Original Article NTP-CERHR Expert Panel Report on the Developmental Toxicity of Soy Infant Formula Gail McCarver, 1 Jatinder Bhatia, 2 Christina Chambers, 3 Robert Clarke, 4 Ruth Etzel, 5 Warren Foster, 6 Patricia Hoyer, 7 J. Steven Leeder, 8 Jeffrey M. Peters, 9 Emilie Rissman, 10 Michael Rybak, 11 Claire Sherman, 13 Jorma Toppari, 13 and Katie Turner 14 1 Medical College of Wisconsin, Milwaukee, Wisconsin 2 Medical College of Georgia, Augusta, Georgia 3 University of California San Diego Medical Center, San Diego, California 4 Georgetown University School of Medicine, Washington, District of Columbia 5 George Washington University Washington, District of Columbia 6 McMaster University, Ontario, Canada 7 University of Arizona, Tucson, Arizona 8 Children’s Mercy Hospitals and Clinics, Kansas City, Missouri 9 Pennsylvania State University, University Park, Pennsylvania 10 University of Virginia School of Medicine, Charlottesville, Virginia 11 U.S. Centers for Disease Control and Prevention, Atlanta, Georgia 12 Cerus Corporation/P-val Gal Consulting, Davis, California 13 University of Turku/Turku University, Turku, Finland 14 RTI International, Research Triangle Park, North Carolina Soy infant formula contains soy protein isolates and is fed to infants as a supplement to or replacement for human milk or cow milk. Soy protein isolates contains estrogenic isoflavones (phytoestrogens) that occur naturally in some legumes, especially soybeans. Phytoestrogens are nonsteroidal, estrogenic compounds. In plants, nearly all phytoestrogens are bound to sugar molecules and these phytoestrogen–sugar complexes are not generally considered hormonally active. Phytoestrogens are found in many food products in addition to soy infant formula, especially soy-based foods such as tofu, soy milk, and in some over-the-counter dietary supplements. Soy infant formula was selected for National Toxicology Program (NTP) evaluation because of (1) the availability of large number of developmental toxicity studies in laboratory animals exposed to the isoflavones found in soy infant formula (namely, genistein) or other soy products, as well as few studies on human infants fed soy infant formula, (2) the availability of information on exposures in infants fed soy infant formula, and (3) public concern for effects on infant or child development. On October 2, 2008 (73 FR 57360), the NTP Center for the Evaluation of Risks to Human Reproduction (CERHR) announced its intention to conduct an updated review of soy infant formula to complete a previous evaluation that was initiated in 2005. Both the current and previous evaluations relied on expert panels to assist the NTP in developing its conclusions on the potential developmental effects associated with the use of soy infant formula, presented in the NTP Brief on Soy Infant Formula. The initial expert panel met on March 15 to 17, 2006, to reach conclusions on the potential developmental and reproductive toxicities of soy infant formula and its predominant isoflavone constituent genistein. The expert panel reports were released for public comment on May 5, 2006 (71 FR 28368). On November 8, 2006 (71 FR 65537), CERHR staff released draft NTP Briefs on Genistein and Soy Formula that provided the NTP’s interpretation of the potential for genistein and soy infant formula to cause adverse reproductive and/or developmental effects in exposed humans. However, CERHR did not complete these evaluations, finalize the briefs, or issue NTP Monographs on these substances based on this initial evaluation. Between 2006 and 2009, a substantial number of new publications related to human exposure or reproductive and/or developmental toxicity were published for these substances. Thus, CERHR determined that updated evaluations of genistein and soy infant formula were needed. However, the current evaluation focuses only on soy infant formula and the potential developmental toxicity of its major isoflavone components, e.g. genistein, daidzein (and estrogenic metabolite, equol), and glycitein. This updated evaluation does not include an assessment on the potential reproductive toxicity of genistein following exposures during adulthood as was carried out in the 2006 evaluation. CERHR narrowed the scope of the evaluation because the assessment of reproductive effects of genistein following exposure to adults was not considered relevant to the consideration of soy infant formula use in infants during Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/bdrb.20314 *Correspondence to: Gail McCarver, Medical College of Wisconsin, Milwaukee, Wisconsin. E-mail: [email protected] Received 4 March 2011; Accepted 17 May 2011 Birth Defects Research (Part B) 92:421–468 (2011) & 2011 Wiley Periodicals, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Original Article
NTP-CERHR Expert Panel Report on theDevelopmental Toxicity of Soy Infant Formula
Gail McCarver,1� Jatinder Bhatia,2 Christina Chambers,3 Robert Clarke,4 Ruth Etzel,5 Warren Foster,6
Patricia Hoyer,7 J. Steven Leeder,8 Jeffrey M. Peters,9 Emilie Rissman,10 Michael Rybak,11 ClaireSherman,13 Jorma Toppari,13 and Katie Turner14
1Medical College of Wisconsin, Milwaukee, Wisconsin2Medical College of Georgia, Augusta, Georgia
3University of California San Diego Medical Center, San Diego, California4Georgetown University School of Medicine, Washington, District of Columbia
5George Washington University Washington, District of Columbia6McMaster University, Ontario, Canada7University of Arizona, Tucson, Arizona
8Children’s Mercy Hospitals and Clinics, Kansas City, Missouri9Pennsylvania State University, University Park, Pennsylvania
10University of Virginia School of Medicine, Charlottesville, Virginia11U.S. Centers for Disease Control and Prevention, Atlanta, Georgia
12Cerus Corporation/P-val Gal Consulting, Davis, California13University of Turku/Turku University, Turku, Finland
14RTI International, Research Triangle Park, North Carolina
Soy infant formula contains soy protein isolates and is fed to infants as a supplement to or replacement for human milkor cow milk. Soy protein isolates contains estrogenic isoflavones (phytoestrogens) that occur naturally in some legumes,especially soybeans. Phytoestrogens are nonsteroidal, estrogenic compounds. In plants, nearly all phytoestrogens arebound to sugar molecules and these phytoestrogen–sugar complexes are not generally considered hormonally active.Phytoestrogens are found in many food products in addition to soy infant formula, especially soy-based foods such astofu, soy milk, and in some over-the-counter dietary supplements. Soy infant formula was selected for NationalToxicology Program (NTP) evaluation because of (1) the availability of large number of developmental toxicity studies inlaboratory animals exposed to the isoflavones found in soy infant formula (namely, genistein) or other soy products, aswell as few studies on human infants fed soy infant formula, (2) the availability of information on exposures in infantsfed soy infant formula, and (3) public concern for effects on infant or child development. On October 2, 2008 (73 FR57360), the NTP Center for the Evaluation of Risks to Human Reproduction (CERHR) announced its intention to conductan updated review of soy infant formula to complete a previous evaluation that was initiated in 2005. Both the currentand previous evaluations relied on expert panels to assist the NTP in developing its conclusions on the potentialdevelopmental effects associated with the use of soy infant formula, presented in the NTP Brief on Soy Infant Formula.The initial expert panel met on March 15 to 17, 2006, to reach conclusions on the potential developmental andreproductive toxicities of soy infant formula and its predominant isoflavone constituent genistein. The expert panelreports were released for public comment on May 5, 2006 (71 FR 28368). On November 8, 2006 (71 FR 65537), CERHRstaff released draft NTP Briefs on Genistein and Soy Formula that provided the NTP’s interpretation of the potential forgenistein and soy infant formula to cause adverse reproductive and/or developmental effects in exposed humans.However, CERHR did not complete these evaluations, finalize the briefs, or issue NTP Monographs on these substancesbased on this initial evaluation. Between 2006 and 2009, a substantial number of new publications related to humanexposure or reproductive and/or developmental toxicity were published for these substances. Thus, CERHR determinedthat updated evaluations of genistein and soy infant formula were needed. However, the current evaluation focuses onlyon soy infant formula and the potential developmental toxicity of its major isoflavone components, e.g. genistein,daidzein (and estrogenic metabolite, equol), and glycitein. This updated evaluation does not include an assessment onthe potential reproductive toxicity of genistein following exposures during adulthood as was carried out in the 2006evaluation. CERHR narrowed the scope of the evaluation because the assessment of reproductive effects of genisteinfollowing exposure to adults was not considered relevant to the consideration of soy infant formula use in infants during
Published online in Wiley Online Library (wileyonlinelibrary.com)DOI: 10.1002/bdrb.20314
*Correspondence to: Gail McCarver, Medical College of Wisconsin,Milwaukee, Wisconsin.E-mail: [email protected] 4 March 2011; Accepted 17 May 2011
Birth Defects Research (Part B) 92:421–468 (2011)& 2011 Wiley Periodicals, Inc.

the 2006 evaluation. To obtain updated information about soy infant formula for the CERHR evaluation, the PubMed(Medline) database was searched from February 2006 to August 2009 with genistein/genistin, daidzein/daidzin,glycitein/glycitin, equol, soy, and other relevant keywords. References were also identified from the bibliographies ofpublished literature. The updated expert panel report represents the efforts of a 14-member panel of government andnongovernment scientists, and was prepared with assistance from NTP staff. The finalized report, released on January15, 2010 (75 FR 2545), reflects consideration of public comments received on a draft report that was released on October19, 2009, for public comment and discussions that occurred at a public meeting of the expert panel held December 16 to18, 2009 (74 FR 53509). The finalized report presents conclusions on (1) the strength of scientific evidence that soy infantformula or its isoflavone constituents are developmental toxicants based on data from in vitro, animal, or human studies;(2) the extent of exposures in infants fed soy infant formula; (3) the assessment of the scientific evidence that adversedevelopmental health effects may be associated with such exposures; and (4) knowledge gaps that will help establishresearch and testing priorities to reduce uncertainties and increase confidence in future evaluations. The Expert Panelexpressed minimal concern for adverse developmental effects in infants fed soy infant formula. This level of concernrepresents a ‘‘2’’ on the five-level scale of concern used by the NTP that ranges from negligible concern (‘‘1’’) to seriousconcern (‘‘5’’). The Expert Panel Report on Soy Infant Formula was considered extensively by NTP staff in preparing the2010 NTP Brief on Soy Infant Formula, which represents the NTP’s opinion on the potential for exposure to soy infantformula to cause adverse developmental effects in humans. The NTP concurred with the expert panel that there isminimal concern for adverse effects on development in infants who consume soy infant formula. This conclusion wasbased on information about soy infant formula provided in the expert panel report, public comments received during thecourse of the expert panel evaluation, additional scientific information made available since the expert panel meeting,and peer reviewer critiques of the draft NTP Brief by the NTP Board of Scientific Counselors (BSC) on May 10, 2010(Meeting materials are available at http://ntp.niehs.nih.gov/go/9741.). The BSC voted in favor of the minimal concernconclusion with 7 yes votes, 3 no votes, and 0 abstentions. One member thought that the conclusion should be negligibleconcern and two members thought that the level of concern should be higher than minimal concern. The NTP’s responseto the May 10, 2010 review (‘‘peer-review report’’) is available on the NTP website at http://ntp.niehs.nih.gov/go/9741.The monograph includes the NTP Brief on Soy Infant Formula as well as the entire final Expert Panel Report on SoyInfant Formula. Public comments received as part of the NTP’s evaluation of soy infant formula and other backgroundmaterials are available at http://cerhr.niehs.nih.gov/evals/index.html. Reports can be obtained from the web site(http://cerhr.niehs.nih.gov/) or from: Kristina A. Thayer, PhD, NIEHS/NTP K2-04, PO Box 12233, Research TrianglePark, NC 27709. E-mail: [email protected]. Birth Defects Res (Part B) 92:421–468, 2011. r 2011 Wiley Periodicals, Inc.
Key words: endocrine disrupters; nutrition; safety assessment
SUMMARY OF CHEMISTRY, USE,AND HUMAN EXPOSURE
In children and adults, exposures to isoflavones occurthrough consumption of soy foods such as tofu, soy milk,soy flour, textured soy protein, tempeh, and miso (FDA,2000b). Infants can be exposed by consuming soy-basedinfant formula, the breast milk of mothers who consumesoy products, or by the use of soy in weaning or‘‘transition’’ foods. Soy oils or soy sauces contain little-to-no genistein (Setchell, 1998; ILSI, 1999). Soy protein canbe used in baked goods, breakfast cereals, pasta,beverages, toppings, meat, poultry, fish products, anddairy-type products including imitation milk and cheese(Soyfoods Associations of North America, 2009; UnitedSoybean Board, 2009). Soybean derivatives are present in60% of processed foods available from U.K. super-markets (UK-Committee-on-Toxicity, 2003). The percent-age of processed foods containing soybeans in the UnitedStates is not known. Exposure to genistein can also occurthrough soy supplements marketed for the beneficialeffects on health, such as improved cardiovascular healthand treatment of menopausal symptoms (Drugstore.com,2009).
On the basis of the sales of soy products, it appearsthat exposure to soy isoflavones in the United States isincreasing and will continue to increase. The SoyfoodsAssociation of America reported that soy food sales haveincreased from $300 million to over $4 billion between
1992 and 2008, attributing this increase to new soy foodcategories being introduced, soy foods being reposi-tioned in the market place, and new customers selectingsoy for health and philosophical reasons (SoyfoodsAssociations of North America, 2009).
The primary isoflavones detected in soy products,including soybeans and soy formula, are derived fromgenistein, daidzein, and to a smaller extent, glycitein.These isoflavones are often referred to as phytoestrogensbecause of their ability to bind to estrogen receptors(ERs) and display weak estrogenic activity comparedwith estradiol (at least based on in vitro model systems),with a relative estrogenic potency of genistein4daid-zein4glycitein (MAFF, 1998; UK-Committee-on-Toxicity,2003; Choi et al., 2008).
The majority of isoflavones in soy formula andunfermented soy products are conjugated to sugarmolecules to form the glycosides genistin, daidzin, andglycitin (Cassidy, 2006; Nielsen and Williamson, 2007).Glucose groups in glycoside compounds can also beesterified with acetyl or malonyl groups to form acetyl-or malonyl glycosides (UK-Committee-on-Toxicity, 2003)(Fig. 1). The terms genistein, daidzein, and glycitein referspecifically to the unconjugated (aglycone) forms of theisoflavones. Small amounts of genistein and daidzein(3.2–5.8%) are present in soy products in their un-conjugated (aglycone) forms (Chen and Rogan, 2004).As a result of bacterial hydrolysis during fermentation,aglycones represent a larger proportion of the isoflavones
422 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

in fermented soy products such as miso, tempeh, andsoybean paste (ILSI, 1999; UK-Committee-on-Toxicity,2003). Because glycosidic compounds are rapidly decon-jugated in the gut to form the biologically active aglyconecompound, exposure to a particular isoflavone istheoretically the sum of the aglycone and respective
glycoside compound concentrations converted on thebasis of molecular weight (MAFF, 1998; UK-Committee-on-Toxicity, 2003). Unfortunately, there is an inconsistency inthe literature where many studies do not clarify whetherthe presented isoflavone levels were normalized on anaglycone basis. Failure to convert the major glycosides,
Fig. 1. Chemical structures of isoflavones found in soy formula.
423NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

i.e. genistin, daidzin, glycitin, to the appropriate aglyconeequivalents can overestimate isoflavone levels or intake byB1.6-fold. Information in the Expert Panel report isnormalized to aglycone equivalents where feasible.
Isoflavone levels in soybeans can vary as a result ofcrop strain, geographic location, climate, and growingconditions (Setchell et al., 1998; UK-Committee-on-Toxicity, 2003). Heating of soy products can causedecarboxylation, deacetylation, or deglycosylation ofglycosides with decomposition of malonyl compoundsto their respective acetylglycosides (Setchell et al., 1998;UK-Committee-on-Toxicity, 2003). Except for alcoholextraction, processing soybeans does not usually reduceisoflavone content (ILSI, 1999).
Soy formula refers to infant food made using soyprotein isolate and other components such as corn syrup,vegetable oils, and sugar (Drugstore.com, 2009). Decadesago soy formula included soy flour. However, in the1950s and 1960s, cases of altered thyroid function, mostlygoiter, were reported in infants fed soy formula. Thisproblem was eliminated by adding more iodine to theformulas and replacing soy flour with soy protein isolate.Although the early reports of goiter in infants fed soyformula have mostly ceased since manufacturers begansupplementing soy formula with iodine in 1959, there isstill concern that the use of soy formula in infants withcongenital hypothyroidism (CH) may decrease theeffectiveness of thyroid hormone replacement therapy,i.e. L-thyroxin. The soy protein isolate is fortified withL-methionine, L-carnitine, and taurine and other nutrients(FDA, 2000a; Bhatia and Greer, 2008). Soy protein isolateincludes phytates (1.5%), which bind minerals andprotease inhibitors, which have antitrypsin, antichymo-trypsin, and antielastin properties (Bhatia and Greer,2008). Phosphorus, calcium, iron, and zinc are added tosoy formula to compensate for phytate binding ofminerals. Heat applied during the processing of soyprotein removes 80 to 90% of protease inhibitor activity.Aluminum is present in soy formulas because of theaddition of mineral salts.
Many aspects of infant formula use are unknown,including what percent of infants are exclusively fedformula compared with what percent are fed a mixtureof infant formula and breast milk. It is also unknownwhat proportion of formula-fed infants are exclusivelyfed soy formula. A 1998 infant-feeding survey conductedby Ross Products Division indicated that 18% of infantsare fed soy formula during the first year of life (Stromet al., 2001). According to market data and hospitaldischarge records, another estimate was that 25% ofnewborns in the United States are fed soy formula(Badger et al., 2002). A study conducted at YaleUniversity assessed formula changes in 189 breast-fedinfants and 184 formula-fed infants and reported that23% of infants in the study received soy formulasometime during the first 4 months of life (Forsythet al., 1985). Based on recent market data, sales of soyformula in the United States represent B12% of the U.S.infant formula dollar sales (personal communicationwith Robert Rankin, Manager of Regulatory and Techni-cal Affairs at the IFC, October 13, 2009). In the UnitedStates between 1999 and 2009, estimates of total soyinfant formula fed decreased from 22.5 to 12.7%calculated based on total formula sold corrected fordifferences in formula cost, i.e. expressed in equivalent
feeding units (public comment from the InternationalFormula Council (IFC), received December 3, 2009 andpersonal communication with Dr. Haley Curtis Stevens,IFC). When sales are considered as a surrogate measureof actual reported usage, these data provide a lowerindication of usage compared with other frequently citedestimates. The usage and sales of soy formula varygeographically ranging from 2 to 7% of infant formulasales in the United Kingdom, Italy, and France, to 13% inNew Zealand (Agostoni et al., 2006; Turck, 2007), and to31.5% in Israel (Berger-Achituv et al., 2005).
A commonly cited reason for use of soy formula is tofeed infants who are allergic to dairy products or areintolerant of lactose, galactose, or cow-milk protein(Essex, 1996; Tuohy, 2003). In May 2008, the AmericanAcademy of Pediatrics (AAP) released an updated policystatement on the use of soy protein-based formulas(Bhatia and Greer, 2008). The overall conclusion of theAAP was that, although isolated soy protein-basedformulas may be used to provide nutrition for normalgrowth and development in term infants, there are fewindications for their use in place of cow milk-basedformula. The only real indications for use are instanceswhere the family prefers a vegetarian diet or for themanagement of infants with galactosemia or primarylactase deficiency (rare). Soy formula is not currentlyrecommended for preterm infants. Similar conclusionswere reached in 2006 by the European Society forPaediatric Gastroenterology Hepatology and Nutrition(ESPGHAN) Committee on Nutrition (Agostoni et al.,2006).
A number of studies in the United States and abroadhave measured total isoflavone levels in infant formulas(see Table 9 of complete final expert panel reportavailable at http://cerhr.niehs.nih.gov/evals/genistein-soy/soyformula/soyformula.html). For formulas fromthe United States, the range of total isoflavone levelsreported in reconstituted or ‘‘ready-to-feed’’ formulaswas 20.9 to 47 mg/liter formula (Franke et al., 1998;Setchell et al., 1998). When normalized to aglyconeequivalents, genistein is the predominant isoflavonefound in soy formula (B58–67%), followed by daidzein(B29–34%) and glycitein (B5–8%). In contrast to theisoflavone content of soybeans and other soy productssuch as soy supplements or soy protein isolates, theisoflavone content in soy formula is less variable. Therange of total isoflavones content in soy formula samplescollected in the United States and other countries is 10 to47 mg/liter (Table 1) (Setchell et al., 1998; Genovese andLajolo, 2002).
Isoflavone exposure through soy formula intake hasbeen estimated in the United States and other countriesbased on total isoflavone levels measured in soyformulas and assumptions of formula intakes and infantbody weights. In the United States, total isoflavone intakeby infants was estimated at 2.3 to 9.3 mg/kg body weight(bw)/day, depending on age of the infant, the estimatedintake for genistein, expressed in aglycone equivalents,ranges from 1.3 to 6.2 mg/kg bw/day. These intakes areseveral orders of magnitude greater than infants whoconsume breast milk or a cows milk-based formula. Soyformula-fed infants have higher daily intakes of genisteinand other isoflavones compared with other populations(excluding regular consumers of soy supplements)(Table 2). However, differences in strategies used to
424 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

develop the intake estimates and sample representative-ness limit the ability to compare across studies, especiallyfor those based on dietary surveys. In addition, iso-flavone intake seems to be inherently highly variable inadult populations and there is support for the notion thatthis variability cannot solely be attributed to differencesin study methods. Recognizing these caveats, the relativeranking of total isoflavone intake appears to be infantsexclusively fed soy formula4vegans4Japanese consum-ing a traditional diet4vegetarians4soy consumers.
Mean blood-based levels of isoflavones in infants fedsoy formulas are considerably higher than other popula-tions, including vegans and Japanese adults (Table 3).For example, concentrations of total genistein in wholeblood samples from U.S. infants fed soy formula are
1455 ng/ml at the 75th percentile (Cao et al., 2009). Thisvalue is almost five times higher than the maximumgenistein concentration detected in plasma in a smallstudy of Japanese men, n 5 6 (Adlercreutz et al., 1994).The mean level of genistein (757 ng/ml) in the blood ofthe soy formula-fed infants reported by Cao et al. (2009)is almost 20 times higher than the mean level of genisteindetected in a sample of vegetarians and vegans inOxford, England (Peeters et al., 2007). Average bloodlevels of total genistein in the soy formula-fed infants areB160 times higher than the mean levels of total genisteinin omnivorous adults in the United States reported byValentin-Blasini (2003) (757 vs. 4.7 ng/ml). A similarpattern is observed based on urinary concentrations ofisoflavones (Table 4).
Table 2Comparison of Estimated U.S. Intake of Genistein and Total Isoflavones in Infants Fed Soy Formula to Other Populations
Daily intake (mg/kg bw/day)
Population Genistein Total isoflavone Reference
U.S. infants (soy formula) 1.3–6.2 2.3–9.3 Table 9 from unabbreviated final expert panel report(http://cerhr.niehs.nih.gov/evals/genistein-soy/SoyFormulaUpdt/FinalEPReport_508.pdf)
U.S. adults (general) 0.005a–0.056b 0.0097a–0.096b Haytowitz et al. (2009)a; Tseng et al. (2008)b; Kirk et al.(1999)
U.S. vegetarians 0.14 0.21European men Mulligan et al. (2007)
Not soy consumers 0.005 0.009Soy consumers 0.057 0.100
European women Mulligan et al. (2007)Not soy consumers 0.004 0.007Soy consumers 0.062 0.112
Vegans (UK) 1.07 Friar and Walker (1998) as cited in Mortensen et al.(2009)
Japanese 0.077a–0.43b 0.67b Fukutake et al. (1996)a as cited in Fitzpatrick (1998);Arai et al. (2000)b
Table 1Comparison of Isoflavone Content in Infant Soy Formulas from Different Countries
Individual isoflavones, mg/liter formula as fed1
(percent of total)
CountryTotal isoflavones, mg/liter
formula as feda Genistein Daidzein Glycitein Reference
US (n 5 20) 20.9a–47b 12.1a–31.2b 7.1a–13.5b 1.7a–2.4b Franke et al. (1998)a; Setchellet al. (1998)b(57.7–66.3%) (B28.8–34%) (B5–8.3%)
UK (n 5 13) 18a–46.7b 10.4a–31.3b 6.7a–12.5b 0.9a–2.8b UK Ministry of Agriculture,Fisheries, and Food (1998)a;Hoey et al. (2004)b
(58.0–67.1%) (26.8–37%) (5–6.1%)
Australia (n 5 4) 17.2–21.9 – – – Knight et al. (1998)New Zealand 17.1–33 11.2–18 5.9–15 – Irvine et al. (1998)(n 5 5) (55–65.4%) (34.6–45%)Brazil 10–27.4 5.9–16.2 2.4–8.6 1.6–2.4 Genovese and Lajolo (2002)(n 5 7) (59.3–59.8%) (23.9–31.7%) (9–16.3%)
1CERHR converted between mg/kg (or mg/g) and mg/liter because the density of prepared formula is similar to water (specific gravityis 1.03) (personal communication from Mead Johnson Medical Nutrition Affairs). – 5 not specified.
425NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

SUMMARY OF PHARMACOKINETICSAND GENERAL TOXICOLOGY
Pharmacokinetics
Genistein, daidzein, and glycitein exist mainly in theirglycosidic forms in unfermented soy foods. Beforeisoflavone glycosides can be absorbed into the systemiccirculation, they must first be hydrolyzed to theiraglycones, which have greater hydrophobicity and lowermolecular weight. Before systemic availability, mostgenistein and daidzein are conjugated with glucuronicacid by uridine diphosphate (UDP)-glucuronosyltrans-ferases; a smaller amount is conjugated to sulfate bysulfotransferases. Conjugation of isoflavones can alsooccur in liver. The glucuronide and sulfate conjugatesenter the systemic circulation, and the majority ofisoflavone compounds in the circulation are present inconjugated form.
Humans. In humans, a considerable amount ofpharmacokinetic information is available for genisteinand daidzein in adults. Less information is available forequol and very little data have been published onglycitein. Human developmental pharmacokinetic dataare lacking; the available ‘‘pharmacokinetic’’ data inhuman infants are essentially equivalent to biomonitor-ing data. The detection of genistein, daidzein, and equolin serum, urine, amniotic fluid, cord blood, and breastmilk in humans demonstrate fetal exposure and absorp-tion into the systemic circulation of infants. Fullpharmacokinetic data on genistein, daidzein, and/orglycitein among human infants following soy formulafeeding have not been published. Rather, data are limitedto single plasma concentrations, generally after chronicsoy formula feeding, which likely represent steady state(although this is uncertain) and spot urinary concentra-tions (Cruz et al., 1994; Setchell et al., 1997; Irvine et al.,
Table 4Comparison of Urinary levels of Genistein, Daidzein, and Equol in Infants Fed Soy Formula to the General
U.S. Population
Urinary concentration (geometric mean), mg/liter or ng/ml
Population Genistein Daidzein Equol Reference
U.S. infants fed soy formula (n 5 27) 5891 5097 2.3 Cao et al. (2009)U.S. children ages 6–11 years (n 5 721–726) 33.8 88.1 11.5 U.S. CDC (2008)U.S. adults ages 40–59 years (n 5 951) 23.7 52.3 8.39 U.S. CDC (2008)
Table 3Comparison of Blood-Based Levels of Genistein and Daidzein in Infants Fed Soy Formula to Other Populations
Average total isoflavone concentration, nM(ng/ml)
Population Sample Genistein Daidzein Reference
U.S. infants (soy formula, n 5 27) Whole blood(spots)
2801 (757), geometricmean
1007 (256), geometricmean)
Cao et al. (2009)
3296 (891), median 1081 (275), median)5384.5 (1455), 75th
percentile2040 (519), 75th
percentileU.S. infants (soy formula, n 5 7) Plasma 2531 (684) 1160 (295) Setchell et al. (1997)U.S. infants (cow milk formula, n 5 30) Whole blood
(spots)52.5 (14.2), geometric
mean(5.5), geometric mean Cao et al. (2009)
U.S. infants (cow milk formula, n 5 7) Plasma 11.7 (3.16), mean 8.1 (2.06), mean Setchell et al. (1997)U.S. infants (breastfed, n 5 20) Whole blood
(spots)40 (10.8), geometric
mean20.8 (5.3), geometric
meanCao et al. (2009)
U.S. infants (breastfed, n 5 7) Plasma 10.3 (2.77) 5.9 (1.49) Setchell et al. (1997)Omnivorous adults in the United States
(n 5 209)Serum 17.4 (4.7) 15.3 (3.9) Valentin-Blasini et al.
(2003)oLOD–(203), range oLOD–(162), range
Vegetarians, Finland (n 5 14) Plasma 17.1 (4.6) 18.5 (4.7) Adlercreutz et al. (1994)Vegans and vegetarians (Oxford, UK,
n 5 70)Plasma 148 (40) 78.7 (20) Peeters et al. (2007)
Japanese women at delivery (n 5 51a or194b)
Serum 26.6 (7.2)a–116.5 (31.5)b 7.1 (1.8)a–50.2 (12.8)b Todaka et al. (2005)a;Nagata et al. (2006)b
Japanese cord serum at delivery (n 5 51a
or 194b)Serum 71.8 (19.4)a–126.9 (34.3)b 16.9 (4.3)a–38.6 (9.81)b Todaka et al. (2005)a;
Nagata et al. (2006)b
Japanese men (traditional diet, n 5 6) Plasma 90.4 (24)–1204 (325),range
58.3 (15)–924 (235),range
Adlercreutz et al. (1994)
426 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

1998; Hoey et al., 2004; Franke et al., 2006; Halm et al.,2007; Franke et al., 2008a; Halm et al., 2008; Cao et al.,2009). No studies of human infants fed genistein,daidzein, and/or glycitein have been reported. Thus,the human pharmacokinetic data are solely based onstudies conducted in adults.
Relatively, few studies include measurement of glyci-tein and it has not been measured in biomonitoringstudies of the general population conducted by the CDCas part of NHANES (U.S. Centers for Disease Controland Prevention, 2008) or measured in the plasma or urineof soy formula-fed infants (Setchell et al., 1997; Irvineet al., 1998; Franke et al., 2006; Cao et al., 2009). Ifglycitein is measured in humans, it is generally followingintentional dosing of subjects with a soy protein orisoflavone supplement.
Before reaching the systemic circulation, most genis-tein and daidzein is conjugated to glucuronic acid byUDP-glucuronosyltransferases; a much smaller amountis conjugated to sulfate by sulfotransferase enzymes(Joannou et al., 1995; Kurzer and Xu, 1997; UK-Committee-on-Toxicity, 2003). Conjugation of genisteinoccurs in the intestine but also has been reported to occurin liver. Based on in vitro data, the enzymes mostinvolved in glucuronidation are: UGT1A1, 1A8, 1A9,1A10 (Tang et al., 2009), whereas the SULT enzymeslikely responsible for sulfation include SULT1A1 andSULT2A1 (Doerge et al., 2000; Chen et al., 2008).However, interpretation of the in vitro data requiresinformation on tissue-specific expression, particularly forhuman intestine and liver. Moreover, to inform thequestion of disposition of soy formula isoflavones, theontogeny of such tissue-specific expression is needed.Only a limited amount of such data are available (Hines,2008) precluding estimates of developmental differencesamong humans, as well as interspecies differencesamong the young.
The glucuronide and sulfate conjugates can enter thesystemic circulation, and it has been reported that themajority of isoflavone compounds in the circulationare present in conjugated form, thus limiting the bio-availability of the aglycone forms of isoflavones. Instudies in which humans were exposed to genistein orisoflavone aglycones at genistein doses of 1 to 16 mg/kgbw, most of the genistein was present in plasma inconjugated form, while free genistein represented 1 to 3%of total plasma genistein levels in most cases (Setchellet al., 2001; Busby et al., 2002; Bloedon et al., 2002).Conjugated isoflavones undergo enterohepatic circula-tion, and on return to the intestine, they are deconjugatedby bacteria with b-glucuronidase or arylsulfatase activity.The metabolites may be reabsorbed or further metabolizedby gut microflora. Isoflavones can undergo furtherbiotransformation that ultimately leads to the formationof 60-hydroxy-O-desmethylangolensin from genistein andO-desmethylangolensin from daidzein (Joannou et al.,1995; Kurzer and Xu, 1997; UK-Committee-on-Toxicity,2003). The metabolic profile varies among individuals,with some individuals producing little or no O-desmethyl-angolensin or equol, an intermediate metabolite ofdaidzein that is biologically active.
Based on the summary of the studies presentedin Nielsen and Williamson (2007) and Cassidy (2006),pharmacokinetic parameters (cmax, tmax, t1/2, area underthe plasma concentration-time curve [AUC]) are similar
for genistein and daidzein. For genistein, the average tmax
was 5.7 hr (range of 3.5–9.3 hr) and for daidzein, it was6.2 hr (range of 4.0–8.3 hr). These estimates are consistentwith the range of peak values of B6 to 8 hr reported byothers for genistein and daidzein following ingestion ofsoy or isoflavones (Pumford et al. (2002); reviewed inWhitten and Patisaul (2001); UK-Committee-on-Toxicity(2003); Larkin et al. (2008). The maximum concentrations(Cmax) of genistein and daidzein, when normalized toingestion of 1mmol/kg bw, were also similar at 0.49 or0.64 mM or 0.50 or 0.54 mM, respectively. The average half-life of elimination (t1/2) estimates from reviews byNielsen and Williamson (2007) and Cassidy (2006) didnot differ between genistein, 9.5 hr (range 6.1–17 hr), anddaidzein, 7.7 hr (range 4.2–16 hr). These estimates aregenerally similar to those presented in a recent review byLarkin (2008); 3 to 9 hr for daidzein and 8 to 11 hr forgenistein after intake of soy foods or pure isoflavoneglycosides. The AUC values, when normalized toingestion of 1mmol/kg bw, were 11 mmol�hr/liter forgenistein and 18mmol�hr/liter for daidzein. Absorptionhalf-lives for both daidzin and genistin, presumably asglucuronides and sulfates of daidzein and genistein,were reported at B1 to 3 hr following intake of foodscontaining r210 mg of each isoflavone or providingdoses of r2 mg/kg bw/day of each isoflavone (Lu et al.,1995; Lu and Anderson, 1998); reviewed in (Whitten andPatisaul, 2001).
Studies in humans that report the bioavailability andother pharmacokinetic parameters of isoflavones ingestedas glycosides versus aglycones have reported conflictingfindings (reviewed in UK-Committee-on-Toxicity, 2003;Cassidy et al., 2006; Nielsen and Williamson, 2007).Although there is no clear explanation for the conflictingfindings on bioavailability of the aglycone and glucoside,one factor may be the differences in the type of isoflavonepreparations administered to subjects, e.g. soybean extractscontaining mixtures of isoflavones, purified single iso-flavone, ingestion in tablet or liquid form (Cassidy, 2006).In addition, comparing the conclusions on bioavailabilityacross studies is difficult because of the variety of indicesused to assess bioavailability, e.g. relative AUC, cmax, tmax,urinary excretion, and recovery, etc. Prasain and Barnes(2007) discussed the various approaches used to assess thebioavailability of isoflavones. In pharmacology (andtoxicology), the term absolute bioavailability refers to theratio of AUC after oral ingestion to the AUC after systemicadministration, i.e. intravenous injection. This is thefraction of the compound absorbed through nonintrave-nous administration compared with the correspondingintravenous administration of the same drug. Thisapproach may underestimate bioavailability at the tissuelevel if circulating isoflavones in the conjugated form areconverted to aglycones by b-glucuronidases and sulfatasessecreted by cells within target tissues. In the nutritionliterature, bioavailability is often assessed by the percent ofadministered isoflavone that is recovered in the urine.Higher degrees of recovery in urine are interpretedas more bioavailability based on the assumption thatthe isoflavone must have been present in bloodbefore reaching the kidney and ultimately being excretedin urine.
In their review, Nielsen and Williamson (2007) con-cluded that despite the apparently contradictory findings,the data are consistent enough to conclude that (1) at
427NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

equivalent doses, the Cmax is higher for genistein anddaidzein following administration as glucosides comparedwith aglycones and (2) the half-life is not significantlydifferent for aglycone and glucoside. In addition, theyconcluded that deglycosylation is required for absorptionbut does not appear to be a rate-limiting step. As notedabove, studies by Setchell et al. (2001) and Rufer et al.(2008) suggest the percent of genistein and daidzeincirculating in the unconjugated forms do not differ basedon administered form. These conclusions support theassumption used by others that because glycosides arequickly deconjugated in the gut to form the activeaglycones, exposure to a particular isoflavone (e.g.genistein) is theoretically the sum of the aglycone andrespective glycoside compound concentrations convertedon the basis of molecular weight (MAFF; Setchell et al.,1998; UK-Committee-on-Toxicity, 2003; Rozman et al.,2006).
No pharmacokinetic studies, defined as measurementof isoflavone concentrations in serial blood or plasmasamples, have been conducted in infants or children.The only data that are interpretable in this context are thedata presented in Setchell et al. (1997) and Cao et al.(2009). These studies reported genistein and daidzeinconcentrations in plasma or blood samples obtained atunspecified times relative to feeding in infants chroni-cally fed soy formula. Usual feeding behavior wasreported to result in mean genistein concentrations of684 and 757 ng/ml, and mean daidzein concentrations of295 and 256 ng/ml by Setchell et al. (1997) and Cao et al.(2009), respectively. Data obtained by Cao et al. (2009)indicated that total genistein concentrations in wholeblood ranged from approximately 13.5 to 3562.9 ng/ml(range data obtained from personal communication withDr. Cao, December 2009). Assuming that the infants wereconsuming soy formula for more than 2 to 3 days, thesevalues approximate a steady-state condition, and thereported concentrations provide a context in whichanimal data can be prioritized and interpreted.
Limited data suggest low levels of equol are detectedin both whole blood and urine of infants, regardless oftype of feeding (Setchell et al., 1997, 1998; Hoey et al.,2004; Cao et al., 2009). Setchell et al. (1997, 1998) detectedequol in all cow-milk formula-fed infants, four of sevensoy formula-fed infants, and one of seven breast-fedinfants. In contrast, Cao et al. (2009) did not detectequol in the blood of any infant, but reported detectionof equol in urine among infants fed cow milk formula(22%) compared with those fed soy formula (5%) orbreast-fed infants (2%). Also, given that equol is presentin breast milk and cow milk (King et al., 1998; Setchellet al., 1998), it does not appear that soy-based formula isassociated with increased exposure to equol relative tocow milk-based formula or breast milk. Collectively,these data indicate that relatively low exposures toequol can be detected not only in infants fed soy formulabut also in infants fed cow milk-based formula andbreast milk. Thus, this level of equol exposure in humaninfants is markedly lower than that reported in animalsfed soy-based diets or daidzein. These data from humaninfants provide a context for interpreting the animaldata presented in the section ‘‘Summary of Develop-mental Toxicity of Soy Infant Formula, IndividualIsoflavones Found in Soy Infant Formula, or Other SoyTreatments.’’
Experimental animals. Genistein is absorbed inrats and mice following oral or subcutaneous (sc)exposure. According to data in Table 5, maximumgenistein levels in blood are obtained within 2 hr ofexposure. A mass-balance study of rats orally treated bygavage with 14C-genistein 4 mg/kg bw reported Vd at1.27 to 1.47 L (Coldham and Sauer, 2000). The ExpertPanel noted that the reported Vd suggests that most ofthe circulating radioactivity was not genistein but theglucuronide. Plasma protein binding ranged from B80 to90%. Radioactivity was distributed throughout the body,with levels in reproductive organs (vagina, uterus, ovary,and prostate) higher than levels in other organs (brain,fat, thymus, spleen, skeletal muscle, and bone). Somestudies demonstrated higher levels of genistein aglyconeversus conjugates within tissues compared with blood,raising the possibility of accumulation or hydrolysis ofaglycones within tissues (Fritz et al., 1998; Chang et al.,2000; Doerge et al., 2000). The Expert Panel noted thatdifferences between free genistein levels in blood andtissues are probably due to differences in how theaglycone and glucuronide compounds partition betweenfat in blood and tissues.
There are two high-quality studies that togetheraddress the impact of route of administration in youngmice (Doerge et al., 2002; Jefferson et al., 2009a,b). Basedon these reports, the dose-adjusted AUC after oralgenistin in CD-1 mice is about half that of the scgenistein. Although there is some inconsistency in theCmax data, both values are relevant to humans as they arewithin the range of blood and plasma concentrationsreported in human infants.
Studies in adult rats and mice fed soy-containing feedreported that blood total equol levels were 2 to 10 timeshigher than blood total genistein and daidzein levels(Brown and Setchell, 2001; Lund et al., 2001; Weber et al.2001). In most cases, total equol levels were lower than orwithin the same order of magnitude as genistein anddaidzein levels in fetuses or pups of rat dams fed soy-based diets (Brown and Setchell, 2001; Weber et al., 2001).The half-life for genistein and its metabolites followingingestion of soy products by rats was reported at 8.8 hr.No data were found for half-life of daidzein and itsmetabolites following ingestion through soy foods.Urinary levels of total genistein and daidzein werereported to be B25% of total equol levels in mice andB50% of total equol levels in rats.
Studies demonstrated placental transfer of genistein tothe rat fetus (Fritz et al., 1998; Doerge et al., 2001; Soucyet al., 2006) and lactational transfer to the rat pupfollowing dietary administration of genistein to the dam(Chang et al., 2000). A study examining placental transferreported higher concentrations of aglycone in fetusescompared with dams, leading the authors to concludethat placental transfer probably involves the aglycone;the finding was said to be consistent with limitedconjugation ability of the fetal rat (Doerge et al., 2001).One study reported that the percentage of free genisteinin milk from the pup stomach (78–97%) was higher thanin milk from the dams’ nipples (57%), suggesting thatgenistein conjugates may be hydrolyzed in the pupstomach (Fritz et al., 1998).
Studies in rats demonstrated the distribution of iso-flavones and metabolites to fetuses during pregnancy orpups during lactation following ingestion of soy-containing
428 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le5
Blo
od
Gen
iste
inL
evel
sin
Ro
den
tsF
edP
hy
toes
tro
gen
-Fre
eD
iets
and
Do
sed
wit
hG
enis
tein
Ser
um
gen
iste
in,
nM
(ng
/m
lo
rmg
/li
ter)
Sp
ecie
san
dst
rain
Ag
ean
dn
um
ber
of
anim
als
Ro
ute
,d
ura
tio
no
fex
po
sure
Do
seT
ota
lA
gly
con
eP
erce
nt
agly
con
eR
efer
ence
Rat
,S
pra
gu
e-D
awle
yD
ams
and
fetu
ses
on
GD
20o
r21
,n
51
dam
(11–
16fe
tuse
s)/
gro
up
Ora
l(g
avag
e),
sin
gle
trea
tmen
to
fd
amo
nG
D20
or
21,
blo
od
coll
ecte
d2
hr
afte
rd
osi
ng
20m
g/
kg
bw
Dam
s:35
40[9
56
]D
ams:
270
[73
]D
ams:
8%D
oer
ge
etal
.(2
001)
Fet
use
s:27
0[7
3]a
Fet
use
s:80
[22
]aF
etu
ses:
31%
34m
g/
kg
bw
Dam
s:54
80[1
48
0]D
ams:
290
[78
]D
ams:
5%F
etu
ses:
190
[51
]F
etu
ses:
60[1
6]
Fet
use
s:34
%75
mg
/k
gb
wD
ams:
4410
[11
91
]D
ams:
780
[211
]D
ams:
18%
Fet
use
s:22
0[5
9]
Fet
use
s:60
[16
]F
etu
ses:
27%
Rat
,S
pra
gu
e-D
awle
yP
ND
1to
2m
ales
and
fem
ales
,n
52
to3
pu
ps
fro
m2
dif
fere
nt
litt
erp
oo
ls
Ora
l(d
iet)
,p
up
sex
po
sed
ind
irec
tly
du
rin
gg
esta
tio
nan
dla
ctat
ion
500
pp
m(B
50m
g/
kg
bw
/d
ay)
tod
ams
1767
307
[48
]47
[13
]53
[27
%b
yC
ER
HR
calc
ula
tio
n]
Do
erg
eet
al.
(200
1)
Rat
,S
pra
gu
e-D
awle
yD
ams
and
PN
D7
and
21p
up
s,n
um
ber
exam
ined
no
tre
po
rted
Ora
l(d
iet)
,d
ams
exp
ose
dd
uri
ng
ges
tati
on
and
lact
atio
n
0D
ams:
6[2
]aD
ams:
6[2
]aD
ams:
100%
Fri
tzet
al.
(199
8)P
ND
7:9
[2]
PN
D7:
9[2
]P
ND
7:10
0%P
ND
21:
6[2
]aP
ND
21:
6[2
]aP
ND
21:
100%
25p
pm
(B2.
2m
g/
kg
bw
/d
ay)
Dam
s:40
[11
]D
ams:
9[2
]D
ams:
23%
PN
D7:
86[2
3]
PN
D7:
16[4
]P
ND
7:19
%P
ND
21:
54[1
5]
PN
D21
:18
[5]
PN
D21
:33
%25
0p
pm
(B22
mg
/k
gb
w/
day
)D
ams:
418
[11
3]
Dam
s:7
[2]
Dam
s:1.
7%P
ND
7:72
6[1
96
]P
ND
7:10
3[2
8]
PN
D7:
14%
PN
D21
:18
10[4
89
]P
ND
21:
120
[32
]P
ND
21:
6.6%
Rat
,S
pra
gu
e-D
awle
yP
ND
21/
PN
D14
0o
ffsp
rin
g,
mal
esan
dfe
mal
es(n
55–
6/g
rou
p)
Ora
l(d
iet)
,d
ams
exp
ose
dd
uri
ng
ges
tati
on
and
lact
atio
n,o
ffsp
rin
gre
ceiv
edd
am’s
die
tat
wea
nin
g
0o
10/o
10[o
3]
–1
to5%
all
do
seg
rou
ps,
bo
thag
esC
han
get
al.
(200
0)5
pp
m[B
0.4
–0.5
mg
/k
gb
w/d
ay]
PN
D21
:–
22[6
](m
ale)
20[5
](f
emal
e)P
ND
140:
–60
[16
](m
ale)
100
[27
](f
emal
e)10
0p
pm
[B8
–1
0m
g/
kg
bw
/day
]P
ND
21:
270
[73
](m
ale)
520
[14
0]
(fem
ale)
PN
D14
0:59
0[1
59
](m
ale)
940
[25
4]
(fem
ale)
500
pp
m[B
40
–50
mg
/k
gb
w/d
ay]
PN
D21
:–
2090
[56
4]
(mal
e)18
70[5
05
](f
emal
e)P
ND
140:
–60
00[1
62
0](m
ale)
7940
[21
44]
(fem
ale)
429NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le5
Co
nti
nu
ed
Ser
um
gen
iste
in,
nM
(ng
/m
lo
rmg
/li
ter)
Sp
ecie
san
dst
rain
Ag
ean
dn
um
ber
of
anim
als
Ro
ute
,d
ura
tio
no
fex
po
sure
Do
seT
ota
lA
gly
con
eP
erce
nt
agly
con
eR
efer
ence
Rat
,S
pra
gu
e-D
awle
yP
ND
91,
fem
ale,
n5
4O
ral
(die
t),
21d
ays
star
tin
gat
PN
D70
750
pp
m22
00[5
84
]40
0[1
08
]18
.2%
San
tell
etal
.(1
997)
Rat
,S
pra
gu
e-D
awle
yP
ND
70,
mal
e,n
58/
gro
up
Ora
l(d
iet)
,d
ams
exp
ose
dd
uri
ng
ges
tati
on
and
lact
atio
n,
and
then
dir
ectl
yu
nti
lP
ND
70
0p
pm
18[5
]a0
0%F
ritz
etal
.(2
002)
25p
pm
(B2.
2m
g/
kg
bw
/d
ay)
167
[14
]6
[2]a
3.6%
250
pp
m(B
22m
g/
kg
bw
/d
ay)
1908
[51
5]
20[5
]1.
0%
Rat
,S
pra
gu
e-D
awle
yP
ND
70,
mal
e,n
58/
gro
up
Ora
l(d
iet)
on
PN
D57
–65
and
gav
age
on
PN
D66
–70
028
[8]a
6[2
]a21
.4%
Fri
tzet
al.
(200
2)25
0p
pm
die
t;22
mg
/k
gb
w/
day
gav
ageb
1785
[48
2]
32[7
]1.
8%
1000
pp
md
iet;
88m
g/
kg
bw
/d
ayg
avag
eb96
40[2
60
2]41
[11
]0.
43%
Rat
,S
pra
gu
e-D
awle
yA
du
lt,
fem
ale
dam
s,n
54/
gro
up
Ora
l(d
iet)
on
GD
7–P
ND
2125
0p
pm
2100
[56
7]
30[8
]1.
4%H
old
eret
al.
(199
9)
PN
D63
,fe
mal
eo
ffsp
rin
g,
n5
10/
gro
up
Ora
l(d
iet)
,ex
po
sed
ind
irec
tly
du
rin
gg
esta
tio
n(f
rom
PN
D7)
thro
ug
hla
ctat
ion
(PN
D21
)an
dth
end
irec
tly
on
PN
D21
–63
250
pp
m13
10[3
54
]38
[10
]2.
9%12
50p
pm
5300
[14
31]
150
[40
]2.
8%
Rat
,S
pra
gu
e-D
awle
yA
du
lt,
mal
ean
dfe
mal
e,n
510
/g
rou
p;
n5
7–10
/g
rou
p
Ora
l(d
iet)
,[d
ura
tio
no
ftr
eatm
ent
no
tcl
earl
yre
po
rted
]
25p
pm
(2m
g/
kg
bw
/d
ay)
r25
0[r
68
]c–
–H
old
eret
al.
(199
9)
250
pp
m(2
0m
g/
kg
bw
/d
ay)
1500
[40
5]
(mal
e)–
–20
00[5
40
](f
emal
e)12
50p
pm
(100
mg
/k
gb
w/
day
)60
00[1
62
0](m
ale)
––
9000
[24
30]
(fem
ale)
Rat
,S
pra
gu
e-D
awle
y11
Wee
ks,
fem
ale,
n5
2–8
Ora
l(d
iet)
for
3w
eek
s,b
egin
nin
gat
8w
eek
so
fag
e
049
[13
]aC
otr
on
eoet
al.
(200
1)25
0p
pm
(B16
mg
/k
gb
w/
day
)11
15[3
01
]13
8[3
7]a
12
1000
pp
m20
31[5
48
]44
6[1
20
]23
Rat
,S
pra
gu
e-D
awle
y11
Wee
ks,
fem
ale,
n5
4–5
SC
for
3w
eek
s,b
egin
nin
gat
8w
eek
so
fag
e,b
loo
dco
llec
ted
16–1
8h
raf
ter
last
inje
ctio
n
04
[1]a
––
Co
tro
neo
etal
.(2
001)
5m
g/
kg
bw
/d
ay45
0[1
22
]–
–16
6m
g/
kg
bw
/d
ay13
80[3
73
]66
2[1
79]a
4850
mg
/k
gb
w/
day
5090
[13
74]
a22
43[6
06]a
44R
at,
Sp
rag
ue-
Daw
ley
PN
D21
,50
,an
d10
0,fe
mal
e,n
56–
9S
C,
sin
gle
do
seg
iven
at21
,50
,o
r10
0d
ays
of
age;
blo
od
coll
ecte
d16
–18
hr
afte
rin
ject
ion
500
mg
/k
gb
w21
day
s:55
58[1
50
1]a
21d
ays:
1956
[52
8]a
21d
ays:
[35
]C
otr
on
eoet
al.
(200
1)50
day
s:39
[11
]50
day
s:16
[4]
50d
ays:
[41
]10
0d
ays:
13[4
]10
0d
ays:
6[2
]10
0d
ays:
[46
]
Mo
use
,C
D-1
PN
D1–
5,m
ale/
fem
ale,
n5
3–8/
sex
/ti
me
per
iod
SC
,P
ND
1–5
blo
od
coll
ecte
db
etw
een
0.5
and
24h
rfo
llo
win
gin
ject
ion
50m
g/
kg
bw
/d
ay30
00[8
10
](m
ale)
B14
00[3
78
]m
ale
31D
oer
ge
etal
.(2
002)
5000
[13
50]
(fem
ale)
B23
00[6
21
]fe
mal
e
430 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

feed by the dam (Brown and Setchell, 2001; Weber et al.,2001). In one study, gestation day (GD) 20.5 fetuses andPND 3.5 pups contained about half the total genisteinconcentration and about one-tenth the total equol concen-tration measured in dams; total daidzein concentrationswere about one-third lower in GD 20.5 fetuses and aboutthe same in PND 3.5 pups compared with dams (Weberet al. 2001). A second study reported that in pups born todams fed a soy-containing diet, total genistein levelsremained steady between birth and PND 12, while totaldaidzein levels were reduced by half during the same timeperiod (Brown and Setchell, 2001). On PND 16, the timewhen pups likely began eating food, there was a 5- to10-fold drop in total genistein and daidzein concentrationscompared with earlier time periods. In contrast, total equollevels in pups decreased steadily following birth.
Genistein glucuronide is the most abundant genisteinmetabolite in rat blood (Coldham and Sauer, 2000).Genistein is conjugated with glucuronide in the intestineand liver, and a study in rats demonstrated that themajority of glucuronidation most likely occurs in theintestine (Sfakianos et al., 1997). With the exception of4-hydroxyphenyl-2-propionic acid, all other urinarygenistein metabolites identified in rats were alsoreported for humans, suggesting pathways common tothe two species. Parent compound was the predominantform of genistein in the uterus, while in prostate the mostabundant form was the metabolite 4-hydroxyphenyl-2-propionic acid. One study reported no evidence thatgenistein aglycone or conjugate levels in blood weresaturated following exposure to dietary genistein at up to1250 ppm.
The Expert Panel noted that comparisons of serumaglycone levels in adult versus fetal or neonatal rodentsof the same study can be made from the rat datapresented in Table 5. A sc dosing study conducted in ratsdemonstrated similar percentages of serum aglycone(35–46%) at PND 21, 50, or 100. One study with gavageexposure demonstrated higher aglycone percentages infetuses (27–34%) than dams (5–18%) on GD 20 or 21(Doerge et al., 2001). A dietary study in which dams werefed 25 or 250 ppm genistein did not consistentlydemonstrate higher percentages of aglycone in dams(1.7–23%) compared with pups on PND 7 (14–19%) orPND 21 (6.6–33%) (Fritz et al., 1998). In an evaluationof all the data in Table 5, it was noted that percentagesof free genistein following oral exposure of adult ratswere usually below 10% but sometimes attained levelsof B20%; percentages of aglycone following direct and/or indirect oral exposure to genistein in rat pups r21days old were reported at 1 to 33%.
In a mass-balance study of rats orally treated bygavage with 4 mg/kg bw 14C-genistein, B65% of thedose was excreted in urine and 33% in feces at 166 hrfollowing dosing (Coldham and Sauer, 2000). Approxi-mately, 90% of the dose was recovered within the first48 hr following dosing. Total clearance was 1.18 mL/minin males and 2.0 mL/min in females. Genistein elimina-tion half-lives have been reported at 2 to 9 hr in rats and 5to 8 hr in mice (Coldham and Sauer, 2000). The ExpertPanel noted an apparent contradiction between the half-lives reported by Chang (2000) (B3–4 hr) and Coldhamand Sauer (2000) (B9–12 hr). The differences in half-livesmay have resulted from dosing regimen. Coldham andSauer used a single low dose of 4 mg/kg bw and ChangM
ou
se,
CD
-1P
ND
1–5,
fem
ale,
n5
4–6/
tim
ep
oin
tO
ral,
blo
od
coll
ecte
d0.
5–48
hr
afte
rla
std
osi
ng
37.5
mg
/k
gb
w/
day
gen
isti
n(a
dm
inis
tere
dd
ose
inag
lyco
ne
equ
ival
ents
)
1920
0[5
18
9],
Cm
ax
5600
[15
13]
,C
max
29%
atC
max
Jeff
erso
net
al.
(200
9a)
Mo
use
,C
D-1
PN
D1–
5,fe
mal
e,n
54–
6/ti
me
po
int
Ora
l,b
loo
dco
llec
ted
0.5–
48h
raf
ter
last
do
sin
g
37.5
mg
/k
gb
w/
day
B10
00[2
70
.2],
Cm
ax
Jeff
erso
net
al.
(200
9a)
Co
nv
ersi
on
sto
mg/
lite
rre
fer
tog
enis
tein
equ
ival
ents
.G
D,
ges
tati
on
ald
ay;
PN
D,
po
stn
atal
day
;sc
,su
bcu
tan
eou
s.aV
alu
esas
sum
edto
be
exp
ress
edin
mea
n7
var
ian
ce[u
nd
efi
ne
d].
bD
ieta
ryan
dg
avag
etr
eatm
ent
pro
vid
edeq
uiv
alen
td
ose
s.[G
av
ag
ed
ose
ssa
idto
be
eq
uiv
ale
nt
tod
ieta
ryd
ose
s,w
hic
hsu
gg
est
sth
at
fee
din
tak
ew
as
ab
ou
t3
6g
/rat
.Th
ise
stim
ate
ap
pe
ars
rea
son
able
toth
eE
xp
ert
Pa
ne
l.].
c Val
ues
esti
mat
edfr
om
ag
rap
hb
yC
ER
HR
.
431NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

et al. (2000) used a high daily dose rate of 50 mg/kg bw.The greatly decreased half-life at the higher dose mayhave resulted in part from saturation of glucuronidationand, hence, reduced enterohepatic circulation. Because itis expected that protein binding is saturated at highgenistein doses, a much smaller portion of the higherdose would be bound to plasma proteins, contributing tothe shorter half-life. In neonatal mice, elimination halflives were reported at 12 to 16 hr for genistein aglyconeand 16 to 19 hr for genistein conjugate.
SUMMARY OF DEVELOPMENTAL TOXICITYOF SOY INFANT FORMULA, INDIVIDUAL
ISOFLAVONES FOUND IN SOY INFANTFORMULA, OR OTHER SOY TREATMENTS
Human Data
No human data were identified on exposures to theindividual isoflavones found in soy formula.
Studies on outcomes after feeding soy formula ininfancy are summarized in Table 6. Most of the studieson growth have not detected differences in growth offull-term infants fed soy and cow-milk formula, but mostof the studies had small sample sizes (Jung and Carr,1977; Steichen and Tsang, 1987; Venkataraman et al.,1992; Mimouni et al., 1993; Lasekan et al., 1999). Agostoniet al. (2007) reported better short-term weight gain ininfants with immunoglobulin E-mediated cow’s milkallergy who were fed a rice or casein hydrolysate formulacompared with soy formula. Some studies reported thatinfants given soy formula had transiently lower bone-mineral content than infants fed cow-milk formula(Kohler et al., 1984) or human milk (Chan et al., 1987);however, other studies reported that serum concentra-tions of bone accretion-related hormones and minerals insoy formula-fed infants demonstrated the appropriatecompensatory response to the vitamin and mineralcontent of the food source (Hillman, 1988; Hillmanet al., 1988; Mimouni et al., 1993). Reports in very lowbirth-weight and preterm babies suggest that soyformula may not support adequate growth and bonedevelopment in these groups (Kulkarni et al., 1984). Soyinfant formula is not marketed for use in preterm infants,and the AAP recommends against its use in this group.
Studies comparing soy- and cow milk-formula feedingin infants with familial predispositions to allergic diseasedid not detect an advantage of one formula over another(Gruskay, 1982; Hillman et al., 1988; Klemola et al., 2002),but most of these studies have demonstrated that breastfeeding was superior to the use of either formula.A series of studies by Chandra (1989, x864; 1989, x765;1991, x865; 1997, x549; 1998, x550) related to allergicdisease were considered by the Expert Panel butclassified as no utility for the evaluation because thevalidity of the articles has been challenged and thescientific integrity cannot be confirmed.
Case reports suggest that hypothyroid infants on oralthyroxine therapy fed soy formula may exhibit lessefficient absorption of the thyroxine from the intestine.This effect has been attributed to fecal wastage withdecreased enterohepatic circulation (Shepard, 1960;Chorazy et al., 1995; Jabbar et al., 1997). Although theaddition of iodine and reduction in fiber content of soyformulas is a partial solution, one article noted that the
problem may occur if formula intolerance increases stoolfrequency (Chorazy et al., 1995). Conrad et al. (2004)reported that infants with CH-fed soy formula had aprolonged elevation of TSH levels compared with infantswith CH-fed nonsoy formula, suggesting that infantswith CH-fed soy formula may require higher levothyr-oxine doses to achieve normal thyroid function tests.
A case–control study of premature thelarche used 130subjects from 552 potentially eligible girls (Freni-Titulaeret al., 1986). Age-matched controls were recruited,consisting in some but not all instances of friends ofthe cases. Parents were interviewed with regard to familyhistory and possible exposures, including the use of soyformula. Multivariate analysis did not show a significantrelationship between premature thelarche and soy-formula feeding. When the analysis was restricted togirls with onset of premature thelarche before 2 years ofage, a significant association with soy-formula feedingwas identified (OR 2.7, 95% CI 1.1–6.8). Other significantfactors included maternal ovarian cysts and consumptionof chicken. Consumption of corn was protective.
Strom et al. (2001) identified 952 adults who hadparticipated as infants in a controlled, nonrandomizedformula trial. There were 248 adults who had been givensoy formula and 563 who had been given cow-milkformula. The subjects were asked about height, weight,sexual maturation, education level, and reproductiveoutcomes. No formula-related differences in height,weight, body-mass index, or sexual maturation wereidentified. Women who had been given soy formula hadlonger menstrual periods (mean 0.37 days longer, 95% CI0.06–0.68 days) and there was a soy formula-associatedincrease in the risk of experiencing severe menstrualdiscomfort (RR 1.7, 95% CI 1.04–3.00 compared with noor mild discomfort, unadjusted). After adjustment formultiple comparisons, these findings were no longerstatistically significant (more than 30 different endpointswere evaluated).
In a study comparing women with and without breastcancer by Boucher et al. (2008), the point estimates of theodds ratios were less than unity. However the variabilityof these estimates indicate that feeding soy formula wasnot statistically significantly associated with developingbreast cancer (soy formula only during first 4 months oflife: OR 5 0.42, 95% CI 5 0.13–1.40; soy formula onlyduring 5–12 months of age: OR 5 0.59, 95% CI 5 0.18–1.90).
Experimental Animal Studies on the IndividualIsoflavones Found in Soy Formula
Studies reporting the most sensitive and apparentlytreatment-related developmental effects are summarizedin Table 7 for oral and parenteral exposures in mice,Table 8 for oral and parenteral exposures in rats, andTable 9 for animal studies in other mammalian species. Inthese tables, dose levels have been converted to mg/kgbw. In general, the most complete information wasavailable from parenteral exposure studies in mice andoral exposure studies in rats. In cases where doses wereconverted to mg/kg bw/day values, ranges were oftenestimated over periods of gestation or lactation or indifferent stages of the offspring’s life.
Pre- and postnatal survival, growth, and gen-eral development endpoints. Oral exposure studiesconducted in rats suggested that genistein exposures can
432 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Table 6Studies of Human Infants Exposed to Soy Formula
Comparison groups Major findings Reference Utilitya
Growth and NutritionInfants with immunoglobulin E-mediated cow’s milk
allergy fed breastmilk (n 5 32) (reference) andinfants randomly assigned to one of three specialformulas during a feeding period of 6 to 12 monthsof age: soy formula (n 5 32), a casein hydrolysate(n 5 31), and a rice hydrolysate (n 5 30)
The 6- to 12-month change in weight-for-age z-scorefor the soy formula group was significantly lowercompared to the rice (0.18, p 5 0.019) and casein(0.16, p 5 0.029) groups. This indicates better short-term weight gain with hydrolyzed products whencompared with soy formula
Agostoni et al.(2007)
Limited
Infants randomized to one of two different soyformulas (n 5 10/group) and 10 breast-fedinfants (for comparisons to 4 months of age)
k bone mineral density and plasma zinc higher in soyformula-fed infants at 4 months of age; unable todetect difference between soy formula-fed infantsand historical controls in these parameters at 6 and12 months
Chan et al.(1987)
Limited
No effect: on measures of growth and serumchemistries
Infants fed soy infant formula (n 5 28) and infantsfed cow-milk formula (n 5 30) for 6 months. Thesoy formula was not methionine-supplementedas is modern soy formula
Soy-fed infants, especially girls, lagged in volumeconsumed and growth from 2 to 4 months of age;k weight gain per fluid ounce of formula;k vomiting; k number of infants hungry afterfeeding; m loose stools
Cherry et al.(1968)
Limited
No effect: on anemia or hematologic parametersInfants on soy formula (n 5 11), cow-milk formula
(n 5 11), or human milk (n 5 9), followed for 1year
Unable to detect a difference in bone width or bonemineral content; differences in serum and urineminerals and bone-related hormones wereconsistent with homeostatic adjustments todifferent mineral content in the food sources
Hillman et al.(1988, 1988)
Limited
Infants on soy formula (n 5 20) and infants on cow-milk formula (n 5 20) for 16 weeks
Unable to detect a difference between the 2 groups ingrowth parameters, hematology measures, or bloodchemistry
Jung and Carr(1977)
Limited
Infants on soy formula (n 5 13), infants on cow-milk formula (n 5 20), and breast-fed infants(n 5 26). Infants selected at 6 weeks of age andfollowed for 1 year
Weight at birth; unable to detect growth differencesbetween groups after 6 weeks of age; soy-fedinfants showed slower bone mineralization at 3months but not thereafter
Kohler et al.(1984)
Limited
Very low birth-weight infants given soy formula,cow-milk formula, or high-calorie cow-milkformula until 3–4 months of age
k serum phosphorus and serum alkaline phosphataseafter three weeks of age
Kulkarni et al.(1984)
Limited
Infants on soy formula with (n 5 73) or without(n 5 73) supplemental nucleotides, and infantsbreast-fed for 2 months and then given cow-milk formula (n 5 67); infants followed to 12months of age
Groups were not comparable in weight and lengthat birth. No group differences in growth weredetected after 6 months of age
Lasekan et al.(1999)
Limited
Infants given 1 of 2 different soy formulas (n 5 21/group), infants given cow-milk formula (n 5 20),and breast-fed infants (n 5 10), followed for 1year; solid food introduced at 3 months inbreast-fed and 4 months in formula-fed groups
m length gain; m serum 1,25-dihydroxyvitamin D in 1of the soy-formula groups at 8 weeks andthereafter; m serum phosphorus in combinedformula groups at 8 weeks. Unable to detect effectof feeding type on weight and head circumference;bone mineral content and bone width; bloodchemistry
Mimouni et al.(1993)
Limited
Infants being breastfed (n 5 401) and infantsassigned to either cow-milk (n 5 839) or soybased formula (n 5 239) based on family historyof allergy. Evaluated monthly until 6 months ofage and bi-monthly thereafter
No differences in length or weight were detectedbetween the 3 feeding groups.
Sellars et al.(1971)
Limited
Infants fed soy formula (n 5 18) or cow-milkformula (n 5 17) for 12 months
k bone mineral content; Unable to detect differencesin energy intake or growth
Steichen andTsang (1987)
Limited
Infants exclusively fed soy formula (n 5 20), cow-milk formula (n 5 19), or breast-fed (n 5 17) for 4months and followed until 6 months of age
m bone mineral content and bone width in soyformula-fed infants compared to breast-fed infants;Unable to detect group differences in growth andserum measurements related to bone accretion
Venkataramanet al. (1992)
Limited
Allergy, immunology, and gastrointestinal effectsInfants with a family history of major allergy
assigned to soy formula (n 5 79) or cow-milkformula (n 5 201), followed 17 years; 48 infantswith a family history of major allergy werebreast fed
Breast-fed children were significantly less likely todevelop allergy after 3 years than soy or cow-milkformula-fed children
Gruskay (1982) Limited
No significant difference in development of allergicdisease by formula type
433NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Table 6Continued
Comparison groups Major findings Reference Utilitya
Infants with cow milk allergy diagnosed at age 2–11 months were randomly assigned toextensively hydrolyzed formula (n 5 90) or soyformula (n 5 80) and followed until 2 years ofage
Parents suspected adverse reaction in 28% of subjectson soy formula and 11% of subjects on extensivelyhydrolyzed formula. Among children younger than6 months, 40% were suspected by parents of havingadverse reaction to soy formula
Klemola et al.(2002)
Limited
Infants with milk allergy fed whey formula(n 5 76) or soy formula (n 5 72), examined atage 2, 3, and 4 years
Non-significant increase in soy IgE levels in soyformula-fed infants (OR 2.28, 95% CI 0.90–5.76;p 5 0.082)
Klemola et al.(2005)
Limited
No effect: prevalence of peanut IgE positivity (OR1.27, 95% CI 0.35–4.71; p 5 0.717)
Children with peanut allergy (n 5 49), childrenwith atopy (n 5 70), and non-allergic children(n 5 140) surveyed for infant feeding history
Soy consumption was a significant and independentrisk factor for peanut allergy (adjusted OR 2.61;95% CI 1.31–5.20)
Lack et al. (2003) Limited
Infants given 1 of 2 different soy formulas (n 5 92,94), one of which contained added nucleotides,and breast-fed children who were weaned tocow-milk formula at 2 months (n 5 81); 1-yearfollow-up
Unable to detect a difference by feeding group in antibodyresponse to immunizations (except Hemophilus influenzab), parent-reported diarrhea, or otitis media. Noconsistent differences were detected in immune status,maturation, or level of immunocompetence betweensoy and cow-milk formula-fed infants
Ostrom et al.(2002), Cordleet al. (2002)
Limited
Infants with cow’s milk allergy fed soy formula(n 5 84, mean starting age 5 7.8 mths) or aextensively hydrolyzed whey formula (HWF)(n 5 84, mean starting age 5 7.5 mths),examined until 48 mths of age
m percentage of energy; mintake of zinc and vitamin E;k riboflavin
Seppo et al.(2005)
Limited
No effect: percentages of abnormally low lab values;percentages of high alkaline phosphatase; growthmeasured between 1 to 4 years; nutritional status
Thyroid functionInfant with congenital hypothyroidism who was
fed soy formula (n 5 1)Oral thyroxine therapy did not decrease thyroid-
stimulating hormone until cow-milk formula wassubstituted for soy formula
Chorazy et al.(1995)
Limited
Infants fed soy formula (n 5 8) and infants fednonsoy formula (n 5 70)
Infants with congenital hypothyroidism fed soyformula had prolonged increase of TSH levelscompared to infants with congenitalhypothyroidism fed non-soy formula
Conrad et al.(2004)
Limited
No effect: on total T4 between groups; weight; height;weight for height
Infants with congenital hypothyroidism who werefed soy formula (n 5 3)
Switching from soy formula to cow-milk formularesulted in increased absorption of thyroidreplacement doses
Jabbar et al.(1997)
Limited
Children with goiter related to soy formula-feeding (n 5 3)
Improvement with discontinuation of soy formula(n 5 2) or addition of iodine (n 5 1)
Shepard (1960) Limited
Reproductive function or tissue effectsAdults with breast cancer (n 5 372) and without
breast cancer (n 5 356) who had been fed soyformula, cow-milk formula, or breastmilk
A reduced, but non-significant, association was foundbetween soy formula intake and breast cancer. Soyformula only during fist 4 months of life: OR 5 0.42,95% 5 0.13–1.40. Soy formula only during 5–12months of age: OR 5 0.59, 95% CI 5 0.18–1.90
Boucher et al.(2008)
Limited
Girls with premature thelarche (n 5 130) and age-matched control subjects, retrospectivequestioning of parents about infant feeding
Unable to detect a significant association overallbetween premature thelarche and soy infantformula intake; restriction of multivariate analysisto subjects with thelarche before age 2 yearsshowed significant association (OR 2.7; 95% CI 1.1–6.8, p 5 0.029)
Freni-Titulaeret al. (1986)
Limited
Adults who had been fed soy (n 5 248) or cow-milk formula (n 5 563) during infancy as part ofa controlled trial; interviewed at 20–34 years ofage
Unable to detect infant feeding-related differences inadult height, weight, body-mass index, or sexualmaturation history; duration of menstrual bleedingwas 0.37 days longer and severe menstrualdiscomfort was more common in women fed withsoy formula than with cow-milk formula
Strom et al.(2001)
Limited
Other endpointsInfants on soy formula (n 5 16, some with
cholesterol added), cow-milk formula (n 5 10),or breast-fed with supplemental cow-milkformula (n 5 12)
Breast-fed infants had the highest serum cholesterollevels and lowest fractional cholesterol synthesisrate. There was a significant inverse relationshipbetween cholesterol intake and fractionalcholesterol synthesis
Cruz et al. (1994) Limited
aThe term utility refers to the applicability to the purpose of drawing conclusions on whether or not a chemical adversely affects reproduction.
434 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011
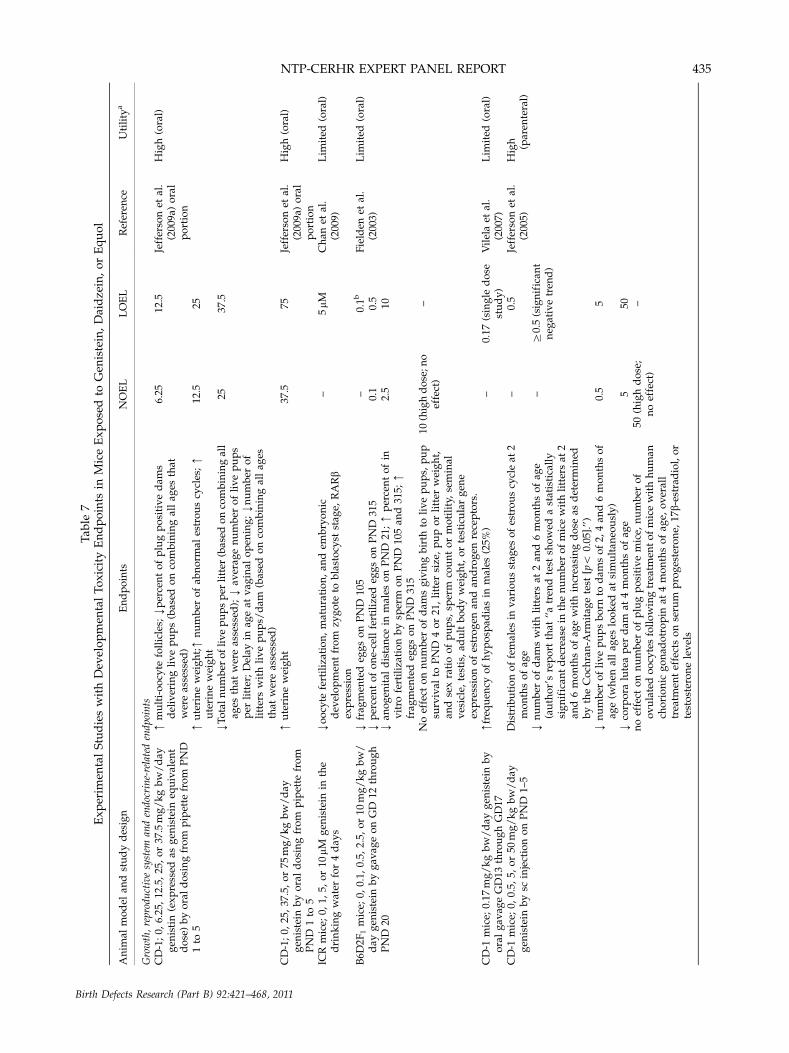
Tab
le7
Ex
per
imen
tal
Stu
die
sw
ith
Dev
elo
pm
enta
lT
ox
icit
yE
nd
po
ints
inM
ice
Ex
po
sed
toG
enis
tein
,D
aid
zein
,o
rE
qu
ol
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity
a
Gro
wth
,re
prod
uct
ive
syst
eman
den
docr
ine-
rela
ted
endp
oin
tsC
D-1
;0,
6.25
,12
.5,
25,
or
37.5
mg
/k
gb
w/
day
gen
isti
n(e
xp
ress
edas
gen
iste
ineq
uiv
alen
td
ose
)b
yo
ral
do
sin
gfr
om
pip
ette
fro
mP
ND
1to
5
mm
ult
i-o
ocy
tefo
llic
les;
kp
erce
nt
of
plu
gp
osi
tiv
ed
ams
del
iver
ing
liv
ep
up
s(b
ased
on
com
bin
ing
all
ages
that
wer
eas
sess
ed)
6.25
12.5
Jeff
erso
net
al.
(200
9a)
ora
lp
ort
ion
Hig
h(o
ral)
mu
teri
ne
wei
gh
t;m
nu
mb
ero
fab
no
rmal
estr
ou
scy
cles
;m
ute
rin
ew
eig
ht
12.5
25
kT
ota
ln
um
ber
of
liv
ep
up
sp
erli
tter
(bas
edo
nco
mb
inin
gal
lag
esth
atw
ere
asse
ssed
);k
aver
age
nu
mb
ero
fli
ve
pu
ps
per
litt
er;
Del
ayin
age
atv
agin
alo
pen
ing
;k
nu
mb
ero
fli
tter
sw
ith
liv
ep
up
s/d
am(b
ased
on
com
bin
ing
all
ages
that
wer
eas
sess
ed)
2537
.5
CD
-1;
0,25
,37
.5,
or
75m
g/
kg
bw
/d
ayg
enis
tein
by
ora
ld
osi
ng
fro
mp
ipet
tefr
om
PN
D1
to5
mu
teri
ne
wei
gh
t37
.575
Jeff
erso
net
al.
(200
9a)
ora
lp
ort
ion
Hig
h(o
ral)
ICR
mic
e;0,
1,5,
or
10mM
gen
iste
inin
the
dri
nk
ing
wat
erfo
r4
day
sk
oo
cyte
fert
iliz
atio
n,
mat
ura
tio
n,
and
emb
ryo
nic
dev
elo
pm
ent
fro
mzy
go
teto
bla
sto
cyst
stag
e,R
ARb
exp
ress
ion
–5mM
Ch
anet
al.
(200
9)L
imit
ed(o
ral)
B6D
2F1
mic
e;0,
0.1,
0.5,
2.5,
or
10m
g/
kg
bw
/d
ayg
enis
tein
by
gav
age
on
GD
12th
rou
gh
PN
D20
kfr
agm
ente
deg
gs
on
PN
D10
5–
0.1b
Fie
lden
etal
.(2
003)
Lim
ited
(ora
l)k
per
cen
to
fo
ne-
cell
fert
iliz
edeg
gs
on
PN
D31
50.
10.
5k
ano
gen
ital
dis
tan
cein
mal
eso
nP
ND
21;m
per
cen
to
fin
vit
rofe
rtil
izat
ion
by
sper
mo
nP
ND
105
and
315;
mfr
agm
ente
deg
gs
on
PN
D31
5
2.5
10
No
effe
cto
nn
um
ber
of
dam
sg
ivin
gb
irth
toli
ve
pu
ps,
pu
psu
rviv
alto
PN
D4
or
21,
litt
ersi
ze,
pu
po
rli
tter
wei
gh
t,an
dse
xra
tio
of
pu
ps,
sper
mco
un
to
rm
oti
lity
,se
min
alv
esic
le,
test
is,
adu
ltb
od
yw
eig
ht,
or
test
icu
lar
gen
eex
pre
ssio
no
fes
tro
gen
and
and
rog
enre
cep
tors
.
10(h
igh
do
se;n
oef
fect
)–
CD
-1m
ice;
0.17
mg
/k
gb
w/
day
gen
iste
inb
yo
ral
gav
age
GD
13th
rou
gh
GD
17m
freq
uen
cyo
fh
yp
osp
adia
sin
mal
es(2
5%)
–0.
17(s
ing
led
ose
stu
dy
)V
ilel
aet
al.
(200
7)L
imit
ed(o
ral)
CD
-1m
ice;
0,0.
5,5,
or
50m
g/
kg
bw
/d
ayg
enis
tein
by
scin
ject
ion
on
PN
D1–
5D
istr
ibu
tio
no
ffe
mal
esin
var
iou
sst
ages
of
estr
ou
scy
cle
at2
mo
nth
so
fag
e–
0.5
Jeff
erso
net
al.
(200
5)H
igh (par
ente
ral)
kn
um
ber
of
dam
sw
ith
litt
ers
at2
and
6m
on
ths
of
age
(au
tho
r’s
rep
ort
that
‘‘a
tren
dte
stsh
ow
eda
stat
isti
call
ysi
gn
ific
ant
dec
reas
ein
the
nu
mb
ero
fm
ice
wit
hli
tter
sat
2an
d6
mo
nth
so
fag
ew
ith
incr
easi
ng
do
seas
det
erm
ined
by
the
Co
chra
n-A
rmit
age
test
[po
0.05
].’’
)
–Z
0.5
(sig
nif
ican
tn
egat
ive
tren
d)
kn
um
ber
of
liv
ep
up
sb
orn
tod
ams
of
2,4
and
6m
on
ths
of
age
(wh
enal
lag
eslo
ok
edat
sim
ult
aneo
usl
y)
0.5
5
kco
rpo
ralu
tea
per
dam
at4
mo
nth
so
fag
e5
50n
oef
fect
on
nu
mb
ero
fp
lug
po
siti
ve
mic
e,n
um
ber
of
ov
ula
ted
oo
cyte
sfo
llo
win
gtr
eatm
ent
of
mic
ew
ith
hu
man
cho
rio
nic
go
nad
otr
op
inat
4m
on
ths
of
age,
ov
eral
ltr
eatm
ent
effe
cts
on
seru
mp
rog
este
ron
e,17b-
estr
adio
l,o
rte
sto
ster
on
ele
vel
s
50(h
igh
do
se;
no
effe
ct)
–
435NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le7
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity
a
CD
-1m
ice;
0o
r50
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
no
nP
ND
1to
5k
imp
lan
tati
on
site
s(b
oth
nu
mb
ero
fm
ice
and
nu
mb
ero
fsi
tes/
mo
use
);k
per
cen
tp
reg
nan
tm
ice;
kn
um
ber
of
corp
ora
lute
a
–50
(sin
gle
do
sest
ud
y)
Jeff
erso
net
al.
(200
5)L
imit
ed(p
aren
tera
l)
CD
-1m
ice;
0o
r50
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
no
nP
ND
1to
5k
nu
mb
ero
fem
bry
os
coll
ecte
dp
erm
ou
sefo
llo
win
gh
CG
adm
inis
trat
ion
;kn
um
ber
and
size
of
imp
lan
tati
on
site
sin
mic
ew
ho
wer
ere
cip
ien
tso
ftr
ansf
erre
db
last
ocy
sts
ob
tain
edfr
om
un
trea
ted
mic
e
–50
(sin
gle
do
sest
ud
y)
Jeff
erso
net
al.
(200
9b)
Lim
ited
(par
ente
ral)
No
effe
cto
nth
en
um
ber
of
ov
ula
ted
egg
s,ti
min
go
fd
evel
op
men
to
rp
erce
nta
ge
of
emb
ryo
sto
reac
hb
last
ocy
stag
e,th
en
um
ber
of
liv
ep
up
s,o
rli
tter
size
50(h
igh
do
se;n
oef
fect
)–
CD
-1;
0,12
.6,
20,
or
25m
g/
kg
bw
/d
ayg
enis
tein
scin
ject
ion
fro
mP
ND
1to
5m
ute
rin
ew
eig
ht
lim
ited
(par
ente
ral)
12.6
20Je
ffer
son
etal
.(2
009a
)sc
po
rtio
n
Lim
ited
(par
ente
ral)
CD
-1m
ice;
0o
r50
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
no
nP
ND
1to
5m
nu
mb
ero
fre
pro
du
ctiv
ele
sio
ns:
no
corp
ora
lute
a,ab
no
rmal
ov
idu
cth
isto
log
y,u
teri
ne
squ
amo
us
met
apla
sia,
cyst
icen
do
met
rial
hy
per
pla
sia
and
ute
rin
eca
rcin
om
a
–50
(sin
gle
do
sest
ud
y)
New
bo
ldet
al.
(200
1)L
imit
ed(p
aren
tera
l)
CD
-1m
ice;
0,0.
5,o
r10
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
no
nG
D15
to18
mb
od
yw
eig
ht
gai
n;
0.5–
1d
ayea
rlie
ro
nse
to
fv
agin
alo
pen
ing
;m
estr
ou
scy
cle
len
gth
,1.
2–2
day
s;m
len
gth
of
die
stru
s
–0.
5N
ikai
do
etal
.(2
004)
kn
um
ber
of
anim
als
wit
hco
rpo
ralu
tea
at4
wee
ks
of
age,
tran
sien
tef
fect
0.5
10
ICR
mic
e;0
or
1000
mg
/k
gb
w/
day
gen
iste
inb
yin
ject
ion
on
PN
D1
to5
kte
stic
ula
rm
RN
Aex
pre
ssio
no
fE
Ra,
ERb,
AR
and
lam
inin
-g2
–10
00(s
ing
led
ose
stu
dy
)A
dac
hi
etal
.(2
004)
Lim
ited
(par
ente
ral)
No
effe
cto
nb
od
yw
eig
ht,
abso
lute
or
rela
tiv
ete
stis
wei
gh
to
rh
isto
log
ical
chan
ges
inth
ete
stes
at12
wee
ks
of
age
1000
(hig
hd
ose
;n
oef
fect
)–
CD
-1p
reg
nan
tm
ice;
0,0.
2o
r2
mg
/k
gb
w/
day
of
gen
iste
inb
yip
inje
ctio
nfr
om
GD
1to
21N
oef
fect
on
gro
sso
rh
isto
log
ical
ano
mal
ies
of
the
ute
rus,
or
Hox
a10
mR
NA
exp
ress
ion
inth
eu
teru
s2
(hig
hd
ose
;n
oef
fect
)–
Ak
bas
etal
.(2
007)
Lim
ited
(par
ente
ral)
CD
-1p
reg
nan
tm
ice;
0o
r2
mg
/k
gb
w/
day
of
dai
dze
inb
yip
inje
ctio
nfr
om
GD
1to
21N
oef
fect
on
gro
sso
rh
isto
log
ical
ano
mal
ies
of
the
ute
rus,
or
Hox
a10
mR
NA
exp
ress
ion
inth
eu
teru
s2
(hig
hd
ose
;n
oef
fect
)–
CD
-1m
ice;
0,1,
10,
or
100mg
/d
ay(0
,0.
5,5,
or
50m
g/
kg
bw
/d
ay)
gen
iste
inb
ysc
inje
ctio
no
nP
ND
1to
5
mm
ult
i-o
ocy
tefo
llic
les
550
Jeff
erso
net
al.
(200
2)L
imit
ed(p
aren
tera
l)
C57
BL
/6,
ERa
and
ERb
mic
e;0,
1,10
,o
r10
0mg
/d
ay(0
,0.
5,5,
or
50m
g/
kg
bw
/d
ay)
gen
iste
inb
ysc
inje
ctio
no
nP
ND
1–5
mm
ult
i-o
ocy
tefo
llic
les
inE
Ra
and
no
effe
ctin
ERb
0.5
5
CD
-1m
ice;
50m
g/
kg
bw
/d
ayg
enis
tein
by
scin
ject
ion
fro
mP
ND
1to
5m
nu
mb
ero
fm
ult
i-o
ocy
tefo
llic
les;
few
ersi
ng
leo
ocy
tes
–50
(sin
gle
do
sest
ud
y)
Jeff
erso
net
al.
(200
6)L
imit
ed(p
aren
tera
l)C
D-1
mic
e;0
or
10m
g/
kg
bw
/d
ayg
enis
tein
by
scin
ject
ion
on
PN
D15
to18
acce
lera
tio
no
fv
agin
alo
pen
ing
–10
(sin
gle
do
sest
ud
y)
Nik
aid
oet
al.
([20
05)
Lim
ited
(par
ente
ral)
No
effe
ct:
on
bo
dy
wei
gh
t,es
tro
us
cycl
ing
,p
oly
ov
ula
ro
var
ian
foll
icle
s,m
orp
ho
log
ical
abn
orm
alit
ies
inv
agin
alo
ru
teri
ne
epit
hel
ium
,o
rm
amm
ary
gla
nd
dev
elo
pm
ent
10(h
igh
do
se;n
oef
fect
)–
ICR
mic
e;0,
7,71
,an
d71
4m
g/
kg
bw
/d
ayg
enis
tein
by
scin
ject
ion
on
PN
D1
to5
kA
Rm
RN
Aex
pre
ssio
nin
test
is7
71S
hib
ayam
aet
al.
(200
1)L
imit
ed(p
aren
tera
l)k
ERa
mR
NA
exp
ress
ion
inte
stis
7171
4 –
436 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

No
effe
cto
nte
stis
wei
gh
t,sp
erm
cou
nt,
or
sper
mm
oti
lity
at12
wee
ks
of
age
714
(hig
hd
ose
;n
oef
fect
)M
ale
Han
-NM
RI
mic
e;0,
0.1
or
1m
g/
day
(50
or
500
mg
/k
gb
w/
day
)g
enis
tein
by
scin
ject
ion
on
PN
D1
to3
kv
entr
alp
rost
ate
wei
gh
tin
adu
lth
oo
d(h
isto
log
ical
chan
ges
ob
serv
ed)
–50
Str
auss
etal
.(1
998)
Lim
ited
(par
ente
ral)
kco
agu
lati
ng
gla
nd
wei
gh
t;m
his
tolo
gic
abn
orm
alit
ies
(hy
per
pla
sia
and
dis
org
aniz
atio
no
fth
eep
ith
eliu
mo
fth
ep
rost
atic
coll
ecti
ng
du
cts,
ven
tral
lob
es,
and
sem
inal
ves
icle
s);m
fib
rom
usc
ula
rst
rom
aan
din
flam
mat
ory
cell
sin
po
ster
ior
per
iure
thra
lre
gio
n
5050
0
No
effe
cto
nc-
fos
mR
NA
exp
ress
ion
inp
rost
atic
ure
thra
500
(hig
hd
ose
;n
oef
fect
)–
Mam
mar
ygl
and
deve
lopm
ent
and
carc
inog
enes
isC
D-1
mic
e;0
orB
0.7–
0.8
mg
/k
gb
w/
day
gen
iste
inb
yin
ject
ion
on
GD
15to
20m
mam
mar
yg
lan
dep
ith
elia
lar
eao
nP
ND
35(b
ut
no
to
nP
ND
25o
rP
ND
46);m
den
sity
of
TE
Bs
on
PN
D35
and
45;
mP
ND
25b
od
yw
eig
ht;
del
ayed
eye
op
enin
g;
acce
lera
ted
tim
ing
of
vag
inal
op
enin
g
–B
0.7–
0.8
(sin
gle
do
sele
vel
)H
ilak
ivi-
Cla
rke
etal
.(1
998)
Lim
ited
(par
ente
ral)
No
effe
cto
nn
um
ber
of
off
spri
ng
bo
rn,
PN
D1
bo
dy
wei
gh
t,d
iffe
ren
tiat
ion
of
bre
ast
tiss
ue
(ass
esse
du
sin
gd
ensi
tyo
fT
EB
san
dlo
bu
loal
veo
lar
un
its)
,se
rum
17b-
estr
adio
lo
res
tro
us
cycl
icit
y
B0.
7–0.
8(h
igh
do
se;
no
effe
ct)
–
CD
-1m
ice;
0,0.
5,5,
or
50m
g/
kg
bw
/d
ayg
enis
tein
by
scin
ject
ion
on
PN
D1
to5
Mam
mar
yg
lan
dh
orm
on
ere
cep
tor
lev
els:m
PR
pro
tein
exp
ress
ion
at5
wee
ks,
kE
Ra
mR
NA
at5
and
6w
eek
s(c
om
bin
ed)
–0.
5P
adil
la-B
ank
set
al.
(200
6)L
imit
ed(p
aren
tera
l)
md
uct
alel
on
gat
ion
at6
wee
ks;
mE
Rb
mR
NA
inm
amm
ary
gla
nd
at5
and
6w
eek
s(c
om
bin
ed)b
–0.
5b
kn
um
ber
of
TE
Bs
at6
wee
ks;k
nu
mb
ero
fb
ran
chp
oin
tsat
5w
eek
s(b
ut
no
tat
6w
eek
s);
mam
mar
yg
lan
dm
orp
ho
log
yat
9m
on
ths:
kal
veo
lar
dev
elo
pm
ent,
dil
ated
and
/o
rd
ilat
edb
ead
edd
uct
s
0.5
5
kn
um
ber
of
TE
Bs
at5
wee
ks;
km
amm
ary
gla
nd
du
ctal
gro
wth
at5
or
6w
eek
s;k
aver
age
area
of
mam
mar
yg
lan
dat
5o
r6
wee
ks;
alte
red
estr
ou
scy
clic
ity
550
kn
um
ber
of
pu
ps
per
litt
er,
litt
erw
eig
hts
,p
up
wei
gh
ts,
%p
up
ssu
rviv
ing
un
til
wea
nin
g5
50(n
oli
ve
pu
ps
del
iver
ed)
No
effe
cto
nci
rcu
lati
ng
lev
els
of
estr
adio
lo
rp
rog
este
ron
e,li
tter
size
,li
tter
wei
gh
ts,
calc
ula
ted
aver
age
pu
pw
eig
hts
,o
r%
of
pu
ps
surv
ivin
gu
nti
lw
ean
ing
50(h
igh
do
se;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
;50
mg/
day
(au
tho
rses
tim
ated
that
the
do
ses
rece
ived
wer
e1.
25to
3.3
mg
/k
gb
wg
enis
tein
by
scin
ject
ion
on
PN
D8
to20
km
amm
ary
epit
hel
ial
den
sity
;kte
rmin
alen
db
ud
nu
mb
ers;
min
crea
sed
lob
ulo
alv
eola
rst
ruct
ure
s;at
8w
eek
so
fag
e;m
BR
CA
1ex
pre
ssio
n;m
ERa
exp
ress
ion
inlo
bu
les
at3
and
8w
eek
s
–50
(sin
gle
do
sest
ud
y)
Cab
anes
etal
.(2
004)
Lim
ited
(par
ente
ral)
Oth
er/m
echa
nis
tic
Av
ym
ice;
250
mg
/k
gg
enis
tein
ind
iet
bef
ore
mat
ing
thro
ug
hp
reg
nan
cyan
dla
ctat
ion
;as
sess
men
to
fh
eter
ozy
go
us
via
ble
yel
low
ago
uti
(Av
y/a
)o
ffsp
rin
g
Sh
ift
inco
atco
lor
ph
eno
typ
eto
war
ds
the
pse
ud
o-a
go
uti
ph
eno
typ
ean
dre
du
ced
bo
dy
wei
gh
tin
pse
ud
o-a
go
uti
ph
eno
typ
e;h
yp
erm
eth
yla
tio
n
–25
0(s
ing
led
ose
stu
dy
)D
oli
no
yet
al.
(200
6)L
imit
ed(o
ral)
No
effe
cto
nli
tter
size
,m
ean
pu
pw
eig
ht,
per
cen
tsu
rviv
alo
rse
xra
tio
250
(hig
hd
ose
;n
oef
fect
)–
437NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le7
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity
a
Av
yfe
mal
em
ice;
wer
eas
sig
ned
too
ne
of
fou
rm
od
ifie
dd
iets
for
two
wee
ks
pri
or
tom
atin
gw
ith
Av
y/a
mal
esan
dth
rou
gh
ou
tp
reg
nan
cyan
dla
ctat
ion
:(1
)m
od
ifie
dA
IN-9
3d
iet
(co
rno
ilsu
bst
itu
ted
for
soy
bea
no
il);
(2)
mo
dif
ied
AIN
-93
die
tsu
pp
lem
ente
dw
ith
50m
gB
PA
/k
gd
iet;
(3)
mo
dif
ied
AIN
-93
die
tsu
pp
lem
ente
dw
ith
50m
gB
PA
and
250
mg
gen
iste
in/
kg
die
t;an
d(4
)m
od
ifie
dA
IN-9
3d
iet
sup
ple
men
ted
wit
h50
mg
BP
A/
kg
die
tan
dm
eth
yl
do
no
rco
mp
ou
nd
s(4
.3m
gfo
lic
acid
,0.
53m
gv
itam
inB
12,
5g
bet
ain
e,an
d7.
97g
cho
lin
ech
lori
de/
kg
die
t)
Mat
ern
ald
ieta
ryB
PA
sig
nif
ican
tly
shif
ted
the
coat
colo
rd
istr
ibu
tio
no
fg
enet
ical
lyid
enti
cal
d22
Av
y/a
off
spri
ng
tow
ard
the
yel
low
coat
colo
rp
hen
oty
pe;
mat
ern
alsu
pp
lem
enta
tio
nw
ith
met
hy
ld
on
ors
or
gen
iste
inre
sto
red
the
coat
colo
rd
istr
ibu
tio
nin
the
BP
A-e
xp
ose
do
ffsp
rin
gto
that
ob
serv
edin
the
con
tro
ls
–25
0(s
ing
led
ose
stu
dy
)D
oli
no
yet
al.
(200
7)L
imit
ed(o
ral)
No
effe
cts:
litt
ersi
ze,
surv
ival
,w
ean
wei
gh
t,g
eno
typ
icra
tio
or
sex
rati
o25
0(h
igh
do
se;
no
effe
ct)
–
C57
Bl/
6m
ice;
0,25
,25
0,o
r12
50p
pm
gen
iste
inin
die
tfr
om
GD
14to
lact
atio
nv
iad
am(m
g/
kg
fee
d;
est
ima
ted
by
the
stu
dy
au
tho
rsto
pro
vid
eg
en
iste
in0
,2
,2
0,
or
10
0m
g/k
gb
w/
da
yto
a2
5-g
mm
ou
se)
Alt
ered
imm
un
ece
llen
dp
oin
ts,
e.g
.,v
ario
us
typ
eso
fth
ym
ocy
tes
and
sple
no
cyte
s(m
any
did
no
td
isp
lay
aco
nsi
sten
tp
atte
rno
fre
spo
nse
);m
mal
ep
up
bo
dy
wei
gh
ts;
msp
leen
wei
gh
tin
mal
es
–2b
Gu
oet
al.
(200
6)L
imit
ed(o
ral)
mm
ater
nal
bo
dy
wei
gh
t;m
mat
ern
alsp
leen
wei
gh
t2
250
ER
E-t
K-L
UC
tran
sgen
icm
ale
mic
e;0
or
50m
g/
kg
gen
iste
into
dam
sb
yo
ral
gav
age
on
PN
D4
mlu
cife
rase
acti
vit
yin
liv
er,
lun
g,
hea
rt,
thy
mu
s,te
stis
,an
db
rain
of
pu
ps
–50
(sin
gle
do
sest
ud
y)
Mo
nta
ni
etal
.(2
008)
Lim
ited
(ora
l)
ER
E-t
K-L
UC
mic
e;0,
0.5,
5,an
d50
mg
/k
gg
enis
tein
on
PN
D4
by
ora
lg
avag
eto
lact
atin
gd
am
mlu
cife
rase
acti
vit
yin
the
test
eso
fm
ale
pu
ps
550
Mo
nta
ni
etal
.(2
009)
Lim
ited
(ora
l)
CD
-1m
ice;
0o
r50
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
no
nP
ND
1to
5H
yp
erm
eth
yla
tio
nin
Nsb
p1p
rom
ote
rC
Gis
lan
d(C
G1)
inan
age
dep
end
ent
man
ner
–50
(sin
gle
do
sest
ud
y)
Tan
get
al.
(200
8)L
imit
ed(p
aren
tera
l)
m,k
5S
ign
ific
ant
incr
ease
,d
ecre
ase.
aT
he
term
uti
lity
refe
rsto
the
app
lica
bil
ity
toth
ep
urp
ose
of
dra
win
gco
ncl
usi
on
so
nw
het
her
or
no
ta
chem
ical
adv
erse
lyaf
fect
sre
pro
du
ctio
n.
bD
idn
ot
dis
pla
ya
do
se–r
esp
on
se.
438 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le8
Ex
per
imen
tal
Stu
die
sw
ith
Dev
elo
pm
enta
lT
ox
icit
yE
nd
po
ints
inR
ats
Ex
po
sed
toG
enis
tein
,D
aid
zein
,o
rE
qu
ol
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity�
Gro
wth
,re
prod
uct
ive
syst
eman
den
docr
ine-
rela
ted
endp
oin
tsS
pra
gu
e-D
awle
yra
t;0,
5,10
0,o
r50
0p
pm
gen
iste
inin
die
td
uri
ng
pre
gn
ancy
and
lact
atio
n;
on
eh
alf
of
mal
eo
ffsp
rin
gco
nti
nu
edo
nth
eg
enis
tein
die
tsat
wea
nin
g(G
/G
);se
con
dh
alf
of
mal
eo
ffsp
rin
gw
ere
giv
enco
ntr
ol
die
tsat
wea
nin
g(G
/C
);m
ult
igen
erat
ion
des
ign
.(I
nta
ke
sa
ssu
me
dto
be
sim
ila
rto
tho
sein
NC
TR
(NC
TR
,2
00
5)o
fw
hic
hth
isst
ud
yw
as
ap
art
:m
ale
s:0
,0
.3,
7,
35
mg
/kg
bw
/da
y;
fem
ale
s:0
,0
.4,
9,
44
mg
/kg
bw
/day
;fe
mal
es
du
rin
gla
cta
tio
n:
0.7
,1
5,
an
d7
8m
g/k
gb
w/d
ay)
kE
Rb
pro
tein
ind
ors
ola
tera
lp
rost
ate
of
F1
mal
esat
PN
D14
0(G
Cg
rou
po
nly
)–
5p
pm
Dal
uet
al.
(200
2)L
imit
ed(o
ral)
mse
rum
DH
Tle
vel
sin
F1
mal
esat
PN
D14
0(b
oth
G/
Gan
dG
/C
gro
up
s)5
pp
m10
0p
pm
mse
rum
test
ost
ero
ne
lev
els
inF
1m
ales
atP
ND
140
(bo
thG
/G
and
G/
Cg
rou
ps)
100
pp
m50
0p
pm
Sp
rag
ue-
Daw
ley
rat;
0,5,
25,
100,
250,
625,
and
1250
pp
mg
enis
tein
thro
ug
hd
iet
du
rin
gp
reg
nan
cyan
dla
ctat
ion
and
un
til
PN
D50
ino
ffsp
rin
g.
(Me
an
do
ses:
0.3
1,1
.7,
5.7
,1
5,
34
,8
3m
g/k
gb
w/d
ay
inp
reg
na
nt
da
ms;
0.5
6,
2.8
,11
,3
0,
73
,1
38
inla
ctat
ing
da
ms;
B0
.6,
3.0
,1
2,
30
,7
2,
an
d1
80
mg
/kg
bw
/day
inp
up
sa
fte
rw
ea
nin
g.)
Acc
eler
ated
vag
inal
op
enin
g;m
rela
tiv
ev
agin
alw
eig
ht
–S
ign
ific
ant
lin
ear
tren
dD
elcl
os
etal
.(2
001)
Lim
ited
(ora
l)H
yp
ertr
op
hy
of
mam
mar
yal
veo
lian
dd
uct
sin
mal
esat
PN
D50
c;
abn
orm
alit
ies
of
sper
mat
og
enes
is5
pp
m25
pp
m
His
top
ath
olo
gy
inv
agin
aat
PN
D50
250
pp
m62
5p
pm
Alv
eola
rp
roli
fera
tio
nin
mam
mar
yo
ffe
mal
esat
PN
D50
;m
ren
altu
bu
lem
iner
aliz
atio
nin
mal
esan
dfe
mal
es10
0p
pm
250
pp
m
kd
ams
del
iver
ing
litt
ers
andk
litt
erb
od
yw
eig
hts
atb
irth
;kre
lati
ve
ven
tral
pro
stat
ew
eig
ht
atP
ND
50;
pro
stat
ein
flam
mat
ion
;h
isto
pat
ho
log
yin
ov
arie
sat
PN
D50
;k
po
stn
atal
bo
dy
wei
gh
tsin
mal
esan
dfe
mal
es
625
pp
m12
50p
pm
Del
ayed
eye
op
enin
gan
dea
ru
nfo
ldin
g62
5p
pm
1250
pp
m(s
ign
ific
ant
lin
ear
tren
d)
No
effe
cto
nm
ater
nal
bo
dy
wei
gh
td
uri
ng
the
lact
atio
np
erio
d,
ges
tati
on
len
gth
,li
tter
size
,p
rop
ort
ion
of
liv
ep
up
s,se
xra
tio
of
pu
ps,
ano
gen
ital
dis
tan
ceo
nP
ND
2,p
rep
uti
alse
par
atio
n;
or
test
icu
lar
sper
mh
ead
or
epid
idy
mal
sper
mco
un
ts
(hig
hd
ose
;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rat;
0,5,
100,
500
pp
mg
enis
tein
ind
iet;
mu
ltig
ener
atio
nal
des
ign
(mal
es:
0,0.
3,7,
35m
g/
kg
bw
/d
ay;
fem
ales
,av
erag
een
tire
feed
ing
per
iod
:0,
0.5,
10,
51m
g/
kg
bw
/d
ay;
fem
ales
,n
on
lact
atin
g:
0,0.
4,9,
44;
and
fem
ales
,la
ctat
ing
:0,
0.7,
15,
and
78m
g/
kg
bw
/d
ay)
kp
up
wei
gh
tat
bir
th(F
5)
(no
dir
ect
gen
iste
intr
eatm
ent)
–5
pp
mN
CT
R(2
008)
Lim
ited
(ora
l)M
amm
ary
gla
nd
hy
per
pla
sia
inm
ales
(F1,
F2,
F3);k
mal
ep
up
wei
gh
td
uri
ng
lact
atio
np
erio
d(F
3)
5p
pm
100
pp
m
kli
tter
size
(F2);
dis
rup
ted
estr
ou
scy
cles
,i.
e.,m
len
gth
or
nu
mb
ero
fab
no
rmal
cycl
es(F
1);
incr
ease
dcy
cle
len
gth
(F1,
F2);k
bo
dy
wei
gh
tat
vag
inal
op
enin
g(F
1,
F2,
F3)
and
acce
lera
ted
vag
inal
op
enin
g(F
1,
F2);
del
ayed
test
icu
lar
dec
ent
(F3
);k
ano
gen
ital
dis
tan
cein
fem
ales
(F1,
F2);k
ano
gen
ital
dis
tan
cem
ales
(F1);k
fem
ale
pu
pw
eig
ht
du
rin
gla
ctat
ion
per
iod
(F1,
F3,
F4);k
mal
ep
up
wei
gh
td
uri
ng
lact
atio
np
erio
d(F
2,
F4);k
infe
edco
nsu
mp
tio
nin
adu
ltfe
mal
es(F
0,
F1,
and
F4)
100
pp
m50
0p
pm
439NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le8
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity�
kli
tter
size
(F1,
F3)
–S
ign
ific
ant
neg
ativ
eli
nea
rtr
end
kan
og
enit
ald
ista
nce
infe
mal
es(F
3)
–10
0a
km
ale
pu
pw
eig
ht
du
rin
gla
ctat
ion
per
iod
(F1)
?5
pp
mN
oef
fect
:o
nm
atin
g,
fert
ilit
y,o
rp
reg
nan
cyin
dic
esin
any
gen
erat
ion
;o
nd
ura
tio
n;
on
of
ges
tati
on
;o
nre
sorp
tio
ns
site
sin
anim
als
that
did
no
tb
eco
me
pre
gn
ant;
on
ov
aria
nfo
llic
leco
un
to
rsp
erm
par
amet
ers
500
pp
m(h
igh
do
se;
no
effe
ct)
–
CD
sS
DIG
Sra
t;0,
20,2
00,o
r10
00p
pm
gen
iste
inin
die
t(m
ean
:1
.7,
18
,a
nd
90
mg
/kg
bw
/day
)fr
om
GD
15to
PN
D10
kb
od
yw
eig
ht
inm
ales
at11
wee
ks
of
age
–1.
7M
asu
tom
iet
al.
(200
3)
Lim
ited
(ora
l)m
rela
tiv
eb
rain
wei
gh
t;m
rela
tiv
ep
itu
itar
yw
eig
ht
–1.
7a
mre
lati
ve
adre
nal
wei
gh
t1.
718
kab
solu
tep
itu
itar
yw
eig
ht;k
bo
dy
wei
gh
tg
ain
inm
ales
on
PN
D21
–42
1890
Wis
tar
(RO
RO
)ra
ts;
0,20
,15
0,o
r10
00m
g/
kg
bw
/d
ayg
enis
tein
fro
mG
D6–
20b
yg
avag
em
pu
pm
ort
alit
y–
20a
McC
lain
etal
.(2
007)
Lim
ited
(ora
l)V
isce
ral
mal
form
atio
ns
(art
ery
ori
gin
var
ian
t)15
010
00(o
nli
tter
inci
den
ceb
asis
)N
oef
fect
on
nu
mb
ero
fd
eliv
ered
pu
ps,
imp
lan
tati
on
or
reso
rpti
on
site
s,o
rex
tern
alm
alfo
rmat
ion
1000
(hig
hd
ose
;n
oef
fect
)–
Wis
tar
(RO
RO
)ra
ts;
0,5,
50,
100,
500
mg
/k
gb
w/
day
gen
iste
infr
om
GD
5–21
inth
ed
iet
mp
ost
-im
pla
nta
tio
nlo
ss,k
nu
mb
ero
ffe
tuse
sp
erfe
mal
e,k
feta
lb
od
yw
eig
ht
(per
litt
erb
asis
),in
cid
ence
skel
etal
oss
ific
atio
n
100
500
Ald
erle
yP
ark
rat;
0,0.
2,o
r2
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
nd
uri
ng
PN
D1–
6an
d4
and
40m
g/
kg
bw
/d
ayb
yg
avag
eo
nP
ND
7–21
(sc
do
ses
wer
ed
eter
min
edto
be
equ
ival
ent
tog
avag
ed
ose
so
f4
and
20m
g/
kg
bw
/d
ay);
on
ep
art
of
the
stu
dy
exam
inin
gS
DN
-PO
Ad
ose
dan
imal
sd
uri
ng
the
sam
ep
erio
dw
ith
scan
do
ral
do
ses
equ
ival
ent
to4
and
40m
g/
kg
bw
/d
ayb
yo
ral
exp
osu
re
Ad
van
ced
vag
inal
op
enin
g,
per
sist
ent
vag
inal
corn
ific
atio
n,
ute
rotr
op
hic
resp
on
se,k
seru
mp
rog
este
ron
ein
fem
ales
,kb
od
yw
eig
ht
infe
mal
es
440
ora
lP
ND
7–21
;20
PN
D1–
6(e
qu
ival
ent
toan
scin
ject
ion
do
seo
f2
mg
/k
gb
w/
day
)
Lew
iset
al.
(200
3)
Nag
aoet
al.
(200
1)
Hig
h(o
ral)
Hig
h(o
ral)
mS
DN
PO
Av
olu
me
infe
mal
es4
40
Sp
rag
ue-
Daw
ley
rat;
0,12
.5,
25,
50,
or
100
mg
/k
gb
w/
day
gen
iste
inb
yg
avag
eo
nP
ND
1–5.
kfe
rtil
ity
ind
ex(g
enis
tein
-tre
ated
fem
ales
mat
edw
ith
un
trea
ted
mal
es);
po
lyo
vu
lar
foll
icle
sin
wea
nli
ng
fem
ales
;k
epid
idy
mal
wei
gh
t;k
bo
dy
wei
gh
tso
fm
ales
at18
wee
ks
of
age;
(kb
od
yw
eig
hts
of
fem
ales
at9
wee
ks
of
age)
–12
.5
kn
orm
ales
tro
us
cycl
e(n
ot
do
sed
epen
den
t)–
12.5
a
kn
um
ber
of
imp
lan
tsp
erli
tter
5010
0k
bo
dy
wei
gh
tso
fb
oth
sex
esat
1,2,
3,5,
7,9
and
18(m
ales
on
ly)
wee
ks
of
age
(kb
od
yw
eig
hts
of
bo
thse
xes
at50
mg
/k
gb
w/
day
fro
m5,
7,9,
and
18(m
ales
on
ly)
wee
ks
of
age)
–10
0a
440 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Hy
per
tro
ph
yo
fu
teri
ne
my
om
etri
um
–50
(rep
ort
edin
anim
als
atlo
wer
do
sele
vel
s,b
ut
no
tst
atis
tica
lly
anal
yze
d)
mat
reti
cfo
llic
les,k
corp
us
lute
um
18w
eek
s,h
yp
ertr
op
hy
of
lum
inal
and
gla
nd
ula
rep
ith
elia
lce
lls,
andk
inn
um
ber
of
ute
rin
eg
lan
ds
–10
0(r
epo
rted
inan
imal
sat
low
erd
ose
lev
els,
bu
tn
ot
stat
isti
call
yan
aly
zed
)N
oef
fect
:M
ales
-n
oef
fect
inti
me
top
rep
uti
alse
par
atio
n,
cop
ula
tio
n,
or
fert
ilit
y;
inn
um
ber
of
imp
lan
tso
rn
um
ber
of
reso
rpti
on
sin
sire
dp
reg
nan
cies
;se
rum
test
ost
ero
ne;
epid
idy
mal
sper
mco
nce
ntr
atio
n;
or
test
icu
lar
his
tolo
gic
chan
ges
Fem
ales
-n
oef
fect
inag
eat
vag
inal
op
enin
g
100
(hig
hd
ose
;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rat;
0o
rd
iet
con
tain
ing
0.5
g/
kg
gen
iste
in(5
00
mg
/kg
die
t;a
uth
ors
est
ima
ted
inta
kes
of
10
mg
/da
yd
uri
ng
pre
gn
ancy
an
d1
8.5
mg
/da
yd
uri
ng
lact
ati
on
;th
ise
qu
als
B2
6–
47
mg
/kg
bw
/da
yb
ase
do
nth
ere
po
rte
db
od
yw
eig
ht
ofB
38
0g
ind
am
s)
No
effe
ct:
on
pre
gn
ancy
ou
tco
me;
pu
pb
od
yw
eig
ht
du
rin
gp
reg
nan
cy,
lact
atio
n,
or
afte
rw
ean
ing
;o
rgan
wei
gh
tsin
off
spri
ng
(ute
rus,
ov
ary,
test
es,
epid
idy
mid
esat
PN
D15
,49
and
75)
B26
–47
(hig
hd
ose
;n
oef
fect
)
–T
ou
sen
etal
.(2
006)
Lim
ited
(ora
l)
Sp
rag
ue-
Daw
ley
dam
sw
ere
fed
die
tco
nta
inin
g0
or
5p
pm
gen
iste
infr
om
GD
17th
rou
gh
ou
tth
ela
ctat
ion
per
iod
up
toP
ND
70in
off
spri
ng
.(E
xp
osu
rein
off
spri
ng
est
imate
datB
0.6
8m
g/
kg
bw
/day
ov
er
the
life
tim
e.)
Ch
ang
esin
ov
aria
nh
isto
log
yat
PN
D21
and
70;k
bo
dy
wei
gh
tin
PN
D70
fem
ales
(co
nti
nu
ou
sex
po
sure
toP
ND
70);m
bo
dy
wei
gh
tin
PN
D70
fem
ales
(ex
po
sure
pri
or
toP
ND
21)
–B
0.68
b(s
ing
led
ose
)A
wo
niy
iet
al.
(199
8)
Lim
ited
(ora
l)
No
effe
ctin
fem
ales
on
PN
D70
on
seru
mL
H,
FS
H,
17b-
estr
adio
l,p
rog
este
ron
e,o
var
ian
or
ute
rin
ew
eig
ht
B0.
68b
(sin
gle
do
se;
no
effe
ct)
–
Pre
gn
ant
Sp
rag
ue-
Daw
ley
rats
wer
efe
dd
iets
con
tain
ing
0,20
,o
r10
0p
pm
gen
iste
in(0
,2
0,
or
87
mg
/kg
bw
/day
)fr
om
con
cep
tio
nth
rou
gh
po
stn
atal
life
(GD
1to
PN
D56
).
man
og
enit
ald
ista
nce
infe
mal
es;k
age
and
wei
gh
tat
vag
inal
op
enin
g;m
ute
rus
wei
gh
to
nP
ND
21;m
rela
tiv
ete
stis
wei
gh
to
nP
ND
21;m
rela
tiv
ete
stis
wei
gh
to
nP
ND
56;k
ven
tral
pro
stat
ew
eig
ht
2087
Cas
ano
va
etal
.(1
999)
Lim
ited
(ora
l)
No
effe
ct:
on
imp
lan
tati
on
site
sp
erd
am;x
of
liv
ep
up
sp
erli
tter
;o
rli
tter
wei
gh
tat
bir
th;
pro
po
rtio
no
fm
ales
wit
hre
tain
edn
ipp
les;
age
or
wei
gh
tat
pre
pu
tial
sep
arat
ion
;o
rw
eig
ht
(ab
solu
teo
rre
lati
ve)
of
the
test
is(P
ND
21o
r56
)o
rv
entr
alp
rost
ate
(PN
D56
)as
litt
erm
ean
s
87(h
igh
do
se;n
oef
fect
)–
Lo
ng
Ev
ans
rat;
0o
r15
mg
/k
gb
w/
day
gen
iste
inb
yg
avag
eo
nG
D14
toP
ND
21.
mu
teri
ne
pro
ges
tero
ne
rece
pto
rex
pre
ssio
nin
gla
nd
ula
rep
ith
elia
lce
lls
–15
(sin
gle
do
sest
ud
y)
Hu
gh
eset
al.
(200
4)L
imit
ed(o
ral)
No
effe
ct:
ute
rin
elu
min
alep
ith
elia
lce
llh
eig
ht,
ute
rin
ep
roli
fera
tio
n,
ERa
exp
ress
ion
inu
teri
ne
lum
inal
or
gla
nd
ula
rep
ith
elia
lce
lls,
or
pro
ges
tero
ne
rece
pto
rex
pre
ssio
nin
lum
inal
epit
hel
ial
cell
s
15(h
igh
do
se;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rat;
at0,
25,
250,
or
1250
pp
mg
enis
tein
inth
ed
iet
fro
mG
D7
tow
ean
ing
via
dam
and
dir
ectl
yto
off
spri
ng
afte
rw
ean
ing
thro
ug
hP
ND
50(B
2,
20
,2
00
mg
/kg
bw
/day
)
Hep
atic
CY
P3A
enzy
me
exp
ress
ion
inm
ales
;al
tere
dfo
rmat
ion
of
test
ost
ero
ne
met
abo
lite
sin
liv
erm
icro
som
es,
i.e.
,D
HT
/3-
dio
lan
d7a
-OH
B2
ZB
20a
Lau
ren
zan
aet
al.
(200
2)
Lim
ited
(ora
l)
441NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011
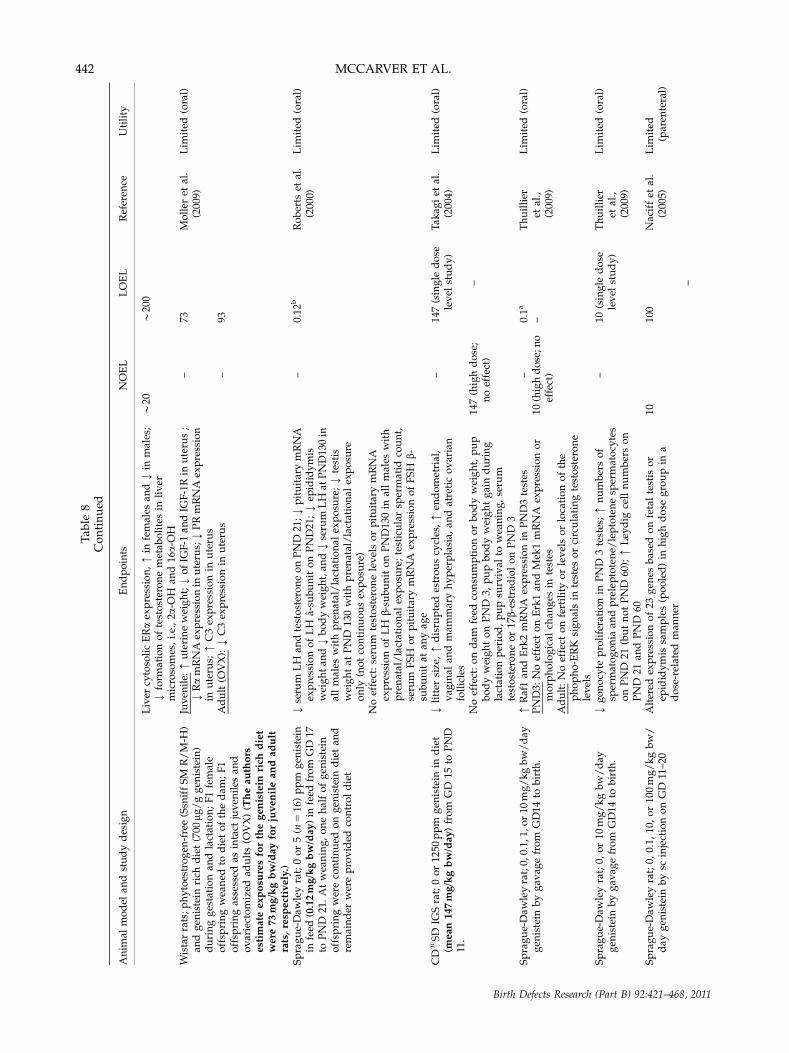
Tab
le8
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity�
Liv
ercy
toso
lic
ERa
exp
ress
ion
,m
infe
mal
esan
dk
inm
ales
;k
form
atio
no
fte
sto
ster
on
em
etab
oli
tes
inli
ver
mic
roso
mes
,i.
e.,
2a-O
Han
d16a-
OH
B20
B20
0
Wis
tar
rats
;p
hy
toes
tro
gen
-fre
e(S
snif
fS
MR
/M
-H)
and
gen
iste
inri
chd
iet
(700
mg/
gg
enis
tein
)d
uri
ng
ges
tati
on
and
lact
atio
n;
F1
fem
ale
off
spri
ng
wea
ned
tod
iet
of
the
dam
;F
1o
ffsp
rin
gas
sess
edas
inta
ctju
ven
iles
and
ov
arie
cto
miz
edad
ult
s(O
VX
)(T
he
au
tho
rse
stim
ate
ex
po
sure
sfo
rth
eg
en
iste
inri
chd
iet
we
re7
3m
g/k
gb
w/d
ay
for
juv
enil
ea
nd
ad
ult
rats
,re
spec
tiv
ely
.)
Juv
enil
e:m
ute
rin
ew
eig
ht;k
of
IGF
-1an
dIG
F-1
Rin
ute
rus
;k
Ra
mR
NA
exp
ress
ion
inu
teru
s;k
PR
mR
NA
exp
ress
ion
inu
teru
s;m
C3
exp
ress
ion
inu
teru
s
–73
Mo
ller
etal
.(2
009)
Lim
ited
(ora
l)
Ad
ult
(OV
X):k
C3
exp
ress
ion
inu
teru
s–
93
Sp
rag
ue-
Daw
ley
rat;
0o
r5
(n5
16)
pp
mg
enis
tein
infe
ed(0
.12
mg
/kg
bw
/day
)in
feed
fro
mG
D17
toP
ND
21.
At
wea
nin
g,
on
eh
alf
of
gen
iste
ino
ffsp
rin
gw
ere
con
tin
ued
on
gen
iste
ind
iet
and
rem
ain
der
wer
ep
rov
ided
con
tro
ld
iet
kse
rum
LH
and
test
ost
ero
ne
on
PN
D21
;k
pit
uit
ary
mR
NA
exp
ress
ion
of
LH
a-su
bu
nit
on
PN
D21
;k
epid
idy
mis
wei
gh
tan
dk
bo
dy
wei
gh
t,an
dk
seru
mL
Hat
PN
D13
0in
all
mal
esw
ith
pre
nat
al/
lact
atio
nal
exp
osu
re;k
test
isw
eig
ht
atP
ND
130
wit
hp
ren
atal
/la
ctat
ion
alex
po
sure
on
ly(n
ot
con
tin
uo
us
exp
osu
re)
–0.
12b
Ro
ber
tset
al.
(200
0)L
imit
ed(o
ral)
No
effe
ct:
seru
mte
sto
ster
on
ele
vel
so
rp
itu
itar
ym
RN
Aex
pre
ssio
no
fL
Hb-
sub
un
ito
nP
ND
130
inal
lm
ales
wit
hp
ren
atal
/la
ctat
ion
alex
po
sure
;te
stic
ula
rsp
erm
atid
cou
nt,
seru
mF
SH
or
pit
uit
ary
mR
NA
exp
ress
ion
of
FS
Hb-
sub
un
itat
any
age
CD
sS
DIG
Sra
t;0
or
1250
pp
mg
enis
tein
ind
iet
(me
an1
47
mg
/kg
bw
/day
)fr
om
GD
15to
PN
D11
.
kli
tter
size
,m
dis
rup
ted
estr
ou
scy
cles
,m
end
om
etri
al,
vag
inal
and
mam
mar
yh
yp
erp
lasi
a,an
dat
reti
co
var
ian
foll
icle
s
–14
7(s
ing
led
ose
lev
elst
ud
y)
Tak
agi
etal
.(2
004)
Lim
ited
(ora
l)
No
effe
ct:
on
dam
feed
con
sum
pti
on
or
bo
dy
wei
gh
t,p
up
bo
dy
wei
gh
to
nP
ND
3,p
up
bo
dy
wei
gh
tg
ain
du
rin
gla
ctat
ion
per
iod
,p
up
surv
ival
tow
ean
ing
,se
rum
test
ost
ero
ne
or
17b-
estr
adio
lo
nP
ND
3
147
(hig
hd
ose
;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rat;
0,0.
1,1,
or
10m
g/
kg
bw
/d
ayg
enis
tein
by
gav
age
fro
mG
D14
tob
irth
.m
Raf
1an
dE
rk2
mR
NA
exp
ress
ion
inP
ND
3te
stes
–0.
1aT
hu
illi
eret
al.,
(200
9)
Lim
ited
(ora
l)P
ND
3:N
oef
fect
on
Erk
1an
dM
ek1
mR
NA
exp
ress
ion
or
mo
rph
olo
gic
alch
ang
esin
test
es10
(hig
hd
ose
;no
effe
ct)
–
Ad
ult
:N
oef
fect
on
fert
ilit
yo
rle
vel
so
rlo
cati
on
of
the
ph
op
ho
-ER
Ksi
gn
als
inte
stes
or
circ
ula
tin
gte
sto
ster
on
ele
vel
sS
pra
gu
e-D
awle
yra
t;0,
or
10m
g/
kg
bw
/d
ayg
enis
tein
by
gav
age
fro
mG
D14
tob
irth
.k
go
no
cyte
pro
life
rati
on
inP
ND
3te
stes
;m
nu
mb
ers
of
sper
mat
og
on
iaan
dp
rele
pto
ten
e/le
pto
ten
esp
erm
ato
cyte
so
nP
ND
21(b
ut
no
tP
ND
60);m
Ley
dig
cell
nu
mb
ers
on
PN
D21
and
PN
D60
–10
(sin
gle
do
sele
vel
stu
dy
)T
hu
illi
eret
al.,
(200
9)
Lim
ited
(ora
l)
Sp
rag
ue-
Daw
ley
rat;
0,0.
1,10
,o
r10
0m
g/
kg
bw
/d
ayg
enis
tein
by
scin
ject
ion
on
GD
11–2
0A
lter
edex
pre
ssio
no
f23
gen
esb
ased
on
feta
lte
stis
or
epid
idy
mis
sam
ple
s(p
oo
led
)in
hig
hd
ose
gro
up
ina
do
se-r
elat
edm
ann
er
1010
0N
acif
fet
al.
(200
5)L
imit
ed(p
aren
tera
l)
–
442 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

No
effe
ct:
mat
ern
alb
od
yw
eig
ht;
nu
mb
erli
ve
fetu
ses/
litt
er;
gro
ssh
isto
pat
ho
log
ical
effe
cts
on
feta
lte
stis
or
epid
idy
mis
100
(hig
hd
ose
;n
oef
fect
)
Lo
bu
nd
-Wis
tar
rat;
0o
r25
0m
gg
enis
tein
/k
gA
IN-
76d
iet.
Fo
ur
gro
up
so
fg
enis
tein
-ex
po
sed
mal
es:
con
tro
l(G
rou
pA
),b
irth
–P
ND
35(G
rou
pB
),P
ND
90—
11m
on
ths
(Gro
up
C);
bir
thth
rou
gh
ou
tli
fe(G
rou
pD
)
Res
po
nse
toN
MU
inch
emic
ally
-cas
trat
edra
tsim
pla
nte
dw
ith
asi
last
icca
psu
leco
nta
inin
gte
sto
ster
on
e:k
po
orl
yd
iffe
ren
tiat
edp
rost
atic
tum
ors
inG
rou
ps
C&
D(r
egu
lati
on
of
AR
/A
kt/
PT
EN
axis
sug
ges
ted
asm
ole
cula
rm
ech
anis
ms)
–25
0m
gg
enis
tein
/k
gA
IN-7
6d
iet
(sin
gle
do
sele
vel
)
Wan
get
al.
(200
9)L
imit
ed(o
ral)
N-m
eth
yln
itro
sou
rea
(NM
U)
use
dto
ind
uce
pro
stat
eca
nce
rat
PN
D70
Sp
rag
ue-
Daw
ley
rat;
0,30
0,o
r80
0p
pm
gen
iste
inin
die
td
uri
ng
pre
gn
ancy
and
lact
atio
nan
du
pto
PN
D90
ino
ffsp
rin
g;
(me
an
ex
po
sure
s:2
5a
nd
53
mg
/kg
bw
/day
ind
am
sa
nd
30
an
d8
4m
g/k
gb
w/d
ay
inp
up
s.)
kb
irth
wei
gh
to
ffe
mal
eo
ffsp
rin
g;
acce
lera
ted
vag
inal
op
enin
g–
25Y
ou
etal
.(2
002a
)L
imit
ed(o
ral)
kb
od
yw
eig
ht
atP
ND
21an
dP
ND
110
inm
ales
and
fem
ales
;m
pit
uit
ary
wei
gh
tin
mal
es;k
age
atp
rep
uti
alse
par
atio
nw
hen
usi
ng
bo
dy
wei
gh
tas
cov
aria
te;m
mam
mar
yg
lan
db
ran
chin
gan
dP
CN
Aim
mu
no
stai
nin
go
fm
amm
ary
epit
hel
ial
cell
sin
mal
es
3084
No
effe
ct:
liv
er,
bra
in,
ven
tral
pro
stat
e,te
stis
,o
ru
teri
ne
wei
gh
tsat
PN
D21
or
PN
D11
0;n
oh
isto
log
icef
fect
sin
mal
eo
rfe
mal
eti
ssu
es
84(h
igh
do
se;n
oef
fect
)–
Lo
ng
Ev
ans
rats
;0,
10m
g/
kg
bw
gen
iste
inb
ysc
inje
ctio
nP
ND
0–3
Ad
van
ced
vag
inal
op
enin
g;
alte
red
estr
ou
scy
clin
gb
y10
wee
ks;
kG
nR
Hac
tiv
atio
n;k
AV
PV
KIS
Sd
ensi
ty–
10(s
ing
led
ose
lev
el)
Bat
eman
and
Pat
isau
l(2
009)
Lim
ited
(par
ente
ral)
No
effe
ct:
on
AR
CK
ISS
fib
erd
ensi
ty10
(sin
gle
do
sele
vel
)–
Sp
rag
ue-
Daw
ley
rats
;0,
250mg
gen
iste
inb
ysc
inje
ctio
nev
ery
12h
rb
egin
nin
go
nP
ND
1fo
ra
tota
lo
f4
inje
ctio
ns
(ass
um
ing
aS
pra
gu
e-
Da
wle
yra
tp
up
we
igh
s7
.5g
,e
ach
do
sew
ou
ldb
e3
3.3
mg
/kg
bw
/day
.T
wo
inje
ctio
ns
pe
rd
ay
for
ato
tal
of
66
.7m
g/k
gb
w/d
ayg
en
iste
in)
No
effe
ct:
on
5-H
Tfi
ber
den
sity
;o
nco
lab
elin
go
fV
MN
v1
ERa
and
Hu
C/
D66
.7(s
ing
led
ose
lev
el)
–P
atis
aul
etal
.(2
008)
Lim
ited
(par
ente
ral)
Alp
k:
AP
fSD
rat:
0,0.
2,o
r2
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
nd
uri
ng
PN
D1–
6an
d4
and
40m
g/
kg
bw
/d
ayb
yg
avag
eo
nP
ND
7–21
(sc
do
ses
wer
ed
eter
min
edto
be
equ
ival
ent
tog
avag
ed
ose
so
f4
and
20m
g/
kg
bw
/d
ay);
on
ep
art
of
the
stu
dy
exam
inin
gS
DN
-PO
Ad
ose
dan
imal
sd
uri
ng
the
sam
ep
erio
dw
ith
scan
do
ral
do
ses
equ
ival
ent
to4
and
40m
g/
kg
bw
/d
ayb
yo
ral
exp
osu
re
Ad
van
ced
vag
inal
op
enin
g,k
bo
dy
wei
gh
tin
fem
ales
fro
mP
ND
57to
13w
eek
so
fag
e,p
ersi
sten
tv
agin
alco
rnif
icat
ion
,an
dk
seru
mp
rog
este
ron
e
440
Lew
iset
al.
(200
3)L
imit
ed(p
aren
tera
l)
mu
teri
ne
wei
gh
tat
PN
D22
(fo
llo
win
gsc
inje
ctio
no
nP
DN
1–6
on
ly)
0.2
2
mS
DN
PO
Av
olu
me
infe
mal
es,m
ute
rin
ew
eig
ht
atP
ND
224
40
Sp
rag
ue-
Daw
ley
rat;
0,0.
1,10
,o
r10
0m
g/
kg
bw
/d
ayg
enis
tein
by
scin
ject
ion
on
GD
11to
20A
lter
edex
pre
ssio
no
f22
7g
enes
bas
edin
po
ole
dfe
tal
ov
ary
and
ute
rin
esa
mp
les;
66g
enes
resp
on
ded
insi
mil
arp
atte
rnto
eth
iny
les
trad
iol
and
bis
ph
eno
lA
1010
0N
acif
fet
al.
(200
2)L
imit
ed(p
aren
tera
l)
443NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le8
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity�
No
effe
ct:
on
mat
ern
alb
od
yw
eig
ht,
nu
mb
ero
fli
ve
fetu
ses/
litt
er,
gro
sso
rh
isto
pat
ho
log
icef
fect
so
no
var
yo
ru
teru
s
100
(hig
hd
ose
;n
oef
fect
)–
Wis
tar
rat;
0o
r1
mg
/d
ay[B
19
mg
/kg
]g
enis
tein
by
scin
ject
ion
fro
mP
ND
1to
5E
arly
on
set
of
vag
inal
op
enin
g,
alte
red
estr
ou
scy
clin
g;k
bo
dy
wei
gh
tin
fem
ales
;k
ov
aria
nw
eig
ht;k
nu
mb
ero
fco
rpo
ralu
tea
–B
19(s
ing
led
ose
stu
dy
)K
ou
ki
etal
.(2
003)
Lim
ited
(par
ente
ral)
B19
(hig
hd
ose
;n
oef
fect
)–
Wis
tar
rat;
0o
r1
mg
/d
ay[B
19
mg
/kg
]d
aid
zein
by
scin
ject
ion
fro
mP
ND
1to
5E
arly
on
set
of
vag
inal
op
enin
g,
alte
red
estr
ou
scy
clin
g;k
bo
dy
wei
gh
tin
fem
ales
;k
ov
aria
nw
eig
ht;k
nu
mb
ero
fco
rpo
ralu
tea
–B
19(s
ing
led
ose
stu
dy
)K
ou
ki
etal
.(2
003)
Lim
ited
(par
ente
ral)
CR
CD
rat;
0,5,
or
25m
g/
anim
al/
day
gen
iste
inb
ysc
inje
ctio
nfr
om
GD
16to
20(1
5a
nd
75
mg
/kg
bw
/day
)
kb
irth
wei
gh
to
ffe
mal
es15
75L
evy
etal
.(1
995)
Lim
ited
(par
ente
ral)
kan
og
enit
ald
ista
nce
inm
ales
;d
elay
edv
agin
alo
pen
ing
15a
No
effe
ct:
vo
lum
eo
fS
DN
-PO
Ain
eith
erse
x75
(hig
hd
ose
;no
effe
ct)
0o
r50
0m
g/
kg
bw
gen
iste
inb
ysc
inje
ctio
no
nP
ND
16,
18,
and
20m
ute
rin
ew
eig
ht,m
seru
m17
b-es
trad
iol,
andk
seru
mp
rog
este
ron
eo
nP
ND
21;k
ERa
pro
tein
;k
and
rog
enre
cep
tor
pro
tein
;m
pro
ges
tero
ne
rece
pto
rp
rote
in(b
oth
iso
form
s)
–50
0(s
ing
led
ose
stu
dy
)C
otr
on
eoet
al.
(200
1)
Lim
ited
(par
ente
ral)
Bra
inan
dbe
havi
orC
Ds
(SD
)IG
Sra
t;0,
20,
200,
or
1000
pp
mg
enis
tein
ind
iet
fro
mG
D15
toP
ND
10(m
ean
est
ima
ted
inta
kes
of
1.3
–2
.1,
13
.7–
23
.0,
an
d6
6.6
–11
3.1
mg
/k
gb
w/d
ay)
No
effe
ct:
on
pro
po
rtio
no
fp
itu
itar
yce
lls
stai
nin
gfo
rL
H,
FS
H,
and
pro
lact
in66
.6–1
13.1
(hig
hd
ose
;n
oef
fect
)
–M
asu
tom
iet
al.
(200
4b)
Lim
ited
(ora
l)
Sp
rag
ue-
Daw
ley
rat;
0,5,
100,
and
500
pp
mg
enis
tein
ind
iet
thro
ug
hg
esta
tio
nan
dla
ctat
ion
and
ino
ffsp
rin
gu
nti
lP
ND
140
(0,
0.3
1,5
.7,
34
mg
/kg
bw
/da
y)
mca
lbin
din
-po
siti
ve
cell
sin
SD
N-P
OA
inm
ales
–0.
31S
call
etet
al.
(200
4)L
imit
ed(o
ral)
Sp
rag
ue-
Daw
ley
rat;
0o
r10
00p
pm
gen
iste
inin
die
tfr
om
GD
15to
PN
D11
(B6
6.6
–11
3.1
mg
/kg
bw
/day
)
No
effe
ct:
gen
eex
pre
ssio
no
fE
Ra,
ERb,
pro
ges
tero
ne
rece
pto
r,an
dst
ero
idre
cep
tor
coac
tiv
ato
rin
the
hy
po
thal
amic
pre
op
tic
area
B66
.6–1
13.1
(sin
gle
do
sest
ud
y;
no
effe
ct)
Tak
agi
etal
.(2
005)
Lim
ited
(ora
l)
CR
CD
rat;
0,10
0,o
r10
00mg
gen
iste
ino
nP
ND
1–10
0.1,
or
1,m
g/
day
by
scin
ject
ion
on
PN
D1–
10.
(Me
anE
12
an
d11
7m
g/k
gb
w/d
ay)
No
n-d
ose
-rel
ated
min
LH
secr
etio
n–
12a
Fab
eran
dH
ug
hes
(199
1)
Lim
ited
(par
ente
ral)
mS
DN
-PO
Av
olu
me
infe
mal
es12
117
CR
CD
rat;
0,0.
001,
0.01
,0.
1,0.
200,
0.4,
0.5,
or
1.0
mg
0,1,
10,
100,
200,
400,
500,
or
1000
mgg
enis
tein
by
scin
ject
ion
on
PN
D1–
10.
(Me
anE
0.1
2,
1.2
,1
2,
23
,4
7,
58
,a
nd
117
mg
/kg
bw
/d
ay.
)
mG
nR
H-i
nd
uce
dL
Hse
cret
ion
–0.
12F
aber
and
Hu
gh
es(1
991)
Lim
ited
(par
ente
ral)
mS
DN
-PO
Av
olu
me
4758
444 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Sp
rag
ue-
Daw
ley
rat;
0o
r25
0mg
/p
up
of
gen
iste
inb
ysc
inje
ctio
ntw
ice
ad
ayo
nP
ND
1an
d2
(B1
00
mg
/kg
bw
/day
)
min
nu
mb
ero
fA
VP
Vce
lls
po
siti
ve
for
tyro
sin
eh
yd
rox
yla
sein
mal
es;k
nu
mb
ero
fA
VP
Vce
lls
po
siti
ve
for
bo
thty
rosi
ne
hy
dro
xy
lase
and
ER
ain
fem
ales
;k
%o
fA
VP
Vce
lls
wit
hE
Ra
inb
oth
mal
esan
dfe
mal
es
–B
100
(sin
gle
do
sest
ud
y)
Pat
isau
let
al.
(200
6)L
imit
ed(p
aren
tera
l)
Sp
rag
ue-
Daw
ley
mal
era
t;0
or
250mg
/p
up
of
gen
iste
inb
ysc
inje
ctio
ntw
ice
ad
ayo
nP
ND
1an
d2
(B10
0m
g/
kg
bw
/d
ay)
mca
lbin
din
imm
un
ore
ativ
ece
lls
inC
AL
B-S
DN
inm
ales
;mA
VP
Vv
olu
me
inm
ales
–B
100
(sin
gle
do
sest
ud
y)
Pat
isau
let
al.
(200
7)L
imit
ed(p
aren
tera
l)N
oef
fect
:S
DN
or
CA
LB
-SD
Nv
olu
me;
nu
mb
ero
fG
nR
Hn
euro
ns,
Fo
sex
pre
ssio
nB
100
(hig
hd
ose
;n
oef
fect
)
–
Lo
ng
Ev
ans
mal
era
t;0
or
10m
g/
kg
of
gen
iste
inb
ysc
inje
ctio
nfr
om
bir
th(P
ND
0)to
PN
D3
No
effe
ct:
KIS
Sim
mu
no
reac
tiv
en
euro
ns
inth
eA
VP
Van
dA
RC
inm
ales
10(h
igh
do
se;n
oef
fect
)–
Pat
isau
let
al.
(200
9)L
imit
ed(p
aren
tera
l)L
on
gE
van
sm
ale
rat;
0o
r10
mg
/k
go
feq
uo
lb
ysc
inje
ctio
nfr
om
bir
th(P
ND
0)to
PN
D3
No
effe
ct:
KIS
Sim
mu
no
reac
tiv
en
euro
ns
inth
eA
VP
Van
dA
RC
inm
ales
10(h
igh
do
se;n
oef
fect
)–
Pat
isau
let
al.
(200
9)L
imit
ed(p
aren
tera
l)M
amm
ary
glan
dde
velo
pmen
tan
dca
rcin
ogen
esis
Sp
rag
ue-
Daw
ley
rat,
die
tco
nta
inin
gr
5p
pm
each
of
gen
iste
inan
dd
aid
zein
or
5,10
0,o
r50
0p
pm
gen
iste
in,
mu
ltig
ener
atio
nal
des
ign
(Th
eF
0g
ener
atio
nw
asex
po
sed
fro
msi
xw
eek
so
fag
eu
nti
lte
rmin
atio
n;
the
F1
and
F2
gen
erat
ion
sw
ere
exp
ose
dfr
om
con
cep
tio
nth
rou
gh
term
inat
ion
;th
eF
3g
ener
atio
nw
asex
po
sed
fro
mco
nce
pti
on
thro
ug
hw
ean
ing
on
PN
D21
and
then
pla
ced
on
the
con
tro
ld
iet
un
til
term
inat
ion
;th
eF
4g
ener
atio
nw
asn
ot
dir
ectl
yex
po
sed
toth
ep
rep
ared
die
ts).
Asu
bse
to
fm
ale
wea
nli
ng
F1
and
F3
anim
als
fro
mth
em
ult
igen
erat
ion
stu
die
sw
ere
assi
gn
edto
2-y
ear
stu
die
s,th
ree
exp
osu
red
ura
tio
ns
wer
est
ud
ied
:co
nti
nu
ou
sex
po
sure
fro
mco
nce
pti
on
thro
ug
h2
yea
rs(F
1C–
F1
gen
erat
ion
),ex
po
sure
fro
mco
nce
pti
on
thro
ug
hP
ND
140
foll
ow
edb
yco
ntr
ol
die
tu
nti
lte
rmin
atio
n(F
1T14
0—F
1g
ener
atio
n),
exp
osu
refr
om
con
cep
tio
nth
rou
gh
wea
nin
gat
PN
D21
foll
ow
edb
yco
ntr
ol
die
tu
nti
lte
rmin
atio
n(F
3T21
—F
3g
ener
atio
n).
All
surv
ivin
gan
imal
sw
ere
kil
led
afte
r2
yea
rsan
dco
mp
lete
nec
rop
sies
wer
ep
erfo
rmed
Mam
mar
yg
lan
dh
yp
erp
lasi
ain
mal
esin
stu
dy
term
inat
edo
nP
ND
140
(F1,
F2)
5p
pm
100
pp
mL
aten
dre
sse
etal
.(2
009)
Lim
ited
(ora
l)
Mam
mar
yg
lan
dh
yp
erp
lasi
ain
mal
esin
2-y
ear
stu
dy
(F1T
140)
100
pp
m50
0p
pm
No
effe
ct:
mam
mar
yg
lan
dfi
bro
ma,
fib
road
eno
ma,
neo
pla
sia,
or
pat
tern
of
fem
iniz
atio
nin
mal
era
ts;
du
ctal
hy
per
pla
sia
atan
yd
ose
500
pp
m(h
igh
do
se;
no
effe
ct)
–
Sp
rag
ue-
Daw
ley
rat;
0,30
0,o
r80
0p
pm
gen
iste
inin
die
td
uri
ng
ges
tati
on
and
lact
atio
nm
IGF
-1re
cep
tor
300
pp
m80
0p
pm
Yo
uet
al.
(200
2b)
Lim
ited
(ora
l)m
bra
nch
es,
term
inal
end
bu
ds,
and
late
ral
bu
ds
inm
ale
off
spri
ng
300
pp
m(p
50.
06)
800
pp
m
No
effe
ct:
seru
mp
rola
ctin
800
pp
m(h
igh
do
se;
no
effe
ct)
–
445NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le8
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity�
Sp
rag
ue-
Daw
ley
rat;
0,25
,o
r25
0p
pm
gen
iste
inin
die
t(B
0,
2.2
,a
nd
22
mg
/kg
bw
/day
)d
uri
ng
pre
gn
ancy
and
lact
atio
n.
At
PN
D50
,al
lfe
mal
esw
ere
adm
inis
tere
dd
imet
hy
lben
zan
thra
cen
e(D
MB
A)
toin
du
cem
amm
ary
gla
nd
tum
ors
kD
MB
A-i
nd
uce
dtu
mo
rs;k
typ
e1
lob
ule
s–
2.2
Fri
tzet
al.
(199
8)L
imit
ed(o
ral)
kte
rmin
alen
db
ud
s2.
222
No
adv
erse
effe
cts
on
rep
rod
uct
ive
dev
elo
pm
ent
inm
ales
or
fem
ales
;n
um
ber
of
typ
eII
lob
ule
s;D
NA
lab
elin
go
fm
amm
ary
end
bu
ds
or
term
inal
du
cts
22(h
igh
do
se;n
oef
fect
)
Sp
rag
ue-
Daw
ley
rat;
0,15
,15
0,o
r30
0m
g(a
gly
con
eeq
uiv
alen
t)/
kg
gen
iste
inin
die
td
uri
ng
pre
gn
ancy
and
lact
atio
nv
iaa
cro
ss-
fost
ered
dam
.(e
stim
ate
dm
ate
rna
lin
tak
ea
t1
–1
.5,
10
–1
5,
an
d2
0–
30m
g/k
gb
w/d
ay
).A
tP
ND
47,
ap
ort
ion
of
fem
ales
wer
ead
min
iste
red
dim
eth
ylb
enza
nth
race
ne
(DM
BA
)to
ind
uce
mam
mar
yg
lan
dtu
mo
rs
kp
rop
ort
ion
surv
ivin
gto
17w
eek
sin
F1
off
spri
ng
foll
ow
ing
DM
BA
trea
tmen
to
nP
ND
471–
1.5
10–1
5H
ilak
ivi-
Cla
rke
etal
.(2
002)
Lim
ited
(ora
l)
kse
rum
17b-
estr
adio
lat
8w
eek
s;k
nu
mb
ers
of
lob
ule
s;m
TE
Bs;
mm
amm
ary
tum
ors
inF
1o
ffsp
rin
gfo
llo
win
gD
MB
Atr
eatm
ent
on
PN
D47
10–1
520
–30
No
effe
ct:
litt
ersi
ze;
po
stn
atal
pu
pw
eig
ht
gai
n;
tum
or
late
ncy
or
mu
ltip
lici
tyin
F1
off
spri
ng
foll
ow
ing
DM
BA
trea
tmen
to
nP
ND
47
20–3
0(h
igh
do
se;
no
effe
ct)
–
Sp
rag
ue-
Daw
ley
rat;
0,30
0,80
0p
pm
gen
iste
inin
die
tfr
om
GD
0th
rou
gh
ges
tati
on
and
lact
atio
n,
off
spri
ng
on
sam
ed
iet
asd
ams
afte
rw
ean
ing
(PN
D22
)u
nti
lP
ND
90
Mo
der
ate
ing
lan
du
lar
size
and
tiss
ue
den
sity
of
mam
mar
yg
lan
ds
inm
ales
–30
0p
pm
Wan
get
al.
(200
6)L
imit
ed(o
ral)
kse
rum
IGF
-I(f
emal
e)30
0p
pm
800
pp
mN
oef
fect
:es
trad
iol,
test
ost
ero
ne,
GH
,F
SH
,L
H,
or
pro
lact
inin
fem
ales
800
pp
m(h
igh
do
se;
no
effe
ct)
–
Sp
rag
ue-
Daw
ley
rat;
0o
r10
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
no
nP
ND
2–8
Mam
mar
yg
lan
ds
sho
wed
evid
ence
of
lact
atio
nw
ith
cyst
icd
uct
ald
ilat
atio
n,
aty
pic
alep
ith
elia
lh
yp
erp
lasi
a,an
dm
icro
calc
ific
atio
ns;
insi
tud
uct
alca
rcin
om
aw
asid
enti
fied
in2/
5an
imal
sex
amin
ed
–10
(sin
gle
do
sest
ud
y)
Fo
ster
etal
.(2
004)
Lim
ited
(par
ente
ral)
Sp
rag
ue-
Daw
ley
rats
;0
orB
0.1
mg
/k
gb
w/
day
gen
iste
inb
yin
ject
ion
on
GD
15–2
0(‘
‘Ex
per
imen
t1’
’);
eval
uat
edfe
mal
eo
ffsp
rin
g
min
cid
ence
of
mam
mar
ytu
mo
rsin
F1
off
spri
ng
foll
ow
ing
DM
BA
trea
tmen
to
nP
ND
45;m
mam
mar
yE
Rp
rote
inco
nte
nt
on
PN
D35
;k
pro
tein
kin
ase
Cac
tiv
ity
on
PN
D45
(bu
tn
ot
PN
D21
)
–B
0.1
(sin
gle
do
sele
vel
)H
ilak
ivi-
Cla
rke
etal
.(1
999a
)
Lim
ited
(par
ente
ral)
On
PN
D45
,fe
mal
esw
ere
adm
inis
tere
dd
imet
hy
lben
zan
thra
cen
e(D
MB
A)
toin
du
cem
amm
ary
gla
nd
can
cer
No
effe
ct:
nu
mb
ero
fp
up
s/li
tter
;P
ND
2b
od
yw
eig
ht;
late
ncy
totu
mo
rd
evel
op
men
t,n
um
ber
of
tum
ors
per
anim
al,
or
nu
mb
ero
ftu
mo
rssh
ow
ing
reg
ula
rg
row
th
B0.
1(h
igh
do
se;
no
effe
ct)
–
Sp
rag
ue-
Daw
ley
rats
;0,B
0.5,
orB
1.5
mg
/k
gb
w/
day
gen
iste
inb
ysc
inje
ctio
no
nG
D15
–20
(‘‘E
xp
erim
ent
2’’)
;ev
alu
ated
fem
ale
off
spri
ng
kb
od
yw
eig
ht
atP
ND
35(b
ut
no
tea
rlie
ro
rla
ter
tim
ep
oin
ts)
–B
0.5
Hil
akiv
i-C
lark
eet
al.
(199
9a)
Lim
ited
(par
ente
ral)
On
PN
D50
,fe
mal
esw
ere
adm
inis
tere
dd
imet
hy
lben
zan
thra
cen
e(D
MB
A)
toin
du
cem
amm
ary
gla
nd
can
cer
min
cid
ence
of
mam
mar
ytu
mo
rsin
F1
off
spri
ng
foll
ow
ing
DM
BA
trea
tmen
to
nP
ND
50;m
mam
mar
yE
Rp
rote
inco
nte
nt
on
PN
D50
B0.
5B
1.5
446 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Lat
ency
totu
mo
rd
evel
op
men
t,n
um
ber
of
tum
ors
per
anim
al,
or
nu
mb
ero
ftu
mo
rssh
ow
ing
reg
ula
rg
row
thB
1.5
(hig
hd
ose
;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rats
;0
or
20mg
gen
iste
inb
ysc
inje
ctio
no
nP
ND
7,10
,14
,17
,20
(do
sele
vel
ran
ge
ofB
2m
g/
kg
bw
on
PN
D7
toB
0.7
mg
/k
gb
wo
nP
ND
7);
fem
ale
off
spri
ng
eval
uat
ed
ktu
mo
rm
ult
ipli
city
;k
per
cen
tag
eo
fp
roli
fera
tin
gtu
mo
rsin
F1
off
spri
ng
foll
ow
ing
DM
BA
trea
tmen
to
nP
ND
45;m
lob
ula
rd
iffe
ren
tiat
ion
,k
term
inal
end
du
ctd
ensi
ty,m
alv
eola
rb
ud
den
sity
–B
0.7–B
2(s
ing
led
ose
lev
el)
Hil
akiv
i-C
lark
eet
al.
(199
9b)
Lim
ited
(par
ente
ral)
On
PN
D45
,fe
mal
esw
ere
adm
inis
tere
dd
imet
hy
lben
zan
thra
cen
e(D
MB
A)
toin
du
cem
amm
ary
gla
nd
can
cer
No
effe
ct:
bo
dy
wei
gh
tg
ain
;E
Rp
rote
inle
vel
sin
mam
mar
yg
lan
d;
inci
den
ceo
fm
amm
ary
tum
ors
,tu
mo
rla
ten
cyin
F1
off
spri
ng
foll
ow
ing
DM
BA
trea
tmen
to
nP
ND
45
B0.
7–B
2(h
igh
do
se;
no
effe
ct)
–
Oth
er/m
echa
nis
tic
Sp
rag
ue-
Daw
ley
rat;
0,5,
100,
or
500mg
/g
gen
iste
inin
feed
du
rin
gg
esta
tio
nan
dla
ctat
ion
(B0
.75,
15
,a
nd
75
mg
/kg
bw
/da
yfo
rw
ea
nli
ng
sa
ndB
0.5
,1
0,
an
d5
0m
g/k
gb
w/d
ay
ina
du
ltfe
ma
les)
kth
yro
idp
ero
xid
ase
acti
vit
yin
bo
thse
xes
–B
0.5–
0.75
Ch
ang
and
Do
erg
e(2
000)
Lim
ited
(ora
l)N
oef
fect
:se
rum
lev
els
of
T3,
T4,
and
TS
HB
50–7
5(h
igh
do
se;
no
effe
ct)
–
Sp
rag
ue-
Daw
ley
rat;
0,5,
100,
or
500
pp
mg
enis
tein
ind
iet
(B0
.0,
0.4
,8
an
d4
0m
gg
en
iste
in/k
g/
da
y),
mu
ltig
ener
atio
nal
stu
dy
des
ign
(F0–
F4
wit
hF
4n
ever
exp
ose
d)
F1
and
F2:
kb
od
yw
eig
ht
of
bo
thse
xes
0.4
8F
erg
uso
net
al.
(200
9)
Lim
ited
(ora
l)A
lter
atio
ns
inse
xu
ald
imo
rph
ism
of
inta
ke
of
3%so
diu
mso
luti
on
40(h
igh
do
se;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rat;
0,25
,25
0,o
r12
50p
pm
gen
iste
in(0
,2,
20,
and
100
mg
/k
gb
w/
day
)in
die
tfr
om
GD
7,d
uri
ng
ges
tati
on
and
lact
atio
n,
un
til
PN
D77
ino
ffsp
rin
g
msa
lin
ein
ges
tio
nin
bo
thm
ales
and
fem
ales
2010
0F
lyn
net
al.
(200
0)L
imit
ed(o
ral)
kD
amb
od
yw
eig
ht;k
feed
inta
ke;
ko
ffsp
rin
gw
eig
ht
PN
D42
-PN
D77
250
1250
No
effe
ct:
on
ges
tati
on
ald
ura
tio
n,
tota
lp
up
s/li
tter
;li
ve
pu
ps/
litt
er;
or
sex
rati
o12
50(h
igh
do
se;
no
effe
ct)
Sp
rag
ue-
Daw
ley
rat;
0,30
0,o
r80
0p
pm
gen
iste
inin
die
t(B
26
an
dB
69
mg
/kg
bw
/day
)d
uri
ng
ges
tati
on
and
lact
atio
n.
Alt
ered
sple
enn
atu
ral
kil
ler
cell
acti
vit
yin
mal
ean
dfe
mal
eo
ffsp
rin
g;k
nu
mb
eran
dp
erce
nta
ge
of
CD
41C
D8
thy
mo
cyte
sin
mal
eo
ffsp
rin
g;k
per
cen
tag
eo
fC
D41
CD
8�
thy
mo
cyte
sin
fem
ale
off
spri
ng
a
–26
Gu
oet
al.
(200
2)L
imit
ed(o
ral)
kb
od
yw
eig
ht
inm
ale
and
fem
ale
off
spri
ng
;k
nu
mb
eran
dp
erce
nta
ge
of
CD
4�C
D8�
thy
mo
cyte
san
dp
erce
nta
ge
of
CD
41C
D81
thy
mo
cyte
sin
fem
ale
off
spri
ng
2669
No
effe
ct:
sple
eno
rth
ym
us
size
inm
ale
or
fem
ale
off
spri
ng
69(h
igh
do
se;n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rat;
0,25
,25
0,o
r12
50p
pm
die
tary
gen
iste
in(B
2,
20
,a
nd
10
0m
g/k
gb
w/
da
y)
fro
mG
D7
thro
ug
hla
ctat
ion
kb
on
em
arro
wD
NA
syn
thes
is;m
colo
ny
form
ing
un
its
incu
bat
edw
ith
gra
nu
locy
tem
acro
ph
age
colo
ny
stim
ula
tin
gfa
cto
rin
bo
ne
mar
row
cell
sin
fem
ales
–2a
Gu
oet
al.
(200
5)L
imit
ed(o
ral)
kco
lon
yfo
rmin
gu
nit
sin
cub
ated
wit
hg
ran
ulo
cyte
mac
rop
hag
eco
lon
yst
imu
lati
ng
fact
or
inb
on
em
arro
wce
lls
inm
ales
220
kco
lon
yfo
rmin
gu
nit
sin
cub
ated
wit
hm
acro
ph
age
colo
ny
stim
ula
tin
gfa
cto
rin
bo
ne
mar
row
cell
sin
mal
es2
20a
kre
cov
ered
bo
ne
mar
row
cell
sin
fem
ales
2010
0N
oef
fect
:b
od
yw
eig
ht
of
mal
era
tes
100
(hig
hd
ose
;n
oef
fect
)–
Lo
ng
Ev
ans
mal
era
t;0,
5,o
r30
0m
gg
enis
tein
/k
gfe
ed(0
.42
an
d2
5m
g/k
gb
w/d
ay)
by
mat
ern
alex
po
sure
du
rin
gp
reg
nan
cyan
dla
ctat
ion
sple
enco
un
to
fto
tal
Tce
lls,k
thy
mu
sco
un
tso
fC
D4�
CD
8�
cell
s,m
thy
mu
sco
un
tso
fC
D41
CD
81ce
lls
–0.
42K
lein
etal
.(2
002)
Lim
ited
(ora
l)
kte
sto
ster
on
ele
vel
sa–
0.42
a
447NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

adversely affect prenatal endpoints such as growth andpossibly survival. The most consistent and sensitiveprenatal endpoint was lower pup birth weight, whichwas reported at Z300 ppm genistein (Z25 mg/kg bw/day in dams during pregnancy) administered in diet(You et al., 2002a); lower pup birth weight was seen inother studies at higher dose levels (Flynn et al., 2000,2008). A lower number of mated dams delivering litterswas reported in one study at 1250 ppm genistein in diet(83 mg/kg bw/day in dams during pregnancy; Delcloset al., 2001). Decreased live litter size was reported in twostudies at Z500 ppm in diet (44 mg/kg bw/day in damsduring pregnancy) (2008; Takagi et al., 2004). In ratsorally treated by gavage with genistein during theneonatal period, a lower pregnancy rate was observedat Z12.5 mg/kg bw/day and lower numbers of implantswere observed at 100 mg/kg bw/day (Nagao et al., 2001).None of the studies assessed structural malformations.
Oral exposure studies examining postnatal develop-ment in rats suggested that genistein exposures canresult in lower and delayed growth. In well-designedmultiple dose-level studies, lower pup weight or weightgain during the lactation period were observed withexposures in diet given to dams from early-to-midgestation through lactation (Delclos et al., 2001; Youet al., 2002a; Delclos et al., 2009). The lowest effect level inthese studies was Z100 ppm genistein (Z11 mg/kg bw/day in dams during lactation) in the NCTR multi-generation study (2008). Lower body weight was alsoshown with gavage dosing of pups with Z100 mg/kgbw/day during the lactation period (Nagao et al., 2001).One multiple-dose level study with gestational andlactation exposure reported trends for developmentaldelay and significant delays in eye and ear opening at1250 ppm genistein (Z138 mg/kg bw/day in damsduring lactation) (Delclos et al., 2001). None of thestudies reported adverse effects on postnatal survival.
Reproductive endpointsMouse: Female mice: There is some evidence that
genistein affects endocrine-mediated endpoints in femalemice. Disrupted estrous cycles were seen in miceexposed neonatally by oral (Jefferson et al., 2009a) or sc(Nikaido et al., 2004; Jefferson et al., 2005) dosing. Earlyvaginal opening was observed following fetal (Nikaidoet al., 2004) or PND15-19 (Nikaido et al., 2005) exposurevia sc injections to dams or pups, respectively. A lowernumber of live pups was observed in mice exposedneonatally by oral (Jefferson et al., 2009a) or sc dosing(Jefferson et al., 2005, 2009b). Following neonatal–prepubertal sc exposures, abnormal uterine and ovarianhistology was observed (Newbold et al., 2001; Nikaidoet al., 2005). Additionally, multiple oocyte ovarianfollicles were observed following neonatal sc exposure(Jefferson et al., 2002, 2006), although the relevance tohumans is unclear.
Male mice: A shorter anogenital distance, adjusted forbody weight, was observed in 21-day-old male offspringof dams exposed to 10 mg/kg bw/day genistein fromGD 12 to PND 20 by gavage (Fielden et al., 2003),suggesting an antiandrogenic effect of treatment. Sinceanogenital distance was not consistently evaluated in theavailable mouse studies, it is not clear whether this is areproducible effect of genistein in mice. Anogenitaldistance was not altered consistently in male ratsexposed to genistein during development (see below).
Tab
le8
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsN
OE
LL
OE
LR
efer
ence
Uti
lity�
msp
leen
cou
nts
of
CD
81ce
lls
0.42
25N
oef
fect
:sp
leen
cou
nts
of
CD
41ce
lls,
thy
mu
sco
un
tso
fC
D41
CD
8�,C
D4�
CD
81ce
lls,
inte
rleu
kin
-4o
rin
terf
ero
n-g
by
cult
ure
dly
mp
ho
cyte
s
25(h
igh
do
se,n
oef
fect
)–
Sp
rag
ue-
Daw
ley
rat;
0,0.
1,1,
or
10m
g/
kg
bw
/d
ayg
enis
tein
by
gav
age
on
GD
14-
PN
D0
mex
pre
ssio
no
fP
DG
Fa-
and
PD
GFb-
rece
pto
rm
RN
Ain
test
eso
fP
ND
3–
0.1
Th
uil
lier
etal
.(2
003)
Lim
ited
(ora
l)
mex
pre
ssio
no
fP
DG
Fa-
rece
pto
rm
RN
Ain
ters
titi
um
and
PD
GFb-
rece
pto
rm
RN
Ain
inte
rsti
tiu
m,
cen
tral
and
per
iph
eral
sem
inif
ero
us
cord
s,an
dg
on
ocy
tes
atP
ND
3
110
m,k
5S
ign
ific
ant
incr
ease
,d
ecre
ase.
aD
idn
ot
dis
pla
ya
do
se-r
esp
on
se.
bT
he
Ex
per
tP
anel
has
lim
ited
con
fid
ence
inth
eac
cura
cyo
fth
ed
ose
det
erm
inat
ion
inth
isst
ud
y.� T
he
term
uti
lity
refe
rsto
the
app
lica
bil
ity
toth
ep
urp
ose
of
dra
win
gco
ncl
usi
on
so
nw
het
her
or
no
ta
chem
ical
adv
erse
lyaf
fect
sre
pro
du
ctio
n.
448 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

A higher incidence of hypospadias in male fetuses wasobserved following oral exposure of dams to 0.17 mg/kgbw/day genistein between GD 13 and 17 (Vilela et al.,2007). No other studies in mice or rats in which maleoffspring were exposed to genistein during late gestationhave reported a higher incidence of hypospadias.Hyperplasia was reported in the prostate and seminalvesicles of adult mice that had been treated sc with500 mg/kg bw/day genistein on PND 1 to 3 (Strausset al., 1998). Conversely, the relative weights of prostatelobes were lower in adult mice from the same studyfollowing sc treatment with Z50 mg/kg bw/day genis-tein on PND 1 to 3 (Strauss et al., 1998). No effects onsperm count or motility were reported and in vitrofertilization was not lower following oral or sc exposuresof dams or developing offspring to genistein (Shibayamaet al., 2001). There were also no consistent effects on testisweights or testicular morphology following oral or scexposure of male mice (Shibayama et al., 2001; Adachiet al., 2004).
Rat: Female rat: Oral exposure studies suggest thatgenistein can affect endocrine-mediated reproductiveendpoints in female rats. Trends or significant effectson accelerated vaginal opening were observed in severalstudies by oral (Casanova et al., 1999; Delclos et al., 2001;Dalu et al., 2002; You et al., 2002a; Lewis et al., 2003,2008;) or sc injection (Kouki et al., 2003) exposure duringthe prenatal or neonatal period. The lowest genisteineffect level for alterations in vaginal opening wasZ300 ppm (Z30 mg/kg bw/day in pups) in the studyof You et al. (2002a). One study reported a higher numberof polyovular follicles in 21-day-old rats following directgavage dosing with Z12.5 mg/kg bw/day genisteinduring the neonatal period (Nagao et al., 2001). Abnor-mal estrous cycles were reported by oral (2008; Takagiet al., 2004) or sc (Kouki et al., 2003) exposure routes.Smaller litter sizes were seen with oral transplacentalexposure (2008; Takagi et al., 2004). Direct ovarian anduterine effects (including morphological changes) wereobserved in several studies with oral or sc exposureduring the prenatal or neonatal period (Awoniyi et al.,1998; Casanova et al., 1999; Cotroneo et al., 2001; Delcloset al., 2001; Nagao et al., 2001; Naciff et al., 2002; Kouki
et al., 2003; Lewis et al., 2003; Takagi et al., 2004; Molleret al., 2009). Effects of genistein on uterine weight arereported in ‘‘Summary of Pharmacokinetics and GeneralToxicology,’’ which describes in vitro and in vivo studiesof estrogenic activity. No other consistent effects onfemale reproductive organ weights were reported.
Male rat: The majority of effects of genistein reportedin the reproductive system of male rats were on theprostate and male mammary glands (see below for adiscussion of the mammary data). Chronic inflammationof the dorsolateral prostate on PND 50 was reportedfollowing mid-gestational, lactational, and postweaningexposure to dietary genistein at 180 mg/kg bw/day(dose in offspring) (Delclos et al., 2001). Lower ventralprostate weight was observed at PND 50 to 56 followingdietary exposure to Z87 mg/kg bw/day genistein (dosein offspring) (Casanova et al., 1999; Delclos et al., 2001).A limited number of studies examined effects of genisteinexposure during development on hormone levels, and theresults were variable in males and females.
There were no consistent effects on testicular andepididymal weights or testicular morphology in maleoffspring. Lower gonocyte proliferation at PND 3, highernumbers of spermatogonia and preleptotene/leptotenespermatocytes were observed at PND 21, and higherLeydig cell numbers at PND 21 and PND 60 were observedin male offspring of dams given 10 mg/kg bw/daygenistein from GD 14 to birth by gavage (Thuillier et al.,2009). With the exception of one study reporting greaterseverity of abnormal spermatogenesis on PND 50 in malerats given a dietary dose of 180 mg/kg bw/day genisteinfrom GD7–PND 50, which may have been related to theperipubertal status of the rats (Delclos et al., 2001), no otherstudies reported adverse effects on sperm count and/ormotility at genistein doses up to 35 mg/kg bw/daygenistein in diet with exposure during gestation, lactation,and postweaning (2008) or 100 mg/kg bw/day by gavageduring the neonatal period (Nagao et al., 2001). A multi-generation study that included exposures in males duringprenatal and postnatal development reported smaller littersizes at dietary doses of 51 mg/kg bw/day in dams, but noadverse effects on fertility in males at dietary doses up to35 mg/kg bw/day genistein (Delclos et al., 2009).
Table 9Experimental Studies with Developmental Toxicity Endpoints in Other Species Exposed to Genistein, Daidzein, or Equol
Animal model and study design Endpoints NOEL LOEL Reference Utility�
Piglets; 0, 1, or 14 mg/liter genistein informula for 10 days [B0, 0.1–0.4, and2–3 mg/kg bw/day]
k trefoil faction mRNA in stomach – B0.1–0.4 Chen et al.(2005)
Limited(oral)
k enterocyte proliferation and a trend for kenterocyte migration
B0.1–0.4 B2–3
No effect: body weight; intestinal size; jejunavillous morphometric parameters;electrophysiological/clinical measurements(ion, glucose, glutamine transport in jejunumor ileum); jejuna disaccharide, lactase, sucrose;apoptosis in intestinal villi; ERa or ERbexpression in jejunum or ileum; phospho-srcTyr 416 protein expression in jejunum; trefoilfaction mRNA in jejunum or ileum
B2–3 (highdose; noeffect)
–
m,k5 Significant increase, decrease.aBold values represent dose conversions calculated or estimated by CERHR.�The term utility refers to the applicability to the purpose of drawing conclusions on whether or not a chemical adversely affects reproduction.
449NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le10
Stu
die
sin
Ex
per
imen
tal
An
imal
sE
xp
ose
dto
So
yF
orm
ula
,S
oy
Die
ts,
or
Oth
erS
oy
Ex
po
sure
sD
uri
ng
Dev
elo
pm
ent
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsR
efer
ence
Uti
lity
Gro
wth
,re
prod
uct
ive
syst
eman
den
docr
ine-
rela
ted
endp
oin
ts(R
at)
Wis
tar
rat;
soy
pro
tein
-bas
edd
iet
or
sam
ed
iet
wit
his
ofl
avo
nes
rem
ov
edfr
om
pre
-mat
ing
thro
ug
hp
reg
nan
cyan
dla
ctat
ion
So
yp
rote
in-b
ased
die
t:m
acti
vit
yd
uri
ng
dar
kp
erio
dat
3m
on
ths,
bu
tn
ot
6m
on
ths;m
sen
siti
vit
yto
acet
ylc
ho
lin
eat
6m
on
ths
(no
effe
ctat
3m
on
ths)
;m
esen
teri
car
teri
esfr
om
fem
ales
sig
nif
ican
tly
mo
red
iste
nsi
ble
at6
mo
nth
s(u
naf
fect
edat
3m
on
ths)
Do
ug
las
etal
.(2
006)
Lim
ited
(ora
l)
No
effe
ct:
bo
dy
wei
gh
tin
off
spri
ng
;es
tro
us
cycl
ing
;to
tal
or
HD
Lch
ole
ster
ol
or
trig
lyce
rid
esin
3-o
r6-
mo
nth
old
off
spri
ng
;b
loo
dp
ress
ure
or
hea
rtra
te;
mes
ente
ric
arte
ryd
iam
eter
Sp
rag
ue-
Daw
ley
rat
fed
soy
and
alfa
lfa-
free
die
t;D
ams
trea
ted
dai
lyb
yg
avag
ew
ith
0,10
,50
,10
0,15
0,20
0m
g/
kg
bw
/d
ayso
yis
ofl
avo
nes
(80%
,g
enis
tin
:d
aid
zin
:g
lyci
tin
513
:5:2
)fr
om
PN
D5
to10
Die
tsco
nta
inin
gso
yis
ofl
avo
nes
:m
ute
rin
ew
eig
ht
(Z15
0m
g/
kg
bw
/d
ay);m
end
om
etri
alth
ick
nes
s(Z
150
mg
/k
gb
w/
day
);m
thic
kn
ess
of
ute
rin
elu
men
(Z10
0m
g/
kg
bw
/d
ay);m
ov
aria
nw
eig
ht
(Z10
0m
g/
kg
bw
/d
ay);m
seru
mes
trad
iol
(Z50
mg
/k
gb
w/
day
);k
seru
mp
rog
este
ron
e(1
00an
d20
0m
g/
kg
bw
/d
ay;m
inte
nsi
tyo
fP
CN
Ast
ain
ing
ino
var
yan
du
teru
s(Z
150
mg
/k
gb
w/
day
);k
PR
mR
NA
exp
ress
ion
ino
var
y(Z
150
mg
/k
gb
w/
day
);m
ER
mR
NA
exp
ress
ion
ino
var
y(Z
150
mg
/k
gb
w/
day
);k
ER
mR
NA
exp
ress
ion
inu
teru
s(Z
150
mg
/k
gb
w/
day
)
Liu
etal
.(2
008)
Hig
h(o
ral)
No
effe
ct:
pu
pw
eig
ht;
nu
mb
ero
fu
teri
ne
gla
nd
sW
ista
rra
tfe
dca
sein
-bas
edd
iet
or
sam
ed
iet
enri
ched
wit
h0.
87m
g/
gto
tal
iso
flav
on
es[i
sofl
av
on
ein
tak
eo
fB
40
mg
/bw
/day
]d
uri
ng
ges
tati
on
and
lact
atio
n.
At
wea
nin
g,
hal
fo
fth
eF
1p
up
sfr
om
each
mat
ern
ald
iet
gro
up
wer
eas
sig
ned
toei
ther
the
con
tro
ld
iet
or
iso
flav
on
een
rich
edd
iet
for
ato
tal
of
4tr
eatm
ent
gro
up
s(C
-C,
C-I
F,IF
-C,
IF-I
F)
F1
gen
erat
ion
:m
bo
dy
wei
gh
to
fal
lra
tsre
ceiv
ing
iso
flav
on
een
rich
edd
iet;m
ute
rin
ew
eig
ht
fro
m6
to18
mo
nth
s,b
ut
no
tat
24m
on
ths
(IF
-C,
IF-I
Fg
rou
ps)
;p
rolo
ng
edp
erio
do
fin
crea
sin
gb
on
em
iner
ald
ensi
tyo
rB
MD
(IF
-C,
IF-I
Fg
rou
ps)
;m
tota
lri
gh
tfe
mu
rB
MD
and
dis
tal
fem
ur
met
aph
yse
alzo
ne
BM
Dat
24m
on
ths
(IF
-C,
IF-I
Fg
rou
ps)
;m
bo
ne
stre
ng
thw
ith
age
(IF
-C,
IF-I
Fg
rou
ps)
Mar
do
net
al.
(200
8)L
imit
ed(o
ral)
No
effe
ct:
age
atw
hic
hp
eak
bo
ne
mas
sw
asac
hie
ved
;fe
mo
ral
size
;p
lasm
ao
steo
calc
in;
uri
ne
deo
xy
py
rid
ino
lin
e;d
ista
lfe
mu
rd
iap
hy
seal
zon
eB
MD
Lo
ng
Ev
ans
rat;
off
spri
ng
of
dam
sfe
da
die
to
f0,
5,50
,50
0,o
r10
00p
pm
iso
flav
on
esfr
om
GD
12u
nti
lw
ean
ing
atP
ND
21
F1
off
spri
ng
of
dam
sfe
dd
iets
con
tain
ing
iso
flav
on
es:m
PN
D5
bo
dy
wei
gh
to
fm
ale
pu
ps
(5an
d50
pp
m);m
PN
D21
bo
dy
wei
gh
to
fm
ales
(5,
50an
d10
00p
pm
);m
PN
D5
ano
gen
ital
dis
tan
ce(A
GD
)in
mal
ep
up
s(5
,50,
and
500
pp
m)
[AG
Dd
iffe
ren
ces
wer
en
ot
app
aren
tw
hen
no
rmal
ized
for
the
pu
pb
od
yw
eig
hts
];m
PN
D21
test
icu
lar
wei
gh
t,ab
solu
tep
aire
d(Z
5p
pm
);m
test
icu
lar
wei
gh
t,re
lati
ve
(Z50
pp
m)
PN
D21
;m
test
icu
lar
wei
gh
t,ab
solu
te(Z
50p
pm
)P
ND
90;m
seru
mte
sto
ster
on
e(
5p
pm
)at
PN
D21
;k
seru
mte
sto
ster
on
e(5
00an
d10
00p
pm
)at
PN
D21
;m
seru
mte
sto
ster
on
ele
vel
san
dsi
gn
ific
antl
yh
igh
erte
stic
ula
rte
sto
ster
on
ese
cret
ion
PN
D90
(100
0);k
test
ost
ero
ne
pro
du
ctio
np
erL
eyd
igce
llw
as(5
00an
d10
00)
PN
D90
;k
ERa
mR
NA
lev
els
of
the
ven
tral
pro
stat
eo
fP
ND
90
Ak
ing
bem
iet
al.
(200
7)L
imit
ed(o
ral)
No
effe
ct:
litt
ersi
ze;
pu
pw
eig
ht;
sex
rati
os;
PN
D90
bo
dy
wei
gh
t;w
eig
hts
of
the
epid
idy
mis
,d
ors
ola
tera
lan
dv
entr
alp
rost
ate,
bu
lbo
ure
thra
lg
lan
ds
and
sem
inal
ves
icle
PN
D90
Wis
tar
rat;
dam
sg
iven
15.5
%so
y-m
eal
die
to
rso
y-f
ree
die
tp
rio
rto
and
du
rin
gm
atin
g,
pre
gn
ancy
,an
dla
ctat
ion
(n5
7–29
,d
epen
din
go
nen
dp
oin
t)
F1
off
spri
ng
of
dam
sfe
dso
y-m
eal
die
t:m
Ser
toli
cell
nu
clea
rv
olu
me
on
PN
D18
;m
sper
mat
ocy
te/
Ser
toli
cell
nu
clea
rv
olu
me
on
PN
D18
and
PN
D25
;kb
od
yw
eig
ht
and
kte
stis
wei
gh
to
nP
ND
90–9
5;m
pla
sma
FS
Ho
nP
ND
90–9
5
Ata
nas
sov
aet
al.
(200
0)L
imit
ed(o
ral)
Wis
tar
rats
;m
ale
off
spri
ng
of
dam
sfe
da
soy
free
or
soy
mea
l-co
nta
inin
gd
iet
du
rin
gg
esta
tio
nan
dla
ctat
ion
(to
tal
iso
flav
on
eco
nte
nt
inb
reed
ing
and
mai
nte
nan
ced
iets
wer
e31
1an
d26
4mg
/g
die
t)
F1
off
spri
ng
of
dam
sfe
dso
y-m
eal
die
t:m
PN
D60
pla
sma
LH
and
pro
lact
in;
Go
rsk
iet
al.
(200
6)L
imit
ed(o
ral)
No
effe
ct:
rep
rod
uct
ive
org
anw
eig
hts
PN
D22
or
PN
D60
;
450 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Sp
rag
ue-
Daw
ley
rats
,tw
ow
eek
so
fag
e[f
ee
dn
ot
des
crib
ed];
sin
gle
dai
lyd
ose
of
gen
iste
in(4
0m
g/
kg
BW
)b
ysc
inje
ctio
nfo
r3
day
s
mex
pre
ssio
nle
vel
so
fg
enes
inim
mat
ure
rat
ute
rus
(m9.
97%
)H
on
get
al.
(200
6)L
imit
ed(p
aren
tera
l)
Lo
ng
Ev
ans
rat;
dam
s(n
55/
gro
up
)g
iven
ph
yto
estr
og
en-c
on
tain
ing
die
t(6
00mg
/g
die
t)o
rp
hy
toes
tro
gen
-fre
efi
shm
eal-
bas
edd
iet
thro
ug
ho
ut
pre
gn
ancy
and
lact
atio
n.
Off
spri
ng
wea
ned
tod
am’s
die
t,ex
cep
tat
PN
D80
wh
eno
ne-
hal
fo
fp
hy
toes
tro
gen
-co
nta
inin
gd
iet
fed
off
spri
ng
wer
esw
itch
edto
ph
yto
estr
og
en-f
ree
die
t
F1
off
spri
ng
of
dam
sfe
dp
hy
toes
tro
gen
-co
nta
inin
gd
iet:
del
ayed
vag
inal
op
enin
g;k
bo
dy
wei
gh
to
nP
ND
50an
d12
0in
mal
ean
dfe
mal
eo
ffsp
rin
g;m
inp
rost
ate
wei
gh
t(a
bso
lute
and
rela
tiv
e);m
SD
N-P
OA
inp
hy
toes
tro
gen
die
tm
ales
com
par
edto
con
tro
lm
ales
and
mal
essw
itch
edfr
om
ph
yto
estr
og
end
iet
top
hy
toes
tro
gen
-fre
ed
iet
atP
ND
80
Lu
nd
etal
.(2
001)
Lim
ited
(ora
l)
Sp
rag
ue-
Daw
ley
rat;
pre
gn
ant
rats
(n5
15/
gro
up
)g
iven
CR
F-1
,a
soy
-bas
edd
iet
(gen
isti
nat
10.2
mg
/10
0g
die
tan
dd
aid
zin
at8.
7m
g/
100
gd
iet)
,or
NIH
-07,
aso
y-f
ree
die
t
F1
off
spri
ng
inso
y-b
ased
die
t(C
RF
-1)
gro
up
:mP
ND
2m
ale
pu
pb
od
yw
eig
ht
on
PN
D2;
mP
ND
21fe
mal
ep
up
bo
dy
wei
gh
t;k
ano
gen
ital
dis
tan
cead
just
edfo
rb
od
yw
eig
ht
infe
mal
es
Mas
uto
mi
etal
.(2
004a
)L
imit
ed(o
ral)
Sp
rag
ue-
Daw
ley
rat;
mal
es(n
56–
12/
gro
up
)fr
om
mu
ltig
ener
atio
nst
ud
yth
atco
nti
nu
edm
ale
off
spri
ng
on
1o
f6
die
tsg
iven
toth
eir
par
ents
:A
IN-9
3G(s
oy
-fr
ee,
case
in-b
ased
die
t),
Co
ntr
ol
die
t1so
yp
rote
in(i
sofl
avo
nes
31.7
mg
/k
gfe
ed),
or
Co
ntr
ol
die
t1so
yp
rote
in1
soy
-iso
flav
on
eco
nce
ntr
ate
(No
vas
oys
)at
36.1
,74
.5,
235.
8,o
r10
46.6
mg
/k
gfe
ed
mT
esti
sw
eig
ht,
PN
D28
in3
hig
hes
tis
ofl
avo
ne
gro
up
s(t
ota
lis
ofl
avo
ne
inta
keZ
2m
g/
kg
bw
/d
ay)
McV
eyet
al.
(200
4a,b
)L
imit
ed(o
ral)
mS
eru
mte
sto
ster
on
ean
dd
ihy
dro
test
ost
ero
ne,
PN
D12
0in
2h
igh
est
iso
flav
on
eg
rou
ps
(to
tal
iso
flav
on
ein
tak
eZ
6.3
mg
/k
gb
w/
day
)m
Tes
ticu
lar
3b-h
yd
rox
yst
ero
idd
ehy
dro
gen
ase
acti
vit
y,P
ND
28,
in2
hig
hes
tis
ofl
avo
ne
gro
up
s[B
ased
on
PN
D28
test
isw
eig
ht:
NO
AE
L1.
2m
g/
kg
bw
/d
ayan
dL
OA
EL
2.0
mg
/k
gb
w/
day
]N
oef
fect
:m
ean
bo
dy
wei
gh
to
rfe
edco
nsu
mp
tio
nat
any
tim
ed
uri
ng
the
stu
dy
;W
ista
r-d
eriv
edra
t;d
ams
(n5
12/
gro
up
)g
iven
assi
gn
edd
iet
du
rin
gp
reg
nan
cyan
dla
ctat
ion
,an
do
ffsp
rin
gw
ean
edto
assi
gn
edd
iet
thro
ug
hP
ND
68.
Die
tas
sig
nm
ents
(dam
/o
ffsp
rin
g)
wer
e:
Eff
ects
com
pare
dto
RM
3/R
M1
Con
trol
diet
:O
du
met
al.
(200
1)L
imit
ed
RM
3/R
M1
(sta
nd
ard
soy
-bas
edd
iets
)A
IN-7
6/A
IN-7
6(s
oy
-an
dal
falf
a-fr
eed
iet)
AIN
-76A
/A
IN-7
6A(s
oy
free
die
td
uri
ng
pre
gn
ancy
and
lact
atio
n):m
age
atte
stis
des
cen
t;k
age
atp
rep
uti
alse
par
atio
n;k
age
ato
nse
to
fv
agin
alo
pen
ing
;P
ND
26fe
mal
es:m
bo
dy
wei
gh
t,u
teri
ne
wei
gh
tan
dv
agin
alw
eig
ht;
PN
D68
mal
es:m
bo
dy
wei
gh
t;m
liv
eran
dk
idn
eyw
eig
ht;k
test
isan
dep
idid
ym
alw
eig
ht;
PN
D12
8m
ales
:mb
od
yw
eig
ht;k
test
isw
eig
ht.
PN
D14
0–14
4fe
mal
es:m
bo
dy
wei
gh
t;m
kid
ney
wei
gh
t;k
ute
rin
ew
eig
ht
RM
3/A
IN-7
6A(s
oy
die
td
uri
ng
pre
gn
ancy
and
soy
free
die
td
uri
ng
lact
atio
n):k
age
atte
stis
RM
3/A
IN-7
6AN
oef
fect
:ey
eo
pen
ing
Glo
bal
2016
/G
lob
al20
16(s
oy
-an
dal
falf
a-fr
eed
iet)
Pu
rin
a50
01/
Pu
rin
a50
01(b
ased
on
soy
bea
ns
and
alfa
lfa)
Wis
tar
rat;
soy
-co
nta
inin
gd
iet,
soy
-fre
ed
iet,
and
soy
-fre
ed
iet
sup
ple
men
ted
wit
hm
eth
ion
ine
and
L-l
ysi
ne
km
atin
gef
fici
ency
for
F0
anim
als;k
mea
nli
ver
wei
gh
tat
60d
ays
inF
1fe
mal
es;m
LH
and
pro
lact
inat
60d
ays;
dif
fere
nce
inte
sto
ster
on
ele
vel
no
tco
nfi
rmed
stat
isti
call
yP
astu
szew
ska
etal
.(2
008)
Lim
ited
No
effe
ct:
feed
inta
ke
and
bo
dy
wei
gh
tfo
rF
0g
ener
atio
n;
acti
vit
yo
fh
epat
icen
zym
es;
tota
lch
ole
ster
ol
Gro
wth
,R
epro
duct
ive
Sys
tem
and
En
docr
ine-
Rel
ated
En
dpoi
nts
(Mou
se)
CD
-1m
ice;
F0
dam
sd
uri
ng
ges
tati
on
and
lact
atio
nan
do
ffsp
rin
gaf
ter
wea
nin
gw
ere
fed
aso
y-b
ased
die
t(P
MI
5008
or
PM
I50
01)
or
soy
-fre
ed
iet
(PM
I5K
96)
F1
off
spri
ng
of
dam
sfe
dso
y-b
ased
die
t(P
MI
5008
or
PM
I50
01)
com
par
edto
soy
-fre
ed
iet
(PM
I5K
96):k
seru
mes
trad
iol
lev
els
infe
tuse
s;m
bir
thw
eig
ht
of
pu
ps;k
PN
D26
and
PN
D90
bo
dy
wei
gh
t(b
ut
no
effe
ctat
PN
D20
);k
ute
rin
ew
eig
ht
and
epit
hel
ial
cell
hei
gh
tin
PN
D20
fem
ales
inre
spo
nse
toes
trad
iol
stim
ula
tio
n;k
ute
rin
ew
eig
ht
and
epit
hel
ial
cell
hei
gh
tin
PN
D26
fem
ales
;la
ter
on
set
of
fert
ilit
yin
fem
ales
(46.
5v
ersu
s44
.7d
ays)
;k
pro
stat
ew
eig
ht
inm
ales
;b
ette
rg
luco
secl
eara
nce
inad
ult
mal
es;
mte
stes
,ep
idid
ym
ides
,se
min
alv
esic
les,
liv
eran
dri
gh
tk
idn
eyw
eig
ht
inm
ales
;k
wei
gh
to
fg
on
adal
and
ren
alfa
tp
ads
inan
imal
s;k
seru
mle
pti
nin
PN
D26
fem
ales
and
PN
D90
inm
ales
and
fem
ales
Ru
hle
net
al.
(200
8)L
imit
ed(o
ral)
451NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le10
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsR
efer
ence
Uti
lity
No
effe
ct:
seru
mes
trad
iol
lev
els
pre
gn
ant
dam
s;P
ND
20se
rum
lep
tin
;b
asal
blo
od
glu
cose
lev
els
and
glu
cose
clea
ran
cein
adu
ltfe
mal
es;
mea
np
up
wei
gh
to
rth
en
um
ber
of
pu
ps
pro
du
ced
by
fem
ales
on
eith
erd
iet
C3H
mic
e:F
0g
ener
atio
nfe
da
stan
dar
dco
ntr
ol
die
t[a
ssu
me
dto
con
tain
som
eso
y]
or
aco
ntr
ol
die
t12%
(So
yL
ifes
)su
pp
lem
ent
(IS
Fd
iet)
du
rin
gp
reg
nan
cyan
dla
ctat
ion
;F
1fe
dth
esa
me
die
tsas
thei
rp
aren
tsp
ost
-wea
nin
g
F1
off
spri
ng
inIS
Fd
iet
gro
up
:ac
cele
rate
dv
agin
alo
pen
ing
;k
bo
dy
wei
gh
t,si
zean
dan
og
enit
ald
ista
nce
infe
mal
esat
the
day
of
vag
inal
op
enin
g;k
size
and
bo
dy
wei
gh
to
nP
ND
42;
loss
of
sex
ual
dim
orp
his
min
DN
Am
eth
yla
ton
inli
ver
Gu
erre
ro-
Bo
sag
na
etal
.(2
008)
Lim
ited
(ora
l)
No
effe
ct:p
up
sex
rati
oo
rli
tter
size
;an
og
enit
ald
ista
nce
inm
ales
or
fem
ales
on
PN
D42
;p
up
size
or
wei
gh
tat
PN
D7,
PN
D14
,o
rP
ND
21;
ER
ap
rom
ote
rin
liv
er;
gen
der
dif
fere
nce
so
rw
ith
inm
ales
or
fem
ales
inA
cta1
inp
ancr
eas;
lack
of
met
hy
lati
on
of
c-fo
sp
rom
oto
rH
an-N
MR
Im
ice;
Dam
sfe
dso
yd
iet
or
soy
-fre
ed
iet
du
rin
gp
reg
nan
cyan
dla
ctat
ion
;m
ales
inea
chli
tter
wer
eco
nti
nu
edo
nth
ed
am’s
die
to
rsw
itch
edto
the
op
po
site
die
t;co
llec
tio
nat
2an
d12
mo
nth
so
fag
e(m
ales
)
2m
on
th:
Mak
ela
etal
.(1
995)
Lim
ited
(ora
l)m
coag
ula
tin
gg
lan
dan
dco
mb
ined
pro
stat
elo
be
rela
tiv
ew
eig
hts
inm
ales
fed
aso
yd
iet
afte
rw
ean
ing
,fo
llo
win
ga
soy
-fre
ed
iet
giv
ento
the
dam
du
rin
gp
reg
nan
cyan
dla
ctat
ion
;m
rela
tiv
ew
eig
ht
of
the
pro
stat
elo
bes
wh
enco
mp
ared
tom
ales
pla
ced
on
aso
y-f
ree
die
taf
ter
wea
nin
g,
reg
ard
less
of
wh
ich
die
tth
eir
dam
sh
adb
een
fed
;S
oy
feed
ingm
rela
tiv
ew
eig
ht
of
coag
ula
tin
gg
lan
d(4
6%),
do
rsal
ater
alp
rost
ate
(81%
)12
mo
nth
:S
oy
feed
ingm
rela
tiv
ere
pro
du
ctiv
eo
rgan
wei
gh
ts-
ven
tral
pro
stat
e(4
0%),
coag
ula
tin
gg
lan
d(6
0%),
do
rsal
pro
stat
e(6
3%),
sem
inal
ves
icle
(35%
)an
dte
stes
(20%
);N
oef
fect
:o
nb
od
yw
eig
ht
Gro
wth
,re
prod
uct
ive
syst
eman
den
docr
ine-
rela
ted
endp
oin
ts(n
onhu
man
prim
ate)
Mar
mo
set
mo
nk
eys;
7se
tso
fco
-tw
ins
wh
ere
on
etw
inw
asfe
da
cow
-mil
kb
ased
form
ula
(SM
AG
oldt
)an
dth
eo
ther
was
fed
soy
infa
nt
form
ula
(Wy
soys
)fr
om
day
4o
r5
tod
ay35
to45
;fo
rmu
lau
sew
aso
nly
par
tial
asm
on
key
sal
son
urs
ed(e
stim
ated
tore
sult
inis
ofl
avo
ne
inta
ke
bet
wee
n40
and
87%
of
the
rep
ort
edin
tak
efo
ra
4-m
on
tho
ldh
um
anex
clu
siv
ely
fed
soy
form
ula
)
Eff
ects
inso
yin
fan
tfo
rmu
lag
rou
pu
pto
120
to13
8w
eek
so
fag
e:m
test
isw
eig
ht;m
Ser
toli
cell
and
Ley
dig
cell
nu
mb
ers/
test
isT
anet
al.
(200
6)L
imit
ed(o
ral)
No
effe
ct:
bo
dy
wei
gh
t;w
eig
hts
of
thy
mu
s,sp
leen
,p
rost
ate,
sem
inal
ves
icle
s,p
itu
itar
y;
pen
ile
len
gth
,ti
min
go
fo
nse
to
fp
ub
erty
;p
lasm
ate
sto
ster
on
e;fe
rtil
ity
Mar
mo
set
mal
em
on
key
s;co
-tw
ins
wh
ere
on
etw
inw
asfe
da
cow
-mil
kb
ased
form
ula
(SM
AG
oldt
)an
dth
eo
ther
was
fed
soy
infa
nt
form
ula
(Wy
soys
)fr
om
day
4–5
tod
ay35
–45
(n5
7–14
twin
pai
rs,
dep
end
ing
on
age
of
eval
uat
ion
);fo
rmu
lau
sew
aso
nly
par
tial
asm
on
key
sal
son
urs
ed(e
stim
ated
tore
sult
inis
ofl
avo
ne
inta
ke
ofB
1.6–
3.5
mg
/k
gb
w/
day
)
Eff
ects
inso
yin
fan
tfo
rmu
lag
rou
pu
pto
PN
D35
–45:
kp
lasm
ate
sto
ster
on
eat
PN
D35
–45
;m
Ley
dig
cell
s/te
stis
atP
ND
35–4
5S
har
pe
etal
.(2
002)
Lim
ited
(ora
l)
No
effe
ct:
bo
dy
wei
gh
t;fo
rmu
lain
tak
e;p
lasm
ate
sto
ster
on
eP
ND
18–2
0;te
stis
wei
gh
t,S
erto
lice
llan
dg
erm
cell
nu
mb
er/
test
isat
PN
D35
–45;
17a-
hy
dro
xy
lase
/C
17–20
lyas
est
ain
ing
inte
stis
and
imm
un
op
osi
tiv
ece
lls
forb-
sub
un
itL
Ho
rF
SH
inp
itu
itar
yse
ctio
ns
atP
ND
35–4
5
Mam
mar
ygl
and
deve
lopm
ent
and
carc
inog
enes
isS
pra
gu
e-D
awle
yra
tfe
dA
IN-9
3Gd
iet
inw
hic
hso
yb
ean
oil
was
rep
lace
db
yco
rno
ilan
dth
ep
rote
inso
urc
ew
asei
ther
case
in,
wh
ey,
or
soy
pro
tein
iso
late
con
tain
ing
iso
flav
on
es43
0m
g/
kg
die
t(g
enis
tein
276
mg
/k
gd
iet
and
dai
dze
in13
2m
g/
kg
die
t)[B
20
.4g
en
iste
inm
g/k
gb
w/d
ay].
At
wea
nin
g,
the
F1
and
F2
off
spri
ng
wer
efe
dth
esa
me
die
tsas
thei
rd
ams
and
con
tin
ued
tore
ceiv
e
F1
and
/o
rF
2fe
mal
esin
soy
-pro
tein
gro
up
:k
bo
dy
wei
gh
tg
ain
at8
wee
ks
(bo
thg
ener
atio
ns)
;ac
cele
rate
dv
agin
alo
pen
ing
(PN
D37
vs
PN
D38
);m
late
ncy
tod
evel
op
mam
mar
yg
lan
dtu
mo
rs,
inte
rms
of
the
nu
mb
ero
fp
ost
-tre
atm
ent
day
sfo
r50
%o
fra
tsto
dev
elo
pat
leas
t1
tum
or
(F1
and
F2,
com
par
edto
case
in);k
late
ncy
tod
evel
op
mam
mar
yg
lan
dtu
mo
rs,
inte
rms
of
the
nu
mb
ero
fp
ost
-tre
atm
ent
day
sfo
r50
%o
fra
tsto
dev
elo
pat
leas
t1
tum
or
(F2,
com
par
edto
wh
ey);k
per
cen
tag
eo
fra
tsw
ith
at
Hak
kak
etal
.(2
000)
Lim
ited
(ora
l)
452 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

the
die
tsth
rou
gh
ou
tth
eir
life
tim
es.
At
PN
D50
,ra
tsg
avag
edw
ith
80m
g/
kg
DM
BA
and
the
dev
elo
pm
ent
of
pal
pab
lem
amm
ary
tum
ors
asse
ssed
leas
t1
mam
mar
ytu
mo
rat
the
end
of
the
stu
dy
(F2);m
tum
ors
/ra
tco
mp
ared
tow
hey
die
tg
rou
p(F
2)
No
effe
ct:
rela
tiv
eo
rgan
wei
gh
t;es
tro
us
cycl
e;su
cces
sfu
lb
reed
ing
;se
xra
tio
;li
tter
size
;ti
me
tofi
rst
tum
or;
%o
fra
tsw
ith
aden
oca
rcin
om
ao
rtu
mo
rw
ith
intr
adu
ctal
pro
life
rati
on
;m
edia
ntu
mo
rv
olu
me;
tum
ors
/ra
tco
mp
ared
toca
sein
die
tg
rou
pS
pra
gu
e-D
awle
yra
t;fe
mal
eF
1o
ffsp
rin
go
fF
0p
aren
tsfe
dA
IN-9
3Gd
iets
con
tain
ing
eith
erca
sein
or
soy
-pro
tein
iso
late
fro
mG
D4
tola
ctat
ion
,at
wea
nin
gfe
mal
esw
ere
wea
ned
toth
ed
iet
of
the
dam
.A
tP
ND
50,
rats
ivin
ject
edw
ith
50m
g/
kg
MN
Uan
dfo
llo
wed
un
til
115
day
sfo
llo
win
gM
NU
trea
tmen
t
Fem
ale
F1
off
spri
ng
of
F0
par
ents
fed
soy
-pro
tein
iso
late
:kin
cid
ence
of
rats
wit
hat
leas
t1
mam
mar
yg
lan
dtu
mo
ran
dm
tum
or
late
ncy
inra
tsfe
dth
eso
y-p
rote
ind
iet;k
inci
den
ceo
fd
uct
alca
rcin
om
ain
situ
bu
ta
hig
her
inci
den
ceo
fin
filt
rati
ng
du
ctal
carc
ino
ma;
mP
Ran
dH
ER
-2/n
eu(i
nv
olv
edin
cell
pro
life
rati
on
)g
ene
exp
ress
ion
:k
seru
mp
rog
este
ron
ele
vel
s;m
apo
pto
tic
cell
sin
anM
CF
-7cu
ltu
rein
cub
ated
wit
hse
ra
Sim
men
etal
.(2
005)
Lim
ited
(ora
l)
No
effe
ct:
cell
pro
life
rati
on
or
apo
pto
sis;
seru
mes
trad
iol
FV
Bm
ice
fed
(I)
con
tro
ld
iet
(F1
fem
ales
),(I
I)27
0m
gis
ofl
avo
ne/
kg
die
tth
rou
gh
ou
tla
ctat
ion
(PN
D0–
21),
(III
)27
0m
gis
ofl
avo
ne/
kg
die
tfr
om
wea
nin
gth
rou
gh
the
end
of
the
stu
dy,
and
(IV
)E
2d
uri
ng
the
lact
atio
np
erio
d
Eff
ects
inis
ofl
avo
ne-
trea
ted
gro
up
s(G
rou
ps
IIan
d/
or
III)
:m
bra
nch
ing
inm
amm
ary
tree
s(G
rou
ps
II/
III)
;m
ino
ver
all
bra
nch
ing
inju
ven
ile
mam
mar
yg
lan
d(P
ND
28;
Gro
up
sII
/II
I));m
TE
Baf
ter
po
stw
ean
ing
iso
flav
on
etr
eatm
ent
(Gro
up
III)
;k
TE
Bs
atP
ND
42–4
3af
ter
po
stw
ean
ing
(Gro
up
sII
I)
Th
om
sen
etal
.(2
006)
Lim
ited
(ora
l)
No
effe
ct:
PN
Ds
70–7
3;le
vel
so
fp
lasm
aes
trad
iol;
gen
eex
pre
ssio
np
rofi
les;
gen
eex
pre
ssio
nle
vel
wh
enco
mp
arin
gp
ost
wea
nin
gis
ofl
avo
ne
exp
osu
reto
E2
exp
osu
re
Bra
inan
dbe
havi
orL
on
gE
van
rat;
pre
gn
ant
dam
sfe
da
die
tco
nta
inin
gp
hy
toes
tro
gen
gly
cosi
des
600mg
/g
die
t(p
hy
to-6
00)
or
ap
hy
toes
tro
gen
-fre
ed
iet
thro
ug
ho
ut
the
ges
tati
on
and
lact
atio
np
erio
d.
Off
spri
ng
wer
ew
ean
edto
the
sam
ed
iet
asth
ed
am.
On
PN
D80
,h
alf
the
off
spri
ng
fed
ph
yto
-600
wer
esw
itch
edto
ap
hy
toes
tro
gen
-fre
ed
iet
for
40d
ays
pri
or
toco
llec
tio
nat
PN
D12
0
Eff
ects
inp
hy
to-6
00):k
bo
dy
wei
gh
t,ab
solu
tep
rost
ate
wei
gh
tan
dre
lati
ve
pro
stat
ew
eig
ht
PN
D75
;mA
VP
Vn
ucl
eus
vo
lum
ein
mal
essw
itch
edto
the
ph
yto
estr
og
en-f
ree
die
to
nP
ND
80
Lep
har
tet
al.
(200
1)L
imit
ed(o
ral)
No
effe
ct:
bra
inar
om
atas
eac
tiv
ity
inA
PV
Nan
dan
tero
ven
tral
per
iven
tric
ula
rn
ucl
eus
vo
lum
ein
fem
ales
un
affe
cted
by
chan
ge
ind
iet,
PN
D11
0;se
xu
ald
imo
rph
ism
inA
VP
Vv
olu
me;
Lo
ng
Ev
ans
rat;
Dam
sg
iven
ph
yto
estr
og
en-c
on
tain
ing
die
t(6
00mg
/g
die
t)o
rp
hy
toes
tro
gen
-fre
efi
shm
eal-
bas
edd
iet
thro
ug
ho
ut
pre
gn
ancy
and
lact
atio
n.
Off
spri
ng
wea
ned
tod
am’s
die
t
F1
off
spri
ng
of
dam
sfe
dp
hy
toes
tro
gen
-co
nta
inin
gd
iet:k
PN
D50
bo
dy
wei
gh
t;m
PN
D50
wat
erin
tak
e;m
tim
esp
end
ino
pen
arm
san
dn
um
ber
of
entr
ies
into
the
op
enar
ms
of
elev
ated
plu
s-m
aze
app
arat
us
Lu
nd
and
Lep
har
t(2
001)
Lim
ited
(ora
l)
Sp
rag
ue-
Daw
ley
rats
;D
ams
fed
ap
hy
toes
tro
gen
con
tain
ing
die
t(2
00mg
/g
die
t,g
enis
tein
95mg
/g
,g
lyci
tin
17mg
/g
,an
dd
aid
zein
82mg
/g
)o
rp
hy
toes
tro
gen
free
die
tfr
om
GD
0–20
Fet
use
so
fd
ams
fed
ph
yto
estr
og
enco
nta
inin
gd
iet:k
calb
ind
inp
rote
inex
pre
ssio
nin
the
med
ial
bas
alh
yp
oth
alam
us
and
pre
op
tic
area
infe
mal
es(a
nd
dis
pla
yed
mo
reo
fa
sex
ual
dim
orp
his
mco
mp
ared
toan
imal
sin
the
ph
yto
estr
og
en-f
ree
die
tg
rou
p);
Tay
lor
etal
.(1
999)
Lim
ited
(ora
l)
No
effe
ct:
litt
ersi
ze,
ano
gen
ital
dis
tan
ce,
feta
lb
od
yw
eig
ht;
calb
ind
inp
rote
inex
pre
ssio
nin
mal
es
Oth
er/M
echa
nis
tic
Fem
ale
New
Zea
lan
dra
bb
its
(8m
on
ths
old
)fe
d(G
rou
p1)
soy
and
alfa
lfa
free
die
t(G
rou
p2)
soy
and
alfa
lfa
free
die
tp
lus
5m
g/
kg
bw
/d
ayay
of
soy
iso
flav
on
es(I
SG
O5)
,(G
rou
p3)
aso
yan
dal
falf
afr
eed
iet,
and
(Gro
up
4)d
iet
con
tain
ing
18%
soy
mea
lp
lus
aco
rnst
arch
pla
ceb
o(S
1)
(B13
mg
/k
gb
w/
day
ayo
fis
ofl
avo
nes
).P
up
sw
ean
edat
5w
eek
so
ldan
dfe
dsa
me
die
tas
mo
ther
kw
eig
ht
inIS
F20
gro
up
at13
wee
ks
of
age;
kfo
od
con
sum
pti
on
inIS
F20
gro
up
at13
wee
ks
of
age
Car
do
soan
dB
ao(2
007)
Lim
ited
(ora
l)
No
effe
ct:
org
anw
eig
hts
;h
isto
pat
ho
log
ical
dif
fere
nce
sin
the
test
eso
rac
cess
ory
sex
gla
nd
s;ag
eth
atm
ales
exh
ibit
edth
em
ou
nti
ng
refl
exo
rin
the
tim
eo
fre
acti
on
;
Rab
bit
s;P
reg
nan
td
oes
fed
(1)
con
tro
ld
iet
(aso
yan
dal
falf
afr
eed
iet)
,(2
)co
ntr
ol
die
tsu
pp
lem
ente
dw
ith
10m
g/
kg
bw
/d
ayay
of
soy
iso
flav
on
es,(
3)co
ntr
ol
die
tsu
pp
lem
ente
dw
ith
20m
g/
kg
bw
/d
ayay
iso
flav
on
es,
or
(4)
ad
iet
con
tain
ing
18%
soy
mea
l
No
effe
ct:
on
dam
feed
inta
ke,
du
rati
on
of
ges
tati
on
,or
litt
ersi
ze;o
ffsp
rin
gb
od
yw
eig
ht,
his
top
ath
olo
gy
of
the
test
eso
rep
idid
ym
ides
;o
rgan
wei
gh
ts;
fem
iniz
atio
no
rd
emas
culi
niz
atio
n;
mal
ese
xu
alb
ehav
ior;
ejac
ula
tev
olu
me;
sper
mm
oti
lity
,v
igo
r,co
nce
ntr
atio
n
Car
do
soan
dB
ao(2
008)
Lim
ited
(ora
l)
453NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tab
le10
Co
nti
nu
ed
An
imal
mo
del
and
stu
dy
des
ign
En
dp
oin
tsR
efer
ence
Uti
lity
Ou
tbre
dC
D-1
mic
efe
dA
IN-9
3Gd
iets
dev
oid
of
estr
og
enic
com
po
un
ds;
F1
off
spri
ng
trea
ted
by
scin
ject
ion
wit
hco
rno
il,
dai
dze
in(2
mg
/k
gb
w/
day
ay),
gen
iste
in(5
mg
/k
gb
w/
day
ay),
dai
dze
inp
lus
gen
iste
in(2
mg
dai
dze
in1
5m
gg
enis
tein
/k
g/
day
),o
rd
ieth
yls
tilb
estr
ol
(2m
g/
kg
bw
/d
ayay
);ad
min
iste
red
each
mo
rnin
gfr
om
PN
D1
toP
ND
5(5
do
ses)
var
iety
of
effe
cts
on
bo
ne
mea
sure
s,i.
e.,
bo
ne
min
eral
con
ten
t,b
on
em
iner
ald
ensi
ty,
mic
roar
chit
ectu
reth
atw
ere
inte
rpre
ted
as‘‘
po
siti
ve’
’ef
fect
sal
tho
ug
hth
eco
mb
inat
ion
of
gen
iste
inan
dd
aid
zein
was
no
tco
nsi
der
edto
hav
ea
gre
ater
ben
efit
than
the
ind
ivid
ual
trea
tmen
ts
Kal
ud
jero
vic
etal
.(2
009)
Lim
ited
(par
ente
ral)
No
effe
ct:
bo
dy
wei
gh
t
Pig
lets
(n5
16fr
om
4li
tter
s)ra
nd
om
lyas
sig
ned
48%
soy
bea
nm
eal-
bas
edli
qu
idd
iet
or
cow
-mil
kb
ased
die
t;p
rote
inso
urc
eco
nti
nu
edaf
ter
wea
nin
gu
nti
l56
day
so
fag
e
kb
od
yw
eig
ht
gai
nat
28d
ays
of
age;
inte
stin
alv
illi
sho
rten
edat
28b
ut
no
t56
day
so
fag
e;m
IgG
tite
rsto
soy
inb
oth
age
gro
up
so
fp
igle
tsfe
dso
yb
ean
com
par
edto
mil
k-
pro
tein
die
ts
Li
etal
.(1
990)
Lim
ited
(ora
l)
No
effe
cts
of
die
to
nse
rum
con
cen
trat
ion
so
fzi
nc,
sele
niu
m,
iro
n,
and
cop
per
wer
ed
etec
ted
at28
and
56d
ays
of
age.
;N
osi
gn
ific
ant
dif
fere
nce
sin
skin
thic
kn
ess
foll
ow
ing
intr
ader
mal
inje
ctio
no
fso
yo
rm
ilk
pro
tein
sw
ere
ob
serv
edb
etw
een
the
soy
bea
n-
and
mil
k-d
iet
gro
up
sat
27an
d55
day
so
fag
e;o
nb
last
og
enic
resp
on
ses
of
per
iph
eral
or
inte
stin
ally
mp
ho
cyte
sco
llec
ted
atei
ther
age
and
cult
ure
dw
ith
pu
rifi
edso
yp
rote
ins,
ph
yto
hem
agg
luti
nin
,o
rp
ok
ewee
dS
pra
gu
e-D
awle
yra
ts;
Ex
per
imen
t1:
Pre
gn
ant
rats
fed
AIN
-93G
(CA
S)
die
t.O
nP
ND
15li
tter
sas
sig
ned
to:
con
tro
l(C
AS
)d
iet,
SP
I1d
iet
(286
mg
/k
gg
enis
tein
and
226
mg
/k
gd
aid
zein
,80
mg
tota
lis
ofl
avo
nes
/k
gB
W/
day
),S
PI-
die
t,C
1G
die
t(2
50m
g/
kg
gen
iste
in),
C1
Gd
iet
(250
mg
/k
gd
aid
zein
);p
up
sfe
dto
PN
D33
Eff
ects
inS
PI
die
tg
rou
pco
mp
ared
toca
sein
con
tro
l:m
Ser
um
gen
iste
inle
vel
so
fm
ale
rats
inth
eC
1G
die
tg
rou
p(2
700
nm
ol/
lite
r)th
anth
eS
PI1
die
tg
rou
p(8
08n
mo
l/li
ter)
atP
ND
50;k
bo
dy
wei
gh
tan
dse
rum
IGF
-1co
nce
ntr
atio
ns,
and
alte
red
gen
eex
pre
ssio
np
atte
rnw
ith
ano
ver
all
pat
tern
of
incr
ease
dex
pre
ssio
no
fh
epat
icg
enes
reg
ula
ted
by
PP
ARa,
PP
ARg,
and
LX
Ra
and
dec
reas
edex
pre
ssio
no
fg
enes
reg
ula
ted
by
SR
EB
P-1
c
Ro
nis
etal
.(2
009)
Lim
ited
(ora
l)
Ex
per
imen
t2:
Mal
era
tsre
ceiv
edA
IG-9
36(C
AS
die
t),
Wes
tern
case
ind
iet,
or
Wes
tern
SP
I1d
iet
fro
mP
ND
24to
64
mw
eig
ht
gai
n,
per
cen
tb
od
yfa
t,p
erce
nt
liv
erw
eig
ht;m
seru
min
suli
nco
nce
ntr
atio
n;m
insu
lin
sen
siti
vit
y;m
exp
ress
ion
of
hep
atic
gen
esre
gu
late
db
yP
PA
Ra,
PP
ARg,
and
LX
Ra;
kex
pre
ssio
no
fg
enes
reg
ula
ted
by
SR
EB
P-1
cA
du
ltfe
mal
ecy
no
mo
lgu
sm
on
key
s(n
519
)an
do
ffsp
rin
g(n
525
)fe
dty
pic
alA
mer
ican
die
t(T
AD
)d
iet
wit
hp
rote
ind
eriv
edfr
om
soy
or
TA
Dd
iet
wit
hth
ep
rote
ind
eriv
edfr
om
case
in-l
acta
lbu
min
.O
ffsp
rin
gco
nti
nu
edT
AD
die
tfe
dto
mo
ther
saf
ter
wea
nin
g.T
AD
soy
die
tp
rov
ided
iso
flav
on
eag
lyco
ne-
equ
ival
ent
do
seo
f18
0m
g/
mo
nk
ey/
day
(B40
–50%
gen
iste
in,
40%
dai
dze
in,
and
10–1
5%g
lyci
tein
)
mse
rum
iso
flav
on
eco
nce
ntr
atio
nin
TA
Dso
yd
iet
mo
nk
eys;
(Off
spri
ng
con
sum
ing
TA
Dca
sein
had
sim
ilar
iso
flav
on
eco
nce
ntr
atio
ns
toad
ult
sco
nsu
min
gT
AD
case
in,
bu
to
ffsp
rin
gco
nsu
min
gT
AD
soy
had
hig
her
con
cen
trat
ion
sth
anth
ead
ult
fem
ales
con
sum
ing
TA
Dso
y);k
fru
cto
sam
ine
con
cen
trat
ion
s;k
bo
dy
wei
gh
tat
1an
d2
yea
rso
fag
e(o
ffsp
rin
g);m
insu
lin
inm
ales
com
par
edw
ith
fem
ales
;m
HD
LC
infe
mal
esco
mp
ared
wit
hm
ales
;k
glu
cose
AU
Can
dfa
ster
dis
app
eara
nce
of
glu
cose
(off
spri
ng
);k
insu
lin
resp
on
ses
tog
luco
sech
alle
ng
e
Wag
ner
etal
.(2
009)
Lim
ited
(ora
l)
No
effe
ct:
bo
dy
wei
gh
t,T
C,
TG
,ap
oB
-C,
glu
cose
(ad
ult
s);
bo
dy
wei
gh
tat
bir
th(o
ffsp
rin
g);
HD
LC
con
cen
trat
ion
;fa
stin
gg
luco
sean
din
suli
nco
nce
ntr
atio
n
� Th
ete
rmu
tili
tyre
fers
toth
eap
pli
cab
ilit
yto
the
pu
rpo
seo
fd
raw
ing
con
clu
sio
ns
on
wh
eth
ero
rn
ot
ach
emic
alad
ver
sely
affe
cts
rep
rod
uct
ion
.
454 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

The majority of oral exposure studies reported no effectson anogenital distance after adjustment for body weightin male and female rats following genistein doses up to180 mg/kg bw/day (doses in offspring) administeredduring gestation, lactation, and/or postweaning develop-ment (Delclos et al., 2001; You et al., 2002a; Masutomiet al., 2003; McClain et al., 2007). Furthermore, a shorteranogenital distance, adjusted for body weight, wasreported in F1 male and F1 and F2 female rats on PND2 in a multigeneration study that included exposuresduring prenatal and postnatal development at dietarydoses of 51 mg/kg bw/day (Delclos et al., 2009). Incontrast, longer anogenital distance at birth was reportedfor female rats exposed to a diet containing both genisteinand daidzein (16 mg genistein and 14 mg daidzein per100 g of feed) during gestation, lactation, and postweaningdevelopment (Casanova et al., 1999).
Most studies reported no effect on age at preputialseparation at dietary doses up to 180 mg/kg bw/daygenistein in offspring (Casanova et al., 1999; Delcloset al., 2001; Masutomi et al., 2003; Takagi et al., 2004)or 100 mg/kg bw/day by gavage (Nagao et al., 2001)following direct or indirect exposure during the gesta-tional, lactational, and/or postweaning periods. How-ever, a dietary dose of 84 mg/kg bw/day genistein(in offspring) enhanced the effect of methoxychlor indelaying preputial separation when administered GD1 -PND 21 (You et al., 2002a).
Effects of developmental exposure to genistein on pupweight were variable; however, few studies reported anassociation with lower fetal body weight (McClain et al.,2007), lower body weight of male and/or female pupsduring lactation (Dalu et al., 2002, 2008), and lowerpostweaning or adult body weight or weight gain inmales and/or females (Awoniyi et al., 1998; Delclos et al.,2001; Nagao et al., 2001; Masutomi et al., 2003; Nagaoet al., 2008) at oral doses of 35 to 180 mg/kg bw/daygenistein.
Mammary gland development and carcinogenesisMouse: No effects on mammary growth or differ-
entiation in adult mice were reported following gavageexposure of their dams with up to 10 mg/kg bw/dayduring mid gestation through lactation (Fielden et al.,2002). Two studies examined the effects of genisteininjection in CD-1 mice. In a single dose study, Hilakivi-Clarke (1998) administered 0.7 to 0.8 mg/kg bw/day onGD 15 to 20 to pregnant dams and measured morpho-logical changes in the mammary gland at variouspostnatal times in the female offspring. These investiga-tors found significantly higher mammary gland epithe-lial area on PND 35 (but not PND 25 or PND 46; and apersistently higher density of terminal end buds whenmeasured on PND 35 and 45. Padilla-Banks et al. (2006)administered 0, 0.5, 5, or 50 mg/kg bw/day on PND 1 to5 and reported higher mammary gland ductal elongationat 6 weeks. The physiological relevance of effects theyobserved at the highest exposures 50 mg/kg bw/day isuncertain. However, lower numbers of terminal endbuds (at 6 weeks) reduced alveolar development (LOEL5 mg/kg bw/day), and increased mammary gland PRand ERb and lower ERa expression levels also werereported (occurred at LOEL 0.5 mg/kg bw/day). Neitherof these studies included assessments of mammarycarcinogenesis but higher numbers of terminal end budsis generally associated with an increased susceptibility to
mammary neoplasia (Hilakivi-Clarke et al., 1999a). Thechanges reported in the normal mammary glands at thehigher doses and later exposures by Padilla-Banks et al.(2006) are generally associated with a reduced suscepti-bility to mammary carcinogenesis. The use of singledoses and the route of administration (injection) arelimiting factors for several of the studies noted above.
Rat: Hypertrophy/hyperplasia of mammary struc-tures was reported following dietary genistein exposureduring periods including mid-to-late gestation and/orthe neonatal stage, at doses Z100 ppm in males(Z5.7 mg/kg bw/day in dams and 7–12 mg/kg bw/day in offspring) (Delclos et al., 2001; Latendresse et al.,2008, 2009) and 1250 ppm in females (Z83 mg/kg bw/day in dams and 180 mg/kg bw/day in offspring;Delclos et al., 2001; Takagi et al., 2004). Higher ductalbranching of the mammary glands was also reported in22 day-old male rats that had been exposed to 84 mg/kgbw/day genistein (dose in offspring) from GD 1 to PND22 (You et al., 2002a).
Smaller numbers of terminal end buds/ducts andgreater numbers of lobules in mammary glands werereported in adult female rats that received genistein by scinjection during development (Lamartiniere et al.,1995a,b; Murrill et al., 1996). Inconsistent effects onmammary structures were observed in adult rats thatwere exposed to genistein through diet during thedevelopmental period, with one study reporting lowernumbers of terminal end buds and lobules (Fritz et al.,1998) and another study reporting no effects onmammary structures of females (You et al., 2002b).Numbers of chemically induced mammary tumors werelower in rats sc treated during postnatal developmentwith 500 mg/kg bw/day genistein (Lamartiniere et al.,1995a,b; Murrill et al., 1996). In the only oral dose studyexamining the effects of genistein exposure on chemi-cally induced mammary tumors, dietary exposure toZ25 ppm genistein (B2.2 mg/kg bw/day) during gesta-tion and lactation reduced dimethylbenzanthracene-induced tumorignicity in adult females (Fritz et al., 1998).
Males: A multigenerational, dose response study inmale rats reported mammary gland hyperplasias at thetermination of the study on PND 140 in both the F1 andF2 generations (LOEL 5 100 ppm; Latendresse et al.(2009). Up to the highest dose tested (500 ppm; all dosesadministered p.o.) no effects were reported with respectto mammary gland neoplasia or ductal hyperplasia. Youet al. (2002) reported greater branching and numbers ofterminal end buds and lateral buds in the mammaryglands of male offspring (dams exposed orally to 0, 25, or800 ppm during pregnancy and lactation) that were ofborderline significance (p 5 0.06) at 300 ppm and signi-ficant at 800 ppm exposure (You et al., 2002b). Dietaryexposure from GD0 through gestation and lactation(offspring on same diet after weaning) to 0, 300, or800 ppm genistein produced greater glandular size(LOEL 5 300 ppm) in male offspring (Wang et al., 2006).
Females–GD0 through lactation exposures: Fritz(1998) reported no effects on chemically induced mam-mary carcinogenesis in rats exposed to dietary genistein(0, 2.2, 22 mg/kg bw/day during pregnancy and lacta-tion). Exposure (parenteral) to 1.5 mg/kg bw/day on GD15 to 20 (but not 0.5 mg/kg bw/day) was associatedwith a higher incidence of mammary tumors (DMBA;B40 mg/kg bw; PND 50) and greater mammary gland of
455NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

total ER binding (measured on PND 50) fHilakivi-Clarke, 1999 x173g. Hilakivi-Clarke f, 2002 x31g adminis-tered 1 to 1.5, 10 to 15, or 20 to 30 mg/kg bw/day duringpregnancy and lactation and reported lower numbers oflobules, higher numbers of terminal end buds, andhigher numbers of mammary tumors (DMBA; B50 mg/kg; PND 47) only with the highest dose. In a single doselevel study (0.1 mg/kg bw/day, sc, GD 15–20), Hilakivi-Clarke (1999b) reported a higher incidence of mammarytumors in F1 offspring (DMBA; B50 mg/kg; PND45)associated with a higher number of mammary ERbinding sites (measured on PND 35) and lower proteinkinase C activity (PND45 but not PND 21)–experiment 1;and a higher incidence of mammary tumors and shorterlatency–experiment 2.
Females–postnatal exposures: Subcutaneous admin-istration of 10 mg/kg bw/day genistein on PND 2 to 8showed evidence of lactation with cystic ductal dilation,atypical epithelial hyperplasia, microcalcifications, andin situ ductal carcinomas (two of five animals examined)in a study reported by Foster et al. (2004). Hilakivi-Clarkeet al. (1999b) reported lower tumor multiplicity (DMBA;B50 mg/kg; PND45), greater lobular development,lower TEB density, increased alveolar bud densityfollowing parenteral administration of genistein (LOEL0.7–2 mg/kg bw/day). The use of single dose levels, theroute of administration (parenteral), and the occasionaluse of small numbers of animals are limiting factors formany of the studies noted above.
Studies of genistein/soy exposure conducted inrodents were largely designed to test hypotheses relatedto how the physiology of the normal mammary glandmight be modified to affect susceptibility to mammarycarcinogenesis. Few were designed to test a standardtoxicological hypothesis and with the rigor requiredfor detailed toxicological analysis, e.g. dose responsefeatures are often not well represented among the studydesigns. The route of administration and dosing variessignificantly across studies, with some using exposuresor regimens that are not readily seen to be relevant tohuman exposures. Hence, many studies were eitherexcluded or judged to be of limited utility.
Despite the limitations, there are some potential trendsevident in the studies. The potential for an effect of thetiming of exposure on outcome is a relatively consistentobservation when viewed across all studies. Althoughthe precise role of each developmental time of exposureis unclear, there is evidence suggesting that in uteroexposure only may increase later susceptibility tomammary carcinogenesis, while later exposure (birth toonset of sexual maturity) may be protective. Exposureonly during normal adulthood (after sexual maturation)may have no major effect on susceptibility to carcinogen-esis in the mammary gland. The limited data on cross-generational effects are also interesting but insufficient toallow for any definitive conclusion.
It is difficult to derive any overall conclusion on a clearassociation of soy exposure with mammary carcinogenesis.When compared with the limited human data from themeta-analyses, the animal model data support the poten-tial for a modestly protective effect for some exposures,e.g. childhood/adolescent exposure might have a smallreduction in risk. A protective effect of exposure in pre-existing breast cancers is potentially more problematic. Thelimited but possibly more consistent data from in vitro and
in vivo studies using low (perhaps more physiologicallyrelevant) exposures in breast cancer models show thatgenistein can act as an estrogen. Consistent with thisactivity, a small number of studies suggest that genisteinmay reduce responsiveness to antiestrogens or aromataseinhibitors. These data likely explain the increasing practiceamong medical oncologists to advise against soy/isofla-vone consumption for breast cancer patients, as furtherreflected in the guidelines proposed by the AmericanCancer Society (see http://www.cancer.org/docroot/ETO/content/ETO_5_3x_Soybean.asp). For the moment,this would appear to be a reasonable precaution.
Brain and behaviorRat: Treatment with genistein has been reported to
increase the size of the sexually dimorphic nucleus(SDN) of the preoptic area, as quantified by Nisslstaining, in adult female rats. Lewis et al. (2003) observeda greater size of the SDN in adult females following scadministration of 2 mg genistein/kg bw during PND 1 to6 (day of birth was PND 1) followed by oral consumptionof 40 mg genistein/kg bw/day (LOEL and NOEL: 40 and4 mg/kg bw/day oral, respectively) from PND 7 to 21.The dose of genistein increased the SDN in femalesrelative to control; however, it was not as effective as ahigh dose of DES (10 mg/kg bw/day) which produced afemale SDN equal to the size of the control male (Lewiset al., 2003). Faber (1991, 1993) conducted two studies inwhich the SDN was examined (Faber and Hughes, 1991,1993). In the first study, pups were injected (sc) fromPND 1 to 10 with 0, 0.1, or 1 mg of genistein, includingpositive controls DES and Zearalenone. Both DES and1 mg genistein (LOEL 12 mg/kg bw/day oral) increasedthe size of the SDN in females to that of males (Faber andHughes, 1991). In the second study (Faber and Hughes,1993), a range of lower doses (0, 0.001, 0.01, 0.1, 0.2, 0.4,0.5, or 1 mg sc) were administered daily from PND 1 to10. In this study, only the two highest doses of genistein(0.5 and 1 mg; corresponding to dosages of 58 and117 mg/kg bw/day sc, respectively) increased the SDNin adult females. The only other study that simply usedNissl staining to examine the SDN did not find anyeffects of much higher doses of genistein In contrast, scinjections of genistein at 5 or 25 mg (corresponding to 15and 75 mg/kg bw/day) did not affect the size of theSDN; however, the sc injections were given to gestatingdams on GD 16 to 20 so the level of genistein the embryoswere exposed to is unclear (Levy et al., 1995).
Another way to visualize the SDN is by usingimmunocytochemistry to stain the region for Calbindind28k. Scallet et al. (2004) fed dams a casein-based dietsupplemented with 0, 5, 100, or 500 ppm genistein(corresponding to 0, 0.31, 5.7, and 34 mg/kg bw/day;LOEL40.31 mg/kg bw/day oral) from conceptionthrough lactation, and the F1 offspring were continuedon these diets until examination at PND140. Maleoffspring exposed to the genistein treatments had a largerSDN as compared with the control males, while there wasno effect of treatment in female offspring. Patisaul et al.(2007) injected pups on PND 1 to 4 with 0.25 mg ofgenistein (LOEL B100 mg/kg bw/day) and found higherCalbindin-SDN volumes in genistein-treated males inadulthood. In contrast, to the Calbindin result, there wasno effect of genistein on the Nissl-stained SDN in thefemale rats relative to controls. These two studies (Scalletet al., 2004; Patisaul et al., 2007) used Sprague-Dawley rats
456 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

that were fed a base diet of soy-free diet. All the studies inthe previous paragraph involved rats fed standard diet(which contains phytoestrogens and soy), except onestudy (Lewis et al., 2003) that referenced diet R&M No 3(Special Diet Services, Stepfield Witham, Essex) containing100 to 110 ppm genistein. In addition, the other studiesused different rat model; CD (Faber and Hughes, 1991,1993; Levy et al., 1995) and Alderley Park rats (Lewis et al.,2003). Thus, strain/stock and diet might play major roleson the differentiation of the SDN and its sensitivity togenistein. In addition, the data suggest that the classical‘‘biomarker’’ of the SDN, calbindin d28k, is not alwaysinformative for the responses of all the neurons in theSDN. Finally, the finding that genistein exposure canincrease the SDN in males but not in females is not aspredicted if genistein was acting only as an estrogenagonist and speaks to a complexity in the actions ofgenistein.
Another SDN that has been examined is the ante-roventral periventricular nucleus of the hypothalamus(AVPV), which is typically larger in females than inmales. Patisaul et al. (2006) evaluated the AVPV bymeasuring tyrosine hydroxylase (TH) levels by immu-nocytochemistry. Pups received a total of four injectionssc of genistein (0.25 mg; LOEL B100 mg/kg bw/day sc)with one injection administered every 12 hr for the first 2days after birth. On PND 19, the brains were collectedand TH-positive cells were analyzed in the AVPV.Treatment lowered the number of TH cells co-expressingERa in the female brain closer to male cell numbervalues, while genistein increased the number of TH cellsin the AVPV of males closer to female values. These datasuggest that when endogenous estrogens are present (inthe PND 1–2 male) genistein has an anti-estrogenic effect.
Patisaul et al. (2009) reported there was no effect ofgenistein on the number of kisspeptin (KISS) immunor-eactive cells in the AVPV of adult males followinginjections of genistein (10 mg/kg) from PND 1 to 4.In another study (Bateman and Patisaul, 2009), femaleLong Evans rats were treated for the first 4 days afterbirth with estradiol, genistein (same dose as above),either an ERa- or ERb-specific agonist, or equol. Alltreatments were in oil given by sc injections. Inadulthood, females were ovariectomized and givensequential estradiol then progesterone to promote LHrelease. The numbers of co-labelled cfos/GnRH cells waslower in the early genistein-treated females than controls.In addition, kisspeptin immunoreactivity was lower inthe AVPV by the early genistein treatment. The ERbagonist had less of an effect on these measures than theERa agonist. This suggests that genistein can have itsactions via ERa.
Effects of genistein on other aspects of the hypotha-lamic–pituitary–gonad axis have been examined by fewstudies. Masutomi et al. (2004b) fed pregnant rats a dietcontaining genistein (0, 20, 200, and 1000 ppm; NOELrange of 66.6 to 113.1 mg/kg bw/day oral) from GD15until PND10. When pituitaries were analyzed forgonadotropin subtypes at 11 weeks of age, no effect ofgenistein was found. In another study, pregnant damswere fed a diet of 0 or 1000 ppm genistein (LOEL113.1 mg/kg/bw/day oral) from GD15 to PND10. OnPND10, the hypothalami (including the SDN) werecollected and RNA was used for real time RT-PCRdeterminations of PR, ERa, ERb, and SRC-1 (Takagi
et al., 2005). There were sex differences in ERa(females4males) and PR (males4females), but genisteinhad no effect on any of the measures.
Other endpoints and mechanistic studies. Onestudy reported lower thymocyte subsets and changes innatural killer cell activity in rats on PND 22 followingdietary exposure of dams during gestation and lactation(Guo et al., 2002). A second study found changes inthymocyte numbers suggesting augmented cell-mediated immunity in PND 70 rats the dams of whichhad been given dietary genistein during pregnancy andlactation (Klein et al., 2002). The inconsistency in the datadetracts from the utility of the developmental immuno-toxicology data set.
Most of the mechanistic studies used high sc doses.The most widely studied mechanistic effect was expres-sion of estrogen, progesterone, and androgen receptors inreproductive organs of rodents. In studies with gesta-tional and lactational exposure of dams, effects onoffspring were only observed with sc dosing. LowerERa and androgen receptor and increased progesteronereceptor expression were observed following sc injectionof rats with 500 mg/kg bw/dose on 3 days during thelate lactation period (Cotroneo et al., 2001). In mouseovary, increased ERa expression was noted at lowerdoses (r10mg/day) and lower expression was noted at ahigher dose (100mg/pup/day) following neonatal scexposure (Jefferson et al., 2002).
Two studies in which mice were sc injected withgenistein in the neonatal period reported reductions inexpression of testicular ERa (Z7 mg/kg bw) and andro-gen receptor (Z71 mg/kg bw/day) (Adachi et al., 2004),but no effect was reported following maternal dietarygenistein exposure during gestation and lactation at upto 10 mg/kg bw/day (Fielden et al., 2003). In a studyexamining ER expression in rats exposed through dietfrom gestation through weaning or adulthood, resultswere somewhat variable in different generations andoften not dose-related, but lower expressions were notedfor ERa (Z25 ppm) and ERb (Z100 ppm) (Dalu et al.,2002). A transgenic mouse model has been engineered toexpress luciferase activity as a reporter of ER transcrip-tional activity, and studies have shown that oral gavageof genistein to dams on PND4 results in lower luciferaseactivity in the male offspring confirming lactationalexposure to genistein is able to activate ER (Montaniet al., 2008, 2009).
Results of estrogen or progesterone receptor expres-sion in mammary gland following oral or sc exposure inrats were variable, with no obvious patterns related todose or period of exposure observed (Cotroneo et al.,2002; You et al., 2002b; Cabanes et al., 2004). One series ofstudies was interpreted by authors as suggesting thatacute sc exposure of immature animals to genistein500 mg/kg bw results in increased differentiation ofimmature terminal end buds, leading to a greaternumber of lobules, thought to be more resistant tocarcinogens, during adulthood (Lamartiniere, 2000). Itappeared that the effects were mediated through ERs,which regulate both progesterone receptor and EGFreceptor. Upregulation of EGF receptor (EGFR) inimmature rats does not occur through tyrosine phos-phorylation. EGFR is downregulated in adult rats, and ithas been hypothesized that a less active EGF-signalingpathway in adulthood suppresses mammary cancer
457NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

development. A third study reported upregulatedexpression of BRCA1, a tumor suppressor gene involvedin DNA damage repair, following sc exposure of rats togenistein during the lactational period (Cabanes et al.,2004).
Experimental Animal Studies of Soy Formulaor other Soy Exposures During Development
Almost 30 experimental animal studies are summar-ized in Table 10, one study was judged to have highutility and the rest were found to be of limited utility forthe evaluation process.
Growth. The effect of exposure to dietary soy onpup body weight was inconsistent among studies. In astudy by Masutomi et al. (2004a) using Sprague-Dawleyrats, female offspring in the soy-diet group showed aincreased body weight on PND 21, greater body weightgain between PND 10 and 21, and shorter weight-adjusted anogenital distance compared with the controlgroup. Male offspring in the soy-diet group showed anincreased body weight on PND 2 and greater bodyweight gain between PND 10 and 21. In this study,exposure to dietary soy was likely limited to the damssince they were given a soy-based or a soy-free diet untilPND 10, before the time when pups will have eaten solidfood. Increased pup body weights before weaning werealso reported in male rats by Akingbemi et al. (2007), andin both female and male rats by Odum (2001). In thestudies by Akingbemi et al. (2007) and Odum et al.(2001), rats were exposed to soy during gestation andlactation. The body weight was greater in male offspringon PND 5 and PND 21 (Akingbemi et al., 2007), and inboth male and female offspring from birth to PND 13(Odum et al., 2001).
When body weight was measured after weaning, theopposite effect was observed with numerous studiesreporting a decreased adult body weight associated withdevelopmental exposures to soy-based diets (e.g. de-creased body weight in male and female rats at PND 50and PND 120 (Lund et al., 2001), decreased body weightin male rats at PND 75 (Lephart et al., 2001), decreasedbody weight in male rats at PND 90 (Atanassova et al.,2000), decreased body weight in male rats at PND 280(Gorski et al., 2006). Odum et al. (2001) observedincreased body weight in male and female rats frombirth to PND 13, no effect around weaning (i.e. PND13–21), and decreased body weight in males from PND26 to PND 128 and in females from PND 26 to PND 140when a standard soy-based diet was given to the damsduring gestation and lactation, and then to offspringpostnatally. Ruhlen et al. (2008) reported the samepattern in CD-1 mice, with increased birth weight, noeffect of body weight around weaning (PND 20 and 26),and decreased body weight with decreased renal andgonadal fat pad weights at PND 90 for F1 offspring ofCD-1 mice exposed to a soy-based diet when comparedwith a animal fed a soy-free diet.
There are also several studies that report no change inadult body weight following developmental exposure toa soy-based diet in rats (McVey et al., 2004b; Akingbemiet al., 2007; Pastuszewska et al., 2008). Mardon et al.(2008) reported body weight in rats given a soy-baseddiet with observations covering longer exposure periodsthan most studies, and including data on 2-year-old rats.
There was no effect of soy consumption on body weightin rats from birth to 18 months of age; however, there wasa increased body weight and body fat mass observed in24 month-old rats when a soy-based diet was given to thedam during gestation and lactation, and/or to the pupsfrom weaning to 24 months of age (Mardon et al., 2008).
There was no effect of soy intake on body weight atPND 35 to 45 in marmoset monkeys given soy-basedformula from days 4 or 5 until weaning at PND 35 to 45compared with controls fed a cow’s milk-based formula(Sharpe et al., 2002; Tan et al., 2006). There was also noeffect on body weight from birth to 6 months of age incynomolgus monkeys when a soy-based diet was givento pregnant monkeys and the offspring were fed thesame soy-based diet (Wagner et al., 2009). In contrast,body weight was decreased in older cynomolgusmonkeys (1 and 2 years of age) exposed to the soy-baseddiet developmentally and with continued maintenance(Wagner et al., 2009).
Female: In all experimental animal studies, exposureto soy proteins was via the oral route. However, findingswere inconsistent between the studies. In the mouse, onestudy reported an acceleration in the age at vaginalopening (Guerrero-Bosagna et al., 2008). In anotherstudy, Ruhlen et al. (2008) reported a delay in the ageat which the female mice produced their first litter. Bothstudies involved prenatal and postnatal exposure to soyproteins in the feed. Body weight was decreased in bothstudies, and there was less fetal circulating 17b-estradiol(Ruhlen et al., 2008). In addition, estradiol-stimulateduterine weight was decreased in PND20 females on thehigh-soy diet relative to those on the control diet.
In Wistar and Sprague-Dawley rats, exposure to soy inthe feed during the prenatal and postnatal periods waslinked to early vaginal opening, greater body weightsand uterine weights (Odum et al., 2001) relative to rats onsoy-free diets, with the exception of the females on theGlobal 2016 diet (soy-free) in which the age of vaginalopening onset was not different than in the soy-fedcontrols (Odum et al., 2001). Body weight was decreasedand vaginal opening onsets were delayed, relative tocontrols, in Long Evans rats exposed to isoflavones in thediet during the prenatal and postnatal period (Lundet al., 2001) and 264). Circulating 17b-estradiol serumlevels were increased in neonatal rats exposed via thedam on PND 5 to 10 (Liu et al., 2008).
The expert panel noted that due to inconsistentfindings it is difficult to draw conclusions about effectsof soy protein on female growth, reproductive system,and endocrine-related endpoints. However, a consistenteffect of genistein and soy protein in female mice andrats was an acceleration of the timing of vaginal opening.This finding may have clinical relevance for timing ofpuberty in girls.
Male: Several rodent studies compared reproductiveendpoints in male offspring of dams given soy-based andsoy-free diets during pregnancy and lactation with pupsweaned to their dam’s diet (Makela et al., 1995; Lundet al., 2001; Odum et al., 2001; McVey et al., 2004a,b). Theresults reported in these studies were inconsistent.Odum et al. (2001) reported advancement of pubertallandmarks in males (and females). Testis weight wasreported to be decreased at PND 90 to 95 by dietary soyin one study (Atanassova et al., 2000) and increased bydietary soy in rats at PND 28 (McVey et al., 2004b).
458 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Increased testicular weight was reported in 68- and128-day-old offspring of rats given a soy-based dietduring pre- and postnatally; epididymal weight was alsoincreased at PND 68 (Odum et al., 2001). Greater weightof the testes, epididymides, and seminal vesicles werealso reported in adult offspring (i.e., at PND 90) ofpregnant mice fed a soy-based diet during gestation,lactation, and given to offspring post-weaning (Ruhlenet al., 2008). Greater testicular and epididymal weightswere reported at PND 160 in male offspring of rats fed asoy meal-containing diet; however, testicular and epidi-dymal weights were not altered in older (i.e. PND 280) oryounger rats (PND 22 or 60) from the same study (Gorskiet al., 2006). Several studies have reported decreasedadult prostate weight in mice (Ruhlen et al., 2008) or rats(Lephart et al., 2001; Lund et al., 2001) exposed to soy- orphytoestrogen-based diets pre- and postnatally whencompared with animals raised on a soy-free diet. Otherstudies have reported no change in prostate weightfollowing exposure pre- and postnatally in rats (Gorskiet al., 2006; Akingbemi et al., 2007) or during lactation inmarmosets (Tan et al., 2006). These differences mayreflect different isoflavone contents of the diets used indifferent studies or other differences in the compositionof the feed. The studies of McVey et al. (2004a,b) permit adose–response evaluation of dietary isoflavone supple-mentation and developmental effects. These investiga-tors used a soy-free diet to which was added soy proteinand soy isoflavones at five exposure levels (plus thesoy-free diet control). The most sensitive endpoint wasincreased testis weight on PND 28 in rats that wereexposed during pregnancy, lactation, and weaning to adietary-isoflavone intake Z2 mg/kg bw/day; however, itis not known if this is related to changes in spermatogen-esis. The increased testis weight did not show a dose–response relationship.
The study of Sharpe et al. (2002), extended by Tan et al.(2006) involved the feeding of marmoset co-twins witheither a soy infant formula or a cow-milk formula. Theauthors expected the use of co-twins to permit pairedanalyses and to decrease variability between animalsassigned to different treatments. The study authorsreported decreased levels of plasma testosterone onPND 35 to 45, increased testis weight, and increasednumbers of Sertoli and Leydig cells at 120 to 138 weeks ofage. Interpretation of this study was complicated bydesign since the offspring were also nursed by theirmothers on a daily basis.
Mammary gland development and carcinogen-esis. Thomsen et al. (2006) fed FVB mice either controldiet, 270 mg isoflavone/kg diet throughout lactation(PND 0–21), or 270 mg isoflavone/kg diet from weaningthrough the end of the study. Effects in both isoflavone-treated groups included increased branching in mam-mary trees and branching in juvenile mammary glands.In the weaning through study termination group, thenumbers of terminal end buds during the early post-weaning phase were initially increased, but weresignificantly decreased by PND 42 to 43. An increase inthe number of terminal end buds would be expected toincrease susceptibility to carcinogenesis but the laterreduction would eliminate/reverse this effect. Thus,timing of carcinogen exposure would likely affect theapparent estimates of changes in the mammary gland’ssusceptibility to carcinogenesis.
Hakkak et al. (2000) fed Sprague-Dawley rats AIN-93Gdiets in which soybean oil was replaced by corn oil andthe protein source was either casein, whey, or soy-proteinisolate containing isoflavones 430 ppm diet (genistein276 ppm diet or B20.4 genistein mg/kg bw/day anddaidzein 132 ppm diet). At weaning, the F1 and F2offspring were fed the same diets as their dams andcontinued to receive the diets throughout their lifetimes.Accelerated vaginal opening was observed on days PND37 vs. PND 38. At PND 50, rats were orally treated bygavage with 80 mg/kg DMBA and the development ofpalpable mammary tumors assessed. The authors re-ported a shorter latency to develop mammary glandtumors (F1 and F2) and a greater number of tumors/ratcompared with the whey diet group (F2). Also inSprague–Dawley rats, Simmen et al. (2005) fed femaleF1 offspring of F0 parents fed AIN-93G diets containingeither casein or soy-protein isolate from GD 4 to lactation,at weaning females were weaned to the diet of the dam.At PND 50, rats iv injected with 50 mg/kg MNU andfollowed until 115 days following MNU treatment.The investigators reported a lower mammary tumorincidence (rats with at least one mammary gland tumor)and a longer tumor latency in rats fed the soy-proteindiet. Although the incidence of ductal carcinoma in situwas lower, a higher incidence of infiltrating ductalcarcinoma was observed.
Three primary tumorigenesis endpoints are usuallymeasured. Latency (time to appearance) and incidence(number of animals with one or more mammary tumors)are likely reasonable measures with respect to the humandisease. Multiplicity (number of tumors/animal) may beless widely applicable but is potentially relevant ascontralateral breast cancers arise in some women.Although the data from the rodent studies are mixed intheir general outcomes with respect to changes inmammary gland differentiation and cancer susceptibility,several of the exposures appear to induce changes thatincrease susceptibility to carcinogenesis. These may beestrogenic effects, as E2 can produce the same outcomesand other markers of estrogenicity are evident, e.g.accelerated vaginal opening and increased the terminalend buds. Although the weight of evidence suggestadverse effects from exposure, it is difficult to arrive at acompelling determination of the lifetime effects onmammary cancer risk. Some of the changes that couldaffect risk are time dependent and modifications thatcould either increase or decrease risks are reported.
Brain and behavior. In three studies, the mainmanipulation was phytoestrogens in the diet. The onlybehavioral study was conducted in Long Evans rats(Lund and Lephart, 2001) with life-long (starting beforeconception) exposure to phytoestrogen-free or phytoes-trogen-containing (600 mg/g) diet. As adults, males andfemales were tested in the elevated plus maze foranxiety-like behavior. Rats on the phytoestrogen-contain-ing diet were less anxious and more active in the mazethan the rats not exposed to phytoestrogens. Lephartet al. (2001) used the same stock and all rats wereingested with phytoestrogen-containing diet (600 mg/g).At PND 80, one half of the animals on the phytoestrogendiet were switched to phytoestrogen-free food on PN 80then all rats were killed on PN 120. The AVPV wasexamined by Nissl stain and the expected sex difference(female4male) was only noted in the animals that
459NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

remained on phytoestrogen-containing diet. Notablymales moved to phytoestrogen-free diet had larger AVPVvolumes than the control males that remained onphytoestrogen-containing diet. This demonstrates thatthe neuronal changes can occur in the adult brain in theAVPV when phytoestrogens are eliminated from diet, butthis occurred only in males. Finally, another sexuallydimorphic region, the SDN, was examined in Sprague-Dawley rats by Taylor et al. (1999). Dams were placed onphytoestrogen-free or phytoestrogen-containing (200mg/g)diet when mated and fetuses were taken on GD 20.Western blots were used to quantify calbindin in themedial hypothalamus (most of the calbindin in this regionwould be in the SDN). The normal sex difference(male4female) was noted only in fetuses from thephytoestrogen-consuming dams. On the phytoestrogen-free diet, females had significantly more calbindin proteinthan the phytoestrogen-ingesting females. Neither of thesesex differences is related to the expression of anxiety.Interestingly, both of these classically described neuraldifferences require phytoestrogens in diet.
CONCLUSIONS
* Evidence is sufficient to conclude that genisteinproduces developmental toxicity in male and femalemice and rats. The panel considered oral or parenteralexposure between birth and weaning at dose levels(between 37.5 and 50 mg/kg bw/day in mice;between 25 and 100 ppm in rats) that produce bloodlevels similar to those reported in children. In femalemice dosed via the oral or parenteral routes, adverseeffects were manifested as histomorphologicalchanges of the reproductive tract and the mammarygland, a decrease in fertility, and/or a decrease innumbers of live pups per litter. In a multigenerationalstudy in rats with dietary exposure to genistein,changes were observed in mammary gland morphol-ogy in both sexes (25 ppm males and 100 ppmfemales). Although not a definitive marker of devel-opmental toxicity, consistent changes in the age ofvaginal opening were seen in mice and rats. Somebiological changes in a discrete brain region in femalerats have been documented at 40 mg/kg bw/day. Theexperimental animal data are considered relevant tothe assessment of human risk.
* Evidence for daidzein, equol, and glycitein is insuffi-cient due to a paucity of relevant studies. However,relative ER binding activity of equol and daidzein arebroadly similar to genistein.
* Evidence is insufficient to conclude that soy infantformula or other soy exposures, including soy-baseddiets, produces or does not produce developmentaltoxicity in experimental animals. Although a fewstudies have examined the developmental effects ofsoy infant formula or other soy exposures and somehave identified potential developmental effects, thesestudies have yet to be replicated. The experimentalanimal data are considered relevant to the assessmentof human risk.
* Evidence is insufficient to conclude that soy infantformula produces or does not produce toxicity withinfant exposure in girls or boys at recommended
intakes manifested by the following endpoints: bonemineral density, gastrointestinal effects, allergy/immunology, thyroid function, reproductive end-points, cholesterol, diabetes mellitus, and cognitivefunction.
* Evidence is sufficient to conclude that use of soyinfant formula in healthy full-term infants does notimpair growth during infancy. This conclusion isbased on a large number of studies of small samplesize that consistently show similar growth trajectoriesof anthropometric measurement.
OVERALL CONCLUSIONS ANDRESEARCH NEEDS
Human Exposure
Infant exposure to isoflavones occurs primarilythrough the consumption of soy formula. The isoflavonestypically found in soy formula are genistein (58–67%),daidzein (29–34%), and glycitein (5–8%) with themajority of isoflavones occurring as glycosides. Totalisoflavone levels found in soy formula worldwide was 10to 47 mg/liter formula (‘‘ready-to-feed’’ equivalent).These levels are two orders of magnitude higher thanthose observed in casein-based formula or breast milk.
The degree to which infants are exclusively fed soyformula versus a combination of soy and nonsoy formulaand/or breast milk is unclear. It is estimated thatupwards of 25% of newborns or infants in the UnitedStates are fed soy formula at some point. Exposure to soyformula also varies depending on developmental stage(e.g. weaning), and cultural variations in soy formulaand soy product usage are known to exist. Prenatalexposure to soy isoflavones via maternal soy intake isalso possible.
Recent sales of soy formula in the United Statesrepresent B12% of the total dollar sales for infantformula (personal communication with Robert Rankin,Manager of Regulatory and Technical Affairs at the IFC,October 13, 2009). U.S. sales data also suggest a 50%reduction in soy formula use over the last decade(public comment from the IFC, received December 3,
2009 and personal communication with Dr. Haley CurtisStevens, IFC). When sales are considered as a surrogatemeasure of actual reported usage, these data providea lower indication of usage compared with otherfrequently cited estimates. The usage and sales of soyformula also varies geographically ranging from 2 to 7%of infant formula sales in the United Kingdom, Italy,and France, to 13% in New Zealand (Agostoni et al., 2006;Turck, 2007),and to 31.5% in Israel (Berger-Achituv et al.,2005).
In the United States, total isoflavone intake by infantswas estimated at 2.3 to 9.3 mg/kg bw/day, depending onage of the infant. The estimated intake for genistein,expressed in aglycone equivalents, ranges from 1.3 to6.2 mg/kg bw/day. These intakes are several orders ofmagnitude greater than those infants who consumebreast milk or a cow’s milk-based formula. Soy for-mula-fed infants have higher daily intakes of genisteinand other isoflavones compared with other populations(excluding regular consumers of soy supplements)(Table 2).
460 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Guidelines regarding the use of soy formula have beenissued by both the AAP (Bhatia and Greer, 2008), theESPGHAN Committee on Nutrition (Agostoni et al.,2006), and reviewed in the literature (Scientific Commit-tee on Food, 2003; Turck, 2007; Thygarajan and Burks,2008). In general the exclusive use of soy formula overnonsoy and/or breast milk is not recommended. Theonly real indications for use are instances where thefamily prefers a vegetarian diet or for the management ofinfants with galactosemia or primary lactase deficiency(rare). Soy formula is not currently recommended forpreterm infants.
Mean blood-based levels of isoflavones in infants fedsoy formulas are considerably higher than other popula-tions, including vegans and Japanese adults (Table 3).For example, concentrations of total genistein in wholeblood samples from U.S. infants fed soy formula are1455 ng/ml at the 75th percentile, ranging from 13.5 to3562.9 ng/ml (Cao et al., 2009) (personal communicationwith Dr. Yang Cao, NIEHS). The geometric mean value oftotal genistein in the infants fed soy infant formula(757 ng/ml) was 53.3 and 70.1 times higher comparedwith levels measured in infants fed cow milk formula(14.2 ng/ml) or breastmilk (10.8 ng/ml), respectively.The maximum obtained value in the Cao et al. (2009)study is approximately 11 times higher than themaximum genistein concentration (325 ng/ml) detectedin plasma in a small study of Japanese men, n 5 6(Adlercreutz et al., 1994). Average blood levels of totalgenistein in the soy formula-fed infants (757 ng/ml) areB160 times higher than the mean levels of total genisteinin omnivorous adults in the United States (4.7 ng/ml)reported by Valentin-Blasini (Valentin-Blasini et al., 2003).
Pharmacokinetics
There are no pharmacokinetic data for individualisoflavones following administration of soy-based infantformula to infants or children. Thus, reliable estimates ofexposure, as defined by AUC, are not available andtherefore preclude meaningful comparisons of exposurebetween infants receiving recommended intakes onnormal feeding schedules and experimental animalmodels. There are currently no estimates of variabilityin exposure to individual isoflavones (genistein, daid-zein, equol, and glycitein), or pharmacokinetic para-meters describing the disposition of those isoflavones,following recommended intakes of soy-based infantformula. Given the heterogeneity of the human infantpopulation, population studies incorporating measuresof systemic exposure (i.e. accurately timed plasmasamples and quantitative urinary recoveries) are essen-tial to identify a potentially susceptible subgroup, if oneexists.
Nevertheless, human data demonstrate that hydrolysisof genistin, the glycoside conjugate of genistein that ispresent in soy formula, occurs in both adults and infantssuch that systemic exposure to genistein occurs (Setchellet al., 1997; Cassidy, 2006; Cao et al., 2009). Themechanism of deconjugation may involve bacterialhydrolysis in the gastrointestinal tract, although somedata suggest that the glycoside conjugate may beabsorbed. Irrespective of the mechanism, systemicexposure to genistein has been documented in soyformula-fed infants, with total genistein concentrations
(isoflavone and conjugates) ranging from 13.5 to3562.9 ng/ml (Cao et al., 2009) (personal communicationwith Dr. Yang Cao, NIEHS). Although it is generallyaccepted that the majority of isoflavone readily availablein accessible body fluids (e.g. blood and urine) is in aconjugated form, the relative proportion of measurableisoflavone that is available as aglycone, glucuronide, orsulfate has not been determined for infants or childrenfed soy-based formula. Thus, the age-related differencesin glucuronidation and sulfation of individual isofla-vones are unknown, and comparison of the relativeexposure to unconjugated forms of isoflavones betweeninfants and adults is not possible.
Equol exposure in human infants following daidzeinintake is relatively low compared with animals of acomparable developmental stage. This observation isrelevant for the risk assessment of daidzein, but not ofimportance to the risk assessment of soy formula due tothe detection of equol in infants independent of feedingtype.
Genistein, daidzein, and equol all have weak affinityfor ERs, when compared with an endogenous agonistsuch as estradiol, and the ability to activate ER-dependent transcription. There is also evidence suggest-ing that many of the developmental and reproductiveendpoints observed in experimental models could bemediated through ER-dependent mechanisms given thecentral role of ERs in modulating these systems andthe similarity of effects observed as compared with thosefound in response to estradiol. However, there arelimited studies to date that have definitively establishedthe essential role of ERs in relevant toxic endpoints(e.g. insufficient studies using null mice, siRNAs, etc.).One study has demonstrated that ERb is required forgenistein-induced multioocyte follicles in C57BL/6 mice,as this effect is not found in genistein-treated ERb-nullmice (Jefferson et al., 2002). However, this study alsodemonstrated that inhibition of tyrosine kinase activityby genistein could also contribute to the mechanisms ofother effects resulting from genistein exposure in thismodel. This finding is consistent with gene expressionprofiling studies showing that the signaling mechanismsinduced by selective ER agonists are substantivelydifferent than those resulting from treatment withisoflavones. This suggests that other receptor-dependentand/or receptor-independent (e.g. epigenetic) mechan-isms could contribute to the etiology of isoflavone-induced toxicities.
Developmental Hazards
Humans. With the exception of conclusions forgrowth (discussed below), the overall evidence wasconsidered insufficient to reach a conclusion on whetherthe use of soy infant formula produces or does notproduce developmental toxicity with infant exposure ingirls or boys at recommended intake levels. Commonlyencountered limitations included the nonrandom orunspecified method of assignment to feeding groups,the use of self-selected breast- and formula-feedingmothers, failure to control for the reasons for which soyformula was used, and the early and inconsistentintroduction of solid foods. Studies that comparedoutcomes in children randomized to soy or cow-milkformula were considered the most reliable, particularly
461NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

when parents and outcome assessors were masked toformula assignment. The studies also were evaluatedbased upon adequate sample size by gender and feedinggroup, the presence or absence of longitudinal follow-up,validation of exposure to soy formula, and appropriateadjustment for potential confounding variables. Theevidence was considered sufficient to conclude that theuse of soy infant formula in healthy full-term infantsdoes not impair growth during infancy. This conclusionis based upon a large number of studies of small samplesize that consistently show similar growth trajectories ofanthropometric measurements.
The expert panel had several remarks related to thequality of the available information for specificendpoints:
* Full-term healthy infants fed soy formula havecomparable growth as compared with full-term,healthy infants fed breast-milk or cow-milk formula.Soy formula feeding may not support the growth ofpremature infants and causes an increased incidenceof rickets. Since the AAP does not recommend the useof soy formula for premature infants, current litera-ture on the subject is scant.
* Soy infant formula may or may not cause reproduc-tive toxicity in boys and girls based on currentevidence. Preliminary data addressed in the panel’sreport do not allow firm conclusions on this effect.Limited retrospective data suggest that soy formula-fed infants may demonstrate premature thelarche(the start of breast development at the beginning ofpuberty) (Freni-Titulaer et al., 1986). The only otherstudy considered to have limited utility that includedreproductive parameters did not have sufficientpower to rule out increased risks (Strom et al., 2001).
* Soy infant formula may or may not cause adverseeffects on thyroid function in male or female infantsand children. A special cohort of infants and childrenwith CH-fed soy infant formula demonstrated a delayof TSH levels to return to normal after adequatetreatment; these children may need increased doses oflevothyroxine and closer follow-up. However, thestudies that specifically targeted infants and childrenwith CH were case-studies, which results in limitedinferences.
* All studies of gastrointestinal effects reviewed for thisreport were classified as having no utility. However,extensive reviews by AAP and ESPGHAN havereported adverse effects in a subset of infants withdocumented cow milk protein allergy; infants withdocumented cow milk protein-induced enteropathyor enterocolitis frequently are as sensitive to soyprotein and should not be given soy protein formulas.
* Sensitivity to soy protein during human infancy andchildhood may occur, but it appears to affect a verysmall subset of the population. Villous atrophy wasreported with the use of soy formulas, but theseinferences were of no utility due to the study designsbeing case reports.
* One cholesterol study was reviewed for this report(Cruz et al., 1994) and it was classified as havinglimited utility. The fractional cholesterol synthesisrates were significantly greater in infants fed soyformula (12.02% per day) as compared with humanbreast milk fed infants (2.97% per day). This rate for
soy formula-fed infants was not statistically signi-ficantly different when compared with cow-milk fedinfants. These findings suggest that infants respond todifferent dietary cholesterol intakes through alteredcholesterol synthesis rates.
* One diabetes study was reviewed for this report (Fortet al., 1986). The design of the study was a retro-spective case–control, which examined feeding his-tories of diabetic children versus controls to study theeffect of breast feeding on the development ofdiabetes. Retrospective collection of infant feedinginformation on average of 14 years was used toclassify feeding groups. As a result of this type ofclassification, twice as many diabetic children hadbeen said to have been fed soy formulas comparedwith control. Poor recall is also associated with thelack of information regarding the duration andquantity of soy formula feeding. Furthermore, soyformula exposure was said to have been mixed withcow milk formula exposure. No association betweendiabetes and exposure to soy formula was found. As aresult of these deficiencies, the study was classified asno utility.
* Two cognitive function studies were reviewed for thisreport (Malloy and Berendes, 1998; Jing et al., 2008)and were considered of no utility. One studycompared cognitive performance in children whowere fed soy formula or human breast milk. Afteradjustment of confounding variables, no differenceswere found in the outcome measures. An additionalfactor which makes this study of no utility is thepossibility of recall bias (9–10 year span). The secondstudy compared soy formula-fed infants to cow milkformula-fed infants. Spectral EEG was used to assessbrain development. Duration of soy feeding was onemonth. None of the spectral variables differedbetween the exposure groups. The duration of feedingfor 1 month may not have been sufficient to discerndifferences in EEG patterns.
Experimental animals. Although experiments de-signed to provide information of potential health risks inhumans are ideally conducted in the same species, thereare numerous reasons why this is often not possible orpractical. The purist might then conclude that we cannotdo any relevant research. However, there are numerousexamples of both translational uses of animal studies todevelop targeted treatments and also of pure basicscience work to inform human studies. Animal modelsare considered to be useful in hazard identification sincethey allow investigators to control for potential con-founders and conditions of exposure that cannot beachieved in epidemiology or clinical studies and thuscausal associations and mechanistic pathways can beaddressed. ERs are expressed in key target tissues,regulate estrogen function in a similar manner in bothrodents and humans, and oral administration of phy-toestrogens results in similar circulating levels in bothrodents and humans.
Animal studies have been used for risk assessment andfor the development of new hypotheses about mechan-isms of action. Experiments in mice, rats, pigs, andnonhuman primates are routinely used to assess effectsof various compounds on endpoints including but notlimited to: reproduction, neural development, behavior,
462 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

and toxicity. In the case of soy and isoflavone studies,routes of exposure have included oral and sc adminis-tration. The expert panel tried to preferentially evaluatestudies that used oral administration since it best mimicsthe human route of exposure. However, sc injectionstudies were considered when they used dose levelswhich produced blood levels representative of thosemeasured following human exposure were also consid-ered. Most weight in the assessment was given to thosestudies that exposed developing animals to soy, or inmost cases the specific isoflavone genistein, solely duringthe period from birth to weaning. During this window ofexposure, which approximates the timing of soy formulaingestion in infants, genistein can affect the following indeveloping animals: puberty onset, neural differentia-tion, reproductive tract morphology, and mammarygland organization.
Although the porcine model was seen to haveimportant advantages over rodents as a model of humanexposure because they do not produce equol fromdaidzein, the panel found that there was a paucity ofstudies using the porcine model. Furthermore, althoughhuman infants do not produce comparably highquantities of equol, they are exposed to daidzein whichhas a similar affinity for the ERs. Although it isrecognized that rodents do not produce SHBG, theyproduce other sex hormone binding proteins. Pharmaco-kinetic issues were considered by the panel, and keyoral and sc injection studies were shown to producesimilar blood concentrations and circulating patterns(Doerge et al., 2002; Jefferson et al., 2009a). Takentogether these results suggest that the sc route ofadministration can be used for hazard identificationstudies. The panel notes that all the animal modelsinclude exposure to genistein.
Level of Concern Conclusions
The Expert Panel has minimal concern for adversedevelopmental effects in infants fed soy infant formula.
This conclusion is based on:
* Lack of clarity on whether studies in experimentalanimals treated with genistein only can be extrapo-lated to infants fed soy infant formula, i.e. exposure toa single isoflavone versus soy infant formula
* Interpretation of findings from experimental animalsas demonstrating adverse effects, i.e. advancedvaginal opening, effects on the mammary gland inthe context of interspecies comparisons
* Although there are a large number of experimentalanimal studies published on genistein or soy, there areonly a limited number of studies where experimentalanimals were treated only during the relevant lifestage of birth to weaning. Multigenerational studiesdo not permit discerning effects attributed to gesta-tional or lactational exposure.
* However, few studies in experimental animals andone study in humans reported effects related to thereproductive system and this elevates the concernfrom ‘‘negligible’’ to ‘‘minimal.’’
* Studies of sufficient quality in humans have not beenconducted to address the concerns raised from theexperimental animal findings or to identify pre-viously unrecognized endpoints.
The panel was in favor of the conclusion by a tally of10 yes and 2 no votes (The expert panel had 14 membersbut the Chair, Dr. Gail McCarver, does not vote andDr. Michael Rybak was not present on the day of thevote.).
Dissenting opinions:One committee member, Dr. Jatinder Bhatia, expressed
negligible concern for adverse developmental effects ininfants fed soy infant formula based on a lack of findingin humans and lack of experimental data in animals fedsoy formula.
One committee member, Dr. Ruth Etzel, expressedsome concern for adverse developmental effects ininfants fed soy infant formula. Dr. Etzel supported herlevel of concern conclusion based on the following threepoints: (1) There are sufficient signals of adversereproductive and developmental effects in experimentalanimals to worry about long-term effects on develop-ment of infants; (2) Infants are exquisitely sensitive to theeffects of exogenous chemicals during early life, andnaturally occurring genistein in soy formula has a muchstronger estrogenic effect than non-natural estrogeniccompounds on the developing reproductive system; and(3) the absence of evidence of an effect in human studiesis not the same as evidence of absence of an effect,particularly given the paucity of human data to informthe conclusion.
Critical Data Gaps and Research Needs
Pharmacokinetics. Knowledge deficits and researchneeds can be summarized as follows:
1. Available pharmacokinetic data in human infantsessentially are equivalent to biomonitoring data, andpermit crude contextual assessment of the animaldata. Full pharmacokinetic data are needed in infantsfed soy formula to inform the questions of interest.
2. Although plasma/blood data approximating steadystate have been collected, the temporal relationshipwith feeding is unknown. In the absence of fullpharmacokinetic profiles, accurate timing of blood/plasma sampling relative to feeding would be animprovement over the available data.
3. From a purely scientific perspective, pharmacokineticdata for a range of doses with individual isoflavoneswould generate the most interpretable data withrespect to understanding the dose–response relation-ship between isoflavones and reproductive outcomes.However, this is unlikely to ever be practical orethically possible as isoflavones are administered tothe population of interest, infants, as a complexbiological mixture. Human ethics committees may bereluctant to approve investigations involving com-pounds of uncertain toxic potential, and parents areunlikely to consent to their infants participating insuch a study. Pharmacokinetic studies followingadministration of soy-based formulas may be moreacceptable to both human ethics committees andparents.
4. The adult data indicate that glucuronidation andsulfation are the two primary pathways for isoflavonebiotransformation. However, high-quality quantitativeassessment of their relative importance is not
463NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

available, including the extent of variability ofglucuronidation and sulfation in relevant populations.
5. In vitro data on the enzymes most important forisoflavone disposition in humans are limited. Further-more, the tissue distribution of those isoforms(especially in liver and intestine) and the relativelevel of expression of individual isoforms in targettissues are unknown. This knowledge deficit pre-cludes further investigations of ontogeny as well aspharmacokinetic modeling studies.
6. Studies to firmly establish whether ERs are, or are not,required for isoflavone-dependent toxicity are needed.
7. Studies to determine the relative role of epigeneticmechanisms that may or may not be required forisoflavone-dependent toxicity are needed.
Human epidemiological.
1. Based upon the current state of the science, a basic(but difficult) question to address is, ‘‘If ingestion ofsoy formula results in altered onset of puberty, and/oralteration of reproductive organs, then what wouldthe cumulative exposure of soy formula have to be toobserve such effects?’’
2. Larger (in terms of sample size) and longer (timespan) longitudinal, prospective cohort studies areneeded, e.g. a longitudinal study that capturessoy exposure from birth through puberty. This islikely to be quite challenging due to current indica-tions for soy formula and declining prevalence of use.One opportunity would be to collect appropriatemeasures of soy protein exposure in the NationalChildren’s Study.
3. Human studies should include endpoints that havebeen used in previous studies (growth, bone mineraldensity, and cognitive performance), in addition toreproductive endpoints that reflect the greater timespan of the study, such as onset of puberty. The firstendpoints in the series collected should reflect a‘‘pure’’ exposure, i.e. minimal cross-feeding withbreastmilk or cows milk-based formula.
4. Case–control studies should include longer termendpoints such as breast cancer. Again, longer termendpoints could be quite challenging due to thedietary drift that occurs in the short term that willalmost certainly occur in the long-term.
5. Continued observational studies of thyroid function ininfants fed soy infant formulas are needed.
6. Studies that combine clinical outcomes with objectivemeasures of soy protein exposure are needed.
Experimental animal. Although animal studiesare essential to hazard identification and to addresspotential mechanisms of action, numerous data gapswere identified in this assessment.
1. There is a paucity of studies that examined the effectof soy formula as opposed to individual compounds.Use of full foods is thought to account for potentialinteractions among chemical constituents of theformula and thus is more representative of humanexposure than studies employing single soyphytoestrogens.
2. There are scant studies that examined the effects ofisoflavones other than genistein.
3. Although rodents are widely used in hazard identifi-cation there are important limitations to rats and miceas models for human exposure. Differences incomparative physiology can have important implica-tions to the model selection and the relevance of thedata generated. Of note, rodents unlike human infantsproduce significant amounts of equol from daidzein.The total estrogenicity of soy needs to be morecarefully considered in future studies.
4. The effects of soy need to be examined in the mostrelevant period of development.
5. Activities other than estrogenicity of soy productsneed to be evaluated at exposure concentrationsrepresentative of human exposure.
6. The impact of species differences in bioavailability ofsex steroid and isoflavones may influence the potencyof soy in rodent models and requires furtherinvestigation.
7. There is limited information concerning the effect ofearly life exposures to soy proteins and isoflavones onanimal susceptibility to subsequent chemical insults inlater life. This question addresses the concern thatearly life exposure to isoflavones or soy proteins mayhave a programming effect that alters risk forhormonally dependent diseases such as breast cancer.
8. More studies examining the effects of soy or iso-flavone exposure during infancy on developmentaleffects on puberty are needed. The relevance of somedevelopmental effects to human health, such asvaginal opening (time of puberty onset) in rodents,is uncertain. More studies, including additionalmeasures of puberty (e.g. ovulation, gonadotropinsecretion, and onset of estrus cyclicity) in rodents andhuman studies on timing of sexual maturation, areneeded.
9. The long-term effects of soy or isoflavones duringinfancy should be addressed in animal and humanstudies of fertility, reproductive senescence (meno-pause), and life span.
10. Animal and human studies that assess effects of soyor isoflavones from birth to weaning on nonrepro-ductive behaviors are needed.
REFERENCES
Adachi T, Ono Y, Koh KB, et al. 2004. Long-term alteration of geneexpression without morphological change in testis after neonatalexposure to genistein in mice: toxicogenomic analysis using cDNAmicroarray. Food Chem Toxicol 42:445–452.
Adlercreutz H, Fotsis T, Watanabe S, et al. 1994. Determination of lignansand isoflavonoids in plasma by isotope dilution gas chromatography-mass spectrometry. Cancer Detect Prev 18:259–271.
Agostoni C, Axelsson I, Goulet O, et al. 2006. Soy protein infant formulaeand follow-on formulae: a commentary by the ESPGHAN Committeeon Nutrition. J Pediatr Gastroenterol Nutr 42:352–361.
Agostoni C, Fiocchi A, Riva E, et al. 2007. Growth of infants withIgE-mediated cow’s milk allergy fed different formulas in thecomplementary feeding period. Pediatr Allergy Immunol 18:599–606.
Akingbemi BT, Braden TD, Kemppainen BW, et al. 2007. Exposure tophytoestrogens in the perinatal period affects androgen secretion bytesticular Leydig cells in the adult rat. Endocrinology 148:4475–4488.
Atanassova N, McKinnell C, Turner KJ, et al. 2000. Comparative effects ofneonatal exposure of male rats to potent and weak (environmental)estrogens on spermatogenesis at puberty and the relationship toadult testis size and fertility: evidence for stimulatory effects of lowestrogen levels. Endocrinology 141:3898–3907.
Awoniyi CA, Roberts D, Veeramachaneni DN, et al. 1998. Reproductivesequelae in female rats after in utero and neonatal exposure to thephytoestrogen genistein. Fertil Steril 70:440–447.
464 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Badger TM, Ronis MJ, Hakkak R, et al. 2002. The health consequences ofearly soy consumption. J Nutr 132:559S–565S.
Bateman HL, Patisaul HB. 2009. Disrupted female reproductive physio-logy following neonatal exposure to phytoestrogens or estrogenspecific ligands is associated with decreased GnRH activationand kisspeptin fiber density in the hypothalamus. Neurotoxicology29(6):988–997.
Berger-Achituv S, Shohat T, Romano-Zelekha O, et al. 2005. Widespreaduse of soy-based formula without clinical indications. J PediatrGastroenterol Nutr 41:660–666.
Bhatia J, Greer F. 2008. Use of soy protein-based formulas in infantfeeding. Pediatrics 121:1062–1068.
Bloedon LT, Jeffcoat AR, Lopaczynski W, et al. 2002. Safety andpharmacokinetics of purified soy isoflavones: single-dose adminis-tration to postmenopausal women. Am J Clin Nutr 76:1126–1137.
Boucher BA, Cotterchio M, Kreiger N, Thompson LU. 2008. Soy formulaand breast cancer risk. Epidemiology 19:165–166.
Brown NM, Setchell KD. 2001. Animal models impacted by phytoestro-gens in commercial chow: implications for pathways influenced byhormones. Lab Invest 81:735–747.
Busby MG, Jeffcoat AR, Bloedon LT, et al. 2002. Clinical characteristicsand pharmacokinetics of purified soy isoflavones: single-doseadministration to healthy men. Am J Clin Nutr 75:126–136.
Cabanes A, Wang M, Olivo S, et al. 2004. Prepubertal estradiol andgenistein exposures up-regulate BRCA1 mRNA and reduce mammarytumorigenesis. Carcinogenesis 25:741–748.
Cao YA, Calafat AM, Doerge DR, et al. 2009. Isoflavones in urine,saliva, and blood of infants: data from a pilot study on theestrogenic activity of soy formula. J Expo Sci Environ Epidemiol19:223–234.
Casanova M, You L, Gaido KW, et al. 1999. Developmental effects ofdietary phytoestrogens in Sprague-Dawley rats and interactions ofgenistein and daidzein with rat estrogen receptors alpha and beta invitro. Toxicol Sci 51:236–244.
Cassidy A. 2006. Factors affecting the bioavailability of soy isoflavones inhumans. J AOAC Int 89:1182–1188.
Cassidy A, Albertazzi P, Lise Nielsen I, et al. 2006. Critical review ofhealth effects of soyabean phyto-oestrogens in post-menopausalwomen. Proc Nutr Soc 65:76–92.
Chan GM, Leeper L, Book LS. 1987. Effects of soy formulas on mineralmetabolism in term infants. Am J Dis Child 141:527–530.
Chandra RK. 1997. Five-year follow-up of high-risk infants with familyhistory of allergy who were exclusively breast-fed or fed partialwhey hydrolysate, soy, and conventional cow’s milk formulas.Journal of Pediatric Gastroenterology and Nutrition 24(4):380–388.
Chandra RK. 1998. Five-year follow up of high-risk infants with familyhistory of allergy exclusively breast-fed or fed partial wheyhydrolysate, soy and conventional cow’s milk formulas. NutritionResearch 18(8):1395–1411.
Chandra RK, Hamed A. 1991. Cumulative incidence of atopic disordersin high risk infants fed whey hydrolysate, soy, and conventional cowmilk formulas. Ann Allergy 67(2 Pt 1):129–132.
Chandra RK, Puri S, Hamed A. 1989. Influence of maternal diet duringlactation and use of formula feeds on development of atopic eczemain high risk infants. BMJ 299(6693):228–230.
Chandra RK, Puri S, Suraiya C, Cheema PS. 1986. Influence of maternalfood antigen avoidance during pregnancy and lactation on incidenceof atopic eczema in infants. Clin Allergy 16(6):563–569.
Chandra RK, Singh G, Shridhara B. 1989. Effect of feeding wheyhydrolysate, soy and conventional cow milk formulas on incidenceof atopic disease in high risk infants. Ann Allergy 63(2):102–106.
Chang HC, Churchwell MI, Delclos KB, et al. 2000. Mass spectrometricdetermination of Genistein tissue distribution in diet-exposedSprague-Dawley rats. J Nutr 130:1963–1970.
Chen A, Rogan WJ. 2004. Isoflavones in soy infant formula: a review ofevidence for endocrine and other activity in infants. Annu Rev Nutr24:33–54.
Chen Y, Huang C, Zhou T, Chen G. 2008. Genistein induction of humansulfotransferases in HepG2 and Caco-2 cells. Basic Clin PharmacolToxicol 103:553–559.
Choi SY, Ha TY, Ahn JY, et al. 2008. Estrogenic activities of isoflavonesand flavones and their structure-activity relationships. Planta Med74:25–32.
Chorazy PA, Himelhoch S, Hopwood NJ, et al. 1995. Persistenthypothyroidism in an infant receiving a soy formula case reportand review of the literature. Pediatrics 96:148–150.
Coldham NG, Sauer MJ. 2000. Pharmacokinetics of [(14)C]Genisteinin the rat: gender-related differences, potential mechanisms ofbiological action, and implications for human health. Toxicol ApplPharmacol 164:206–215.
Conrad SC, Chiu H, Silverman BL. 2004. Soy formula complicatesmanagement of congenital hypothyroidism. Arch Dis Child 89:37–40.
Cotroneo MS, Wang J, Eltoum IA, Lamartiniere CA. 2001. Sex steroidreceptor regulation by genistein in the prepubertal rat uterus. MolCell Endocrinol 173:135–145.
Cotroneo MS, Wang J, Fritz WA, et al. 2002. Genistein action in theprepubertal mammary gland in a chemoprevention model. Carcino-genesis 23:1467–1474.
Cruz MLA, Wong WW, Mimouni F, et al. 1994. Effects of infant nutritionon cholesterol synthesis rates. Pediatr Res 35:135–140.
Dalu A, Blaydes BS, Bryant CW, et al. 2002. Estrogen receptor expressionin the prostate of rats treated with dietary genistein. J Chromatogr777:249–260.
Delclos KB, Bucci TJ, Lomax LG, et al. 2001. Effects of dietary genisteinexposure during development on male and female CD (Sprague-Dawley) rats. Reprod Toxicol (Elmsford, NY) 15:647–663.
Delclos KB, Weis CC, Bucci TJ, et al. 2009. Overlapping but distinct effectsof genistein and ethinyl estradiol (EE(2)) in female Sprague-Dawleyrats in multigenerational reproductive and chronic toxicity studies.Reprod Toxicol 27:117–132.
Doerge DR, Chang HC, Churchwell MI, Holder CL. 2000. Analysis ofsoy isoflavone conjugation in vitro and in human blood usingliquid chromatography-mass spectrometry. Drug Metab Dispos28:298–307.
Doerge DR, Churchwell MI, Chang HC, et al. 2001. Placental transfer ofthe soy isoflavone genistein following dietary and gavage adminis-tration to Sprague Dawley rats. Reprod Toxicol 15:105–110.
Doerge DR, Twaddle NC, Banks EP, et al. 2002. Pharmacokinetic analysisin serum of genistein administered subcutaneously to neonatal mice.Cancer Lett 184:21–27.
Drugstore.com. 2009. Formulation information for soy protein-basedinfant formulas. Available at http://www.drugstore.com (see ‘‘Baby& Mom’’). Accessed March 10, 2009.
Essex C. 1996. Phytoestrogens and soy based infant formula: risks remaintheoretical. Br Med J 313:507–508.
Faber KA, HughesJr CL. 1991. The effect of neonatal exposure todiethylstilbestrol, genistein, and zearalenone on pituitary respon-siveness and sexually dimorphic nucleus volume in the castratedadult rat. Biol Reprod 45:649–653.
Faber KA, HughesJr CL. 1993. Dose-response characteristics of neonatalexposure to genistein on pituitary responsiveness to gonadotropinreleasing hormone and volume of the sexually dimorphic nucleus ofthe preoptic area (SDN-POA) in postpubertal castrated female rats.Reprod Toxicol 7:35–39.
FDA. 2000a. Infant formula. 21 CFR 107.FDA. 2000b. Soy: Health claims for soy protein, questions about
other components. Available at http://vm.cfsan.fda.gov/Bdms/fdsoypr.htmlxhealth. Food and Drug Administration.
Fielden MR, Fong CJ, Haslam SZ, Zacharewski TR. 2002. Normalmammary gland morphology in pubertal female mice following inutero and lactational exposure to genistein at levels comparable tohuman dietary exposure. Toxicol Lett 133:181–191.
Fielden MR, Samy SM, Chou KC, Zacharewski TR. 2003. Effect of humandietary exposure levels of genistein during gestation and lactation onlong-term reproductive development and sperm quality in mice.Food Chem Toxicol 41:447–454.
Flynn KM, Ferguson SA, Delclos KB, Newbold RR. 2000. Effects ofgenistein exposure on sexually dimorphic behaviors in rats. ToxicolSci 55:311–319.
Forsyth BW, McCarthy PL, Leventhal JM. 1985. Problems of early infancyformula changes and mothers beliefs about their infants. J Pediatr106:1012–1017.
Fort P, Lanes R, Dahlem S, et al. 1986. Breast feeding andinsulin-dependent diabetes mellitus in children. J Am Coll Nutr 5:439–441.
Foster WG, Younglai EV, Boutross-Tadross O, et al. 2004. Mammary glandmorphology in sprague-dawley rats following treatment with anorganochlorine mixture in utero and neonatal genistein. Toxicol Sci77:91–100.
Franke AA, Custer LJ, Tanaka Y. 1998. Isoflavones in human breast milkand other biological fluids. Am J Clin Nutr 68:1466S–1473S.
Franke AA, Halm BM, Custer LJ, et al. 2006. Isoflavones inbreastfed infants after mothers consume soy. Am J Clin Nutr 84:406–413.
Franke AA, Halm BM, Ashburn LA. 2008a. Isoflavones in children andadults consuming soy. Arch Biochem Biophys 476:161–170.
Franke AA, Halm BM, Ashburn LA. 2008b. Urinary isoflavones areincreased in adults, but decreased in children, consuming soy whenon oral antibiotic therapy. Nutr Cancer 60:627–635.
Freni-Titulaer LW, Cordero JF, Haddock L, et al. 1986. Prematurethelarche in Puerto Rico. A search for environmental factors. Am JDis Child 140:1263–1267.
Fritz WA, Coward L, Wang J, Lamartiniere CA. 1998. Dietary genistein:perinatal mammary cancer prevention, bioavailability and toxicitytesting in the rat. Carcinogenesis 19:2151–2158.
465NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Genovese MI, Lajolo FM. 2002. Isoflavones in soy-based foods consumedin Brazil: levels, distribution, and estimated intake. J Agric FoodChem 50:5987–5993.
Gorski K, Taciak M, Romanowicz K, Misztal T. 2006. Differential effects ofsoy-containing diets on the reproductive tissues growth andreproductive hormone secretion in male rats. Reprod Biol 6:275–290.
Gruskay FL. 1982. Comparison of breast, cow, and soy feedings in theprevention of onset of allergic disease: a 15-year prospective study.Clin Pediatr (Phila) 21:486–491.
Guerrero-Bosagna CM, Sabat P, Valdovinos FS, et al. 2008. Epigenetic andphenotypic changes result from a continuous pre and post nataldietary exposure to phytoestrogens in an experimental population ofmice. BMC Physiol 8:17.
Guo TL, Zhang XL, Bartolucci E, et al. 2002. Genistein and methoxychlormodulate the activity of natural killer cells and the expression ofphenotypic markers by thymocytes and splenocytes in F0 and F1generations of Sprague-Dawley rats. Toxicology 172:205–215.
Hakkak R, Korourian S, Shelnutt SR, et al. 2000. Diets containing wheyproteins or soy protein isolate protect against 7,12-dimethylbenz(a)anthracene-induced mammary tumors in female rats. CancerEpidemiol Biomarkers Prev 9:113–117.
Halm BM, Ashburn LA, Franke AA. 2007. Isoflavones from soyafoods are more bioavailable in children than adults. Br J Nutr98:998–1005.
Halm BM, Franke AA, Ashburn LA, et al. 2008. Oral antibiotics decreaseurinary isoflavonoid excretion in children after soy consumption.Nutr Cancer 60:14–22.
Hilakivi-Clarke L, Cho E, Clarke R. 1998. Maternal genistein exposuremimics the effects of estrogen on mammary gland development infemale mouse offspring. Oncol Rep 5:609–616.
Hilakivi-Clarke L, Cho E, Onojafe I, et al. 1999a. Maternal exposure togenistein during pregnancy increases carcinogen-induced mammarytumorigenesis in female rat offspring. Oncol Rep 6:1089–1095.
Hilakivi-Clarke L, Onojafe I, Raygada M, et al. 1999b. Prepubertalexposure to zearalenone or genistein reduces mammary tumorigenesis.Br J Cancer 80:1682–1688.
Hilakivi-Clarke L, Cho E, Cabanes A, et al. 2002. Dietary modulation ofpregnancy estrogen levels and breast cancer risk among female ratoffspring. Clin Cancer Res 8:3601–3610.
Hillman LS. 1988. Bone mineral content in term infants fed humanmilk, cow milk-based formula, or soy-based formula. J Pediatr113:208–212.
Hillman LS, Chow W, Salmons SS, et al. 1988. Vitamin D metabolism,mineral homeostasis, and bone mineralization in term infants fedhuman milk, cow milk-based formula, or soy-based formula.J Pediatr 112:864–874.
Hines RN. 2008. The ontogeny of drug metabolism enzymes andimplications for adverse drug events. Pharmacol Ther 118:250–267.
Hoey L, Rowland IR, Lloyd AS, et al. 2004. Influence of soya-based infantformula consumption on isoflavone and gut microflora metaboliteconcentrations in urine and on faecal microflora composition andmetabolic activity in infants and children. Br J Nutr 91:607–616.
ILSI. 1999. Safety assessment and potential health benefits of foodcomponents based on selected scientific criteria. ILSI North AmericaTechnical Committee on Food Components for Health Promotion.Crit Rev Food Sci Nutr 39:203–316.
Irvine CH, Shand N, Fitzpatrick MG, Alexander SL. 1998. Daily intakeand urinary excretion of genistein and daidzein by infants fed soy- ordairy-based infant formulas. Am J Clin Nutr 68:1462S–1465S.
Jabbar MA, Larrea J, Shaw RA. 1997. Abnormal thyroid function tests ininfants with congenital hypothyroidism: the influence of soy-basedformula. J Am Coll Nutr 16:280–282.
Jefferson WN, Couse JF, Padilla-Banks E, et al. 2002. Neonatal exposureto genistein induces estrogen receptor (ER)alpha expressionand multioocyte follicles in the maturing mouse ovary: evidencefor ERbeta-mediated and nonestrogenic actions. Biol Reprod67:1285–1296.
Jefferson WN, Padilla-Banks E, Newbold R. 2005. Adverse effects onfemale development and reproduction in CD-1 mice followingneonatal exposure to the phytoestrogen genistein at environmentallyrelevant doses. Biol Reprod 73:798–806.
Jefferson W, Newbold R, Padilla-Banks E, Pepling M. 2006. Neonatalgenistein treatment alters ovarian differentiation in the mouse:inhibition of oocyte nest breakdown and increased oocyte survival.Biol Reprod 74:161–168.
Jefferson WN, Doerge D, Padilla-Banks E, et al. 2009a. Oral exposure togenistin, the glycosylated form of genistein, during neonatal lifeadversely affects the female reproductive system. Environ HealthPerspect 117:1883–1889.
Jefferson WN, Padilla-Banks E, Goulding EH, et al. 2009b. Neonatalexposure to genistein disrupts ability of female mouse reproductivetract to support preimplantation embryo development and implan-tation. Biol Reprod 80:425–431.
Jing H, Pivik RT, Gilchrist JM, Badger TM. 2008. No difference indicatedin electroencephalographic power spectral analysis in 3- and 6-month-old infants fed soy- or milk-based formula. Matern ChildNutr 4:136–145.
Joannou GE, Kelly GE, Reeder AY, et al. 1995. A urinary profile study ofdietary phytoestrogens. The identification and mode of metabolismof new isoflavonoids. J Steroid Biochem Mol Biol 54:167–184.
Jung AL, Carr SL. 1977. A soy protein formula and a milk-based formula.A comparative evaluation in milk-tolerant infants showed nosignificant nutritional differences. Clin Pediatr (Phila) 16:982–985.
King RA, Mano MM, Head RJ. 1998. Assessment of isoflavonoidconcentrations in Australian bovine milk samples. J Dairy Res65:479–489.
Klein SL, Wisniewski AB, Marson AL, et al. 2002. Early exposure togenistein exerts long-lasting effects on the endocrine and immunesystems in rats. Mol Med 8:742–749.
Klemola T, Vanto T, Juntunen-Backman K, et al. 2002. Allergy tosoy formula and to extensively hydrolyzed whey formula in infantswith cow’s milk allergy: a prospective, randomized study with afollow-up to the age of 2 years. J Pediatr 140:219–224.
Kohler L, Meeuwisse G, Mortensson W. 1984. Food intake and growthof infants between six and twenty-six weeks of age on breastmilk, cow’s milk formula, or soy formula. Acta Paediatr Scand73:40–48.
Kouki T, Kishitake M, Okamoto M, et al. 2003. Effects of neonataltreatment with phytoestrogens, genistein and daidzein, on sexdifference in female rat brain function: estrous cycle and lordosis.Horm Behav 44:140–145.
Kulkarni PB, Dorand RD, Bridger WM, et al. 1984. Rickets in prematureinfants fed different formulas. South Med J 77:13–16, 20.
Kurzer MS, Xu X. 1997. Dietary phytoestrogens. Annu Rev Nutr17:353–381.
Lamartiniere CA. 2000. Protection against breast cancer with genistein:a component of soy. Am J Clin Nutr 71:1705S–1707S; discussion1708S–1709S.
Lamartiniere CA, Moore J, Holland M, Barnes S. 1995a. Neonatalgenistein chemoprevents mammary cancer. Proc Soc Exp Biol Med208:120–123.
Lamartiniere CA, Moore JB, Brown NM, et al. 1995b. Genisteinsuppresses mammary cancer in rats. Carcinogenesis 16:2833–2840.
Larkin T, Price WE, Astheimer L. 2008. The key importance of soyisoflavone bioavailability to understanding health benefits. Crit RevFood Sci Nutr 48:538–552.
Lasekan JB, Ostrom KM, Jacobs JR, et al. 1999. Growth of newborn,term infants fed soy formulas for 1 year. Clin Pediatr (Phila) 38:563–571.
Latendresse JR, Bucci TJ, Olson G, et al. 2009. Genistein and ethinylestradiol dietary exposure in multigenerational and chronic studiesinduce similar proliferative lesions in mammary gland of maleSprague-Dawley rats. Reprod Toxicol 28:342–353.
Lephart ED, Adlercreutz H, Lund TD. 2001. Dietary soy phytoestrogeneffects on brain structure and aromatase in Long-Evans rats.Neuroreport 12:3451–3455.
Levy JR, Faber KA, Ayyash L, Hughes Jr CL. 1995. The effect of prenatalexposure to the phytoestrogen genistein on sexual differentiation inrats. Proc Soc Exp Biol Med 208:60–66.
Lewis RW, Brooks N, Milburn GM, et al. 2003. The effects of thephytoestrogen genistein on the postnatal development of the rat.Toxicol Sci 71:74–83.
Liu Z, Zhang X, Li L, et al. 2008. Effects of lactational exposure to soyisoflavones on reproductive system in neonatal female rats. BasicClin Pharmacol Toxicol 102:317–324.
Lu LJ, Anderson KE. 1998. Sex and long-term soy diets affect themetabolism and excretion of soy isoflavones in humans. Am J ClinNutr 68:1500S–1504S.
Lu LJ, Grady JJ, Marshall MV, et al. 1995. Altered time course of urinarydaidzein and genistein excretion during chronic soya diet in healthymale subjects. Nutr Cancer 24:311–323.
Lund TD, Lephart ED. 2001. Dietary soy phytoestrogens produceanxiolytic effects in the elevated plus-maze. Brain Res 913:180–184.
Lund TD, Rhees RW, Setchell KD, Lephart ED. 2001. Altered sexuallydimorphic nucleus of the preoptic area (SDN-POA) volume inadult Long-Evans rats by dietary soy phytoestrogens. Brain Res914:92–99.
MAFF. 1998. Plant oestrogens in soya-based infant formulae. Reportnr 167. London, UK: Ministry of Agriculture, Fisheries, andFood. Available at: http://archive.food.gov.uk/maff/archive/food/infsheet/1998/no167/167phy.htm
Makela SI, Pylkkanen LH, Santti RS, Adlercreutz H. 1995. Dietarysoybean may be antiestrogenic in male mice. J Nutr 125:437–445.
Malloy MH, Berendes H. 1998. Does breast-feeding influence intelligencequotients at 9 and 10 years of age? Early Hum Dev 50:209–217.
466 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011

Mardon J, Mathey J, Kati-Coulibaly S, et al. 2008. Influence of lifelong soyisoflavones consumption on bone mass in the rat. Exp Biol Med(Maywood) 233:229–237.
Masutomi N, Shibutani M, Takagi H, et al. 2003. Impact of dietaryexposure to methoxychlor, genistein, or diisononyl phthalate duringthe perinatal period on the development of the rat endocrine/reproductive systems in later life. Toxicology 192:149–170.
Masutomi N, Shibutani M, Takagi H, et al. 2004a. Dietary influence on theimpact of ethinylestradiol-induced alterations in the endocrine/reproductive system with perinatal maternal exposure. ReprodToxicol 18:23–33.
Masutomi N, Shibutani M, Takagi H, et al. 2004b. Alteration of pituitaryhormone-immunoreactive cell populations in rat offspring aftermaternal dietary exposure to endocrine-active chemicals. ArchToxicol 78:232–240.
McClain RM, Wolz E, Davidovich A, et al. 2007. Reproductive safetystudies with genistein in rats. Food Chem Toxicol 45:1319–1332.
McVey MJ, Cooke GM, Curran IH. 2004a. Altered testicular microsomalsteroidogenic enzyme activities in rats with lifetime exposure to soyisoflavones. J Steroid Biochem Mol Biol 92:435–446.
McVey MJ, Cooke GM, Curran IH. 2004b. Increased serum and testicularandrogen levels in F1 rats with lifetime exposure to soy isoflavones.Reprod Toxicol 18:677–685.
Mimouni F, Campaigne B, Neylan M, Tsang RC. 1993. Bone mineraliza-tion in the first year of life in infants fed human milk, cow-milkformula, or soy-based formula. J Pediatr 122:348–354.
Moller FJ, Zierau O, Hertrampf T, et al. 2009. Long-term effects of dietaryisoflavones on uterine gene expression profiles. J Steroid BiochemMol Biol 113:296–303.
Montani C, Penza M, Jeremic M, et al. 2008. Genistein is an efficientestrogen in the whole-body throughout mouse development. ToxicolSci 103:57–67.
Montani C, Penza M, Jeremic M, et al. 2009. Estrogen receptor-mediatedtranscriptional activity of genistein in the mouse testis. Ann N YAcad Sci 1163:475–477.
Murrill WB, Brown NM, Zhang JX, et al. 1996. Prepubertal genisteinexposure suppresses mammary cancer and enhances gland differ-entiation in rats. Carcinogenesis 17:1451–1457.
Naciff JM, Jump ML, Torontali SM, et al. 2002. Gene expression profileinduced by 17alpha-ethynyl estradiol, bisphenol A, and genistein inthe developing female reproductive system of the rat. Toxicol Sci68:184–199.
Nagao T, Yoshimura S, Saito Y, et al. 2001. Reproductive effects in maleand female rats of neonatal exposure to genistein. Reprod Toxicol15:399–411.
National Toxicology Program (NTP). 2008. NTP MultigenerationalReproductive Study of Genistein (CAS No. 446-72-0) in Sprague-Dawley Rats (Feed Study). Natl Toxicol Program Tech Rep Ser539:1–266.
Newbold RR, Banks EP, Bullock B, Jefferson WN. 2001. Uterineadenocarcinoma in mice treated neonatally with genistein. CancerRes 61:4325–4328.
Nielsen IL, Williamson G. 2007. Review of the factors affectingbioavailability of soy isoflavones in humans. Nutr Cancer 57:1–10.
Nikaido Y, Danbara N, Tsujita-Kyutoku M, et al. 2005. Effects ofprepubertal exposure to xenoestrogen on development of estrogentarget organs in female CD-1 mice. In Vivo 19:487–494.
Nikaido Y, Yoshizawa K, Danbara N, et al. 2004. Effects of maternalxenoestrogen exposure on development of the reproductive tract andmammary gland in female CD-1 mouse offspring. Reprod Toxicol18:803–811.
Odum J, Tinwell H, Jones K, et al. 2001. Effect of rodent diets on thesexual development of the rat. Toxicol Sci 61:115–127.
Padilla-Banks E, Jefferson WN, Newbold RR. 2006. Neonatal exposureto the phytoestrogen genistein alters mammary gland growthand developmental programming of hormone receptor levels.Endocrinology 147:4871–4882.
Pastuszewska B, Taciak M, Ochtabinska A, et al. 2008. Nutritionalvalue and physiological effects of soya-free diets fed to ratsduring growth and reproduction. J Anim Physiol Anim Nutr (Berl)92:63–74.
Patisaul HB, Fortino AE, Polston EK. 2006. Neonatal genistein orbisphenol-A exposure alters sexual differentiation of the AVPV.Neurotoxicol Teratol 28:111–118.
Patisaul HB, Fortino AE, Polston EK. 2007. Differential disruption ofnuclear volume and neuronal phenotype in the preoptic area byneonatal exposure to genistein and bisphenol-A. Neurotoxicology28:1–12.
Patisaul HB, Todd KL, Mickens JA, Adewale HB. 2009. Impact of neonatalexposure to the ERalpha agonist PPT, bisphenol-A or phytoestrogenson hypothalamic kisspeptin fiber density in male and female rats.Neurotoxicology 30:350–357.
Peeters PH, Slimani N, van der Schouw YT, et al. 2007. Variations inplasma phytoestrogen concentrations in European adults. J Nutr137:1294–1300.
Prasain JK, Barnes S. 2007. Metabolism and bioavailability of flavonoidsin chemoprevention: current analytical strategies and futureprospectus. Mol Pharm 4:846–864.
Pumford SL, Morton MM, Turkes A, Griffiths K. 2002. Determination ofthe isoflavonoids genistein and daidzein in biological samples by gaschromatography-mass spectrometry. Ann Clin Biochem 39:281–292.
Rozman KK, Bhatia J, Calafat AM, et al. 2006. NTP-CERHR expert panelreport on the reproductive and developmental toxicity of genistein.Birth Defects Res B Dev Reprod Toxicol 77:485–638.
Rufer CE, Bub A, Moseneder J, et al. 2008. Pharmacokinetics of thesoybean isoflavone daidzein in its aglycone and glucoside form:a randomized, double-blind, crossover study. Am J Clin Nutr87:1314–1323.
Ruhlen RL, Howdeshell KL, Mao J, et al. 2008. Low phytoestrogenlevels in feed increase fetal serum estradiol resulting in the ‘‘fetalestrogenization syndrome’’ and obesity in CD-1 mice. EnvironHealth Perspect 116:322–328.
Scallet AC, Divine RL, Newbold RR, Delclos KB. 2004. Increased volumeof the calbindin D28k-labeled sexually dimorphic hypothalamusin genistein and nonylphenol-treated male rats. Toxicol Sci 82:570–576.
Scientific Committee on Food. 2003. Report on the revision of essentialrequirements of infant formulae and follow-up formulae (adoptedon April 4, 2003). SCF/CS/NUT/IF/65, Final May 18, 2003 Availableat: http://ec.europa.eu/food/food/labellingnutrition/children/formulae_en.htm.
Setchell KD. 1998. Phytoestrogens: the biochemistry, physiology, andimplications for human health of soy isoflavones. Am J Clin Nutr68:1333S–1346S.
Setchell KD, Zimmer-Nechemias L, Cai J, Heubi JE. 1997. Exposure ofinfants to phyto-oestrogens from soy-based infant formula. Lancet350:23–27.
Setchell KD, Zimmer-Nechemias L, Cai J, Heubi JE. 1998. Isoflavonecontent of infant formulas and the metabolic fate of thesephytoestrogens in early life. Am J Clin Nutr 68:1453S–1461S.
Setchell KD, Brown NM, Desai P, et al. 2001. Bioavailability of pureisoflavones in healthy humans and analysis of commercial soyisoflavone supplements. J Nutr 131:1362S–1375S.
Sfakianos J, Coward L, Kirk M, Barnes S. 1997. Intestinal uptake and biliaryexcretion of the isoflavone genistein in rats. J Nutr 127:1260–1268.
Sharpe RM, Martin B, Morris K, et al. 2002. Infant feeding with soyformula milk: effects on the testis and on blood testosterone levels inmarmoset monkeys during the period of neonatal testicular activity.Hum Reprod 17:1692–1703.
Shepard T. 1960. Soybean goiter. N Engl J Med 262:1099–1103.Shibayama T, Fukata H, Sakurai K, et al. 2001. Neonatal exposure to
genistein reduces expression of estrogen receptor alpha andandrogen receptor in testes of adult mice. Endocr J 48:655–663.
Simmen RC, Eason RR, Till SR, et al. 2005. Inhibition of NMU-inducedmammary tumorigenesis by dietary soy. Cancer Lett 224:45–52.
Soucy NV, Parkinson HD, Sochaski MA, Borghoff SJ. 2006. Kinetics ofGenistein and its Conjugated Metabolites in Pregnant Sprague-Dawley Rats Following Single and Repeated Genistein Administration.Toxicol Sci 90:230–240.
Soyfoods Associations of North America. 2009. Soyfood sales &trends. Available at: http://www.soyfoods.org/products/sales-and-trends. Accessed October 10, 2009. Soyfoods Association of NorthAmerica.
Steichen JJ, Tsang RC. 1987. Bone mineralization and growth interm infants fed soy-based or cow milk-based formula. J Pediatr110:687–692.
Strauss L, Makela S, Joshi S, et al. 1998. Genistein exerts estrogen-likeeffects in male mouse reproductive tract. Mol Cell Endocrinol144:83–93.
Strom BL, Schinnar R, Ziegler EE, et al. 2001. Exposure to soy-basedformula in infancy and endocrinological and reproductive outcomesin young adulthood. JAMA 286:807–814.
Takagi H, Shibutani M, Lee KY, et al. 2004. Lack of modifying effectsof genistein on disruption of the reproductive system byperinatal dietary exposure to ethinylestradiol in rats. Reprod Toxicol18:687–700.
Takagi H, Shibutani M, Lee KY, et al. 2005. Impact of maternal dietaryexposure to endocrine-acting chemicals on progesterone receptorexpression in microdissected hypothalamic medial preoptic areas ofrat offspring. Toxicol Appl Pharmacol 208:127–136.
Tan KA, Walker M, Morris K, et al. 2006. Infant feeding with soy formulamilk: effects on puberty progression, reproductive function andtesticular cell numbers in marmoset monkeys in adulthood. HumReprod 21:896–904.
467NTP-CERHR EXPERT PANEL REPORT
Birth Defects Research (Part B) 92:421–468, 2011

Tang L, Singh R, Liu Z, Hu M. 2009. Structure and concentration changesaffect characterization of UGT isoform-specific metabolism ofisoflavones. Mol Pharmacol 6:1466–1482.
Taylor H, Quintero EM, Iacopino AM, Lephart ED. 1999. Phytoestrogensalter hypothalamic calbindin-D28k levels during prenatal develop-ment. Brain Res Dev Brain Res 114:277–281.
Thomsen AR, Almstrup K, Nielsen JE, et al. 2006. Estrogenic effect of soyisoflavones on mammary gland morphogenesis and gene expressionprofile. Toxicol Sci 93:357–368.
Thuillier R, Manku G, Wang Y, Culty M. 2009. Changes in MAPKpathway in neonatal and adult testis following fetal estrogenexposure and effects on rat testicular cells. Microsc Res Tech 87:1–26.
Thygarajan A, Burks AW. 2008. American Academy of Pediatricsrecommendations on the effects of early nutritional interventionson the development of atopic disease. Curr Opin Pediatr 20:698–702.
Tuohy PG. 2003. Soy infant formula and phytoestrogens. J Paediatr ChildHealth 39:401–405.
Turck D. 2007. Soy protein for infant feeding: what do we know? CurrOpin Clin Nutr Metab Care 10:360–365.
U.S. Centers for Disease Control and Prevention. 2008. National report onbiochemical indicators of diet and nutrition in the U.S. population1999–2002. Atlanta, GA: National Center for EnvironmentalHealth; July 2008. Available at: http://www.cdc.gov/nutritionreport/index.html
UK-Committee-on-Toxicity. 2003. Phytoestrogens and health. London,UK: Committee on Toxicity of Chemicals in Food, ConsumerProducts and the Environment. Available at: http://www.food.gov.uk/multimedia/pdfs/phytoreport0503
United Soybean Board. 2009. 16th annual survey: consumer attitudes aboutnutrition. Available at: http://www.soyfoods.org/wp/wp-content/up-loads/2009/ConsumerAttitudes2009.pdf. Accessed October 10, 2009.
Valentin-Blasini L, Blount BC, Caudill SP, Needham LL. 2003. Urinary andserum concentrations of seven phytoestrogens in a human referencepopulation subset. J Expo Anal Environ Epidemiol 13:276–282.
Venkataraman PS, Luhar H, Neylan MJ. 1992. Bone mineral metabolismin full-term infants fed human milk, cow milk-based, and soy-basedformulas. Am J Dis Child 146:1302–1305.
Vilela ML, Willingham E, Buckley J, et al. 2007. Endocrine disruptors andhypospadias: role of genistein and the fungicide vinclozolin. Urology70:618–621.
Wagner JD, Jorgensen MJ, Cline JM, et al. 2009. Effects of soy vs. caseinprotein on body weight and glycemic control in female monkeys andtheir offspring. Am J Primatol. 2009 Sep; 71(9):802–811.
Wang XJ, Bartolucci-Page E, Fenton SE, You L. 2006. Altered mammarygland development in male rats exposed to genistein and methoxy-chlor. Toxicol Sci 91:93–103.
Weber KS, Setchell KD, Lephart ED. 2001. Maternal and perinatal brainaromatase: effects of dietary soy phytoestrogens. Brain Res Dev BrainRes 126:217–221.
Whitten PL, Patisaul HB. 2001. Cross-species and interassay comparisonsof phytoestrogen action. Environ Health Perspect 109:5–20.
You L, Casanova M, Bartolucci EJ, et al. 2002a. Combined effects ofdietary phytoestrogen and synthetic endocrine-active compound onreproductive development in Sprague-Dawley rats: genistein andmethoxychlor. Toxicol Sci 66:91–104.
You L, Sar M, Bartolucci EJ, McIntyre BS. 2002b. Modulation of mammarygland development in prepubertal male rats exposed to genisteinand methoxychlor. Toxicol Sci 66:216–225.
468 MCCARVER ET AL.
Birth Defects Research (Part B) 92:421–468, 2011
Related Documents











