NMDA receptor activation modulates programmed cell death during early post-natal retinal development: a BDNF-dependent mechanism Rodrigo A. P. Martins, Mariana S. Silveira, Marco R. Curado, Angela I. Police and Rafael Linden Laboratorio de Neurogenese, Instituto de Biofı ´sica Carlos Chagas Filho, UFRJ, Rio de Janeiro, Brazil Abstract Glutamate is a classical excitotoxin of the central nervous system (CNS), but extensive work demonstrates neuropro- tective roles of this neurotransmitter in developing CNS. Mechanisms of glutamate-mediated neuroprotection are still under scrutiny. In this study, we investigated mediators of glutamate-induced neuroprotection, and tested whether this neurotransmitter controls programmed cell death in the developing retina. The protective effect of N-methyl-D-aspar- tate (NMDA) upon differentiating cells of retinal explants was completely blocked by a neutralizing antibody to brain-derived neurotrophic factor (BDNF), but not by an antibody to neurot- rophin-4 (NT-4). Consistently, chronic activation of NMDA receptor increased the expression of BDNF and trkB mRNA, as well as BDNF protein content, but did not change the con- tent of NT-4 mRNA in retinal tissue. Furthermore, we showed that in vivo inactivation of NMDA receptor by intraperitoneal injections of MK-801 increased natural cell death of specific cell populations of the post-natal retina. Our results show that chronic activation of NMDA receptors in vitro induces a BDNF- dependent neuroprotective state in differentiating retinal cells, and that NMDA receptor activation controls programmed cell death of developing retinal neurons in vivo. Keywords: amacrine cells, development, excitotoxicity, glu- tamate, neurotrophins, programmed cell death. J. Neurochem. (2005) 95, 244–253. The development of the central nervous system (CNS) is regulated by both cell-intrinsic and cell-extrinsic factors (Cepko 1999; Dyer and Cepko 2001; Pallas 2001; Coskun and Luskin 2002; Zhang et al. 2002; Ferguson and Slack 2003; Job and Tan 2003; Marquardt 2003). Various classes of extracellular molecules, such as growth factors, neuro- peptides and neurotransmitters, have been implicated in the extrinsic control of CNS development (Cameron et al. 1998; Nguyen et al. 2001; Linden et al. 2005). Besides its classical role as a mediator of synaptic transmission, the excitatory neurotransmitter glutamate has important roles during development of the CNS, such as the control of cell proliferation, as well as neuronal migration and survival (Komuro and Rakic 1993; LoTurco et al. 1995; Haydar et al. 2000). Although it has been reported that both activation of glutamate receptors and neuronal activity regulate cell survival during development of the CNS, the mechanisms of these effects are only partially understood. It has been shown that the ability of glutamate to induce cell survival may depend on the modulation of the responsiveness of neuronal cells to trophic factors. As an example, survival of cerebellar granule neurons depends on the activation of N-methyl-D-aspartate (NMDA) receptors in vitro and in vivo and, at least in vitro, it was shown that this protective effect is mediated through the regulation of the expression of brain- derived neurotrophic factor (BDNF) (Marini et al. 1998; Bhave et al. 1999; Monti and Contestabile 2000). BDNF, similar to other neurotrophins, has a well-defined ability to promote neuronal survival during development, through binding to the high-affinity tyrosine kinase receptor TrkB, and activation of intracellular signalling pathways that either regulate gene expression or directly modulate the cell death Received January 22, 2005; revised manuscript received May 25, 2005; accepted June 2, 2005. Address correspondence and reprint requests to Rodrigo A. P. Martins, CCS, Sala G2-019, Cidade Universita ´ria, Rio de Janeiro, RJ, 21949– 900, Brazil. E-mail: [email protected] Abbreviations used: BDNF, brain-derived neurotrophic factor; CNS, central nervous system; GCL, ganglion cells layer; iINL, immature inner nuclear layer; IPL, inner plexiform layer; NBL, neuroblastic layer; NMDA, N-methyl-D-aspartate; NT-4, neurotrophin-4; PCD, programmed cell death; VACC, voltage-activated calcium channels. Journal of Neurochemistry , 2005, 95, 244–253 doi:10.1111/j.1471-4159.2005.03360.x 244 ȑ 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

NMDA receptor activation modulates programmed cell deathduring early post-natal retinal development: a BDNF-dependentmechanism
Rodrigo A. P. Martins, Mariana S. Silveira, Marco R. Curado, Angela I. Police and Rafael Linden
Laboratorio de Neurogenese, Instituto de Biofısica Carlos Chagas Filho, UFRJ, Rio de Janeiro, Brazil
Abstract
Glutamate is a classical excitotoxin of the central nervous
system (CNS), but extensive work demonstrates neuropro-
tective roles of this neurotransmitter in developing CNS.
Mechanisms of glutamate-mediated neuroprotection are still
under scrutiny. In this study, we investigated mediators of
glutamate-induced neuroprotection, and tested whether this
neurotransmitter controls programmed cell death in the
developing retina. The protective effect of N-methyl-D-aspar-
tate (NMDA) upon differentiating cells of retinal explants was
completely blocked by a neutralizing antibody to brain-derived
neurotrophic factor (BDNF), but not by an antibody to neurot-
rophin-4 (NT-4). Consistently, chronic activation of NMDA
receptor increased the expression of BDNF and trkB mRNA,
as well as BDNF protein content, but did not change the con-
tent of NT-4 mRNA in retinal tissue. Furthermore, we showed
that in vivo inactivation of NMDA receptor by intraperitoneal
injections of MK-801 increased natural cell death of specific
cell populations of the post-natal retina. Our results show that
chronic activation of NMDA receptors in vitro induces a BDNF-
dependent neuroprotective state in differentiating retinal cells,
and that NMDA receptor activation controls programmed cell
death of developing retinal neurons in vivo.
Keywords: amacrine cells, development, excitotoxicity, glu-
tamate, neurotrophins, programmed cell death.
J. Neurochem. (2005) 95, 244–253.
The development of the central nervous system (CNS) isregulated by both cell-intrinsic and cell-extrinsic factors(Cepko 1999; Dyer and Cepko 2001; Pallas 2001; Coskunand Luskin 2002; Zhang et al. 2002; Ferguson and Slack2003; Job and Tan 2003; Marquardt 2003). Various classesof extracellular molecules, such as growth factors, neuro-peptides and neurotransmitters, have been implicated in theextrinsic control of CNS development (Cameron et al. 1998;Nguyen et al. 2001; Linden et al. 2005). Besides itsclassical role as a mediator of synaptic transmission, theexcitatory neurotransmitter glutamate has important rolesduring development of the CNS, such as the control of cellproliferation, as well as neuronal migration and survival(Komuro and Rakic 1993; LoTurco et al. 1995; Haydaret al. 2000).
Although it has been reported that both activation ofglutamate receptors and neuronal activity regulate cellsurvival during development of the CNS, the mechanismsof these effects are only partially understood. It has beenshown that the ability of glutamate to induce cell survivalmay depend on the modulation of the responsiveness ofneuronal cells to trophic factors. As an example, survival
of cerebellar granule neurons depends on the activation ofN-methyl-D-aspartate (NMDA) receptors in vitro and in vivoand, at least in vitro, it was shown that this protective effect ismediated through the regulation of the expression of brain-derived neurotrophic factor (BDNF) (Marini et al. 1998;Bhave et al. 1999; Monti and Contestabile 2000). BDNF,similar to other neurotrophins, has a well-defined ability topromote neuronal survival during development, throughbinding to the high-affinity tyrosine kinase receptor TrkB,and activation of intracellular signalling pathways that eitherregulate gene expression or directly modulate the cell death
Received January 22, 2005; revised manuscript received May 25, 2005;accepted June 2, 2005.Address correspondence and reprint requests to Rodrigo A. P. Martins,
CCS, Sala G2-019, Cidade Universitaria, Rio de Janeiro, RJ, 21949–900, Brazil. E-mail: [email protected] used: BDNF, brain-derived neurotrophic factor; CNS,
central nervous system; GCL, ganglion cells layer; iINL, immature innernuclear layer; IPL, inner plexiform layer; NBL, neuroblastic layer;NMDA, N-methyl-D-aspartate; NT-4, neurotrophin-4; PCD, programmedcell death; VACC, voltage-activated calcium channels.
Journal of Neurochemistry, 2005, 95, 244–253 doi:10.1111/j.1471-4159.2005.03360.x
244 � 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253

machinery. (for review see Kaplan and Miller 2000; Huangand Reichardt 2003).
The control of BDNF expression was also defined as a keyevent for the maintenance of neuronal survival in other cellpopulations of the developing CNS. Depolarization ofembryonic cortical neurons in vitro leads to calcium influxthrough voltage-activated calcium channels (VACC), activa-tion of the CREB transcription factor, and an increase in theexpression of BDNF (Shieh et al. 1998; Tao et al. 1998).Other studies indicated that the expression of another TrkBligand, NT-4, is also up-regulated by activation of glutama-tergic receptors. Systemic injection of kainate, a glutamat-ergic agonist, increases the expression of NT-4 in the spinalcord (Scarisbrick et al. 1999).
Enhancement of neuronal survival by increased neuronalactivity may, however, occur through alternative mechanisms.Using an in vitro model of isolated retinal ganglion cells,Meyer-Franke et al. (1998) reported that depolarizationrapidly induces the translocation of TrkB receptors fromintracellular stores to the plasma membrane. A similarmechanism was described in cultures of embryonic hippo-campal neurons. High frequency tetanic stimulation of thosecells resulted in an increase in the surface expression of TrkBreceptor, depending on the activation of NMDA receptors andcalcium influx (Du et al. 2000). Conversely, it was shown that,in cultured cortical cells, the increase in trophic factorresponsiveness after depolarization depends on the regulationof trkBmRNAand protein expression (Kingsbury et al. 2003).
In the mature retina, glutamate is the neurotransmitter usedin the radial pathway, which mediates signalling fromphotoreceptors to the ganglion cells (Massey and Redburn1987; Barnstable 1993; Thorenson and Witkovsky 1999).Although glutamate receptors, as well as both vesicular anduptake glutamate transporters, are expressed at early stagesof rodent retinal development (Pow and Barnett 1999;Grunder et al. 2000a,b; Johansson et al. 2000; Johnson et al.2003; Sucher et al. 2003), the roles of this neurotransmitterduring retinal development are not completely understood.Previous studies showed that BDNF, NT-4 and the TrkBreceptor are also expressed during early development of theretina (Perez and Caminos 1995; Rickman and Brecha 1995;Cellerino and Kohler 1997; Nakazawa et al. 2002). It isreasonable to suggest that, as observed in other CNS areas,the expression of these neurotrophins or their receptors mayalso be regulated by glutamate during retinal development.
In a previous study, we showed that glutamate is a survivalfactor for both differentiating and proliferative cells in thedeveloping retina. It was also shown that this protectiveeffect of glutamate against induced cell death was mediatedby NMDA receptors and depended on TrkB receptoractivation (Rocha et al. 1999). Therefore, the neurotrophindependence for the glutamate protective effect may resultfrom NMDA-induced regulation of either one or both of theTrkB ligands (BDNF or NT-4), or up-regulation of the TrkB
receptor itself. One goal of this work was to distinguishamong these possibilities. In addition, it was reported that theNMDA receptor is a physiological regulator of naturallyoccurring cell death in several areas of the developing CNS(Brenneman et al. 1990; Gould et al. 1994; Ikonomidouet al. 1999; Monti and Contestabile 2000; Fiske and Brunjes2001; Adams et al. 2004). We therefore also tested whetherthe activity of NMDA receptors regulates naturally occurringcell death in the developing retina in vivo.
Experimental procedures
Materials
Hepes, NMDA were from Sigma (St Louis, MO, USA), basal
medium of Eagle (BME), dNTP mix, fetal calf serum (FCS), Taq
polymerase, Trizol reagent were from Invitrogen (Carlsbad, CA,
USA), Rnase-free Dnase I was from Ambion (Austin, TX, USA),
BDNF antibody was from Promega (Madison, WI, USA), NT-4
antibody was from R & D Systems (Minneapolis, MN, USA), NT-4
was provided by Regeneron Pharmaceuticals (New York, NY, USA).
Tissue culture
All experimental procedures with animals were approved by the
Committee on Animal Experimentation of the Institute of
Biophysics Carlos Chagas Filho, based on the currently accepted
international rules. Lister hooded rats were used in this study.
Retinal explants were prepared as previously described (de Araujo
and Linden 1993). Briefly, rat pups were killed instantaneously by
decapitation, their eyes were removed and the retinas dissected and
cut into fragments of approximately 1 mm2. Retinal explants were
cultured in BME containing 5% FCS, 20 mM HEPES (pH 7.4) in
Erlenmeyer flasks kept in an orbital shaker at 37�C. For RT-PCR or
ELISA, the explants were cultured for 3, 6 or 18 h in the presence or
absence of NMDA. For the experiments with excitotoxic cell death,
retinal explants were exposed to 6 mM glutamate for 6 h following
18 h of pretreatment with the indicated compounds.
In vivo experiments
Rat pups at post-natal day 6 (P6) were injected intraperitonealy with
either saline or the non-competitive NMDA receptor antagonist,
MK-801 (1 mg/kg of body weight). After 24 h, the animals were
killed, and their retinae were histologically processed for quantifi-
cation of cell death.
Histology
Retinal tissue was fixed with 4% paraformaldehyde in sodium
phosphate buffer pH 7.4, and cryoprotected in 20% sucrose. Then,
explants were oriented under a dissecting microscope in an
aluminum chamber filled with OCT embedding medium and
10 lM transverse sections were cut in a cryostat as previously
described (Rehen et al. 1996). Sections were mounted and stained
with neutral red.
Quantification of cell death and imaging
Dead cells were identified by their condensed and deeply stained
chromatin among normal neighbouring cells in retinal sections
NMDA and programmed cell death in developing retina 245
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253

(Rehen et al. 1996; Rocha et al. 1999). At post-natal day 3 (P3), the
outer stratum of the rat retina contains differentiating amacrine cells
close to the inner plexiform layer (IPL), corresponding to the inner
portion of the inner nuclear layer (iINL), as well as the neuroblastic
layer (NBL), which contains proliferating and undifferentiated post-
mitotic cells (see Fig. 1a, Linden et al. 1999). At that developmental
stage, more than 80% of excitotoxic-sensitive cells are differentiating
cells localized close to the IPL and are found scattered among round
cells, which are both typical criteria to identify differentiating
amacrine cells of the post-natal retina. (Weidman and Kuwabara
1968; Rocha et al. 1999). For these reasons, excitotoxic-induced celldeath experiments were done at P3. To quantify induced cell death,
four randomly selected explants were analyzed for each group in each
experiment. Counts of pyknotic profiles were made at 1000 · mag-
nification in three random fields of 0.0148 mm2 within the outer
stratum of each explants. For quantification of cell death in the in vivoexperiments, histological procedures were the same as above, and
both eyes of each animal were totally sectioned. All sections
representative of the central third of both retinas of each animal were
analyzed, to avoid variation because of the central-peripheryc
gradient of retinal development. Inner INL (iINL) or outer (oINL)
were defined as the inner and outer halves of the INL, respectively (see
Fig. 6). Images were captured with an Axiocam HR digital camera
(Zeiss), mounted on an Axiophot microscope.
Immunohistochemistry
Retinal sections were fixed in paraformaldehyde [4% in phosphate-
buffered saline (PBS)], washed, and treated with hydrogen peroxide
(3% in PBS) before incubation in blocking solution (1% bovine
serum albumin in PBS). The sections were then treated with primary
antibodies anti-TrkB (1 : 500, no. AB5372, Chemicon, Temecula,
CA, USA) or anti-BDNF (1 : 100, #AB1534, Chemicon, USA),
washed with PBS and incubated with a biotin-conjugated secondary
antibody (1 : 500, goat anti-rabbit IgG; Vector Laboratories,
Burlingame, CA, USA). After secondary antibody binding, sections
were treated with an avidin–biotin–peroxidase complex (Vectastain
ABC, Vector Laboratories) followed by Cy-3 tyramide detection
(Perkin Elmer, Boston, MA, USA) according to the manufacturers’
instructions. For nuclear staining, sections were incubated with
DAPI (Sigma). Images were captured with an Axiocam HR digital
camera (Zeiss), mounted on an Axiophot microscope.
Semiquantitative RT-PCR
Retinal explants exposed to NMDA (60–100 lM) for various time
periods were washed with cold sodium phosphate buffer pH 7.4,
and total RNA was extracted using Trizol reagent following the
instructions of the manufacturer. Then, RNA was treated with
Rnase-free Dnase I for 10 min at 37�C. RNA was then precipitated
with 3 M sodium acetate/ethanol, centrifuged, washed with 70%
ethanol and ressuspended in DEPC-treated water. One microgram of
RNA was reverse transcribed using a First Strand cDNA synthesis
kit (Amersham Pharmacia Biotech, Piscataway, NJ, USA). cDNA
was amplified in a 50-lL PCR reaction, containing 5 lL of cDNA,
0.2 mM dNTPs, 1.5 U of Taq polymerase, 1 mM MgCl2 and 1 lM of
each primer. After 5 min at 94�C, all amplifications were performed
for 1 min at 94�C, 1 min at 58�C and 1 min at 72�C, followed by a
final extension of 10 min at 72�C. The total number of cycles for
each PCR was determined based on saturation curves of each
reaction and, in all cases, it was below the saturation of the
reactions. Specific primer sequences, number of cycles and product
size (in bp) were as follows: Glyceraldehyde-3-phosphate dehy-
drogenase (GAPDH) 5¢-CGTCTTCACCACCATGGAGA-3¢(forward) and 5¢-CGGCCATCACGCCACAGTTT-3¢ (reverse),
30 cycles, 310; BDNF 5¢-AAACATGTCTATGAGGGTTCGGC-3¢(forward) and 5¢-GCCACTATCTTCCCCTTTTAATGG-3¢ (re-
verse), 30 cycles, 343; NT-4 5¢-CGTCAGTACTTCTTCGA-GACGCGCTGC-3¢ (forward) and 5¢-GCAGACGCAAGCGGTGT-CGAT-3¢ (reverse), 35 cycles, 191 and trkB 5¢-TGCTGTGGTGGT-GATTGCCTCTGTG-3¢ (forward) and 5¢-GTTCTCTCCTACCAA-GCAGTTCCGG-3¢ (reverse), 27 cycles, 746. The latter primers
amplified all trkB receptor isoforms. Aliquots of PCR reactions were
eletrophoresed, visualized using ethidium bromide and photo-
graphed under UV illumination using a Kodak DC 120 digital
camera. Densitometry was done using 1D gel analysis software from
Kodak, and data for each time point were expressed as the ratio
between the BDNF, trkB or NT-4 band and the (GAPDH) internal
control band.
ELISA
BDNF protein was quantified in retinal tissue extracts, using
a quantitative two-site immunoassay system (BDNF EMax
immunoassay system, Promega). Retinal explants treated
with NMDA (100 lM) for various time periods were lysed
Fig. 1 NMDA-induced neuroprotection against glutamate-induced cell
death in cultured retinal explants. (a) Representative drawing of major
cell types in the rat retina at P3. (b) Representative photomicrographs
of retinal explant cultured for 24 h in control medium, or (c) treated
with glutamate (6 mM in the last 6 h of culture). Treatment with glu-
tamate specifically induces cell death in the iINL, as identified by the
condensed pyknotic profiles (arrowheads). (d) NMDA (60–100 lM)
pretreatment protects against glutamate-induced cell death and
(e) addition of an anti-BDNF antibody (10 lg/mL) during NMDA pre-
treatment prevents NMDA-induced neuroprotection. GCL, ganglion
cells layer; iINL, immature inner nuclear layer; IPL, inner plexiform
layer; NBL, neuroblastic layer. Scale bar: 20 lm.
246 R. A. P. Martins et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253
and detection of BDNF was made as indicated by the
manufacturer.
Statistics
Data were analyzed using GraphPad Prism software (GraphPad
Software Inc., San Diego, CA, USA). Analysis of variance was
applied followed by Tukey’s post-hoc test. The significance level
adopted was p < 0.05.
Results
NMDA-mediated neuroprotection in the retina depends
on BDNF, but not on NT-4
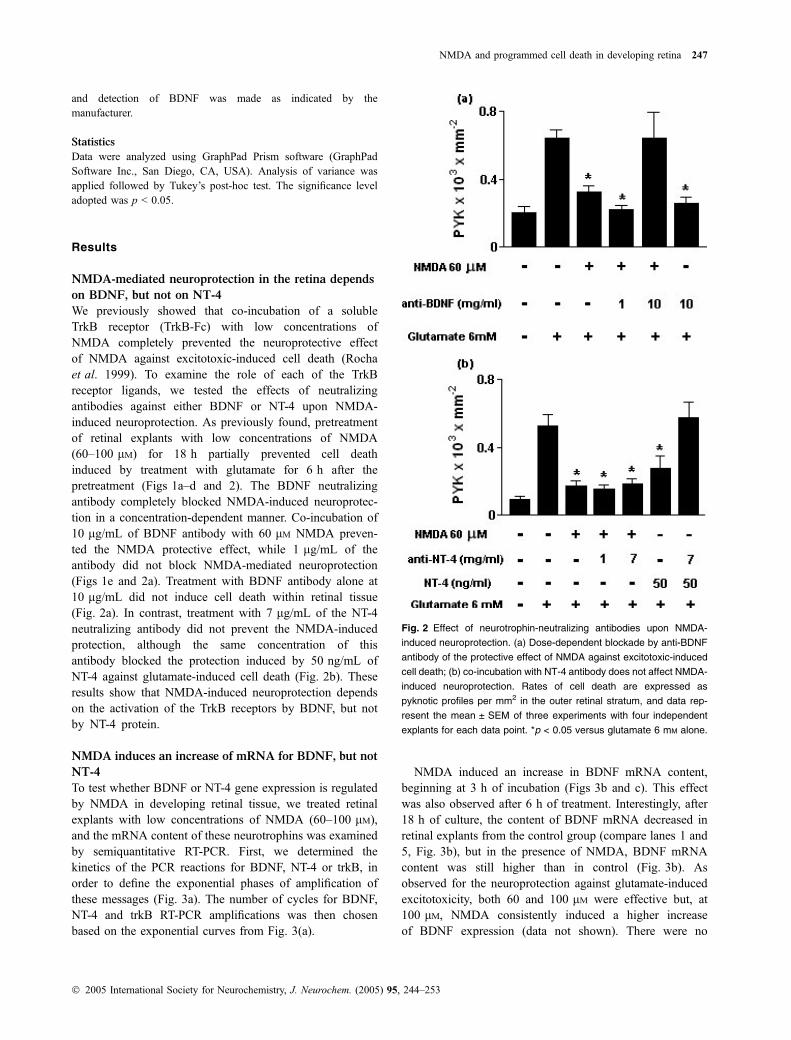
We previously showed that co-incubation of a solubleTrkB receptor (TrkB-Fc) with low concentrations ofNMDA completely prevented the neuroprotective effectof NMDA against excitotoxic-induced cell death (Rochaet al. 1999). To examine the role of each of the TrkBreceptor ligands, we tested the effects of neutralizingantibodies against either BDNF or NT-4 upon NMDA-induced neuroprotection. As previously found, pretreatmentof retinal explants with low concentrations of NMDA(60–100 lM) for 18 h partially prevented cell deathinduced by treatment with glutamate for 6 h after thepretreatment (Figs 1a–d and 2). The BDNF neutralizingantibody completely blocked NMDA-induced neuroprotec-tion in a concentration-dependent manner. Co-incubation of10 lg/mL of BDNF antibody with 60 lM NMDA preven-ted the NMDA protective effect, while 1 lg/mL of theantibody did not block NMDA-mediated neuroprotection(Figs 1e and 2a). Treatment with BDNF antibody alone at10 lg/mL did not induce cell death within retinal tissue(Fig. 2a). In contrast, treatment with 7 lg/mL of the NT-4neutralizing antibody did not prevent the NMDA-inducedprotection, although the same concentration of thisantibody blocked the protection induced by 50 ng/mL ofNT-4 against glutamate-induced cell death (Fig. 2b). Theseresults show that NMDA-induced neuroprotection dependson the activation of the TrkB receptors by BDNF, but notby NT-4 protein.
NMDA induces an increase of mRNA for BDNF, but not
NT-4
To test whether BDNF or NT-4 gene expression is regulatedby NMDA in developing retinal tissue, we treated retinalexplants with low concentrations of NMDA (60–100 lM),and the mRNA content of these neurotrophins was examinedby semiquantitative RT-PCR. First, we determined thekinetics of the PCR reactions for BDNF, NT-4 or trkB, inorder to define the exponential phases of amplification ofthese messages (Fig. 3a). The number of cycles for BDNF,NT-4 and trkB RT-PCR amplifications was then chosenbased on the exponential curves from Fig. 3(a).
NMDA induced an increase in BDNF mRNA content,beginning at 3 h of incubation (Figs 3b and c). This effectwas also observed after 6 h of treatment. Interestingly, after18 h of culture, the content of BDNF mRNA decreased inretinal explants from the control group (compare lanes 1 and5, Fig. 3b), but in the presence of NMDA, BDNF mRNAcontent was still higher than in control (Fig. 3b). Asobserved for the neuroprotection against glutamate-inducedexcitotoxicity, both 60 and 100 lM were effective but, at100 lM, NMDA consistently induced a higher increaseof BDNF expression (data not shown). There were no
Fig. 2 Effect of neurotrophin-neutralizing antibodies upon NMDA-
induced neuroprotection. (a) Dose-dependent blockade by anti-BDNF
antibody of the protective effect of NMDA against excitotoxic-induced
cell death; (b) co-incubation with NT-4 antibody does not affect NMDA-
induced neuroprotection. Rates of cell death are expressed as
pyknotic profiles per mm2 in the outer retinal stratum, and data rep-
resent the mean ± SEM of three experiments with four independent
explants for each data point. *p < 0.05 versus glutamate 6 mM alone.
NMDA and programmed cell death in developing retina 247
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253
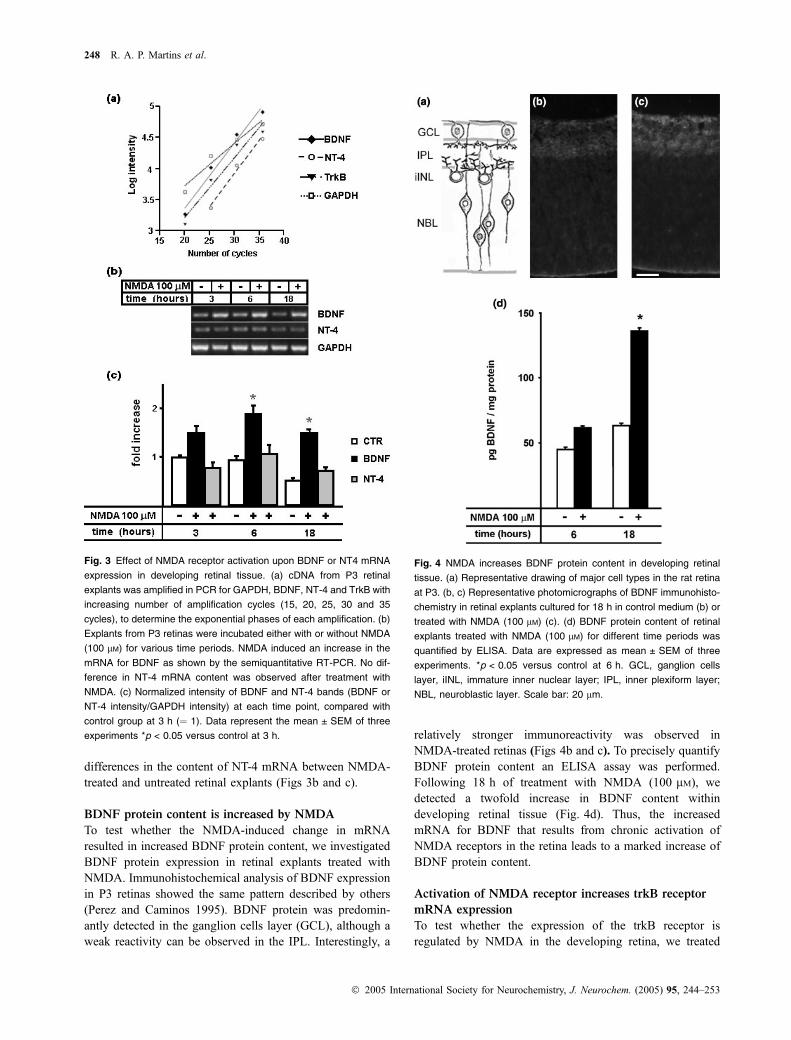
differences in the content of NT-4 mRNA between NMDA-treated and untreated retinal explants (Figs 3b and c).
BDNF protein content is increased by NMDA
To test whether the NMDA-induced change in mRNAresulted in increased BDNF protein content, we investigatedBDNF protein expression in retinal explants treated withNMDA. Immunohistochemical analysis of BDNF expressionin P3 retinas showed the same pattern described by others(Perez and Caminos 1995). BDNF protein was predomin-antly detected in the ganglion cells layer (GCL), although aweak reactivity can be observed in the IPL. Interestingly, a
relatively stronger immunoreactivity was observed inNMDA-treated retinas (Figs 4b and c). To precisely quantifyBDNF protein content an ELISA assay was performed.Following 18 h of treatment with NMDA (100 lM), wedetected a twofold increase in BDNF content withindeveloping retinal tissue (Fig. 4d). Thus, the increasedmRNA for BDNF that results from chronic activation ofNMDA receptors in the retina leads to a marked increase ofBDNF protein content.
Activation of NMDA receptor increases trkB receptor
mRNA expression
To test whether the expression of the trkB receptor isregulated by NMDA in the developing retina, we treated
Fig. 3 Effect of NMDA receptor activation upon BDNF or NT4 mRNA
expression in developing retinal tissue. (a) cDNA from P3 retinal
explants was amplified in PCR for GAPDH, BDNF, NT-4 and TrkB with
increasing number of amplification cycles (15, 20, 25, 30 and 35
cycles), to determine the exponential phases of each amplification. (b)
Explants from P3 retinas were incubated either with or without NMDA
(100 lM) for various time periods. NMDA induced an increase in the
mRNA for BDNF as shown by the semiquantitative RT-PCR. No dif-
ference in NT-4 mRNA content was observed after treatment with
NMDA. (c) Normalized intensity of BDNF and NT-4 bands (BDNF or
NT-4 intensity/GAPDH intensity) at each time point, compared with
control group at 3 h (¼ 1). Data represent the mean ± SEM of three
experiments *p < 0.05 versus control at 3 h.
Fig. 4 NMDA increases BDNF protein content in developing retinal
tissue. (a) Representative drawing of major cell types in the rat retina
at P3. (b, c) Representative photomicrographs of BDNF immunohisto-
chemistry in retinal explants cultured for 18 h in control medium (b) or
treated with NMDA (100 lM) (c). (d) BDNF protein content of retinal
explants treated with NMDA (100 lM) for different time periods was
quantified by ELISA. Data are expressed as mean ± SEM of three
experiments. *p < 0.05 versus control at 6 h. GCL, ganglion cells
layer, iINL, immature inner nuclear layer; IPL, inner plexiform layer;
NBL, neuroblastic layer. Scale bar: 20 lm.
248 R. A. P. Martins et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253
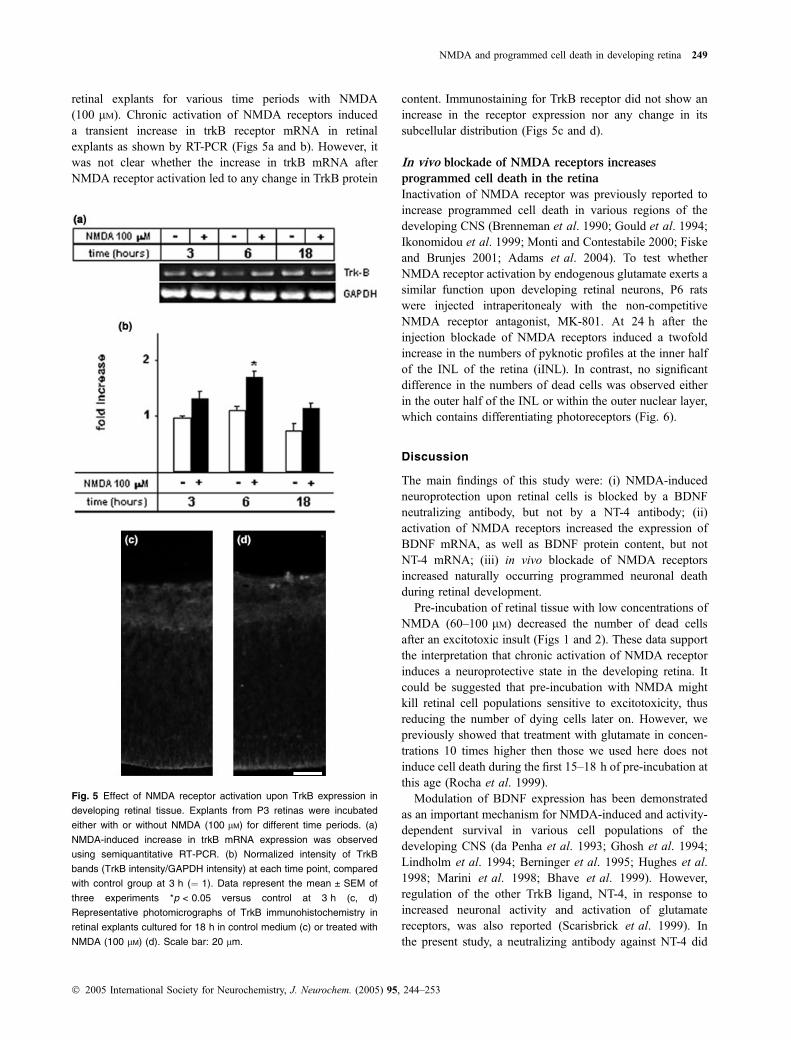
retinal explants for various time periods with NMDA(100 lM). Chronic activation of NMDA receptors induceda transient increase in trkB receptor mRNA in retinalexplants as shown by RT-PCR (Figs 5a and b). However, itwas not clear whether the increase in trkB mRNA afterNMDA receptor activation led to any change in TrkB protein
content. Immunostaining for TrkB receptor did not show anincrease in the receptor expression nor any change in itssubcellular distribution (Figs 5c and d).
In vivo blockade of NMDA receptors increases
programmed cell death in the retina
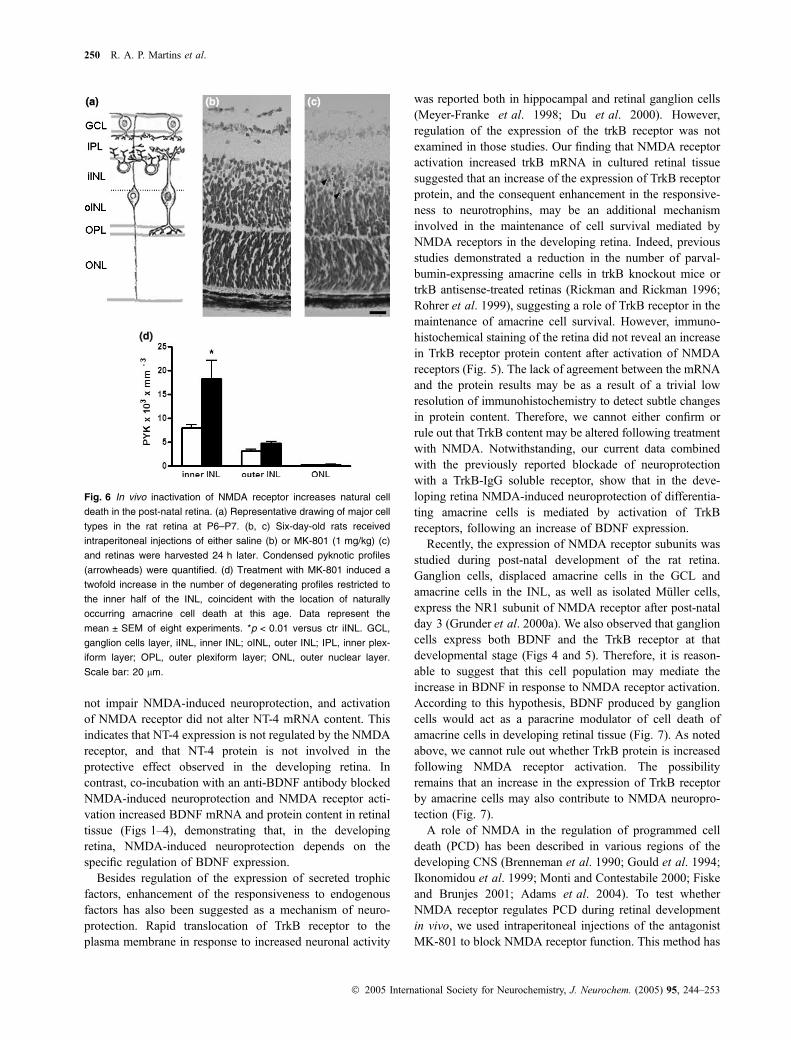
Inactivation of NMDA receptor was previously reported toincrease programmed cell death in various regions of thedeveloping CNS (Brenneman et al. 1990; Gould et al. 1994;Ikonomidou et al. 1999; Monti and Contestabile 2000; Fiskeand Brunjes 2001; Adams et al. 2004). To test whetherNMDA receptor activation by endogenous glutamate exerts asimilar function upon developing retinal neurons, P6 ratswere injected intraperitonealy with the non-competitiveNMDA receptor antagonist, MK-801. At 24 h after theinjection blockade of NMDA receptors induced a twofoldincrease in the numbers of pyknotic profiles at the inner halfof the INL of the retina (iINL). In contrast, no significantdifference in the numbers of dead cells was observed eitherin the outer half of the INL or within the outer nuclear layer,which contains differentiating photoreceptors (Fig. 6).
Discussion
The main findings of this study were: (i) NMDA-inducedneuroprotection upon retinal cells is blocked by a BDNFneutralizing antibody, but not by a NT-4 antibody; (ii)activation of NMDA receptors increased the expression ofBDNF mRNA, as well as BDNF protein content, but notNT-4 mRNA; (iii) in vivo blockade of NMDA receptorsincreased naturally occurring programmed neuronal deathduring retinal development.
Pre-incubation of retinal tissue with low concentrations ofNMDA (60–100 lM) decreased the number of dead cellsafter an excitotoxic insult (Figs 1 and 2). These data supportthe interpretation that chronic activation of NMDA receptorinduces a neuroprotective state in the developing retina. Itcould be suggested that pre-incubation with NMDA mightkill retinal cell populations sensitive to excitotoxicity, thusreducing the number of dying cells later on. However, wepreviously showed that treatment with glutamate in concen-trations 10 times higher then those we used here does notinduce cell death during the first 15–18 h of pre-incubation atthis age (Rocha et al. 1999).
Modulation of BDNF expression has been demonstratedas an important mechanism for NMDA-induced and activity-dependent survival in various cell populations of thedeveloping CNS (da Penha et al. 1993; Ghosh et al. 1994;Lindholm et al. 1994; Berninger et al. 1995; Hughes et al.1998; Marini et al. 1998; Bhave et al. 1999). However,regulation of the other TrkB ligand, NT-4, in response toincreased neuronal activity and activation of glutamatereceptors, was also reported (Scarisbrick et al. 1999). Inthe present study, a neutralizing antibody against NT-4 did
Fig. 5 Effect of NMDA receptor activation upon TrkB expression in
developing retinal tissue. Explants from P3 retinas were incubated
either with or without NMDA (100 lM) for different time periods. (a)
NMDA-induced increase in trkB mRNA expression was observed
using semiquantitative RT-PCR. (b) Normalized intensity of TrkB
bands (TrkB intensity/GAPDH intensity) at each time point, compared
with control group at 3 h (¼ 1). Data represent the mean ± SEM of
three experiments *p < 0.05 versus control at 3 h (c, d)
Representative photomicrographs of TrkB immunohistochemistry in
retinal explants cultured for 18 h in control medium (c) or treated with
NMDA (100 lM) (d). Scale bar: 20 lm.
NMDA and programmed cell death in developing retina 249
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253
not impair NMDA-induced neuroprotection, and activationof NMDA receptor did not alter NT-4 mRNA content. Thisindicates that NT-4 expression is not regulated by the NMDAreceptor, and that NT-4 protein is not involved in theprotective effect observed in the developing retina. Incontrast, co-incubation with an anti-BDNF antibody blockedNMDA-induced neuroprotection and NMDA receptor acti-vation increased BDNF mRNA and protein content in retinaltissue (Figs 1–4), demonstrating that, in the developingretina, NMDA-induced neuroprotection depends on thespecific regulation of BDNF expression.
Besides regulation of the expression of secreted trophicfactors, enhancement of the responsiveness to endogenousfactors has also been suggested as a mechanism of neuro-protection. Rapid translocation of TrkB receptor to theplasma membrane in response to increased neuronal activity
was reported both in hippocampal and retinal ganglion cells(Meyer-Franke et al. 1998; Du et al. 2000). However,regulation of the expression of the trkB receptor was notexamined in those studies. Our finding that NMDA receptoractivation increased trkB mRNA in cultured retinal tissuesuggested that an increase of the expression of TrkB receptorprotein, and the consequent enhancement in the responsive-ness to neurotrophins, may be an additional mechanisminvolved in the maintenance of cell survival mediated byNMDA receptors in the developing retina. Indeed, previousstudies demonstrated a reduction in the number of parval-bumin-expressing amacrine cells in trkB knockout mice ortrkB antisense-treated retinas (Rickman and Rickman 1996;Rohrer et al. 1999), suggesting a role of TrkB receptor in themaintenance of amacrine cell survival. However, immuno-histochemical staining of the retina did not reveal an increasein TrkB receptor protein content after activation of NMDAreceptors (Fig. 5). The lack of agreement between the mRNAand the protein results may be as a result of a trivial lowresolution of immunohistochemistry to detect subtle changesin protein content. Therefore, we cannot either confirm orrule out that TrkB content may be altered following treatmentwith NMDA. Notwithstanding, our current data combinedwith the previously reported blockade of neuroprotectionwith a TrkB-IgG soluble receptor, show that in the deve-loping retina NMDA-induced neuroprotection of differentia-ting amacrine cells is mediated by activation of TrkBreceptors, following an increase of BDNF expression.
Recently, the expression of NMDA receptor subunits wasstudied during post-natal development of the rat retina.Ganglion cells, displaced amacrine cells in the GCL andamacrine cells in the INL, as well as isolated Muller cells,express the NR1 subunit of NMDA receptor after post-natalday 3 (Grunder et al. 2000a). We also observed that ganglioncells express both BDNF and the TrkB receptor at thatdevelopmental stage (Figs 4 and 5). Therefore, it is reason-able to suggest that this cell population may mediate theincrease in BDNF in response to NMDA receptor activation.According to this hypothesis, BDNF produced by ganglioncells would act as a paracrine modulator of cell death ofamacrine cells in developing retinal tissue (Fig. 7). As notedabove, we cannot rule out whether TrkB protein is increasedfollowing NMDA receptor activation. The possibilityremains that an increase in the expression of TrkB receptorby amacrine cells may also contribute to NMDA neuropro-tection (Fig. 7).
A role of NMDA in the regulation of programmed celldeath (PCD) has been described in various regions of thedeveloping CNS (Brenneman et al. 1990; Gould et al. 1994;Ikonomidou et al. 1999; Monti and Contestabile 2000; Fiskeand Brunjes 2001; Adams et al. 2004). To test whetherNMDA receptor regulates PCD during retinal developmentin vivo, we used intraperitoneal injections of the antagonistMK-801 to block NMDA receptor function. This method has
Fig. 6 In vivo inactivation of NMDA receptor increases natural cell
death in the post-natal retina. (a) Representative drawing of major cell
types in the rat retina at P6–P7. (b, c) Six-day-old rats received
intraperitoneal injections of either saline (b) or MK-801 (1 mg/kg) (c)
and retinas were harvested 24 h later. Condensed pyknotic profiles
(arrowheads) were quantified. (d) Treatment with MK-801 induced a
twofold increase in the number of degenerating profiles restricted to
the inner half of the INL, coincident with the location of naturally
occurring amacrine cell death at this age. Data represent the
mean ± SEM of eight experiments. *p < 0.01 versus ctr iINL. GCL,
ganglion cells layer, iINL, inner INL; oINL, outer INL; IPL, inner plex-
iform layer; OPL, outer plexiform layer; ONL, outer nuclear layer.
Scale bar: 20 lm.
250 R. A. P. Martins et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253

been used in other studies (Ikonomidou et al. 1999; Montiand Contestabile 2000), and has the advantage of avoidingintraocular lesions, which may lead to spurious release oftrophic factors that regulate retinal cell death at this stage ofdevelopment (Mansour-Robaey et al. 1994; Cusato et al.2002). Our data showed that loss of NMDA receptor functionin vivo caused an increase in cell death in the inner half of theINL. At that stage of development (1 week post-natal),amacrine cells occupy that portion of the INL, and this is thepeak period of naturally occurring cell death among thisneuronal population (Weidman and Kuwabara 1968; Hors-burgh and Sefton 1987). Taken together, these observationsindicate that the NMDA receptor regulates PCD of amacrinecells in vivo. BDNF was also shown to regulate PCD ofamacrine cells at that same developmental stage (Cusatoet al. 2002; Pinzon-Duarte et al. 2004). Interestingly, studiesin other CNS regions suggested that the physiologicaldependence of NMDA receptor activation for neuronalsurvival occurs through the regulation of BDNF expressionand the activity of the transcription factor CREB (Montiet al. 2002; Hansen et al. 2004). Thus, it is likely that, as
observed in vitro, NMDA can regulate PCD of amacrine cellsthrough the regulation of the expression of BDNF. Otherphysiological roles for the NMDA receptor have beendemonstrated during retinal development, such as NMDAreceptor-mediated regulation of the formation of retinalcircuitry (Tian and Copenhagen 2001; Wong and Wong2001). Our results reveal a novel function for the NMDAreceptor during retinal development that is the control ofnaturally occurring cell death.
In conclusion, our results indicate that NMDA receptor-mediated neuroprotection in developing retinal tissuedepends on the regulation of the expression of BDNF. Inaddition, we showed that physiological activity of theNMDA receptor regulates programmed cell death duringretinal development in vivo.
Acknowledgements
CNPq, FAPERJ, PRONEX-MCT and CAPES-ProDoc supported
this research. We thank Jose Nilson dos Santos, Levi R. de Amorim,
J. F. Tiburcio and Gildo dos Santos for technical assistance, Drs
Maurıcio Caetano, Marılia Guimaraes for coaching with BDNF
mRNA measurements and Dr Marcelo Santiago for assistance with
imaging. RL is a fellow of the John Simon Guggenheim Foundation.
References
Adams S. M., Rivero Vaccari J. C. and Corriveau R. A. (2004)Pronounced cell death in the absence of NMDA receptors in thedeveloping somatosensory thalamus. J. Neurosci. 24, 9441–9450.
de Araujo E. G. and Linden R. (1993) Trophic factors produced byretinal cells increase the survival of retinal ganglion cells in vitro.Eur. J. Neurosci. 5, 1181–1188.
Barnstable C. J. (1993) Glutamate and GABA in retinal circuitry. Curr.Opin. Neurobiol. 3, 520–525.
Berninger B., Marty S., Zafra F., da Penha B. M., Thoenen H. andLindholm D. (1995) GABAergic stimulation switches fromenhancing to repressing BDNF expression in rat hippocampalneurons during maturation in vitro. Development 121, 2327–2335.
Bhave S. V., Ghoda L. and Hoffman P. L. (1999) Brain-derived neuro-trophic factor mediates the anti-apoptotic effect of NMDA incerebellar granule neurons: signal transduction cascades and site ofethanol action. J. Neurosci. 19, 3277–3286.
Brenneman D. E., Forsythe I. D., Nicol T. and Nelson P. G. (1990)N-methyl-D-aspartate receptors influence neuronal survival in devel-oping spinal cord cultures. Brain Res. Dev. Brain Res. 51, 63–68.
Cameron H. A., Hazel T. G. and McKay R. D. (1998) Regulation ofneurogenesis by growth factors and neurotransmitters. J. Neuro-biol. 36, 287–306.
Cellerino A. and Kohler K. (1997) Brain-derived neurotrophic factor/neurotrophin-4 receptor TrkB is localized on ganglion cells anddopaminergic amacrine cells in the vertebrate retina. J. Comp.Neurol. 386, 149–160.
Cepko C. L. (1999) The roles of intrinsic and extrinsic cues and bHLHgenes in the determination of retinal cell fates. Curr. Opin. Neu-robiol. 9, 37–46.
Coskun V. and Luskin M. B. (2002) Intrinsic and extrinsic regulation ofthe proliferation and differentiation of cells in the rodent rostralmigratory stream. J. Neurosci. Res. 69, 795–802.
Fig. 7 Possible cellular interactions mediating NMDA-induced neuro-
protection. The expression pattern of BDNF, NMDA and TrkB
receptors during early retinal development and the data obtained in
this study suggest a paracrine neuroprotective effect of NMDA upon
amacrine cells survival in developing retina, that occurs through an
increase in the expression of BDNF by ganglion cells and activation of
TrkB receptors expressed by amacrine cells. The second symbol
depicting TrkB in the amacrine cell at the left indicates the possibility
that TrkB receptor is also up-regulated.
NMDA and programmed cell death in developing retina 251
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253

Cusato K., Bosco A., Linden R. and Reese B. E. (2002) Cell death in theinner nuclear layer of the retina is modulated by BDNF. Brain Res.Dev. Brain Res. 139, 325–330.
Du J., Feng L., Yang F. and Lu B. (2000) Activity- and Ca(2+)-dependent modulation of surface expression of brain-derivedneurotrophic factor receptors in hippocampal neurons. J. Cell Biol.150, 1423–1434.
Dyer M. A. and Cepko C. L. (2001) Regulating proliferation duringretinal development. Nat. Rev. Neurosci. 2, 333–342.
Ferguson K. L. and Slack R. S. (2003) Growth factors: can they promoteneurogenesis? Trends Neurosci. 26, 283–285.
Fiske B. K. and Brunjes P. C. (2001) NMDA receptor regulation of celldeath in the rat olfactory bulb. J. Neurobiol. 47, 223–232.
Ghosh A., Carnahan J. and Greenberg M. E. (1994) Requirement forBDNF in activity-dependent survival of cortical neurons. Science263, 1618–1623.
Gould E., Cameron H. A. and McEwen B. S. (1994) Blockade ofNMDA receptors increases cell death and birth in the developingrat dentate gyrus. J. Comp. Neurol. 340, 551–565.
GrunderT.,KohlerK.,KalettaA. andGuenther E. (2000a)The distributionand developmental regulation of NMDA receptor subunit proteins inthe outer and inner retina of the rat. J. Neurobiol. 44, 333–342.
Grunder T., Kohler K. and Guenther E. (2000b) Distribution anddevelopmental regulation of AMPA receptor subunit proteins in ratretina. Invest. Ophthalmol. Vis. Sci. 41, 3600–3606.
Hansen H. H., Briem T., Dzietko M. et al. (2004) Mechanisms leading todisseminated apoptosis following NMDA receptor blockade in thedeveloping rat brain. Neurobiol. Dis. 16, 440–453.
Haydar T. F., Wang F., Schwartz M. L. and Rakic P. (2000) Differentialmodulation of proliferation in the neocortical ventricular andsubventricular zones. J. Neurosci. 20, 5764–5774.
Horsburgh G. M. and Sefton A. J. (1987) Cellular degeneration andsynaptogenesis in the developing retina of the rat. J. Comp. Neurol.263, 553–566.
Huang E. J. and Reichardt L. F. (2003) Trk receptors: roles in neuronalsignal transduction. Annu. Rev. Biochem. 72, 609–642.
Hughes P. E., Young D., Preston K. M., Yan Q. and DragunowM. (1998)Differential regulation by MK801 of immediate-early genes, brain-derived neurotrophic factor and trk receptor mRNA induced by akindling after-discharge. Brain Res. Mol. Brain Res. 53, 138–151.
Ikonomidou C., Bosch F., Miksa M., Bittigau P., Vockler J., DikranianK., Tenkova T. I., Stefovska V., Turski L. and Olney J. W. (1999)Blockade of NMDA receptors and apoptotic neurodegeneration inthe developing brain. Science 283, 70–74.
Job C. and Tan S. S. (2003) Constructing the mammalian neocortex: therole of intrinsic factors. Dev. Biol. 257, 221–232.
Johansson K., Bruun A., Torngren M. and Ehinger B. (2000) Develop-ment of glutamate receptor subunit 2 immunoreactivity in postnatalrat retina. Vis. Neurosci. 17, 737–742.
Johnson J., Tian N., Caywood M. S., Reimer R. J., Edwards R. H. andCopenhagen D. R. (2003) Vesicular neurotransmitter transporterexpression in developing postnatal rodent retina: GABA and gly-cine precede glutamate. J. Neurosci. 23, 518–529.
Kaplan D. R. and Miller F. D. (2000) Neurotrophin signal transduction inthe nervous system. Curr. Opin. Neurobiol. 10, 381–391.
Kingsbury T. J., Murray P. D., Bambrick L. L. and Krueger B. K. (2003)Ca(2+)-dependent regulation of TrkB expression in neurons.J. Biol. Chem. 278, 40 744–40 748.
Komuro H. and Rakic P. (1993) Modulation of neuronal migration byNMDA receptors. Science 260, 95–97.
Linden R., Rehen S. K. and Chiarini L. B. (1999) Apoptosis in devel-oping retinal tissue. Prog. Retina. Eye Res. 18, 133–165.
Linden R., Martins R. A. P. and Silveira M. (2005) Control ofprogrammed cell death by neurotransmitters and neuropeptides in
the developing mammalian retina. Prog. Retinal Eye Res. 24,457–491.
Lindholm D., Castren E., Berzaghi M., Blochl A. and Thoenen H. (1994)Activity-dependent and hormonal regulation of neurotrophinmRNA levels in the brain – implications for neuronal plasticity.J. Neurobiol. 25, 1362–1372.
LoTurco J. J., Owens D. F., Heath M. J., Davis M. B. and Kriegstein A.R. (1995) GABA and glutamate depolarize cortical progenitor cellsand inhibit DNA synthesis. Neuron 15, 1287–1298.
Mansour-Robaey S., Clarke D. B., Wang Y. C., Bray G. M. and AguayoA. J. (1994) Effects of ocular injury and administration of brain-derived neurotrophic factor on survival and regrowth of axotomizedretinal ganglion cells. Proc. Natl Acad. Sci. USA 91, 1632–1636.
Marini A. M., Rabin S. J., Lipsky R. H. and Mocchetti I. (1998)Activity-dependent release of brain-derived neurotrophic factorunderlies the neuroprotective effect of N-methyl-D-aspartate.J. Biol. Chem. 273, 29 394–29 399.
Marquardt T. (2003) Transcriptional control of neuronal diversificationin the retina. Prog. Retin Eye Res. 22, 567–577.
Massey S. C. and Redburn D. A. (1987) Transmitter circuits in thevertebrate retina. Prog. Neurobiol. 28, 55–96.
Meyer-Franke A., Wilkinson G. A., Kruttgen A., Hu M., Munro E.,Hanson M. G. Jr, Reichardt L. F. and Barres B. A. (1998) Depo-larization and cAMP elevation rapidly recruit TrkB to the plasmamembrane of CNS neurons. Neuron 21, 681–693.
Monti B. and Contestabile A. (2000) Blockade of the NMDA receptorincreases developmental apoptotic elimination of granule neuronsand activates caspases in the rat cerebellum. Eur. J. Neurosci. 12,3117–3123.
Monti B., Marri L. and Contestabile A. (2002) NMDA receptor-dependent CREB activation in survival of cerebellar granule cellsduring in vivo and in vitro development. Eur. J. Neurosci. 16,1490–1498.
Nakazawa T., Nakano I., Sato M., Nakamura T., Tamai M. and Mori N.(2002) Comparative expression profiles of Trk receptors and Shc-related phosphotyrosine adapters during retinal development:potential roles of N-Shc/ShcC in brain-derived neurotrophic factorsignal transduction and modulation. J. Neurosci. Res. 68, 668–680.
Nguyen L., Rigo J. M., Rocher V., Belachew S., Malgrange B., RogisterB., Leprince P. and Moonen G. (2001) Neurotransmitters as earlysignals for central nervous system development. Cell Tissue Res.305, 187–202.
Pallas S. L. (2001) Intrinsic and extrinsic factors that shape neocorticalspecification. Trends Neurosci. 24, 417–423.
da Penha B. M., Cooper J., Castren E., Zafra F., Sofroniew M., ThoenenH. and Lindholm D. (1993) Cholinergic regulation of brain-derivedneurotrophic factor (BDNF) and nerve growth factor (NGF) but notneurotrophin-3 (NT-3) mRNA levels in the developing rat hippo-campus. J. Neurosci. 13, 3818–3826.
Perez M. T. and Caminos E. (1995) Expression of brain-derived neu-rotrophic factor and of its functional receptor in neonatal and adultrat retina. Neurosci. Lett. 183, 96–99.
Pinzon-Duarte G., Arango-Gonzalez B., Guenther E. and Kohler K.(2004) Effects of brain-derived neurotrophic factor on cell survival,differentiation and patterning of neuronal connections and Mullerglia cells in the developing retina. Eur. J. Neurosci. 19, 1475–1484.
Pow D. V. and Barnett N. L. (1999) Changing patterns of spatial buff-ering of glutamate in developing rat retinae are mediated by theMuller cell glutamate transporter GLAST. Cell Tissue Res. 297,57–66.
Rehen S. K., Varella M. H., Freitas F. G., Moraes M. O. and Linden R.(1996) Contrasting effects of protein synthesis inhibition and ofcyclic AMP on apoptosis in the developing retina. Development122, 1439–1448.
252 R. A. P. Martins et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253

Rickman D. W. and Brecha N. C. (1995) Expression of the proto-oncogene, trk, receptors in the developing rat retina. Vis. Neurosci.12, 215–222.
Rickman D. W. and Rickman C. B. (1996) Suppression of trkBexpression by antisense oligonucleotides alters a neuronal pheno-type in the rod pathway of the developing rat retina. Proc. NatlAcad. Sci USA 93, 12 564–12 569.
Rocha M., Martins R. A. and Linden R. (1999) Activation of NMDAreceptors protects against glutamate neurotoxicity in the retina:evidence for the involvement of neurotrophins. Brain Res. 827,79–92.
Rohrer B., Korenbrot J., LaVail M., Reichardt L. and Xu B. (1999) Roleof neurotrophin receptor TrkB in the maturation of rod photore-ceptors and establishment of synaptic transmission to the innerretina. J.Neurosci. 19, 8919–8930.
Scarisbrick I. A., Isackson P. J. and Windebank A. J. (1999) Differentialexpression of brain-derived neurotrophic factor, neurotrophin-3,and neurotrophin-4/5 in the adult rat spinal cord: regulation by theglutamate receptor agonist kainic acid. J. Neurosci. 19, 7757–7769.
Shieh P. B., Hu S. C., Bobb K., Timmusk T. and Ghosh A. (1998)Identification of a signaling pathway involved in calcium regula-tion of BDNF expression. Neuron 20, 727–740.
Sometani A., Kataoka H., Nitta A., Fukumitsu H., Nomoto H. andFurukawa S. (2001) Transforming growth factor-b1 enhances
expression of brain-derived neurotrophic factor and its receptor,TrkB, in neurons cultured from rat cerebral cortex. J. Neurosci.Res. 66, 369–376.
Sucher N. J., Kohler K., Tenneti L., Wong H. K., Grunder T., Fauser S.,Wheeler-Schilling T., Nakanishi N., Lipton S. A. and Guenther E.(2003) N-methyl-D-aspartate receptor subunit NR3A in the retina:developmental expression, cellular localization, and functionalaspects. Invest. Ophthalmol. Vis. Sci. 44, 4451–4456.
Tao X., Finkbeiner S., Arnold D. B., Shaywitz A. J. and Greenberg M. E.(1998) Ca2+ influx regulates BDNF transcription by a CREB familytranscription factor-dependent mechanism. Neuron 20, 709–726.
Thorenson W. B. and Witkovsky P. (1999) Glutamate receptors andcircuits in the vertebrate retina. Prog. Retin Eye Res. 18, 765–810.
Tian N. and Copenhagen D. R. (2001) Visual deprivation alters devel-opment of synaptic function in inner retina after eye opening.Neuron 32, 439–449.
Weidman T. A. and Kuwabara T. (1968) Postnatal development of therat retina. An electron microscopic study. Arch. Ophthalmol. 79,470–484.
Wong W. T. and Wong R. O. (2001) Changing specificity of neuro-transmitter regulation of rapid dendritic remodeling during syn-aptogenesis. Nat. Neurosci. 4, 351–352.
Zhang S. S., Fu X. Y. and Barnstable C. J. (2002) Molecular aspects ofvertebrate retinal development. Mol. Neurobiol. 26, 137–152.
NMDA and programmed cell death in developing retina 253
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 244–253
Related Documents











