Research Report Novelty-induced correlation between visual neurons and the hippocampal theta rhythm in sleep and wakefulness Marisa Pedemonte * , Juan P. Gambini, Ricardo A. Velluti Neurofisiologı ´a, Departamento de Fisiologı ´a, Facultad de Medicina, Universidad de la Repu ´blica, Ave. General Flores 2125, 11800 Montevideo, Uruguay Accepted 29 July 2005 Available online 24 October 2005 Abstract Various rhythms have been shown to affect sensory processing such as the waking –sleep cycle and the hippocampal theta waves. Changes in the firing of visual lateral geniculate nucleus neurons have been reported to be dependent on the animal’s behavioral state. The lateral geniculate extracellular neuronal firing and hippocampal field activity were recorded in chronically implanted animals to analyze the relationship during quiet wakefulness and sleep associated with stimulation shifts that may introduce novelty. During wakefulness, a change in light flash stimulation pattern (stimuli frequency shift, stimuli on and off) caused an increment in the theta band power in 100% of the cases and a phase-locking of the spikes in 53% of the recorded neurons. During slow wave sleep, there were no consistent changes in the theta power notwithstanding 13% of the neurons exhibited phase-locking, i.e., novelty may induce changes in the temporal correlation of visual neuronal activity with the hippocampal theta rhythm in sleep. The present results suggest that visual processing in slow wave sleep exists, while auditory information and learning were reported during slow wave sleep in animals and newborn humans. The changes in the theta power as well as in the neuronal phase-locking amount indicate that in slow wave sleep, the ability of the hippocampus to detect/process novelty, although present, may be decreased. This is consistent with the noticeable decrease in awareness of the environment during sleep. D 2005 Elsevier B.V. All rights reserved. Theme: Sensory systems Topic: Visual psychophysics and behavior Keywords: Sensory processing; Attention; Visual unit; Phase-locking; Lateral geniculate; Sleep– wakefulness 1. Introduction Neuronal activity in sensory systems is dependent on several factors that modulate incoming information from specific environmental and body channels. These neurons may be part of different brain networks/cell assemblies that shape their activity, of which a meaningful modulator is the animal’s behavioral state. The central nervous system (CNS) is continuously receiving sensory inputs during the 24-h waking – sleep cycle [37,38]. Although sensory processing occurs without interruptions, it is known that neuronal activity, unitary and field evoked, in every sensory system can change depending on the sleeping or waking brain [36]. In the visual system, lateral geniculate nucleus (LGn) neuronal activity shifts were reported to be sleep state- dependent [7,20,21,24]. Within the waking –sleep rhythm, other cycles exist that modulate neuronal activity with a different temporal scale, the ultradian hippocampal theta rhythm being a conspicuous example. Theta activity, 4 to 10 –12 cycles per second (cps), is always present in the CNS electrophysiological activity even in slow wave sleep as shown by Fourier analysis [12,37]. Furthermore, it has been related to several brain processes; it was found to be involved in motor activities during both wakefulness and paradoxical sleep [4,11,19], as well as in sensory processing related to the motor context 0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2005.07.069 * Corresponding author. Fax: +598 2 924 87 84. E-mail address: [email protected] (M. Pedemonte). Brain Research 1062 (2005) 9 – 15 www.elsevier.com/locate/brainres

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/brainres
Brain Research 106
Research Report
Novelty-induced correlation between visual neurons and the hippocampal
theta rhythm in sleep and wakefulness
Marisa Pedemonte*, Juan P. Gambini, Ricardo A. Velluti
Neurofisiologıa, Departamento de Fisiologıa, Facultad de Medicina, Universidad de la Republica, Ave. General Flores 2125,
11800 Montevideo, Uruguay
Accepted 29 July 2005
Available online 24 October 2005
Abstract
Various rhythms have been shown to affect sensory processing such as the waking–sleep cycle and the hippocampal theta waves.
Changes in the firing of visual lateral geniculate nucleus neurons have been reported to be dependent on the animal’s behavioral state. The
lateral geniculate extracellular neuronal firing and hippocampal field activity were recorded in chronically implanted animals to analyze the
relationship during quiet wakefulness and sleep associated with stimulation shifts that may introduce novelty. During wakefulness, a change
in light flash stimulation pattern (stimuli frequency shift, stimuli on and off) caused an increment in the theta band power in 100% of the
cases and a phase-locking of the spikes in 53% of the recorded neurons. During slow wave sleep, there were no consistent changes in the
theta power notwithstanding 13% of the neurons exhibited phase-locking, i.e., novelty may induce changes in the temporal correlation of
visual neuronal activity with the hippocampal theta rhythm in sleep. The present results suggest that visual processing in slow wave sleep
exists, while auditory information and learning were reported during slow wave sleep in animals and newborn humans. The changes in the
theta power as well as in the neuronal phase-locking amount indicate that in slow wave sleep, the ability of the hippocampus to detect/process
novelty, although present, may be decreased. This is consistent with the noticeable decrease in awareness of the environment during sleep.
D 2005 Elsevier B.V. All rights reserved.
Theme: Sensory systems
Topic: Visual psychophysics and behavior
Keywords: Sensory processing; Attention; Visual unit; Phase-locking; Lateral geniculate; Sleep–wakefulness
1. Introduction
Neuronal activity in sensory systems is dependent on
several factors that modulate incoming information from
specific environmental and body channels. These neurons
may be part of different brain networks/cell assemblies that
shape their activity, of which a meaningful modulator is the
animal’s behavioral state. The central nervous system (CNS)
is continuously receiving sensory inputs during the 24-h
waking–sleep cycle [37,38]. Although sensory processing
occurs without interruptions, it is known that neuronal
0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2005.07.069
* Corresponding author. Fax: +598 2 924 87 84.
E-mail address: [email protected] (M. Pedemonte).
activity, unitary and field evoked, in every sensory system
can change depending on the sleeping or waking brain [36].
In the visual system, lateral geniculate nucleus (LGn)
neuronal activity shifts were reported to be sleep state-
dependent [7,20,21,24].
Within the waking–sleep rhythm, other cycles exist that
modulate neuronal activity with a different temporal scale,
the ultradian hippocampal theta rhythm being a conspicuous
example. Theta activity, 4 to 10–12 cycles per second (cps),
is always present in the CNS electrophysiological activity
even in slow wave sleep as shown by Fourier analysis
[12,37]. Furthermore, it has been related to several brain
processes; it was found to be involved in motor activities
during both wakefulness and paradoxical sleep [4,11,19], as
well as in sensory processing related to the motor context
2 (2005) 9 – 15

M. Pedemonte et al. / Brain Research 1062 (2005) 9–1510
[13,18]. It was also found to be involved in spatio-temporal
learning [27,42], association of discontiguous events [41],
and learning of temporal sequences [22,25]. A role of the
theta rhythm in learning and memory has been proposed
from different viewpoints [1,5,8,15,17,40]. This hippo-
campal rhythm has also been linked to modulation of
autonomic processes such as heart rate [29,31].
In previous reports, we found that LGn visual units show
phase-locking to the theta rhythm during short periods of
wakefulness, slow wave sleep, and paradoxical sleep
[10,37]. Moreover, other sensory neurons such as auditory
ones also showed phase-locking with theta rhythm at
different levels of the auditory pathway including the
primary cortex. Thus, the theta rhythm was postulated as a
temporal organizer of sensory processing [28,30]. Proces-
sing of many sensory modalities were also related to the
theta rhythm, e.g., tactile [26], pain [39], and olfactory [23].
The notion that theta rhythm changes are dependent on
the animal’s attention state during behavior was suggested
by Kemp and Kaada (1975) [16]. Vinogradova (2001) [40]
summarizes the modulating influences of theta rhythm in
two ways: (1) as a regulatory pattern, linking the hippo-
campus to the brain-stem structures, sensing the attention
level, and introducing primary information about the
changes in the environment; and (2) as an informational
pattern acting on specific systems with reciprocal inter-
actions with the neocortex.
The available information, cross-correlating visual neu-
rons activity to the theta rhythm – phase-locking – leads us
to analyze changes in response to varying visual stimuli.
The main hypothesis proposes the hippocampal theta
participating in the visual unitary input temporal processing
(as an internal zeitgeber), i.e., phase-locking linked with
stimulus shifts – frequency, stim on, stim off – in
wakefulness and particularly during slow wave sleep.
2. Materials and methods
Adult guinea pigs (Cavia porcellus, n = 12) weighing
450–550 g were chronically implanted under pentobarbital
anesthesia (35 mg/kg, ip) with macroelectrodes to record the
CA1 hippocampus electrogram (Hipp: A = 6; L = 2; H =
�4), electrocorticogram (ECoG), and electromyogram of
the neck muscles (EMG), for behavioral control. Two light
metal bars were cemented to the animal’s skull. This
procedure permitted, after a week of recovery, the painless
reproduction of the stereotaxic position for unit assessment
during physiological behavior. The animal’s head was thus
firmly held, with the body hanging on a canvas, during the
recording sessions. A small hole (2 mm diameter) was
drilled in the skull over the LGn (A = 5–7; L = 4–6; H = 5–
9), covered with antibiotic cream and left open for glass
microelectrode penetrations. After a week of recovery,
animals were trained to be placed on a hammock during
2–3 days, when they showed to have normal sleep–waking
cycles. All experimental procedures were conducted in
accordance with the Medical School Committee for Animal
Research (Montevideo) and with the NIH Guide for the
Care and Use of Laboratory Animals.
During the recording sessions, flashes of light (Grass
Photo-stimulator TS 22; 2–16/s, 30 cm distant) were
applied as visual stimuli, e.g., changing stimulus frequency,
stopping or initiating it. The pupils were dilated by atropine,
the eyelids were always open, even during sleep as it
normally occurs in these animals. A glass micropipette –
filled with 3 M sodium acetate and Sky-blue solution – was
advanced through the skull hole using a micro-manipulator
(Narishige MO-8) searching for evoked extracellular unit
firing. Flash synchronizing signal, neuronal discharge,
ECoG, EMG, and Hipp were amplified, acquired, and
stored for off-line processing by computer software (Spike2,
Cambridge Electronic Design). Light stimulation frequency
ranged from 1 to 20/s; light on and light off were also an
applied pattern. Flash light frequency shifts (120 changes
recorded in 32 neurons) were analyzed; 60 of them during
wakefulness and 60 during slow wave sleep. Sixty percent
of the light stimulation shifts increased the frequency, 25%
decreased, and 15% were shifts from stimulus on to off.
Epochs without any movement and for which there was no
change in the behavioral state before and after light
frequency change were considered for processing.
Three successive temporal windows (5 s duration each)
were selected for subsequent processing: the first one,
immediately prior to the flash frequency change; the second
and the third are successive windows immediately after the
stimulus shifts. Neuronal firing was digitized with a voltage
window level and analyzed with peri-stimulus time histo-
grams to be characterized as a visual one. The hippocampal
electrogram was band-pass filtered (between 0.5 and 100
Hz), sampled at 500 Hz, and processed using autocorrela-
tion and power spectrum analysis. The cross-correlation
between hippocampal field activity and spikes was assessed
by spike-triggered averaging. In order to analyze the theta
band power shifts, the power spectra before flash stimula-
tion rate changes were taken as the reference (100%). The
Student’s t test was used for statistical validation of the
observed changes immediately before and after the stimulus
shifts.
At the end of the recording sessions, sites were
iontophoretically injected with Sky-blue; after no more than
three successful recordings, the animals were perfused with
10% formalin solution and the brains were removed for
micro-anatomical analysis.
3. Results
Epochs during which changes in the visual stimulation
rate occurred were analyzed in 32 neurons: 14 units during
both quiet wakefulness and slow wave sleep, 9 units during
quiet wakefulness and 9 units during slow wave sleep.

M. Pedemonte et al. / Brain Research 1062 (2005) 9–15 11
All the flash frequency stimulation rate changes present
during quiet wakefulness (n = 60) provoked increments in
the theta band power ranging between 110% and 553%
(mean = 183 T 102%). The power spectra at 100%
correspond to the period just previous to the light
stimulation change (Fig. 1, arrows A). The animal was
head-restrained while any possible movements were con-
trolled through the neck muscles EMG and direct observa-
tion. Periods with movements were discarded from the
analysis.
Results were different during slow wave sleep: 60 flash
stimulation shifts provoked changes in the theta band power
spectra, incrementing the power in 57% (34 out of 60),
while 43% (26 out of 60) of the light stimulation shifts
induced power decrements. The power of the theta band
after flash frequency changes varied from 34% to 372%
(mean = 106 T 48%; Fig. 1, slow wave sleep, arrow B).
Averages and their standard deviation both during quiet
wakefulness and slow wave sleep are shown in the Fig. 1
inset (bars) showing only significant changes in wakefulness
(P < 0.005).
The temporal correlation between visual units and theta
rhythm (phase-locking) was different in quiet wakefulness
compared to slow wave sleep. Fifty-three percent (32 out of
60) of the light stimulation changes evoked a cross-
correlation (phase-locking) between the LGn neuronal firing
and the Hipp theta rhythm during wakefulness. On the other
hand, only 13% of the flash stimulation changes (8 out of
Fig. 1. Hippocampal theta band power spectra shifts (4–12 Hz) evoked by flash f
(SWS). Sixty episodes of light stimulation rate changes were studied during wak
windows around the beginning of the stimulation change. Arrows A, theta power s
as 100%. Arrows B, each black point represents the percentage of the theta power
ones. During wakefulness, all the stimulation changes provoke a significant ( P <
during slow wave sleep, no significant shift appears (ranging from 34% to 372%). U
nucleus recording loci and the electrode positioning. Lower insert shows the pow
60) evoked a phase-locking of the neuronal discharge with
the theta rhythm during slow wave sleep.
Fig. 2 shows an example of neuronal discharge during
wakefulness throughout two light stimulation frequency
shifts. The post-stimulus time histogram (PSTH) character-
izes the neuron as a visual one. The theta waves increased
their amplitude and power after the flash rate changed
(windows B and E, raw recording, power spectra, and Hipp
autocorrelation) and the unit firing showed the correspond-
ing phase-locking (windows B and E, cross-correlation).
The same stimulation paradigm was applied 25 times to
study the unit discharge related to theta rhythm and the
rhythm power spectrum. The same light stimulation
frequency shifts (2 to 4 and 4 to 8 flashes/s) were applied
21 times during slow wave sleep with different quantitative
results exhibiting both lower theta phase-locking and
increasing/decreasing theta power. These results are indica-
tive that the changes are related to the behavioral shift and
not to the stimulation frequency variations.
An example during slow wave sleep – evoked and
spontaneous activity – is shown in Fig. 3 in which phase-
locking appeared when the light stimulation was off.
Although the theta power spectrum did not increase after
light stimulation went off (B, power spectra), the LGn unit
began to show phase-locking with the theta rhythm (B,
cross-correlation). This figure is intended to show that
phase-locking is not only provoked by flash stimulating
frequency shifts but also associated to flash stimulation off.
requency stimulation changes during wakefulness (W) and slow wave sleep
efulness and sixty during slow wave sleep. An analysis was made with 5-s
pectrum in the windows previous to the flash stimulus change is considered
spectra in the windows after stimulus changes, compared with the previous
0.005) increment in the theta power (ranging from 110% to 553%), while
pper inset: drawing showing the hippocampus (Hipp) and lateral geniculate
er values averaged with the corresponding standard error.
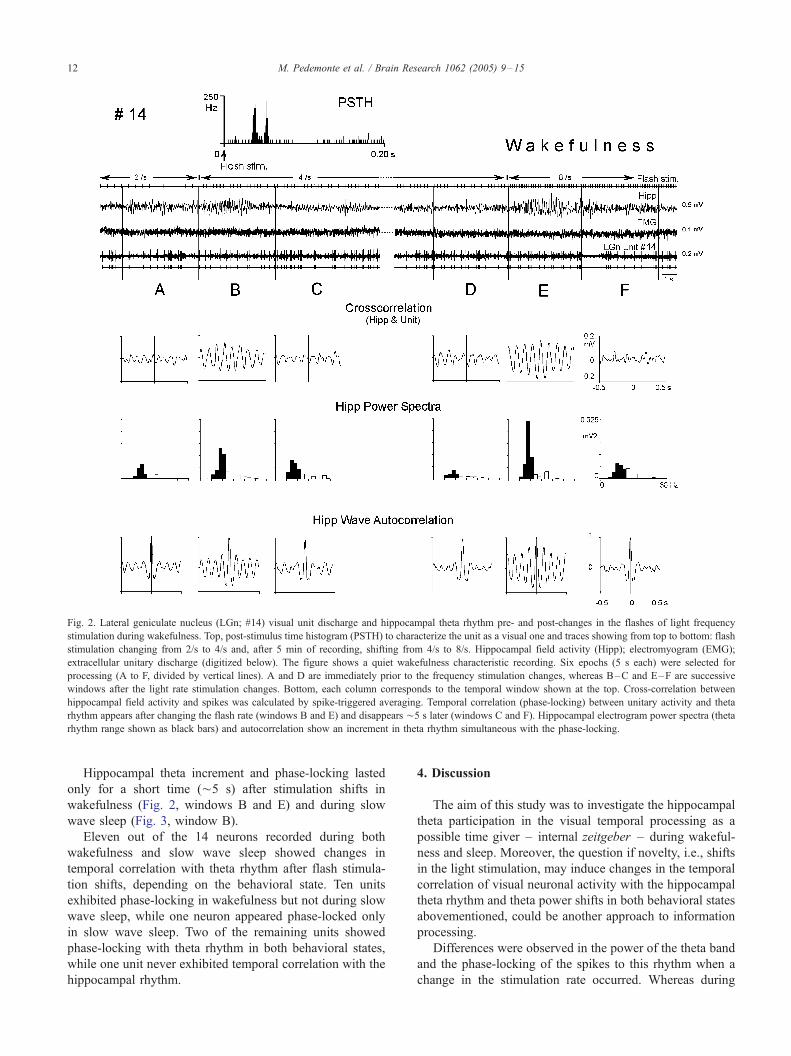
Fig. 2. Lateral geniculate nucleus (LGn; #14) visual unit discharge and hippocampal theta rhythm pre- and post-changes in the flashes of light frequency
stimulation during wakefulness. Top, post-stimulus time histogram (PSTH) to characterize the unit as a visual one and traces showing from top to bottom: flash
stimulation changing from 2/s to 4/s and, after 5 min of recording, shifting from 4/s to 8/s. Hippocampal field activity (Hipp); electromyogram (EMG);
extracellular unitary discharge (digitized below). The figure shows a quiet wakefulness characteristic recording. Six epochs (5 s each) were selected for
processing (A to F, divided by vertical lines). A and D are immediately prior to the frequency stimulation changes, whereas B–C and E–F are successive
windows after the light rate stimulation changes. Bottom, each column corresponds to the temporal window shown at the top. Cross-correlation between
hippocampal field activity and spikes was calculated by spike-triggered averaging. Temporal correlation (phase-locking) between unitary activity and theta
rhythm appears after changing the flash rate (windows B and E) and disappears ¨5 s later (windows C and F). Hippocampal electrogram power spectra (theta
rhythm range shown as black bars) and autocorrelation show an increment in theta rhythm simultaneous with the phase-locking.
M. Pedemonte et al. / Brain Research 1062 (2005) 9–1512
Hippocampal theta increment and phase-locking lasted
only for a short time (¨5 s) after stimulation shifts in
wakefulness (Fig. 2, windows B and E) and during slow
wave sleep (Fig. 3, window B).
Eleven out of the 14 neurons recorded during both
wakefulness and slow wave sleep showed changes in
temporal correlation with theta rhythm after flash stimula-
tion shifts, depending on the behavioral state. Ten units
exhibited phase-locking in wakefulness but not during slow
wave sleep, while one neuron appeared phase-locked only
in slow wave sleep. Two of the remaining units showed
phase-locking with theta rhythm in both behavioral states,
while one unit never exhibited temporal correlation with the
hippocampal rhythm.
4. Discussion
The aim of this study was to investigate the hippocampal
theta participation in the visual temporal processing as a
possible time giver – internal zeitgeber – during wakeful-
ness and sleep. Moreover, the question if novelty, i.e., shifts
in the light stimulation, may induce changes in the temporal
correlation of visual neuronal activity with the hippocampal
theta rhythm and theta power shifts in both behavioral states
abovementioned, could be another approach to information
processing.
Differences were observed in the power of the theta band
and the phase-locking of the spikes to this rhythm when a
change in the stimulation rate occurred. Whereas during

Fig. 3. Lateral geniculate nucleus (LGn; #17) visual unit discharge and the hippocampal theta rhythm associated to a light stimulation shift during slow wave
sleep. Top, post-stimulus time histogram (PSTH) characterizes the unit as a visual one and traces showing from top to bottom are: flash synchronizing signal,
changing from 16/s to no stimulation (spontaneous activity); hippocampal electrogram (Hipp); electromyogram (EMG); extracellular unitary discharges (with
their corresponding digitized traces below). This recording is characteristic of slow wave sleep because of the high amplitude and low frequency in the Hipp
field activity and the EMG low activity. As in Fig. 2, three epochs (5 s each) were selected for processing (A to C, divided by vertical lines); A is immediately
previous to the end of flash stimulation while B–C are successive windows without flashes (spontaneous firing). In the lower part, each column corresponds to
the temporal window shown above. Cross-correlation between hippocampal field activity and spikes was calculated by spike-triggered averaging. Temporal
correlation (phase-locking) between unitary discharge and theta rhythm appears after changing the flash stimulation (window B) and disappears some seconds
(¨5 s) later (window C). Hipp power spectra (theta rhythm range as black bars) exhibit two frequency peaks after the flash shift. Hipp wave autocorrelogram
does not show any dominant frequency after stimulation shift.
M. Pedemonte et al. / Brain Research 1062 (2005) 9–15 13
wakefulness 100% of the cases showed an increase in the
theta band power, the changes observed during slow wave
sleep were different exhibiting increasing and decreasing
theta power. In addition, 53% of the neurons showed theta
phase-locking during quiet wakefulness whereas this phe-
nomenon was observed in only 13% of the neurons during
slow wave sleep.
The type 2 theta in the restrained guinea pig, i.e., the
theta we are recording, occurs during sensory processing in
an alert animal [33]; it can be recorded in slow wave sleep
during which theta rhythm is present as shown in the
literature [12] as well as in our previous [30,37] and in the
present report power spectra analysis in slow wave sleep.
Moreover, in addition to the well-established relationship
between theta and motor activity, a non-motor process
appears as contributing to theta of which a much likely
candidate is a sensory process [35].
The consistent increment of theta power when the
stimulus changes occurred in quiet wakefulness is in
accordance with the idea that the hippocampal theta senses
and expresses a different level of attention, considering the
hippocampus as a novelty detector and comparator [9,14,40].
A double interconnected function has been postulated for
this rhythm, i.e., selective attention and protection from
interference, and the global function of selective information
transfer into cortical memory storage [34,43].

M. Pedemonte et al. / Brain Research 1062 (2005) 9–1514
The present results showing an induction of phase-
locking (temporal correlation) as a consequence of the
stimulation change supports the notion that the theta rhythm
serves as a temporal organizer for specific sensory process-
ing as previously postulated for the auditory and visual
systems [10,28,30,37,38]. In support of our proposal for
theta rhythm acting as a temporal organizer, oscillatory
influences have been reported transforming an asymmetric
rate code into a temporal code, playing a critical role in
temporal sequence learning by compressing and replaying
in a relevant temporal order [25].
Theta power increment and phase-locking are maintained
for approximately 5 s after stimulus shift (see windows B
and E in Fig. 2). This duration, about 5 s, is in accordance
with previous results showing a few seconds (5–10 s)
duration epochs with theta increment and LGn neuronal
firing phase-locked with it, appearing at random without any
known significant environmental change [37]. Furthermore,
Vinogradova (2001) [40] showed inhibitory responses of
hippocampal CA3 neurons after auditory stimulation that
disappeared after 4–6 s because of gradual habituation, a
well-known theta-associated phenomenon.
The present results suggest that visual processing in slow
wave sleep exists, to some extent, although different than
during wakefulness. The changes in the theta power as well
as in the neuronal phase-locking amount indicate that in
slow wave sleep, the ability of the hippocampus to detect
novelty, although present, may be decreased. This is
consistent with the noticeable decrease in awareness of the
environment during sleep while auditory learning was
reported during slow wave sleep in newborn humans [6],
supporting the slow wave sleep capacity to process
information [37].
The differences we observed between the quiet wakeful-
ness and sleep, recording from the same neuron, agree with
the hypothesis that induced activity, the theta rhythm in this
particular case, plays a functional role reflecting changes in
the parameters controlling dynamic interactions within and
between brain structures [2,3,32].
Since visual LGn and auditory neuronal firing do not
cease in wakefulness nor in slow wave sleep [10,37],
either analyzing the evoked activity (Fig. 3, window A) or
the spontaneous one (Fig. 3, windows B and C), we
postulate that the temporal pattern of spikes and theta
phase-locking are important factors that may reflect
whether or not the brain is aware of temporal changes in
the environment, i.e., processing information, even during
slow wave sleep.
Acknowledgments
We are grateful to Prof. Peter M. Narins (UCLA) and Dr.
Jose L. Pena (Caltech) for reading the manuscript and their
very valuable suggestions. Grants: Partially supported by the
Program for Basic Science Development (PEDECIBA),
Ministerio de Educacion y Cultura—Universidad de la
Republica—United Nations Development Program (UNDP).
References
[1] W.R. Adey, C.W. Dunlop, C.E. Hendrix, Hippocampal slow waves
distribution and phase relations in the course of approach learning,
Arch. Neurol. 3 (1960) 74–90.
[2] E. Basar, S. Karakas, Event-related oscillations in the brain, in: E.
Basar (Ed.), Brain Functions and Oscillations, Springer-Verlag, Berlin,
1998, pp. 147–151.
[3] M. Bastiaansen, P. Hagoort, Event-induced theta responses as
a window on the dynamics of memory, Cortex 39 (2003)
967–992.
[4] W. Buno, J.C. Velluti, Relationship of hippocampal theta cycle with
bar pressing during self-stimulation, Physiol. Behav. 19 (1977)
615–621.
[5] A.P. Burgess, J.H. Gruzelier, Short duration synchronization of human
theta rhythm during recognition memory, Neurol. Rep. 8 (1997)
1039–1042.
[6] M. Cheour, O. Martynova, R. Naatanen, R. Erkkola, M. Sillanpaa, P.
Kero, A. Raz, M-L. Kaipio, J. Hiltunen, O. Aaltonen, J. Savela, H.
Hamalainen, Speech sounds learned by sleeping newborns, Nature
415 (2002) 599–600.
[7] A.M. Coenen, H.J. Gerrits, A.J. Vendrik, Analysis of the response
characteristics of optic tract and geniculate units and their mutual
relationships, Exp. Brain Res. 15 (1972) 452–471.
[8] M. Doppelmayr, W. Klimesch, J. Schwaiger, P. Auinger, T. Winkler,
Theta synchronization in the human EEG and episodic retrieval,
Neurosci. Lett. 257 (1998) 41–44.
[9] H. Eichenbaum, To cortex: thanks for the memories, Neuron 19 (1997)
481–484.
[10] J.P. Gambini, R.A. Velluti, M. Pedemonte, Hippocampal theta rhythm
synchronized visual neurons in sleep and waking, Brain Res. 926
(2002) 137–141.
[11] E. Garcıa-Austt, Hippocampal level of neural integration, in: E.
Ajmone-Marsan, F. Reinoso-Suarez (Eds.), Cortical Integration
Basic Archicortical and Cortical Association Levels of Neuronal
Integrations, IBRO Monograph Series, Raven Press, New York,
1984, pp. 91–104.
[12] J.M. Gaztelu, M. Romero-Vives, V. Abraira, E. Garcıa-Austt, Hippo-
campal EEG theta power density is similar during slow-wave sleep
and paradoxical sleep. A long-term study in rats, Neurosci. Lett. 172
(1994) 31–34.
[13] E. Grastyan, K. Lissak, I. Madarasz, Hippocampal activity during the
development of conditioned reflex, Electroencephalogr. Clin. Neuro-
physiol. 11 (1959) 409–430.
[14] S. Grossberg, J.W.L. Merrill, A neural network model of adaptively
timed reinforcement learning and hippocampal dynamics, Cogn. Brain
Res. 1 (1992) 3–38.
[15] M.J. Kahana, R. Sekuler, J.B. Caplan, M. Kirschen, J.R. Madsen,
Human theta oscillations exhibit task dependence during virtual maze
navigation, Nature 399 (1999) 781–784.
[16] I.R. Kemp, B.R. Kaada, The relation of hippocampal theta activity to
arousal, attentive behaviour and somato-motor movements in unre-
strained cats, Brain Res. 95 (1975) 323–342.
[17] W. Klimesch, EEG alpha and theta oscillations reflect cognitive and
memory performance: a review and analysis, Brain Res. Rev. 29
(1999) 169–195.
[18] R. Kramis, C.H. Vanderwolf, B.H. Bland, Two types of hippocampal
rhythmical slow activity in both the rabbit and the rat: relations to
behaviour and effects atropine, diethyl ether, urethane and pentobar-
bital, Exp. Neurol. 49 (1975) 58–85.
[19] J. Lerma, E. Garcıa-Austt, Hippocampal theta rhythm during
paradoxical sleep. Effects of afferent stimuli and phase-relationships

M. Pedemonte et al. / Brain Research 1062 (2005) 9–15 15
with phasic events, Electroencephalogr. Clin. Neurophysiol. 60
(1985) 46–54.
[20] M.S. Livingstone, D.H. Hubel, Effects of sleep and arousal on the
processing of visual information in the cat, Nature 291 (1981)
554–561.
[21] L. Maffei, G. Morruzi, G. Rizzolati, Influence of sleep and wakeful-
ness on the response of lateral geniculate units to sinewave photic
stimulation, Arch. Ital. Biol. 103 (1965) 609–622.
[22] J.C. Magee, A prominent role for intrinsic neuronal properties in
temporal coding, Trends Neurosci. 26 (2003) 14–16.
[23] T.W. Margrie, A.T. Schaefer, Theta oscillation coupled spike latencies
yield computational vigour in a mammalian sensory system, J. Physiol.
546 (2003) 363–374.
[24] R. McCarley, O. Benoit, G. Barrionuevo, Lateral geniculate nucleus
unitary discharge in sleep waking: state- and rate-specific aspects,
J. Neurophysiol. 50 (1983) 798–817.
[25] M.R. Mehta, A.K. Lee, M.A. Wilson, Role of experience and
oscillations in transforming a rate code into a temporal code, Nature
417 (2002) 741–745.
[26] A. Nunez, I. de, E. Garcıa-Austt, Relationships of nucleus reticularis
pontis oralis neuronal discharge with sensory and carbachol evoked
hippocampal theta rhythm, Exp. Brain Res. 87 (1991) 303–308.
[27] J. O’Keefe, M.L. Recce, Phase relationship between hippocampal
place units and EEG theta rhythm, Hippocampus 3 (1993) 317–330.
[28] M. Pedemonte, J.L. Pena, R.A. Velluti, Firing of inferior colliculus
auditory neuron is phase-locked to the hippocampus theta rhythm
during paradoxical sleep and waking, Exp. Brain Res. 112 (1996)
41–46.
[29] M. Pedemonte, A. Rodrıguez, R.A. Velluti, Hippocampal theta waves
as an electrocardiogram rhythm timer in paradoxical sleep, Neurosci.
Lett. 276 (1999) 5–8.
[30] M. Pedemonte, L. Perez-Perera, J.L. Pena, R.A. Velluti, Sleep and
wakefulness auditory processing: cortical units vs. hippocampal theta
rhythm, Sleep Res. Online 4 (2001) 51–57.
[31] M. Pedemonte, N. Goldstein-Daruech, R.A. Velluti, Temporal corre-
lation between heart rate, medullary units and hippocampal theta
rhythm in anesthetized, sleeping and awake guinea pigs, Auton.
Neurosci.: Basic Clin. 107 (2003) 99–104.
[32] G. Pfurtscheller, F.H. Lopes da Silva, Event-related EEG–MEG
synchronization and desynchronization: basic principles, Clin. Neuro-
physiol. 110 (1999) 1842–1857.
[33] R.S. Sainsbury, A. Heynen, C.P. Montoya, Behavioral correlates of
hippocampal type 2 theta in the rat, Physiol. Behav. 39 (1987)
513–519.
[34] L.R. Squire, Memory and the hippocampus: a synthesis from finding
with rats, monkeys and humans, Psychol. Rev. 99 (1992) 195–231.
[35] H. van Lier, A.M. Coenen, W.H. Drinkenburg, Behavioral transitions
modulate hippocampal electroencephalogram correlates of open field
behavior in the rat: support for a sensory–motor function of hippo-
campal rhythmical synchronous activity, J. Neurosci. 23 (2003)
2459–2465.
[36] R.A. Velluti, Interactions between sleep and sensory physiology,
J. Sleep Res. 6 (1997) 61–77.
[37] R.A. Velluti, M. Pedemonte, In vivo approach to the cellular
mechanisms for sensory processing in sleep and wakefulness, Cell.
Mol. Neurobiol. 22 (2002) 501–516.
[38] R.A. Velluti, J.L. Pena, M. Pedemonte, Reciprocal actions between
sensory signals and sleep, Biol. Signals Recept. 9 (2000) 297–308.
[39] R.P. Vertes, B. Kocsis, Brainstem–diencephalo–septohippocampal
systems controlling the theta rhythm of the hippocampus, Neuro-
science 81 (1997) 893–926.
[40] O.S. Vinogradova, Hippocampus as comparator: role of the two input
and two output systems of the hippocampus in selection and
registration of information, Hippocampus 11 (2001) 578–598.
[41] G.W. Wallestein, H. Eichenbaum, M.E. Hasselmo, The hippocampus
as an associator of discontiguous events, Trends Neurosci. 21 (1998)
317–323.
[42] J. Winson, Loss of hippocampal theta rhythm results in spatial
memory deficit in the rat, Science 201 (1978) 160–163.
[43] E.R. Wood, P.A. Dudchenko, H. Eichenbaum, The global record of
memory in hippocampal neuronal activity, Nature 397 (1999)
613–616.
Related Documents











