Send Orders for Reprints to [email protected] 150 Current Medicinal Chemistry, 2015, 22, 150-164 Novel Strategies for Preventing Diabetes and Obesity Complications with Natural Polyphenols C. Carpéné* ,a,b,# , S. Gomez-Zorita b , S. Deleruyelle a,b and M.A. Carpéné b a Institute of metabolic and cardiovascular diseases (I2MC), team 3, Institut National de la Santé et de la Re- cherche Médicale, INSERM Unit 1048, Toulouse, France; b Université Paul Sabatier Toulouse 3, Toulouse, France Abstract: During the last years, the list of resveratrol effects has grown in parallel with the number of other members of the polyphenol family described to modulate glucose or lipid handling. In the same time, more than ten human studies on the influence of resveratrol supplementation on two related metabolic diseases, obesity and diabetes, have indicated that impressive beneficial effects co-exist with lack of demonstration of clinical relevance, irrespective of the daily dose ingested (0.075 to 1.5 g per capita) or the number of studied patients. Such contrasting observations have been proposed to depend on the degree of insulin resistance of the patients incorporated in the study. To date, no definitive conclusion can be drawn on the antidiabetic or antiobesity benefits of res- veratrol. On the opposite, studies on animal models of diabesity consistently indicated that resveratrol impairs diverse in- sulin actions in adipocytes, blunting glucose transport, lipogenesis and adipogenesis. Since resveratrol also favours lipoly- sis and limits the production of proinflammatory adipokines, its administration in rodents results in limitation of fat depo- sition, activation of hexose uptake into muscle, improvement of insulin sensitivity, and facilitation of glucose disposal. Facing to a somewhat disappointing extrapolation to man of these promising antidiabetic and antiobesity properties, atten- tion must be paid to re-examine resveratrol targets, especially those attainable after polyphenol ingestion and to re-define the responses to low doses. In this context, human adipocytes are proposed as a convenient model for the screening of "novel" polyphenols that can reproduce, outclass, or reinforce resveratrol metabolic actions, Moreover, the use of combi- nation of polyphenols is proposed to treat diabesity complications in view of recently reported synergisms. Lastly, multidisciplinar approaches are recommended for future investigations, considering the wide range of polyphenol actions that induce body fat reduction, liver disease mitigation, muscle function improvement, cardiovascular and renal protec- tion. Keywords: Insulin resistance, glucose transport, adipogenesis, adipokines, energy balance. # Author’s Profile: Dr Christian Carpéné is an Investigator at the French National Institute of Health and Medical Research (INSERM), in the Cardiometabolic Diseases Institute of Toulouse, France. He reported for the first time a link between I 2 - imidazoline binding sites and monoamine oxidases. He is studying semicarbazide-sensitive amine oxidases, dietary amines and polyphenols having antidiabetic or antiobesity properties. INTRODUCTION Diabetes is one of the major health problems worldwide, and more than 80% of diabetic patients suffer from type 2 diabetes. This type of diabetes is a complication of insulin resistance, a state in which insulin exhibits a reduced ability to enhance glucose utilisation in its major target tissues: skeletal muscle(s) and adipose tissue(s). The accompanying increases in the blood levels of glucose and insulin are the cause of many deleterious complications, including cardio- vascular diseases (retinal injury, hypertension, atherosclero- sis), dyslipidemia, fatty liver and renal diseases. To date, the growing proportion of the adult population being either overweight or obese is involved in the currently increased prevalence of (type 2) non-insulin dependent diabetes melli- tus. As most of the obesity-related complications, diabetes can be linked to excessive fat stores via the disturbances of two main features of adipose tissue: its endocrine function and its metabolic capacity to handle glucose and fatty acids, *Address correspondence to this author at the Institut des Maladies Métaboliques et Cardiovasculaires, I2MC, INSERM U 1048 - Eq n°3, CHU Rangueil, Bat. L4, BP 84225, 31432 Toulouse Cedex 4, France; Tel: (33) 5 61 32 56 36; E-mail: [email protected] the major sources of energy for the body. These two regu- lated processes are summarized below, while it must be kept in mind that other aspects of adipose tissue physiopathology are disturbed under insulin resistance (for further details, please consult [1]): • On the one hand, adipose tissue secretes numerous bioac- tive peptides and factors, collectively known as adipoki- nes, which play important roles in glucose homeostasis, appetite regulation and blood pressure control, but which also participate in the regulation of low-grade tissue and systemic inflammation [2]. • On the other hand, adipose cells activate their glucose uptake mainly upon insulin stimulation, the key hypogly- caemic hormone involved in the regulation of blood glu- cose concentrations within a narrow range around 5-6 mM. Although adipose tissue (AT) is not quantitatively as important as skeletal muscles for peripheral glucose dis- posal, it is of utmost qualitative importance: any defect of insulin responsiveness in adipose depots alters the overall glucose tolerance [3]. In addition, excess of adipose cells plays a key role in lipotoxicity by increasing the release of free fatty acids in situations where there is not a demand from the organism for such energy supply. 1875-533X/15 $58.00+.00 © 2015 Bentham Science Publishers

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Send Orders for Reprints to [email protected]
150 Current Medicinal Chemistry, 2015, 22, 150-164
Novel Strategies for Preventing Diabetes and Obesity Complications with Natural Polyphenols
C. Carpéné*,a,b,#, S. Gomez-Zoritab, S. Deleruyellea,b and M.A. Carpénéb
aInstitute of metabolic and cardiovascular diseases (I2MC), team 3, Institut National de la Santé et de la Re-cherche Médicale, INSERM Unit 1048, Toulouse, France; bUniversité Paul Sabatier Toulouse 3, Toulouse, France
Abstract: During the last years, the list of resveratrol effects has grown in parallel with the number of other members of the polyphenol family described to modulate glucose or lipid handling. In the same time, more than ten human studies on the influence of resveratrol supplementation on two related metabolic diseases, obesity and diabetes, have indicated that impressive beneficial effects co-exist with lack of demonstration of clinical relevance, irrespective of the daily dose ingested (0.075 to 1.5 g per capita) or the number of studied patients. Such contrasting observations have been proposed to depend on the degree of insulin resistance of the patients incorporated in the study. To date, no definitive conclusion can be drawn on the antidiabetic or antiobesity benefits of res-veratrol. On the opposite, studies on animal models of diabesity consistently indicated that resveratrol impairs diverse in-sulin actions in adipocytes, blunting glucose transport, lipogenesis and adipogenesis. Since resveratrol also favours lipoly-sis and limits the production of proinflammatory adipokines, its administration in rodents results in limitation of fat depo-sition, activation of hexose uptake into muscle, improvement of insulin sensitivity, and facilitation of glucose disposal. Facing to a somewhat disappointing extrapolation to man of these promising antidiabetic and antiobesity properties, atten-tion must be paid to re-examine resveratrol targets, especially those attainable after polyphenol ingestion and to re-define the responses to low doses. In this context, human adipocytes are proposed as a convenient model for the screening of "novel" polyphenols that can reproduce, outclass, or reinforce resveratrol metabolic actions, Moreover, the use of combi-nation of polyphenols is proposed to treat diabesity complications in view of recently reported synergisms. Lastly, multidisciplinar approaches are recommended for future investigations, considering the wide range of polyphenol actions that induce body fat reduction, liver disease mitigation, muscle function improvement, cardiovascular and renal protec-tion.
Keywords: Insulin resistance, glucose transport, adipogenesis, adipokines, energy balance. #Author’s Profile: Dr Christian Carpéné is an Investigator at the French National Institute of Health and Medical Research (INSERM), in the Cardiometabolic Diseases Institute of Toulouse, France. He reported for the first time a link between I2-imidazoline binding sites and monoamine oxidases. He is studying semicarbazide-sensitive amine oxidases, dietary amines and polyphenols having antidiabetic or antiobesity properties.
INTRODUCTION Diabetes is one of the major health problems worldwide,
and more than 80% of diabetic patients suffer from type 2 diabetes. This type of diabetes is a complication of insulin resistance, a state in which insulin exhibits a reduced ability to enhance glucose utilisation in its major target tissues: skeletal muscle(s) and adipose tissue(s). The accompanying increases in the blood levels of glucose and insulin are the cause of many deleterious complications, including cardio-vascular diseases (retinal injury, hypertension, atherosclero-sis), dyslipidemia, fatty liver and renal diseases. To date, the growing proportion of the adult population being either overweight or obese is involved in the currently increased prevalence of (type 2) non-insulin dependent diabetes melli-tus. As most of the obesity-related complications, diabetes can be linked to excessive fat stores via the disturbances of two main features of adipose tissue: its endocrine function and its metabolic capacity to handle glucose and fatty acids,
*Address correspondence to this author at the Institut des Maladies Métaboliques et Cardiovasculaires, I2MC, INSERM U 1048 - Eq n°3, CHU Rangueil, Bat. L4, BP 84225, 31432 Toulouse Cedex 4, France; Tel: (33) 5 61 32 56 36; E-mail: [email protected]
the major sources of energy for the body. These two regu-lated processes are summarized below, while it must be kept in mind that other aspects of adipose tissue physiopathology are disturbed under insulin resistance (for further details, please consult [1]): • On the one hand, adipose tissue secretes numerous bioac-
tive peptides and factors, collectively known as adipoki-nes, which play important roles in glucose homeostasis, appetite regulation and blood pressure control, but which also participate in the regulation of low-grade tissue and systemic inflammation [2].
• On the other hand, adipose cells activate their glucose uptake mainly upon insulin stimulation, the key hypogly-caemic hormone involved in the regulation of blood glu-cose concentrations within a narrow range around 5-6 mM. Although adipose tissue (AT) is not quantitatively as important as skeletal muscles for peripheral glucose dis-posal, it is of utmost qualitative importance: any defect of insulin responsiveness in adipose depots alters the overall glucose tolerance [3]. In addition, excess of adipose cells plays a key role in lipotoxicity by increasing the release of free fatty acids in situations where there is not a demand from the organism for such energy supply.
1875-533X/15 $58.00+.00 © 2015 Bentham Science Publishers

Using Natural Polyphenols in Diabetes and Obesity Current Medicinal Chemistry, 2015, Vol. 22, No. 1 151
Moreover, insulin resistance, by increasing the demand on the pancreatic �-cell to secrete high amounts of insulin, plays an important role in the progressive �-cell failure of type 2 diabetes while it also alters various other organs and tissues [4]. However, this review will focus reader's attention on peripheral glucose utilisation, especially in fat tissues and muscles, since it is the main metabolic defect occurring un-der insulin resistance, which is responsible of excessive hy-perglycaemia. Therefore, enhancing insulin sensitivity and especially improving its stimulation of glucose uptake into target tissues is critical for limiting the deleterious effects of insulin resistance. Since liver is rather limiting its glucose production in response to insulin, and since hepatic glucose uptake mainly depends on the GLUT2 carrier (which works proportionally with blood glucose level), the liver must be considered different from the so-called "insulin-sensitive tissues" (mainly skeletal or cardiac muscle and AT), regard-ing glucose transport regulation. Limiting too high and/or too long excursions above the normoglycaemia can be obtained by the activation of the regulated transport of glucose into these target tissues (an approach also applying for type 1 diabetic patients with defective insulin supply). To this aim, it is necessary to directly increase at the cell surface the en-zymatic activity of the glucose transporters of the GLUT family and/or to correct of their decreased insulin respon-siveness.
Physical exercise is known to increase glucose uptake in muscle, almost in an insulin-independent manner, though it is not the case for AT. Consequently, the current policy that seeks to combat metabolic and cardiovascular diseases policy by reducing sedentary life and modifying behaviour and food habits of citizens is highly recommendable for prevention, public health and economics. However, physi-cal exercise is definitely one of the best modes of preven-tion, but it is often difficult to manage for many diabetic and obese individuals and remains a disease treatment that is barely able to lead to complete recovery. That explains why diabetic patients require long-term drug treatment and/or nutritional care, including the 90 % of them who were previously obese.
Unfortunately, the pharmacological treatments of diabe-tes and obesity have not improved the global situation as much as expected, due to adverse reactions found with many of the antidiabetic or antiobesity drugs prescribed so far. Since these drugs have to be administered during very long periods to treat chronic cardiovascular and metabolic diseases herein summarized as "diabesity", it is easily con-ceivable that they have increased risk to develop unwanted adverse effects when compared to drugs requiring single or short-term administration. For example, in the treatment of obesity, sibutramine and rimonabant were withdrawn re-garding to the risk of hypertension and depression, respec-tively. For diabetes care, the glucose lowering effect of the sulfonylureas is not durable enough for treating throughout lifespan [4], while restrictions have appeared on the use of the "glitazones" regarding to possible cardiac effects. Simi-larly, serious doubts about pancreatic risk (acute pancreati-tis or cancer) have been raised for novel antidiabetic drugs acting as receptor agonists for the glucagon-like peptide (GLP-1) or as inhibitors of its degradation (DPP4 inhibi-tors). In this context, various compounds of plant origin have been studied in the treatment of experimental models
of obesity, diabetes and dyslipidemia [5-8]. Currently, most of these phytochemicals or related food items are consid-ered as a good alternative to antidiabetic and/or antiobesity drugs, especially by the companies selling dietary supple-ments. In the case of resveratrol, the emblematical "gold standard" of natural polyphenols, the results of basic re-search in animal models were in total agreement with its beneficial properties detected by epidemiological studies (Mediterranean diet or "french paradox"): prolonged inges-tion of this agent may limit the complications of diabetes [9, 10] or obesity [11, 12]. Resveratrol has been consis-tently demonstrated to exert numerous beneficial effects in animal models of obesity or diabetes, as already reviewed in [13]. By contrast, demonstrative clinical data of resvera-trol therapeutic interest in human diabetes and obesity are extremely limited.
The review will begin by a rapid presentation of the die-tary polyphenol family, followed by an overview of recent data from human studies about the cardiometabolic effects of resveratrol. Then, novel interpretations will be drawn from the numerous studies of the beneficial effects of resveratrol in rodents with obesity and diabetes. Next, in vitro effects of resveratrol and natural polyphenols will be re-examined with a special attention to the adipocyte model. After that, novel perspectives to improve the benefits/risk ratio of the use of polyphenols in the treatment or prevention of diabesity will be proposed, owing to the large number of members in this family, and to the recently tested combinations with other agents. Lastly, a multidisciplinary approach will be recom-mended for future studies, on the basis of the growing list of resveratrol actions in experimental models: body fat reduc-tion, mitigation of liver disease, inhibition of inflammation and of platelet activation, maintenance of bone mass, cardio-protection, neuroprotection, etc.
THE LARGE FAMILY OF DIETARY POLYPHENOLS IN A BRIEF DIGEST
Polyphenol family can be classified in 5 classes: stil-benes, flavonoids, lignans, phenolic acids, and other poly-phenols. These classes are divided in subclasses, and num-bering their members is rather difficult since these natural compounds are often found in plants under the form of gly-cosides, i.e. attached to sugars of more or less complex com-position.
The most recognized polyphenol, namely resveratrol, belongs to the stilbene class. Stilbenes are composed of two benzene rings linked by two carbones. (Fig. 1) shows the trans-resveratrol only. It must be noted that this form counts only for 21 % of the mean daily ingestion of res-veratrol in man, according to the epidemiological EPIC study (European Prospective Investigation into Cancer) [14]. The remaining ingested forms being cis-resveratrol (6 %) or the glycoside forms, namely the trans-piceid (54 %) and cis-piceid (19 % of the mean daily ingestion of resveratrol). All these components of the overall dietary resveratrol (averaging from 0.1 to 1 mg/d) are essentially brought by red wine consumption. In fact, resveratrol is a phytoalexin (a compound produced by plants in response to infection or other stresses) present in grapes and then found in wine, and can been found to a lower extend in various edible plants, as reviewed in: [10]. Two other examples of

152 Current Medicinal Chemistry, 2015, Vol. 22, No. 1 Carpéné et al.
the Stilbene class are reported: pterostilbene and piceatan-nol. Other agents studied in the field of obesity/diabetes research belong to the wide class of flavonoids: they are structurally based on a skeleton of two phenyl rings linked by a propane bond, with multiple cyclisation and substitu-tion possibilities, leading to polycyclic molecules. The most known representatives are flavonols (quercetin, kaempferol, myricetin), isoflavonoids (genistein, daidzein), flavanones (hesperidin, hesperetin, naringenin, naringin), flavanols (catechins, procyanidins, theaflavins), anthocyan-ins (cyanidin), chalcones and dihydrochalcones (phloridzin also known as phlorizin) (Fig. 1). While the latter com-pound is a well-established inhibitor of sodium/glucose transporters, reducing intestinal absorption or renal reab-sorption of glucose and consequently lowering blood glu-cose levels, the other flavonoids (especially those which are not methylated) have been reported to inhibit �-glucosidases [15, 16]. Glucosidase inhibition lowers the rate of glucose absorption through delayed carbohydrate digestion time, and is useful in reducing the postprandial hyperglycemia, the major origin of diabetic complications. Lignans have a chemical structure based on two mono-lignol units: (C6-C3)2. Lignans belong to the classes of phytoestrogens and have as representative member pro-posed for dietary supplementation sesamin, found in seeds. Phenolic acids are not necessarily poly-phenolic. Neverthe-less, they are based on the phenol structure and comprise a carboxylic acid group. In the phenolic acid class, several members have been studied for their antidiabetic and anti-oxidant properties: gallic acid, chlorogenic acid, and caf-feic acid (do not make confusion with caffeine, a xanthine alkaloid also found in seeds, fruits and leaves). Notewor-thy, caffeic acid and derivatives have been recently shown to substantially inhibit �-glucosidases [16]. Lastly, the "other polyphenols" class is the most diverse class, encom-passing alkylphenols, coumarins, phenolic terpenes, cur-cuminoids, etc. Curcumin is an example reported in (Fig. 1)just to illustrate that a 7-carbon chain can link the two phe-nols in such natural products. Its antiobesity effects have been reviewed in [17].
To obtain much more data on the hundreds of polyphe-nols described in various foods or drinks and on their me-tabolites generated upon ingestion in animals or man, the Phenol-Explorer database [18] is publicly available and can be found online at: http://www.phenol-explorer.eu.
To have an almost exhaustive list of the beneficial effects for each bioactive compound of the polyphenol family, please consult [6], since the present review will treat them rather collectively, highlighting only several examples.
CLINICAL STUDIES OF RESVERATROL EFFECTS ON GLUCOSE HANDLING: AN OVERVIEW
Only limited clinical data are available concerning res-veratrol potential effects on insulin sensitivity and glucose handling. Several of the human studies published so far about the influence of orally administered resveratrol either under the form of pills, tablets or supplemented via en-riched plant extracts, are consistent with previous observa-tions made on animal models of obesity or diabetes. They confirm the beneficial effects of the stilbene on glucose disposal.
OH
OHHO
Resveratrol
OH
OO
CH3 CH3
Pterostilbene
HO
OH
OH
OH
Piceatannol
HO
OH
O OH
OH
HO
O
Quercetin
HO
HO
O OH
OH
Kaempferol
HO
O
O
OH
DaidzeinHO
O
O
OH
Genistein
OH
O
HO
OH
OH
HOO
HO OH
O
OH
Phloridzin
OH
OHHO
OHO
Gallic acid
HO
OH
HO
O
OH
OH
O
HO
O
Chlorogenic acid
OH
OH3C
HO
OH3C
O O
Curcumin
OH
HO
OHO
Caffeic acid
Fig. (1). Structural formulae of several dietary polyphenols. Stil-benes are represented by resveratrol, pterostilbene and piceatannol. Quercetin, kaempferol, daidzein, genistein and phloridzin are typi-cal molecules of the flavonoid class. Also shown are representatives of phenolic acids (gallic, chlorogenic and caffeic acid acid) and of curcuminoids (curcumin).
A study performed in Hungary with type 2 diabetic pa-tients reported a decreased insulin resistance in those of 19 adults with impaired glucose tolerance (IGT) who received resveratrol for 4 weeks when compared to placebo [19]. An-other pilot study was performed in USA with open-label administration for 4 weeks in 10 older adults with IGT re-ceiving either 1, 1.5, or 2 g resveratrol/d. The fasting plasma glucose was unchanged, but its peak-increase after the inges-tion of standard mixed meal and the overall hyperglycaemic excursion declined after treatment when compared to control [20]. In the Netherlands, a group of 11 obese middle-aged

Using Natural Polyphenols in Diabetes and Obesity Current Medicinal Chemistry, 2015, Vol. 22, No. 1 153
men were treated with resveratrol (0.15 g/d) for 1 month in a randomized double-blind crossover study. An improvement in a variety of metabolic parameters was reported, including decreased circulating glucose and insulin and modestly en-hanced insulin sensitivity. Systolic blood pressure, hepatic fat and markers of inflammation were reduced, together with reduced leptin levels. Thus, the authors qualified the resvera-trol effects as "calorie restriction-like"[21], a name previ-ously used for animal studies [22, 23]. Of note, the activities or levels of resveratrol intracellular targets were increased in skeletal muscle, namely the adenosine monophosphate-activated protein kinase (AMPK), the deacetylating enzyme sirtuin-1 (SIRT-1), and the peroxisome proliferator-activated receptor � coactivator-1� (PGC-1�) [21]. Furthermore, lipolysis was modestly decreased in adipose tissue. Addi-tionally, the same resveratrol supplementation was able in one month to decrease the proportion of large adipocytes and to increase the small adipocytes of abdominal subcutaneous adipose tissue together with changes found by microarray analysis that suggested an enhanced adipogenesis, according to the authors [24]. This latter observation in humans is puz-zling since the polyphenols have been constantly described to inhibit in vitro adipocyte differentiation in murine and human models, at least when tested at 10-100 �M [25]. However, these clinical observations [24] converge with basic research demonstrating resveratrol-induced lipid breakdown in fat cells [26].
The effects of resveratrol supplementation were studied in a double-blind manner on a larger cohort in Iran (n = 66) and it has been observed that, when given at 1 g/day for one month and half, the polyphenol induced a decrease of fasting blood glucose and insulin in type 2 diabetic patients [27]. More importantly, glycated haemoglobin (HbA1c), a recog-nized circulating marker of bouts of hyperglycaemia, was reduced after resveratrol. Even a lower dose of orally in-gested resveratrol (0.25 g/d, for 3 months) has been reported to ameliorate the glycaemic control in 62 diabetic patients enrolled in India for an open-label, randomized trial, which showed a modest decrease in blood glucose levels and in HbA1c in the treated group [28].
However, not all the human clinical trials performed so far have reported so significant beneficial effects of resvera-trol on insulin sensitivity and glucose handling in obese sub-jects. A randomized, double-blind, crossover design, testing the influence of one-month resveratrol supplementation (0.15 g/d) on incretin hormone responses in 10 obese sub-jects failed to demonstrate any impact on fasting plasma concentrations but indicated that the supplementation solely limited the postprandial glucagon elevation [29]. Similarly, resveratrol did not modulate insulin sensitivity when admin-istered to 8 men at 1 to 2 g/d during two weeks, while it ex-hibited a tendency to limit overproduction of lipoproteins involved in hypertriglyceridemia [30].
When clinical studies included relatively healthy (nondiabetic) subjects, more discordant results were ob-tained. One of these elegant studies, using a large panel of metabolic tests in a randomized, double-blind design per-formed in Denmark, did not bring evidence of any improve-ment of glucose handling [31]. Indeed, the oral resveratrol given at 1.5 g/d in obese old patients weighing approx 107-
115 kg - but otherwise healthy - did not modulate the en-dogenous glucose production, the glucose turnover and its oxidation rate. Again, it did not improve insulin sensitivity (tested as hyperinsulinemic euglycaemic clamp) [31]. Re-garding lipid metabolism, fat content was not reduced, while energy expenditure and lipid oxidation were not increased, leading the authors to raise doubt about the justification of resveratrol as a human nutritional supplement in metabolic disorders. In fact, one can consider that its rather difficult to substantiate the antidiabetic potential of resveratrol in volun-teers who exhibited unaltered glucose tolerance. Relative to the obesity aspect, 4-week treatment with resveratrol did not exhibit any of the improvement that could be expected in the selected patients, who where overtly obese. In fact, it is still needed to establish whether the supplementation with poly-phenols is useful for prevention in overweight or prediabetic consumers and whether it can delay both the onset and sever-ity of hyperglycaemia in diabesity.
Other trials, studying lower doses of resveratrol, failed to bring evidence of any beneficial action. Resveratrol, at 0.075 g/day during 12 weeks in nonobese postmenopausal women with normal glucose tolerance, failed to increase insulin sen-sitivity and to demonstrate significant metabolic effects [32]. In a study on the effects of extracts of Polygonum cuspida-tum (also known as Fallopia japonica or Japanese knotweed, a plant particularly rich in resveratrol, piceid, flavonoids and phenolic compounds) [33], the planned protocol resulted in the ingestion of 0.04 g resveratrol/d during 6 weeks in 10 nonobese nondiabetic young subjects. A decrease of TNF�,a proinflammatory cytokine that is deleterious for insulin sensitivity was evidenced after treatment in plasma, together with lowered levels of IL-6 and C-reactive protein. Other markers of antiinflammatory and antioxidant actions were detected but not any change more directly related to glucose handling.
Lastly, a meta-analysis of 11 of the randomized con-trolled trials published so far concluded that resveratrol con-sumption significantly reduces the circulating levels of fast-ing glucose, insulin and haemoglobin A1c, and diminishes insulin resistance in diabetic subjects [34]. However, the meta-analysis also pointed out that resveratrol does not affect glycaemic control of non-diabetic volunteers, as stated above [34]. Thus, the beneficial action of resveratrol has not been univocally proven in recent human studies, at least regarding to its usefulness as a preventive agent delaying the onset of insulin resistance. An influence of diabetes on the pharma-cokinetic behavior of polyphenols has been proposed [8], and might play a role in the reported discrepancies about resveratrol action in type 2 diabetic patients vs normoglyce-mic individuals. Consequently, the demonstration of resvera-trol prophylactic properties needs much more complex ap-proaches than the pilot studies or the clinical studies achieved so far. In fact, the promise of resveratrol to combat diabesity is far from being totally blunted although its ad-ministration in humans did not always result in the impres-sive recovery reported in animal models. Albeit this aspect is well commented in [35], it must be repeated here that the "negative" results from human studies do not totally disagree with basic research on animal models, since it has been es-tablished that resveratrol is more active in models with dis-turbed glucose handling than in insulin normoresponsive

154 Current Medicinal Chemistry, 2015, Vol. 22, No. 1 Carpéné et al.
rodents [36]. In this context, the delay in the dissemination of observations from pharmaceutical companies that aimed at improving the polyphenol bioavailability and that have not completed yet the valorisation of their own probe-of-concept, might indicate that resveratrol does not easily con-firm its promising clinical utility. Hopefully, resveratrol seems to be well tolerated and almost no marked toxicity has been reported. Thus, the major pitfalls in the extrapolation to man of the positive effects found in animal models are re-lated to the following questions: • What is the optimal dose? • What are the efficient targets in rodents and are they func-
tional as well in humans? • Is resveratrol bioavailability compatible with efficient
access to such targets?
A while after being characterized as antioxidants [37], phenolic coumponds have been recognized for their plei-otropic effects. In the case of resveratrol, many of the (rare) dose-responses reported so far were not merely sigmoidal, even in simple in vitro assays, as it should be the case for any given selective receptor agonist. This indicates that the polyphenol targets are multiple, and that their activation or inhibition depends on the doses and models studied. Addi-tional basic studies are therefore needed to solve such dose-response issues. Meanwhile, a correct dose translation from rats or mice to humans is required, as already proposed [5]. The duration and pulsatility of exposure to the polyphenols is also of importance, especially in relation with the hormesis hypothesis [10, 38]. Hence, several findings observed in pio-neering studies cannot be considered as dogmas and unex-pected novel divergences might be observed when further comparing human and animal studies. This looks to be the case for the AMPK inhibition observed with 100 �M res-veratrol in human skeletal muscle cells [39], which appar-ently contrast with the AMPK activation found in muscle of resveratrol-treated rodents [40]. Another shadow in the dem-onstration of resveratrol therapeutic insterest is related to the pharmacokinetics and the metabolism of the stilbene [41, 42], which may affect its efficacy according to factors that deserve to establish. In this view, further studying the recip-rocal interplay between resveratrol and the intestinal micro-biote should be of interest [43]. Meanwhile, as documented above, it must be kept in mind that resveratrol is only one of the members of the polyphenol family and that improvement of the benefit/risk ratio for treating diabesity can be obtained by searching other candidates within the other plant-derived polyphenols.
NOVEL INTERPRETATIONS OF THE BENEFICIAL EFFECTS OF POLYPHENOLS EVIDENCED IN RO-DENTS WITH OBESITY AND DIABETES
Resveratrol has demonstrated promising effects on glu-cose metabolism improvement and on fat deposition limita-tion in rodent models essentially since 2006, when two inde-pendent studies led to similar conclusions: the natural stil-bene improves health in mice with obesity and diabetes in-duced by a high fat diet, via the activation of SIRT-1, PGC-1�, and AMPK [22, 44]. The subsequent beneficial effects on oxidative phosphorylation and mitochondrial biogenesis
in muscle were then confirmed by various approaches in diabetic and obese rodents [23, 45], but it still remains un-clear whether they are dependent on a SIRT-1- mediated activation of AMPK [46] or whether it is the AMPK activa-tion by the polyphenol which indirectly activates SIRT-1 [40]. Also widely recognized was the demonstration of an increased capacity of muscle to transport glucose, argued either by enhanced expression of insulin-sensitive glucose transporters (GLUT4) [45] and by increased responsiveness to insulin [23]. An improvement of the diabetic conditions was obtained with various doses, while influence on nondia-betic nonobese rodents was questionable [36, 47] since not all the reports indicated that the polyphenol enhances insulin sensitivity in "healthy" laboratory rodents [48]. Other bene-ficial effects of resveratrol supplementation were reported in obese and insulin-resistant rodents, such as reduction of liver steatosis [49-51] or lowering of hypertension [49]. Again, no clear dose-dependency was evidenced, and the maximum of improvement of the disturbances studied was not obtained with the maximal dose tested [11, 50].
Albeit not reducing in all circumstances the body weight gain, resveratrol was repeatedly evidenced to limit fat white deposition in visceral and subcutaneous adipose tissues [12, 48, 51-53]. Resveratrol also promotes brown fat activation [48]. Such effects cannot be qualified as insulin-like, since the pancreatic hormone tends to increase fat deposition and to transform brown adipose tissue into a less thermogenic white fat depot. Thus, a question raised by such observations is: how can resveratrol improve the overall glucose disposal in the organism while it reduces fat accumulation in the adi-pocytes? The body generally stores excessive calorie intake under the form of triglycerides by using glucose and either circulating or de novo synthesized free fatty acids in a proc-ess named lipogenesis. In the case of an inhibition of the latter in AT, an adaptation often occurs, consisting in ectopic fat deposition, such as liver steatosis (a phenomenom that does not occur with resveratrol since hepatic lipids are re-duced). Other adaptative responses may encompass in-creased energy expenditure and decreased metabolic effi-ciency. These two events are far for being excluded under resveratrol supplementation. Of note, calorie restriction is modulating these physiological processes in the opposite direction, in order to maintain energy balance as close as possible to the equilibrium. In this view, resveratrol is unable to potentiate the AT loss induced by energy restriction [54], arguing therefore that it is not simply a "calorie-restriction mimetic" [55].
Considering increased energy consumption as one of the responses to resveratrol, it can be easily stated that it occurs mainly in skeletal muscle or in brown AT in ro-dents. Taking into account that brown AT is hardly func-tional in humans, we therefore propose that ongoing re-search efforts should be focused on the differential effects of resveratrol on muscle cells vs white adipose cells. In other words, resveratrol must be considered as a partition-ing agent dispatching the energy flux to catabolism in mus-cle rather than to storage in AT. Curiously, other AMPK activators are already recognized for such properties [56] and used (or misused?) by athletes.
Using Natural Polyphenols in Diabetes and Obesity Current Medicinal Chemistry, 2015, Vol. 22, No. 1 155
A RE-EXAMINATION OF RESVERATROL IN VITROEFFECTS WITH SPECIAL ATTENTION TO THE ADIPOCYTE MODEL
Among the pleitropic effects of polyphenols [57], their actions on cultured preadipocytes or on mature fat cells iso-lated from normoglycemic and obese/diabetic models have led to the univocal conclusion that they limit lipid accumula-tion, at least when tested at 10-100 �M. Therefore, short- and long-term effects of polyphenols on fat cells will be re-examined below since they will be proposed thereafter as a basis for further screening studies aiming at identifying other molecules that can reproduce or even improve the metabolic actions of the most studied resveratrol. Such novel ap-proaches are still needed since the extrapolation to man of the oral antiobesity and antidiabetic properties of polyphe-nols is not definitely validated. Again, it is worth mentioning that studying the effects of lower doses has not been suffi-ciently addressed.
Understanding how insulin recruits the glucose carriers GLUT4 at the cellular level and activate glucose consump-tion is critically important for seeking solutions to diabetes. In fact, hexoses are hydrophilic molecules that can cross plasma membrane only via transporters of the GLUT family (12 genes with distinct expression profile and biochemically different products). Among them, GLUT4 is usually hidden in intracellular vesicles and becomes functional only once translocated to cell surface, essentially upon insulin activa-tion. Since insulin is also a growth factor that regulates many anabolic processes, studying other insulin-sensitive re-sponses is also instrumental for preventing the complications other than hyperglycaemia occurring in insulin resistance and obesity. In this way, the influence of several polyphenols will be successively discussed for various insulin-regulated processes in the adipocyte: glucose transport, lipogenesis, lipolysis, adipogenesis and adipokine secretion.
Fat cell is a convenient model to measure glucose trans-port, a key event for increasing glucose disposal in response to insulin, which is also the first step for lipogenesis or glycogen synthesis. Measuring such rapid biological event consists in quantifying the incorporation (on 5 to 10-min periods) of a non-metabolizable glucose analogue into intact cells or tissues. The labeled 2-deoxyglucose (2-DG) is usually chosen for such uptake assays [58, 59]. In AT, the response is highly sensitive to acute insulin activation, and the fat cells, freshly isolated by collagenase or liberase digestion, can be distributed in numer-ous vials to test in parallel various doses, conditions or agents. Muscles or cardiomyocytes also respond to insulin, but do not permit to deal with the same elevated number of parallel ob-servations. Albeit this latter disadvantage can be overcome in many basic research laboratories having experience on meta-bolic exploration, the muscle biopsies are invasive in man, and difficult to obtain, while human fat cells can be obtained as surgical waste from patients undergoing liposuction or lipec-tomy in plastic surgery departments, with easier approval of appropriate ethics committees.
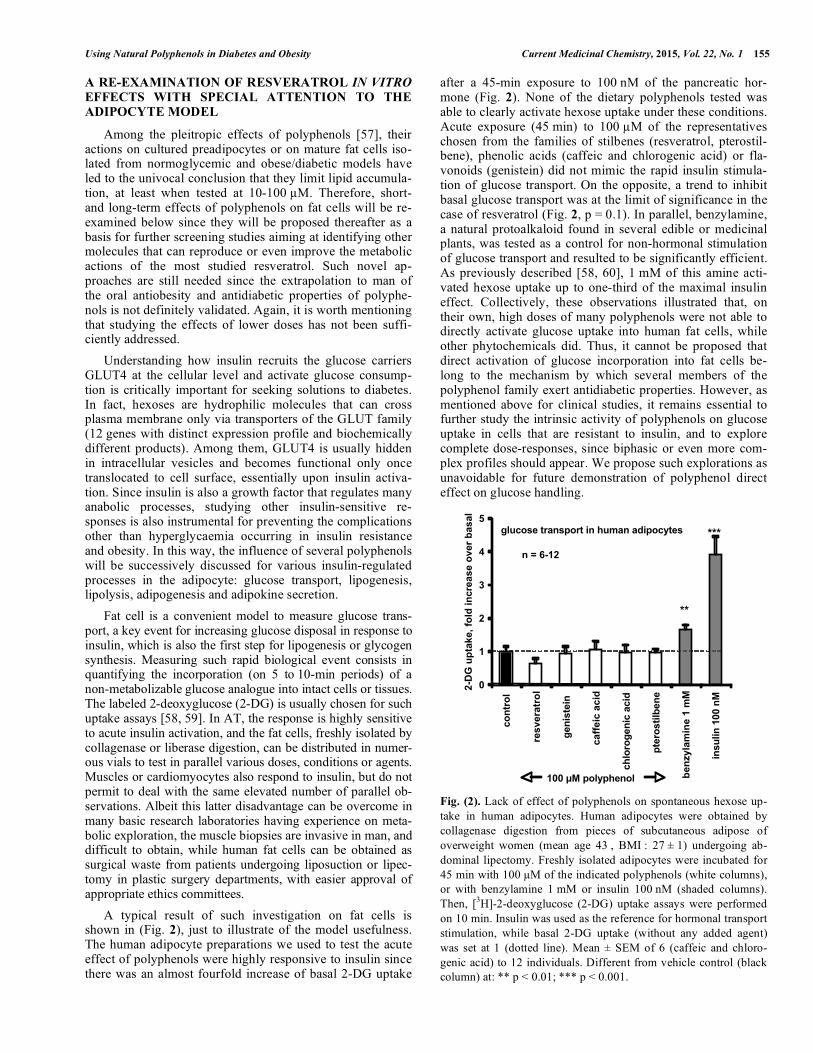
A typical result of such investigation on fat cells is shown in (Fig. 2), just to illustrate of the model usefulness. The human adipocyte preparations we used to test the acute effect of polyphenols were highly responsive to insulin since there was an almost fourfold increase of basal 2-DG uptake
after a 45-min exposure to 100 nM of the pancreatic hor-mone (Fig. 2). None of the dietary polyphenols tested was able to clearly activate hexose uptake under these conditions. Acute exposure (45 min) to 100 �M of the representatives chosen from the families of stilbenes (resveratrol, pterostil-bene), phenolic acids (caffeic and chlorogenic acid) or fla-vonoids (genistein) did not mimic the rapid insulin stimula-tion of glucose transport. On the opposite, a trend to inhibit basal glucose transport was at the limit of significance in the case of resveratrol (Fig. 2, p = 0.1). In parallel, benzylamine, a natural protoalkaloid found in several edible or medicinal plants, was tested as a control for non-hormonal stimulation of glucose transport and resulted to be significantly efficient. As previously described [58, 60], 1 mM of this amine acti-vated hexose uptake up to one-third of the maximal insulin effect. Collectively, these observations illustrated that, on their own, high doses of many polyphenols were not able to directly activate glucose uptake into human fat cells, while other phytochemicals did. Thus, it cannot be proposed that direct activation of glucose incorporation into fat cells be-long to the mechanism by which several members of the polyphenol family exert antidiabetic properties. However, as mentioned above for clinical studies, it remains essential to further study the intrinsic activity of polyphenols on glucose uptake in cells that are resistant to insulin, and to explore complete dose-responses, since biphasic or even more com-plex profiles should appear. We propose such explorations as unavoidable for future demonstration of polyphenol direct effect on glucose handling.
Fig. (2). Lack of effect of polyphenols on spontaneous hexose up-take in human adipocytes. Human adipocytes were obtained by collagenase digestion from pieces of subcutaneous adipose of overweight women (mean age 43 , BMI : 27 ± 1) undergoing ab-dominal lipectomy. Freshly isolated adipocytes were incubated for 45 min with 100 �M of the indicated polyphenols (white columns), or with benzylamine 1 mM or insulin 100 nM (shaded columns). Then, [3H]-2-deoxyglucose (2-DG) uptake assays were performed on 10 min. Insulin was used as the reference for hormonal transport stimulation, while basal 2-DG uptake (without any added agent) was set at 1 (dotted line). Mean ± SEM of 6 (caffeic and chloro-genic acid) to 12 individuals. Different from vehicle control (black column) at: ** p < 0.01; *** p < 0.001.
glucose transport in human adipocytes
0
1
2
3
4
5co
ntro
l
resv
erat
rol
geni
stei
n
caffe
ic a
cid
chlo
roge
nic
acid
pter
ostil
bene
benz
ylam
ine
1 m
M
insu
lin 1
00 n
M
100 µM polyphenol
2-D
G u
ptak
e, fo
ld in
crea
se o
ver b
asal
n = 6-12
**
***
156 Current Medicinal Chemistry, 2015, Vol. 22, No. 1 Carpéné et al.
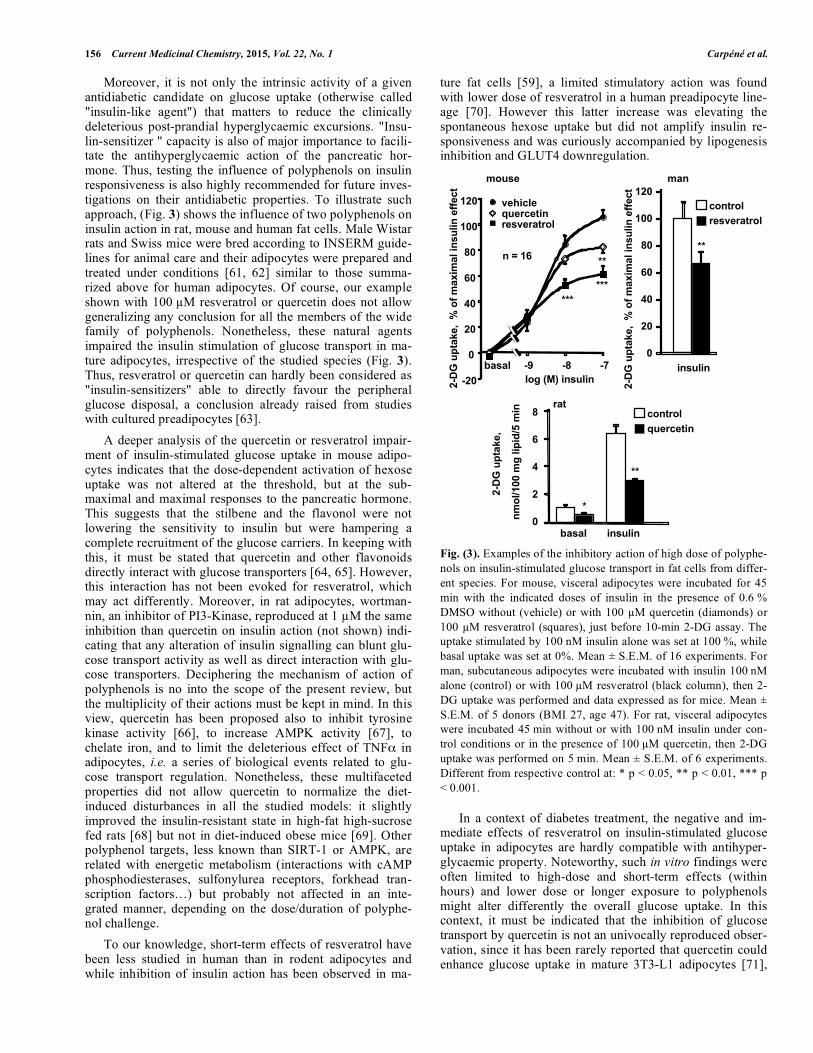
Moreover, it is not only the intrinsic activity of a given antidiabetic candidate on glucose uptake (otherwise called "insulin-like agent") that matters to reduce the clinically deleterious post-prandial hyperglycaemic excursions. "Insu-lin-sensitizer " capacity is also of major importance to facili-tate the antihyperglycaemic action of the pancreatic hor-mone. Thus, testing the influence of polyphenols on insulin responsiveness is also highly recommended for future inves-tigations on their antidiabetic properties. To illustrate such approach, (Fig. 3) shows the influence of two polyphenols on insulin action in rat, mouse and human fat cells. Male Wistar rats and Swiss mice were bred according to INSERM guide-lines for animal care and their adipocytes were prepared and treated under conditions [61, 62] similar to those summa-rized above for human adipocytes. Of course, our example shown with 100 �M resveratrol or quercetin does not allow generalizing any conclusion for all the members of the wide family of polyphenols. Nonetheless, these natural agents impaired the insulin stimulation of glucose transport in ma-ture adipocytes, irrespective of the studied species (Fig. 3). Thus, resveratrol or quercetin can hardly been considered as "insulin-sensitizers" able to directly favour the peripheral glucose disposal, a conclusion already raised from studies with cultured preadipocytes [63].
A deeper analysis of the quercetin or resveratrol impair-ment of insulin-stimulated glucose uptake in mouse adipo-cytes indicates that the dose-dependent activation of hexose uptake was not altered at the threshold, but at the sub-maximal and maximal responses to the pancreatic hormone. This suggests that the stilbene and the flavonol were not lowering the sensitivity to insulin but were hampering a complete recruitment of the glucose carriers. In keeping with this, it must be stated that quercetin and other flavonoids directly interact with glucose transporters [64, 65]. However, this interaction has not been evoked for resveratrol, which may act differently. Moreover, in rat adipocytes, wortman-nin, an inhibitor of PI3-Kinase, reproduced at 1 �M the same inhibition than quercetin on insulin action (not shown) indi-cating that any alteration of insulin signalling can blunt glu-cose transport activity as well as direct interaction with glu-cose transporters. Deciphering the mechanism of action of polyphenols is no into the scope of the present review, but the multiplicity of their actions must be kept in mind. In this view, quercetin has been proposed also to inhibit tyrosine kinase activity [66], to increase AMPK activity [67], to chelate iron, and to limit the deleterious effect of TNF� in adipocytes, i.e. a series of biological events related to glu-cose transport regulation. Nonetheless, these multifaceted properties did not allow quercetin to normalize the diet-induced disturbances in all the studied models: it slightly improved the insulin-resistant state in high-fat high-sucrose fed rats [68] but not in diet-induced obese mice [69]. Other polyphenol targets, less known than SIRT-1 or AMPK, are related with energetic metabolism (interactions with cAMP phosphodiesterases, sulfonylurea receptors, forkhead tran-scription factors…) but probably not affected in an inte-grated manner, depending on the dose/duration of polyphe-nol challenge.
To our knowledge, short-term effects of resveratrol have been less studied in human than in rodent adipocytes and while inhibition of insulin action has been observed in ma-
ture fat cells [59], a limited stimulatory action was found with lower dose of resveratrol in a human preadipocyte line-age [70]. However this latter increase was elevating the spontaneous hexose uptake but did not amplify insulin re-sponsiveness and was curiously accompanied by lipogenesis inhibition and GLUT4 downregulation.
Fig. (3). Examples of the inhibitory action of high dose of polyphe-nols on insulin-stimulated glucose transport in fat cells from differ-ent species. For mouse, visceral adipocytes were incubated for 45 min with the indicated doses of insulin in the presence of 0.6 % DMSO without (vehicle) or with 100 �M quercetin (diamonds) or 100 �M resveratrol (squares), just before 10-min 2-DG assay. The uptake stimulated by 100 nM insulin alone was set at 100 %, while basal uptake was set at 0%. Mean ± S.E.M. of 16 experiments. For man, subcutaneous adipocytes were incubated with insulin 100 nM alone (control) or with 100 �M resveratrol (black column), then 2-DG uptake was performed and data expressed as for mice. Mean ± S.E.M. of 5 donors (BMI 27, age 47). For rat, visceral adipocytes were incubated 45 min without or with 100 nM insulin under con-trol conditions or in the presence of 100 �M quercetin, then 2-DG uptake was performed on 5 min. Mean ± S.E.M. of 6 experiments. Different from respective control at: * p < 0.05, ** p < 0.01, *** p < 0.001.
In a context of diabetes treatment, the negative and im-mediate effects of resveratrol on insulin-stimulated glucose uptake in adipocytes are hardly compatible with antihyper-glycaemic property. Noteworthy, such in vitro findings were often limited to high-dose and short-term effects (within hours) and lower dose or longer exposure to polyphenols might alter differently the overall glucose uptake. In this context, it must be indicated that the inhibition of glucose transport by quercetin is not an univocally reproduced obser-vation, since it has been rarely reported that quercetin could enhance glucose uptake in mature 3T3-L1 adipocytes [71],
***
mouse
2-D
G u
ptak
e, %
of m
axim
al in
sulin
effe
ct
vehiclequercetinresveratrol
n = 16
***
-20
0
20
40
60
80
100
120
basal -9 -8 -7log (M) insulin
**
man
0
20
40
60
80
100
120
insulin
controlresveratrol
2-D
G u
ptak
e, %
of m
axim
al in
sulin
effe
ct
**
controlquercetin
nmol
/100
mg
lipid
/5 m
in
6
8
2
4
0insulinbasal
rat
**
*2-
DG
upt
ake,

Using Natural Polyphenols in Diabetes and Obesity Current Medicinal Chemistry, 2015, Vol. 22, No. 1 157
(as well as kaempferol), without being proadipogenic. Simi-larly, resveratrol was not found to reduce the insulin-stimulated glucose transport in rat adipocytes [72] or cul-tured human preadipocytes [70]. Nevertheless, the most con-vincing in vitro argument agreeing with in vivo antidiabetic action is that resveratrol and other polyphenols directly acti-vate glucose uptake into skeletal muscle, at least in cultured myocytes either of the L6 [73-75] or the C2C12 lineages [6]. Increased consumption of oxygen in muscle fibres [44] in resveratrol-treated mice is believed to be concomitant with increased glucose consumption. An increase of the phos-phorylated forms of insulin-receptor-substrate (IRS-1), and of AKT was found in muscles from insulin-resistant mice treated with resveratrol [76]. The resveratrol-induced glucose uptake was dependent on AMPK activation, but independent from phosphatidylinositol-3 kinase (PI3K) signalling [6], and sensitive to nicotinamide or splitomycin, two poorly selective sirtuin inhibitors. Though GLUT4 translocation has been questioned in response to revesratrol, it has been re-cently demonstrated in response to piceatannol, another stil-bene [75]. Accordingly, in vivo supplementation with phy-tosterols seems to traduce that AMPK activation is involved in the activation of glucose uptake in muscle cells, since the improvement of glucose tolerance by resveratrol is lost in AMPK-deficient mice [40]. Lastly, in vivo administration of myricetin (a flavonol) or green tea catechins triggered an increase of glucose uptake in skeletal muscles and a decrease of glucose uptake in AT, as reviewed in [6]. The fact that a given agent increases glucose utilisation in one tissue but not in another could appear astonishing at the first glance, but corresponds to already observed redistribution of metabolic fluxes. Several molecules have been shown to exhibit such partitioning properties, such as the �3-adrenergic agonists, able to activate glucose transport in rat skeletal muscle [77], while they hamper insulin activation of hexose uptake in white adipocytes [78].
After these statements about muscle, the major compo-nent of peripheral glucose disposal, it appears less amazing to consider that polyphenols can exhibit antidiabetic capaci-ties, owing to their positive effects on glucose utilisation by myocytes whereas they do not favour insulin stimulation of glucose uptake into fat cells. In this view, polyphenols also lower peroxisome proliferator-activated receptor gamma (PPAR�) activity via SIRT-1 activation [63] and further im-pair insulin action in fat cells. Though not facilitating the demonstration of their antidiabetic effect, these integrated effects have another advantage: contrarily to thiazolidinedi-ones or other agents that improve glucose handling under diabetic situations at the expense of an enlargement of fat depots, the polyphenols are not expected to facilitate fat ac-cretion. This counter-action of the anabolic effect of insulin in fat cells might be involved in the limitation of fat store enlargement found after resveratrol administration in animal studies [53]. Since reduction of fat deposition is accompa-nied by a reduction of proinflammatory adipokines, it im-proves the overall peripheral insulin sensitivity. The impair-ment of fat deposition is also in total agreement with the repeatedly reported antiadipogenic effects of resveratrol and other stilbenes or flavonoids (quercetin, garcinol, pterostil-bene, piceatannol) in 3T3-L1 cells [26, 63, 67, 79-82]. In a preliminary study comparing such antiadipogenic effects of
stilbenes, Visitin A, a resveratrol tetramer was the most effi-cient blocker of adipocyte differentiation, acting via preadi-pocyte cell cycle arrest [83].
In vitro adipogenic differentiation is a complex process not only orchestrated by insulin since preadipocytes require to be primed by corticoids, isobutylmethylxanthine, and other growth factors to fulfil adipogenesis and fat storage specialization [1]. As far as we know, most of the demon-strations of the antiadipogenic action of resveratrol or deriva-tives have been obtained with 3T3-L1 preadipocytes, which require such complex priming. Thus, it remains unclear whether resveratrol impairs insulin action only or whether it prevents other factors to trigger lipid deposition. With the objective of studying the interplay resveratrol vs insulin, we used the 3T3 F442A murine preadipocyte lineage, which depends only on insulin to differentiate into lipid-laden adi-pocytes [84]. (Fig. 4) clearly shows that, when cultured for 8 days post-confluence under optimal conditions, save for insulin, these cells accumulate less than one-third of the lip-ids they contain in the presence of 50 nM insulin. Daily res-veratrol addition at 20 �M final concentration in the culture medium deeply limited insulin-induced lipid accumulation, as well as it impaired protein synthesis and clonal expansion (post-confluent mitosis). Anyhow, resveratrol was also highly active in the absence of the pancreatic hormone: it deeply lowered all measured parameters, therefore exhibiting strong antiadipogenic property per se and suggesting a cyto-toxic behaviour (Fig. 4). It can be mentioned at this stage that hydrogen peroxide and reactive oxygen species are somewhat proadipogenic [85], making the antiadipogenic effect of resveratrol closely related to its antioxidant capacity rather than to its selective antiinsulin properties. Hence, many other polyphenols are antiadipogenic, being among them: resveratrol tetramer [83] or caffeic acid [86].
Moreover, resveratrol exerts antilipogenic effect in rat [87] or mouse [62] mature adipocytes and in differentiated human preadipocytes [70] differently from other antioxi-dants. It inhibits the expression of fatty acid synthase (FAS) [70, 88] and activation the fatty acid beta-oxidation pathway [89], thus exerting a coordinate action to lower the intracel-lular storage of lipids.
Regarding lipolysis regulation, resveratrol has been re-ported to activate triglyceride breakdown in rodent [87] and human [59] adipocytes , in a manner that depends on AMPK activation and which results in the stimulation of ATGL (adipose triglyceride lipase) rather than HSL (hormone-sensitive lipase) activity [90]. Accordingly, resveratrol alter-ated the antilipolytic effect of insulin [87], while it enhanced the lipolytic effect of beta-adrenoceptor agonists [87], as did quercetin [91]. Thus, resveratrol induces harmonized re-sponses in fat cell: limiting FAS activity and triacylglycerol assembly, together with glucose transport impairment, and increased lipolysis. These multiple properties therefore largely participate to its antiobesity effects, at least in several animal models [52].
Regarding the expression and secretion of adipokines, it is now widely accepted that resveratrol modulates in a coor-dinated manner (again) many of these factors allowing a crosstalk between AT and the entire organism. Resveratrol inhibits leptin secretion [92] and resistin expression of adi-

158 Current Medicinal Chemistry, 2015, Vol. 22, No. 1 Carpéné et al.
pocytes [89]. Likewise, its partial impairment of both ex-pression and secretion of proinflammatory adipokines such as monocyte chemoattractant protein-1 (MCP-1), interleu-kins (IL1�, IL6, IL8…) and others (PAI-1…) occurs through the SIRT-1 pathway, in explants of human adipose tissue [93], visceral fat cells [94], or in a human preadipocyte line-age [70]. On the opposite, the improvement by resveratrol of adiponectin expression [94] is depending on AMPK activa-tion [95] and surely participates in the mitigation of insulin resistance observed in vivo. With the same consequences, resveratrol and quercetin were confirmed to attenuate the TNF�-induced production of inflammatory adipokines and reduction of adiponectin expression, together with a block-ade of TNF�-induced inhibition of PPAR� [96]. Of note, the capacity to regulate adipokine production, increasing the beneficial adiponectin and reducing the deleterious ones, has been extended to resveratrol metabolites (glucuronides or sulfates), indicating that they may also participate to the benefits of resveratrol ingestion [97].
Fig. (4). Antiadipogenic effect of resveratrol in differentiating 3T3-F442A preadipose cells. Triglyceride accumulation (upper panel) or protein content (middle panel) was expressed as percentage of the optimal condition of adipocyte differentiation, on post confluent-preadipocytes cultured for 8 days in the presence of insulin 50 nM. Resveratrol 20 �M was tested, without (negative control, black column) or with insulin (optimal condition, shaded columns). Lower panel: cell number at day 8. Mean ± SEM of 4-6 determina-tions. Different from corresponding culture condition without res-veratrol at: ** p < 0.01; *** p < 0.001.
To summarize, numerous in vitro and in vivo observa-tions are converging to attest that resveratrol inhibits glucose transport in adipose cells and increases it in skeletal muscle (for review, see: [98]). Other polyphenols have been ascribed
with the same dual action, e.g. green tea catechins (myricetin, quercetin and catechin-gallate), which have been reported to inhibit glucose transport in intestine, too [6, 64]. This latter observation point out that resveratrol and deriva-tives might be useful in preventing and treating diabetes not only on the basis of their complex actions on "insulin-sensitive tissues" but also by modulating many other aspects of food efficiency, including in the gut-brain axis.
SEARCHING FOR SYNERGISM WITHIN POLY-PHENOLS OR WITH OTHER ANTIDIABETIC OR ANTIOBESITY MOLECULES
The potential health impact of resveratrol expected from its multiple in vitro actions is considerably hampered by its low bioavailability. In reality, the polyphenol is subject to an intense metabolism and only low plasma concentrations of resveratrol are detected following oral administration. Once ingested, resveratrol rapidly generates glucuronide and sul-fate conjugates [41, 42]. In gut, resveratrol has been de-scribed also to induce overexpresion of proglugacon mRNAs [99] and to be metabolized by bacteria [43]. Although sev-eral resveratrol metabolites exert a biological activity on adipocytes [97], numerous investigations have been per-formed to find new tools that increase the stilbene bioavail-ability or that improve its effectiveness. The alternative ways of resveratrol administration solving the problems of its lim-ited water solubility [100], its trans-intestinal passage or the concerns about its degradation in food [11] are not treated here. Only combination with other molecules or modification of its chemical structure will be considered.
It has been established that the co-administration of sev-eral dietary polyphenols induces an inhibition of resveratrol metabolism by a competition mechanism towards the me-tabolizing enzymes, and consequently increases its bioavail-ability [101]. For instance, combining resveratrol (100 mg/kg) with piperine (10 mg/kg) enhances its bioavailability because piperine inhibits its glucuronidation, thereby slow-ing its elimination. With that combination, an increase of the maximum serum concentrations was observed in mice [102]. Likewise, quercetin inhibits resveratrol sulfation and glucu-ronidaton and thus might indirectly increase the low bioavailability of the stilbene. However, the therapeutical effect of such combinations, expected to augment the degree of exposure to resveratrol, has not been evaluated on insulin sensitivity or on obesity, if one does not take into account the studies performed with plant extracts containing multiple components. Nevertheless, other combinations have been tested, owing to other rationale. In fact, it is the antiadipo-genic effect of several polyphenols that is the basis for the demonstration of potentiation (or synergism) with resvera-trol.
Noteworthy, an in vitro study showed that quercetin and resveratrol strongly inhibited the differentiation of preadi-pose cells when combined together at a low concentration (25 �M), which by itself was marginally active to impair lipid accumulation [103]. Such findings were in line with previous demonstrations of the antiadipogenic effects of quercetin [67] or resveratrol [79], and with the fact that each of them activate AMPK when used separately [67, 73]. The novel insight was that these two polyphenols acted in syner-
Adipogenesis of 3T3 F442A cells
020406080
100120
( % o
f ins
ulin
)
020406080
100120
( % o
f ins
ulin
)
0
200
400
600
800
- +resveratrol 20 µM:
( x
1000
)
triglycerides/well
proteins/well
cell number/well
negative control + insulin 50 nM
- +
***
***
***
***
***
*

Using Natural Polyphenols in Diabetes and Obesity Current Medicinal Chemistry, 2015, Vol. 22, No. 1 159
gism when combined, leading to propose that such combina-tion is instrumental for treating obesity. Moreover, the com-bination of resveratrol + quercetin induced apoptosis in ma-ture adipocytes [103]. Regrettably, all these in vitro effects of quercetin plus resveratrol were hardly transposable to invivo beneficial effects.
Genistein, another flavonoid, shares similar inhibitory properties in adipocytes, reducing the lipogenic and the an-tilipolytic effects of insulin. The combination of resveratrol with genistein enhanced in vitro effects such as inhibiting adipogenesis, inducing lipolysis and triggering apoptosis in 3T3-L1adipocytes (at 25 to 100 �M) [104]. Even better, the "triple" combination of 50-100�M of resveratrol plus quer-cetin and genistein was strongly antiadipogenic in murine and human preadipocytes, resulting in an authentic syner-gism, i.e. an effect of the combination of agents that was more than a simple addition of their respective effects [25]. Moreover, when tested in old rats together with Vitamin D, these phytochemicals limited excessive weight gain [52]. A research group from a company aiming at developing sirtuin activators reported synergistic effects of leucine and resvera-trol in cultured preadipocytes, which were explained by the fact that both agents increased mitochondrial biogenesis and fat oxidation via SIRT-1 dependent pathways [105]. More importantly, a 6-week supplementation with leucine and res-veratrol improved insulin sensitivity and limited adiposity in diet-induced obese mice [105]. The same group extended its observation of such potentiation leucine-resveratrol on fatty acid oxidation to other polyphenols, especially the phenolic acids. Additionally, synergisms were found during in vitroand in vivo approaches with other agents, such as a leucine metabolite, methylxanthines and metformin [106]. To our knowledge, no data on human studies are available about this promising combined therapeutic approach
However, another combination has been studied recently in 18 overweight volunteers, consisting in supplementation with epigallocatechin-gallate together with resveratrol (282 mg/day + 200 mg/day) for 3 days [107]. Such treatment in-creased fasting and postprandial energy expenditure, proba-bly by improving the altered lipid oxidation that it is associ-ated with obesity and type 2 diabetes. However, the effect of each molecule alone was not determined and it is therefore unknown whether such observation was related to a syner-gistic effect or was due only to one of the molecules. Never-theless, the addition of a mixture of isoflavones to epigallo-catechin-gallate and resveratrol further enhanced the circu-lating indexes of lipolysis, suggesting that long-term sup-plementation with a mixture of polyphenols plus other agents may improve metabolic flexibility in men.
Conjugated linoleic acids have also been considered to improve metabolic health, and to facilitate lipid handling. Consequently, they were logically tested together with res-veratrol in obese and diabetic animal models. The admini-stration of a combination of trans-10, cis-12 conjugated lino-leic acid (CLA) with resveratrol at 30 mg/kg/d during 6 weeks in obese rats unexpectedly demonstrated that their combination reduced the effectiveness of each compound in lowering body fat content, while it maintained the positive effect of resveratrol on glycaemic control [108]. Hence, this combination attenuated the individual effects of resveratrol
and CLA on triacylglycerol metabolism in adipose tissue [109]. In vitro studies demonstrated that the combination of these agents did not increase their own delipidating effect in 3T3 adipocytes [110]. Furthermore, an antagonism was found between these two classes of compounds regarding inflammation and insulin resistance in human adipocytes, the fatty acid being rather proinflammatory whereas the stilbene was antiinflammatory [111]. Although negative, this attempt to obtain a synergistic antiobesity or antidiabetic combina-tion indicated at least that the in vitro properties tested on adipocyte models (e.g. adipogenesis inhibition) [26] might be relevant for further in vivo studies of the antiobesity effect of polyphenols and related molecules.
Phenelzine is a well-recognized monoamine oxidase (MAO) inhibitor, which has been evidenced to inhibit adipo-genesis in diverse preadipocyte models in a MAO-independent manner [84]. Since we found that this drug also inhibited lipogenesis in mature fat cells [61] - though by un-known mechanisms - we recently tested the effect of resvera-trol and phenelzine combination in mice fed a high-fat diet. However, the low doses chosen, did not exhibit any syner-gism relative to that achieved by each molecule separately in terms of body weight gain, fat content or glucose tolerance [62]. Nevertheless, post-hoc verification in mouse adipocytes showed that the antilipogenic effects of resveratrol and of phenelzine were not additive. Again, this seems to indicate that antiadipogenic and antilipogenic effects could be useful for primary screening before developing polyphenol-based experimental therapies. At last, the subtle difference between multiple combinatory treatment with well-defined molecules and the oral supplementation with poorly chemically defined dietary supplements or with often adulterated medicinal plant extracts must be maintained in order to define whether there is or not any synergism among the chosen active principles [112].
Analogues of resveratrol may overcome the limitations of its pharmacologic efficacy by different ways: or they are more potent in activating or repressing the cellular key tar-gets able to correct the metabolic dysregulations of diabesity, or they have substantially greater intestinal absorption and enhanced hepatic stability. When considering the limits of resveratrol bioavailability and pharmacodynamic activity, one understands easily why congeners or analogues of res-veratrol have attracted so much attention recently.
One of these analogues is pterostilbene. The bioavailabil-ity of this methylated polyphenol is greater than that of res-veratrol because the two methoxy groups of pterostilbene make the molecule more lipophilic, and increase its oral ab-sorption and cellular uptake. After ingestion, pterostilbene presents a bioavailability of 95 %, whereas resveratrol is limited at 20 % [113, 114]. Thus, pterostilbene reaches higher plasma concentrations. In spite of that, there are very less studies describing its effects in metabolic diseases than with resveratrol. Regarding diabetes and insulin resistance, it has been reported that a plant very rich in pterostilbene (Pterocarpus marsupium) almost normalized the glycaemia and insulinemia altered by sucrose feeding in rodents [115]. Furthermore, other studies have also shown a decrease of hbAIc, and various changes in enzyme expressions related to the antioxidant potential of pterostilbene that confered a pro-

160 Current Medicinal Chemistry, 2015, Vol. 22, No. 1 Carpéné et al.
tective effect against diabetic complications such as liver and kidney damage [116]. Of note, pterostilbene exhibits antiadi-pogenic and antiproliferative effects on preadipocytes [117].
Another natural polyphenol, structurally close to resvera-trol, is piceatannol. This hydroxylated analogue and metabo-lite of resveratrol is able to inhibit adipogenesis, too [82]. As mentioned above for resveratrol, piceatannol enhances glu-cose uptake in muscle cells and, in vivo, it delays the onset of hyperglycaemia in genetically obese and diabetic mice [75]. Interestingly, its antitumor, antioxidant, and antiinflamma-tory properties have been reviewed recently, as well as its ability to bind the quinone reductase already reported to in-teract with resveratrol [118], but its impact on metabolic dysregulations deserves to be further studied [119].
Many other derivatives have been the subject of scarce studies (oxyresveratrol, isorhapontigenin, parthenocissin A…) or of early patenting and the dissemination to the scien-tific community of their beneficial effect, and medical appli-cation, if any, belongs to ongoing research. Therefore, res-veratrol, either under the form of combinations with other phytochemicals, or taken as skeleton for substituted ana-logues, will continue to provide potential novel potent thera-pies for both obesity and diabetes. As precised in the intro-duction, physical activity, which is a good prevention and treatment for diabesity, deserves also to be considered with resveratrol supplementation [7], alone or combined with other polyphenols.
MUTIDISCIPLINAR PERSPECTIVES TO IMPROVE THE BENEFITS/RISKS RATIO OF POLYPHENOLS FOR DIABESITY TREATMENT OR PREVENTION
Obesity and diabetes are major public health problems in developed societies, affecting more than 15% of the adult population and increasing with age. Indeed, both obesity and diabetes are important primarily as risk factors for serious complications such as cardiovascular (coronary ischemia, hypertension, heart failure, etc), liver (steatosis, cirrhosis), kidney and many other diseases, including cancer. The grow-ing list of polyphenol actions in experimental models of each of these complications indicates that polyphenols are valu-able candidates for improving cardiometabolic health of obese and/or diabetic patients. Their antiinflammatory ac-tions resemble to that of a class previously used antidiabetic agents, the glitazones or PPAR� agonists. However, these latter agents contributed to a vicious circle of weight gain and fat accretion, by facilitating the lipid storage or excess energy intake into the adipose mass, a mechanism that is not activated by polyphenols. Similarly, polyphenols seem to activate the conversion of white- to brown-adipocytes [48] as did other antidiabetic and antiobesity candidates, namely the �3-adrenergic agonists, which have never concretized their promising therapeutic advantage. Differently from such ago-nists, resveratrol and related agents do not act on a single receptor in target cells, but affect various key enzymes and transcription factors, like those of the sirtuin family. Albeit non-shivering thermogenesis does not occupy the same im-portance in the energy balance in man than in rodents, fur-ther studies are needed to assess whether polyphenols in-crease the proportion and functionality of beige or brite adi-pocytes. Fortunately, polyphenols tend to impair adipocyte
enlargement and seems to facilitate fat cell shrinking, with-out promoting ectopic fat deposition in the liver, as it was the case for conjugated forms of linoleic acid, suggested for a while as antiobesity dietary supplements. The polyphenols are therefore believed to increase energy expenditure or re-duce metabolic efficiency by still not totally defined mecha-nisms but which likely include enhanced lipid turnover.
As discussed above, a direct insulin-like or insulin-sensitizer action of polyphenols in AT is probably not in-volved in their clinical antidiabesity effect. However, by modifying the life cycle of adipocytes, polyphenols should also modify secretion of adipokines and might limit the metabolic disorders associated to excessive fat depots. Albeit polyphenols are surely exerting numerous effects in the body elsewhere than in AT, we propose adipose cells as a conven-ient tool for further screening of derivatives or polyphenol combinations endowed with improved therapeutic properties, compared with resveratrol. These cells are easily available and they exhibit various pathways already documented to be sensitive to polyphenol actions: hexose transport, lipogene-sis, triglyceride uptake, lipolysis, fatty acid oxidation and thermogenesis. However, due to their cardioprotective ef-fects, to their activation of glucose transport and mitochon-drial activity in myocytes, the polyphenols likely impact both cardiac and skeletal muscle in a beneficial manner that de-serves further studies, too. Lastly, their central actions are far from being totally elucidated, as it is the case for influence on gut bacteria and pancreatic cells. Therefore multidiscipli-nary approaches are necessary for studying all the facets of such compounds already proposed to exert multiple "calorie restriction-like" or "exercise-like" actions.
Finally, obesity and diabetes are two disorders of high prevalence and impact on public health that have in common inflammation and oxidative stress. However, when taken separately, neither the antioxidants nor the inflammatory agents have been proven therapeutically relevant to treat these diseases. "Novel" polyphenols, including oligomers, having validated antioxidant capabilities are supposed to be beneficial [120]. Since preliminary studies have indicated that such compounds may potentiate their actions when ad-ministered simultaneously, achieving greater effects than when given separately. It appears logical to further study the dual action of antioxidant and antiinflammatory approaches. This can be performed by searching the synergistic effect of the more valuable combinations of polyphenols and other compounds, via a first "screening" in fat cells. Then, any further validation in animal models should test whether a potentiation of such antioxidant and antiinflammatory effects is one of the active components leading to such expected antidiabesity properties. Since it has been described that the antiinflammatory and antioxidant properties of resveratrol help in improving cardiovascular and renal complications, one could expect that other dysregulated organs may be im-proved by such synergism. Thus, although combating obe-sity and diabetes requires to perform investigations on ani-mal models with dysregulated energy balance, studies in adipocytes, skeletal muscle cells, cardiomyocytes and hepa-tocytes, and probably in kidney and intestine as well, are highly recommended to retrieve the natural chemical struc-tures having the best ratio benefit/risk for future medical or nutritional use.

Using Natural Polyphenols in Diabetes and Obesity Current Medicinal Chemistry, 2015, Vol. 22, No. 1 161
CONFLICT OF INTEREST
The author(s) confirm that this article content has no con-flicts of interest.
ACKNOWLEDGEMENTS
This work was partly supported by DIOMED project (INTERREG IVB SUDOE 1/P1/E178). Authors acknowl-edge the networking of all members of the frensh-spanish Refbio/POLYFrEsNOL project and of the Consortium Trans Pyrenean of Investigations in Obesity and Diabetes (http:// obesitydiabetesinctp.weebly.com).
REFERENCES [1] Bastard, J. P.; Fève, B. Physiology and Physiopathology of adipose
tissue. 1st ed. Springer-Verlag: Paris, 2013.[2] Duncan, B. B.; Schmidt, M. I.; Pankow, J. S.; Ballantyne, C. M.;
Couper, D.; Vigo, A.; Hoogeveen, R.; Folsom, A. R.; Heiss, G. Atherosclerosis Risk in Communities Study. Low-grade systemic inflammation and the development of type 2 diabetes: the athero-sclerosis risk in communities study. Diabetes, 2003, 52, 1799-1805.
[3] Minokoshi, Y.; Kahn, C.R.; Kahn, B.B. Tissue-specific ablation of the GLUT4 glucose transporter or the insulin receptor challenges assumptions about insulin action and glucose homeostasis. J. Biol. Chem., 2003, 278, 33609-33612.
[4] DeFronzo, R. A. From the triumvirate to the ominous octet: a new paradigm for the treatment of type 2 diabetes mellitus. Diabetes,2009, 58, 773-795.
[5] Borriello, A.; Cucciolla, V.; Della Ragione, F.; Galletti, P. Dietary polyphenols: focus on resveratrol, a promising agent in the preven-tion of cardiovascular diseases and control of glucose homeostasis. Nutr. Metab. Cardiovasc. Dis., 2010, 20, 618-625.
[6] Park, T.; Kim, Y. Phytochemicals as potential agents for prevention and treatment of obesity and metabolic diseases. Anti-Obes. Drug Discov. Develop., 2011, 1, 150-185.
[7] Schrauwen, P.; Timmers, S. Can resveratrol help to maintain meta-bolic health? Proc. Nutr. Soc., 2014, 73, 271-277.
[8] Xiao, J.; Hogger, P. Influence of diabetes on the pharmacokinetic behavior of natural polyphenols. Curr. Drug Metab., 2014, 15, 23-29.
[9] Su, H. C.; Hung, L. M.; Chen, J. K. Resveratrol, a red wine anti-oxidant, possesses an insulin-like effect in streptozotocin-induced diabetic rats. Am. J. Physiol. Endocrinol. Metab., 2006, 290,E1339-E1346.
[10] Baur, J. A.; Sinclair, D. A. Therapeutic potential of resveratrol: the in vivo evidence. Nat. Rev. Drug Discovery, 2006, 5, 493-506.
[11] Maraculla, M. T.; Alberdi, G.; Gomez, S.; Tueros, I.; Bald, C.; Rodriguez, V. M.; Martinez, J. A.; Portillo, M. P. Effects of differ-ent doses of resveratrol on body fat and serum parameters in rats fed a hypercaloric diet. J. Physiol. Biochem., 2009, 65, 369-376.
[12] Kim, S.; Jin, Y.; Choi, Y.; Park, T. Resveratrol exerts anti-obesity effects via mechanisms involving down-regulation of adipogenic and inflammatory processes in mice. Biochem. Pharmacol., 2011,81, 1343-1351.
[13] Szkudelska, K.; Szkudelski, T. Resveratrol, obesity and diabetes. Eur. J. Pharmacol., 2010, 635, 1-8.
[14] Zamora-Ros, R.; Andres-Lacueva, C.; Lamuela-Raventós, R. M.; Berenguer, T.; Jakszyn, P.; Martínez, C.; Sánchez, M. J.; Navarro, C.; Chirlaque, M. D.; Tormo, M. J.; Quirós, J. R.; Amiano, P.; Dor-ronsoro, M.; Larrañaga, N.; Barricarte, A.; Ardanazn, E.; González, C. A. Concentrations of resveratrol and derivatives in foods and es-timation of dietary intake in a Spanish population: European Pro-spective Investigation into Cancer and Nutrition (EPIC)-Spain co-hort. Br. J. Nutr., 2008,100, 188-196.
[15] Xiao, J.; Ni, X.; Kai, G.; Chen, X. A review on structure-activity relationship of dietary polyphenols inhibiting �-amylase. Crit. Rev. Food Sci. Nutr., 2013, 53, 497-506.
[16] Xiao, J.; Kai, G.; Yamamoto, K.; Chen, X. Advance in dietary polyphenols as �-glucosidases inhibitors: a review on structure-activity relationship aspect. Crit. Rev. Food Sci. Nutr., 2013, 53,818-836.
[17] Meydani, M.; Hasan, S. T. Dietary polyphenols and obesity. Nutri-ents, 2010 2, 737-751.
[18] Rothwell, J. A.; Urpi-Sarda, M.; Boto-Ordoñez, M.; Knox, C.; Llorach, R.; Eisner, R.; Cruz, J.; Neveu, V.; Wishart, D.; Manach, C.; Andres-Lacueva, C.; Scalbert, A. Phenol-Explorer 2.0: a major update of the Phenol-Explorer database integrating data on poly-phenol metabolism and pharmacokinetics in humans and experi-mental animals. Database (Oxford), 2012, 2012, bas031.
[19] Brasnyó, P.; Molnár, G. A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I. A.; Mérei, A.; Halmai, R.; Mészáros, L. G.; Sümegi, B.; Wittmann, I. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br. J. Nutr., 2011, 106, 383-389.
[20] Crandall, J. P.; Oram, V.; Trandafirescu, G.; Reid, M.; Kishore, P.; Hawkins, M.; Cohen, H. W.; Barzilai, N. Pilot study of resveratrol in older adults with impaired glucose tolerance. J. Gerontol. A Biol. Sci. Med. Sci., 2012, 67, 1307-1312.
[21] Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R. H.; van de Weijer, T.; Goossens, G.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; Moonen-Kornips, E.; Hesselink, M.; Kunz, I.; Schrau-wen-Hinderling, V.; Blaak, E.; Auwerx, J.; Schrauwen, P. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab., 2011, 14, 612-622.
[22] Baur, J. A.; Pearson, K. J.; Price, N. L.; Jamieson, H. A.; Lerin, C.; Kalra, A.; Prabhu, V. V.; Allard, J. S.; Lopez-Lluch, G.; Lewis, K.; Pistell, P. J.; Poosala, S.; Becker, K. G.; Boss, O.; Gwinn, D.; Wang, M.; Ramaswamy, S.; Fishbein, K. W.; Spencer, R. G.; Lakatta, E. G.; Le Couteur, D.; Shaw, R. J.; Navas, P.; Puigserver, P.; Ingram, D. K.; de Cabo, R.; Sinclair, D. A. Resveratrol im-proves health and survival of mice on a high-calorie diet. Nature,2006, 444, 337-342.
[23] Barger, J. L.; Kayo, T.; Vann, J. M.; Arias, E. B.; Wang, J.; Hacker, T. A.; Wang, Y.; Raederstorff, D.; Morrow, J. D.; Leeu-wenburgh, C.; Allison, D. B.; Saupe, K. W.; Cartee, G. D.; Wein-druch, R.; Prolla, T. A. A low dose of dietary resveratrol partially mimics caloric restriction and retards aging parameters in mice. PLoS One, 2008, 3, e2264.
[24] Konings, E.; Timmers, S.; Boekschoten, M. V.; Goossens, G. H.; Jocken, J. W.; Afman, L. A.; Müller, M.; Schrauwen, P.; Mariman, E. C.; Blaak, E. E. The effects of 30 days resveratrol supplementa-tion on adipose tissue morphology and gene expression patterns in obese men. Int. J. Obes. (Lond), 2014, 38, 470-473.
[25] Park, H. J.; Yang, J. Y.; Ambati, S.; Della-Fera, M. A.; Hausman, D. B.; Rayalam, S.; Baile, C. A. Combined effects of genistein, quercetin, and resveratrol in human and 3T3-L1 adipocytes. J. Med. Food, 2008, 11, 773-783.
[26] Lasa, A.; Churruca, I.; Eseberri, I.; Andrés-Lacueva, C.; Portillo, M. P. Delipidating effect of resveratrol metabolites in 3T3-L adi-pocytes. Mol. Nutr. Food Res., 2012, 56, 1559-1568.
[27] Movahed, A.; Nabipour, I.; Lieben Louis, X.; Thandapilly, S. J.; Yu, L.; Kalantarhormozi, M.; Rekabpour, S. J.; Netticadan, T. An-tihyperglycemic effects of short term resveratrol supplementation in type 2 diabetic patients. Evid. Based Compl. Alternat. Med.,2013, 2013, 851267.
[28] Bhatt, J. K.; Thomas, S.; Nanjan, M. J. Resveratrol supplementa-tion improves glycemic control in type 2 diabetes mellitus. Nutr. Res., 2012, 32, 537-541.
[29] Knop, F. K.; Konings, E.; Timmers, S.; Schrauwen, P.; Holst, J. J.; Blaak, E. E. Thirty days of resveratrol supplementation does not af-fect postprandial incretin hormone responses, but suppresses post-prandial glucagon in obese subjects. Diabet. Med., 2013, 30, 1214-1218.
[30] Dash, S.; Xiao, C.; Morgantini, C.; Szeto, L.; Lewis, G. F. High-dose resveratrol treatment for 2 weeks inhibits intestinal and he-patic lipoprotein production in overweight/obese men. Arterioscler. Thromb. Vasc. Biol., 2013, 33, 2895-901.
[31] Poulsen, M. M.; Vestergaard, P. F.; Clasen, B. F.; Radko, Y.; Christensen, L. P.; Stødkilde-Jørgensen, H.; Møller, N.; Jessen, N.; Pedersen, S. B.; Jørgensen, J. O. High-dose resveratrol supplemen-tation in obese men: an investigator-initiated, randomized, placebo-controlled clinical trial of substrate metabolism, insulin sensitivity, and body composition. Diabetes, 2013, 62, 1186-1195.
[32] Yoshino, J.; Conte, C.; Fontana, L.; Mittendorfer, B.; Imai, S. I.; Schechtman, K. B.; Gu, C.; Kunz, I.; Fanelli, F. R.; Patterson, B.

162 Current Medicinal Chemistry, 2015, Vol. 22, No. 1 Carpéné et al.
W.; Klein, S. Resveratrol supplementation does not improve meta-bolic function in nonobese women with normal glucose tolerance. Cell Metab., 2012, 16, 658-664.
[33] Ghanim, H.; Sia, C. L.; Abuaysheh, S.; Korzeniewsk, i. K.; Pat-naik, P.; Marumganti, A.; Chaudhuri, A.; Dandona, P. An antiin-flammatory and reactive oxygen species suppressive effects of an extract of Polygonum cuspidatum containing resveratrol. J. Clin. Endocrinol. Metab., 2010, 95, E1-E8.
[34] Liu, K.; Zhou, R.; Wang, B.; Mi, M. T. Effect of resveratrol on glucose control and insulin sensitivity: a meta-analysis of 11 ran-domized controlled trials. Am. J. Clin. Nutr., 2014, 99, 1510-1519.
[35] Crandall, J. P.; Barzilai, N. Exploring the promise of resveratrol: where do we go from here? Diabetes, 2013, 62, 1022-1023.
[36] Kang, W.; Hong, H. J.; Guan, J.; Kim, D. G.; Yang, E. J.; Koh, G.; Park, D.; Han, C. H.; Lee, Y. J.; Lee, D. H. Resveratrol improves insulin signaling in a tissue-specific manner under insulin-resistant conditions only: in vitro and in vivo experiments in rodents. Me-tabolism, 2012, 61, 424-433.
[37] Fauconneau, B.; Waffo-Teguo, P.; Huguet, F.; Barrier, L.; Decen-dit, A.; Merillon, J. M. Comparative study of radical scavenger and antioxidant properties of phenolic compounds from Vitis viniferacell cultures using in vitro tests. Life Sci., 1997, 61, 2103-2110.
[38] Ristow, M.; Zarse, K.; Oberbach, A.; Klöting, N.; Birringer, M.; Kiehntopf, M.; Stumvoll, M.; Kahn, C. R.; Blüher, M. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc. Natl. Acad. Sci. USA, 2009, 106, 8665-8670.
[39] Skrobuk, P.; von Kraemer, S.; Semenova, M. M.; Zitting, A.; Koistinen, H. A. Acute exposure to resveratrol inhibits AMPK ac-tivity in human skeletal muscle cells. Diabetologia, 2012, 55,3051-3060.
[40] Um, J. H.; Park, S. J.; Kang, H.; Yang, S.; Foretz, M.; McBurney, M. W.; Kim, M. K.; Viollet, B.; Chung, J. H. AMP-activated pro-tein kinase-deficient mice are resistant to the metabolic effects of resveratrol. Diabetes, 2010, 59, 554-563.
[41] Cottart, C.-H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.-L. Resveratrol bioavailability and toxicity in humans. Mol. Nutr. Food Res., 2010, 54, 7-16.
[42] Brown, V. A.; Patel, K. R.; Viskaduraki, M.; Crowell, J. A.; Per-loff, M.; Booth, T. D.; Vasilinin, G.; Sen, A.; Schinas, A. M.; Pic-cirilli, G.; Brown, K.; Steward, W. P.; Gescher, A.; Brenner, D. E. Repeat dose study of the cancer chemopreventive agent resveratrol in healthy volunteers: safety, pharmacokinetics, and effect on the insulin-like growth factor axis. Cancer Res., 2010, 70, 9003-9011.
[43] Bode, L. M.; Bunzel, D.; Huch, M.; Cho, G. S.; Ruhland, D.; Bun-zel, M.; Bub, A.; Franz, C. M.; Kulling, S. E. In vivo and in vitrometabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr., 2013, 97, 295-309.
[44] Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; Geny, B.; Laakso, M.; Puigserver, P.; Auwerx, J. Resveratrol im-proves mitochondrial function and protects against metabolic dis-ease by activating SIRT1 and PGC-1alpha. Cell, 2006, 127, 1109-1122.
[45] Do, G. M.; Jung, U. J.; Park, H. J.; Kwon, E. Y.; Jeon, S. M.; McGregor, R. A.; Choi, M. S. Resveratrol ameliorates diabetes-related metabolic changes via activation of AMP-activated protein kinase and its downstream targets in db/db mice. Mol. Nutr. Food Res., 2012, 56, 1282-1291.
[46] Price, N. L.; Gomes, A. P.; Ling, A. J.; Duarte, F. V.; Martin-Montalvo, A.; North, B. J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J. S.; Hubbard, B. P.; Varela, A. T.; Davis, J. G.; Varamini, B.; Hafner, A.; Moaddel, R.; Rolo, A. P.; Coppari, R.; Palmeira, C. M.; de Cabo, R.; Baur, J. A.; Sinclair, D. A. SIRT1 is required for AMPK activation and the beneficial effects of resvera-trol on mitochondrial function. Cell Metab., 2012, 15, 675-690.
[47] Louis, X. L.; Thandapilly, S. J.; MohanKumar, S. K.; Yu, L.; Tay-lor, C. G.; Zahradka, P.; Netticadan, T. Treatment with low-dose resveratrol reverses cardiac impairment in obese prone but not in obese resistant rats. J. Nutr. Biochem., 2012, 23, 1163-1169.
[48] Andrade, J. M.; Frade, A. C.; Guimarães, J. B.; Freitas, K. M.; Lopes, M. T.; Guimarães, A. L.; de Paula, A. M.; Coimbra, C. C.; Santos, S. H. Resveratrol increases brown adipose tissue ther-mogenesis markers by increasing SIRT1 and energy expenditure and decreasing fat accumulation in adipose tissue of mice fed a standard diet. Eur. J. Nutr., 2014. [Epub ahead of print].
[49] Rivera, L.; Moron, R.; Zarzuelo, A.; Galisteo, M. Long-term res-veratrol administration reduces metabolic disturbances and lowers blood pressure in obese Zucker rats. Biochem. Pharmacol. 2009,77, 1053-1063.
[50] Gómez-Zorita, S.; Fernández-Quintela, A.; Macarulla, M. T.; Aguirre, L.; Hijona, E.; Bujanda, L.; Milagro, F.; Martínez, J. A.; Portillo, M. P. Resveratrol attenuates steatosis in obese Zucker rats by decreasing fatty acid availability and reducing oxidative stress. Br. J. Nutr., 2012, 107, 202-210.
[51] Cho, S. J.; Jung, U. J.; Choi, M. S. Differential effects of low-dose resveratrol on adiposity and hepatic steatosis in diet-induced obese mice. Br. J. Nutr., 2012, 108, 2166-2175.
[52] Baile, C. A.; Yang, J. Y.; Rayalam, S.; Hartzell, D. L.; Lai, C. Y.; Andersen, C.; Della-Fera, M. A. Effect of resveratrol on fat mobili-zation. Ann. N. Y. Acad. Sci., 2011, 1215, 40-47.
[53] Alberdi, G.; Rodríguez, V. M.; Miranda, J.; Macarulla, M. T.; Arias, N.; Andrés-Lacueva, C.; Portillo, M. P. Changes in white adipose tissue metabolism induced by resveratrol in rats. Nutr. Me-tab. (Lond), 2011, 8, 29.
[54] Alberdi, G.; Macarulla, M. T.; Portillo, M. P.; Rodríguez, V. M. Resveratrol does not increase body fat loss induced by energy re-striction. J. Physiol. Biochem., 2014, 70, 639-646.
[55] Barger, J. L. An adipocentric perspective of resveratrol as a calorie restriction mimetic. Ann. N. Y. Acad. Sci., 2013, 1290, 122-129.
[56] Narkar, V. A.; Downes, M.; Yu, R. T.; Embler, E.; Wang, Y. X.; Banayo, E.; Mihaylova, M. M.; Nelson, M. C.; Zou, Y.; Juguilon, H.; Kang, H.; Shaw, R. J.; Evans, R. M. AMPK and PPARdelta agonists are exercise mimetics. Cell, 2008, 134, 405-415.
[57] Delmas, D.; Xiao, J. Natural polyphenols properties: chemopreven-tive and chemosensitizing activities. Anticancer Agents Med. Chem., 2012, 12, 835.
[58] Mercader, J.; Iffiú-Soltesz, Z.; Brenachot, X.; Földi, A.; Dunkel, P.; Balogh, B.; Attané, C.; Valet, P.; Mátyus, P.; Carpéné, C. SSAO substrates exhibiting insulin-like effects in adipocytes as a promis-ing treatment option for metabolic disorders. Future Med. Chem.,2010, 2, 1735-1749.
[59] Gomez-Zorita, S.; Tréguer, K.; Mercader, J.; Carpéné, C. Resvera-trol directly affects in vitro lipolysis and glucose transport in hu-man fat cells. J. Physiol. Biochem., 2013, 69, 585-593.
[60] Carpéné, C.; Bour, S.; Visentin, V.; Pellati, F.; Benvenuti, S.; Igle-sias-Osma, M. C.; Garcia-Barrado, M. J.; Valet, P. Amine oxidase substrates for impaired glucose tolerance correction. J. Physiol. Biochem., 2005, 61, 405-419.
[61] Carpéné, C.; Grès, S.; Rascalou, S. The amine oxidase inhibitor phenelzine limits lipogenesis in adipocytes without inhibiting insu-lin action on glucose uptake. J. Neural. Transm., 2013, 120, 997-1004.
[62] Carpéné, C.; Gomez-Zorita, S.; Gupta, R.; Grès, S.; Rancoule, C.; Cadoudal, T.; Mercader, J.; Gomez, A.; Bertrand, C.; Iffiu-Soltész, Z. Combination of low dose of the anti-adipogenic agents resvera-trol and phenelzine in drinking water is not sufficient to prevent obesity in very-high-fat diet-fed mice. Eur. J. Nutr., 2014. [Epub ahead of Print].
[63] Floyd, Z. E.; Wang, Z. Q.; Kilroy, G.; Cefalu, W. T. Modulation of peroxisome proliferator-activated receptor gamma stability and-transcriptional activity in adipocytes by resveratrol. Metabolism,2008, 57, S32-S38.
[64] Strobel, P.; Allard, C.; Perez-Acle, T.; Calderon, R.; Aldunate, R.; Leighton, F. Myricetin, quercetin and catechin-gallate inhibit glu-cose uptake in isolated rat adipocytes. Biochem. J., 2005, 386, 471-478.
[65] Vlachodimitropoulou, E.; Sharp, P. A.; Naftalin, R. J. Quercetin-iron chelates are transported via glucose transporters. Free Radic. Biol. Med., 2011, 50, 934-944.
[66] Shisheva, A.; Shechter, Y. Quercetin selectively inhibits insulin receptor function in vitro and the bioresponses of insulin and insu-linomimetic agents in rat adipocytes. Biochemistry, 1992, 31, 8059-8063.
[67] Ahn, J.; Lee, H.; Kim, S.; Park, J.; Ha, T. The anti-obesity effect of quercetin is mediated by the AMPK and MAPK signaling path-ways. Biochem. Biophys. Res. Commun., 2008, 373, 545-549.
[68] Arias, N.; Macarulla, M. T.; Aguirre, L.; Martínez-Castaño, M. G.; Portillo, M. P. Quercetin can reduce insulin resistance without de-creasing adipose tissue and skeletal muscle fat accumulation. Genes Nutr., 2014, 9, 361.

Using Natural Polyphenols in Diabetes and Obesity Current Medicinal Chemistry, 2015, Vol. 22, No. 1 163
[69] Stewart, L. K.; Wang, Z.; Ribnicky, D.; Soileau, J. L.; Cefalu, W. T.; Gettys, T. W. Failure of dietary quercetin to alter the temporal progression of insulin resistance among tissues of C57BL/6J mice during the development of diet-induced obesity. Diabetologia,2009, 52, 514-523.
[70] Fischer-Posovszky, P.; Kukulus, V.; Tews, D.; Unterkircher, T.; Debatin, K. M.; Fulda, S.; Wabitsch, M. Resveratrol regulates hu-man adipocyte number and function in a Sirt1-dependent manner. Am. J. Clin. Nutr., 2010, 92, 5-15.
[71] Fang, X. K.; Gao, J.; Zhu, D. N. Kaempferol and quercetin isolated from Euonymus alatus improve glucose uptake of 3T3-L1 cells without adipogenesis activity. Life Sci., 2008, 62, 615-622.
[72] Szkudelska, K.; Nogowski, L.; Szkudelski, T. Resveratrol and genistein as adenosine triphosphate-depleting agents in fat cells. Metabolism, 2011, 60, 720-729.
[73] Breen, D. M.; Sanli, T.; Giacca, A.; Tsiani, E. Stimulation of mus-cle cell glucose uptake by resveratrol through sirtuins and AMPK. Biochem. Biophys. Res. Commun., 2008, 374, 117-122.
[74] Lee, E. S.; Uhm, K. O.; Lee, Y. M.; Han, M.; Lee, M.; Park, J. M.; Suh, P. G.; Park, S. H.; Kim, H. S. CAPE (caffeic acid phenethyl ester) stimulates glucose uptake through AMPK (AMP-activated protein kinase) activation in skeletal muscle cells. Biochem. Bio-phys. Res. Commun., 2007, 361, 854-858.
[75] Minakawa, M.; Miura, Y.; Yagasaki, K. Piceatannol, a resveratrol derivative, promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and suppresses blood glucose levels in type 2 diabetic model db/db mice. Biochem. Biophys. Res. Commun., 2012, 422, 469-475.
[76] Chen, S.; Li, J.; Zhang, Z.; Li, W.; Sun, Y.; Zhang, Q.; Feng, X.; Zhu, W. Effects of resveratrol on the amelioration of insulin resis-tance in KKAy mice. Can. J. Physiol. Pharmacol., 2012, 90, 237-242.
[77] Liu, Y. L.; Cawthorne, M. A.; Stock, M. J. Biphasic effects of the �-adrenoceptor agonist, BRL 37344, on glucose utilisation in rat isolated skeletal muscle. Br. J. Pharmacol., 1996, 117, 1355-1361.
[78] Carpéné, C.; Chalaux, E.; Lizarbe, M.; Estrada, A.; Mora, C.; Palacin, M.; Zorzano, A.; Lafontan, M.; Testar, X. �3-adrenergic receptors are responsible for the adrenergic inhibition of insulin-stimulated glucose transport in rat adipocytes. Biochem. J., 1993,296, 99-105.
[79] Rayalam, S.; Yang, J.-Y.; Ambati, S.; Della-Fera, M. A.; Baile, C. A. Resveratrol induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Phytother. Res., 2008, 22, 1367-1371.
[80] Chen, S.; Li, Z.; Li, W.; Shan, Z.; Zhu, W. Resveratrol inhibits cell differentiation in 3T3-L1 adipocytes via activation of AMPK. Can. J. Physiol. Pharmacol., 2011, 89, 793-799.
[81] Kang, N. E.; Ha, A. W.; Kim, J. Y.; Kim, W. K. Resveratrol inhib-its the protein expression of transcription factors related adipocyte differentiation and the activity of matrix metalloproteinase in mouse fibroblast 3T3-L1 preadipocytes. Nutr. Res. Pract., 2012, 6,499-504.
[82] Kwon, J. Y.; Seo, S. G.; Heo, Y. S.; Yue, S.; Cheng, J. X.; Lee, K. W.; Kim, K. H. Piceatannol, natural polyphenolic stilbene, inhibits adipogenesis via modulation of mitotic clonal expansion and insu-lin receptor-dependent insulin signaling in early phase of differen-tiation. J. Biol. Chem., 2012, 287, 11566-11578.
[83] Kim, S. H.; Park, H. S.; M.S., L.; Cho, Y.; Kim, Y. S.; Hwang, J. T.; Sung, M. J.; Kim, M. S.; Kwon, D. Y. Vitisin A inhibits adipo-cyte differentiation through cell cycle arrest in 3T3-L1 cells. Bio-chem. Biophys. Res. Commun., 2008, 372, 108-113.
[84] Chiche, F.; Le Guillou, M.; Chetrite, G.; Lasnier, F.; Dugail, I.; Carpéné, C.; Moldes, M.; Fève, B. The antidepressant phenelzine alters differentiation of cultured human and mouse preadipocytes. Mol. Pharmacol., 2009, 75, 1052-1061.
[85] Lee, H.; Lee, Y. J.; Choi, H.; Ko, E. H.; Kim, J. W. Reactive oxy-gen species facilitate adipocyte differentiation by accelerating mi-totic clonal expansion. J. Biol. Chem., 2009, 284, 10601-10609.
[86] Juman, S.; Yasui, N.; Okuda, H.; Ueda, A.; Negishi, H.; Miki, T.; Ikeda, K. Caffeic acid phenethyl ester suppresses the production of adipocytokines, leptin, tumor necrosis factor -alpha and resistin, during differentiation to adipocytes in 3T3-L1 cells. Biol. Pharm. Bull., 2011, 34, 490-494.
[87] Szkudelska, K.; Nogowski, L.; Szkudelski, T. Resveratrol, a natu-rally occurring diphenolic compound, affects lipogenesis, lipolysis and antilipolytic action of insulin in isolated rat adipocytes. J. Ster-oid Biochem. Mol. Biol., 2009, 113, 17-24.
[88] Pandey, P. R.; Okuda, H.; Watabe, M.; Pai, S. K.; Liu, W.; Koba-yashi, A.; Xing, F.; Fukuda, K.; Hirota, S.; Sugai, T.; Wakabayashi, G.; Koeda, K.; Kashiwaba, M.; Suzuki, K.; Chiba, T.; Endo, M.; Fujioka, T.; Tanji, S.; Mo , Y. Y.; Cao, D.; Wilber, A. C.; Watabe, K. Resveratrol suppresses growth of cancer stem-like cells by in-hibiting fatty acid synthase. Breast Cancer Res. Treat., 2011, 130,387-398.
[89] Mercader, J.; Palou, A.; Bonet, M. L. Resveratrol enhances fatty acid oxidation capacity and reduces resistin and Retinol-Binding Protein 4 expression in white adipocytes. J. Nutr. Biochem., 2011,22, 828-834.
[90] Lasa, A.; Schweiger, M.; Kotzbeck, P.; Churruca, I.; Simón, E.; Zechner, R.; Del Puy Portillo, M. Resveratrol regulates lipolysis via adipose triglyceride lipase. J. Nutr. Biochem., 2012, 23, 379-384.
[91] Kuppusamy, U. R.; Das, N. P. Potentiation of beta-adrenoceptor agonist-mediated lipolysis by quercetin and fisetin in isolated rat adipocytes. Biochem. Pharmacol., 1994, 47, 521-529.
[92] Szkudelska, K.; Nogowski, L.; Szkudelski, T. The inhibitory effect of resveratrol on leptin secretion from rat adipocytes. Eur. J. Clin. Invest., 2009, 39, 899-905.
[93] Olholm, J.; Paulsen, S. K.; Cullberg, K. B.; Richelsen, B.; Peder-sen, S. B. Anti-inflammatory effect of resveratrol on adipokine ex-pression and secretion in human adipose tissue explants. Int. J. Obes. (Lond), 2010, 34, 1546-1553.
[94] Costa, C. S.; Rohden, F.; Hammes, T. O.; Margis, R.; Bortolotto, J. W.; Padoin, A. V.; Mottin, C. C.; Guaragna, R. M. Resveratrol upregulated SIRT1, FOXO1, and adiponectin and downregulated PPAR�1-3 mRNA expression in human visceral adipocytes. Obes. Surg., 2011, 21, 356-361.
[95] Wang, A.; Liu, M.; Liu, X.; Dong, L. Q.; Glickman, R. D.; Slaga, T. J.; Zhou, Z.; Liu, F. Up-regulation of adiponectin by resveratrol: the essential roles of the Akt/FOXO1 and AMP-activated protein kinase signaling pathways and DsbA-L. J. Biol. Chem., 2011, 286,60-66.
[96] Chuang, C. C.; Martinez, K.; Xie, G.; Kennedy, A.; Bumrungpert, A.; Overman, A.; Jia, W.; McIntosh, M. K. Quercetin is equally or more effective than resveratrol in attenuating tumor necrosis factor alpha-mediated inflammation and insulin resistance in primary hu-man adipocytes. Am. J. Clin. Nutr., 2010, 92, 1511-1521.
[97] Eseberri, I.; Lasa, A.; Churruca, I.; Portillo, M. P. Resveratrol metabolites modify adipokine expression and secretion in 3T3-L1 pre-adipocytes and mature adipocytes. PLoS One, 2013, 8, e63918.
[98] Szkudelski, T.; Szkudelska, K. Anti-diabetic effects of resveratrol. Ann. N. Y. Acad. Sci., 2011, 1215, 34-39.
[99] Dao, T. M.; Waget, A.; Klopp, P.; Serino, M.; Vachoux, C.; Pechere, L.; Drucker, D. J.; Champion, S.; Barthélemy, S.; Barra, Y.; Burcelin, R.; Sérée, E. Resveratrol increases glucose induced GLP-1 secretion in mice: a mechanism which contributes to the glycemic control. PLoS One, 2011, 6, e20700.
[100] Wong, Y. T.; Gruber, J.; Jenner, A. M.; Ng, M. P.; Ruan, R.; Tay, F. E. Elevation of oxidative-damage biomarkers during aging in F2 hybrid mice: protection by chronic oral intake of resveratrol. Free Radic. Biol. Med., 2009, 46, 799-809.
[101] De Santi, C.; Pietrabissa, A.; Mosca, F.; Pacifici, G. M. Glucuroni-dation of resveratrol, a natural product present in grape and wine, in the human liver. Xenobiotica, 2000, 30, 1047-1054.
[102] Johnson, J. J.; Nihal, M.; Siddiqui, I. A.; Scarlett, C. O.; Bailey, H. H.; Mukhtar, H.; Ahmad, N. Enhancing the bioavailability of res-veratrol by combining it with piperine. Mol. Nutr. Food Res., 2011,55, 1169-1176.
[103] Yang, J. Y.; Della-Fera, M. A.; Rayalam, S.; Ambati, S.; Hartzell, D. L.; Park, H. J.; Baile, C. A. Enhanced inhibition of adipogenesis and induction of apoptosis in 3T3-L1 adipocytes with combinations of resveratrol and quercetin. Life Sci., 2008, 82, 1032-1039.
[104] Rayalam, S.; Della-Fera, M. A.; Yang, J. Y.; Park, H. J.; Ambati, S.; Baile, C. A. Resveratrol potentiates genistein's antiadipogenic and proapoptotic effects in 3T3-L1 adipocytes. J. Nutrition, 2007,137, 2668-2673.
[105] Bruckbauer, A.; Zemel, M. B.; Thorpe, T.; Akula, M. R.; Stuckey, A. C.; Osborne, D.; Martin, E. B.; Kennel, S.; Wall, J. S. Synergis-tic effects of leucine and resveratrol on insulin sensitivity and fat metabolism in adipocytes and mice. Nutr. Metab. (Lond.), 2012, 9,77.
[106] Bruckbauer, A.; Zemel, M. B. Synergistic effects of polyphenols and methylxanthines with leucine on AMPK/sirtuin-mediated metabolism in muscle cells and adipocytes. PLoS One, 2014, 9, e89166.

164 Current Medicinal Chemistry, 2015, Vol. 22, No. 1 Carpéné et al.
[107] Most, J.; Goossens, G. H.; Jocken, J. W.; Blaak, E. E. Short-term supplementation with a specific combination of dietary polyphe-nols increases energy expenditure and alters substrate metabolism in overweight subjects. Int. J. Obes. (Lond.), 2014, 38, 698-706.
[108] Arias, N.; Macarulla, M. T.; Aguirre, L.; Martínez-Castaño, M. G.; Gómez-Zorita, S.; Miranda, J.; Martínez, J. A.; Portillo, M. P. The combination of resveratrol and conjugated linoleic acid is not useful in preventing obesity. J. Physiol. Biochem., 2011, 67, 471-477.
[109] Arias, N.; Miranda, J.; Macarulla, M. T.; Aguirre, L.; Fernandez-Quintela, A.; Andres-Lacueva, C.; Urpi-Sarda, M.; Portillo, M. P. The combination of resveratrol and conjugated linoleic acid attenu-ates the individual effects of these molecules on triacylglycerol me-tabolism in adipose tissue. Eur. J. Nutr., 2014, 53, 575-582.
[110] Lasa, A.; Miranda, J.; Churruca, I.; Simón, E.; Arias, N.; Milagro, F.; Martínez, J. A.; Portillo, M. P. The combination of resveratrol and CLA does not increase the delipidating effect of each molecule in 3T3-L1 adipocytes. Nutr. Hosp., 2011, 26, 997-1003.
[111] Kennedy, A.; Overman, A.; Lapoint, K.; Hopkins, R.; West, T.; Chuang, C. C.; Martinez, K.; Bell, D.; McIntosh, M. Conjugated li-noleic acid-mediated inflammation and insulin resistance in human adipocytes are attenuated by resveratrol. J. Lipid Res., 2009, 50,225-232.
[112] Herranz-López, M.; Fernández-Arroyo, S.; Pérez-Sanchez, A.; Barrajón-Catalán, E.; Beltrán-Debón, R.; Menéndez, J. A.; Alonso-Villaverde, C.; Segura-Carretero, A.; Joven, J.; Micol, V. Syner-gism of plant-derived polyphenols in adipogenesis: perspectives and implications. Phytomedicine, 2012, 19, 253-261.
[113] Kapetanovic, I. M.; Muzzio, M.; Huang, Z.; Thompson, T. N.; McCormick, D. L. Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother. Pharmacol., 2011, 68,593-601.
[114] Lin, H. S.; Yue, B. D.; Ho, P. C. Determination of pterostilbene in rat plasma by a simple HPLC�UV method and its application in preclinical pharmacokinetic study. Biomed. Chromatography,2009, 23, 1308-1315.
[115] Grover, J. K.; Vats, V.; Yadav, S. S. Pterocarpus marsupium ex-tract (Vijayasar) prevented the alteration in metabolic patterns in-duced in the normal rat by feeding an adequate diet containing fructose as sole carbohydrate. Diabetes Obes. Metab., 2005, 7, 414-420.
[116] Satheesh, M. A.; Pari, L. The antioxidant role of pterostilbene in streptozotocin nicotinamide induced type 2 diabetes mellitus in Wistar rats. J. Pharm. Pharmacol., 2006, 58, 1483-1490.
[117] Hsu, C. L.; Lin, Y. J.; Ho, C. T.; Yen, G. C. Inhibitory effects of garcinol and pterostilbene on cell proliferation and adipogenesis in 3T3-L1 cells. Food Funct., 2012, 3, 49-57.
[118] Hsieh, T. C.; Bennett, D. J.; Lee, Y. S.; Wu, E.; Wu, J. M. In silicoand biochemical analyses identify quinone reductase 2 as a target of piceatannol. Curr. Med. Chem. 2013, 20, 4195-4202.
[119] Piotrowska, H.; Kucinska, M.; Murias, M. Biological activity of piceatannol: leaving the shadow of resveratrol. Mutat. Res., 2012,750, 60-82.
[120] He, S.; Yan, X. From resveratrol to its derivatives: new sources of natural antioxidant. Curr. Med. Chem., 2013, 20, 1005-1017.
Received: April 30, 2014 Revised: June 17, 2014 Accepted: August 02, 2014
Related Documents











