Gene expression pattern Notch pathway genes are expressed in mammalian ovarian follicles Joshua Johnson a , Tamara Espinoza a , Robert W. McGaughey a,b , Alan Rawls a,b , Jeanne Wilson-Rawls a,b, * a Department of Biology, Arizona State University, Tempe, AZ 85287-1501, USA b Graduate Program in Molecular and Cellular Biology, Arizona State University, Tempe, AZ 85287-1501, USA Received 9 March 2001; received in revised form 26 July 2001; accepted 13 August 2001 Abstract Folliculogenesis is the process of development of ovarian follicles that ultimately results in the release of fertilizable oocytes at ovulation. This is a complex program that involves the proliferation and differentiation of granulosa cells. Granulosa cells are necessary for follicle growth and support the oocyte during folliculogenesis. Genes that regulate the proliferation and differentiation of granulosa cells are beginning to be elucidated. In this study, the expression patterns of Notch receptor genes and their ligands, which have been shown to regulate cell-fate decisions in many systems during development, were examined in the mammalian ovary. In situ hybridization data showed that Notch2, Notch3, and Jagged2 were expressed in an overlapping pattern in the granulosa cells of developing follicles. Jagged1 was expressed in oocytes exclusively. Downstream target genes of Notch also were expressed in granulosa cells. These data implicate the Notch signaling pathway in the regulation of mammalian folliculogenesis. q 2001 Elsevier Science Ireland Ltd. All rights reserved. Keywords: Notch; Jagged; Ovary; Follicle; Granulosa 1. Results and discussion During the reproductive lifespan of mammals there is continuous follicle development within the ovary. Follicu- logenesis involves the proliferation of granulosa cells into a multilaminar epithelium and the development of a fluid filled antrum. Granulosa cells differentiate into two cell types, mural granulosa cells that lie close to the basal lamina and extend to the antrum, and cumulus granulosa cells that surround the oocyte. The growth and maturation of the oocyte is tightly coordinated with the proliferation and differentiation of granulosa cells (Gosden et al., 1997; Drummond and Findlay, 1999; Rodgers et al., 1999; Erick- son and Shimasaki, 2000). The Notch genes encode highly conserved transmem- brane receptors (Kimble and Simpson 1997; Lewis, 1998; Artavanis-Tsakonis et al., 1999) that control cell-fate deci- sions between adjacent cells that may or may not be equiva- lent. Notch was initially identified in Drosophila (Wharton et al., 1985), and homologs have been found in C. elegans, and vertebrates (Notch1-4) (Weinmaster et al., 1991, 1992; Franco del Amo et al., 1992; Lardelli and Lendahl, 1993; Lardelli et al., 1994; Uyttendaele et al., 1996). There are two families of Notch ligands that are also transmembrane proteins. In vertebrates, they are Deltalike-1, -3, and -4 (Dll-1, 3, 4) (Bettenhausen et al., 1995; Dunwoodie et al., 1997; Shutter et al., 2000), and Jagged1 and -2 (Lindsell et al., 1995; Shawber et al., 1996). As both Notch and its ligands are transmembrane proteins, signaling only occurs between closely apposed cells. Activation of Notch by ligand binding triggers clea- vage of the receptor, releasing the intracellular domain, which then translocates to the nucleus (Schroeter et al., 1998; Kopan et al., 1996; Blaumueller et al., 1997; Struhl and Adachi, 1998). In the nucleus, Notch forms transcrip- tional complexes with transcription factors of the CSL family (Weinmaster, 1997; Greenwald, 1998). The expres- sion of Notch1-4 in the ovary was determined by in situ hybridization of ovaries from 6 week old mice using gene-specific digoxygenin-labeled antisense RNA probes. Gene expression was analyzed using the follicle staging scheme of Pedersen and Peters (1968). Notch2 transcripts were initially detected in type 3b follicles (Fig. 1A,B) and this gene was expressed up to type 8. There was a loss of Notch2 expression in the cells that are coincident with the sites of initial antral pocket formation and in periantral gran- ulosa cells (Fig. 2A, black arrows). Oocytes did not express Notch2 (Fig. 1B). Notch3 was expressed in an overlapping Mechanisms of Development 109 (2001) 355–361 0925-4773/01/$ - see front matter q 2001 Elsevier Science Ireland Ltd. All rights reserved. PII: S0925-4773(01)00523-8 www.elsevier.com/locate/modo * Corresponding author. Department of Biology, Box 871501, Tempe, AZ 85287-1501, USA. Tel.: 11-480-727-6675; fax: 11-480-965-2519. E-mail address: [email protected] (J. Wilson-Rawls).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Gene expression pattern
Notch pathway genes are expressed in mammalian ovarian follicles
Joshua Johnsona, Tamara Espinozaa, Robert W. McGaugheya,b,Alan Rawlsa,b, Jeanne Wilson-Rawlsa,b,*
aDepartment of Biology, Arizona State University, Tempe, AZ 85287-1501, USAbGraduate Program in Molecular and Cellular Biology, Arizona State University, Tempe, AZ 85287-1501, USA
Received 9 March 2001; received in revised form 26 July 2001; accepted 13 August 2001
Abstract
Folliculogenesis is the process of development of ovarian follicles that ultimately results in the release of fertilizable oocytes at ovulation.
This is a complex program that involves the proliferation and differentiation of granulosa cells. Granulosa cells are necessary for follicle
growth and support the oocyte during folliculogenesis. Genes that regulate the proliferation and differentiation of granulosa cells are
beginning to be elucidated. In this study, the expression patterns of Notch receptor genes and their ligands, which have been shown to
regulate cell-fate decisions in many systems during development, were examined in the mammalian ovary. In situ hybridization data showed
that Notch2, Notch3, and Jagged2 were expressed in an overlapping pattern in the granulosa cells of developing follicles. Jagged1 was
expressed in oocytes exclusively. Downstream target genes of Notch also were expressed in granulosa cells. These data implicate the Notch
signaling pathway in the regulation of mammalian folliculogenesis. q 2001 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Notch; Jagged; Ovary; Follicle; Granulosa
1. Results and discussion
During the reproductive lifespan of mammals there is
continuous follicle development within the ovary. Follicu-
logenesis involves the proliferation of granulosa cells into a
multilaminar epithelium and the development of a fluid
filled antrum. Granulosa cells differentiate into two cell
types, mural granulosa cells that lie close to the basal lamina
and extend to the antrum, and cumulus granulosa cells that
surround the oocyte. The growth and maturation of the
oocyte is tightly coordinated with the proliferation and
differentiation of granulosa cells (Gosden et al., 1997;
Drummond and Findlay, 1999; Rodgers et al., 1999; Erick-
son and Shimasaki, 2000).
The Notch genes encode highly conserved transmem-
brane receptors (Kimble and Simpson 1997; Lewis, 1998;
Artavanis-Tsakonis et al., 1999) that control cell-fate deci-
sions between adjacent cells that may or may not be equiva-
lent. Notch was initially identified in Drosophila (Wharton
et al., 1985), and homologs have been found in C. elegans,
and vertebrates (Notch1-4) (Weinmaster et al., 1991, 1992;
Franco del Amo et al., 1992; Lardelli and Lendahl, 1993;
Lardelli et al., 1994; Uyttendaele et al., 1996). There are two
families of Notch ligands that are also transmembrane
proteins. In vertebrates, they are Deltalike-1, -3, and -4
(Dll-1, 3, 4) (Bettenhausen et al., 1995; Dunwoodie et al.,
1997; Shutter et al., 2000), and Jagged1 and -2 (Lindsell et
al., 1995; Shawber et al., 1996).
As both Notch and its ligands are transmembrane
proteins, signaling only occurs between closely apposed
cells. Activation of Notch by ligand binding triggers clea-
vage of the receptor, releasing the intracellular domain,
which then translocates to the nucleus (Schroeter et al.,
1998; Kopan et al., 1996; Blaumueller et al., 1997; Struhl
and Adachi, 1998). In the nucleus, Notch forms transcrip-
tional complexes with transcription factors of the CSL
family (Weinmaster, 1997; Greenwald, 1998). The expres-
sion of Notch1-4 in the ovary was determined by in situ
hybridization of ovaries from 6 week old mice using
gene-specific digoxygenin-labeled antisense RNA probes.
Gene expression was analyzed using the follicle staging
scheme of Pedersen and Peters (1968). Notch2 transcripts
were initially detected in type 3b follicles (Fig. 1A,B) and
this gene was expressed up to type 8. There was a loss of
Notch2 expression in the cells that are coincident with the
sites of initial antral pocket formation and in periantral gran-
ulosa cells (Fig. 2A, black arrows). Oocytes did not express
Notch2 (Fig. 1B). Notch3 was expressed in an overlapping
Mechanisms of Development 109 (2001) 355–361
0925-4773/01/$ - see front matter q 2001 Elsevier Science Ireland Ltd. All rights reserved.
PII: S0925-4773(01)00523-8
www.elsevier.com/locate/modo
* Corresponding author. Department of Biology, Box 871501, Tempe,
AZ 85287-1501, USA. Tel.: 11-480-727-6675; fax: 11-480-965-2519.
E-mail address: [email protected] (J. Wilson-Rawls).
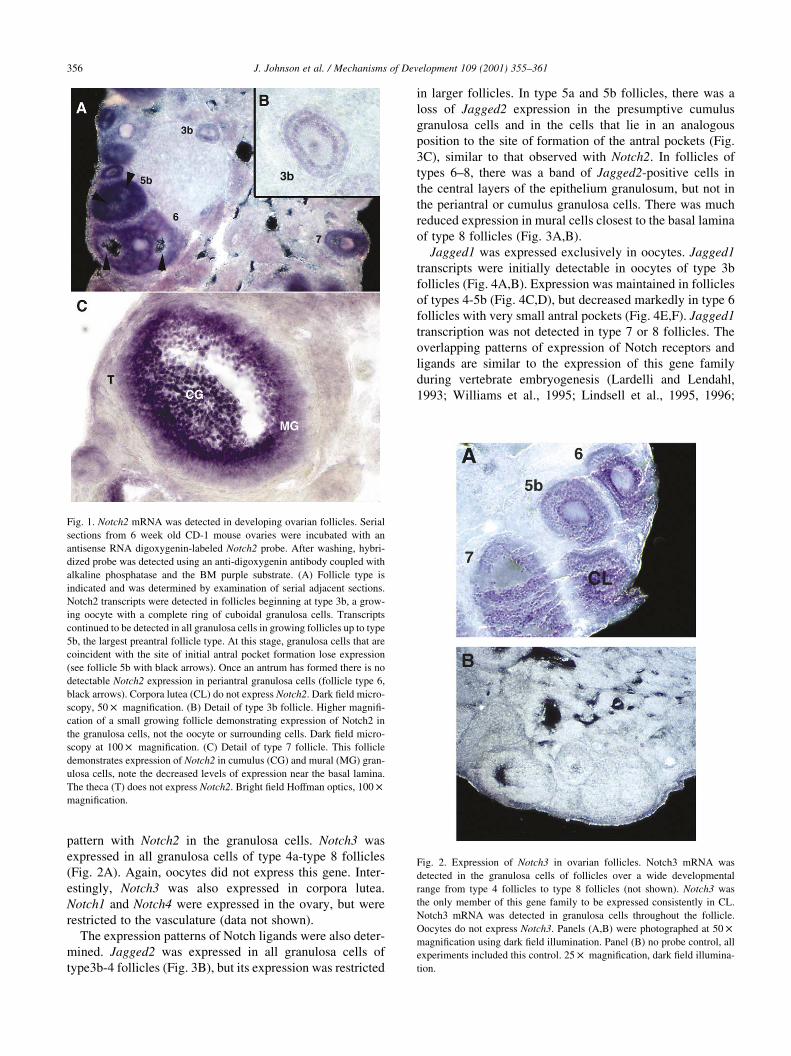
pattern with Notch2 in the granulosa cells. Notch3 was
expressed in all granulosa cells of type 4a-type 8 follicles
(Fig. 2A). Again, oocytes did not express this gene. Inter-
estingly, Notch3 was also expressed in corpora lutea.
Notch1 and Notch4 were expressed in the ovary, but were
restricted to the vasculature (data not shown).
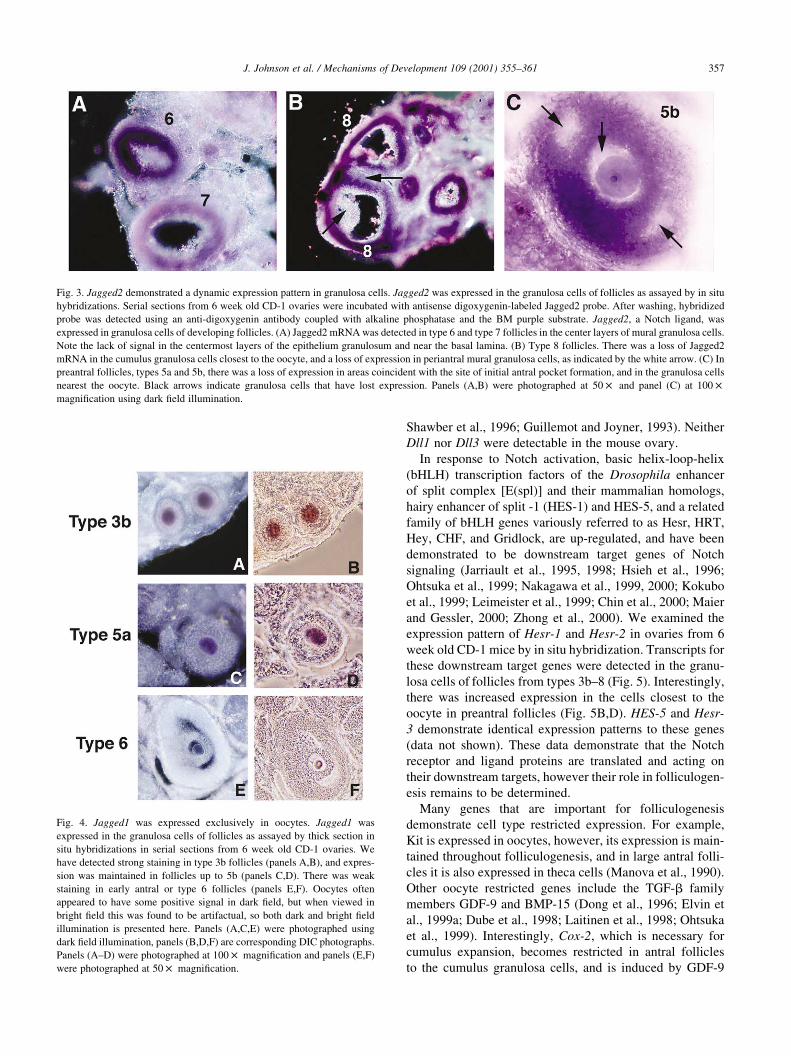
The expression patterns of Notch ligands were also deter-
mined. Jagged2 was expressed in all granulosa cells of
type3b-4 follicles (Fig. 3B), but its expression was restricted
in larger follicles. In type 5a and 5b follicles, there was a
loss of Jagged2 expression in the presumptive cumulus
granulosa cells and in the cells that lie in an analogous
position to the site of formation of the antral pockets (Fig.
3C), similar to that observed with Notch2. In follicles of
types 6–8, there was a band of Jagged2-positive cells in
the central layers of the epithelium granulosum, but not in
the periantral or cumulus granulosa cells. There was much
reduced expression in mural cells closest to the basal lamina
of type 8 follicles (Fig. 3A,B).
Jagged1 was expressed exclusively in oocytes. Jagged1
transcripts were initially detectable in oocytes of type 3b
follicles (Fig. 4A,B). Expression was maintained in follicles
of types 4-5b (Fig. 4C,D), but decreased markedly in type 6
follicles with very small antral pockets (Fig. 4E,F). Jagged1
transcription was not detected in type 7 or 8 follicles. The
overlapping patterns of expression of Notch receptors and
ligands are similar to the expression of this gene family
during vertebrate embryogenesis (Lardelli and Lendahl,
1993; Williams et al., 1995; Lindsell et al., 1995, 1996;
J. Johnson et al. / Mechanisms of Development 109 (2001) 355–361356
Fig. 1. Notch2 mRNA was detected in developing ovarian follicles. Serial
sections from 6 week old CD-1 mouse ovaries were incubated with an
antisense RNA digoxygenin-labeled Notch2 probe. After washing, hybri-
dized probe was detected using an anti-digoxygenin antibody coupled with
alkaline phosphatase and the BM purple substrate. (A) Follicle type is
indicated and was determined by examination of serial adjacent sections.
Notch2 transcripts were detected in follicles beginning at type 3b, a grow-
ing oocyte with a complete ring of cuboidal granulosa cells. Transcripts
continued to be detected in all granulosa cells in growing follicles up to type
5b, the largest preantral follicle type. At this stage, granulosa cells that are
coincident with the site of initial antral pocket formation lose expression
(see follicle 5b with black arrows). Once an antrum has formed there is no
detectable Notch2 expression in periantral granulosa cells (follicle type 6,
black arrows). Corpora lutea (CL) do not express Notch2. Dark field micro-
scopy, 50 £ magnification. (B) Detail of type 3b follicle. Higher magnifi-
cation of a small growing follicle demonstrating expression of Notch2 in
the granulosa cells, not the oocyte or surrounding cells. Dark field micro-
scopy at 100 £ magnification. (C) Detail of type 7 follicle. This follicle
demonstrates expression of Notch2 in cumulus (CG) and mural (MG) gran-
ulosa cells, note the decreased levels of expression near the basal lamina.
The theca (T) does not express Notch2. Bright field Hoffman optics, 100 £
magnification.
Fig. 2. Expression of Notch3 in ovarian follicles. Notch3 mRNA was
detected in the granulosa cells of follicles over a wide developmental
range from type 4 follicles to type 8 follicles (not shown). Notch3 was
the only member of this gene family to be expressed consistently in CL.
Notch3 mRNA was detected in granulosa cells throughout the follicle.
Oocytes do not express Notch3. Panels (A,B) were photographed at 50 £
magnification using dark field illumination. Panel (B) no probe control, all
experiments included this control. 25 £ magnification, dark field illumina-
tion.
Shawber et al., 1996; Guillemot and Joyner, 1993). Neither
Dll1 nor Dll3 were detectable in the mouse ovary.
In response to Notch activation, basic helix-loop-helix
(bHLH) transcription factors of the Drosophila enhancer
of split complex [E(spl)] and their mammalian homologs,
hairy enhancer of split -1 (HES-1) and HES-5, and a related
family of bHLH genes variously referred to as Hesr, HRT,
Hey, CHF, and Gridlock, are up-regulated, and have been
demonstrated to be downstream target genes of Notch
signaling (Jarriault et al., 1995, 1998; Hsieh et al., 1996;
Ohtsuka et al., 1999; Nakagawa et al., 1999, 2000; Kokubo
et al., 1999; Leimeister et al., 1999; Chin et al., 2000; Maier
and Gessler, 2000; Zhong et al., 2000). We examined the
expression pattern of Hesr-1 and Hesr-2 in ovaries from 6
week old CD-1 mice by in situ hybridization. Transcripts for
these downstream target genes were detected in the granu-
losa cells of follicles from types 3b–8 (Fig. 5). Interestingly,
there was increased expression in the cells closest to the
oocyte in preantral follicles (Fig. 5B,D). HES-5 and Hesr-
3 demonstrate identical expression patterns to these genes
(data not shown). These data demonstrate that the Notch
receptor and ligand proteins are translated and acting on
their downstream targets, however their role in folliculogen-
esis remains to be determined.
Many genes that are important for folliculogenesis
demonstrate cell type restricted expression. For example,
Kit is expressed in oocytes, however, its expression is main-
tained throughout folliculogenesis, and in large antral folli-
cles it is also expressed in theca cells (Manova et al., 1990).
Other oocyte restricted genes include the TGF-b family
members GDF-9 and BMP-15 (Dong et al., 1996; Elvin et
al., 1999a; Dube et al., 1998; Laitinen et al., 1998; Ohtsuka
et al., 1999). Interestingly, Cox-2, which is necessary for
cumulus expansion, becomes restricted in antral follicles
to the cumulus granulosa cells, and is induced by GDF-9
J. Johnson et al. / Mechanisms of Development 109 (2001) 355–361 357
Fig. 3. Jagged2 demonstrated a dynamic expression pattern in granulosa cells. Jagged2 was expressed in the granulosa cells of follicles as assayed by in situ
hybridizations. Serial sections from 6 week old CD-1 ovaries were incubated with antisense digoxygenin-labeled Jagged2 probe. After washing, hybridized
probe was detected using an anti-digoxygenin antibody coupled with alkaline phosphatase and the BM purple substrate. Jagged2, a Notch ligand, was
expressed in granulosa cells of developing follicles. (A) Jagged2 mRNA was detected in type 6 and type 7 follicles in the center layers of mural granulosa cells.
Note the lack of signal in the centermost layers of the epithelium granulosum and near the basal lamina. (B) Type 8 follicles. There was a loss of Jagged2
mRNA in the cumulus granulosa cells closest to the oocyte, and a loss of expression in periantral mural granulosa cells, as indicated by the white arrow. (C) In
preantral follicles, types 5a and 5b, there was a loss of expression in areas coincident with the site of initial antral pocket formation, and in the granulosa cells
nearest the oocyte. Black arrows indicate granulosa cells that have lost expression. Panels (A,B) were photographed at 50 £ and panel (C) at 100 £
magnification using dark field illumination.
Fig. 4. Jagged1 was expressed exclusively in oocytes. Jagged1 was
expressed in the granulosa cells of follicles as assayed by thick section in
situ hybridizations in serial sections from 6 week old CD-1 ovaries. We
have detected strong staining in type 3b follicles (panels A,B), and expres-
sion was maintained in follicles up to 5b (panels C,D). There was weak
staining in early antral or type 6 follicles (panels E,F). Oocytes often
appeared to have some positive signal in dark field, but when viewed in
bright field this was found to be artifactual, so both dark and bright field
illumination is presented here. Panels (A,C,E) were photographed using
dark field illumination, panels (B,D,F) are corresponding DIC photographs.
Panels (A–D) were photographed at 100 £ magnification and panels (E,F)
were photographed at 50 £ magnification.
(Sirois et al., 1992; Sirois, 1995; Elvin et al., 1999b). In the
ovary, kit ligand is expressed only by granulosa cells, and it
becomes restricted to the mural population in antral follicles
(Manova et al., 1993; Joyce et al., 1999). While there are
genes whose expression is restricted to the oocyte or specific
populations of granulosa cells, there are none that have
identical expression patterns to that of Jagged1 or Jagged2.
Nor are there any reports of genes that demonstrate a loss of
expression in areas coincident with the sites of formation of
the antral pockets, as observed for Notch2 and Jagged2.
Atresia in ovarian follicles is marked by the apoptotic
death of the granulosa cells (Hsu and Hsueh, 1998;
McGee et al., 1998). Notch signaling inhibits apoptosis in
hematopoietic cells (Varnum-Finney et al., 1998; Jones et
al., 1998; Deftos et al., 1998; Jehn et al., 1999; Shelly et al.,
1999; Han et al., 2000). Further, a Notch2 mutant that is
signaling defective demonstrated increased apoptosis
during embryogenesis (Hamada et al., 1999). We reasoned
that it was likely that the Notch genes were expressed in
growing, not atretic follicles. Since, treatment with exogen-
ous gonadotropin (PMSG) will induce the proliferation and
differentiation of granulosa cells and increase the number of
developing follicles by rescuing them from atresia, Notch2
expression was compared with TUNEL and BrdU labeling
of ovaries. Mice were given PMSG intraperitoneally and
ovaries were harvested 6 h post-treatment. Serial ovary
sections were alternately placed on slides for in situ hybri-
dization to detect Notch2, or for TUNEL labeling to detect
apoptotic cells. As a comparison, mice were injected with
BrdU to label proliferating cells 1 h before the ovaries were
harvested.
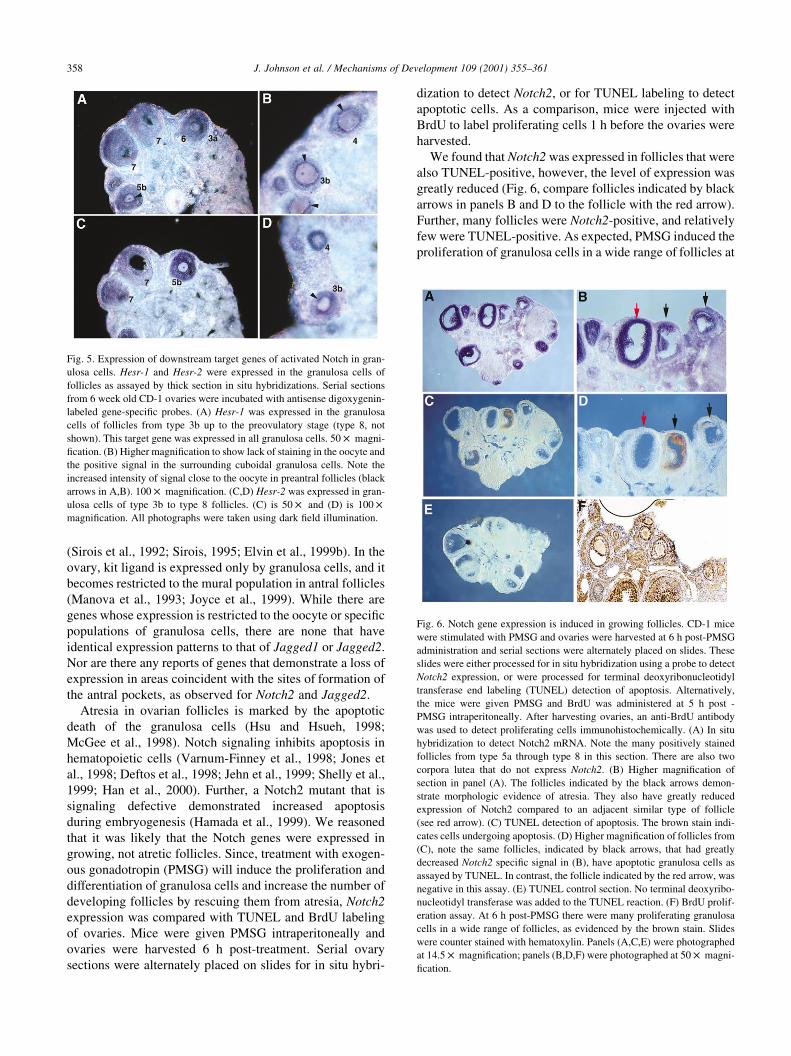
We found that Notch2 was expressed in follicles that were
also TUNEL-positive, however, the level of expression was
greatly reduced (Fig. 6, compare follicles indicated by black
arrows in panels B and D to the follicle with the red arrow).
Further, many follicles were Notch2-positive, and relatively
few were TUNEL-positive. As expected, PMSG induced the
proliferation of granulosa cells in a wide range of follicles at
J. Johnson et al. / Mechanisms of Development 109 (2001) 355–361358
Fig. 5. Expression of downstream target genes of activated Notch in gran-
ulosa cells. Hesr-1 and Hesr-2 were expressed in the granulosa cells of
follicles as assayed by thick section in situ hybridizations. Serial sections
from 6 week old CD-1 ovaries were incubated with antisense digoxygenin-
labeled gene-specific probes. (A) Hesr-1 was expressed in the granulosa
cells of follicles from type 3b up to the preovulatory stage (type 8, not
shown). This target gene was expressed in all granulosa cells. 50 £ magni-
fication. (B) Higher magnification to show lack of staining in the oocyte and
the positive signal in the surrounding cuboidal granulosa cells. Note the
increased intensity of signal close to the oocyte in preantral follicles (black
arrows in A,B). 100 £ magnification. (C,D) Hesr-2 was expressed in gran-
ulosa cells of type 3b to type 8 follicles. (C) is 50 £ and (D) is 100 £
magnification. All photographs were taken using dark field illumination.
Fig. 6. Notch gene expression is induced in growing follicles. CD-1 mice
were stimulated with PMSG and ovaries were harvested at 6 h post-PMSG
administration and serial sections were alternately placed on slides. These
slides were either processed for in situ hybridization using a probe to detect
Notch2 expression, or were processed for terminal deoxyribonucleotidyl
transferase end labeling (TUNEL) detection of apoptosis. Alternatively,
the mice were given PMSG and BrdU was administered at 5 h post -
PMSG intraperitoneally. After harvesting ovaries, an anti-BrdU antibody
was used to detect proliferating cells immunohistochemically. (A) In situ
hybridization to detect Notch2 mRNA. Note the many positively stained
follicles from type 5a through type 8 in this section. There are also two
corpora lutea that do not express Notch2. (B) Higher magnification of
section in panel (A). The follicles indicated by the black arrows demon-
strate morphologic evidence of atresia. They also have greatly reduced
expression of Notch2 compared to an adjacent similar type of follicle
(see red arrow). (C) TUNEL detection of apoptosis. The brown stain indi-
cates cells undergoing apoptosis. (D) Higher magnification of follicles from
(C), note the same follicles, indicated by black arrows, that had greatly
decreased Notch2 specific signal in (B), have apoptotic granulosa cells as
assayed by TUNEL. In contrast, the follicle indicated by the red arrow, was
negative in this assay. (E) TUNEL control section. No terminal deoxyribo-
nucleotidyl transferase was added to the TUNEL reaction. (F) BrdU prolif-
eration assay. At 6 h post-PMSG there were many proliferating granulosa
cells in a wide range of follicles, as evidenced by the brown stain. Slides
were counter stained with hematoxylin. Panels (A,C,E) were photographed
at 14.5 £ magnification; panels (B,D,F) were photographed at 50 £ magni-
fication.

this same timepoint (Fig. 6F). Since, many follicles were
Notch-positive and few were TUNEL-positive, it is likely
that Notch2 expression is upregulated in developing, not
atretic follicles. However, a role in the induction of apop-
tosis cannot be ruled out.
Our data demonstrate that Notch family genes are
expressed in mammalian ovarian follicles. The Notch signal-
ing pathway has been repeatedly demonstrated to play a
crucial developmental role in tissues where it is expressed,
therefore, we propose that Notch signaling is implicated in
regulating folliculogenesis. Perturbation of this pathway in
developing follicles will be necessary to provide insight into
its functional role(s) during folliculogenesis.
2. Materials and methods
2.1. Animal use and exogenous hormone treatment
CD-1 mice were used for all studies and were 6 weeks old
(42 days) unless otherwise stated. For all in situ hybridiza-
tion experiments, ovaries were immediately explanted into
ice-cold PBS, then processed for individual experiments as
described below. For animals stimulated with pregnant
mare’s serum gonadotropin (PMSG) female mice were
injected intraperitoneally with 5 IU of PMSG (Sigma
Chemical Co., St. Louis, MO) and ovaries were collected
6 h post hormone treatment.
2.2. Staging of follicles
Follicles were staged using the scheme of Pedersen and
Peters (1968). This scheme sets out eight types of follicles
based on the size of the oocyte and the number of granulosa
cells in the widest cross-section. Type 1 is a small oocyte with
no attached follicle cells. Type 3a is a small oocyte with a
complete ring of cells, but not more than 20 cells are present
in the largest cross-section. Type 3b is a growing oocyte with
21–60 follicle cells, the largest preantral follicle is type 5b,
this stage has a fully grown oocyte (70 mm diameter) and up
to 4000 follicle cells. Early antral follicles are types 6 and 7,
and Graafian follicles, are type 8. All follicles were staged
from serial adjacent sections using this scheme.
2.3. Thick section in situ hybridization
Sections for in situ hybridization were prepared after Xu
and Wilkinson (1995). Ovaries were fixed overnight in 4%
paraformaldehyde in phosphate buffered saline (PBS) at
48C, then were embedded in paraffin after stepwise dehydra-
tion in ethanol and Hemo-DE tissue clearing agent (Fisher
Biotech, Pittsburgh, PA). Twenty to thirty micron (mm)
sections were cut perpendicular to the axis of entry of the
ovarian blood vessels. Sections were then dewaxed, rehy-
drated, and in situ hybridizations were done as described in
Wilson-Rawls et al. (1999). The sections were hybridized
with antisense digoxygenin-11-UTP-labeled (Roche Mole-
cular Biochemicals, Indianapolis, IN) RNA probes specific
to each of the Notch signaling pathway genes, at 628C.
Detection of the hybridized probe was done using an alka-
line-phosphatase conjugated anti-digoxygenin antibody to
recognize hybridized probe after washing (Roche Molecular
Biochemicals, Indianapolis, IN). The color reaction used to
visualize gene expression was done using the BM purple
alkaline phosphatase substrate (Roche Molecular Biochem-
icals, Indianapolis, IN). Control samples were prepared to
determine non-specific background labeling by omitting
probe from the hybridization buffer. Further controls
included testing sense probes on ovary sections and all anti-
sense probes were tested on embryo controls for specificity.
2.4. Terminal deoxynucleotidyl transferase-mediated
dUTP-fluorescein end labeling (TUNEL) detection of
apoptosis
TUNEL detection of apoptosis in ovary sections was
performed as per Li et al. (1998), with the following modi-
fications: 10 nM fluorescein-labeled UTP (Roche Molecular
Biochemicals, Indianapolis, IN) was used in the end-label-
ing reaction mixture, and horseradish-peroxidase-conju-
gated anti-fluorescein antibody (Roche Molecular
Biochemicals, Indianapolis, IN) diluted 1:100 in the digox-
ygenin blocking reagent was used to visualize. Negative
control samples were prepared by omitting TdT enzyme
(Promega Corp., Madison, WI) from the labeling reactions.
2.5. BrdU labeling of ovaries
Mice were injected with PMSG as described above. Thirty
minutes before the collection of the ovaries a suspension of
50 mg BrdU and 10 mg FUDR/kg body weight, was injected
intraperitoneally. Ovaries were collected, fixed and paraffin-
embedded as described above, then sectioned to 7 mm.
Detection of BrdU/FUDR incorporation was performed
using a streptavidin-biotin BrdU detection kit (ZYMED,
South San Francisco, CA) per the manufacturer’s instruc-
tions, using the optional heat-induced epitope retrieval
step. Color reactions were done using the horseradish perox-
idase substrate, DAB, resulting in a dark brown stain in
proliferating cells. The slides were counterstained with
hematoxylin.
Acknowledgements
We would like to thank Drs R.L. Johnson and H. Kokubo
for Hesr gene probes. We would like to thank Drs B.
Vanderhyden and P. Hunt for helpful discussions. We
would also like to acknowledge imaging support from the
W.M. Keck Bioimaging and Life Sciences Visualization
laboratories at ASU. A.R. is supported by grants from the
Muscular Dystrophy Association and NSF. J.W.R. is
supported by NIH, T.E. is supported by a MARC program
grant to ASU from the NIH.
J. Johnson et al. / Mechanisms of Development 109 (2001) 355–361 359

References
Artavanis-Tsakonis, S., Rand, M.D., Lake, R.J., 1999. Notch Signaling:
Cell fate control and signal integration in development. Science 284,
770–776.
Bettenhausen, B., Hrabe de Angelis, M., Guenet, J.L., Gosler, A., 1995.
Transient and restricted expression during mouse embryogenesis of
Dll1, a murine gene closely related to Drosophila Delta. Development
21, 2407–2418.
Blaumueller, C.M., Qi, H., Zagouras, P., Artavanis-Tsakonis, S., 1997.
Intracellular cleavage of Notch leads to a heterodimeric receptor on
the plasma membrane. Cell 90, 281–291.
Chin, M.T., Maemura, K., Fukumoto, S., Jain, M.K., Layne, M.D., Wata-
nabe, M., Hsieh, C.M., Lee, M.E., 2000. Cardiovascular basic helix
loop helix factor 1, a novel transcriptional repressor expressed prefer-
entially in the developing and adult cardiovascular system. J. Biol.
Chem. 275, 6381–6387.
Deftos, M., He, Y.W., Ojala, E.W., Bevan, M.J., 1998. Correlating notch
signaling with thymocyte maturation. Immunity 9, 777–786.
Dong, J.D., Albertini, F., Nishimori, K., Kumar, T.R., Lu, N., Matzuk,
M.M., 1996. Growth differentiation factor-9 is required during early
ovarian folliculogenesis. Nature 383, 531–535.
Drummond, A.E., Findlay, J.K., 1999. The role of estrogen in folliculogen-
esis. Mol. Cell Endocrinol. 151, 57–64.
Dube, J.L., Wang, P., Elvin, J., Lyons, K.M., Celeste, A.J., Matzuk, M.M.,
1998. The bone morphogenetic protein 15 gene is X-linked and
expressed in oocytes. Mol. Endocrinol. 12, 1809–1817.
Dunwoodie, S.L., Henrique, D., Harrison, S.M., Beddington, R.S., 1997.
Mouse Dll3: a novel divergent Delta gene which may complement the
function of other Delta homologues during early pattern formation on
the mouse embryo. Development 16, 3065–3076.
Elvin, J.A., Yan, C., Wang, P., Nishimori, K., Matzuk, M.M., 1999a. Mole-
cular characterization of the follicle defects in the growth differentiation
factor 9-deficient ovary. Mol. Endocrinol. 13, 1018–1034.
Elvin, J.A., Clark, A.T., Wang, P., Wolfman, N.M., Matzuk, M.M., 1999b.
Paracrine actions of growth factor-9 in the mammalian ovary. Mol.
Endocrinol. 13, 1035–1048.
Erickson, G.F., Shimasaki, S., 2000. The role of the oocyte in folliculogen-
esis. Trends Endocrinol. Metab. 11, 193–198.
Franco del Amo, F., Smith, D.E., Swiatek, P.J., Gendron-Maguire, M.,
Greenspan, R.J., McMahon, A.P., Gridley, T., 1992. Expression pattern
of Motch, a mouse homolog of Drosophila Notch, suggests an important
role in early postimplantation mouse development. Development 115,
737–744.
Gosden, R., Krapez, J., Briggs, D., 1997. Growth and development of the
mammalian oocyte. Bioessays 19, 875–882.
Greenwald, I., 1998. Lin-12/Notch signaling: lessons from worms and flies.
Genes Dev. 12, 1751–1762.
Guillemot, F., Joyner, A.L., 1993. Dynamic expression of the murine
Achaete-Scute homologue Mash-1 in the developing nervous system.
Mech. Dev. 42, 171–185.
Hamada, Y., Kadokawa, Y., Okabe, M., Ikawa, M., Coleman, J.R., Tsuji-
moto, Y., 1999. Mutation in ankyrin repeats of the mouse Notch2 gene
induces early embryonic lethality. Development 126, 3415–3424.
Han, W., Ye, Q., Moore, M.A., 2000. A soluble form of human Delta-like -
1 inhibits differentiation of hematopoietic progenitor cells. Blood 95,
1616–1625.
Hsieh, J.J.-D., Henkel, T., Salmon, P., Robey, E., Peterson, M.G., Hayward,
S.D., 1996. Truncated mammalian Notch1 activates CBF1/RBP-Jk-
repressed genes by a mechanism resembling that of Epstein-Barr
virus EBNA2. Mol. Cell Biol. 16, 952–959.
Hsu, S.Y., Hsueh, A.J., 1998. Intracellular mechanisms of ovarian cell
apoptosis. Mol. Cell Endocrinol. 145, 21–25.
Jarriault, S., Brou, C., Logeat, F., Schroeter, E.H., Kopan, R., Israel, A.,
1995. Signalling downstrean of activated mammalian Notch. Nature
377, 355–358.
Jarriault, S., Le Bail, O., Hirsinger, E., Pourquie, O., Logeat, F., Strong,
C.F., Brou, C., Seidah, N.G., Israel, A., 1998. Delta-1 activation of
notch-1 signaling results in HES-1 transactivation. Mol. Cell Biol. 18,
7423–7431.
Jehn, B.M., Bilke, W., Pear, W.S., Osborne, B.A., 1999. Protective effects
of notch-1 on TCR-induced apoptosis. J. Immunol. 162, 635–638.
Jones, P., May, G., Healy, L., Brown, J., Hoyne, G., Delassus, S., Enver, T.,
1998. Stromal expression of Jagged 1 promotes colony formation by
fetal hematopoietic progenitor cells. Blood 92, 1505–1511.
Joyce, I.M., Pendola, F.L., Wigglesworth, K., Eppig, J.J., 1999. Oocyte
regulation of kit ligand expression in mouse ovarian follicles. Dev.
Biol. 214, 342–353.
Kimble, J., Simpson, P., 1997. The lin-12/Notch signaling pathway and its
regulation. Annu. Rev. Cell. Dev. Biol. 13, 333–361.
Kokubo, H., Lun, Y., Johnson, R.L., 1999. Identification and expression of
a novel family of bHLH cDNAs related to Drosophila Hairy and Enhan-
cer of Split. Biochem. Biophys. Res. Commun. 260, 459–465.
Kopan, R., Schroeter, E.H., Weintraub, H., Nye, J.S., 1996. Signal trans-
duction by activated mNotch: importance of proteolytic processing and
its regulation by the extracellular domain. Proc. Natl. Acad. Sci. USA
93, 1683–1688.
Laitinen, M., Vuojolainen, K., Jaatinen, R., Ketola, I., Aaltonen, J., Lehto-
nen, E., Heikinheimo, M., Ritvos, O., 1998. A novel growth differentia-
tion factor-9 (GDF-9) related factor is co-expressed with GDF-9 in
mouse oocytes during folliculogenesis. Mech. Dev. 78, 135–140.
Lardelli, M., Lendahl, U., 1993. MotchA and MotchB-Two mouse Notch
homologues coexpressed in a wide variety of tissues. Exp. Cell Res.
204, 364–372.
Lardelli, M., Dahlstrand, J., Lendahl, U., 1994. The novel Notch homologue
mouse Notch3 lacks specific epidermal growth factor-repeats and is
expressed in proliferating neuroepithelium. Mech. Dev. 46, 123–136.
Leimeister, C., Externbrink, A., Klamt, B., Gessler, M., 1999. Hey genes: a
novel subfamily of hairy- and Enhancer of split related genes specifi-
cally expressed during mouse embryogenesis. Mech. Dev. 85, 173–177.
Lewis, J., 1998. Notch signalling and the control of cell fate choices in
vertebrates. Semin. Cell Dev. Biol. 9, 584–589.
Li, J., Kim, J.M., Li, M., Miyazaki, T., Mackenzie, A., Korneluk, R.G.,
Tsang, B.K., 1998. Expression of inhibitor of apoptosis proteins (IAPs)
in rat granulosa cells during ovarian follicular development and atresia.
Endocrinology 139, 1321–1328.
Lindsell, C.E., Shawber, C.J., Boulter, J., Weinmaster, G., 1995. Jagged: a
mammalian ligand that activates Notch1. Cell 80, 909–917.
Lindsell, C.E., Boulter, J., diSibio, G., Gossler, A., Weinmaster, G., 1996.
Expression patterns of Jagged, Delta1, Notch1, Notch2, and Notch3
genes identify ligand-receptor pairs that may function in neural devel-
opment. Mol. Cell. Neurosci. 8, 14–27.
Manova, K., Nocka, K., Besmer, P., Bachvarova, R.F., 1990. Gonadal
expression of c-kit encoded at the W locus of the mouse. Development
110, 1057–1069.
Manova, K., Huang, E.J., Angeles, M., De Leon, V., Sanchez, S., Prono-
vost, M., Besmer, P., Bachvarova, R.F., 1993. The expression pattern of
the c-kit ligand in gonads of mice supports a role for the c-kit receptor in
oocyte growth and in proliferation of spermatogonia. Dev. Biol. 57, 85–
99.
Maier, M.M., Gessler, M., 2000. Comparative analysis of the human and
mouse Hey1 promoter: Hey genes are new Notch target genes.
Biochem. Biophys. Res. Commun. 275, 652–660.
McGee, E.A., Hsu, S.Y., Kaipia, A., Hsueh, A.J., 1998. Cell death and
survival during ovarian follicle development. Mol. Cell Endocrinol.
140, 15–18.
Nakagawa, O., Nakagawa, M., Richardson, J.A., Olson, E.N., Srivasatava,
D., 1999. HRT1, HRT2, and HRT3: a new subclass of bHLH transcrip-
tion factors marking specific cardiac, somitic, and pharyngeal arch
segments. Dev. Biol. 26, 72–84.
Nakagawa, O., McFadden, D.G., Nakagawa, M., Yanagisawa, H., Hu, T.,
Srivastava, D., Olson, E.N., 2000. Members of the HRT family of basic
J. Johnson et al. / Mechanisms of Development 109 (2001) 355–361360

helix-loop-helix proteins act as transcriptional repressors downstream
of Notch signaling. Proc. Natl. Acad. Sci. USA 25, 13655–13660.
Ohtsuka, T., Ishibiashi, M., Gradwohl, F., Nakanishi, S., Guillemot, F.,
Kageyama, R., 1999. Hes1 and Hes5 as Notch effectors in mammalian
neuronal differentiation. EMBO J. 18, 2196–2207.
Pedersen, T., Peters, H., 1968. Proposal for a classification of oocytes and
follicles in the mouse ovary. J. Reprod. Fert. 17, 555–557.
Rodgers, R.J., Lavranos, T.C., van Wezel, H.F., Irving-Rodgers, H.F.,
1999. Development of the ovarian follicular epithelium. Mol. Cell
Endocrinol. 151, 170–171.
Schroeter, E.H., Kisslinger, J.A., Kopan, R., 1998. Notch-1 signaling
requires ligand-induced proteolytic release of intracellular domain.
Nature 393, 382–386.
Shawber, C., Boulter, J., Lindsell, C.E., Weinmaster, G., 1996. Jagged2: a
serrate-like gene expressed during rat embryogenesis. Dev. Biol. 180,
370–376.
Shelly, L., Fuchs, C., Miele, L., 1999. Notch-1 inhibits apoptosis in murine
erythroleukemia cells and is necessary for differentiation induced by
hybrid polar compounds. J. Cell. Biochem. 73, 164–175.
Shutter, J.R., Scully, S., Fan, W., Richards, W.G., Kitajewski, J.,
Deblandre, G.A., Kitner, C.R., Stark, K.L., 2000. DII4, a novel Notch
ligand expressed in arterial endothelium. Genes Dev. 14, 1313–1318.
Sirois, J., Simmons, D.L., Richards, J.S., 1992. Hormonal regulation of
messenger ribonucleic acid encoding a novel isoform of prostaglandin
endoperoxide H synthase in rat preovulatory follicles. J. Biol. Chem.
267, 11586–11592.
Sirois, J., 1995. Induction of prostaglandin endoperoxide H synthase-2 by
human chorionic gonadotropin in bovine preovulatory follicles in vivo.
Endocrinology 135, 406–409.
Struhl, G., Adachi, A., 1998. Nuclear access and action of notch in vivo.
Cell 93, 649–660.
Uyttendaele, H., Marazzi, G., Wu, G., Yan, Q., Sassoon, D., Kitajewski, J.,
1996. Notch4/int-3, a mammary proto-oncogene, is an endothelial cell-
specific mammalian Notch gene. Development 122, 2251–2259.
Varnum-Finney, B., Purton, L.E., Yu, M., Brashem-Stein, C., Flowers, D.,
Staats, S., Moore, K.A., Le Roux, I., Mann, R., Gray, G., Artavanis-
Tsakonis, S., 1998. The Notch ligand, Jagged1, influences the devel-
opment of primitive hematopoietic precursor cells. Blood 91, 4084–
4091.
Wharton, K.A., Johansen, K.M., Xu, T., Artavanis-Tsakonis, S., 1985.
Nucleotide sequence from the neurogenic locus Notch implies a gene
product that shares homology with proteins containing EGF-like
repeats. Cell 43, 567–581.
Weinmaster, G., 1997. The ins and outs of notch signaling. Mol. Cell.
Neurosci. 9, 91–102.
Weinmaster, G., Roberts, V.J., Lemke, G., 1991. A homolog of Drosophila
Notch expressed during mammalian development. Development 113,
199–205.
Weinmaster, G., Roberts, V.J., Lemke, G., 1992. Notch2: a second mamma-
lian Notch gene. Development 116, 931–941.
Williams, R., Lendahl, U., Lardelli, M., 1995. Complementary and combi-
natorial patterns of Notch gene family expression during early mouse
development. Mech. Dev. 53, 357–368.
Wilson-Rawls, J., Hurt, C.R., Parsons, S.M., Rawls, A., 1999. Differential
regulation of epaxial and hypaxial muscle development by paraxis.
Development 126, 5217–5229.
Xu, Q., Wilkinson, D., 1995. Whole-mount in situ hybridization of thick
tissue sections. Zebrafish Sci. Monitor 3 (6), 5.
Zhong, T.P., Rosenber, M., Mohideen, M.A., Weinstein, B., Fishman,
M.C., 2000. Gridlock, an HLH gene required for assembly of the
aorta in zebrafish. Science 287, 1820–1824.
J. Johnson et al. / Mechanisms of Development 109 (2001) 355–361 361
Related Documents











