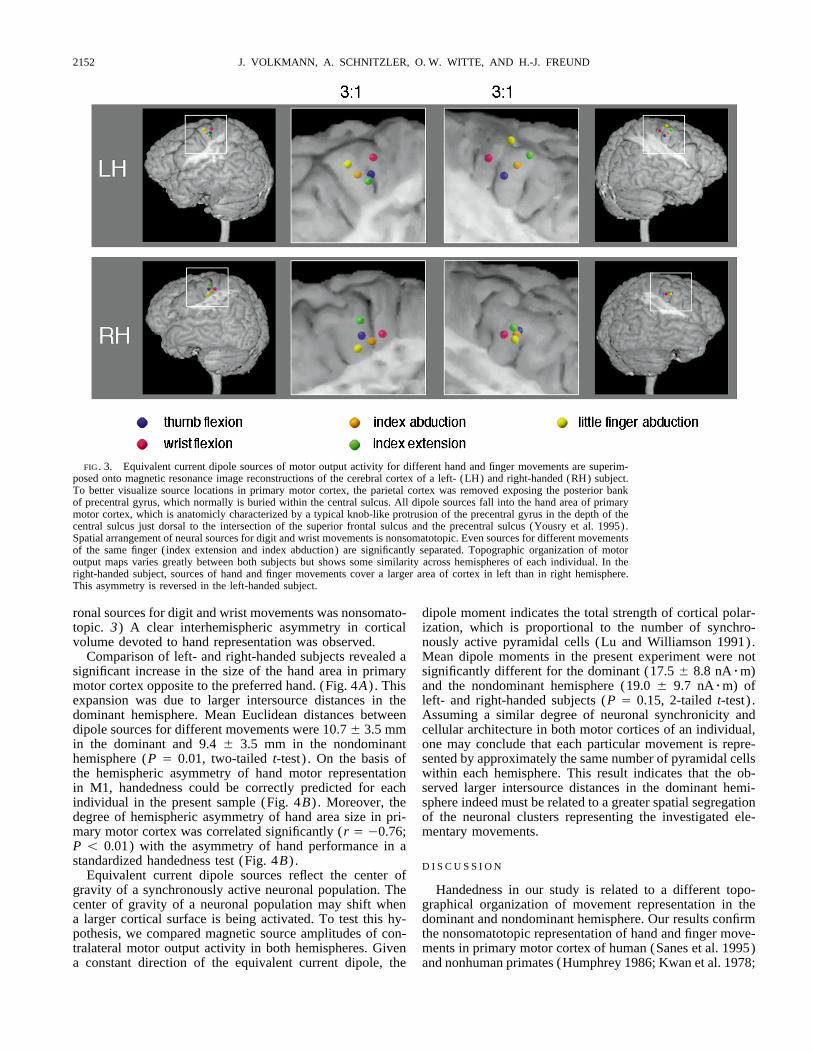
FIG. 3. Equivalent current dipole sources of motor output activity for different hand and finger movements are superim-posed onto magnetic resonance image reconstructions of the cerebral cortex of a left- (LH) and right-handed (RH) subject.To better visualize source locations in primary motor cortex, the parietal cortex was removed exposing the posterior bankof precentral gyrus, which normally is buried within the central sulcus. All dipole sources fall into the hand area of primarymotor cortex, which is anatomicly characterized by a typical knob-like protrusion of the precentral gyrus in the depth of thecentral sulcus just dorsal to the intersection of the superior frontal sulcus and the precentral sulcus (Yousry et al. 1995).Spatial arrangement of neural sources for digit and wrist movements is nonsomatotopic. Even sources for different movementsof the same finger (index extension and index abduction) are significantly separated. Topographic organization of motoroutput maps varies greatly between both subjects but shows some similarity across hemispheres of each individual. In theright-handed subject, sources of hand and finger movements cover a larger area of cortex in left than in right hemisphere.This asymmetry is reversed in the left-handed subject.
ronal sources for digit and wrist movements was nonsomato- dipole moment indicates the total strength of cortical polar-ization, which is proportional to the number of synchro-topic. 3) A clear interhemispheric asymmetry in cortical
volume devoted to hand representation was observed. nously active pyramidal cells (Lu and Williamson 1991).Mean dipole moments in the present experiment were notComparison of left- and right-handed subjects revealed a
significant increase in the size of the hand area in primary significantly different for the dominant (17.5 { 8.8 nArm)and the nondominant hemisphere (19.0 { 9.7 nArm) ofmotor cortex opposite to the preferred hand. (Fig. 4A) . This
expansion was due to larger intersource distances in the left- and right-handed subjects (P Å 0.15, 2-tailed t-test) .Assuming a similar degree of neuronal synchronicity anddominant hemisphere. Mean Euclidean distances between
dipole sources for different movements were 10.7 { 3.5 mm cellular architecture in both motor cortices of an individual,one may conclude that each particular movement is repre-in the dominant and 9.4 { 3.5 mm in the nondominant
hemisphere (P Å 0.01, two-tailed t-test) . On the basis of sented by approximately the same number of pyramidal cellswithin each hemisphere. This result indicates that the ob-the hemispheric asymmetry of hand motor representation
in M1, handedness could be correctly predicted for each served larger intersource distances in the dominant hemi-sphere indeed must be related to a greater spatial segregationindividual in the present sample (Fig. 4B) . Moreover, the
degree of hemispheric asymmetry of hand area size in pri- of the neuronal clusters representing the investigated ele-mentary movements.mary motor cortex was correlated significantly (r Å 00.76;
P õ 0.01) with the asymmetry of hand performance in astandardized handedness test (Fig. 4B) . D I S C U S S I O N
Equivalent current dipole sources reflect the center ofgravity of a synchronously active neuronal population. The Handedness in our study is related to a different topo-
graphical organization of movement representation in thecenter of gravity of a neuronal population may shift whena larger cortical surface is being activated. To test this hy- dominant and nondominant hemisphere. Our results confirm
the nonsomatotopic representation of hand and finger move-pothesis, we compared magnetic source amplitudes of con-tralateral motor output activity in both hemispheres. Given ments in primary motor cortex of human (Sanes et al. 1995)
and nonhuman primates (Humphrey 1986; Kwan et al. 1978;a constant direction of the equivalent current dipole, the
J800-7/ 9k27$$ap07 03-18-98 20:08:34 neupa LP-Neurophys

HANDEDNESS AND MOTOR CORTEX ASYMMETRY 2153
source with a radius of Ç8 mm. Given the observed inter-source distances in the present study and the limited numberof movements investigated, a significant degree of overlapmust indeed be assumed. From a bird’s eye perspective astaken by MEG, our finding of topographically organizedneuronal clusters in M1 subserving different hand and fingeractions supports the concept of a functional grouping ofdifferent muscle representations at a cortical site into syner-gies that make kinesiological sense (Humphrey 1986; Nudoet al. 1992). In such an arrangement, an overlap of differentmovement representations would be economical by sharingmuscle representations that are used as common elementsin different synergies.
Our finding of an enlarged hand representation in motorcortex opposite to the preferred hand is in agreement withprevious studies using transcranial magnetic stimulation inhumans. Triggs et al. (1994) described a lateralization ofmotor cortex excitability, which correlated with handedness.Wassermann et al. (1992) investigated the cortical topogra-phy of the abductor pollicis brevis muscle using focal mag-netic stimulation with a figure-eight–shaped coil and founda larger area of representation in the dominant hemisphere.A more complete picture of hemispheric differences of motorcortical output maps was obtained by intracortical micro-stimulation in nonhuman primates (Nudo et al. 1992). Nudoet al. related—in complete agreement with our findings inhuman subjects—that individual distal forelimb representa-tions in monkeys were highly idiosyncratic but that the repre-sentational topography varied less between hemispheres ofeach animal. Forelimb representation opposite to the pre-ferred hand of the monkey was generally larger in total area
FIG. 4. A : right-handed subjects exhibited stronger leftward asymmetry and boundary length.of hand motor representation, whereas left-handed subjects showed a com- It is reasonable to assume that the expansion of handparable degree of right-hemispheric dominance as indicated by significant motor cortex in the dominant hemisphere provides the neu-group differences in the asymmetry index of hand area size in motor cortex
ral substrate for a more efficient processing of motor output(P õ 0.0007, 2-tailed t-test) . B : for each subject, an asymmetry index ofto the preferred hand. Higher efficiency can be achievedhand performance (R 0 L/R / L) was computed from the performance
measures in a standardized hand dexterity test (HDT). Scatterplot illustrates through an increase in the total number of elements in athe strong linear correlation (r Å 00.76; P Å 0.0082) between the behav- neural assembly and by a tighter intraareal connectivity.ioral asymmetry of hand performance and the degree of hemispheric asym-
In agreement with this concept, motor output maps oppo-metry of hand area size in primary motor cortex.site to the preferred hand of the squirrel monkey werenot only expanded but also showed a higher degree of
Nudo et al. 1992; Schieber and Hibbard 1993). Intracortical fragmentation and spatial complexity (Nudo et al. 1992) .microstimulation experiments have shown multiple, overlap- Such an increase in spatial complexity may allow for betterping representations of distal and proximal forelimb muscles interaction between neuronal clusters at a cortical site rep-or movements throughout the entire arm area of M1 in mon- resenting muscles that are used in close temporal contigu-keys (Humphrey 1986; Kwan et al. 1978; Nudo et al. 1992). ity. The behavioral consequence of such improved interac-These studies, however, did not provide any insight into how tion may be a more refined motor skill repertoire of thenaturally occurring hand or finger movements make use of preferred hand.such fragmented motor output maps. Even fairly simple In a previous study with functional magnetic resonancemanual skills—such as investigated in the present study— imaging ( fMRI) Kim et al. (1993) described a hemi-involve complex muscle synergies that consist of main ago- spheric asymmetry in the functional activation of the hu-nists for the primary movement and muscles involved in man motor cortex during contralateral and ipsilateral fin-postural stabilization. The present study revealed a signifi- ger movements. Whereas right motor cortex was activatedcant spatial segregation of dipole sources for different hand mostly during contralateral movements, the left motor cor-and finger movements. It is important to emphasize that tex was activated substantially during ipsilateral move-dipole sources reflect the center of gravity of a coherently ments in left-handed subjects and even more so in right-active neuronal population, which may in fact overlap. From handed subjects. These results could not be replicated inmeasured intracortical current densities it has been estimated the present study using neuromagnetic recordings. Wethat a dipole moment of 20 nArm corresponds to cortical found that in both left- and right-handed subjects the con-activation extending over a surface area of Ç200 mm2 (Lu tralateral motor cortex activation was clearly dominant
over the ipsilateral motor cortex activation. There was noand Williamson 1991). This would correspond to a circular
J800-7/ 9k27$$ap07 03-18-98 20:08:34 neupa LP-Neurophys

J. VOLKMANN, A. SCHNITZLER, O. W. WITTE, AND H.-J. FREUND2154
REFERENCESsignificant difference in the degree of ipsilateral motorAMUNTS, K., SCHLAUG, G., SCHLEICHER, A., STEINMETZ, H., DABRINGHAUS,cortex activity during left- or right-hand movements.
A., ROLAND, P. E., AND ZILLES, K. Asymmetry in the human motor cortexPooled data of both handedness groups revealed a ten-and handedness. Neuroimage 4: 216–222, 1996.dency for a stronger contribution of ipsilateral motor cor- ANNETT, M. The growth of manual preference and speed. Br. J. Psychol.
tex to movements of the nonpreferred hand. This finding 61: 545–558, 1970.CARTER-SALTZMAN, L. Biological and sociocultural effects on handedness:rather indicates a differential effect of hand preference on
comparison between biological and adoptive families. Science 209:the hemispheric asymmetry of motor cortical output than1263–1265, 1980.of the hemisphere per se. One may speculate about meth-
FOX, P. W., HERSHBERGER, S. L., AND BOUCHARD, T. J. Genetic and envi-odological differences between both studies. While the ronmental contributions to the acquisition of a motor skill. Nature 384:motor field in MEG most likely reflects the synchronous 356–358, 1996.
HAALAND, K. Y. AND HARINGTON, D. L. Hemispheric asymmetry of move-discharges of pyramidal cells during corticospinal outflowment. Curr. Opin. Neurobiol. 6: 796–800, 1996.from motor cortex, fMRI provides an indirect measure of
HAMALAINEN, M., HARI, R., ILMONIEMI, R. J., KNUUTILA, J., AND LOUNAS-neuronal activity and integrates due to its limited time MAA, O. V. Magnetoencephalography—theory, instrumentation, and ap-resolution over motor cortical outflow and reafferent ac- plication to noninvasive studies of the working human brain. Rev. Modern
Phys. 65: 413–497, 1993.tivity.HUMPHREY, D. R. Representation of movements and muscles within theOur study demonstrates that individual variability in hand
primate precentral motor cortex: historical and current perspectives.motor performance covaries with features of hand movement FASEB J. 45: 2687–2699, 1986.representation in primary motor cortex. It does not explain, JANCKE, L. The hand performance test with a modified time limit instruction
enables the examination of hand performance asymmetries in adults.however, the mechanism underlying this asymmetry. APercept. Mot. Skills 82: 735–738, 1996.short- and long-term plasticity of sensorimotor cortical rep-
KARNI, A., MEYER, G., JEZZARD, P., ADAMS, M. M., TURNER, R., ANDresentations in relation to motor learning and training has UNGERLEIDER, L. G. Functional MRI evidence for adult motor cortexbeen demonstrated in several previous studies (Karni et al. plasticity during motor skill learning. Nature 377: 155–158, 1995.
KIM, S.-G., ASHE, J., HENDRICH, K., ELLERMAN, J. M., MERKLE, H., UGUR-1995; Pascual-Leone et al. 1994). This use-dependent plas-BIL, K., AND GEORGOPOULOS, A. P. Functional magnetic resonance im-ticity has been attributed to changes in cortical excitabilityaging of motor cortex: hemispheric asymmetry and handedness. Sciencethrough the unmasking of preexisting, but unused, synaptic 261: 615–617, 1993.
connections (Pascual-Leone et al. 1994). In contrast, in the KWAN, H. C., MACKAY, W. A., MURPHY, J. T., AND WONG, Y. C. Spatialorganization of precentral cortex in awake primates. II. Motor outputs.present study hand representations in motor cortex were ex-J. Neurophysiol. 41: 1120–1131, 1978.panded due to a shift in the centers of cortical activation.
LU, Z. AND WILLIAMSON, S. J. Spatial extent of coherent sensory-evokedThis finding rather indicates an underlying structural asym- cortical activity. Exp. Brain Res. 84: 411–416, 1991.metry such as described by a recent in vivo morphometry NATHAN, P. W., SMITH, M. C., AND DEACON, P. The corticospinal tracts in
man: course and location of fibres at different segmental levels. Brainstudy, which found a larger surface of dorsolateral motor113: 303–324, 1990.cortex in the dominant hemisphere of left- and right-handed
NUDO, R. J., JENKINS, W. M., MERZENICH, M. M., PREJEAN, T., ANDsubjects (Amunts et al. 1996). GRENDA, R. Neurophysiological correlates of hand preference in primaryIt has been suggested that handedness may be determined motor cortex of adult squirrel monkeys. J. Neurosci. 12: 2918–2947,
1992.genetically (Carter-Saltzman 1980). Motor asymmetries al-PASCUAL-LEONE, A., GRAFMAN, J., AND HALLETT, M. Modulation of corticalready can be observed in neonates, and handedness as de-
motor output maps during development of implicit and explicit knowl-fined by preference and performance scores is measurable edge. Science 263: 1287–1289, 1994.in children as young as 3 yr (Annett 1970). Recently, twin SANES, J. N., DONOGHUE, J. P., THANGARAJ, V., EDELMAN, R. R., AND WAR-
studies revealed the significance of genetic contributions to ACH, S. Shared neural substrates controlling hand movements in humanmotor cortex. Science 268: 1775–1777, 1995.individual differences in motor skill acquisition. Both, motor
SCHIEBER, M. H. AND HIBBARD, L. S. How somatotopic is the motor cortexperformance and rate of learning of a rotatory pursuit taskhand area? Science 261: 489–492, 1993.
were found to be highly heritable (Fox et al. 1996). The TRIGGS, W. J., CALVANIO, R., MACDONNEL, R.A.L., CROS, D., AND CHIAPPA,contribution of environmental factors to the development of K. H. Physiological motor asymmetry in human handedness: evidence
from transcranial magnetic stimulation. Brain Res. 636: 270–276, 1994.hand preference and underlying brain asymmetries thus mayWASSERMANN, E. M., MCSHANE, L. M., HALLETT, M., AND COHEN, L. G.be limited. We therefore would like to suggest that the func-
Noninvasive mapping of muscle representations in human motor cortex.tional asymmetry of hand movement representation in motor Electroencephalogr. Clin. Neurophysiol. 85: 1–8, 1992.cortex, that we have reported here, is more likely a prerequi- WHITE, L. E., ANDREWS, T. J., HULETTE, C., RICHARDS, A., GROELLE, M.,
PAYDARFAR, J., AND PURVES, D. Structure of the human sensorimotorsite than a consequence of handedness.system. I. Morphology and cytoarchitecture of the central sulcus. Cereb.Cortex 7: 18–30, 1997.
WITELSON, S. F. Hand and sex differences in the isthmus and genu of theThis study was supported by Deutsche Forschungsgemeinschaft Grant human corpus callosum. A postmortem morphological study. Brain 112:
SFB194, Z2. 799–835, 1989.Address for reprint requests: J. Volkmann, Dept. of Neurology, Heinrich- YOUSRY, T., SCHMID, U. D., JASSOY, A. G., SCHMIDT, D., EISNER, W. E.,
Heine-University, Moorenstr. 5, D-40225 Duesseldorf, Germany. REULEN, H.-J., REISER, M. F., AND LISSNER, J. Topography of the corticalhand area: prospective study with functional MR imaging and directmotor mapping at surgery. Radiology 195: 23–29, 1995.Received 1 October 1997; accepted in final form 2 December 1997.
J800-7/ 9k27$$ap07 03-18-98 20:08:34 neupa LP-Neurophys
Related Documents











