Novel RasGRF1-derived Tat-fused peptides inhibiting Ras-dependent proliferation and migration in mouse and human cancer cells Elena Sacco a, ⁎, David Metalli a , Michela Spinelli a , Romilde Manzoni a , Maria Samalikova a , Rita Grandori a , Andrea Morrione b , Silvio Traversa c , Lilia Alberghina a , Marco Vanoni a, ⁎⁎ a Department of Biotechnology and Biosciences, University of Milano-Bicocca, Piazza della Scienza 2, 20126 Milano, Italy b Department of Urology, the Endocrine Mechanisms and Hormone Action Program, Kimmel Cancer Center, Thomas Jefferson University, 233 South 10th Street, BLSB Room 620, Philadelphia, PA, 19107, USA c Creabilis Therapeutics Srl, Bioindustry Park Silvano Fumero, Via Ribes 5, Colleretto Giacosa (TO), Italy abstract article info Article history: Received 30 March 2011 Accepted 9 May 2011 Available online xxxx Keywords: Ras inhibitors Ras-dependent transformation Anticancer agents Drug design Peptide engineering Cell-penetrating peptides Drug delivery Mutations of RAS genes are critical events in the pathogenesis of different human tumors and Ras proteins represent a major clinical target for the development of specific inhibitors to use as anticancer agents. Here we present RasGRF1-derived peptides displaying both in vitro and in vivo Ras inhibitory properties. These peptides were designed on the basis of the down-sizing of dominant negative full-length RasGRF1 mutants. The over-expression of these peptides can revert the phenotype of K-RAS transformed mouse fibroblasts to wild type, as monitored by several independent biological readouts, including Ras–GTP intracellular levels, ERK activity, morphology, proliferative potential and anchorage independent growth. Fusion of the RasGRF1- derived peptides with the Tat protein transduction domain allows their uptake into mammalian cells. Chemically synthesized Tat-fused peptides, reduced to as small as 30 residues on the basis of structural constraints, retain Ras inhibitory activity. These small peptides interfere in vitro with the GEF catalyzed nucleotide dissociation and exchange on Ras, reduce cell proliferation of K-RAS transformed mouse fibroblasts, and strongly reduce Ras-dependent IGF-I-induced migration and invasion of human bladder cancer cells. These results support the use of RasGRF1-derived peptides as model compounds for the development of Ras inhibitory anticancer agents. © 2011 Elsevier Inc. All rights reserved. 1. Introduction Ras proteins are monomeric small guanine nucleotide-binding proteins whose structure is conserved in eukaryotes. They bind both GDP and GTP with similar affinity and possess low intrinsic GTPase activity. When bound to GDP, Ras proteins are in an inactive state unable to activate downstream pathways, while in the GTP-bound state, they regulate key cellular processes, in higher eukaryotes including cell proliferation, differentiation, migration and survival (Hingorani and Tuveson, 2003; Mitin et al., 2005; Vetter and Wittinghofer, 2001). Ras– GTP levels in vivo are controlled by the competing activities of Guanine nucleotide Exchange Factors (GEFs) that, by catalyzing the nucleotide- bound dissociation on Ras, promote GDP/GTP exchange inducing Ras activation and GTPase Activating Proteins (GAPs) that, by increasing the intrinsic GTPase activity of Ras, promote the hydrolysis of bound-GTP leading to Ras inactivation (Bos et al., 2007). In mammalian cells, three major Ras-specific GEF classes exist: Sos, RasGRF1/Cdc25 Mm and GRP/Cal-DAG-GEF [reviewed in (Quilliam et al., 2002; Zheng and Quilliam, 2003)]. Despite the different domain organization and the involvement in different signal transduction pathways, they share a conserved catalytic domain containing all the structural determinants for Ras binding and Ras-bound nucleotide exchange reaction (Freedman et al., 2006, 2009; Sacco et al., 2005, 2006). Mammalian Ras-specific GAPs include several multi-domain proteins, such as p120-RasGAP (RASA1), neurofibromin (NF1), p135 SynGAP, GAP1 protein (GAP1m or RASA2), GAP1IP4BP (or RASA3), CAPRI (or RASA4), and RASAL [reviewed in (Bernards, 2003; Pamonsinlapatham et al., 2009)]. Mutated RAS genes encoding constitutively active GTP-bound Ras proteins, insensitive to GAP action or self-sufficient (i.e., GEF- independent) in guanine nucleotide exchange activity, are found in several precancerous states and in 30% of human tumors, with a great incidence in colon (45%) and pancreatic (90%) adenocarcinomas (Chin et al., 1999; DeNicola and Tuveson, 2009; Karnoub and Weinberg, 2008; Wittinghofer and Waldmann, 2000). An updated Biotechnology Advances xxx (2011) xxx–xxx ⁎ Correspondence to: E. Sacco, Department of Biotechnology and Biosciences, University of Milano-Bicocca, Piazza della Scienza 2, 20126, Milano, Italy. Tel.: + 39 02 64483534; fax: +39 02 64483565. ⁎⁎ Correspondence to: M. Vanoni, Department of Biotechnology and Biosciences, University of Milano-Bicocca, Piazza della Scienza 2, 20126, Milano, Italy. Tel.: + 39 02 64483525; fax: + 39 02 64483565. E-mail addresses: [email protected] (E. Sacco), [email protected] (A. Morrione), [email protected] (S. Traversa), [email protected] (M. Vanoni). JBA-06410; No of Pages 11 0734-9750/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.biotechadv.2011.05.011 Contents lists available at ScienceDirect Biotechnology Advances journal homepage: www.elsevier.com/locate/biotechadv Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fused peptides inhibiting Ras-dependent proliferation and migration in mouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.biotechadv.2011.05.011

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biotechnology Advances xxx (2011) xxx–xxx
JBA-06410; No of Pages 11
Contents lists available at ScienceDirect
Biotechnology Advances
j ourna l homepage: www.e lsev ie r.com/ locate /b iotechadv
Novel RasGRF1-derived Tat-fused peptides inhibiting Ras-dependent proliferationand migration in mouse and human cancer cells
Elena Sacco a,⁎, David Metalli a, Michela Spinelli a, Romilde Manzoni a, Maria Samalikova a, Rita Grandori a,Andrea Morrione b, Silvio Traversa c, Lilia Alberghina a, Marco Vanoni a,⁎⁎a Department of Biotechnology and Biosciences, University of Milano-Bicocca, Piazza della Scienza 2, 20126 Milano, Italyb Department of Urology, the Endocrine Mechanisms and Hormone Action Program, Kimmel Cancer Center, Thomas Jefferson University, 233 South 10th Street, BLSB Room 620,Philadelphia, PA, 19107, USAc Creabilis Therapeutics Srl, Bioindustry Park Silvano Fumero, Via Ribes 5, Colleretto Giacosa (TO), Italy
⁎ Correspondence to: E. Sacco, Department of BioUniversity of Milano-Bicocca, Piazza della Scienza 2, 202 64483534; fax: +39 02 64483565.⁎⁎ Correspondence to: M. Vanoni, Department of BUniversity of Milano-Bicocca, Piazza della Scienza 2, 20164483525; fax: +39 02 64483565.
E-mail addresses: [email protected] (E. Sacco), a(A. Morrione), [email protected] (S. Traversa),(M. Vanoni).
0734-9750/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.biotechadv.2011.05.011
Please cite this article as: Sacco E, et al, Novmouse and human cancer cells, Biotechnol
a b s t r a c t
a r t i c l e i n f oArticle history:Received 30 March 2011Accepted 9 May 2011Available online xxxx
Keywords:Ras inhibitorsRas-dependent transformationAnticancer agentsDrug designPeptide engineeringCell-penetrating peptidesDrug delivery
Mutations of RAS genes are critical events in the pathogenesis of different human tumors and Ras proteinsrepresent amajor clinical target for the development of specific inhibitors to use as anticancer agents. Here wepresent RasGRF1-derived peptides displaying both in vitro and in vivo Ras inhibitory properties. Thesepeptides were designed on the basis of the down-sizing of dominant negative full-length RasGRF1 mutants.The over-expression of these peptides can revert the phenotype of K-RAS transformed mouse fibroblasts towild type, as monitored by several independent biological readouts, including Ras–GTP intracellular levels,ERK activity, morphology, proliferative potential and anchorage independent growth. Fusion of the RasGRF1-derived peptides with the Tat protein transduction domain allows their uptake into mammalian cells.Chemically synthesized Tat-fused peptides, reduced to as small as 30 residues on the basis of structuralconstraints, retain Ras inhibitory activity. These small peptides interfere in vitro with the GEF catalyzednucleotide dissociation and exchange on Ras, reduce cell proliferation of K-RAS transformed mousefibroblasts, and strongly reduce Ras-dependent IGF-I-induced migration and invasion of human bladdercancer cells. These results support the use of RasGRF1-derived peptides as model compounds for thedevelopment of Ras inhibitory anticancer agents.
technology and Biosciences,0126, Milano, Italy. Tel.: +39
iotechnology and Biosciences,26, Milano, Italy. Tel.: +39 02
[email protected]@unimib.it
l rights reserved.
el RasGRF1-derived Tat-fused peptides inhibAdv (2011), doi:10.1016/j.biotechadv.2011.0
© 2011 Elsevier Inc. All rights reserved.
1. Introduction
Ras proteins are monomeric small guanine nucleotide-bindingproteins whose structure is conserved in eukaryotes. They bind bothGDP and GTP with similar affinity and possess low intrinsic GTPaseactivity.When bound toGDP, Ras proteins are in an inactive state unableto activate downstream pathways, while in the GTP-bound state, theyregulate key cellular processes, in higher eukaryotes including cellproliferation, differentiation, migration and survival (Hingorani andTuveson, 2003; Mitin et al., 2005; Vetter and Wittinghofer, 2001). Ras–GTP levels in vivo are controlled by the competing activities of Guaninenucleotide Exchange Factors (GEFs) that, by catalyzing the nucleotide-bound dissociation on Ras, promote GDP/GTP exchange inducing Ras
activation and GTPase Activating Proteins (GAPs) that, by increasing theintrinsic GTPase activity of Ras, promote the hydrolysis of bound-GTPleading to Ras inactivation (Bos et al., 2007).
Inmammalian cells, threemajor Ras-specific GEF classes exist: Sos,RasGRF1/Cdc25Mm and GRP/Cal-DAG-GEF [reviewed in (Quilliam etal., 2002; Zheng and Quilliam, 2003)]. Despite the different domainorganization and the involvement in different signal transductionpathways, they share a conserved catalytic domain containing all thestructural determinants for Ras binding and Ras-bound nucleotideexchange reaction (Freedman et al., 2006, 2009; Sacco et al., 2005,2006). Mammalian Ras-specific GAPs include several multi-domainproteins, such as p120-RasGAP (RASA1), neurofibromin (NF1), p135SynGAP, GAP1 protein (GAP1m or RASA2), GAP1IP4BP (or RASA3),CAPRI (or RASA4), and RASAL [reviewed in (Bernards, 2003;Pamonsinlapatham et al., 2009)].
Mutated RAS genes encoding constitutively active GTP-bound Rasproteins, insensitive to GAP action or self-sufficient (i.e., GEF-independent) in guanine nucleotide exchange activity, are found inseveral precancerous states and in 30% of human tumors, with a greatincidence in colon (45%) and pancreatic (90%) adenocarcinomas(Chin et al., 1999; DeNicola and Tuveson, 2009; Karnoub andWeinberg, 2008; Wittinghofer and Waldmann, 2000). An updated
iting Ras-dependent proliferation and migration in5.011

2 E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
review on the incidence of RAS mutation in human cancer is reportedin Pamonsinlapatham et al. (2009). The presence of oncogenic RASmutants is also a predictive marker of resistance to conventionalanticancer therapies (Di Fiore et al., 2010; Nazarian et al., 2010; Rizzoet al., 2010; Tentler et al., 2010).
Ras oncogenic mutants induce physiological changes in cells, typicalof tumor transformation: (i) self-sufficiency of proliferation from themitogenic signal by the activation of Raf1/MAPK signaling, (ii) resistanceto apoptotic stimuli by the deregulation of Akt/BAD or GSK3 signaling,(iii) ability to migrate and to invade adjacent tissues, ultimately leadingto metastasis formation, mediated by Rho/Rac and RalGEF activation,(iv) ability to promote angiogenesis in particular to assure the survival ofcancer cells within the tumor mass by the activation of the HIF1-alphaexpression (Karnoub andWeinberg, 2008).
Aberrant Ras signaling also contributes to the pathogenesis ofhuman developmental disorders, including Costello, Noonan andCardio-Facio-Cutaneous syndromes (Aoki et al., 2008; Schubbert et al.,2007; Tidyman and Rauen, 2009; Zenker, 2009). Such syndromes areusually associated with gain-of-function Ras mutations, even thoughthe levels of K-Ras activation is less pronounced than in the oncogenicK-Ras proteins (Schubbert et al., 2007).
Ras proteins have been proposed as major clinical targets for anti-cancer therapy (Adjei, 2001, 2008). Several strategies have beenexplored in order to inhibit aberrant Ras signaling: (i) down-regulation of Ras expression achieved by antisense DNAs or siRNAs(Brummelkamp et al., 2002; Jeong et al., 2010; Zhang et al., 2006);(ii) targeting of Ras mutants using vaccine (Abou-Alfa et al., 2010);(iii) inhibition ofmembrane localizationof Ras by inhibitors of farnesyl-,geranylgeranyl- and palmitoyl-transferases (Bustinza-Linares et al.,2010; Dekker and Hedberg, 2010; Ducker et al., 2006; Le Moulec et al.,2009; Leonard, 1997; Mesa, 2006; Morgillo and Lee, 2006);(iv) enhancement of GTPase activity of oncogenic Ras mutants(Ahmadian et al., 1999); (v) inhibition of interaction between Ras andits downstream effectors (Tanaka and Rabbitts, 2010; Waldmann et al.,2004); (vi) inhibition of Ras downstream effectors (Roberts and Der,2007;Wong, 2009); (vii) inhibition of the nucleotide exchange reactionon Ras by small organic molecules (Airoldi et al., 2007; Palmioli et al.,2009a,b; Sacco et al., 2011). However, most of such strategies have notproduced the expected results in clinical trials (Morgillo and Lee, 2006;Stathopoulos et al., 2008). To date, no selective and specific inhibitor ofthe oncogenic Ras proteins is available for routine clinical use.
We have previously reported dominant negativemutants of the Ras-specificGEF, RasGRF1. Suchmutants, RasGRF1W1056E andRasGRF1T1184E
(DN-GEFs, 1256 aa) display Ras inhibitory properties both in vitro and invivo. In vitro these mutants can efficiently compete with wild-type GEFfor binding toRas anddisrupt theRas/GEF complex (Vanoni et al., 1999).Over-expression of full lengthmolecules harboring suchmutations in K-RAS transformed mouse fibroblasts efficiently attenuates Ras signalingand reverts their transformed phenotype to normal on the basis ofseveral independent readouts, including intracellular Ras–GTP level,morphology, proliferative potential, anchorage-independent growth,transcriptional reprogramming and metabolic alterations (Bossù et al.,2000; Chiaradonna et al., 2006a,b).
Herewe showthatRasGRF1-derivedpeptides– as small as 30 residues– centered on residue 1056 retain Ras inhibitory properties – both in vitroand in vivo – similarly to the full length parental protein and inhibit Ras-dependent proliferation andmigration in mouse and human cancer cells.These results support the use of RasGRF1-derived peptides as leadcompounds for the development of Ras inhibitory anticancer agents.
2. Materials and methods
2.1. Recombinant and genetic manipulation
Recombinant DNA manipulations were performed according tostandard methods (Sambrook et al., 1989). The RasGRF1-derived AB-
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
encoding region (encoding residues 1024–1090 of RasGRF1/Cdc25Mm
mature protein) was amplified by PCR using the primers: 5′-CGGGATCCATGGAAAACCACCCAGCCCTGGAG-3′ and 5′-CCGGAATTC-TACTCGTTTCGGATAATTTCTGAAGC-3′ containing respectively BamHIand EcoRI restriction sites and inserted in the expression vector formammalian cells pcDNA3 (Invitrogen).
The sequence encoding the peptide His-Tat-AB described in Fig. 3Awas amplified by PCR using the primers: 5′-AACTGCAGGATCCATATG-CACCACCATCATCATCACGGGTATGGACGCAAGAAACGACGT-CAAAGGCGGAGAGGTGAAAACCACCCAGCCCTGGAG-3′ containingBamHI and NdeI restriction sites and 5′-CCGGAATTCTACTCGTTTCG-GATAATTTCTGAAGC-3′, containing EcoRI restriction site. Such se-quence was subcloned in the expression vector for Escherichia colipET21a (EMD Biosciences), usingNdeI and EcoRI and in pcDNA3, usingBamHI and EcoRI.
2.2. Cell lines and cell biology methods
Parental NIH3-T3 (from ATCC) and K-RAS transformed (NIH-RAS)mouse fibroblasts, obtained and characterized as described in Bossù etal. (2000) according to a procedure described by Pulciani et al. (1985),were routinely grown at 37 °C in a humidified atmosphere of 5% CO2
in Dulbecco's modified Eagle's medium (D-MEM) supplemented with10% Newborn Calf Serum (NCS), 2 mM glutamine, 100 units/mlpenicillin and 100 mg/ml streptomycin. Cells were passaged usingtrypsin–EDTA (Invitrogen).
Invasive urothelial carcinoma-derived human 5637 cell lines wereobtained from the American Type Culture Collection (Manassas, VA).Cells were maintained in RPMI medium supplemented with 10% fetalbovine serum. Migration experiments were performed in serum-freemedium (SFM) that is D-MEM supplemented with 0.1% bovine serumalbumin and 50 μg/ml of transferrin (Sigma-Aldrich Corp., St.Louis,MO).
2.2.1. Stable transfectionCell lines stably and constitutively expressing RasGRF1-derivatives
were obtained by the transfection of NIH-RAS cells with pcDNA3-based constructs by calcium phosphate method followed by growth inselective medium containing 0.7 mg/ml geneticin (G418, Sigma).After 15–20 days of selection, clones were selected by the cloning ringtechnique. Successful transfection was confirmed by RT-PCR (kitInvitrogen) using specific primers as described above.
2.2.2. Fos-luciferase assayParental and K-RAS transformed mouse fibroblasts were transient-
ly cotransfected with 0.33 μg fos-luciferase plasmid and 1.0 μgpcDNA3-derived plasmid for expression of RasGRF1 derivatives aspreviously described (Vanoni et al., 1999), using Lipofectaminereagent (Invitrogen). After transfection, cells were starved for 24 hin serum-free medium supplemented with 4 mg/ml transferrin and0.346 ng/ml sodium selenite and collected. Luciferase activity wasassayed using the luciferase assay system with reporter lysis buffer(Promega) and normalized to protein content determined byBradford assay (Bio-Rad reagent).
2.2.3. Ras–GTP pull down assayRas–GTP intracellular level was assayed by a pull down assay using
the Ras binding domain of Raf1 (RBD) bound to glutathione–sepharose essentially [as described in (Taylor and Shalloway,1996)]. 0.4 mg of total clarified extracts was incubated with 30 mlof RBD-bound glutathione–sepharose 1 h at 4 °C under gentleshaking. After abundant washes in Shalloway buffer (Taylor andShalloway, 1996), bound Ras–GTP was eluted with reducing Laemmlisample buffer (Laemmli, 1970), loaded on 12% SDS-PAGE anddetected by western blotting analysis with anti-H-Ras 259 antibody(Santa Cruz Biotechnology). Ras–GTP levels were determined by
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011

3E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
densitometric scanning and quantification with J-Image (rsbweb.-nih.gov/ij/). Data are expressed after normalization to the level of totalRas or vinculin, determined by western blot analysis of total clarifiedextracts using respectively anti-H-Ras 259 or anti-vinculin H-300antibodies (Santa Cruz Biotechnology).
2.2.4. MAPK activityMouse fibroblasts were plated (6000 cells/cm2) in 60-mm tissue
culture dishes. After 18 h several concentrations of peptide of interest(or water) were added to the cell culture. After 24 h from treatment,both plate-adherent and in suspension cells were harvested in lysisbuffer from PathSscan Sandwich ELISA kit (Cell Signaling). Thedetection of endogenous levels of Phospho-p44/42 MAPK wasperformed according to manufacturer's instructions, and the resultswere normalized on total protein content.
2.2.5. Soft agar colony formation assayStable transfectants or control cellswereplated (1.750 cells) in60-mm
tissue culture dishes containing 0.33% top agar and 0.5% bottom agar(Bacto agar; Difco, Detroit, MI, USA) supplemented with DMEM10% NCS,2 mMglutamine, 100 units/ml penicillin and 100 mg/ml streptomycin. Indetail 1.5 ml of amixture of 4 ml prewarmed (40 °C) 5×D-MEM(GIBCO),2 mlNCS, 0.2 ml200 mMglutamine, 0.2 ml 10,000 units/mlpenicillin and10,000 mg/ml streptomycinand6 mLprewarmed(40 °C)2.2%BactoAgarperplate (Difco) (0.66%final agar)weremixedwith1.5 mlcell suspension(3500 cells) and seeded over a 0.5% agar/D-MEM prelayer (4 ml) in a 60-mmdish. Semisolid 0.33% feeder layers (1.5 ml)were overlayed on top ofthe solidified layers. Cells were re-fed every 3±4 days with completemedium. After 3 weeks of incubation colonies were stained with MTT(Sigma) and counted.
2.2.6. Immunofluorescence microscopyNIH3T3 cells (6000/cm2) were plated on cover slip pre-treated
with 0.2% gelatin. After 24 h, cells were treated with Tat-engineeredpeptides (0.5 μM) for the indicated times. After treatments cells werefixed in 4% paraformaldehyde/PBS, permeabilized with 0.1% Triton X-100/PBS, blocked with 10% goat serum/PBS and probed with primaryantibody anti-histidine (1:100) (Santa Cruz Biotechnology CA, USA)in 10% goat serum/PBS for 1 h at room temperature. Samples werethen probed with Alexa Fluor 488 goat anti-rabbit IgG (1:400)(Molecular Probes/Invitrogen) in 10% goat serum/PBS for 30 min atroom temperature. Cover slips were mounted in DABCO-MOVIOL(Sigma) and counterstained with Dapi (Vector Laboratories).
The cover glasses were analyzed under a Nikon C600 ECLIPSEmicroscope equipped with a b/w Leica DG350F CCD camera. Theacquired images were processed in Adobe Photoshop 7.0.1 withadjustments of brightness and contrast.
2.2.7. Proliferation assaysFor growth kinetic and MTS experiments cells were plated into
respectively 6-well or 96-well flat-bottomed culture plates at thedensity of 3000 cells/cm2. At 18 h after seeding, predeterminedconcentrations of peptide of interest (or water) were added to the cellculture. After 24, 48, and 72 h from treatment, cells were harvestedand counted by Coulter Counter to obtain growth curves or treated for1 h with MTS tetrazolium reagent [from kit CellTiter 96® AQueousOneSolution Cell Proliferation Assay (PROMEGA)] that is bioreducedby metabolically active cells into a colored formazan product that issoluble in tissue culture medium. The quantity of formazan product,as measured by the absorbance at 490 nm recorded with a 96-wellplate reader, is directly proportional to the number of living cells inculture.
2.2.8. Migration and invasion assayMigration and invasion assays were performed as described in
Lovat et al. (2009) and Metalli et al. (2010), using Boyden chambers
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
24-well inserts with 8 μm PET membrane (for migration) or BDBioCoat™Matrigel™Matrix inserts (for invasion) (Becton Dickinson).Cells were grown in Serum FreeMedium (SFM) or SFM supplementedwith 50 ng/ml of IGF-I, used as a chemoattractant in the lowerchamber. 500 nM Tat-loopAB was added both in the upper and lowerchambers at the moment of cell seeding.
2.3. Protein purification and Ras-bound nucleotide dissociation assay
His-Tat-AB peptide, used in the cell-transduction experiments,was produced as described in Supplementary data.
Tat-loopAB and Tat-loopAB2cys peptides, synthesized by GenScript,were dissolved in water (W4502 SIGMA) at 1 mM final concentration.
Recombinant N-terminal His-tagged H-Ras protein, used in thenucleotide dissociation assays, was expressed in E. coli M15 [pREP4]strain harboring a pQE™-derived plasmid (Qiagen) and purified byaffinity chromatography using a Ni2+-NTA column (Qiagen) accord-ing to manufacturer's suggestions. The N-terminal GST-taggedRasGRF1 catalytic domain (aa 976–1262 of the mature protein),used in the nucleotide dissociation assays, was purified by glutathi-one–sepharose chromatography (Amersham Bioscience) as described(Martegani et al., 1992).
The exchange reaction between GDP and fluorescent MANT-GDP(Molecular Probes) on Ras was performed by adding directly in anUV-cuvette 0.25 μM His-tagged H-Ras–GDP, and an opportuneconcentration of peptide of interest, in a buffer composed by 40 mMHepes (pH 7.5), 5 mM DTE, and 10 mM MgCl2. After 300 s ofincubation, 1.5 μM MANT-GDP and 0.0625 μM GST-tagged RasGRF1catalytic domain (GEF) were added to catalyze the exchange. Thedissociation reaction was performed by adding to 0.25 μMHis-taggedH-Ras-MANT-GDP, preincubated for 300 s with the peptide, 200 μMGDP and 0.0625 μM GST-tagged RasGRF1 catalytic domain (GEF). Thefluorescence measurements were done at a temperature of 25 °Cusing a Perkin-Elmer LS45 luminescence spectrometer with anexcitation wavelength of 366 nm and emission wavelength of442 nm. Each reaction was monitored for at least 1500 s. Eachexperimental curve was fitted to a non linear ‘growth-sigmoidalHill’ curve (n=1) with OriginPro 8.0 software. In the graph themaximum value of relative fluorescence (100 on Y axis) represent thefully loaded Ras status obtained as start point in dissociation reactionand plateau of an exchange curve.
2.4. Mass spectrometry experiments
Mass-spectrometry measurements were performed on a hybridquadrupole-Time-of-Flight (Q-TOF) instrument (QSTAR ELITE, Ap-plied Biosystems, Foster City, CA, USA), equipped with a nano-ESIsample source. Metal-coated borosilicate capillaries (Proxeon, Oden-se, DK), with medium-length emitter tip of 1-mm internal diameter,were used to infuse the sample. The instrument was calibrated usingthe renine-inhibitor (1757.9 Da) (Applied Biosystems, Foster City, CA,USA) and its fragment (109.07 Da) as standards. Spectra wereacquired in the 1500–3000 m/z range, with accumulation time of1 s, ion-spray voltage of 1200–1500 V, declustering potential of 80 V,and instrument interface of 50 °C. Spectra were averaged over a timeperiod of at least 3 min. Data analysis was performed by the programAnalyst QS 2.0 (Applied Biosystems, Foster City, CA, USA). The sampleswere prepared in 5 mM ammonium acetate pH 6.5.
3. Results and discussion
3.1. Design of a first generation of peptides derived from the RasGRF1WE
Ras inhibitory mutant
RasGRF1 is a Ras activator that promotes guanine nucleotideexchange on Ras, as described in Section 1. We previously
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011
4 E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
demonstrated that the substitution of Trp1056 with glutamate turnsfull-length RasGRF1 into a dominant negative protein (RasGRF1WE),which inhibits Ras both in vitro (Vanoni et al., 1999) and in vivo (Bossùet al., 2000; Chiaradonna et al., 2006b). This mutant is catalyticallyinactive, but retains affinity and specificity for Ras, efficientlycompeting with wild-type GEFs, thus down-regulating Ras signaling.
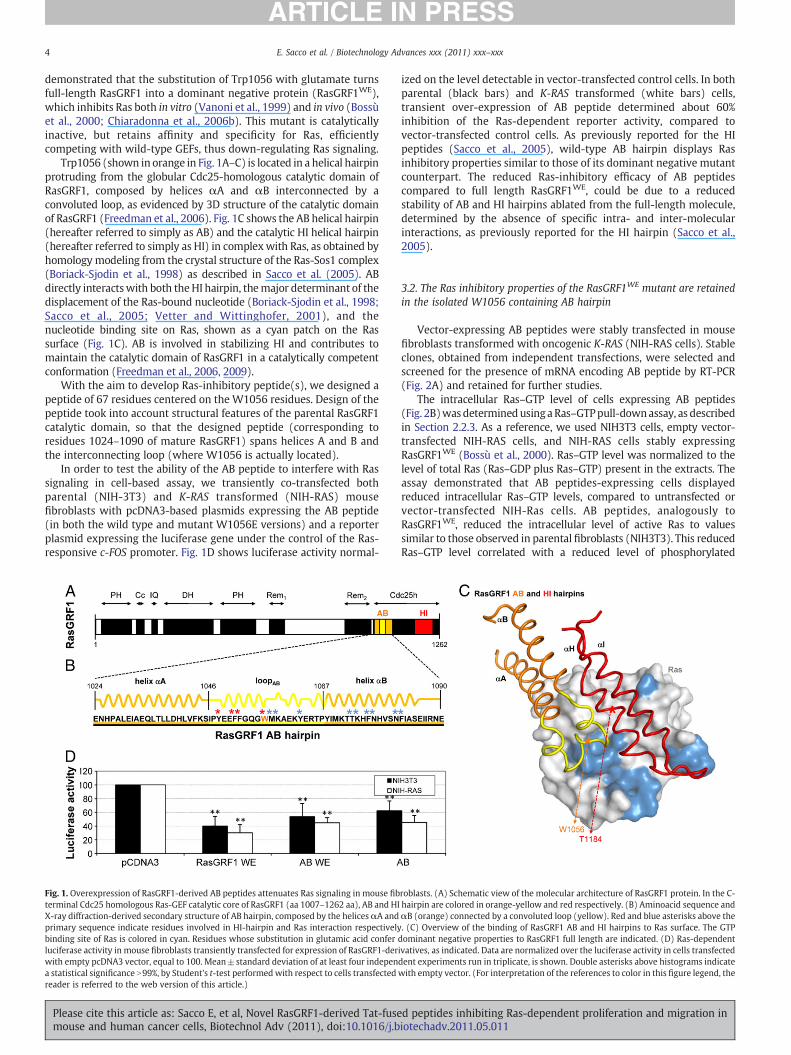
Trp1056 (shown in orange in Fig. 1A–C) is located in a helical hairpinprotruding from the globular Cdc25-homologous catalytic domain ofRasGRF1, composed by helices αA and αB interconnected by aconvoluted loop, as evidenced by 3D structure of the catalytic domainof RasGRF1 (Freedman et al., 2006). Fig. 1C shows the AB helical hairpin(hereafter referred to simply as AB) and the catalytic HI helical hairpin(hereafter referred to simply as HI) in complexwith Ras, as obtained byhomology modeling from the crystal structure of the Ras-Sos1 complex(Boriack-Sjodin et al., 1998) as described in Sacco et al. (2005). ABdirectly interactswith both theHI hairpin, themajor determinant of thedisplacement of the Ras-bound nucleotide (Boriack-Sjodin et al., 1998;Sacco et al., 2005; Vetter and Wittinghofer, 2001), and thenucleotide binding site on Ras, shown as a cyan patch on the Rassurface (Fig. 1C). AB is involved in stabilizing HI and contributes tomaintain the catalytic domain of RasGRF1 in a catalytically competentconformation (Freedman et al., 2006, 2009).
With the aim to develop Ras-inhibitory peptide(s), we designed apeptide of 67 residues centered on the W1056 residues. Design of thepeptide took into account structural features of the parental RasGRF1catalytic domain, so that the designed peptide (corresponding toresidues 1024–1090 of mature RasGRF1) spans helices A and B andthe interconnecting loop (where W1056 is actually located).
In order to test the ability of the AB peptide to interfere with Rassignaling in cell-based assay, we transiently co-transfected bothparental (NIH-3T3) and K-RAS transformed (NIH-RAS) mousefibroblasts with pcDNA3-based plasmids expressing the AB peptide(in both the wild type and mutant W1056E versions) and a reporterplasmid expressing the luciferase gene under the control of the Ras-responsive c-FOS promoter. Fig. 1D shows luciferase activity normal-
Fig. 1. Overexpression of RasGRF1-derived AB peptides attenuates Ras signaling in mouse fibterminal Cdc25 homologous Ras-GEF catalytic core of RasGRF1 (aa 1007–1262 aa), AB and HIX-ray diffraction-derived secondary structure of AB hairpin, composed by the helicesαA andprimary sequence indicate residues involved in HI-hairpin and Ras interaction respectivelybinding site of Ras is colored in cyan. Residues whose substitution in glutamic acid conferluciferase activity in mouse fibroblasts transiently transfected for expression of RasGRF1-derwith empty pcDNA3 vector, equal to 100. Mean±standard deviation of at least four indepena statistical significance N99%, by Student's t-test performedwith respect to cells transfectedreader is referred to the web version of this article.)
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
ized on the level detectable in vector-transfected control cells. In bothparental (black bars) and K-RAS transformed (white bars) cells,transient over-expression of AB peptide determined about 60%inhibition of the Ras-dependent reporter activity, compared tovector-transfected control cells. As previously reported for the HIpeptides (Sacco et al., 2005), wild-type AB hairpin displays Rasinhibitory properties similar to those of its dominant negative mutantcounterpart. The reduced Ras-inhibitory efficacy of AB peptidescompared to full length RasGRF1WE, could be due to a reducedstability of AB and HI hairpins ablated from the full-length molecule,determined by the absence of specific intra- and inter-molecularinteractions, as previously reported for the HI hairpin (Sacco et al.,2005).
3.2. The Ras inhibitory properties of the RasGRF1WE mutant are retainedin the isolated W1056 containing AB hairpin
Vector-expressing AB peptides were stably transfected in mousefibroblasts transformed with oncogenic K-RAS (NIH-RAS cells). Stableclones, obtained from independent transfections, were selected andscreened for the presence of mRNA encoding AB peptide by RT-PCR(Fig. 2A) and retained for further studies.
The intracellular Ras–GTP level of cells expressing AB peptides(Fig. 2B)wasdeterminedusingaRas–GTPpull-downassay, as describedin Section 2.2.3. As a reference, we used NIH3T3 cells, empty vector-transfected NIH-RAS cells, and NIH-RAS cells stably expressingRasGRF1WE (Bossù et al., 2000). Ras–GTP level was normalized to thelevel of total Ras (Ras–GDP plus Ras–GTP) present in the extracts. Theassay demonstrated that AB peptides-expressing cells displayedreduced intracellular Ras–GTP levels, compared to untransfected orvector-transfected NIH-Ras cells. AB peptides, analogously toRasGRF1WE, reduced the intracellular level of active Ras to valuessimilar to those observed in parental fibroblasts (NIH3T3). This reducedRas–GTP level correlated with a reduced level of phosphorylated
roblasts. (A) Schematic view of the molecular architecture of RasGRF1 protein. In the C-hairpin are colored in orange-yellow and red respectively. (B) Aminoacid sequence andαB (orange) connected by a convoluted loop (yellow). Red and blue asterisks above the. (C) Overview of the binding of RasGRF1 AB and HI hairpins to Ras surface. The GTPdominant negative properties to RasGRF1 full length are indicated. (D) Ras-dependentivatives, as indicated. Data are normalized over the luciferase activity in cells transfecteddent experiments run in triplicate, is shown. Double asterisks above histograms indicatewith empty vector. (For interpretation of the references to color in this figure legend, the
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011

Fig. 2. Overexpression of RasGRF1-derived AB peptides reverts the transforming phenotype of NIH-RAS mouse fibroblasts by attenuating Ras-dependent biological responses. (A) PCRproducts obtained with specific primers for RasGRF1-derived AB hairpin. The PCR was performed using as template cDNAs produced from RNA isolated from different stably-transfectedNIH-RAS cells. (B) Upper panel: total Ras level in cell lysates. Lower panel: Ras–GTP eluted from GST–RBD–glutathione–sepharose, preincubated with cell lysates, as described inSection2.2.3. Data are froma representative experiment. Central panel: quantificationof theRas–GTPamountafternormalization over total Ras. Data are normalizedover theRasGTP/totalRas ratio in cells transfected with empty vector taken equal to 100. Data shown are mean±standard deviation of two independent experiments. (C) Phospho-p44/42MAPK level in celllysates, determined byELISA assay as described in Section 2.2.4. Data are normalized on the Phospho-MAPK level in cells transfectedwith empty vector taken equal to 100. Data shown aremean±standard deviation of three independent experiments. (E) Cell staining of colony formation in soft-agar. Lowmagnification showed. The number of colonies is expressed over thenumber of colonies formedby vector-transfected cells normalized to100. (D)Morphological analysis of the different cell lines. The inset represents a colorimetric assayof acidproducedbythe same cells after 200 h of growth. Double asterisk above histograms indicates a statistical significance N99%, by Student's t test performedwith respect to cells transfected with emptyvector.
5E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
p42/p44 MAPK (Fig. 2C), i.e., the major Ras downstream pathwayinvolved in cell proliferation (Karnoub and Weinberg, 2008).
Consistent with the efficient down-regulation of the Ras pathwayin NIH-RAS cells expressing AB peptides, the cells showed morpho-logical features more similar to NIH3T3 cells than to untransfected orempty vector-transfected K-RAS transformed fibroblasts. NIH-RASfibroblasts expressing AB peptides showed a proliferative capacitysimilar to parental cells. In fact, while NIH-RAS originated confluentmultilayered cultures that strongly acidified the growthmedium after120 h (Fig. 2D insert), AB-expressing NIH-RAS cells were growtharrested by contact inhibition after reaching confluence, similar tonormal cells or the previously characterized NIH-RAS cells stablyexpressing RasGRF1WE (Bossù et al., 2000; Chiaradonna et al., 2006b).AB-expressing NIH-RAS cells are also less rifrangent than NIH-RASand, by being better adherent to the plate substrate, loose the spindle-shape typical of transformed fibroblasts (Fig. 2D).
To further characterize the biological effect of the over-expressionof AB peptides, we performed soft agar assays. Fig. 2E showsrepresentative soft-agar plates for the various cell lines with theaverage number of colonies obtained from at least three independent
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
experiments. AB peptides strongly reduced the ability of NIH-Ras cellsto form colonies in soft-agar to a level similar to parental NIH3T3 orNIH-Ras cells stably expressing RasGRF1WE.
In summary, the expression of AB peptides reverts the transformedphenotype of K-RAS expressing fibroblasts on the basis of severalindependent readouts, including intracellular Ras–GTP level, mor-phology, proliferative potential and anchorage-independent growth.The biological effects elicited by expression of the AB peptides is closeto that elicited by expression of full length RasGRF1WE, indicating thatthe AB peptide carries most, if not all, the biochemical determinantsrequired to down-regulate Ras in Ras-dependent transformedfibroblasts.
3.3. Tat-fused AB peptides are delivered into mammalian cells andattenuate Ras-signaling
Residues 47–57 of Tat (Tat47–57) from HIV virus (Dietz and Bahr,2004; Kilic et al., 2004) form a positively charged domain(YGRKKRRQRRR) that confers cell-penetrating properties to a widerange of proteins, also used in therapeutic applications [as reviewed in
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011
6 E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
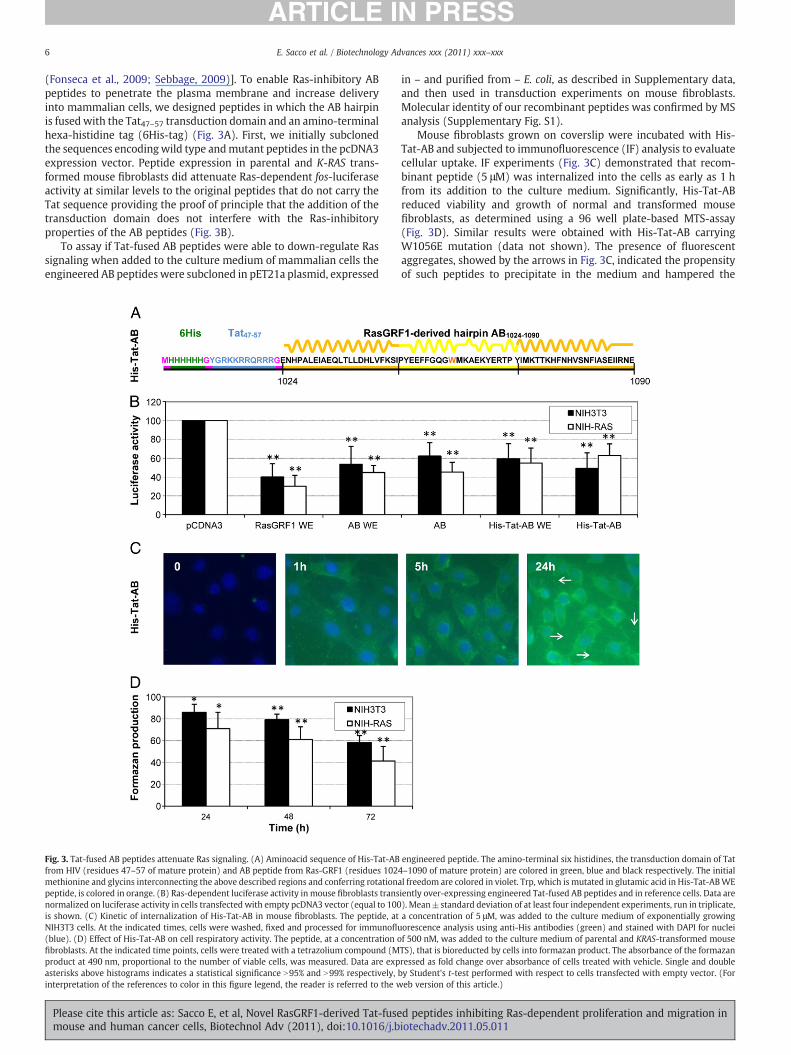
(Fonseca et al., 2009; Sebbage, 2009)]. To enable Ras-inhibitory ABpeptides to penetrate the plasma membrane and increase deliveryinto mammalian cells, we designed peptides in which the AB hairpinis fusedwith the Tat47–57 transduction domain and an amino-terminalhexa-histidine tag (6His-tag) (Fig. 3A). First, we initially subclonedthe sequences encodingwild type andmutant peptides in the pcDNA3expression vector. Peptide expression in parental and K-RAS trans-formed mouse fibroblasts did attenuate Ras-dependent fos-luciferaseactivity at similar levels to the original peptides that do not carry theTat sequence providing the proof of principle that the addition of thetransduction domain does not interfere with the Ras-inhibitoryproperties of the AB peptides (Fig. 3B).
To assay if Tat-fused AB peptides were able to down-regulate Rassignaling when added to the culture medium of mammalian cells theengineered AB peptides were subcloned in pET21a plasmid, expressed
Fig. 3. Tat-fused AB peptides attenuate Ras signaling. (A) Aminoacid sequence of His-Tat-ABfrom HIV (residues 47–57 of mature protein) and AB peptide from Ras-GRF1 (residues 102methionine and glycins interconnecting the above described regions and conferring rotationapeptide, is colored in orange. (B) Ras-dependent luciferase activity in mouse fibroblasts transnormalized on luciferase activity in cells transfected with empty pcDNA3 vector (equal to 100is shown. (C) Kinetic of internalization of His-Tat-AB in mouse fibroblasts. The peptide, atNIH3T3 cells. At the indicated times, cells were washed, fixed and processed for immunoflu(blue). (D) Effect of His-Tat-AB on cell respiratory activity. The peptide, at a concentration ofibroblasts. At the indicated time points, cells were treated with a tetrazolium compound (Mproduct at 490 nm, proportional to the number of viable cells, was measured. Data are expasterisks above histograms indicates a statistical significance N95% and N99% respectively,interpretation of the references to color in this figure legend, the reader is referred to the w
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
in – and purified from – E. coli, as described in Supplementary data,and then used in transduction experiments on mouse fibroblasts.Molecular identity of our recombinant peptides was confirmed by MSanalysis (Supplementary Fig. S1).
Mouse fibroblasts grown on coverslip were incubated with His-Tat-AB and subjected to immunofluorescence (IF) analysis to evaluatecellular uptake. IF experiments (Fig. 3C) demonstrated that recom-binant peptide (5 μM) was internalized into the cells as early as 1 hfrom its addition to the culture medium. Significantly, His-Tat-ABreduced viability and growth of normal and transformed mousefibroblasts, as determined using a 96 well plate-based MTS-assay(Fig. 3D). Similar results were obtained with His-Tat-AB carryingW1056E mutation (data not shown). The presence of fluorescentaggregates, showed by the arrows in Fig. 3C, indicated the propensityof such peptides to precipitate in the medium and hampered the
engineered peptide. The amino-terminal six histidines, the transduction domain of Tat4–1090 of mature protein) are colored in green, blue and black respectively. The initiall freedom are colored in violet. Trp, which is mutated in glutamic acid in His-Tat-ABWEiently over-expressing engineered Tat-fused AB peptides and in reference cells. Data are). Mean±standard deviation of at least four independent experiments, run in triplicate,a concentration of 5 μM, was added to the culture medium of exponentially growingorescence analysis using anti-His antibodies (green) and stained with DAPI for nucleif 500 nM, was added to the culture medium of parental and KRAS-transformed mouseTS), that is bioreducted by cells into formazan product. The absorbance of the formazanressed as fold change over absorbance of cells treated with vehicle. Single and doubleby Student's t-test performed with respect to cells transfected with empty vector. (Foreb version of this article.)
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011

7E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
possibility to use higher concentration in IF and cell growthexperiments.
3.4. Down-sizing Tat-fused AB peptides: design, chemical synthesis andin vitro Ras inhibitory activity
In order to bypass solubility problems of Tat-fused AB peptidesand/or improve their pharmacokinetic and pharmacodynamic prop-erties, we generated truncated versions lacking one or two of thethree secondary structural elements of the RasGRF1 moiety (Fig. 4A).The ability of such smaller peptides to attenuate Ras signaling wasanalyzed by fos-luciferase assay on transfected parental and K-RAStransformedmouse fibroblasts. As shown in Fig. 4B, this assay allowedus to identify the smallest AB-derived peptide that attenuates Ras-dependent luciferase activity and retains Ras-inhibitory property ofparental His-Tat-AB peptide. Such peptide, His-Tat-loopAB (1046–1066 residues of RasGRF1 mature protein), spans dominant negativemutation-susceptible Trp1056 harboring region.
By means of its small dimension, such peptide, lacking the amino-terminal 6His-tag (Fig. 5A), was produced by chemical synthesis, as aguarantee for reproducible production and purity. Such synthesizedpeptidewas soluble in water (W4502 SIGMA) up to a concentration of1 mM.
ESI-MS experiments confirmed that Tat-loopAB was able to bind toRas–GDP complex, without promoting dissociation of the Ras-boundnucleotide. In fact, the deconvoluted mass spectrum of a mixture of50 μM peptide and 10 μM Ras–GDP complex revealed peaks corre-sponding to the ternary complexes Ras–GDP/peptide, while no signals
Fig. 4. A Tat-fused down-sizedABpeptide (His-Tat-loopAB) retains Ras-inhibitory property of paHis-Tat-AB peptide (B) Ras-dependent luciferase activity in mouse fibroblasts transiently oveLuciferase activity of reference cells is also reported. Data are normalizedon the luciferase activitat least three independent experiments, each conducted in triplicate, is shown. Single and douband N99% respectively by Student's t-test, performed with respect to cells transfected with em
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
corresponding to the nucleotide-free Ras/peptide complexes werepresent (Fig. 5B–C).
We next tested the ability of Tat-loopAB to interfere in vitro withthe GEF-catalyzed dissociation of Ras-bound nucleotide and itsexchange with nucleotides present in the buffer. Dissociation of Ras-bound MantGDP (fluorescently labeled nucleotide) and exchange ofMantGDP with Ras-bound GDP were performed as described inSection 2.3 and in Lenzen et al. (1998), Palmioli et al. (2009ab) andSacco et al. (2006). Time course of representative dissociation andexchange reactions obtained in presence of 0.5 μM Tat-loopAB peptide(i.e. a 2:1 ratio with Ras) are reported in Fig. 5D and E, respectively.The Tat-loopAB peptide effectively reduced the rate of both GEF-catalyzed dissociation and exchange, likely by competing with thewild-type GEF RasGRF1 for Ras binding thereby interfering with itscatalytic activity (see also Section 4).
3.5. Chemically synthesized Tat-loopAB peptide inhibits Ras-dependentcell proliferation and migration in transformed cell lines
We then evaluated the Tat-loopAB peptide for its Ras-inhibitoryefficacy.
First, we monitored the effect of peptide on Ras-dependentproliferation of K-RAS transformed mouse fibroblasts. Cells wereplated at a density of 3000 cells/cm2 and left to adhere to the wells for18 h. Then, cells were treated with the peptide, directly added in theculture medium, and counted at 24, 48 and 72 h after treatment. Bycomparing growth curves (Fig. 6A) and morphological analysis(Fig. 6B) of cells treated with 0.5 μM Tat-loopAB with controls, not-
rental one. (A) Schematic representation of down-sizing of RasGRF1-derivedABmoiety ofr-expressing Tat-fused down-sized AB peptides, or their parental peptide (His-Tat-AB).y in cells transfectedwith empty pcDNA3, takenequal to 100.Mean±standarddeviation ofle asterisks above histograms indicate that the inhibition has a statistical significance N95%pty vector.
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011

Fig. 5. Chemically synthesized Tat-loopAB peptide binds to Ras–GDP and inhibits GEF-catalyzed Ras-bound nucleotide dissociation and exchange. (A) Aminoacid sequence ofchemically synthesized Tat-loopAB. The theoretical Molecular Weight of Tat-loopAB is 4205.79 Da. Mass deconvoluted spectra of nano-ESI-TOF analysis of (B) 5 μM Ras–GDP and(C) 10 μM Ras–GDP with 50 μM Tat-loopAB. The dashed peaks corresponds to the calculated positions of the Ras-peptide complexe without GDP. Samples were infused in 5 mMammonium acetate, pH 6.5. The experiments were performed in duplicate. Effect of Tat-loopAB on GEF-catalyzed Ras-bound nucleotide dissociation (E) and exchange (F). Thedissociation reaction onH-Raswas obtained adding to 0.25 μMRas-mantGDP preicubated for 300 s with 500 nMTat-loopAB, 200 μMGDP and 0.025 μMRasGRF1. The exchange reactionon H-Ras was obtained adding to 0.25 μM H-RasGDP, preincubated for 300 s with 500 nM Tat-loopAB, 1.25 μM mantGDP and 0.025 μM RasGRF1. Each reaction was monitored for1500 s. The graph shows representative experimental curves and their fitting to non linear ‘growth-sigmoidal Hill’ curves (thin lines). Themaximum value of relative fluorescence (100on Y axis) represents the fully charged Ras status obtained as start point in dissociation reaction and plateau of an exchange curve.
8 E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
treated cells or cells treated with vehicle buffer, the peptidesignificantly reduced cell proliferation. The peptide likely exerted acytotoxic effect, since a lot of floating cells appeared in the culturemedium of treated cells, as compared with controls. The reducedproliferative potential of cells after peptide treatment correlated withtheir reduced activation of the MAPK proteins. In fact, as shown inFig. 6C, 24 hours treatment with increasing concentrations of Tat-loopAB peptide reducedMAPK activation in a dose dependent manner.
We then determined the effect of peptides on the migration andinvasion ability of urothelial-carcinoma-derived 5637 cells, whichstrongly migrate and invade after IGF-I stimulation in a Ras andMAPK-dependent fashion (Metalli et al., 2010).
IGF-I-induced migration (black bars) and invasion (white bars) of5637 urothelial-carcinoma derived cells was severely reduced by0.5 μM Tat-loopAB peptide (Fig. 6D) indicating that Ras-targeting maycontribute to inhibit the invasive and possibly metastatic phenotypein bladder cancer.
4. Conclusions
In mammalian cells, Ras proteins control transduction pathwaysleading to proliferation, differentiation, migration and survival(Hingorani and Tuveson, 2003) and aberrant Ras signaling correlateswith proliferative and developmental disorders (Karnoub andWeinberg, 2008; Schubbert et al., 2007). Activating RAS mutationsoccur in approximately 30% of human tumors and RAS oncogenes areinvolved in both early steps and progression of tumorigenesis (Caulinet al., 2004; Chin et al., 1999; DeNicola and Tuveson, 2009; Lemoine etal., 1989; Pasca di Magliano et al., 2006; Vakiani and Solit, 2011).Hence, oncogenic Ras proteins are major clinical targets for anti-cancer therapies. Despite the development of several strategies forattenuating aberrant Ras-signaling, to date no selective and specific
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
Ras inhibitor is available for clinical use (Adjei, 2008; Vakiani andSolit, 2011). It is therefore essential to develop novel approaches andmolecules to efficiently inhibit Ras activity as a therapeutic approach.
Using as a guide the structure of the complex between nucleotide-free Ras and hSos1, a Ras activator whose Ras binding catalyticdomain shares strong homology with that of RasGRF1, and theproperties of dominant-negative RasGRF1 derivatives, Ras inhibitorypeptides have been designed. Delivery into mammalian cells has beenachieved through fusion of the peptide to the Hiv Tat penetratingpeptide. The Ras-inhibitory peptides bind the Ras–GDP complex,reduce GEF-catalyzed dissociation and exchange nucleotide on Ras invitro and reduce Ras-dependent biological responses in transformedcells, such as proliferation in K-RAS transformedmouse fibroblasts andmigration and invasion in 5637 human urothelial cancer cells.
A class of Ras inhibitors that shares a similar mode of action – i.e.,interference with GEF-catalyzed Ras-bound nucleotide dissociationand exchange in vitro, and attenuation of Ras-dependent proliferationin K-RAS transformed cells – has been characterized by our group(Airoldi et al., 2007; Palmioli et al., 2009a,b). Although these sugar-based Ras inhibitory compounds have been designed on the basis ofmolecules designed by Taveras et al. (1997) to compete with GDP forbinding to the nucleotide binding site of the Ras protein, binding toRas of both the original compound (Taveras et al., 1997) and of thesugar-based derivatives characterized by our group is not competitivewith GDP [(Palmioli et al., 2009a) and (Sacco et al., unpublishedresults)]. Despite their different structural properties, the peptidic andsugar-based inhibitors bind to similar regions on the Ras surface.Residues colored in yellow in panel B indicate Ras residues contactingthe smallest active peptide identified in this study, i.e., the loopbetween the A and B helices, while residues colored in green in panelD indicate Ras residues contacting Ras-inhibitory small sugar-derivedmolecules that span the β-3/α-2 region (Switch 2) of H-Ras–GDP and
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011

Fig. 6. Chemically synthetized Tat-loopAB peptides reduce cell proliferation andMAP kinase activation in KRAS-transformedmouse fibroblasts (NIH-RAS) andmigration and invasionof urothelial carcinoma-derived 5637 cells. (A) Proliferation assays of NIH-RAS cells not treated (circles) or grown inmedium supplemented with either vehicle (triangles) or 0.5 μMTat-loopAB (diamonds). Cells were counted after 24, 48 and 72 h. (B) Morphological analysis of NIH-RAS cells not treated or treated for 48 h with 0.5 μM Tat-loopAB or buffer.(C) Phospho-p44/42 MAPK level in cell lysates from NIH-RAS cells not treated or treated for 48 h with different concentrations of Tat-loopAB, or vehicle. MAPK activation wasassessed by ELISA assay as described in Section 2.2.4. Data are normalized on the Phospho-MAPK level in NIH-RAS cells equal to 100. Data shown are mean+standard deviation oftwo independent experiments, each performed in triplicate. Single and double asterisks above histograms indicate a statistical significance N95% and N99% respectively, by Student'st-test, over cells treated with buffer. (D) Effect of Tat-loopAB on 5637 cells' motility. Experiments of migration and invasion were performed with 0.5 μM Tat-loopAB peptide asdescribed in Section 2.2.8. Values were normalized over the cells incubated in serum-free medium (SFM) without IGF-I, equal to 100. Black and white histograms refer to migrationand invasion data respectively. The values represent the average of two independent experiments, each performed in duplicate. Double asterisks above histograms indicate that theinhibition of migration and invasion exerted by peptide has a statistical significance N99% by Student's t-test, performed with respect to cells treated with IGF1 alone.
9E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
partially overlap with binding sites for wild type GEFs (Fig. 7A, D andE), and Ras effectors, such as p120GAP, RalGDS, NORE1A andPhospholipaseCε (Rodriguez-Viciana and McCormick, 2005). Forcomparison, residues directly contacting RasGRF1 (panel A) and theHI hairpin (panel C) – that also has been shown to act as a Rasinhibitory compound (Sacco et al., 2005) – are also shown.
By 31P NMR spectroscopy two major conformational states havebeen detected in active Ras (Geyer et al., 1996; Spoerner et al., 2005,2010). These states are in dynamic equilibrium and show differentphysiological properties. State 1 has a low affinity for effectors and ispreferentially recognized by GEFs (Kalbitzer et al., 2009), while theaffinity of state 2 for effector proteins (Geyer et al., 1996, 1997;Gronwald et al., 2001; Linnemann et al., 1999; Spoerner et al., 2001,2004, 2005) is larger by a factor of approximately 20 (Spoerner et al.,2001). Recently, it has been demonstrated by isothermal titrationcalorimetry that cyclen complexes impair effector binding to activeRas by stabilization of the weak binding state (state 1) (Rosnizeck etal., 2010). Despite differences in the reported mechanism of action,Ras residues affected by cyclen complexes (i.e., Gly13, Tyr32, Ala59,Gly60, and Gln61) involve residues bound by the sugar-derivedcompounds, as well as residues bound by both AB and HI-derivedpeptides (Fig. 7 E).
The availability of different classes of compounds targeting related,but not identical, regions of Ras and acting through different
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
mechanisms, may allow to explore combinatorial treatments of Ras-dependent cancer cell lines. If these combinatorial approaches shouldprove successful, they could lead to the development of a newgeneration of “hybrid” Ras inhibitory molecules presenting multiple,complementary inhibitory mechanisms and improved selectivity.
Conflict of interest
The authors declare that they have no conflict of interest.
Acknowledgments
The authors wish to thank Alessia Vanzin and Elisa Mazzoleni forhelp in cell-based assays, Stefano Lamperti for help with figures of 3D-structures and Luca DeGioia for stimulating discussion. This work hasbeen supported by grants from Creabilis and MIUR (FAR andItalbionet) to Marco Vanoni. Andrea Morrione was supported by theBenjamin Perkins Bladder Cancer Fund and the Martin Greitzer Fund.
Appendix A. Supplementary data
Supplementary data to this article can be found online at doi:10.1016/j.biotechadv.2011.05.011.
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011
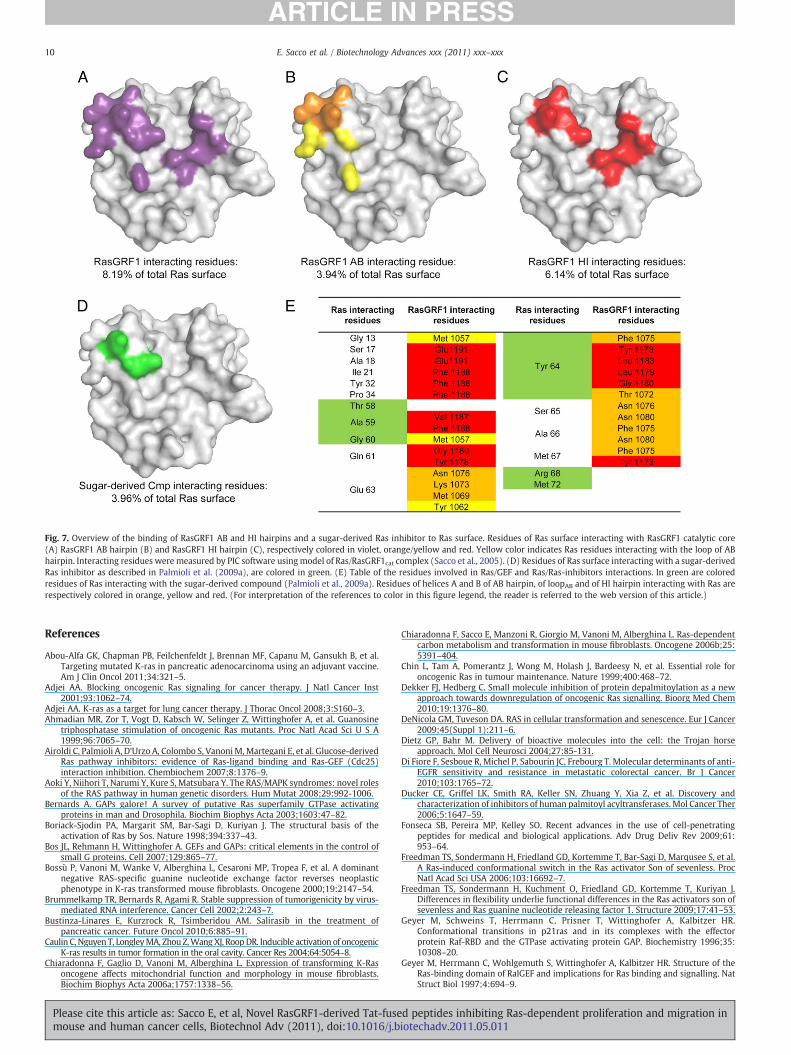
Fig. 7. Overview of the binding of RasGRF1 AB and HI hairpins and a sugar-derived Ras inhibitor to Ras surface. Residues of Ras surface interacting with RasGRF1 catalytic core(A) RasGRF1 AB hairpin (B) and RasGRF1 HI hairpin (C), respectively colored in violet, orange/yellow and red. Yellow color indicates Ras residues interacting with the loop of ABhairpin. Interacting residues were measured by PIC software usingmodel of Ras/RasGRF1cat complex (Sacco et al., 2005). (D) Residues of Ras surface interacting with a sugar-derivedRas inhibitor as described in Palmioli et al. (2009a), are colored in green. (E) Table of the residues involved in Ras/GEF and Ras/Ras-inhibitors interactions. In green are coloredresidues of Ras interacting with the sugar-derived compound (Palmioli et al., 2009a). Residues of helices A and B of AB hairpin, of loopAB and of HI hairpin interacting with Ras arerespectively colored in orange, yellow and red. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
10 E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
References
Abou-Alfa GK, Chapman PB, Feilchenfeldt J, Brennan MF, Capanu M, Gansukh B, et al.Targeting mutated K-ras in pancreatic adenocarcinoma using an adjuvant vaccine.Am J Clin Oncol 2011;34:321–5.
Adjei AA. Blocking oncogenic Ras signaling for cancer therapy. J Natl Cancer Inst2001;93:1062–74.
Adjei AA. K-ras as a target for lung cancer therapy. J Thorac Oncol 2008;3:S160–3.Ahmadian MR, Zor T, Vogt D, Kabsch W, Selinger Z, Wittinghofer A, et al. Guanosine
triphosphatase stimulation of oncogenic Ras mutants. Proc Natl Acad Sci U S A1999;96:7065–70.
Airoldi C, Palmioli A, D'Urzo A, Colombo S, Vanoni M,Martegani E, et al. Glucose-derivedRas pathway inhibitors: evidence of Ras-ligand binding and Ras-GEF (Cdc25)interaction inhibition. Chembiochem 2007;8:1376–9.
Aoki Y, Niihori T, Narumi Y, Kure S, Matsubara Y. The RAS/MAPK syndromes: novel rolesof the RAS pathway in human genetic disorders. Hum Mutat 2008;29:992-1006.
Bernards A. GAPs galore! A survey of putative Ras superfamily GTPase activatingproteins in man and Drosophila. Biochim Biophys Acta 2003;1603:47–82.
Boriack-Sjodin PA, Margarit SM, Bar-Sagi D, Kuriyan J. The structural basis of theactivation of Ras by Sos. Nature 1998;394:337–43.
Bos JL, Rehmann H, Wittinghofer A. GEFs and GAPs: critical elements in the control ofsmall G proteins. Cell 2007;129:865–77.
Bossù P, Vanoni M, Wanke V, Alberghina L, Cesaroni MP, Tropea F, et al. A dominantnegative RAS-specific guanine nucleotide exchange factor reverses neoplasticphenotype in K-ras transformed mouse fibroblasts. Oncogene 2000;19:2147–54.
Brummelkamp TR, Bernards R, Agami R. Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell 2002;2:243–7.
Bustinza-Linares E, Kurzrock R, Tsimberidou AM. Salirasib in the treatment ofpancreatic cancer. Future Oncol 2010;6:885–91.
Caulin C, Nguyen T, LongleyMA, Zhou Z,Wang XJ, RoopDR. Inducible activation of oncogenicK-ras results in tumor formation in the oral cavity. Cancer Res 2004;64:5054–8.
Chiaradonna F, Gaglio D, Vanoni M, Alberghina L. Expression of transforming K-Rasoncogene affects mitochondrial function and morphology in mouse fibroblasts.Biochim Biophys Acta 2006a;1757:1338–56.
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
Chiaradonna F, Sacco E, Manzoni R, Giorgio M, Vanoni M, Alberghina L. Ras-dependentcarbon metabolism and transformation in mouse fibroblasts. Oncogene 2006b;25:5391–404.
Chin L, Tam A, Pomerantz J, Wong M, Holash J, Bardeesy N, et al. Essential role foroncogenic Ras in tumour maintenance. Nature 1999;400:468–72.
Dekker FJ, Hedberg C. Small molecule inhibition of protein depalmitoylation as a newapproach towards downregulation of oncogenic Ras signalling. Bioorg Med Chem2010;19:1376–80.
DeNicola GM, Tuveson DA. RAS in cellular transformation and senescence. Eur J Cancer2009;45(Suppl 1):211–6.
Dietz GP, Bahr M. Delivery of bioactive molecules into the cell: the Trojan horseapproach. Mol Cell Neurosci 2004;27:85-131.
Di Fiore F, Sesboue R, Michel P, Sabourin JC, Frebourg T. Molecular determinants of anti-EGFR sensitivity and resistance in metastatic colorectal cancer. Br J Cancer2010;103:1765–72.
Ducker CE, Griffel LK, Smith RA, Keller SN, Zhuang Y, Xia Z, et al. Discovery andcharacterization of inhibitors of human palmitoyl acyltransferases. Mol Cancer Ther2006;5:1647–59.
Fonseca SB, Pereira MP, Kelley SO. Recent advances in the use of cell-penetratingpeptides for medical and biological applications. Adv Drug Deliv Rev 2009;61:953–64.
Freedman TS, Sondermann H, Friedland GD, Kortemme T, Bar-Sagi D, Marqusee S, et al.A Ras-induced conformational switch in the Ras activator Son of sevenless. ProcNatl Acad Sci USA 2006;103:16692–7.
Freedman TS, Sondermann H, Kuchment O, Friedland GD, Kortemme T, Kuriyan J.Differences in flexibility underlie functional differences in the Ras activators son ofsevenless and Ras guanine nucleotide releasing factor 1. Structure 2009;17:41–53.
Geyer M, Schweins T, Herrmann C, Prisner T, Wittinghofer A, Kalbitzer HR.Conformational transitions in p21ras and in its complexes with the effectorprotein Raf-RBD and the GTPase activating protein GAP. Biochemistry 1996;35:10308–20.
Geyer M, Herrmann C, Wohlgemuth S, Wittinghofer A, Kalbitzer HR. Structure of theRas-binding domain of RalGEF and implications for Ras binding and signalling. NatStruct Biol 1997;4:694–9.
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011

11E. Sacco et al. / Biotechnology Advances xxx (2011) xxx–xxx
Gronwald W, Huber F, Grunewald P, Sporner M, Wohlgemuth S, Herrmann C, et al.Solution structure of the Ras binding domain of the protein kinase Byr2 fromSchizosaccharomyces pombe. Structure 2001;9:1029–41.
Hingorani SR, Tuveson DA. Ras redux: rethinking how and where Ras acts. Curr OpinGenet Dev 2003;13:6-13.
Jeong S, Han SR, Lee YJ, Kim JH, Lee SW. Identification of RNA aptamer specific tomutantKRAS protein. Oligonucleotides 2010;20:155–61.
Kalbitzer HR, Spoerner M, Ganser P, Hozsa C, Kremer W. Fundamental link betweenfolding states and functional states of proteins. J Am Chem Soc 2009;131:16714–9.
Karnoub AE, Weinberg RA. Ras oncogenes: split personalities. Nat Rev Mol Cell Biol2008;9:517–31.
Kilic U, Kilic E, Dietz GP, Bahr M. The TAT protein transduction domain enhances theneuroprotective effect of glial-cell-line-derived neurotrophic factor after opticnerve transection. Neurodegener Dis 2004;1:44–9.
Laemmli UK. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 1970;227:680–5.
Le Moulec S, Loriot Y, Soria JC. Targeting KRAS pathway in NSCLC therapy. Bull Cancer2009;96(Suppl):S69–74.
Lemoine NR, Mayall ES, Wyllie FS, Williams ED, Goyns M, Stringer B, et al. Highfrequency of ras oncogene activation in all stages of human thyroid tumorigenesis.Oncogene 1989;4:159–64.
Lenzen C, Cool RH, Prinz H, Kuhlmann J,Wittinghofer A. Kinetic analysis by fluorescenceof the interaction between Ras and the catalytic domain of the guanine nucleotideexchange factor Cdc25Mm. Biochemistry 1998;37:7420–30.
Leonard DM. Ras farnesyltransferase: a new therapeutic target. J Med Chem 1997;40:2971–90.
Linnemann T, Geyer M, Jaitner BK, Block C, Kalbitzer HR, Wittinghofer A, et al.Thermodynamic and kinetic characterization of the interaction between the Rasbinding domain of AF6 and members of the Ras subfamily. J Biol Chem 1999;274:13556–62.
Lovat F, Bitto A, Xu SQ, Fassan M, Goldoni S, Metalli D, et al. Proepithelin is an autocrinegrowth factor for bladder cancer. Carcinogenesis 2009;30:861–8.
Martegani E, Vanoni M, Zippel R, Coccetti P, Brambilla R, Ferrari C, et al. Cloning byfunctional complementation of a mouse cDNA encoding a homologue of CDC25, aSaccharomyces cerevisiae RAS activator. EMBO J 1992;11:2151–7.
Mesa RA. Tipifarnib: farnesyl transferase inhibition at a crossroads. Expert RevAnticancer Ther 2006;6:313–9.
Metalli D, Lovat F, Tripodi F, Genua M, Xu SQ, Spinelli M, et al. The insulin-like growthfactor receptor I promotes motility and invasion of bladder cancer cells throughAkt- and mitogen-activated protein kinase-dependent activation of paxillin. Am JPathol 2010;176:2997–3006.
Mitin N, Rossman KL, Der CJ. Signaling interplay in Ras superfamily function. Curr Biol2005;15:R563–74.
Morgillo F, Lee HY. Lonafarnib in cancer therapy. Expert Opin Investig Drugs 2006;15:709–19.
Nazarian R, Shi H, Wang Q, Kong X, Koya RC, Lee H, et al. Melanomas acquire resistanceto B-RAF(V600E) inhibition by RTK or N-RAS upregulation. Nature 2010;468:973–7.
Palmioli A, Sacco E, Abraham S, Thomas CJ, Di Domizio A, De Gioia L, et al. Firstexperimental identification of Ras-inhibitor binding interface using awater-solubleRas ligand. Bioorg Med Chem Lett 2009a;19:4217–22.
Palmioli A, Sacco E, Airoldi C, Di Nicolantonio F, D'Urzo A, Shirasawa S, et al. Selectivecytotoxicity of a bicyclic Ras inhibitor in cancer cells expressing K-Ras(G13D).Biochem Biophys Res Commun 2009b;386:593–7.
Pamonsinlapatham P, Hadj-Slimane R, Lepelletier Y, Allain B, Toccafondi M, Garbay C,et al. P120-Ras GTPase activating protein (RasGAP): a multi-interacting protein indownstream signaling. Biochimie 2009;91:320–8.
Pasca di MaglianoM, Sekine S, Ermilov A, Ferris J, Dlugosz AA, Hebrok M. Hedgehog/Rasinteractions regulate early stages of pancreatic cancer. Genes Dev 2006;20:3161–73.
Pulciani S, Santos E, Long LK, Sorrentino V, Barbacid M. Ras gene amplification andmalignant transformation. Mol Cell Biol 1985;5:2836–41.
Quilliam LA, Rebhun JF, Castro AF. A growing family of guanine nucleotide exchangefactors is responsible for activation of Ras-family GTPases. Prog Nucleic Acid ResMol Biol 2002;71:391–444.
Rizzo S, Bronte G, Fanale D, Corsini L, Silvestris N, Santini D, et al. Prognostic vspredictive molecular biomarkers in colorectal cancer: is KRAS and BRAF wild typestatus required for anti-EGFR therapy? Cancer Treat Rev 2010;36(Suppl 3):S56–61.
Roberts PJ, Der CJ. Targeting the Raf-MEK-ERK mitogen-activated protein kinasecascade for the treatment of cancer. Oncogene 2007;26:3291–310.
Rodriguez-Viciana P, McCormick F. Characterization of interactions between ras familyGTPases and their effectors. Methods Enzymol 2005;407:187–94.
Please cite this article as: Sacco E, et al, Novel RasGRF1-derived Tat-fusemouse and human cancer cells, Biotechnol Adv (2011), doi:10.1016/j.b
Rosnizeck IC, Graf T, Spoerner M, Trankle J, Filchtinski D, Herrmann C, et al. Stabilizing aweak binding state for effectors in the human ras protein by cyclen complexes.Angew Chem Int Ed 2010;49:3830–3.
Sacco E, Fantinato S, Manzoni R, Metalli D, De Gioia L, Fantucci P, et al. The isolatedcatalytic hairpin of the Ras-specific guanine nucleotide exchange factor Cdc25Mmretains nucleotide dissociation activity but has impaired nucleotide exchangeactivity. FEBS Lett 2005;579:6851–8.
Sacco E, Metalli D, Busti S, Fantinato S, D'Urzo A,Mapelli V, et al. Catalytic competence ofthe Ras-GEF domain of hSos1 requires intra-REM domain interactions mediated byphenylalanine 577. FEBS Lett 2006;580:6322–8.
Sacco E, Abraham S, Palmioli A, Damore G, Bargna A, Mazzoleni E, et al. Bindingproperties and biological characterization of new sugar-derived Ras ligands. MedChem Commun 2011;2:396–401.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. 2nd edn.Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 1989.
Schubbert S, Shannon K, Bollag G. Hyperactive Ras in developmental disorders andcancer. Nat Rev 2007;7:295–308.
Sebbage V. Cell-penetrating peptides and their therapeutic applications. Biosci Hori2009;2:64–72.
SpoernerM, Herrmann C, Vetter IR, Kalbitzer HR,Wittinghofer A. Dynamic properties ofthe Ras switch I region and its importance for binding to effectors. Proc Natl AcadSci USA 2001;98:4944–9.
Spoerner M,Wittinghofer A, Kalbitzer HR. Perturbation of the conformational equilibriain Ras by selective mutations as studied by 31P NMR spectroscopy. FEBS Lett2004;578:305–10.
Spoerner M, Nuehs A, Ganser P, Herrmann C, Wittinghofer A, Kalbitzer HR.Conformational states of Ras complexed with the GTP analogue GppNHp orGppCH2p: implications for the interaction with effector proteins. Biochemistry2005;44:2225–36.
Spoerner M, Hozsa C, Poetzl JA, Reiss K, Ganser P, Geyer M, et al. Conformational statesof human rat sarcoma (Ras) protein complexed with its natural ligand GTP andtheir role for effector interaction and GTP hydrolysis. J Biol Chem 2010;285:39768–78.
Stathopoulos GP, Androulakis N, Souglakos J, Stathopoulos J, Georgoulias V. Presenttreatment and future expectations in advanced pancreatic cancer. Anticancer Res2008;28:1303–8.
Tanaka T, Rabbitts TH. Interfering with RAS-effector protein interactions prevent RAS-dependent tumour initiation and causes stop–start control of cancer growth.Oncogene 2010;29:6064–70.
Taveras AG, Remiszewski SW, Doll RJ, Cesarz D, Huang EC, Kirschmeier P, et al. Rasoncoprotein inhibitors: the discovery of potent, ras nucleotide exchange inhibitorsand the structural determination of a drug–protein complex. Bioorg Med Chem1997;5:125–33.
Taylor SJ, Shalloway D. Cell cycle-dependent activation of Ras. Curr Biol 1996;6:1621–7.Tentler JJ, Nallapareddy S, Tan AC, Spreafico A, Pitts TM, Morelli MP, et al. Identification
of predictive markers of response to the MEK1/2 inhibitor selumetinib (AZD6244)in K-ras-mutated colorectal cancer. Mol Cancer Ther 2010;9:3351–62.
Tidyman WE, Rauen KA. The RASopathies: developmental syndromes of Ras/MAPKpathway dysregulation. Curr Opin Genet Dev 2009;19:230–6.
Vakiani E, Solit DB. KRAS and BRAF: drug targets and predictive biomarkers. J Pathol2011;223:219–29.
Vanoni M, Bertini R, Sacco E, Fontanella L, Rieppi M, Colombo S, et al. Characterizationand properties of dominant-negative mutants of the ras-specific guaninenucleotide exchange factor CDC25(Mm). J Biol Chem 1999;274:36656–62.
Vetter IR, Wittinghofer A. The guanine nucleotide-binding switch in three dimensions.Science 2001;294:1299–304.
Waldmann H, Karaguni IM, Carpintero M, Gourzoulidou E, Herrmann C, Brockmann C,et al. Sulindac-derived Ras pathway inhibitors target the Ras–Raf interaction anddownstream effectors in the Ras pathway. Angew Chem Int Ed 2004;43:454–8.
Wittinghofer A, Waldmann H. Ras — a molecular switch involved in tumor formation.Angew Chem Int Ed 2000;39:4192–214.
Wong KK. Recent developments in anti-cancer agents targeting the Ras/Raf/MEK/ERKpathway. Recent Pat Anticancer Drug Discov 2009;4:28–35.
Zenker M. Genetic and pathogenetic aspects of Noonan syndrome and relateddisorders. Horm Res 2009;72(Suppl 2):57–63.
Zhang Z, Jiang G, Yang F, Wang J. Knockdown of mutant K-ras expression byadenovirus-mediated siRNA inhibits the in vitro and in vivo growth of lung cancercells. Cancer Biol Ther 2006;5:1481–6.
Zheng Y, Quilliam LA. Activation of the Ras superfamily of small GTPases. Workshop onexchange factors. EMBO Rep 2003;4:463–8.
d peptides inhibiting Ras-dependent proliferation and migration iniotechadv.2011.05.011
Related Documents






![Bulletin 194R Fused and Non-Fused Disconnects...11 Bulletin 194R Fused and Non-Fused Disconnects Product Selection 4th Pole Modules Rated Current [A] Maximum Hp Ratings Fuse Dim. Ref.](https://static.cupdf.com/doc/110x72/5e3f37a7aa25636cac719ea2/bulletin-194r-fused-and-non-fused-disconnects-11-bulletin-194r-fused-and-non-fused.jpg)




