Nitrile-converting enzymes: An eco-friendly tool for industrial biocatalysis Pramod W. Ramteke 1∗ Navodita G. Maurice 1 Babu Joseph 2 Bharat J. Wadher 3 1 Department of Biological Sciences, Sam Higginbotom Institute of Agriculture, Technology and Sciences, Allahabad, India 2 College of Applied Medical Sciences, Shaqra University, Shaqra, Kingdom of Saudi Arabia 3 PG Department of Microbiology, Nagpur University, Nagpur, India Abstract Nitriles are organic compounds bearing a –C≡N group; they are frequently known to occur naturally in both fauna and flora and are also synthesized chemically. They have wide applicability in the fields of medicine, industry, and environmental monitoring. However, the majority of nitrile compounds are considered to be lethal, mutagenic, and carcinogenic in nature and are known to cause potential health problems such as nausea, bronchial irritation, respiratory distress, convulsions, coma, and skeletal deformities in humans. Nitrile-converting enzymes, which are extracted from microorganisms, are commonly termed nitrilases and have drawn the attention of researchers all over the world to combat the toxicity of nitrile compounds. The present review focuses on the utility of nitrile-converting enzymes, sources, classification, structure, properties, and applications, as well as the future perspective on nitrilases. C 2013 International Union of Biochemistry and Molecular Biology, Inc. Volume 60, Number 5, Pages 459–481, 2013 Keywords: biocatalysis, bioremediation, nitriles, nitrilases, Rhodococcus 1. Introduction Nitrile compounds are intermediates, products, by-products, and waste products of agriculture, chemical, pharmaceutical industries, and processing of fossil fuels [1]. Nitriles are found in fruit pits and brassica crops (such as cabbage, brussels sprouts, and cauliflower), which are released during hydrol- ysis. Mandelonitrile, a cyanohydrin produced by ingesting almonds or other fruit pits, releases hydrogen cyanide, which is responsible for the toxicity of cyanogenic glycosides [2]. Over 30 nitrile-containing pharmaceuticals are currently marketed for a diverse variety of medicinal indications with more than Abbreviations: ACN, acrylonitrile; AS, amidase signature; FAAH, fatty acid amide hydrolase; IAA, indole-3-acetic acid; IAN, indoleacetonitrile; IDPN, iminodipropionitrile; IUPAC, International Union of Pure and Applied Chemistry; NBR, nitrile butadiene rubber; NCBI, National Center for Biotechnology Information. ∗ Address for correspondence: Pramod W. Ramteke, PhD, Department of Biological Sciences, Sam Higginbotom Institute of Agriculture, Technology and Sciences, Allahabad 211007, India. Tel.: +91 532 2684572; Fax: +91 532 2684394; e-mail: [email protected]. Received 13 March 2013; accepted 21 June 2013 DOI: 10.1002/bab.1139 Published online 25 October 2013 in Wiley Online Library (wileyonlinelibrary.com) 20 additional nitrile-containing leads in clinical developmental studies [3]. The nitrile group is relatively inert; sometimes it is not metabolized and passes through the body unchanged. In pharmaceuticals, nitriles have diverse applications, from vildagliptin, an antidiabetic drug, to anastrazole, which is used in treating breast cancer. The nitrile mimics functionality present in substrates for enzymes, whereas in other cases the nitrile increases water solubility or decreases susceptibility to oxidative metabolism in the liver [4]. Industrially, nitriles are produced by ammoxidation and hydrocyanation. In ammonox- idation, a hydrocarbon is partially oxidized in the presence of ammonia; this is widely practiced for large-scale conversion for acrylonitrile (ACN) [5]. A nitrile is any organic compound that has a –C≡N functional group (IUPAC). In the industrial litera- ture, the prefix cyano- is used interchangeably with the term nitrile. It is mostly found in superglue (methyl cyanoacrylate) and latex-free laboratory and medical gloves [nitrile butadiene rubber (NBR)]. Inorganic compounds containing the –C≡N group are called cyanides instead of nitriles (NCBI). Although both nitriles and cyanides are the derivatives of cyanide salts, most nitriles are not as toxic as cyanides. Chemical industries make extensive use of various ni- trile compounds for manufacturing a variety of polymers and other chemicals. For example, ACN and adiponitrile are re- quired for the production of polyacrylonitrile and nylon 66 459

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Nitrile-converting enzymes: An eco-friendly
tool for industrial biocatalysis
Pramod W. Ramteke1∗
Navodita G. Maurice1
Babu Joseph2
Bharat J. Wadher3
1Department of Biological Sciences, Sam Higginbotom Institute ofAgriculture, Technology and Sciences, Allahabad, India2College of Applied Medical Sciences, Shaqra University, Shaqra, Kingdomof Saudi Arabia3PG Department of Microbiology, Nagpur University, Nagpur, India
Abstract
Nitriles are organic compounds bearing a –C≡N group; theyare frequently known to occur naturally in both fauna and floraand are also synthesized chemically. They have wideapplicability in the fields of medicine, industry, andenvironmental monitoring. However, the majority of nitrilecompounds are considered to be lethal, mutagenic, andcarcinogenic in nature and are known to cause potential healthproblems such as nausea, bronchial irritation, respiratorydistress, convulsions, coma, and skeletal deformities in
humans. Nitrile-converting enzymes, which are extracted frommicroorganisms, are commonly termed nitrilases and havedrawn the attention of researchers all over the world tocombat the toxicity of nitrile compounds. The present reviewfocuses on the utility of nitrile-converting enzymes, sources,classification, structure, properties, and applications, as wellas the future perspective on nitrilases. C© 2013 International Unionof Biochemistry and Molecular Biology, Inc. Volume 60, Number 5, Pages459–481, 2013
Keywords: biocatalysis, bioremediation, nitriles, nitrilases, Rhodococcus
1. IntroductionNitrile compounds are intermediates, products, by-products,and waste products of agriculture, chemical, pharmaceuticalindustries, and processing of fossil fuels [1]. Nitriles are foundin fruit pits and brassica crops (such as cabbage, brusselssprouts, and cauliflower), which are released during hydrol-ysis. Mandelonitrile, a cyanohydrin produced by ingestingalmonds or other fruit pits, releases hydrogen cyanide, whichis responsible for the toxicity of cyanogenic glycosides [2]. Over30 nitrile-containing pharmaceuticals are currently marketedfor a diverse variety of medicinal indications with more than
Abbreviations: ACN, acrylonitrile; AS, amidase signature; FAAH, fatty acidamide hydrolase; IAA, indole-3-acetic acid; IAN, indoleacetonitrile; IDPN,iminodipropionitrile; IUPAC, International Union of Pure and AppliedChemistry; NBR, nitrile butadiene rubber; NCBI, National Center forBiotechnology Information.∗Address for correspondence: Pramod W. Ramteke, PhD, Department ofBiological Sciences, Sam Higginbotom Institute of Agriculture, Technologyand Sciences, Allahabad 211007, India. Tel.: +91 532 2684572; Fax: +91532 2684394; e-mail: [email protected] 13 March 2013; accepted 21 June 2013DOI: 10.1002/bab.1139Published online 25 October 2013 in Wiley Online Library(wileyonlinelibrary.com)
20 additional nitrile-containing leads in clinical developmentalstudies [3]. The nitrile group is relatively inert; sometimes itis not metabolized and passes through the body unchanged.In pharmaceuticals, nitriles have diverse applications, fromvildagliptin, an antidiabetic drug, to anastrazole, which isused in treating breast cancer. The nitrile mimics functionalitypresent in substrates for enzymes, whereas in other cases thenitrile increases water solubility or decreases susceptibility tooxidative metabolism in the liver [4]. Industrially, nitriles areproduced by ammoxidation and hydrocyanation. In ammonox-idation, a hydrocarbon is partially oxidized in the presence ofammonia; this is widely practiced for large-scale conversion foracrylonitrile (ACN) [5]. A nitrile is any organic compound thathas a –C≡N functional group (IUPAC). In the industrial litera-ture, the prefix cyano- is used interchangeably with the termnitrile. It is mostly found in superglue (methyl cyanoacrylate)and latex-free laboratory and medical gloves [nitrile butadienerubber (NBR)]. Inorganic compounds containing the –C≡Ngroup are called cyanides instead of nitriles (NCBI). Althoughboth nitriles and cyanides are the derivatives of cyanide salts,most nitriles are not as toxic as cyanides.
Chemical industries make extensive use of various ni-trile compounds for manufacturing a variety of polymers andother chemicals. For example, ACN and adiponitrile are re-quired for the production of polyacrylonitrile and nylon 66
459

Biotechnology andApplied Biochemistry
FIG. 1Enzymatic pathways for nitrile hydrolysis.
polymers [6], synthesis of acrylamide [7], nicotinamide [8],and cyanoverlamide [9]. Different types of nitrile compoundsare used in pharmaceuticals, drug intermediates (chiral syn-thons), solvents, feedstock, extractants, pesticides (bromoxynil,dichlobenil), and so on. In organic synthesis of esters, amines,amides, amidines, aldehydes, carboxylic acids, heterocycliccompounds, and ketones, nitriles are the intermediates [10].NBR or nitrile rubber is a copolymer of butadiene and ACN; itoffers high strength and exceptional balance for execution ina wide range of industrial and automotive applications. China,Japan, and the USA are continuing as primary consumers ofrubber. China, in particular, owing to robust growth in domes-tic, industrial goods, and motor vehicles, is expected to exceed645 thousand tons in consumption by 2017 [11].
A majority of the nitrile compounds are found as lethal,mutagenic, and carcinogenic in nature [12] and cause potentialhealth problems, like nausea, bronchial irritation, respiratorydistress, convulsions, coma, and skeletal deformities in humans.The toxicity of acrylonitrile (ACN) in rats resulted in neuro-behavioral changes, correlated with locomotor behavior, motornerves, learning, and memory [13,14]. On the other hand, IDPNwas found to inhibit dopaminergic neurotransmission [15].
1.1. NitrilasesEnzymes are incredible catalysts capable of catalyzing chemicalreactions in a biological cell. Biochemists and microbiologistsview biocatalysis as a field with immense promise for chemicalsynthesis. Enzymes are selective for their substrates, speedup reactions in many cases, and determine the metabolicpathways [16]. Nitrilase was first discovered by Mahadevanand Thiman [17] from plants about 48 years ago. Bioproductionof glycolic acid from glyconitrile has been reported from abacterial isolate of Alcaligenes sp. ECU0401 [17]. Nitrilase isknown to convert indole-3-acetonitrile into indole-3-acetic acid(IAA), a plant growth hormone [18]. The nitrilases involvedin the formation of IAA have been found in plant families,namely Graminae (grasses), Musaceae (banana family), andCruciferae (cabbage family) [19–21]. In the plant Arabidopsisthaliana, two metabolic pathways catalyzed by nitrilase forthe synthesis of IAA were identified [22]. A. thaliana has fournitrilase homologs: NIT1, NIT2, NIT3, and NIT4. NIT4 does not
hydrolyze indoleacetonitrile (IAN) and is highly selective forβ-cyano-l-alanine [23].
Nitrile-converting biocatalysts are now of substantial in-dustrial interest from the perspective of treating toxic nitrileand cyanide containing desecrates and as mediators for thesynthesis of chemicals for industrial applications [24]. Nitri-lases are commercially useful enzymes in the production ofpyrazinoic acid (an antimycobacterial agent), nicotinic acid,isotopically active amino acids from amino nitriles [25], ibupro-fen, and in degradation of bromoxynil and ioxynil [26]. Inindustry, most nitriles are converted chemically to corre-spondingly high value acids and amides. However, chemicalconversion requires either strongly acidic or basic medium,high-energy expenditure, or causes generation of unnecessarytoxic by-products. Thus, the application of enzymes for theorganic chemical process is increasing. Nitrile hydratases werefirst explored as biocatalysts for the synthesis of chemicals, forexample, acrylamide, acrylic acid, and nicotinamide [27]. Al-though the benefits of using enzymes in organic synthesis havebeen fully demonstrated, however, much attention is neededfor systematic management of microorganisms for productionof new enzymes [28]. The nitrile-degrading enzymes consistof nitrilases and amidases. The microbial degradation of ni-triles proceeds via two different enzymatic pathways (Fig. 1).Nitrilase, cyanide dihydratase, and cyanide hydratase catalyzethe direct hydrolysis of nitriles into the corresponding car-boxylic acid and ammonia. Nitrile hydratase converts nitrilesto the corresponding amide, which is then hydrolyzed to thecorresponding carboxylic acid and ammonia by an amidase[29,30]. Nitrilases hydrolyze larger nitriles like alkyl cyanides(RCN), whereas cyanide dihydratases and cyanide hydrataseshydrolyze hydrogen cyanide (HCN) only.
2. Bacteria and Fungi as Sourcesof NitrilaseMicroorganisms isolated from soil and freshwater, as well asmarine sediments, were screened for nitrilase activity [31]. Onlya few fungi are also known to convert nitrile compounds bynitrilase production, such as Fusarium oxysporum, F. melonis,Aspergillus, Penicillium, and so on [32]. The nitrilases fromAspergillus niger CBS 513.88 (NitAn) and Neurospora crassaOR74A (NitNc) were the first fungal arylaceto nitrilases to beexpressed in Escherichia coli [33] and subsequently purified[34]. The enzyme in A. niger K-10 showed broad specificity
460 Nitrile-Converting Enzymes

FIG. 2Chemical hydrolysis of nitriles in harsh and mildconditions.
toward heteroaromatic and aromatic nitriles and was inducedby cyanopyridine [33]. It was also found that indolyacetonitrilewas hydrolyzed by fungi belonging to Fusarium, Giberella,Aspergillus, and Penicillium [29, 35] to 2-aminoproponitrileand 4-amino-4-cyanobutyric acid in basidiomycetes [36,37].
The first bacterial nitrilase was isolated from a soil bac-terium, a pseudomonad, using a ricinine, a naturally occurringnitrile as the sole source of carbon [38, 39]. This enzyme hy-drolyzed ricinine and 2-pyridones to the corresponding acidand ammonia. Two kinds of nitrilases from Arthrobacter werecharacterized, which degrade aromatic nitriles such as ben-zonitriles and p-tolunitile [8]. Nitrilases are homo-oligomersor less frequently monomers and do not contain any metalcofactor [40]. They are assumed to have cysteine residues ator near the active site, which is important for catalytic activityof the enzyme [41]. The reaction mechanism is proposed toinvolve a nucleophilic attack on the nitrile carbon atom by asulfhydryl group of a cysteine residue followed by hydrolysis(Fig. 2). It has been previously reported that aromatic andheterocyclic nitriles in which the cyano group is conjugatedwith a double bond are readily degraded by nitrilase, whereasaliphatic nitriles are catabolized in two stages, via conversionto the corresponding amides and then to acids and ammoniaby nitrile hydratase and amidase, respectively [42]. All theknown nitrilases acting on aromatic nitrilases as substrateswere unable to degrade preferentially aliphatic nitriles [43].
The nitrilase of Rhodococcus rhodochrous J1 was inducedwhen isovaleronitrile was used in the medium [44]. Thesecells were cultured in a medium containing urea and cyclo-hexane carboxamide in the presence of cobalt ions, high- andlow-molecular-weight nitrile hydratases were induced corre-spondingly [45]. Other organisms that express nitrilases wheninduced with isovaleronitrile include Alcaligenes faecalis JM3,Rhodococcus ATCC 39484, and R. rhodochrous K22 [46–48].The nitrilase from R. rhodochrous NCIMB 11216 is induced withpropionitrile as the sole carbon and nitrogen source [49]. A newnitrilase was isolated from Labrenzia aggregate DSM 13394[50], which showed 45% identity with template nitrilase from
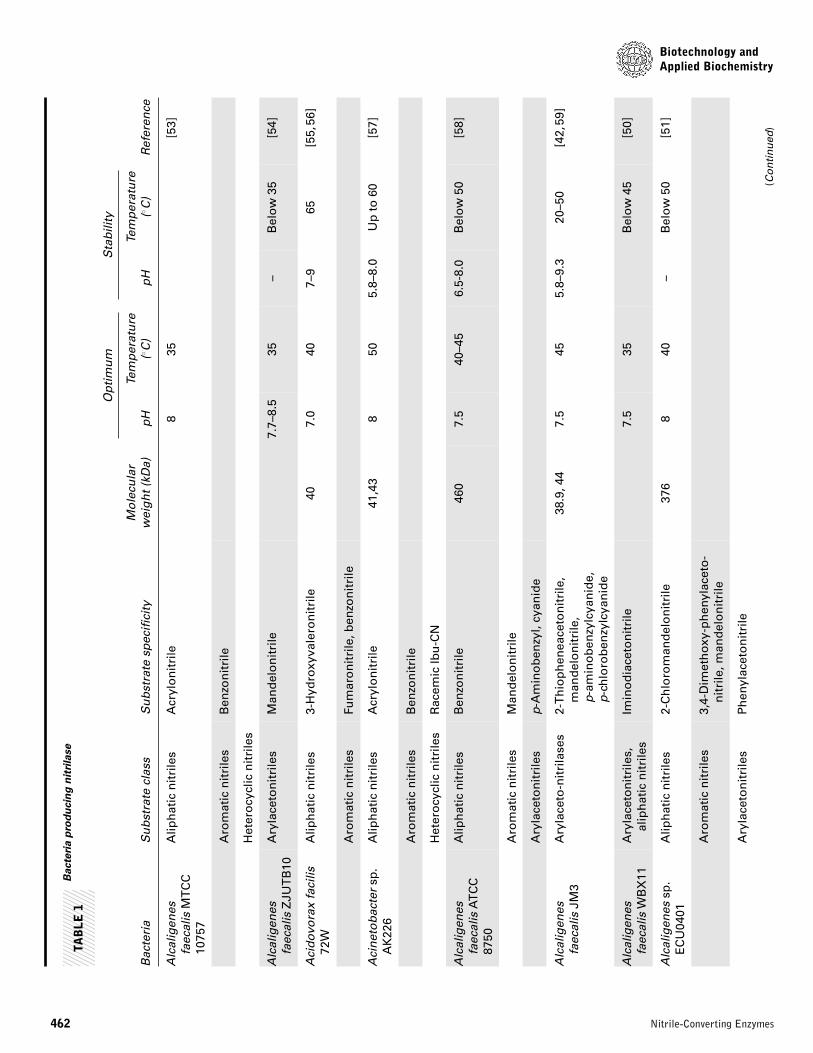
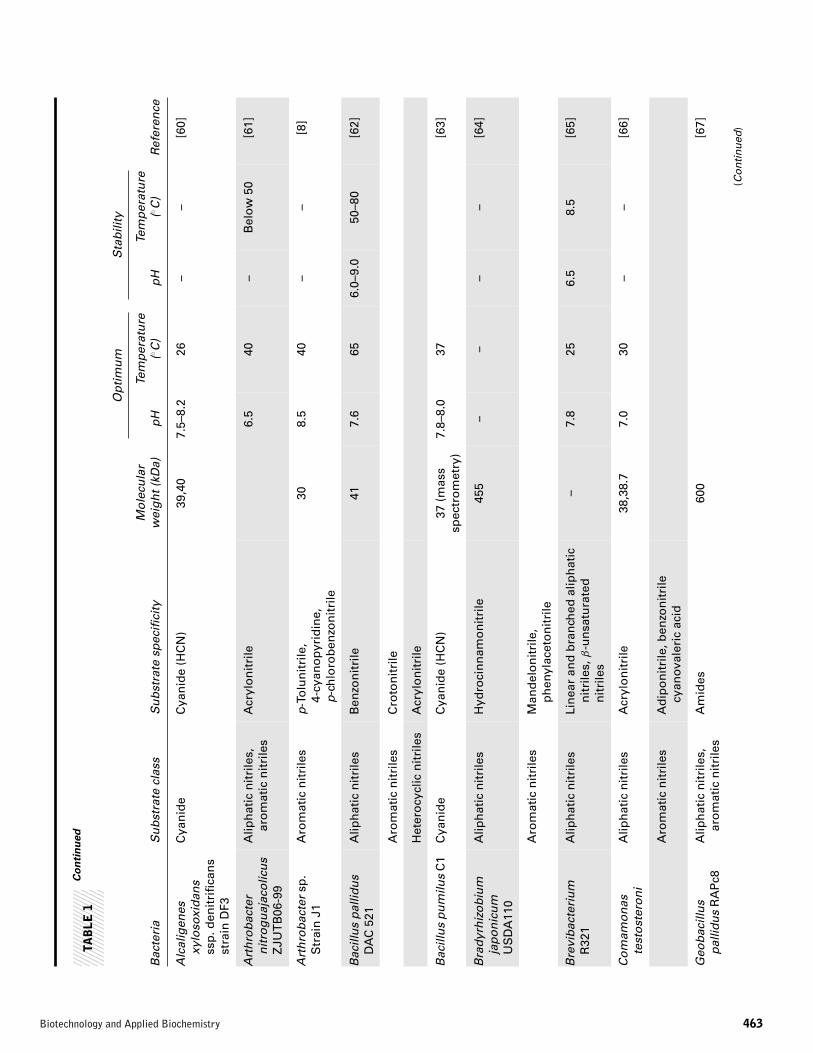
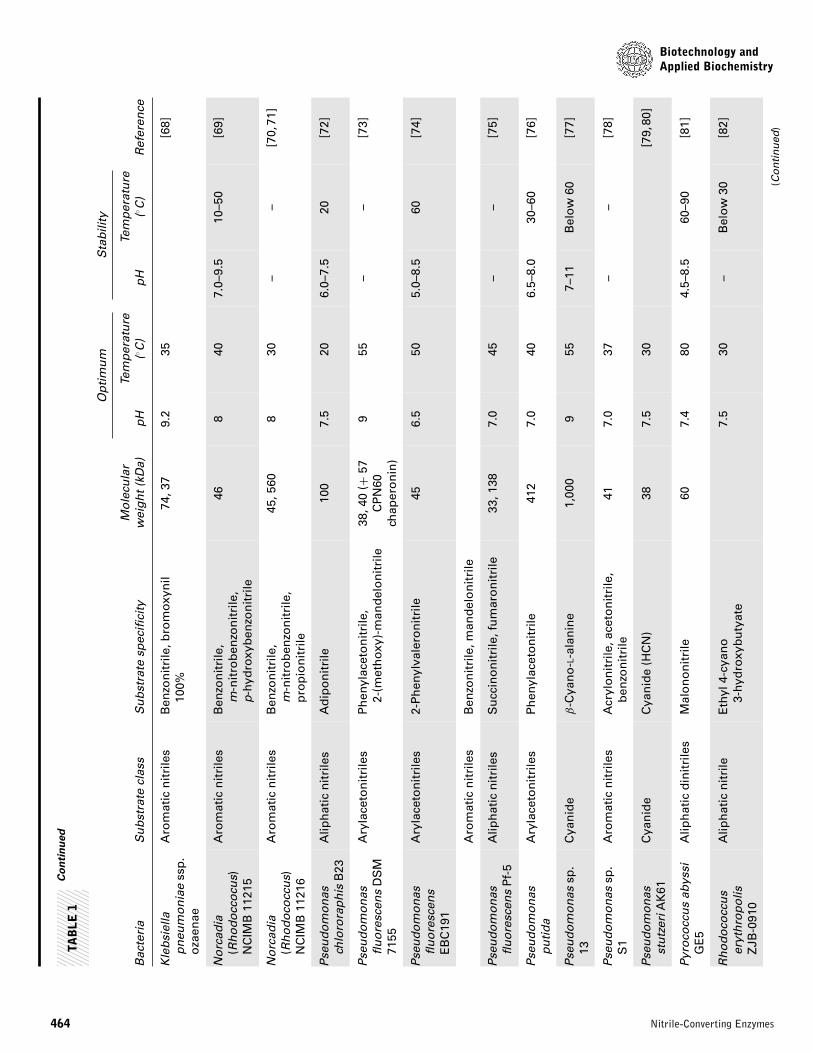
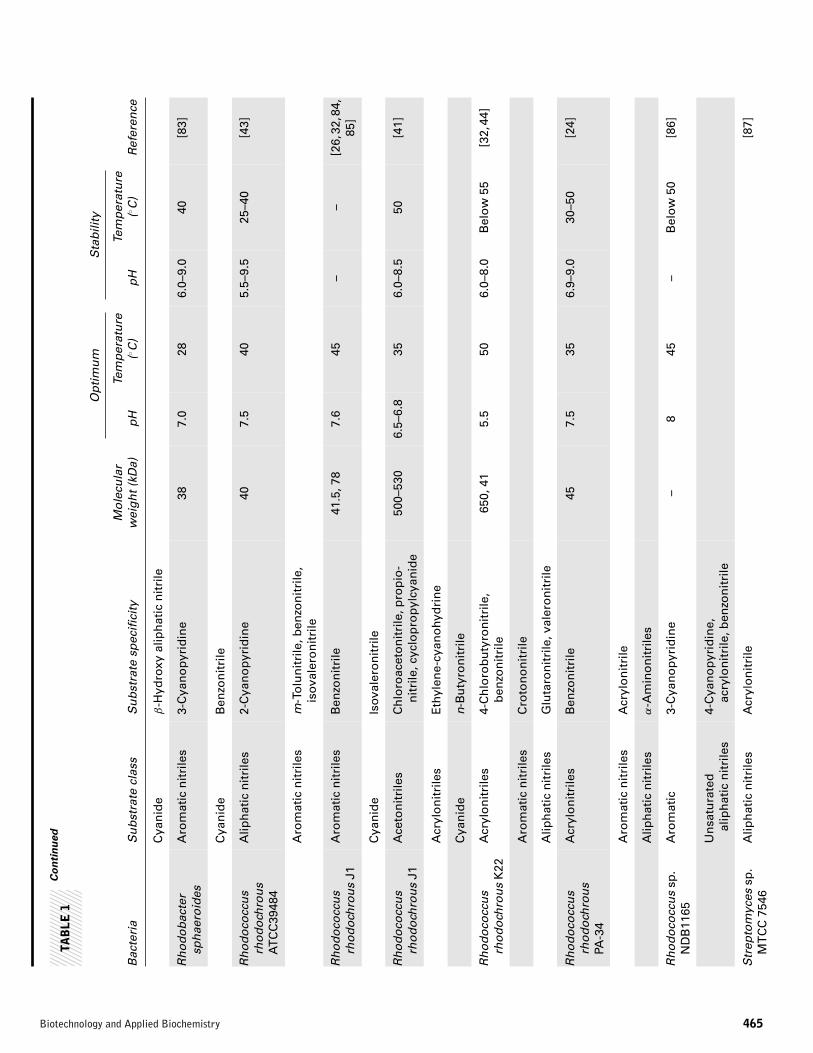
Alcaligenes sp. ECU 0401 [51]. Thus, the number of reportedbacteria with nitrilases rapidly increased in two decades [52].Bacteria exhibiting nitrilase activity were isolated and screenedfor metabolizing synthetic and natural nitriles. The potentialuse of nitrilases in industry has increased the need for screen-ing and characterization of bacterial nitrilase. A list of variousnitrilase producing bacteria is presented in Table 1.
3. Nitrilase SuperfamilyPrimarily, the carbon–nitrogen (C–N) hydrolyzing enzymesincluded nitrilases, cyanide hydratase, aliphatic amidase, andβ-ureidopropionase [88]. These enzymes have been classi-fied into 13 distinct branches based on amino-acid sequenceanalysis as the nitrilase superfamily [89]. Enzymes in thesuperfamily are used by prokaryotes and eukaryotes to hy-drolyze and condense a variety of protein and nonprotein C–Nbonds [90]. Branch 1 enzymes consist of nitrilases, cyanidedihydratases, and cyanide hydratases, which digest only ni-triles. However, the in vivo substrates for nitrilases remainunknown (except for the cyanide dihydratases and cyanidehydratases). The majority of the remaining 12 branches exhibitamidase activity with varying specificity. Branches 2, 3, 4,and 5 consist of aliphatic amidases, amino-terminal amidase,biotinidase, and β-ureidopropionase, respectively. Branch 6enzymes, namely the carbamylases, are specific for the decar-bamylation of d-amino acids. Branch 7 and 8 enzymes consistof prokaryotic and eukaryotic NAD synthetases, which utilizeeither glutamine or free ammonia to catalyze the transferof amide nitrogen to a wide range of substrates. A study onNAD+ synthetase from Mycobacterium tuberculosis showedthat glutamine-amidotransferase activity depends on an amino-terminal nitrilase domain, called a glutamine amide transferdomain [91]. Mutation of the active cysteine in the nitrilasedomain inactivated the NAD+ synthetase from M. tuberculosis.This suggests that the nitrilase domain is sometimes fusedto another domain and may be part of a multifaceted chainof reactions obligatory for the production of metabolites invarious biological systems [89]. Hence, there is need to markthese enzymes and associated domains as a potential sourceof novel antibacterial agents [92]. Only branch 9 enzymes,namely the alipoprotein N-acyltransferases, catalyze an amidecondensation reaction. The members in the superfamily sharea common fold with a conserved catalytic cys-glu-lys site, whichis surrounded by signature sequences conserved within eachbranch but differing between branches [91]. The functions ofenzymes in branches 11, 12, and 13 are unknown.
3.1. Nitrile hydratasesNitrile hydratases (NHases; EC 4.2.1.84) are mononucleariron or cobalt enzymes that catalyze the hydration of a largenumber of miscellaneous nitriles to their analogous amides.NHase is a key enzyme in the bienzymatic pathway for theconversion of nitriles to acids, which converts nitriles to thecorresponding amides. NHases can be classified into two broad
Biotechnology and Applied Biochemistry 461
Biotechnology andApplied Biochemistry
TAB
LE
1B
acte
ria
pro
du
cin
gn
itri
lase
Opt
imum
Sta
bilit
y
Bac
teri
aS
ubst
rate
clas
sS
ubst
rate
spec
ifici
tyM
olec
ular
wei
ght(
kDa)
pHTe
mpe
ratu
re(◦ C
)pH
Tem
pera
ture
(◦ C)
Ref
eren
ce
Alc
alig
enes
faec
alis
MT
CC
1075
7
Alip
hat
icn
itri
les
Acr
ylo
nit
rile
835
[53]
Aro
mat
icn
itri
les
Ben
zon
itri
le
Het
ero
cycl
icn
itri
les
Alc
alig
enes
faec
alis
ZJU
TB
10A
ryla
ceto
nit
rile
sM
and
elo
nit
rile
7.7–
8.5
35–
Bel
ow
35[5
4]
Aci
dovo
rax
faci
lis72
WA
liph
atic
nit
rile
s3-
Hyd
roxy
vale
ron
itri
le40
7.0
407–
965
[55,
56]
Aro
mat
icn
itri
les
Fum
aro
nit
rile
,ben
zon
itri
le
Aci
neto
bact
ersp
.A
K22
6A
liph
atic
nit
rile
sA
cryl
on
itri
le41
,43
850
5.8–
8.0
Up
to60
[57]
Aro
mat
icn
itri
les
Ben
zon
itri
le
Het
ero
cycl
icn
itri
les
Rac
emic
Ibu
-CN
Alc
alig
enes
faec
alis
AT
CC
8750
Alip
hat
icn
itri
les
Ben
zon
itri
le46
07.
540
–45
6.5-
8.0
Bel
ow
50[5
8]
Aro
mat
icn
itri
les
Man
del
on
itri
le
Ary
lace
ton
itri
les
p-A
min
ob
enzy
l,cy
anid
e
Alc
alig
enes
faec
alis
JM3
Ary
lace
to-n
itri
lase
s2-
Th
iop
hen
eace
ton
itri
le,
man
del
on
itri
le,
p-am
ino
ben
zylc
yan
ide,
p-ch
loro
ben
zylc
yan
ide
38.9
,44
7.5
455.
8–9.
320
–50
[42,
59]
Alc
alig
enes
faec
alis
WB
X11
Ary
lace
ton
itri
les,
alip
hat
icn
itri
les
Imin
od
iace
ton
itri
le7.
535
Bel
ow
45[5
0]
Alc
alig
enes
sp.
EC
U04
01A
liph
atic
nit
rile
s2-
Ch
loro
man
del
on
itri
le37
68
40–
Bel
ow
50[5
1]
Aro
mat
icn
itri
les
3,4-
Dim
eth
oxy-
ph
enyl
acet
o-
nit
rile
,man
del
on
itri
le
Ary
lace
ton
itri
les
Ph
enyl
acet
on
itri
le
(Con
tinue
d)
462 Nitrile-Converting Enzymes
TAB
LE
1C
on
tin
ued
Opt
imum
Sta
bilit
y
Bac
teri
aS
ubst
rate
clas
sS
ubst
rate
spec
ifici
tyM
olec
ular
wei
ght(
kDa)
pHTe
mpe
ratu
re(◦ C
)pH
Tem
pera
ture
(◦ C)
Ref
eren
ce
Alc
alig
enes
xylo
soxi
dans
ssp
.den
itri
fica
ns
stra
inD
F3
Cya
nid
eC
yan
ide
(HC
N)
39,4
07.
5–8.
226
––
[60]
Art
hrob
acte
rni
trog
uaja
colic
usZ
JUT
B06
-99
Alip
hat
icn
itri
les,
aro
mat
icn
itri
les
Acr
ylo
nit
rile
6.5
40–
Bel
ow
50[6
1]
Art
hrob
acte
rsp
.S
trai
nJ1
Aro
mat
icn
itri
les
p-To
lun
itri
le,
4-cy
ano
pyr
idin
e,p-
chlo
rob
enzo
nit
rile
308.
540
––
[8]
Bac
illus
palli
dus
DA
C52
1A
liph
atic
nit
rile
sB
enzo
nit
rile
417.
665
6.0–
9.0
50–8
0[6
2]
Aro
mat
icn
itri
les
Cro
ton
itri
le
Het
ero
cycl
icn
itri
les
Acr
ylo
nit
rile
Bac
illus
pum
ilus
C1
Cya
nid
eC
yan
ide
(HC
N)
37(m
ass
spec
tro
met
ry)
7.8–
8.0
37[6
3]
Bra
dyrh
izob
ium
japo
nicu
mU
SD
A11
0
Alip
hat
icn
itri
les
Hyd
roci
nn
amo
nit
rile
455
––
––
[64]
Aro
mat
icn
itri
les
Man
del
on
itri
le,
ph
enyl
acet
on
itri
le
Bre
viba
cter
ium
R32
1A
liph
atic
nit
rile
sLi
nea
ran
db
ran
ched
alip
hat
icn
itri
les,
β-u
nsa
tura
ted
nit
rile
s
–7.
825
6.5
8.5
[65]
Com
amon
aste
stos
tero
niA
liph
atic
nit
rile
sA
cryl
on
itri
le38
,38.
77.
030
––
[66]
Aro
mat
icn
itri
les
Ad
ipo
nit
rile
,ben
zon
itri
lecy
anov
aler
icac
id
Geo
baci
llus
palli
dus
RA
Pc8
Alip
hat
icn
itri
les,
aro
mat
icn
itri
les
Am
ides
600
[67]
(Con
tinue
d)
Biotechnology and Applied Biochemistry 463
Biotechnology andApplied Biochemistry
TAB
LE
1C
on
tin
ued
Opt
imum
Sta
bilit
y
Bac
teri
aS
ubst
rate
clas
sS
ubst
rate
spec
ifici
tyM
olec
ular
wei
ght(
kDa)
pHTe
mpe
ratu
re(◦ C
)pH
Tem
pera
ture
(◦ C)
Ref
eren
ce
Kle
bsie
llapn
eum
onia
ess
p.
oza
enae
Aro
mat
icn
itri
les
Ben
zon
itri
le,b
rom
oxyn
il10
0%74
,37
9.2
35[6
8]
Nor
cadi
a(R
hodo
ccoc
us)
NC
IMB
1121
5
Aro
mat
icn
itri
les
Ben
zon
itri
le,
m-n
itro
ben
zon
itri
le,
p-hy
dro
xyb
enzo
nit
rile
468
407.
0–9.
510
–50
[69]
Nor
cadi
a(R
hodo
cocc
us)
NC
IMB
1121
6
Aro
mat
icn
itri
les
Ben
zon
itri
le,
m-n
itro
ben
zon
itri
le,
pro
pio
nit
rile
45,5
608
30–
–[7
0,71
]
Pseu
dom
onas
chlo
rora
phis
B23
Alip
hat
icn
itri
les
Ad
ipo
nit
rile
100
7.5
206.
0–7.
520
[72]
Pseu
dom
onas
fluor
esce
nsD
SM
7155
Ary
lace
ton
itri
les
Ph
enyl
acet
on
itri
le,
2-(m
eth
oxy)
-man
del
on
itri
le38
,40
(+57
CP
N60
chap
ero
nin
)
955
––
[73]
Pseu
dom
onas
fluor
esce
nsE
BC
191
Ary
lace
ton
itri
les
2-P
hen
ylva
lero
nit
rile
456.
550
5.0–
8.5
60[7
4]
Aro
mat
icn
itri
les
Ben
zon
itri
le,m
and
elo
nit
rile
Pseu
dom
onas
fluor
esce
nsP
f-5
Alip
hat
icn
itri
les
Su
ccin
on
itri
le,f
um
aro
nit
rile
33,1
387.
045
––
[75]
Pseu
dom
onas
putid
aA
ryla
ceto
nit
rile
sP
hen
ylac
eto
nit
rile
412
7.0
406.
5–8.
030
–60
[76]
Pseu
dom
onas
sp.
13C
yan
ide
β-C
yan
o-L
-ala
nin
e1,
000
955
7–11
Bel
ow
60[7
7]
Pseu
dom
onas
sp.
S1
Aro
mat
icn
itri
les
Acr
ylo
nit
rile
,ace
ton
itri
le,
ben
zon
itri
le41
7.0
37–
–[7
8]
Pseu
dom
onas
stut
zeri
AK
61C
yan
ide
Cya
nid
e(H
CN
)38
7.5
30[7
9,80
]
Pyro
cocc
usab
yssi
GE
5A
liph
atic
din
itri
les
Mal
on
on
itri
le60
7.4
804.
5–8.
560
–90
[81]
Rho
doco
ccus
eryt
hrop
olis
ZJB
-091
0
Alip
hat
icn
itri
leE
thyl
4-cy
ano
3-hy
dro
xyb
uty
ate
7.5
30–
Bel
ow
30[8
2]
(Con
tinue
d)
464 Nitrile-Converting Enzymes
TAB
LE
1C
on
tin
ued
Opt
imum
Sta
bilit
y
Bac
teri
aS
ubst
rate
clas
sS
ubst
rate
spec
ifici
tyM
olec
ular
wei
ght(
kDa)
pHTe
mpe
ratu
re(◦ C
)pH
Tem
pera
ture
(◦ C)
Ref
eren
ce
Cya
nid
eβ
-Hyd
roxy
alip
hat
icn
itri
le
Rho
doba
cter
spha
eroi
des
Aro
mat
icn
itri
les
3-C
yan
op
yrid
ine
387.
028
6.0–
9.0
40[8
3]
Cya
nid
eB
enzo
nit
rile
Rho
doco
ccus
rhod
ochr
ous
AT
CC
3948
4
Alip
hat
icn
itri
les
2-C
yan
op
yrid
ine
407.
540
5.5–
9.5
25–4
0[4
3]
Aro
mat
icn
itri
les
m-T
olu
nit
rile
,ben
zon
itri
le,
isov
aler
on
itri
le
Rho
doco
ccus
rhod
ochr
ous
J1A
rom
atic
nit
rile
sB
enzo
nit
rile
41.5
,78
7.6
45–
–[2
6,32
,84,
85]
Cya
nid
eIs
oval
ero
nit
rile
Rho
doco
ccus
rhod
ochr
ous
J1A
ceto
nit
rile
sC
hlo
roac
eto
nit
rile
,pro
pio
-n
itri
le,c
yclo
pro
pyl
cyan
ide
500–
530
6.5–
6.8
356.
0–8.
550
[41]
Acr
ylo
nit
rile
sE
thyl
ene-
cyan
ohy
dri
ne
Cya
nid
en-
Bu
tyro
nit
rile
Rho
doco
ccus
rhod
ochr
ous
K22
Acr
ylo
nit
rile
s4-
Ch
loro
bu
tyro
nit
rile
,b
enzo
nit
rile
650,
415.
550
6.0–
8.0
Bel
ow
55[3
2,44
]
Aro
mat
icn
itri
les
Cro
ton
on
itri
le
Alip
hat
icn
itri
les
Glu
taro
nit
rile
,val
ero
nit
rile
Rho
doco
ccus
rhod
ochr
ous
PA-3
4
Acr
ylo
nit
rile
sB
enzo
nit
rile
457.
535
6.9–
9.0
30–5
0[2
4]
Aro
mat
icn
itri
les
Acr
ylo
nit
rile
Alip
hat
icn
itri
les
α-A
min
on
itri
les
Rho
doco
ccus
sp.
ND
B11
65A
rom
atic
3-C
yan
op
yrid
ine
–8
45–
Bel
ow
50[8
6]
Un
satu
rate
dal
iph
atic
nit
rile
s4-
Cya
no
pyr
idin
e,ac
rylo
nit
rile
,ben
zon
itri
le
Str
epto
myc
essp
.M
TC
C75
46A
liph
atic
nit
rile
sA
cryl
on
itri
le[8
7]
Biotechnology and Applied Biochemistry 465

Biotechnology andApplied Biochemistry
groups. Ferric NHases and cobalt NHases, the metal ions, arevery good catalysts for –CN hydration and are required forthe folding or the stabilization of the subunit polypeptides ofthe enzyme [4]. A large number of microorganisms exhibitingNHase activity have been isolated and the purified enzymecharacterized. All the characterized NHases showed a widerange of substrate specificity and physiochemical properties.The bacterial NHases contain a metal ion in their active site Fe3+
and/or Co3+ [93] and Zn2+ in the enzyme from Myrrotheciumverrucaria [94]. In yeast, nitriles are most likely hydrolyzedvia a nitrile hydratase/amidase pathway as correspondingamides were also accepted as substrate. All the characterizedbacterial nitrile hydratases are composed of two kinds ofsubunits accumulated into heteroligomers (mostly dimmersor tetramers, but also larger subunits containing up to 20subunits) [95]. The molecular level of the catalytic mechanismof the nitrile hydratase is interesting, as the activity of NHasehas unique features when exposed to light [96, 97]. Threedimensional studies carried out on the nonheme ferric nitrilehydratase from Rhodococcus sp. N-771 showed that low spinferric ion is coordinated by the five ligands—three sulfur atomsof cysteine residues and two chain nitrogen atoms. This enzymeis then deactivated by nitric oxide, which binds reversibly to thesixth coordination site, probably the catalytic site. Nitric oxideis held in its position by interaction with three oxygen atomsof the catalytic residues and is released by photoirradiation. Inthe absence of NO, the iron probably coordinates a hydroxideion that acts as a nucleophile attacking the nitrile carbon viaan activated water molecule. As a result, an intermediate isformed, which finally tautomerizes to an amide [95,96].
The actinomycete R. rhodococcus produces two NHasesdepending on the inducer and in the presence of cobalt ions[98]. High- and low-molecular-weight NHases (H- and L-NHases) were selectively induced in the presence of urea andcyclohexane carboxamide [99]. H-NHases exhibited empathyfor aliphatic nitriles, whereas L-NHases acted on aromaticnitriles [43, 98]. The study of these NHases suggested that itcontains cobalt as cofactors. The cobalt NHases have threoninein the active site sequence, whereas ferric NHases containedserine [100]. The different metal cofactors present in thenitrile hydratases can be attributed to the different amino-acidresidues (threonine/serine) in the active site [4].
3.2. AmidasesAn amide bond is of considerable importance in biochemistrybecause many C-terminal amide-containing peptides act ashormones [101] and many are involved in nitrogen metabolismin the prokaryotic as well as eukaryotic cells [102]. Althoughnitrilases break the C–N bond of a nitrile, amidases cleave theC–N bond of an amide to produce the equivalent carboxylicacid and ammonia. These enzymes encompass branch 2 of thenitrilase superfamily and examples embrace amidases fromPseudomonas aeruginosa, R. erythropolis, Helicobacter pylori,and Bacillus stearothermophilus [90]. One amidase (EC3.5.1.4)attracted interest in assorted fields such as neurobiochemistry,
plant physiology, and applied microbiology. This enzyme, whichcatalyzes hydrolysis of an amide to form acid and ammonium,has been shown to be engaged in the following areas: (i)processes inactivating fatty acid amidases as neuronally activesignaling molecules [103]; (ii) biosynthesis of indoleacetic acidas a vital plant hormone [104]; and (iii) enzymatic production inan industrial scale of marketable products [28]. Characteristicsof amidases from diverse sources are dissimilar; some arespecific to aliphatic amides, and others cleave amides ofaromatic origin. Stereoselectivity is usually correlated with theamidases in the bienzymatic pathway [105]. Studies reveal thatthis group of amidases symbolizes a large class, serine–lysinehydrolases [84].
Mutational and comparative modeling studies on theamidase from P. aeruginosa reveal that it has a cys-glu-lyscatalytic triad that is aligned with that of the NitFhit struc-ture (PDB ID, 1EMS) from the worm Caenorhabditis elegans[65,106]. There exists a class of amidases that are structurallyor sequentially alike to nitrilases. The bacteria R. rhodochrousJ1 is reported to have both nitrilase and amidase enzymes,which hydrolyze benzonitrile and benzamide, respectively,using similar catalytic sites [107]. Thus, the reaction mech-anism of both nitrilase and amidase catalyzed reactions isanalogous but the active nucleophile present in enzyme differs.The reaction mechanisms of amidase involve nitrile hydrol-ysis. The carbonyl group of amide undergoes a nucleophilicattack, resulting in the formation of a tetrahedral intermediate,which is renovated to acyl-enzyme with the removal of ammo-nia and subsequently hydrolyzed to acid. All of the differentamidases also exhibit acyl transfer activity in the presence ofhydroxylamine [59].
Wide-spectrum amidases from Brevibacterium sp. strainR312 [108] and P. aeruginosa [109] hydrolyze only short chainamides. These enzymes are made up of four and six indistin-guishable subunits having molecular weights of about 45,000and 35,000 Da, respectively. The other amidases are enantios-elective amidases isolated from Rhodococcus sp. strain R312[104] Delftia acidovorans [110], Rhodococcus sp. N-774 [111],Rhodococcus J1 [112], P. cholororaphis B23 [113], R. erythropo-lis MP50 [114]; they belong to a group of amidases containing aGGSS signature in the amino-acid sequence [115] and are madeup of two identical subunits. There is no sequence similaritybetween the amidase from R. rhodochrous J1 with the amidasefrom P. aeruginosa. However, the nitrilase from R. rhodochrousJ1 has significant sequence similarity with the aliphatic ami-dase from P. aeruginosa and both have cysteine as the activenucleophile [107]. Prominent differences between nitrilasesare apparent in their substrate specificity, enzyme structure,pH, and temperature optima [116]. Amidases from variousmicroorganisms have been described to date; aliphatic (wide-spectrum) amidases acting on aliphatic amides with short acylchains were found in P. aeruginosa [117], Brevibacterium sp.[118], and B. stearothermophilus BR388 [119]. Fungal amidaseshave been reported in Aspergillus and Penicillium sp. and usedin production of essential pharmaceutical commodities. Two
466 Nitrile-Converting Enzymes

unlinked loci gmdA and bzuA have previously been identifiedin Aspergillus nidulans required for the utilization of benza-mide as the sole nitrogen source [120]. Cloning of each genewas done via complementation. The gmdA gene encodes anenvisaged product belonging to the amidase signature (AS) se-quence family that displays similarity to AmdS from A. nidulans[121]. Rahim et al. developed a quick plate for the detectionof acetamidase and A. candidius produced the maximum(397.79 U/mL) acetamidase among various Aspergillus spp. Itproduced inducible intracellular acetamidase, which showedcatabolite repression by glucose at concentrations greater than0.7%. The enzyme was optimally active at 40◦C, pH 5.0, andwas stable up to a concentration of 10 mM [118]. The AS familywas originally identified by primary structure analysis, whichrevealed an exceedingly conserved serine and glycine-richsequence present in several amidases of bacterial and fungalorigin [104, 115]. Proteins containing the AS sequence existin archae [122], eubacterium [111, 112], fungi [120], nema-todes, plants, insects, birds [123], and mammals [102]. Thesubstrate specificities and biological function of these enzymesvary widely, including carbon/nitrogen metabolism in fungithrough the hydrolysis of acetamide [119], generation of prop-erly charged tRNAGln in eubacteria through glutamine [124],and the degradation of neuromodulatory fatty acids [102].Fatty acid amide hydrolase (FAAH) belongs to a large classof hydrolytic enzymes termed the Amidase Signature Familywhose members have a conserved sketch of approximately 130amino acids termed the AS sequence [125].
Aliophatic amidases (EC 3.5.1.4) are enzymes catalyzingthe hydrolysis of short chain amides to produce ammonia andthe corresponding amino acid. Such an amidase, AmiE, hasbeen spotted formerly in H. pylori. The two H. pylori amidaseswere discriminated by different biochemical uniqueness such asoptimum pH or temperature. AmiE hydrolyzed proprionamide,acetamide, acrylamide but not formamide. AmiF showed anunexpected substrate specificity by hydrolyzing formamide.AmiF is thus the first formamidase (EC 3.5.1.49) relatedto aliphatic amidases described. Production of AmiE andAmiF proteins is dependent on the activity of other enzymesinvolved in the nitrogen metabolism of H. pylori (ureaseand arginase, respectively). It strongly suggested that theH. pylori paralogous amidases have advanced to accomplishenzymatic specialization after ancestral gene duplicationand the production of these enzymes helped to maintainintracellular nitrogen balance in H. pylori [126].
Peptide amylase catalyzed amidations of the C-terminalcarboxylic group; peptides were studied using model substrateof a large series of Nα protected di-, tri-, tetra-, and penta-peptides in the presence of ammonium bicarbonate as theammonium source. The maximum yields of amide synthesiswere attained in medium consisting of acetonitrile with 20–25 volume percentage of dimethyl formamide and 3% water.Under these conditions, the substrate specificity of the enzymewas more restricted in the synthetic reactions than amidehydrolysis. Elongation of the peptide chain has a negative
effect on enzyme amidation [69]. The phylogenetic analysis andamino-acid sequence proposed that the novel R. erythropolisenzymes belong to the iron-type NHase family. Some differentresidues in the translated sequences were positioned nearthe residue involved in the stabilization of NHase active site,suggesting that the substitutions could be responsible for thedifferent enzyme activities and substrate specificities [68].The nitrile transforming activities of R. erythropolis strainsfrom widely disconnected geographic locations have beencharacterized and found to have a NHase amidase hydrolyzingsystem [127]. A novel amidase from Blastobacter sp. strainAI7p-4 was found to be associated with bacterial cyclic imidemetabolism. The enzyme functions in the second step ofcyclic imide degradation of monoamidated dicarboxylates(half-amides) to dicarboxylates and ammonia [71]. Enzymeproduction increased in the presence of cyclic imides such asglutarimide and succinimide but not by amide compounds thatare common substrates and inducers of amidases.
Amidases from microorganisms showed a wide diver-sity of substrate specificities. A report on the hydrolysis ofamides containing a bulky substituent at lysis of amide ni-trogen such as tert-butylcarboamide is not available. Thisinability to hydrolyze bulky amides increased the wide use ofamidases for the production of complex compounds. Therefore,microorganisms were screened for hydrolysis of piperazine-2-tert-butylcarboxamide and hydrolytic (amidase) activity wasfound in Pseudomonas sp. MCI3434. The amidase was purifiedfrom cells, which hydrolyzed R-stereoselectivity piperazine-2-tert-butylcarboxamide to form R-piperazine-2-carboxylic acidand was named R-amidase. The gene coding for the R-amidasewas isolated, sequenced, and expressed in E. coli host. Therecombinant protein was purified, characterized, and found tobe a novel amidase with unique substrate specificity [26].
3.3. Aromatic nitrilasesAromatic nitrilases have been purified from the bacteriaPseudomonas sp., Norcadia sp. NCIMB 11215 and NCIMB11216 [32, 57], Arthrobacter sp. strain J1 [8], R. rhodochrousJ1 [109], Klebsiella pneumonae subsp. ozaenae [128], andthe fungus Fusarium solani [32]. This class consists of nitri-lases that hydrolyze benzonitrile and related aromatic sub-strates. The best characterized nitrilase is from R. rhodochrousJ1, which was found to be specific for aromatic or hete-rocylic nitriles such as benzonitrile, 3-chlorobenzonitrile,and 2-thiophenecarbonitrile. This enzyme was purified fromisovaleronitrile-induced cells of R. rhodochrous J1 and ex-hibits regiospecificity by hydrolyzing 1,3-dicyanobenzoate and1,4-dicyanobenzene to 3-cyanobenzoate and 4-cyanobenzoate,respectively [129]. Most of these enzymes are inactive onaliphatic nitriles except for the nitrilase of R. rhodochrous (for-merly Norcadia) NCIMB 11216, which hydrolyzes benzonitrileand propionitrile, an aliphatic nitrile [130]. The nitrilase fromR. rhodochrous J1 could hydrolyze ACN upon substrate-inducedactivation of the native organism with benzonitrile [131]. Thenitrilase from isovaleronitrile-induced cells of Rhodococcus
Biotechnology and Applied Biochemistry 467

Biotechnology andApplied Biochemistry
sp. ATCC 39484 was reported to prefer hydrophobic aromaticsubstrates [44]. Upon binding the substrate, the activity in theenzyme was increased. This suggests that the binding of anaromatic moiety in the active site triggers a conformationalchange that leads to enzyme association and increased activity.All these enzymes prefer aromatic substrates substituted withmethyl or halogen groups at the meta- and para-position, withpoor or no activity with ortho-substitutions [32,44] due to stericconflicts. The only atom tolerated at this position is fluorine,as reported for the nitrilase from the bacterium Norcadia sp.(Rhodochrous) NCIMB 11215 [57].
3.4. Aliphatic nitrilasesAliphatic nitrilases from microorganisms have been purifiedand characterized with different substrate specificity. Thenitrilase from crotonitrile-induced cells of R. rhodochrousK22 has high activity with ACN and glutaronitrile [32]. Thenitrilase from Acinetobacter sp. AK226 showed high activ-ity with benzonitrile and maximum activity with ACN [66].The rate of hydrolysis of 3-phenylpropionitrile versus that ofbenzonitrile is boosted by a factor of 270 in the recombinantAtNIT1 nitrilase of A. thaliana. The AtNIT1 nitrilase prefersaliphatic nitriles and showed increased hydrolysis with in-creasing size of the substrate. This enzyme also produced 95%amide product upon hydrolysis of 3-nitroacrylonitrile. Further,substrate analysis showed that IAN was not the preferredsubstrate for AtNIT1, 2, 3, and 4 nitrilases [78]. The nitri-lases from Acidovorax facilis 72W [133] and Synechocystis sp.strain PCC6803 [58] were reported to hydrolyze fumarodini-trile (trans-1,2-dicyanoethene). Particularly, the enzyme fromthe cyanobacterium Synechocystis sp. strain PCC6803 had120 times superior relative activity with fumarodinitrile com-pared with that monitored with benzonitrile. It also had higheractivity with some aromatic, heterocylic, and aliphatic nitriles.Addition of various concentrations of methanol (0%–20%), anorganic solvent, resulted in increased conversion rates for longchain hydrophobic aliphatic nitriles. Other enzymes in thisclass include those from Comamonas testosterone [25] andPseudomonas sp. S1 [73]. The nitrilase from Pseudomonas sp.S1 was reported to hydrolyze cyanide (KCN) to formic acid inaddition to aliphatic and some aromatic nitriles [74].
3.5. Bromoxynil-specific nitrilaseThe nitrilase of K. pneumoniae subsp. ozaenae is highly spe-cific for the herbicide bromoxynil. The native and recom-binant enzymes could completely convert 3, 5-dibromo-4-hydroxybenzonitrile (bromoxynil) to 3, 5-dibromo-4-hydroxybenzoic acid and use the liberated ammonia as thesole nitrogen source [60]. This enzyme shows no activity atall with benzonitrile but superior activity with bromoxynil,chloroxynil, and ioxynil [68]. The nitrilase of Norcadia sp.NCIMB 11215 was reported to have little but detectable activitywith bromoxynil and ioxynil [57].
3.6. ArylacetonitrilasesThese enzymes comprise the fourth class that does notshow activity with benzonitrile. The nitrilase from A. fae-calis ATCC 8750, which was induced with n-butyronitrile,showed highest activity with p-aminobenzylcyanide [133].This enzyme is enantioselective and produces R-(−)-mandelicacid from mandelonitrile or benzaldehyde and HCN [79]. Thenitrilase from A. faecalis JM3 is reported to hydrolyze aryl-cetonitriles such as 2-thiophenacetonitrile, p-tolyacetonitrile,p-fluorobenzylcyanide, and 3-pyridylacetonitrile with high ac-tivity when p-chlorobenzylcyanide is used as substrate [42].The nitrilase from P. fluorescens DSM 7155 was induced whenphenylacetonitrile was used as the sole carbon source [73].It showed high activity with phenylacetonitrile, but no activ-ity with benzonitrile. The bulk of nitrilases did not produceamide by-product. The arylacetonitrilase from P. fluorescensDSM 7155 produced 3%–5% of the corresponding amideby nitrile hydrolysis. Kiziak et al. [74] purified a recombi-nant enantioselective nitrilase from P. fluorescens EBC191.This enzyme exhibited high activity with para-substitutedphenylyacetonitriles and hydrolyzed heterocyclic and bicyclicarylacetonitriles. The enzyme converted (R, S)-mandelonitrileto R-mandelic acid with an enantiomeric excess (ee) of 31%,whereas 2-phenylpropionitrile was converted to the corre-sponding S-acids and amides with an ee of 65%. The nitrilasealso produced 43% and 28% amide by-products when racemicO-acetylmandelonitrile and (R)–O-acetylmandelonitrile wereused as substrates, respectively. This nitrilase shares 47%amino-acid sequence identity with the arylacetonitrilase fromA. faecalis JM3 and both have very little activity with benzoni-trile and aliphatic nitriles.
3.7. Cyanide dihydratases and cyanide hydratasesCyanide dihydratases and cyanide hydratases are specific forcyanide (HCN) and their products upon hydrolysis are formateand formamide, respectively. The cyanide dihydratase fromA. xylosoxidans subsp. denitrificans [135] was tested with ace-tonitrile, benzonitrile, and propionitrile, which hydrolyzed onlycyanide. Other cyanide dihydratases have also been identifiedin B. pumilus C1 [47], P. fluoroscens NCIMB 11764 [136], andP. stutzeri AK61 [137]. The filamentous fungus Fusarium later-itium produced cyanide hydratase with cyanide as an inducer.This enzyme showed 3,000-fold activity toward KCN as com-pared with benzonitrile [138]. The cyanide hydratases isolatedfrom F. oxysporum N-10 [139] and F. solani [140] hydrolyzedtetracyanonickelate (II) –K2[Ni(CN)4]; TCN˝, a metal–cyanocomplex. The cyanide hydratase from F. oxysporum N-10 had2,330 times more relative activity with KCN than with TCN. Noactivity was detected with benzonitrile.
Cyanide dihydratases are enzymes of the nitrilase super-family, which hydrolyzes cyanide (free form or metal cyanidecomplexes) to formate and ammonia. These enzymes havebeen identified in A. xylosoxidans subsp. denitrificans [45],B. pumillus C1[43, 44], P. fluorescens NCIMB 11764 [45], andP. stutzeri AK61[46]. The enzyme from A. xylosoxidans subsp.
468 Nitrile-Converting Enzymes

denitrificans [47] was induced by cyanide. None of these en-zymes was reported to produce formamide in these reactions.Cyanide hydratase, which hydrolyzes cyanide to formamide,was first identified in the fungus Stemphylium loti, a pathogenof cyanogenic plants [48]. This enzyme is not related to themetal-containing nitrile hydratases, which have no relationwith the nitrilase superfamily. The cyanide hydratases includeenzymes from Gloeoecercospora sorghi [49,88,89], F. lateritium[90], F. solani [91], F. solani IMI 196840 [92], and Leptosphaeriamaculans [93].
4. Enzyme Structure and StructuralHomologyThere are four atomic structures of distant homologs in thenitrilase superfamily. These include the NitFhit protein (PDBID, 1EMS) from Caenorhabditis elegans [105], N-carbamyl-d-amino acid amidohydrolase (DCase or D-NCAase; PDB ID,1ERZ) from Agrobacterium [63, 80, 141], Nit3 CN hydrolase(PDB ID, 1F89) from yeast [142], and the hypothetical proteinPh0642 (PDB ID, 1J31) from Pyrococcus horikoshii [143].Structures of 1EMS and 1ERZ show that these enzymes formtetramers with 222 symmetry. It has two contact surfacesbetween the tetramers that are almost perpendicular to eachother: a larger contact surface, referred to as the A surface,and a smaller contact surface, B. The A surface of 1EMS ismade up of two α-helices and a length of β-sheet, whereasthat of 1ERZ consists of three α-helices. In both structures,surface B is made up of two antiparallel β-sheets, which areexternally located next to the central core of β-sheets in eachmonomer. The worm NitFhit structure is a fusion protein of anitrilase homolog (Nit) and Fhit, a protein that plays a role inapoptosis. The structures of 1F89 and 1J31 share a conservedcatalytic cys-glu-lys triad with 1EMS, 1ERZ, and indeed, withother members in the nitrilase superfamily. These structuresresulted from efforts of structural genomics and their functionsare unidentified. The nitrilase atomic homologs form dimersacross a conserved twofold symmetric interface, which hasbeen called the A surface [144]. The A surface in 1F89 is heldtogether by salt bridges and hydrogen bonds comparable tothose observed in 1EMS and 1ERZ.
Nitrilases and amidases are reported to have a widesubstrate range, which may be because of the shape andvolume of the active site cavity. The volume in 1F89 is largercompared with that of 1ERZ [142]. This was not found in theNitFhit structure because of a mercury atom, which was boundto the active cysteine and might have altered the geometryof the active site [145]. The structure of the amidase, namelyN-carbamoyl-d-amino acid amidohydrolase (D-NCAase) fromAgrobacterium radiobacter, which was solved with boundsubstrate, revealed that there were limited conformationalchanges in structure upon binding the substrate [141]. Thisenzyme belongs to the nitrilase superfamily and allocates aparallel fold with the NitFhit structure. Structural analysis ofits free and substrate-bound forms revealed residues that were
directly bound to the substrate within a large pocket, whereasmodeling studies concluded that this enzyme has a preferencefor bulky substrates with long hydrophobic chains.
Sequence alignment of the nitrilase protein sequencesfrom P. stutzeri AK61, B. pumilus C1, and G. sorghi againstsolved structures using GenTHREADER showed that they sharea similar fold [146]. A dimer model of the cyanide dihydratasefrom P. stutzeri AK61 based on the solved structures wasused to interpret a low-resolution map. This revealed a left-handed 14-subunit spiral and highlighted residues involved instabilizing its quaternary structure. Two major insertions inits sequence relative to those of the Nit domain of 1EMS and1ERZ are also proposed to contribute to spiral formation. Theregion of these interactions is referred to as the C surface and isabsent in the crystal structures. The nitrilase atomic homologsexist as dimers or tetramers, whereas microbial nitrilases existas higher oligomers. The number of subunits required to formone turn of the spiral is 10, and it is suggested that this wouldbe the smallest microbial nitrilase [144].
There is mounting substantiation that formation of thespiral leads to commencement of the enzyme. A pH-dependenttransition from a terminating spiral at pH 8 to a long helix at pH5.4 was observed in the cyanide dihydratase of B. pumilus C1[147]. Interestingly, an increase in enzymatic activity of about10% is also observed at pH 6, whereas no increase in activitywas observed in the enzymes from G. sorghi and P. stutzeriAK61 [148]. The enzymes from G. sorghi and P. stutzeriAK61 do not undergo a structural transition either. In thecase of Rhodococcal nitrilases, however, the nitrilase fromR. rhodochrous J1 showed that the dimers are inactive, but thehigher oligomers have activity [44,57,131]. Evidence suggestsinterplay between activation and oligomerization lays thefoundation for a detailed investigation into this phenomenon.
5. Mechanism of CatalysisMost nitrilases exhibit maximum activity at an optimal pH rangeof 7.5–8.5. The nitrilases of K. pneumoniae subsp. ozaenae andAcinetobacter sp. AK226 were reported to have maximumactivity at pH 9.2 [66,130]. The pH-dependent transition of thecyanide dihydratase from B. pumilus C1 takes place betweena short spiral form and a long regular helix [63]. The activityfor this enzyme was lost at a pH above 8.4, but the presence ofCr3+ and Tb3+ at a concentration of 200 μM restored activityidentical to that at its optimum pH [47].
Site-directed mutagenesis on the catalytic residues in thealiphatic amidase from P. aeruginosa led to inactivity andinstability of the enzyme [106]. This amidase shares a similarstructural and catalytic scaffold to the cracked structures of thenitrilase homologs. Crystal structures of the unbound modifiedbiocatalyst from A. radiobacter, namely N-carbamoyl-d-aminoacid amidohydrolase (DNCase or DCase) and also in complexwith the substrate N-carbamoyl-d-p-hydroxyphenylglycine,were solved at a resolution range of 2.0–2.40 A by molecularreplacement [141]. Circular dichroism studies showed that
Biotechnology and Applied Biochemistry 469

Biotechnology andApplied Biochemistry
mutation of the active Cys172 did not affect the conformationof the active site, secondary structure, or the stability ofthe enzyme. The substrate-bound mutants revealed that thecarbamoyl group of the substrate is close to the thiol groupof cysteine. Further evidence of the role of the active cysteineresidue in catalysis is provided by the effect of inhibitorson enzyme activity. Thiol-complexing agents inactivated thenitrilase of P. fluoroescens DSM 7155, whereas thiol-reducingagents enhanced its activity [149]. Thiol-reducing agents werealso observed to enhance activity in the enzymes of A. faecalisJM3 [133] and R. rhodochrous J1 [107].
Crystal structures of 1EMS [145], 1ERZ [80], and 1F89[142] suggest that catalytic residues are located at the bottomof a pocket and are pointing toward its center [90]. In thesestructures, the active site is located at the bottom of this pocket.These structures also suggest that the residues located in theloops above this pocket play a role in determining specificity[106, 141]. Other studies of the nitrilases of A. faecalis JM3[112], P. stutzeri AK61 [137], and R. rhodochrous J1 [28]using far-UV circular dichroism, kinetic, and molecular massanalyses concluded that there was no significant structuralconformational change upon substitution of the active cysteinewith alanine or serine. However, a study concluded thatmodification of the catalytic glutamate or lysine residuesinduced conformational changes as well as enzyme instability[102].
As early as 1964, a reaction mechanism for hydrolysisof ricinine by a nitrilase of Pseudomonas species had beenproposed [38]. Mechanistic studies of the nitrilase of Rhodococ-cus ATCC 39484 indicated that the reaction proceeds throughcovalent intermediates [44]. Based on the results and knownchemical reactions between thiols and nitriles, the thiol group ofthe cysteine initiates a nucleophilic attack on the nitrile carbonfollowed by protonation of the nitrogen to form a thioimidate[43]. Subsequent two-step hydrolysis of the thioimidate pro-duces the corresponding acid and ammonia. Using ion-spraymass spectroscopy, a covalent intermediate whose mass wasconsistent with either the thioimidate or acylenzyme was de-tected [147]. A modification of the earliest nitrilase reactionmechanism has been proposed [150], in which the tetrahedralintermediate is shown to break down reversibly to producean amide instead of an acid product via the usual route. Thiscorrelates with the observation that some nitrilases producedsmall amounts of amide products with selected substrates.Based on the hydrolysis of different substrates to produceamide or acid products by AtNIT1 nitrilase from A. thaliana,the structural relationship between the type of substrateand amide formation has been postulated [128]. The ratioof acid to amide produced is variable and depends on themicrobial species, substrate structure, incubation time, and theage of the culture batch [147]. An amidase model built accord-ing to the NitFhit structure showed that the catalytic residueswere not only conserved but also aligned well with those of1EMS [106]. Furthermore, equivalent distances between the
catalytic residues from both structures were also observed(Figs. 3 and 4).
6. ImmobilizationEntrapment in hydrogels such as calcium alginate [151, 152],barium alginate [153], pectinate, k-carrageenan, and poly-acrylamide was used for immobilization of nitrile-convertingenzymes [154]. Some of the immobilized preparations showeda marked thermal or chemical stability. Whole cells of Bacillusspecies immobilized in alginate transformed aliphatic nitrilesand dinitriles. This offers advantages like reuse of the biocat-alyst, hence, continuous operation. Immobilized whole cells ofB. pallidus in calcium alginate were found to be more resistantto high concentrations of 3-cyanopyridine and nicotinic acidthan those of free cells [155]. Similarly, polyacrylnitrile immo-bilization of Rhodococcus and A. facilis improved their nitrilehydrolyzing activity during production of acrylic acid [154]and 4-cyanopentanoic acid ammonium salt correspondingly.The alginate provided some thermal stabilization for enzymedurability as monitored with immobilized cells of P. thermophilaand R. rhodococcus J1. Alginate beads containing cells of im-mobilized E. coli could be frequently used for three consecutivebatches and 85% of the initial enzyme activity could be pre-served in second and third batches [156]. It was also notedthat entrapped cells of Candida guillermondi metabolized evensubstrates that were not altered by free cells of the samemicroorganism (cyclopentanecabonitirile, benzonitrile, andbenzamide) [153]. In contrast, it was observed that sometimesimmobilization was injurious to the biocatalyst activity, possiblybecause of imperfect mass transfer across the gel barrier andperhaps also due to a limited oxygen supply to the cells [151].For instance, dried and reswollen polyvinyl alcohol beads withwhole cells of Rhodococcus sp. strain showed the same ac-tivity as nondried beads but amidase activity was augmentedon drying. Freeze-dried and lyophilized cells result in steadyand effortless use as biocatalysts and are more resistant tomicrobial contamination [157].
7. CloningThe first nitrilase gene was cloned in E. coli, encoding abromoxynil-degrading activity from K. penumoniae ozaenae[158]. Also, nitrilase is the first cloned plant enzyme that isproficient in converting an endogenous substrate indole-3-acetonitrile to the plant growth promoting hormone IAA [159]but also shows the ample range of other nitriles. Several othernitrilase genes from various other sources have been cloned likenitA gene (750 bp) from Rhodococcus in E. coli plasmid pNJ1,A. faecalis JM3 in E. coli [28, 108], and Gordan terrae MA-1in E. coli [160]. The nitrilase gene AtNIT1 from Arabidopsisthalina was overexpressed in E. coli. This recombinant AtNIT1has properties comparable to that of the native enzyme and thenitrilase from Brassica napus. Similarly, various NHase, H-,
470 Nitrile-Converting Enzymes
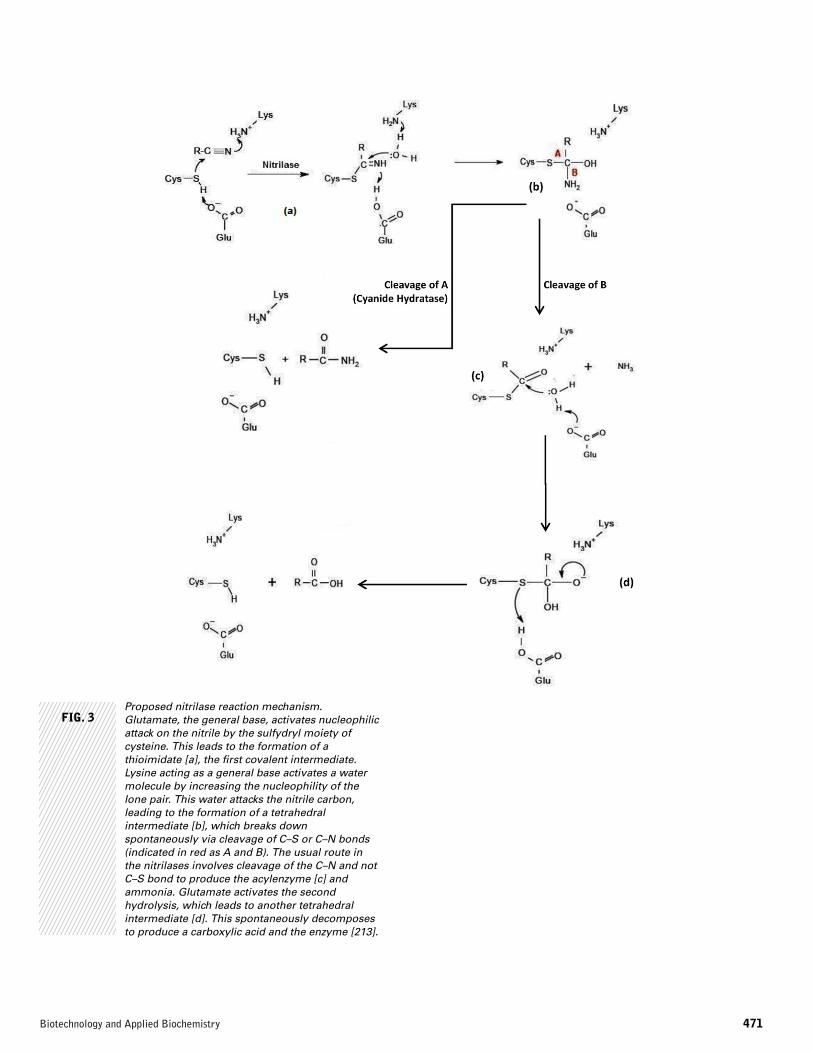
FIG. 3Proposed nitrilase reaction mechanism.Glutamate, the general base, activates nucleophilicattack on the nitrile by the sulfydryl moiety ofcysteine. This leads to the formation of athioimidate [a], the first covalent intermediate.Lysine acting as a general base activates a watermolecule by increasing the nucleophility of thelone pair. This water attacks the nitrile carbon,leading to the formation of a tetrahedralintermediate [b], which breaks downspontaneously via cleavage of C–S or C–N bonds(indicated in red as A and B). The usual route inthe nitrilases involves cleavage of the C–N and notC–S bond to produce the acylenzyme [c] andammonia. Glutamate activates the secondhydrolysis, which leads to another tetrahedralintermediate [d]. This spontaneously decomposesto produce a carboxylic acid and the enzyme [213].
Biotechnology and Applied Biochemistry 471
Biotechnology andApplied Biochemistry
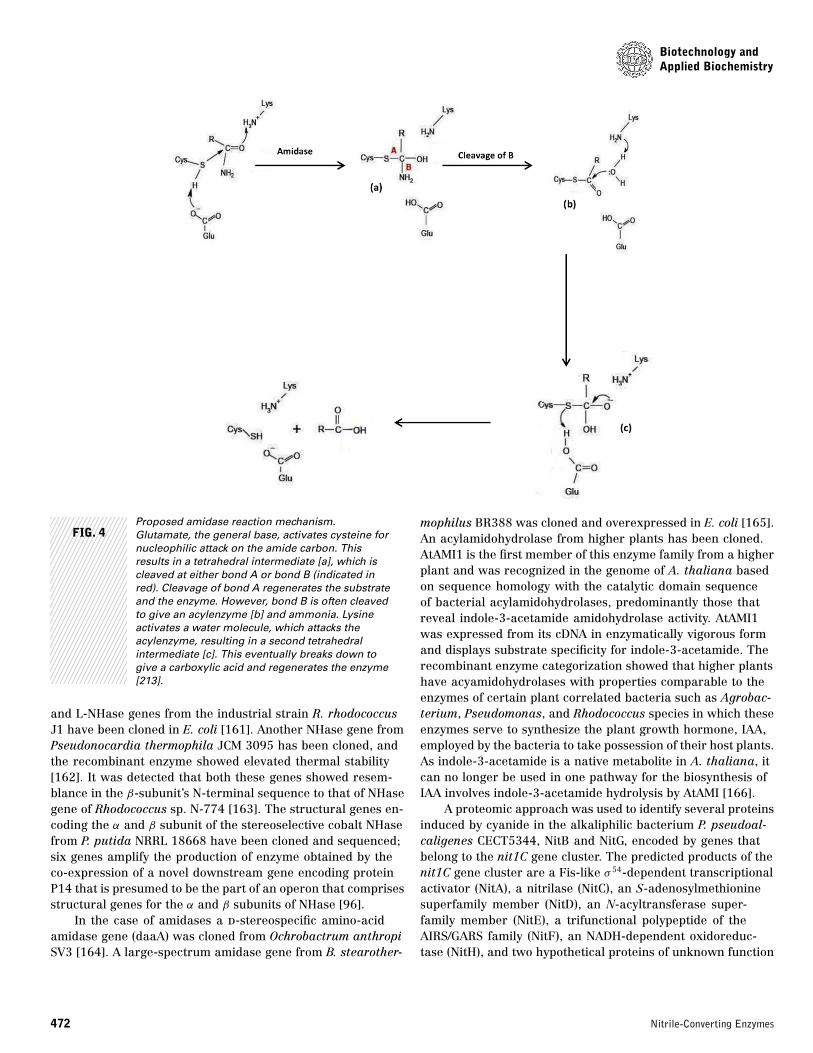
FIG. 4Proposed amidase reaction mechanism.Glutamate, the general base, activates cysteine fornucleophilic attack on the amide carbon. Thisresults in a tetrahedral intermediate [a], which iscleaved at either bond A or bond B (indicated inred). Cleavage of bond A regenerates the substrateand the enzyme. However, bond B is often cleavedto give an acylenzyme [b] and ammonia. Lysineactivates a water molecule, which attacks theacylenzyme, resulting in a second tetrahedralintermediate [c]. This eventually breaks down togive a carboxylic acid and regenerates the enzyme[213].
and L-NHase genes from the industrial strain R. rhodococcusJ1 have been cloned in E. coli [161]. Another NHase gene fromPseudonocardia thermophila JCM 3095 has been cloned, andthe recombinant enzyme showed elevated thermal stability[162]. It was detected that both these genes showed resem-blance in the β-subunit’s N-terminal sequence to that of NHasegene of Rhodococcus sp. N-774 [163]. The structural genes en-coding the α and β subunit of the stereoselective cobalt NHasefrom P. putida NRRL 18668 have been cloned and sequenced;six genes amplify the production of enzyme obtained by theco-expression of a novel downstream gene encoding proteinP14 that is presumed to be the part of an operon that comprisesstructural genes for the α and β subunits of NHase [96].
In the case of amidases a d-stereospecific amino-acidamidase gene (daaA) was cloned from Ochrobactrum anthropiSV3 [164]. A large-spectrum amidase gene from B. stearother-
mophilus BR388 was cloned and overexpressed in E. coli [165].An acylamidohydrolase from higher plants has been cloned.AtAMI1 is the first member of this enzyme family from a higherplant and was recognized in the genome of A. thaliana basedon sequence homology with the catalytic domain sequenceof bacterial acylamidohydrolases, predominantly those thatreveal indole-3-acetamide amidohydrolase activity. AtAMI1was expressed from its cDNA in enzymatically vigorous formand displays substrate specificity for indole-3-acetamide. Therecombinant enzyme categorization showed that higher plantshave acyamidohydrolases with properties comparable to theenzymes of certain plant correlated bacteria such as Agrobac-terium, Pseudomonas, and Rhodococcus species in which theseenzymes serve to synthesize the plant growth hormone, IAA,employed by the bacteria to take possession of their host plants.As indole-3-acetamide is a native metabolite in A. thaliana, itcan no longer be used in one pathway for the biosynthesis ofIAA involves indole-3-acetamide hydrolysis by AtAMI [166].
A proteomic approach was used to identify several proteinsinduced by cyanide in the alkaliphilic bacterium P. pseudoal-caligenes CECT5344, NitB and NitG, encoded by genes thatbelong to the nit1C gene cluster. The predicted products of thenit1C gene cluster are a Fis-like σ 54-dependent transcriptionalactivator (NitA), a nitrilase (NitC), an S-adenosylmethioninesuperfamily member (NitD), an N-acyltransferase super-family member (NitE), a trifunctional polypeptide of theAIRS/GARS family (NitF), an NADH-dependent oxidoreduc-tase (NitH), and two hypothetical proteins of unknown function
472 Nitrile-Converting Enzymes

(NitB and NitG). RT-PCR analysis suggested that nitBCDEFGHgenes were cotranscribed, whereas the regulatory nitA genewas divergently transcribed. The wild-type strain CECT5344showed a nitrilase activity that allows growth on cyanide or hy-droxynitriles. Very recently, nitrilase gene of R. rhodochrous J1was expressed in E. coli using the expression vector, pKK223–3.The recombinant E. coli JM109 cells were able to hydrolyzeenantioselectively 2-methyl-2-propylmalononitrile to form (S)-2-cyano-2-methylpentanoic acid (CMPA) with 96% ee underoptimized conditions; 80 g (S)-CMPA L−1 was produced with amolar yield of 97% at 30◦C after 24 H without any by-products[167].
8. Biocatalytic Activity of Nitriles inAqueous Organic MediaMost of the nitriles are usually found to be hydrophobic innature; hence, organic cosolvents are usually added into thereaction medium restraining a nitrile-converting biocatalyst.Assorted medium alterations were devised and optimized toboost the bioavailability of the substrates for biocatalysts [91].It was shown that nitrile-converting enzymes in both unfinishedand purified forms could be used with a high percentage oforganic solvents in the medium. Cell extracts with a nitrilaseactivity were found to be functional in n-alkane (C-8 or C-16)aqueous biphasic mixtures and in monophasic water saturatedC-6-C-11 n-alkanols [74]. Whereas nitrile hydratase purifiedfrom R. rhodococcus was found to be resistant to alcoholsholding 89–100% of its original activity, it was also noted thatsubstantial activity was present with 50% (v/v) of acetone, 1,4-dioxane, N,N’-dimethylformamide, tetrahydrofuran, dimethylsulfoxide, and ethylene glycol. The remarkable activity of theenzyme to these organic solvents suggests its multifacetedquaternary structure [91].
9. Biotechnological and IndustrialApplicationsBiocatalysis has long been used by biotechnologists as an areawith greater potential for chemical synthesis over conventionalchemical processes [15]. Nitrile-converting enzymes, princi-pally nitrilases, nitrile hydratases, and amidases, are usedin the synthesis of pharmaceuticals and commodity chemi-cals. These versatile biocatalysts have latent applications indiverse fields counting synthetic biocatalysis and bioreme-diation. They are also very important intermediates in theorganic synthesis of amines, amidines, carboxylic acids, es-ters, aldehydes, ketones, and heterocyclic compounds [4].Some nitrilases have also been successfully applied to prac-tical production in food industries, chemical manufacturing,pharmaceutical processes, wastewater treatment, and textileindustries [52].
9.1. Synthesis of chemicalsSeveral organic compounds, including p-aminobenzoic acid,acrylic acid, nicotinic acid, pyrazinoic acid, and IAA, arecommercially produced through the biotransformation of ni-triles using nitrile hydratases and amidase activities (Table 2).Nitrile-converting enzymes can also be used for the manufac-ture of nicotinamide and acrylamide [197]. In recent years,this research resulted in the establishment of a 4,000-ton/yearscale nicotinamide production facility by Lonza AG under li-cense from Mitsubishi Rayonco. This process makes use ofan equivalent biocatalyst for the manufacture of acrylamide,that is, Rhodococcus rhodococcus J1 immobilized in polyacry-lamide. Conversion of 3-cyanopyridine to nicotinamide as wellas nicotinic acid is obtained by using R. rhodococcus J1. A ther-mophilic strain B. pallidus has been reported as a biocatalystfor direct hydrolysis of 3-cyanopyridine into nicotinic acid andexecutes this process at 50–60◦C [198]. NHase has been widelystudied because it is used as a biocatalyst in various chemicalindustries, that is, the industrial production of acrylamide,nicotinamide, and 5-cyanaovaleramide [4]. Another essentialfact of nitrile biocatalysis is the capability of these enzymes tocarry out stereoselective transformations. The optically activecompounds can have a substantial effect on the market ofhigh-value pharmaceuticals (nonsteroidal anti-inflammatorydrugs), agricultural chemicals, and ferroelectric crystals [60].
Petite aliphatic nitriles are usually voluntarily hydrolyzedby the nitrile hydratase/amidase systems. The range of biocata-lysts useful for the enzymatic hydrolysis of these compounds hasbeen extended. Rhodococcus sp. utilized saturated and unsatu-rated aliphatic nitriles as substrates as well as their derivativessubstituted with aromatic moieties [185]. Nitrilases and nitrilehydratase, the enzymes that hydrate nitriles into amides, areusually produced in bacteria along with an amidase activityproviding eventually carboxylic acid and ammonia [199]. Allthese nitrile-metabolizing enzymes hydrolyze a number ofstructurally diverse nitriles. Several commercially significantorganic compounds such as p-aminobenzoic acid, benzamide,acrylamide, nicotinic acid, pyrazinoic acid, thiophenamide, andso on have been prepared from the corresponding nitriles usingmicrobial cells.
The enzymatic regioselective hydrolysis of dinitriles isa process that can rarely be accomplished chemically in asingle step [175]; such exclusive biocatalysis affords usefulintermediates with diverse functional groups. The productsof regioselective hydrolysis of di- and tri-nitriles were ob-tained using whole cells of A. facilis ATCC 55746. The ther-mostable nitrilase produced was used for the regioselectiveconversion of a single nitrile moiety in 2-methylglutaronitrile,2-methyleneglutaronitrile, 2-ethylsuccinonitrile, amlononi-trile, succinonitrile, and glutaronitrile [180, 200]. Heating at50◦C weakens another nitrile-hydrolyzing activity that ledto the formation of dicarboxylic acid monoamides and di-carboxylic acids. The hydrolysis of 2-ethylsuccinonitrile into3-cyanopentanoic acid ammonium salt was catalyzed by the
Biotechnology and Applied Biochemistry 473
Biotechnology andApplied Biochemistry
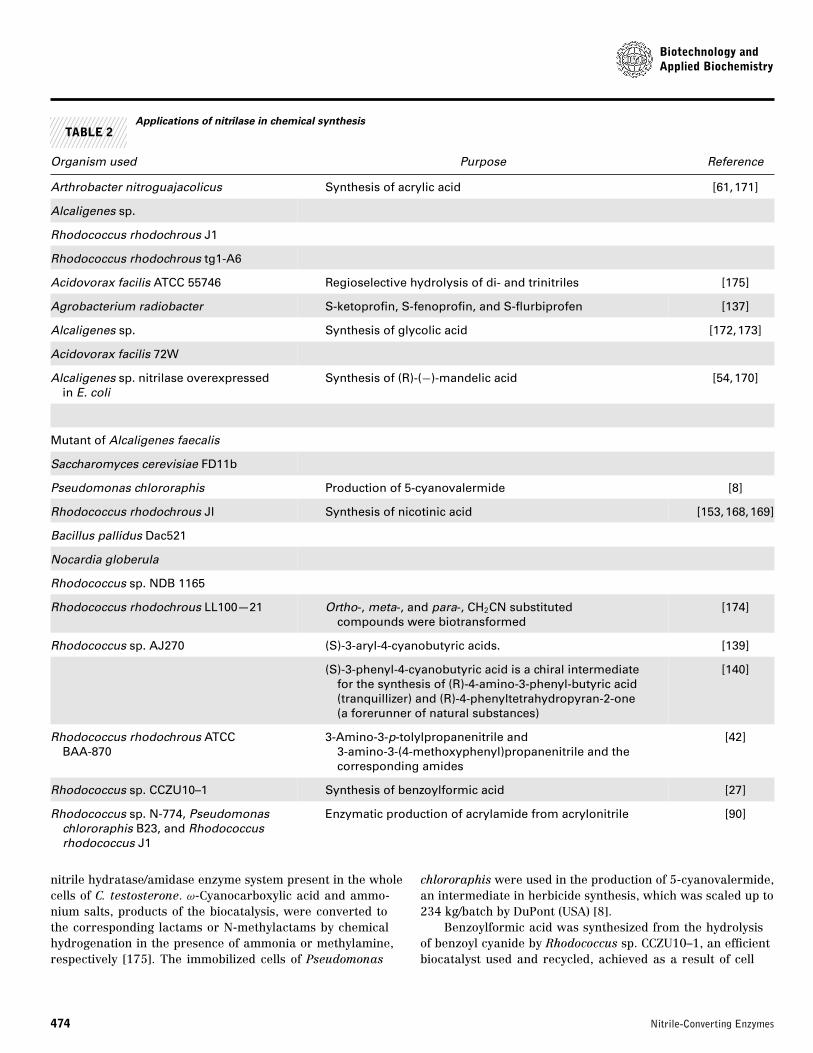
TABLE 2Applications of nitrilase in chemical synthesis
Organism used Purpose Reference
Arthrobacter nitroguajacolicus Synthesis of acrylic acid [61,171]
Alcaligenes sp.
Rhodococcus rhodochrous J1
Rhodococcus rhodochrous tg1-A6
Acidovorax facilis ATCC 55746 Regioselective hydrolysis of di- and trinitriles [175]
Agrobacterium radiobacter S-ketoprofin, S-fenoprofin, and S-flurbiprofen [137]
Alcaligenes sp. Synthesis of glycolic acid [172,173]
Acidovorax facilis 72W
Alcaligenes sp. nitrilase overexpressedin E. coli
Synthesis of (R)-(−)-mandelic acid [54,170]
Mutant of Alcaligenes faecalis
Saccharomyces cerevisiae FD11b
Pseudomonas chlororaphis Production of 5-cyanovalermide [8]
Rhodococcus rhodochrous Jl Synthesis of nicotinic acid [153,168,169]
Bacillus pallidus Dac521
Nocardia globerula
Rhodococcus sp. NDB 1165
Rhodococcus rhodochrous LL100—21 Ortho-, meta-, and para-, CH2CN substitutedcompounds were biotransformed
[174]
Rhodococcus sp. AJ270 (S)-3-aryl-4-cyanobutyric acids. [139]
(S)-3-phenyl-4-cyanobutyric acid is a chiral intermediatefor the synthesis of (R)-4-amino-3-phenyl-butyric acid(tranquillizer) and (R)-4-phenyltetrahydropyran-2-one(a forerunner of natural substances)
[140]
Rhodococcus rhodochrous ATCCBAA-870
3-Amino-3-p-tolylpropanenitrile and3-amino-3-(4-methoxyphenyl)propanenitrile and thecorresponding amides
[42]
Rhodococcus sp. CCZU10–1 Synthesis of benzoylformic acid [27]
Rhodococcus sp. N-774, Pseudomonaschlororaphis B23, and Rhodococcusrhodococcus J1
Enzymatic production of acrylamide from acrylonitrile [90]
nitrile hydratase/amidase enzyme system present in the wholecells of C. testosterone. ω-Cyanocarboxylic acid and ammo-nium salts, products of the biocatalysis, were converted tothe corresponding lactams or N-methylactams by chemicalhydrogenation in the presence of ammonia or methylamine,respectively [175]. The immobilized cells of Pseudomonas
chlororaphis were used in the production of 5-cyanovalermide,an intermediate in herbicide synthesis, which was scaled up to234 kg/batch by DuPont (USA) [8].
Benzoylformic acid was synthesized from the hydrolysisof benzoyl cyanide by Rhodococcus sp. CCZU10–1, an efficientbiocatalyst used and recycled, achieved as a result of cell
474 Nitrile-Converting Enzymes

immobilization in calcium alginate, with a product-to-biocatalyst ratio of 14.26 g benzoylformic acid g−1 dry cellweight cell after 20 cycles of repeated use [27]. A range ofβ-aminonitriles (3-amino-3-phenylpropanenitrile and deriva-tives) were synthesized by reaction of various benzonitrileswith acetonitrile and subsequent reduction of the resultingACN products. These compounds were hydrolyzed to the cor-responding amides using the nitrile biocatalytic activity ofRhodococcus rhodochrous ATCC BAA-870. Results showed thatthe nitrile hydratase enzyme was enantioselective for thesecompounds, in particular, 3-amino-3-p-tolylpropanenitrile and3-amino-3-(4-methoxyphenyl) propanenitrile and the corre-sponding amides [201]. Nitrilases have attracted increasingattention as catalysts for organic chemical processing be-cause of the mild conditions, quantitative yields, absence ofby-products, and in some cases, enantio- or regioselectivitythat results from the reactions.
9.2. Bioremediation and biodegradationSynthetic nitrile compounds are widespread in the environmentin the form of industrial waste. Most of them are toxic, muta-genic, and carcinogenic in nature [11]; thus, there is a need tocontrol their discharge into the environment. In nature, twodiscrete pathways for the degradation of nitriles to carboxylicacids exist. Nitrilase (Nit) catalyzes the direct hydrolysis ofnitrile to carboxylic acid [4], whereas nitrile hydratase (NHase)catalyzes the hydration to nitrile to amide [202], which is thenhydrolyzed to carboxylic acid by amidase. Chemical industriesexploit diverse nitrile compounds for manufacturing of varioussynthetic compounds. In comparison with conventional meth-ods of nitrile conversion to acids and amides requiring high pHand temperature, microbial conversion of nitriles is attractivebecause of its ability to carry out the reactions in a “greener”manner. Also, its capability to carry out chemo-, regio-, andenantioselective transformation makes it a useful tool for in-dustrial catalysis [4]. Thorough research into the applicabilityof nitrile-converting enzymes in synthetic organic chemistrywas hooked up to the innovation of the nitrile hydrates in theearly 1980s and to the revolutionary work of Yamada and Glazyas well as the Nitto Chemical Industry Company [89]. Thesegroups showed the enzymatic production of acrylamide fromACN using nitrile-hydrating whole cells of Rhodococcus sp.N-774, P. chlororaphis B23, and R. rhodococcus J1. This wasthe first study that made use of nitrile-converting biocatalystson an industrial scale [90]. The nitrile hydratases produced bybacteria employed in the industrial manufacture of acrylamideare most lengthily depicted. The amidase in whole cells ofBacillus sp. was found to be resourcefully restrained by ureaor chloroacetone [203]. In contrast to acrylamide production,the manufacture of acrylic acid requires microorganisms witha glut of amidase over nitrile hydratase activity or possessingnitrilase activity [199].
Besides the use of nitrile-hydrolyzing enzymes in biocatal-ysis, the exploration of microbial nitrile metabolism in thechemical and agricultural industries also had a remarkable
environmental impact as the nitriles had different degreesof toxic influence [180]. The flexible biocatalytic nature andapplications of nitrile-converting enzymes are now increasinglydistinguished for the production of numerous vital pharma-ceutical compounds and finer chemicals. With the advantageof their capability to abolish highly toxic nitriles, these nitrile-degrading enzymes play a significant role in defending theenvironment [4]. Nitriles can be degraded from the envi-ronmental contaminants by enzymatic reaction of nitrilase.This degradation is of great significance because most nitrilecompounds are highly toxic, carcinogenic, and mutagenic innature, and are considered harmful to human beings, animals,and plants [204]. Nitrilases are used in biotransformation anddegradation of xenobiotic compounds. Amidases are used in theconversion of acrylamide into acrylic acid and these are usedto produce polymers for application such as superabsorbents,dispersants, and flocculants (Table 3).
Prolonged exposure to nitrile herbicides (dichlobenhil andbromoxynil) results in weight loss, fever, vomiting, headache,and urinary problems [205]. Nitrile-metabolizing enzymescompetently degrade these cyano group containing herbicidesand prevent them from entering the food chain. A. radiobacter,a bromoxynil degrading soil bacterium, is used for the degra-dation of the herbicide under nonsterile batch and continuousconditions. A gene encoding the nitrile-degrading enzyme hasbeen cloned and used to raise herbicide-resistant plants [127].Bromoxynil-resistant transgenic plant resulting from the in-troduction of bromoxynil-specific nitrilase genes into tomatoor tobacco may be available in the market in the near future[195]. A recombinant E. coli strain displaying the dimericnitrilase from K. pneumoniae subsp. ozaenae (NitKp) on thecell surface was constructed using the autodisplay system.The whole-cell biocatalyst was also able to convert 3-fluoro-4-hydroxybenzonitrile and 3,5-dimethyl-4-hydroxybenzonitrileto the corresponding carboxylic acids [206].
Acrylamide, a neurotoxic monomer with all-embracingindustrial applications, was found to be degraded by the mi-croorganisms present in the sultry garden soil. Pseudomonassp. degraded high concentrations of acrylamide (4 g/L) toacrylic acid and ammonia; it could also act on other short chainamides like formamide and acetamide but not on acrylamideanalogs: methylacrylamide and N, N-methylene bisacrylamide[183]. A mixed culture of bacteria containing different nitrile-hydrolyzing enzymes (including NHase, nitrilase, amidase) thatmetabolize effluent containing ACN, fumaronitrile, succinoni-trile, and so on was grown in batch and continuous culture onthese components of waste [179]. The use of these specializedcultures of microorganisms to degrade toxic wastes could proveto be a competent method to the traditional activated sludgesystem. Nitrile-converting whole-cell biocatalysts proved to beuseful in degradation of ACN-butadiene and ACN butadienestyrene [207] and waste effluents from acryloanitrile manufac-ture [208]. For the biodegradation of starch-graft ACN, a strainof Bacillus cereus was isolated showing both extracellular de-polymerizing (α-amylase, peroxidase) activity and intracellular
Biotechnology and Applied Biochemistry 475

Biotechnology andApplied Biochemistry
TABLE 3Application of nitrilase in bioremediation and biodegradation
Organism used Purpose Reference
Actinobacteria Degradation of benzonitrile [176]
Rhodococcus rhodochrous PA-34,Rhodococcus sp.
Hydrolyze chloroxynil and bromoxynil completely, andpartially ioxynil
NDB 1165, and N. globerula NHB-2
Alcaligenes, Bacillus, Stemphylium, Fusarium,Pseudomonas, and Arthrobacter
Nitrile biodegradation [177]
Klebsiella pneumoniae subsp. ozaenae Degradation of bromoxynil to3,5-dibromo-4-hydroxybenzoic acid
[68,178]
Mixed cultures of the bacteria Metabolize effluent containing acrylonitrile,fumaronitrile, succinonitrile
[179]
Mixed cultures of the bacteria Regioselective conversion of a single nitrile moiety in2-methylglutaronitrile, 2-methyleneglutaronitrile,2-ethylsuccinonitrile, amlononitrile, succinonitrile,and glutaronitrile
[180]
Nitrilase bacteria Degradation of acetonitrile, acrylonitrile, succinonitrile,benzonitrile, dichlobenil, bromoxynil, and ioxynil
[52,181]
Nitrilase bacteria Degrade more than 80% of succinonitrile [182]
Pseudomonas sp. Degraded high concentrations of acrylamide [183]
Rhodococcus rhodochrous BX2) along withBacillus subtilis E2, E3, and N4
Acetonitrile-containing wastewater treatment [184]
Rhodococcus sp. Hydrolysis of petite aliphatic nitriles [185]
ACN-degrading activity. Whole cells of Pseudomonas sp. couldbe utilized for biodegradation of both organic and inorganiccyanides [78,151]. Propionitrile, which results in large amountsas a waste from the production of synthetic fibers, was con-verted into ammonium propionate by free or immobilized cellsof R. erythropolis [190]. An acetonitrile-degrading bacterium(R. rhodochrous BX2) along with Bacillus subtilis E2, E3, andN4 was established for acetonitrile-containing wastewatertreatment. Biofilm exhibited strong resistance to acetonitrileloading shock and displayed a typical spatial and structuralheterogeneity and completely depleted the initial concentrationof acetonitrile (800 mg L−1) within 24 hours in a moving-bed-biofilm reactor after operation for 30 days. This approachoffered a novel strategy for enhancing biological oxidation oftoxic pollutants in wastewater and confirms biofilm-formingbacteria can promote the immobilization of contaminant de-grading bacteria in biofilms and can subsequently improve thedegradation of contaminants in wastewater [184].
9.3. Surface modification of polymersACN hydration is also used to amplify the hydrophilicity of thepolyacrylic fibers used in textiles. The amide groups formed in
the fiber by the action of whole-cell biocatalyst Rhodococcusrhodocrous, Brevibacterium imperiale raised the effectivenessof the dying process [207, 209]. One of the most attractivefeatures of nitrile-metabolizing enzymes is their facility toselectivity convert one cyano group of a polynitrile, whichis virtually infeasible using conventional chemical methods.R. rhodococcus K22 catalyzes the conversion of adiponitrileto 5-cyanovaleric acid, which is intermediate in synthesis ofnylon-66 [191]. Ortho-, meta-, and para-, CH2CN substitutedcompounds were biotransformed using whole cell suspensionsof the bacterium R. rhodochrous LL100–21 [174]. Nitrile-converting enzyme met the demands for an efficient productionof (S)-mandelamide, S- and R-2-hydroxy acids using whole cellsof Gordona terrae MA-1[3, 210] and (S)-3-aryl-4-cyanobutyricacids [139]. (S)-3-phenyl-4-cyanobutyric acid is a chiral in-termediate for the synthesis of (R)-4-amino-3-phenyl-butyricacid (tranquillizer) and (R)-4-phenyltetrahydropyran-2-one (aforerunner of natural substances); these optically enrichedproducts were prepared by the use of whole cells of Rhodococ-cus sp. AJ270 [140]. The nitrilase catalyzed process for themanufacture of mandelic acid has been commercialized ona multiton scale (BASF, D) [14]. The S-enantiomers for the
476 Nitrile-Converting Enzymes

TABLE 4Applications of nitrilase in surface modification of polymers
Organism used Purpose Reference
Brevibacterium imperiale CBS 49874,Corynebacterium nitrilophilusATCC21419
Surface modification through selective enzymatichydrolysis
[186]
Commercial nitrilase Direct enzymatic modification of –CN groups into –COOHon fibers
[187]
Escherichia coli BL21 (DE3)/pET-Nit Effectively modify the surface of PAN fibers and membrane [188]
Micrococcus luteus BST20 Use PAN polymers as sole carbon source [189]
Rhodococcus erythropolis Propionitrile degradation to ammonium propionate [190]
Rhodococcus rhodococcus K22 Catalyzes the conversion of adiponitrile to 5-cyanovalericacid, which is intermediate in synthesis of nylon-66
[191]
TABLE 5Pharmaceutical and medical applications of nitrilase
Organism used Purpose Reference
Alcaligenes faecalis JM3, A. faecalis ATCC8750, Alcaligenes sp. ECU0401,Pseudomonas putida, Microbacteriumparaoxydans, Microbacterium liquefaciens
Enzymatic hydrolysis of (R,S)-mandelonitrile into(R)-mandelic acid (general resolving agent,intermediate of the semisynthetic antibioticcefamandol)
[25,42,51,170,192,193]
Comamonas testosterone Hydrolysis of 2-ethylsuccinonitrile into3-cyanopentanoic acid ammonium salt
[175]
Rhodococcus sp. strain R312 Hydroxamic acids for tissue remodeling [194]
Rhodococcus sp. strain R312 Production of α-aminohydroxamic acids, syntheticsiderophores, acetohydroxamic acids
[195]
Sulfolobus solfataricus Renovation of phenylglycine nitrile tophenylgycine, a precursor for semisyntheticcephalosporins and penicillins
[196]
analogous acids as well as those of the other 2-substitutedarylacetic acid and the R-enantiomers of some 2-substitutedarylacetamide could be prepared in enantiopure forms due to ahighly enantioselective amidase present in the Rhodococcus sp.AJ270 strain [140]. Similarly, S-ketoprofin, S-fenoprofin, andS-flurbiprofen were obtained in, for example, >98 g using anA. radiobacter strain producing a reasonably S-selective nitrilehydratase and a stringently S-selective amidase. As previouslyshown, admirable enantioselctivity is natural to plentiful ami-dases, the production of which is linked to that of the nitrilehydratases in nitrile-converting microorganisms [92] (Table 4).
9.4. Pharmaceutical and medical applicationsNitrilases are also used in pharmaceutical and medical appli-cations (Table 5) for the conversion of nicotinamide to nicotinicacid, which proves to be a good additive in food industries aswell as in drugs to lower the level of cholesterol and triglyc-
erides. A new nitrile hydratase was obtained from the cysteineprotease papain by replacing Gln19 in its oxyanion hole witha glutamic acid residue. The obtained enzyme (Gln19 Glupapai), similar to the parent papain, is very stable, active inorganic solvents and at low pH, and specific for peptide-basedsubstrates. It proved useful for preparation of amidra zones[211] that serve as intermediates in the synthesis of key com-ponents of biologically active molecules. Perhaps no enzymehas such an impact on human health as penicillin amidase(PA, EC 3.5.1.11). Its use has facilitated the massive produc-tion of a large group of β-lactam antibiotics for the control ofbacterial infectious diseases. This enzyme is used to hydrolyzeeither benzyl-penicillin (penicillin G) or phenoxymethylpeni-cillin (penicillin V) to produce 6-aminopenicillanic acid (6-APA),the vital preliminary compound for the production of semisyn-thetic penicillins. The semisynthetic penicillin derivatives notonly demonstrate improved properties over penicillin G or V,
Biotechnology and Applied Biochemistry 477

Biotechnology andApplied Biochemistry
such as augmented stability, easier absorption, and fewer sideeffects but also symbolize a practical solution to the problem ofadaptive microbial resistance to antibiotics. The most commonmechanism of microbial resistance to β-lactam antibiotics isbased on a group of enzymes that hydrolyze the β-lactamring of penicillin known as β-lactamases. This type of resis-tance spreads swiftly in bacterial populations by transfer ofgenetic information among populations and natural selection.Consequently, the immense use of antibiotics soon rendersthem incompetent, and new antibiotics need to be prepared totrounce resistant forms. Cephalosporins, another chief group ofβ-lactam antibiotics, have been increasingly used in predilec-tion to penicillin derivatives. However, the recent introductionof certain β-lactamase inhibitors (clavan compounds) in β-lactam antibiotic preparation has reawakened attention in theuse of semisynthetic penicillin derivatives in medicine. Withits tactical value reinstated, PA persists in playing a key rolein the production of useful β-lactam antibiotics for the controlof infectious bacterial diseases [205]. All of the different ami-dases also reveal an acyl transfer activity in the presence ofhydroxylamine:
RCONH2+NH2OH ↔ RCONHOH + NH3
This kind of reaction was previously explained for the wide-spectrum amidase from Rhodococcus sp. strain R312 [65]. Thefinal reaction products (hydroxamic acids) are known to pro-duce high chelating properties. Some of them (particularlyα-aminohydroxamic acid derivatives) are effective inhibitorsof matrix metalloproteases, a family of zinc endopeptidasesinvolved in tissue remodeling [194]. Others (predominantlyα-aminohydroxamic acids, synthetic siderophores, acetohy-droxamic acids, etc.) have also been inspected as antihumanimmunodeficiency virus agents or antimalarial agents or havebeen counseled for treatment or ureaplasma infections andanemia [195]. Apart from medical applications, some hydrox-amic acids (chiefly polymerizable unsaturated hydroxamicacids and midchain or long-chain hydroxamic acids) have alsobeen extensively investigated in wastewater treatment andnuclear technology studies as a way to eradicate contaminat-ing metal ions. FAAH is the mammalian integral membraneenzyme responsible for the catabolism of the fatty acid amidefamily of endogenous signaling lipids [98]. Representative fattyacid amides comprise the endocannabinoid anandamide andthe sleep-inducing lipid oleamide. Fatty acid amides engender afascinating assortment of pharmacological effects in mammals,including sleep and analgesia, which indicates that FAAH mightserve as an attractive choice for pharmaceutical efforts aspiredat influencing endogenous pain and/or sleep-wake systems. Theclarification of FAAH catalytic and structural features wouldprovide an expensive foundation upon which to launch suchmedicinal chemistry endeavors. FAAH is the only differentiatedmammalian member of a large group of enzymes termed theAS family. These enzymes share a highly preserved linearsequence rich in serine and glycine residues that span roughly50 amino acids in length. Regardless of the presence of AS
enzymes in several kingdoms of life, their catalytic features andprobable relatedness to other classes of amidolytic enzymesremain inadequately understood [194].
A stereoselective NHase that renovates phenylglycinenitrile to phenylgycine, a precursor for semisyntheticcephalosporins and penicilins, has been extensively segre-gated [196]. For instance, the enzymatic hydrolysis of (R,S)-mandelonitrile into (R)-mandelic acid (general resolving agent,intermediate of the semisynthetic antibiotic cefamandol, etc.)was demonstrated with bacterial nitrilases from A. faecalisJM3 [42], A. faecalis ATCC 8750 [25], Alcaligenes sp. ECU0401[51, 170], Pseudomonas putida [192, 193], Microbacteriumparaoxydans, or Microbacterium liquefaciens [192], which arecharacterized by their high selectivity for (R)-mandelonitrile.
10. ConclusionThe wide applicability of nitrile-converting biocatalysts, namelynitrilases, in various fields like pharmaceuticals, medicine, andbioremediation, strongly suggests that they are a potentialtool for catalysis. With the advancement in techniques relatedto isolation, screening, and genetic engineering, it is possi-ble to isolate microorganisms with novel characteristics andenhanced thermostability. Current studies demonstrate thatnitrile-converting enzymes with new properties have beenisolated from bacteria and are used for organic synthesis;however, there is a need to screen nitrile-converting enzymesfrom eukaryotes. There is a strong possibility that eukary-otic nitrile-converting enzymes may exhibit wider substratespecificity, stereo-/regioslectivity, and stability; the changedproperties may be due to posttranslational modifications suchas glycosylation.
11. References[1] Martinkova, L., Uhnakova, B., Patek, M., Nesvera, J., and Kren, V. (2009)
Environ. Int. 35, 162–177.[2] Pollak, P., Romeder, G., Hagedorn, F., and Gelbke, H. P. (2002) Nitriles:
Ullmann’s Encyclopedia of Industrial Chemistry, Wiley-VCH, Weinheim.[3] Fleming, F. F., Yao, L., Ravikumar, P. C., Funk, L., and Shook, B. C. (2010) J.
Med. Chem. 53(22), 7902–7917.[4] Banerjee, A., Sharma, R., and Banerjee, U. C. (2002) Appl. Microbiol.
Biotechnol. 60, 30–44.[5] Asano, Y., Fujishiro, K., Tani, Y., and Yamada, H. (1982) Agric. Biol. Chem.
46, 1165–1174.[6] Fleming, F. F. (1999) Nat. Prod. Rep. 16, 597–606.[7] Fleming, F. F., Yao, L., Ravikumar, P. C., Funk, L., and Shook, B. C. (2010) J.
Med. Chem. 53(22), 7902–7917.[8] Bandyopadhyay, A. K., Nagasawa, T., Asano, Y., Fujishiro, K., Tani, Y., and
Yamada, H. (1986) Appl. Environ. Microbiol. 51, 302–306.[9] Hann, E. C., Eisenberg, A., Fager, S. K., Perkins, N. E., Gallagher, F. G.,
Cooper, S. M., Gavagan, J. E., Stieglitz, B., Hennessey, S. M., and DiCosimo,R. (1999) Bioorg. Med. Chem. 7, 2239–2245.
[10] Rubber World Magazine (2011) Global Nitrile Butadiene Rubber Market toExceed 645 Thousand Tons by 2017. Market report for the Rubber Industry.
[11] Pollak, P., Romender, G., Hagedorn, F., and Gelbke, H. P. (1991). In Ullman’sEncyclopedia of Industrial Chemistry, 5th ed., vol A17. (Elvers, B., Hawkins,S., Schulz, G., eds.), pp. 363–376, Wiley-VCH, Weinheim.
478 Nitrile-Converting Enzymes

[12] Solomonson, L. P. (1981). In Cyanide in Biology (Vannesland, B., Conn,E. E., Knowles, C. J., Westley, J., and Wissing, F., eds). pp. 11–28, AcademicPress, London.
[13] Khan, H. A., Alhomdia, A. S., and Arif, I. A. (2009) Toxicol. Sci. 109, 124–131.[14] Cadet, J. L., Braun, T., and Freed, W. J. (1987). Exp. Neurol. 96, 594–560.[15] Schmid, A., Dordick, J. S., Hauer, B., Kiener, A., Wubbolts, M., and Witholt,
B. (2001) Nature 409, 258–268.[16] Trott, S., Bauer, R., Knackmuss, H. J., and Stolz, A. (2001) Microbiology 147,
1815–1824.[17] Mahadevan, S., and Thimann, K. (1964) Arch. Biochem. Biophys. 107, 62–68.[18] Kobayashi, M., Fujiwara, Y., Goda, M., Komeda, H., and Shimizu, S. (1997)
Proc. Natl. Acad. Sci. USA 94, 11986–11991.[19] Kobayashi, M., Suzuki, T., Fujita, T., Masuda, M., and Shimizu, S. (1995) Proc.
Natl. Acad. Sci. USA 92(3), 714–718.[20] Bartling, D., Seedorf, M., Mithofer, A., and Weiler, E. W. (1992) Eur. J.
Biochem. 205(1), 417–424.[21] Bartling, D., Seedorf, M., Schimdt, R. C., and Weiler, E. W. (1994) Proc. Natl.
Acad. Sci. USA 91, 6021–6025.[22] Piotrowski, M., Schonfelder, S., and Weiler, E. W. (2001) J. Biol. Chem.
276(4), 2616–2621.[23] Langdahl, B. B., Bisp, P., and Ingvorsen, K. (1996) Microbiology 142, 145–154.[24] Bhalla, T. C., Miura, A., Wakamoto, A., Ohba, Y., and Furuhasi, K. (1992)
Appl. Microbiol. Biotechnol. 37, 184–190.[25] Yamamoto, K., Oishi, K., Kawakami, K., and Komatsu, K. (1990) Appl.
Environ. Microbiol. 57(10), 3028–3032.[26] Nagasawa, T., Wieser, M., Nakamura, T., Iwahara, H., Yoshida, T., and Gekko,
K. (2000) Eur. J. Biochem. 267, 138–144.[27] He, Y. C., Zhou, Q., Ma, C. L., Cai, Z. Q., Wang, L. Q., Zhao, X. Y., Chen, Q.,
Gao, D. Z., Zheng, M., Wang, X. D., Sun, Q., and Sun, Q. (2012) Bioresour.Technol. 115, 88–95.
[28] Kobayashi, M., Goda, M., and Shimizu, S. (1998) Biochem. Biophys. Res.Commun. 253, 662–68.
[29] Thimann, K. V., and Mahadevan, S. (1964) Arch. Biochem. Biophys. 105,133–141.
[30] Mahadevan, S. (1963) Arch. Biochem. Biophys. 100, 557–558.[31] Harper, D. B. (1977a) Biochem. J. 167, 685–692.[32] Kobayashi, M., Komeda, H., Yanaka, N., Nagasawa, T., and Yamada, H.
(1992) J. Biol. Chem. 267(29), 20746–20751.[33] Kaplan, O., Bezouska, K., Malandra, A., Vesela, A. B., Petrickova, A., Felsberg,
J., Rinagelova, A., Kren, V., and Martinkova, L. (2011) Biotechnol. Lett. 33,309–312.
[34] Petrickova, A., Vesela, A. B., Kaplan, O., Kubac, D., Uhnakova, B., Malandra,A., Felsberg, J., Rinagelova, A., Weyrauch, P., Kren, V., Bezouska, K., andMartinkova, L. (2012) Appl. Microbiol. Biotechnol. 93, 1553–1561.
[35] Strobel, G. A. (1966) J. Biol. Chem. 241, 2618–2621.[36] Hook, R. H., and Robinson, W. G. (1964) J. Biol. Chem. 239(12), 4263–4267.[37] Strobel, G. A. (1967) J. Biol. Chem. 242, 3465–3469.[38] Robinson, W. G., and Hook, R. H. (1964) J. Biol. Chem. 239, 4257–4262.[39] Shimizu, T., and Taguchi, H. (1969) Ferment. Technol. 47, 639–643.[40] Nagasawa, T., Kobayashi, M., and Yamada, H. (1988) Arch. Microbiol. 150,
89–94.[41] Nagasawa,T., Takeuchi, K., and Yamada, H. (1991) Eur. J. Biochem. 196(3),
581–589.[42] Nagasawa, T., Mauger, J., and Yamada, H. (1990) Eur. J. Biochem. 194,
765–772.[43] Stevenson, D. E., Feng, R., Dumas, F., Groleau, D., Mihoc, A., and Storer, A.
C. (1992) Biotechnol. Appl. Biochem. 15, 283–302.[44] Kobayashi, M., Yanaka, N., Nagasawa, T., and Yamada, H. (1990) J. Bacteriol.
172(9), 4807–4815.[45] Hann, E. C., Sigmund, A. E., Fager, S. K., Cooling, F. B., Gavagan, J. E.,
Bramucci, M. G., Chauhan, S., Payne, M. S., and DiCosimo, R. (2004)Tetrahedron 60, 577–581.
[46] Meyers, P. R., Rawlings, D. E., Woods, D. R., and Lindsey, G. G. (1993) J.Bacteriol. 175(19), 6105–6112.
[47] Bork, P., and Koonin, E. V. (1994) Prot. Sci. 3(8), 1344–1346.
[48] Pace, H., and Brenner, C. (2001) Gen. Biol. 2(1), 1–9.[49] Brenner, C. (2002) Curr. Opin. Struct. Biol. 12, 775–782.[50] Zhang, C. S., Zhang, Z. J., Li C. X., Yu, H. L., Zheng, G. W., and Xu, J. H.,
(2012) Appl. Microbiol. Biotechnol. 95(1), 91–99.[51] Zhang, Z. J., Xu, J. H., He, Y. C., Ouyang, L. M., and Liu, Y. Y. (2011)
Bioprocess Biosyst. Eng. 34, 315–322.[52] Gong, J. S., Lu, Z. M., Li H, Shi J. S., Zhou Z. M., and Xu, Z. H. (2012) Microb.
Cell Fact. 11, 142.[53] Nageshwar, Y. V. D., Sheelu, G., Shambhu, R. R., Muluka, H., Mehdi, N.,
Malik, M. S., and Kamal, A. (2011) Bioproc. Biosyst. Eng. 34, 515–523.[54] Xue, Y. P., Xu, S. Z., Liu, Z. Q., Zheng, Y. G., and Shen, Y. C. (2011) J. Ind.
Microbiol. Biotechnol. 38, 337–345.[55] Gavagan, J. E., DiCosimo, R., Eisenberg, A., Fager, S. K., Folsom, P. W., Hann,
E. C., Schneider, K. J., and Fallon, R. D. (1999) Appl. Microbiol. Biotechnol.52, 654–659.
[56] Chauhan, S., Wu, S., Blumerman, S., Fallon, R. D., Gavagan, J. E., DiCosimo,R., and Payne, M. S. (2003) Appl. Microbiol. Biotechnol. 61, 118–122.
[57] Yamamoto, K., and Komatsu, K. (1991) Agri. Biol. Chem. 55, 1459–1466.[58] Yamamoto, K., Fijimatsu, I., and Komatsu, K. (1992) J. Ferment. Bioeng. 73,
425–430.[59] Kobayashi, M., Izui, H., Nagasawa, T., and Yamada, H., (1993) Proc. Natl.
Acad. Sci. USA 90, 247–251.[60] Ingvorsen, K., Hojer-Pedersen, B., and Godtfredsen, S. E., (1991) Appl.
Environ. Microbiol. 57(6), 1783–1789.[61] Shen, M., Zheng, Y. G., and Shen, Y. C., (2009) Process Biochem. 44,781–785.[62] Alamatawah, Q. A., Cramp, R., and Cowan, D. A., (1999) Extremophiles 3,
283–291.[63] Jandhyala, D., Berman, M., Meyers, P. R., Sewell, B. T., Willson, R. C., and
Benedik, M. J. (2003) Appl. Environ. Microbiol. 69(8), 4794–4805.[64] Zhu, D., Mukherjee, C., Biehl, E. R., and Hua, L. (2007) J. Biotechnol. 129,
645–650.[65] Thiery, A., Maestracci, M., Arnaud, A., and Galzy, P., (1986) Zentralbl.
Mikrobiol. 141, 575–582.[66] Levy Schil, S., Soubrier, F., Crutz-Le Coq, A. M., Faucher, D., Crouzet, J., and
Petre, D., (1995) Gene 161, 15–20.[67] Williamson, D. S., Dent, K. C., Weber, B. W., Varsani, A., Frederick, J., Thuku,
R. N., Cameron, R. A., van Heerden, J. H., Cowan, D. A., and Sewell, B. T.(2010) Appl. Microbiol. Biotechnol. 88, 143–153.
[68] Stalker, D. M., Malyi, L. D., and McBride KE. (1988) J. Biol. Chem. 263,6310–6314.
[69] Harper, D. B. (1985) Int. J. Biochem. 17(6), 677–683.[70] Harper, D. B. (1977b) Biochem. J. 165, 309–319.[71] Hoyle, A. J., Bunch, A. W., and Knowles, C. J. (1998) Enz. Microb. Technol.
23, 475–482.[72] Nagasawa, T., Nanba, H., Ryuno, K., Takeuchi, K., and Yamada H. (1987) Eur.
J. Biochem. 162(3), 691–698.[73] Layh, N., Paratt, J., and Willets, A. (1998) J. Mol. Catal. B Enzym. 5(5–6),
467–474.[74] Kiziak, C., Conradt, D., Stolz, A., Mattes, R., and Klein, (2005) J. Microbiol.
151(11), 3639–3648.[75] Kim, J. S., Tiwari, M. K., Moon, H. J., Jeya, M., Ramu, T., Oh, D. K., Kim, I.
W., and Lee, J. K. (2009) Appl. Microbiol. Biotechnol. 83(2), 273–283.[76] Banerjee, A., Kaul, P., and Banerjee, U. (2006) Arch. Microbiol. 184, 407–418.[77] Yanase, H., Sakai, T., and Tonomura, K. (1983) Agric. Biol. Chem. 47, 473–482.[78] Dhillon, J. K., and Shivaraman, N. (1999) Can. J. Microbiol. 45(3), 201–208.[79] Watanabe, A., Yano, K., Ikebukuro, K., and Karube, I. (1998) Microbiology
144, 1677–1682.[80] Sewell, B. T., Berman, M. N., Meyers, P. R., Jandhyala, D., and Benedik, M.
(2003) Structure 11, 1–20.[81] Mueller, P., Egorova, K., Vorgias, C. E., Boutou, E., Trauthwein, H., Verseck,
S., and Antranikian, G. (2006) Protein Expr. Purif. 47, 672–681.[82] Dong, H. P., Liu, Z. Q., Zheng, Y. G., and Shen, Y. C. (2010) Appl. Microbiol.
Biotechnol. 87, 1335–1345.[83] Yang, C, Wang, X., and Wei, D. (2011) Appl. Biochem. Biotechnol. 165,
1556–1567.
Biotechnology and Applied Biochemistry 479

Biotechnology andApplied Biochemistry
[84] Kobayashi, M., Nagasawa, T., and Yamada, H. (1989) Eur. J. Biochem. 182,349–356.
[85] Thuku, R. N., and Sewell, B. T. (2005) Microsc. Soc. South Afr. Proc. 35, 31.[86] Prasad, S., Misra, A., Jangir, V., Awasthi, A., Raj, J., and Bhalla, T. (2007)
World J. Microbiol. Biotechnol. 23, 345–353.[87] Nigam, V. K., Khandelwal, A. K., Gothwal, R. K., Mohan, M. K., Choudhury,
B., Vidyarthi, A. S., and Ghosh, P. (2009) J. Biosci. 34, 21–26.[88] Bellinzoni, M., Buroni, S., Pasca, M. R., Guglierame, P., Arcesi, F., De Rossi,
E., and Riccardi, G. (2005) Res. Microbiol. 156, 173–177.[89] Yamada, H., and Kobayashi, M. (1996) Biosci. Biotechnol. Biochem. 60,
1391–1400.[90] Maier-Greiner, U. H., Obermaier-Skrobranek, B. M. M., and Estermaier, L.
M. (1991) Proc. Natl. Acad. Sci. USA 88, 4260–4264.[91] Mylerova, V., and Martinkova, L. (2003) Curr. Org. Chem. 7(14), 1–17.[92] Endo, I., Okada, M., and Yohda, M. (1999) Trend Biotechnol. 17, 244–248.[93] Popescn, V. C., Munk, E., Fox, B. G., Sanakis, Y., Cummings, J. G., Turner, I.
M., and Nelson, M. J. (2001) Biochemisty 40, 7984–7991.[94] Komeda, H., Hori, Y., Kobayashi, M., and Shimizu, S. (1996) Proc. Natl. Acad.
Sci. USA 90, 247–251.[95] Payne, M. S., Wu, S., Fallon, R. D., Tudor, G., Stieglitz, B., Turner, I. M., Jr,
and Nelson, M. J. (1997) Biochemistry 36, 5447–5454.[96] Celia, E., Cernia, E., Palocci, C., Soro, S., and Turchet, T. (2005) J. Supercrit.
Fluid 33, 193–199.[97] Banerjee, A., Kaul, P., Sharma, R., and Banerjee, U. C. (2003) J. Biomol.
Screen. 8, 559–565.[98] Calvete, J. J., Thole, H. H., Raida, M., Urbane, C., Romero, A., and Grangeiro,
T. B. (1999) Biochim. Biophys. Acta 1430, 367–375.[99] Yamada, K., Hasegawa, T., Minami, E., Shibuya, N., Kosemura, S.,
Yamamura, S., and Hasegawa, K., (2003) J. Plant Physiol. 160, 255–259.[100] Mayaux, J. F., Cerbelaud, E., Soubrier, F., Faucher, D., and Petre, D. (1990) J.
Bacteriol. 172, 6764–6773.[101] Endo, I., and Odaka, M. (2000) Mol. Catal. B: Enzym. 10, 81–86.[102] Novo, C., Tata, R., Clemente, A., and Brown, P. R. (1995) FEBS Lett. 367,
275–279.[103] Novo, C., Farnaud, S., Tata, R., Clemente, A., and Brown, P. R. (2002)
Biochem. J. 365, 731–38.[104] Farnaud, S., Tata, R., Sohi, M. K., Wan, T., Brown, P. R., and Sutton, B. J.
(1999) Biochem. J. 340, 711–714.[105] Pace, H. C., Hodawadekar, S. C., Draganescu, A., Huang, J., Bieganowski,
P., Pekarsky, Y., Croce, C. M., and Brenner, C. (2000) Curr. Biol. 10, 907–917.
[106] Fournand, D., Bigey, F., and Arnaud, A. (1998) Appl. Environ. Biol. 64,2844–2852.
[107] Hashimoto, A. G. (1986) Agric. Wastes 17, 241–261.[108] Gregoriou, M., and Brown, P. R. (1979) Eur. J. Biochem. 96, 101–108.[109] Hirrlinger, B., Stolz, A., and Knackmuss, H. J. (1996) J. Bacteriol. 178(12),
3501–3507.[110] Hongpattarakere, T., Komeda, H., and Asano, Y. (2005) J. Ind. Microbiol.
Biotechnol. 32(11–12), 567–576.[111] Chebrou, H., Bigley, F., Arnaud, A., and Glazy, P. (1996) Biochem. Biophys.
Acta 1298, 285–293.[112] O’Reilly, C. and Turner, P. D. (2003) J. Appl. Microbiol. 95, 1161–1174.[113] Ambler, R. P., Auffret, A. D., and Clarke, P. H. (1987) FEBS Lett. 215, 285–290.[114] Soubrier, F., Levy, Schil, S., Mayaux, J. F., Petre, D., Arnaud, A., and Crouzet,
J. (1992) Gene, 116, 99–104.[115] Cheong, Y. M., Jung, H. K., Joo, Y. S., Kim, S. S., and Kim, Y. S. (2000) IEEE
Trans. Ultrason. Ferroelectric Freq. Control 47(3), 559–564.[116] Hynes, M. J. (1975) J. Gen. Microbiol. 91, 91–109.[117] Fraser, H. B., Hirsh, A. E., Steinmetz, L. M., Scharfe, C., and Feldman, M. W.
(2002) Science 296, 750–752.[118] Rahim, M. A., Saxena, R. K., Gupta, R., Sheoran, A., and Giri, B. (2003)
Process Biochem. 38, 861–866.[119] Wegman, F. M. A., Janssen, M. H. A., van Rantwijk, F., and Sheldon, R. A.
(2001) Adv. Synth. Catal. 343, 559–576.[120] Effenberger, F., and Graef, B. W. (1998) Biotechnology 60, 165–174.
[121] Curnow, A. W., Hong, K., Yuan, R., Kim, S., Martins, O., Winkler, W., Henkin,T. M., and Soll, D. (1997) Proc. Natl. Acad. Sci. USA 94, 11819–11826.
[122] Skouloubris, S., Labigne, A., and De Reuse, H. (1997) Mol. Microbiol. 25(5),989–998.
[123] Cravatt, B. F., Giang, D. K., Mayfield, S. P., Boger, D. L., Lerner, R. A., andGilula, N. B. (1996) Nature 384, 83–87.
[124] Brandao, P. F. B., and Bull, A. T. (2003) Antonie van Leeuwenhoek 84, 89–98.[125] Soong, C. L., Ogawa, J., Honda, M., and Shimizu, S. (1999) Appl. Environ.
Microbiol. 65(4), 1459–1462.[126] Komeda, H., Harada, H., Washika, S., Sakamoto, T., Ueda, M., and Asano, Y.
(2004) Eur. J. Biochem. 271, 1580–1590.[127] Kobayashi, M., Nagasawa, T., and Yamada, H. (1988) Appl. Microbiol.
Biotechnol. 29, 231–233.[128] Osswald, S., Wajant, H., and Effenberger, F. (2002) Eur. J. Biochem. 269,
680–687.[129] Vorwerk, S., Biernacki, S., Hillebrand, H., Janzik, I., Muller, A., Weiler, E. W.,
and Piotrowski, M., (2001) Planta 212, 508–516.[130] Gagavan, J. E., DiCosimo, R., Eisenberg, A., Fager, S. K., Folsom, P. W., Hahn,
E. C., Schneider, K. J., and Fallon, R. D. (1999) Appl. Microbiol. Biotechnol.52, 654–659.
[131] Heinemann, U., Engels, D., Burger, S., Kiziak, C., Mattes, R., and Stolz, A.(2003) Appl. Environ. Microbiol. 69(8), 4359–4366.
[132] McBride, K. E., Kenny, J. W., and Stalker, D. M. (1986) Appl. Environ.Microbiol. 52, 325–330.
[133] Kunz, D. A., Wang, C. S., and Chen, J. L. (1994) Microbiology 140, 1705–1712.[134] Yanase, H., Sakamoto, A., Okamoto, K., Kita, K., and Sato, Y. (2000) Appl.
Microbiol. Biotechnol. 53, 328–334.[135] Barclay, M., Tett, V. A., and Knowles, C. J. (1998) Enzym. Microb. Technol.
23, 321–330.[136] Nakai, T., Hasegawa, T., Yamashita, E., Yamamoto, M., Kumasaka, T., Ueki,
T., Nanba, H., Ikenaka,Y., Takahashi, S., Sato, M., and Tsukihara, T. (2000)Structure 8(7), 729–739.
[137] Chen, C., Chiu, W., Liu, J., Hsu, W., and Wang, W. (2003) J. Biol. Chem.278(28), 26194–26201.
[138] Wang, W., Hsu, W., Chien, F., and Chen, C. (2001) J. Mol. Biol. 306, 251–261.[139] Kumaran, D., Eswaramoorthy, S., Gerchman, S. E., Kycia, H., Studier, F. W.,
and Swaminathan, S. (2003) Proteins 52, 283–291.[140] Sakai, N., Tajika, Y., Yao, M., Watanabe, N., and Tanaka, I. (2004) Proteins.
57, 869–873.[141] Jones, D. T. (1999) J. Mol. Biol. 287, 797–815.[142] Jandhyala, D. M., Wilson, R. C., Sewell, B. T., and Benedik, M. J. (2005) Appl.
Microbiol. Biotechnol. 68(3), 327–335.[143] Stevenson, D. E., Feng, R., and Storer, A. C. (1990) FEBS Lett. 277(1–2),
112–114.[144] Brady, D., Beeton, A., Zeevaart, J., Kgaje, C., van Rantwijk, F., and Sheldon,
R. A. (2004) Appl. Microbiol. Biotechnol. 64, 76–85.[145] Brammar, W. J., and Clarke, P. H. (1964) J. Gen. Microbiol. 37, 307–319.[146] Xu, G., and West, T. P. (1994) Arch. Microbiol. 161, 70–74.[147] Lee, C. K., and Fan, C. H. (1999) Enzym. Microb. Technol. 24, 659–666.[148] Ciskanik, L. M., Wilczek, J. M., and Fallon, R. D. (1995) Appl. Environ.
Microbiol. 61(3), 998–1003.[149] Nolan, L. M., Harnedy, P. A., Turner, P., Hearne, A. B., and O’Reilly, C. (2003)
FEMS Microbiol. Lett. 221, 161–165.[150] Ramakrishna, C., Dave, H., and Ravindranathan, M. (1999) J. Sci. Ind. Res.
58(12), 925–947.[151] Dias, J. C. T., Rezende, R. P., Rosa, C. A., Lachance, M. A., and Linardi, V. R.
(2000) Can. J. Microbiol. 46, 525–531.[152] Hughes, J., Armitage, Y., and Symes, K. C. (1998) Antonie van Leeuwenhoek
74, 107–118.[153] Almatawah, Q. A., and Cowan, D. A. (1999) Enzym. Microb. Technol. 25,
718–724.[154] Namboothiri, I. N. N., Rastogi, N., Ganguly, B., Mobin, S. M., and Cojocaru,
M. (2004) Tetrahedron 60, 1453–1462.[155] Bauer, R., Hirrlinger, B., Layh, N., Stolz, A., and Knackmuss, H. J. (1994)
Appl. Microbiol. Biotechnol. 42, 1–7.
480 Nitrile-Converting Enzymes

[156] Stalker, D. M., and McBride, K. E. (1987) J. Bacteriol. 169, 955–960.[157] Chacko, S., Ramteke, P. W., and Joseph, B. (2012) J. Genet. Eng. Biotechnol.
10, 121–127.[158] Takashima, Y. (1995) European Patent, 95-101282.2.10.[159] Bartling, D., Seedorf, M., Schmidt, R. C., and Weiler, E. W. (1994) Proc. Natl.
Acad. Sci. USA 91, 6021–6025.[160] Ikenaka, Y., Nanba, H., Yajima, K., Yamada, Y., Takano, M., and Takahashi, S.
(1998) Biosci. Biotechnol. Biochem. 62(9), 1668–16671.[161] Yamaki, T., Oikawa, T., and Ito, K. (1997) J. Ferment. Bioeng. 83, 474–477.[162] Cheong, T. K., and Oriel, P. J. (2000) Appl. Biochem. Biotechnol. 84–86,
903–915.[163] Komeda, H., and Asano, Y. (2005) Eur. J. Biochem. 272, 3075–3084.[164] Pollmann, S., Neu, D., and Weiler, E. W. (2003) Phytochemistry 62, 293–300.[165] Nagasawa, T., Shimizu, S., and Yamada, H. (1993) Appl. Microbiol.
Biotechnol. 40, 189–195.[166] Cowan, D. A., Cramp, R. A., Pereira, R. A., and Almatawah, Q. (1998)
Extremophiles 2, 207–216.[167] Yoshida, T., Mitsukura, K., Mizutani, T., Nakashima, R., Shimizu, Y., Kawabata,
H., and Nagasawa, T. (2013) Biotechnol. Lett. 35, 658–688.[168] Mathew, C. D., Nagasawa, T., Kobayashi, M., and Yamada, H. (1988) Appl.
Environ. Microbiol. 54, 1030–1032.[169] Sharma, N. N., Sharma, M., Kumar, H., and Bhalla, T. C. (2006) Process
Biochem. 41, 2078–2081.[170] Zhang, Z. J., Xu, JH., He, YC., Ouyang, L. M., Liu, Y. Y., and Imanaka, T. (2010)
Process Biochem. 45, 887–891.[171] Zabaznaya, E., Kozulin, S., and Voronin, S. (1998) Appl. Biochem. Microbiol.
34, 341–345.[172] He, Y. C., Xu, J. H., Su, J. H., and Zhou, L., (2009) Appl. Biochem. Biotechnol.
160, 1428–1440.[173] Wu, S., Fogiel, A. J., Petrillo, K. L., Jackson, R. E., Parker, K. N., DiCosimo, R.,
Ben-Bassat, A., O’Keefe, D. P., and Payne, M. S. (2008) Biotechnol. Bioeng.99, 717–720.
[174] Hashimoto, A. G. (1984) Agric. Wastes 9, 299–308.[175] Saillenfait, A. M., and Sabate, J. P. (2000) Toxico1. Appl. Pharmacol. 163(2),
149–163.[176] Vesela, A., Franc, M., Pelantova, H., Kubac, D., Vejvoda, V., Sulc, M., Bhalla,
T., Mackova, M., Lovecka, P., Janu, P., Demnerova, K., and Martinkova, L.(2010) Biodegradation 21, 761–770.
[177] Martinkova, L., Uhnakova, B., Patek, M., Nesvera, J., and Kren, V. (2009)Environ. Int. 35, 162–177.
[178] Mc Bride, K. E., Kenny, J. W., and Stalker, D. M. (1986) Appl. Environ.Microbiol. 52, 325–330.
[179] Freyssinet, G., Peleissier, B., Freyssinet, M., and Delon, R. (1996) Crop Res.45, 125–133.
[180] Wyatt, J. M., and Knowles, C. J. (1995) Int. Biodeter. Biodegrad. 35, 227–248.[181] Brady, D. (2010). In Handbook of Green Chemistry (Crabtree, R. H., ed.).
pp. 27–49, Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim.[182] Zhou, X. F., Zhang, Y. L., Xu, D. Q., Cao, W. H., Dai, C. M., Qiang, Z. M., Yang,
Z., and Zhao, J. F. (2008) Water Sci. Technol. 58, 911–918.[183] Christian, H. J., Hollergeschwandner, C., Peitzsch, M., Ress-Loeschke,
M., Hauer, B., Brandao, P. F., Bunch, A., Robinson, G., Bull, A. T., andSyldatk, C. (2001) Fifth International Symposium on Biocatalysis andBiotransformation. Darmstadt, 140.
[184] Li, C., Li, Y., Cheng, X., Feng, L., Xi, C., and Zhang, Y. (2013) Bioresour.Technol. 131, 390–396.
[185] Battistel, E., Morra, M., and Marinetti, M. (2000) Appl. Surf. Sci. 177,32–41.
[186] Tao, J., Lin, G. Q., and Liese A. (2009) Biocatalysis for the PharmaceuticalIndustry: Discovery, Development, and Manufacturing. John Wiley & SonsAsia Ltd.
[187] Matama, T., Carneiro, F., Caparros, C., Gubitz, G. M., and Cavaco-Paulo, A.(2007) Biotechnol. J. 2, 353–360.
[188] Feng, L. L., Zhang, J. F., Luo, H., and Li, Z. (2011) Adv. Mater. Res. 175–176,651–655.
[189] Fischer-Colbrie, G., Matama, T., Heumann, S., Martinkovac, L., Paulob, A. C.,and Guebitz, G. (2007) J. Biotechnol. 129, 62–68.
[190] Holmes, L. B. (1996) Teratology 53, 227–229.[191] Tamura, G., Sakata, K., Nishizuka, S., Maesawa, C., Suzuki, Y., Iwaya, T.,
Terashima, M., Saito, K., and Satodate, R. (1997) Genes ChromosomesCancer 20, 98–102.
[192] Kaul, P., Banerjee, A., Mayilraj, S., and Banerjee, U. C. (2004) Tetrahedron15, 207–211.
[193] Banerjee, A., Dubey, S., Kaul, P., Barse, B., Piotrowski, M., and Banerjee, U.C. (2009) Mol. Biotechnol. 41, 35–41.
[194] Ishikawa, T., Watabe, K., Mukohara, Y., and Nakamura, H. (1996) Biosci.Biotechnol. Biochem. 60(4), 612–615.
[195] Yokozeki, K., Nakamori, S., Eguchi, C., Yamada, K., and Mitsugi, K. (1987)Agric. Biol. Chem. 51(2), 355–362.
[196] Politi, L., Chiancone, E., Giangiacomo, L., Cervoni, L., Scotto d’Abusco, A.,Scorsino, S., and Scandurra, R. (2009) Archaea 2(4), 221–31.
[197] Prasad, A. S., and Satyanarayana, B. (2012) Der. Pharm. Chem. 4(1), 93–99.[198] Dufour, E., Tam, W., Nagler, D. K., Storer, A. C., and Menard, R. (1998) FEBS
Lett. 433, 78–82.[199] Meth-Cohn, O., and Wang, M. X. (1997) J. Chem. Soc. Perkin Trans. 1,
1099–1104.[200] Shanker, R., Ramakrishna, C., and Seth, P. K. (1990) Arch. Microbiol. 154,
192–198.[201] Chhiba, V., Moira, L. B., Mathiba, K., Kwezi, W., and Brady, D. (2012) J. Mol.
Catal. B Enzym. 76, 68–74.[202] Almatawah, Q. A., Cramp, R., and Cowan, D. A. (1999) Extremophiles 3(4),
283–291.[203] Grabam, D., Pereira, R, Barfield, D., and Cowan, D. (2000) Enzym. Microb.
Technol. 26, 368–373.[204] Chen, J., Zheng, R. C., Zheng, Y. G., and Shen, Y. C. (2009) Adv. Biochem.
Eng. Biotechnol. 113, 33–77.[205] Gaffney, T. D., da Costa e Silva, O., Yamada, T., and Kosuge, T. (1990) J.
Bacteriol. 172, 5593–5601.[206] Detzel, C., Maas, R., Tubeleviciute, A., and Jose, J. (2012) Appl. Microbiol.
Biotechnol. 97(11), 4887–4896.[207] Godtfredsen, W. O., and Vangedal, S. (1964) Proc. Chem. Soc. Lond. 188.[208] Hartmeier, W. (1988) Immobilized Biocatalysts. Springer-Verlag, Berlin.[209] Dadd, M. R., Sharp, D. C. A., Pettman, A. J., and Knowles, C. J. (2000) J.
Microbiol. Methods 41, 69–75.[210] Wang, N., Xu, Y., and Lu, D. (2004) AATCC Rev. 4, 28–30.[211] Tauber, M. M., Cavaco-Paulo, A., Robra, K. H., and Gubitz, G. M. (2000) Appl.
Environ. Microbiol. 66, 1634–1638.[212] Thuku, R. N. (2006) The Structure of the Nitrilase from Rhodococcus
rhodochrous J1: Homology Modeling and Three-Dimensional Recon-struction. M.Sc. Minithesis, University of the Western Cape and ElectronMicroscope Unit, University of Cape Town.
Biotechnology and Applied Biochemistry 481
Related Documents











