MOLECULAR PLANT PATHOLOGY (2005) 6 (1), 65–78 DOI: 10.1111/J.1364-3703.2004.00266.X © 2005 BLACKWELL PUBLISHING LTD 65 Blackwell Publishing, Ltd. Nitric oxide contributes both to papilla-based resistance and the hypersensitive response in barley attacked by Blumeria graminis f. sp. hordei ELENA PRATS 1 *, LUIS A. J. MUR 2 , RUTH SANDERSON 1 AND TIMOTHY L. W. CARVER 1 1 Institute of Grassland and Environmental Research, Aberystwyth, Ceredigion, SY23 3EB, UK 2 University of Wales Aberystwyth, Institute of Biological Sciences, Aberystwyth, Ceredigion, SY23 2DA, UK SUMMARY Nonspecific penetration resistance due to papilla formation and race-specific hypersensitive response (HR) can both contribute to Blumeria graminis resistance in barley. Some effective papillae form even in the susceptible cv. Pallas and the isoline P01 carries the additional Mla1 allele conditioning HR. The NO-specific stain DAF-2DA (4,5-diaminofluorescein-2-diacetate) revealed a transient NO generation burst commencing 10 h after inoculation (h.a.i.) in close association with sites of papilla formation in both barley lines. In P01 a burst of NO production throughout some attacked cells was initiated around 10 –12 h.a.i. and this preceded whole-cell autofluorescence indicative of HR. The specificity of DAF-2DA staining was demonstrated by the suppression of staining following application of the NO scavenger C-PTIO (1 H-imidazol- 1-yloxy-2-(4-carboxyphenyl)-4,5-dihydro-4,4,5,5-tetramethyl-3-oxide). In addition, C-PTIO application increased penetration frequencies in both barley lines, indicating a role for NO in papilla-based resistance. Furthermore, C-PTIO application slightly delayed HR in P01 whereas, conversely, application of an NO donor, sodium nitroprusside, slightly accelerated HR in P01 and increased cell death frequency in Pallas. Thus, NO generation is one of the earliest responses of barley epidermal cell defence against B. graminis attack and may be important in both the initiation and the development of effective papillae and cell death due to HR. INTRODUCTION Resistance to pathogens is often associated with elicitation of the hypersensitive response (HR) mediated by the interaction of a host- encoded resistance (R ) gene product with that of a pathogen avirulence (avr ) gene. The active oxygen species (AOS) H 2 O 2 (Chamnongpol et al., 1998; Levine et al., 1994) or (Jabs et al., 1997) have been advanced as key signals in the HR but other studies show that AOS alone can be insufficient to induce cell death (Dorey et al., 1999; Glazener et al., 1996). Recent evidence indicates that AOS can act either in parallel or synergistically with the gaseous radical, nitric oxide (NO), to elicit cell death (Delledonne et al ., 2001; de Pinto et al ., 2002). Rapid NO generation follows inoculation of avirulent but not virulent bacterial strains into soybean (Delledonne et al., 1998) and Arabidopsis cell suspension cultures (Clarke et al., 2000) as well as treatment of tobacco epidermal peels with a necrotizing elicitor (Foissner et al., 2000). In mammalian cells, NO is most frequently generated from the NADPH-dependent conversion of L-arginine to L-citrulline by a nitric oxide synthase (NOS, Alderton et al., 2001). NOS activity may be suppressed by substrate analogues such as N G -monomethyl-L-arginine (L-NMMA) or its methyl-ester (L-NAME; Young et al., 2000). In Arabidopsis, two novel NO-generating complexes have been isolated, and although neither is similar to mammalian NOS, L-NAME inhibits both (Chandok et al., 2003; Guo et al., 2003). L-NMMA treatment suppresses the HR in Arabidopsis (Delledonne et al., 1998) and pathogen resistance in both tobacco and Arabidopsis (Delledonne et al., 1998; Dürner et al., 1998). Expression of a nitric oxide dioxygenase (NOD) in transgenic plants and within challenging avirulent bacteria, to suppress NO levels, also compromised defence associated with HR, leading to reduced resistance (Zeier et al., 2004). Furthermore, NO may elicit cytological responses reminiscent of mammalian apoptosis, again supporting its importance in the plant cell death process. Thus, exogenous application of NO elicits chromatin condensation and DNA fragmenta- tion (Clarke et al., 2000; Pedroso et al., 2000), which in Arabidopsis cell cultures is mediated by caspase-like activity that can be suppressed with a mammalian caspase-1 inhibitor (Clarke et al., 2000). Although great progress has been made in understanding plant NO biology using model plant – pathogen interactions and often using cell culture systems, few studies have assessed its spatio- temporal role in economically important crop–pathogen interactions. Here we have applied such an analysis to barley ( Hordeum vulgare L.) leaf tissues attacked by the barley powdery mildew fungus, Blumeria graminis (DC) Speer f. sp. hordei Marchal. *Correspondence : E-mail: [email protected] O 2 −

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78 DOI 10 1111 J 1364-3703200400266X
copy 2005 BLACKWELL PUBL ISH ING LTD
65
Blackwell Publishing Ltd
Nitric oxide contributes both to papilla-based resistance and the hypersensitive response in barley attacked by
Blumeria graminis
f sp
hordei
E LENA PRATS
1
LU I S A J MUR
2
RUTH SANDERSON
1
AND T IMOTHY L W CARVER
1
1
Institute of Grassland and Environmental Research Aberystwyth Ceredigion SY23 3EB UK
2
University of Wales Aberystwyth Institute of Biological Sciences Aberystwyth Ceredigion SY23 2DA UK
SUMMARY
Nonspecific penetration resistance due to papilla formation andrace-specific hypersensitive response (HR) can both contributeto
Blumeria graminis
resistance in barley Some effective papillaeform even in the susceptible cv Pallas and the isoline P01 carriesthe additional Mla1 allele conditioning HR The NO-specificstain DAF-2DA (45-diaminofluorescein-2-diacetate) revealed atransient NO generation burst commencing 10 h after inoculation(hai) in close association with sites of papilla formation in bothbarley lines In P01 a burst of NO production throughout someattacked cells was initiated around 10ndash12 hai and this precededwhole-cell autofluorescence indicative of HR The specificity ofDAF-2DA staining was demonstrated by the suppression of stainingfollowing application of the NO scavenger C-PTIO (1
H
-imidazol-1-yloxy-2-(4-carboxyphenyl)-45-dihydro-4455-tetramethyl-3-oxide)In addition C-PTIO application increased penetration frequenciesin both barley lines indicating a role for NO in papilla-basedresistance Furthermore C-PTIO application slightly delayed HRin P01 whereas conversely application of an NO donor sodiumnitroprusside slightly accelerated HR in P01 and increasedcell death frequency in Pallas Thus NO generation is one ofthe earliest responses of barley epidermal cell defence against
B graminis
attack and may be important in both the initiation and
the development of effective papillae and cell death due to HR
INTRODUCTION
Resistance to pathogens is often associated with elicitation of thehypersensitive response (HR) mediated by the interaction of a host-encoded resistance (
R
) gene product with that of a pathogen avirulence(
avr
) gene The active oxygen species (AOS) H
2
O
2
(Chamnongpol
et al
1998 Levine
et al
1994) or (Jabs
et al
1997) have been
advanced as key signals in the HR but other studies show that AOSalone can be insufficient to induce cell death (Dorey
et al
1999Glazener
et al
1996) Recent evidence indicates that AOS can acteither in parallel or synergistically with the gaseous radical nitric oxide(NO) to elicit cell death (Delledonne
et al
2001 de Pinto
et al
2002)Rapid NO generation follows inoculation of avirulent but not
virulent bacterial strains into soybean (Delledonne
et al
1998)and
Arabidopsis
cell suspension cultures (Clarke
et al
2000) as wellas treatment of tobacco epidermal peels with a necrotizing elicitor(Foissner
et al
2000) In mammalian cells NO is most frequentlygenerated from the NADPH-dependent conversion of
L
-arginineto
L
-citrulline by a nitric oxide synthase (NOS Alderton
et al
2001)NOS activity may be suppressed by substrate analogues suchas
N
G
-monomethyl-
L
-arginine (L-NMMA) or its methyl-ester (L-NAMEYoung
et al
2000) In
Arabidopsis
two novel NO-generatingcomplexes have been isolated and although neither is similar tomammalian NOS L-NAME inhibits both (Chandok
et al
2003 Guo
et al
2003) L-NMMA treatment suppresses the HR in
Arabidopsis
(Delledonne
et al
1998) and pathogen resistance in both tobaccoand
Arabidopsis
(Delledonne
et al
1998 Duumlrner
et al
1998)Expression of a nitric oxide dioxygenase (NOD) in transgenic plantsand within challenging avirulent bacteria to suppress NO levelsalso compromised defence associated with HR leading to reducedresistance (Zeier
et al
2004) Furthermore NO may elicit cytologicalresponses reminiscent of mammalian apoptosis again supportingits importance in the plant cell death process Thus exogenousapplication of NO elicits chromatin condensation and DNA fragmenta-tion (Clarke
et al
2000 Pedroso
et al
2000) which in
Arabidopsis
cell cultures is mediated by caspase-like activity that can be suppressedwith a mammalian caspase-1 inhibitor (Clarke
et al
2000)Although great progress has been made in understanding plant
NO biology using model plantndashpathogen interactions and oftenusing cell culture systems few studies have assessed its spatio-temporal role in economically important cropndashpathogen interactionsHere we have applied such an analysis to barley (
Hordeum vulgare
L) leaf tissues attacked by the barley powdery mildew fungus
Blumeria graminis
(DC) Speer f sp
hordei
Marchal
Correspondence
E-mail elenapratsbbsrcacuk
O2minus
66
E PRATS
et al
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Blumeria graminis
is an obligate biotroph whose asexual conidiagerminate and develop reasonably synchronously through a highlyordered morphogenetic sequence (reviewed by Green
et al
2002)Emergence of a short primary germ tube (30ndash60 min) is followedby that of the second appressorial germ tube that elongates anddifferentiates a hooked apical appressorium by around 10 hafter inoculation (hai) A penetration peg emerging beneath theappressorium (10ndash12 hai) attempts to breach the plant epidermalcell wall probably combining physical force and enzymaticdegradation If successful it enters the cell lumen where its tipswells (12ndash15 hai) and differentiates (15ndash20 hai) into a maturehaustorium that develops numerous digitate processes overthe next 4ndash5 days This absorbs nutrient from the epidermal cellto support growth of ectophytic secondary hyphae from whichsubsequent generations of haustoria (from 3 days) and conidio-phores (from 4 days) are formed
Two of the mechanisms by which cereals defend against powderymildew are by forming papillaemdashcell wall appositions depositedon the inner surface of epidermal cell walls directly beneathappressoriamdashand by the death of attacked cells Attacked cellsthat form an effective papilla survive and much evidence (reviewedby Zeyen
et al
2002) indicates that papillae present a physicalandor chemical penetration barrier Papillae form where appres-sorium contact attempted penetration causes focused cytologicalresponses including reorganization of the cell cytoskeletonand cytoplasmic streaming leading to formation of a cytoplasmicaggregate from which papilla components are depositedPapillae have a callose matrix containing various inorganic andorganic constituents including proteins and autofluorescentphenolic compounds (Zeyen
et al
2002) A halo of altered stainingaffinity in the epidermal cell wall often surrounds the papilla sitePapillae are formed even in susceptible host genotypes suchas the barley cv Pallas (Lyngkjaeligr
et al
2001) used in the presentstudies but often these papillae are ineffective and the fungalpenetration peg passes through the deposit which remains asthe haustorial neck collar that seals the site of invasion Papillaeffectiveness may relate to the rate of deposition to quantitativeor qualitative characteristics or to a combination of these factorsEarly H
2
O
2
accumulation indicated by DAB (33
prime
-diaminobenzidine)staining (Thordal-Christensen
et al
1997) may be important in cellsignalling to initiate papilla formation and it appears particularlyimportant in effective papilla-based resistance either through itsantimicrobial toxicity or by contributing to papilla-strengtheningthrough the oxidative cross-linking and immobilization of structuralcompounds (reviewed by Huumlckelhoven and Kogel 2003)
In cerealndashpowdery mildew interactions barley alleles at the
Mla
locus which condition HR have been relatively well characterized(Chelkowski
et al
2003 Panstruga and Schulze-Lefert 2002)When avirulent isolates attack
Mla1
barley such as the nearisogenic line P01 derived from cv Pallas (Koslashlster
et al
1986) usedhere HR occurs rapidly before or soon after visible haustoria form
leading to death and collapse of the single attacked epidermalcell Death is preceded by cessation of cytoplasmic streamingfollowed by loss of ability to take up vital dyes or to plasmolyse(Koga
et al
1988) The onset of whole-cell autofluorescence is areliable and convenient indicator of cell death (Bushnell 1981Koga
et al
1988 Lyngkjaeligr
et al
2001 Zeyen
et al
1995) One ofthe first signs of incompatibility conditioned by
Mla1
(Vanacker
et al
2000) or
Mla3
(Thordal-Christensen
et al
1997) is the transientappearance (at 14ndash15 hai) of H
2
O
2
at attachment sites betweenattacked epidermal cells and underlying mesophyll This precedesthe appearance of H
2
O
2
throughout the epidermal cell (at around16 h) which is in turn followed by death (Vanacker
et al
2000)Previous work suggested the possibility that the rapid signalling
events occurring in the challenged epidermis and underlyingmesophyll may arise from NO generation (Vanacker
et al
2000)Here we used confocal laser scanning electron microscopy (CLSM)to visualize NO generation associated with both papilla formationand HR during the interaction of
B graminis
f sp
hordei
andbarley by applying the stain DAF-2DA (45-diaminofluorescein-2-diacetate) which forms fluorescent DAF-2T (diaminotriazoloflu-orescein) in the presence of NO (Kojima
et al
1998ab) ReducingNO levels by treatment with the NO scavenger C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxidepotassium salt) (Delledonne
et al
1998 Garces
et al
2001Neill
et al
2003) suppressed penetration resistance and slightlydelayed HR Conversely when additional NO was providedby sodium nitroprusside (SNP Clarke
et al
2000 Garces
et al
2001 Pedroso
et al
2000) HR was slightly accelerated althoughpenetration resistance was unaffected Our data indicate roles forNO in both papilla defence and cell death
RESULTS
CLSM visualization of NO generation in barley attacked by
B graminis
CSLM after DAF-2DA staining of both Pallas (susceptible) and P01(
Mla1
resistance) revealed intense localized fluorescence indica-tive of NO generation at barley epidermal cell contact sites with
B graminis
f sp
hordei
appressoria Between 6 and 24 hai imagesof 157 and 120 germlings on Pallas and P01 respectively werecaptured before and after staining Visual inspection indicatedthat NO generation was an early response that became abruptlyevident as appressoria matured in both barley lines Thus up to9 hai there was no evidence for NO generation associated withimmature appressorial germ tubes However at 10ndash12 hai beforestrong autofluorescence indicative of phenolic compound accumula-tion was evident (Fig 1A) staining often led to localized pronouncedfluorescence beneath appressoria (Fig 1B) An intense fluorescentfocus subtending appressoria suggested NO accumulation in plantcell cytoplasmic aggregates and papillae and this focus was often
NO and resistanceresponse in barley
67
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
encircled by a less intense zone corresponding to the cell wallhalo (Fig 1B) The absence of DAF-2T fluorescence within thehook-shaped appressoria indicated that NO was primarily localizedto the plant cell However restricted NO generation appearedtransient because intensified fluorescence after staining becameincreasingly difficult to discern as localized autofluorescencedeveloped By 24 hai when intense autofluorescence wasevident (Fig 1C) staining did not appear to increase in intensity(Fig 1D)
Overview images (Fig 1) of localized fluorescence responsesdid not show the depth of NO generation within response sitesand thus three-dimensional models were created from transverseoptical sections made through such sites Figure 2(A) shows arepresentative example of an appressorial attack site examined14 hai As was common at this time (Fig 1) before stainingthe site was non-fluorescent indicating the absence of phenolicautofluorogens (data not shown) but after staining the overviewimage showed a characteristically circular zone of intensefluorescence indicative of NO generation in the region of papillaformation and the halo (as in Fig 1B) The transverse opticalsection suggested that NO generation was integral to the devel-oping papilla with little detectable DAF-2T fluorescence in thesurrounding cell wall and cytoplasm The transmitted light imagewas insufficiently detailed to resolve details of fungal structurebut it seems reasonable to suppose that the thickened region ofNO generation subtended the site of penetration peg emergencefrom the appressorial lobe Figure 2(B) shows before and afterDAF-2DA staining of a representative site of response to appres-sorial attack at 18 hai when deposition of the mature papillawould have been completed As shown previously (Fig 1) inoverview the only fluorescence detected was associated withautofluorescent phenolic compounds which accumulated in thepapilla and halo region and transverse optical sections of the site
were also virtually indistinguishable before and after stainingIt is also notable that the distribution of DAF-2T fluorescence inFig 2(A) and that of autofluorogens in Fig 2(B) was strikinglysimilar in shape and size This provides good evidence that NOdistribution at sites of early response to penetration is closelycorrelated with the subcellular locality in which components ofpapillae and haloes accumulate at maturity
Fluorescence intensity values (corrected for background) fromcontact sites that met the stringent criteria of acceptability (seeExperimental procedures) were obtained before and after DAF-2DA staining These provided objective data on the developmentof autofluorescence and fluorescence due to DAF-2DA stainingwithin separate time intervals over the period 6ndash24 hai (Fig 3)Thus for each individual contact site the fluorescence intensityvalue before and after staining was obtained and the differencebetween these values was calculated for each site Within theseparate time intervals the data for difference were subjected tostatistical analysis (see Experimental procedures) which showedvery similar trends in Pallas and P01 Appressorial germ tubecontact sites showed no localized fluorescence before 10 hai(before appressoria matured) but at 10ndash12 hai when autoflu-orescence was barely detectable in either barley line contactsites showed substantial elevated fluorescence after stainingindicative of NO generation Significant increases in fluorescenceafter staining were also evident at 13ndash14 and at 15ndash16 hai inP01 indicating continued NO production throughout this periodHowever in later samples there was no significant increase influorescence after DAF-2DA staining The data therefore suggesta burst of NO generation commencing very soon after appressorialmaturation but declining to undetectable levels from around 14 haionwards in Pallas and from 16 hai in P01
Interestingly at 17ndash19 hai staining led to a slight decreasein fluorescence intensity in both lines and in Pallas this effect was
Fig 1 Localized NO generation in Pallas (susceptible) barley epidermal cells at the B graminis appressorial germ tube contact site CLSM images of localized responses to B graminis attack 12 and 24 h after inoculation (hai) before and after DAF-2DA staining to reveal NO generation Ap = appressorium contact site C = conidium Ha = halo P = primary germ tube contact site (A) At 12 hai before staining a faint image of the fungus is evident and the P and Ap contact sites show faint autofluorescence (B) As in A after staining NO generation in the region of papilla deposition is indicated by intense fluorescence at the Ap site and fainter fluorescence is present in the halo region (C) At 24 hai before staining the P and Ap sites show strong autofluorescence in papillae and fainter autofluorescent haloes (D) As in C after staining there is no intensification of fluorescence indicating absence of NO Scale bars = 20 microM
68
E PRATS
et al
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
significant This suggests that components of the staining solu-tion either caused quenching or removed autofluorogens fromresponse sites To test this comparisons were made between thefluorescence intensity of individual papilla sites sampled in theperiods 15ndash16 and 17ndash19 hai before and after treatment withbuffer alone or treatment with buffer plus DMSO but omittingDAF-2DA Treatment with buffer alone had no effect on fluores-cence intensity but treatment with buffer plus DMSO led to asignificant decrease in intensity (
P
lt 0001) in papilla sites treated15ndash16 hai (data not shown) Because the data for intensityafter staining (Fig 3) represent the sum of fluorescence due toformation of DAF-2T and autofluorescence any reduction ofautofluorescence due to quenching or removal would decreasethe post-staining intensity value Thus the data probably under-estimate the contribution resulting from DAF-2T formation due to
Fig 3 Quantification of localized fluorescence responses at B graminis appressorium contact sites in Pallas (susceptible) and P01 (Mla1-resistant) barley epidermal cells Localized fluorescence intensities at appressorial contact sites on Pallas and P01 were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
Fig 2 Three-dimensional models of early localized NO generation and mature papilla structure at B graminis appressorial germ tube contact sites in Pallas barley (A) Overview image of an appressorium and host response site at 14 hai stained with DAF-2DA (no autofluorescence was present before staining image not shown) Intense green fluorescence indicates the presence of NO in the central region underlying the appressorial lobe (AL) at the tip of the appressorial germ tube (AGT) This is surrounded by a less intensely stained halo The red line marks the plane of a transverse optical section shown in the lower pane This shows a vertically thickened fluorescent region within the site of papilla deposition and a thinner region of peripheral staining corresponding to the halo (B) Upper images show overviews captured at 18 hai of an appressorium overlying a response site with a mature papilla Autofluorescence in the unstained specimen (left) indicates the presence of phenolic compounds and (right) no further fluorescence after DAF-2DA treatment The red lines mark the position of the transverse optical sections shown in the corresponding lower images which also show that staining did not add to fluorescence The dimensions and shape of autofluorescence associated with the mature papilla and halo correspond closely to the distribution of NO seen earlier (compare with A)
NO and resistanceresponse in barley
69
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
NO generation This does not however invalidate the conclusionthat NO was generated in an early but transient burst
In
Mla1
barley whole-cell autofluorescence indicating celldeath due to HR is first evident around 15ndash18 hai (Koga
et al
1990 Vanacker
et al
2000) but in susceptible Pallas cell deathis very infrequent Here cell death was seen so rarely in Pallasthat no attempt was made to study it In the current study visualinspection of P01 images also showed little evidence of whole-cell autofluorescence until after 15 hai but before this DAF-2DA staining often led to whole-cell fluorescence (Fig 4B)suggesting NO generation throughout some cells under attackfrom appressoria (Fig 4AB) These data suggested that whole-cell NO generation preceded autofluorogen accumulation in cellsundergoing HR (Fig 4A) As for localized responses this whole-cell generation of NO in P01 also appeared transient becomingincreasingly difficult to discern as autofluorescence developedThus by 24 hai when intense whole-cell autofluorescence wasevident (Fig 4C) staining added little if anything to the fluores-cence (Fig 4D)
Fluorescence intensity data from individual whole cells examinedbefore and after staining supported the conclusion that transientwhole-cell NO generation preceded cell death Figure 5 showsthat whole-cell autofluorescence intensities were very low insamples examined before 15 hai Nevertheless the
t
-test ofdifferences between whole-cell fluorescence intensity before and
Fig 4 Whole-cell fluorescence in P01 (Mla1-resistant) barley epidermis following challenge with B graminis CLSM images of whole-cell responses to B graminis attack 14 and 23 hai before and after DAF-2DA staining Ap = appressorium contact site P = primary germ tube contact site (A) At 14 hai before staining the P site shows distinct localized autofluorescence but the Ap site shows little and there is no whole-cell autofluorescence of the attacked cell (arrowhead) A second cell also without whole-cell autofluorescence is marked (double arrowhead) (B) As in A after staining the cell attacked by the appressorium (which was displaced slightly during staining) shows whole-cell NO generation indicated by its whole-cell fluorescence this is also evident in the second cell (double arrowhead) that was attacked out of view (C) At 23 hai before staining the P site shows distinct localized autofluorescence and the cell attacked by the appressorium shows whole-cell autofluorescence (D) As in C after staining there is no apparent intensification of fluorescence indicating absence of NO Scale bars = 50 microm
Fig 5 Quantification of whole-cell fluorescence responses to B graminis attack in P01 (Mla1-resistant) barley epidermal cells Whole-cell fluorescence intensities of attacked epidermal cells were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
70 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
after staining showed significantly increased whole-cell fluores-cence intensity following DAF-2DA at 10ndash12 hai and an evenmore marked and significant effect in the period 13ndash14 haiThis suggested a substantial burst of NO generation at a veryearly stage when the low intensity of whole-cell autofluorescenceindicated that cells remained alive From 15 hai howeverstatistical analysis showed no significant effect of DAF-2DAstaining indicating cessation of NO generation in cells that weredying or dead as indicated by their relatively intense whole-cellautofluorescence
C-PTIO suppresses NO accumulation as visualized by CLSM
The work above assumed that intensified fluorescence afterDAF-2DA staining was due to NO To test this CLSM was used ina second study of P01 over the critical period 10ndash19 hai buthere leaf segments were first bathed in the NO scavenger C-PTIObefore applying DAF-2DA
As previously (Fig 3) Fig 6 shows that the fluorescence intensityof papilla sites obtained before treatment indicating autofluores-cence intensity was low at 10ndash12 hai but increased thereafterHowever when treated with C-PTIO there was no detectableincrease in intensity of localized fluorescence following subse-quent DAF-2DA staining even over the period 10ndash14 hai And
as previously (Fig 3) staining samples 17ndash19 hai caused a lossof fluorescence
When C-PTIO was applied before staining no cells showed whole-cell fluorescence after C-PTIODAF-2DA treatment unless theyshowed whole-cell autofluorescence before treatment Over theperiod 10ndash19 hai 20 cells showing faint or obvious whole-cellautofluorescence were imaged and there was no statisticallysignificant increase in whole-cell fluorescence after treatment evenin the 10ndash14 hai period when the previous experiment showedmarked intensification (Fig 5) Together these observationsindicate that C-PTIO effectively removed NO from P01 barley cellsundergoing HR
L-NAME does not suppress NO accumulation as visualized by CLSM
Mammalian NOS is effectively inhibited by L-NAME which alsoinhibits some NO-related defensive responses in certain dicotyle-donous plants (Chandok et al 2003 Duumlrner et al 1998 Guo et al2003) In a similar experiment to that performed with C-PTIO(above) L-NAME was applied for 30 min prior to applying DAF-2DAin samples examined over the critical period 10ndash19 hai Figure 7shows data obtained from papilla sites demonstrating that thetemporal pattern of autofluorescence development was verysimilar to that observed in earlier experiments (Figs 3 and 6)intensifying from low to much higher levels over this period
Fig 6 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of the NO scavenger C-PTIO and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before C-PTIODAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
Fig 7 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of L-NAME and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before L-NAMEDAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
NO and resistanceresponse in barley 71
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Patterns of NO generation were also very similar to those seenin untreated leaves (Fig 3) again indicating a transient burst ofgeneration Hence DAF-2DA staining led to significant increasesin fluorescence intensity indicative of NO generation in papillasites over the period 10ndash14 hai a near-significant increase wasfound in samples examined 15ndash16 hai but thereafter therewas no evidence for NO generation Therefore unlike C-PTIO therewas no evidence for an immediate effect of L-NAME treatment onNO generation at sites of papilla formation Furthermore althoughthe number of cells undergoing HR was too small for reliablestatistical analyses there was no indication that L-NAME affectedwhole-cell fluorescence responses Thus as in untreated leaves(Fig 5) treatment did not diminish the early whole-cell fluores-cence revealed by DAF-2DA staining
Light and fluorescence microscope analyses of effects of C-PTIO L-NAME and SNP on B graminis development and barley epidermal cell responses to attack
To investigate the role(s) of NO generated in the interaction ofB graminis with barley an NO scavenger (C-PTIO) a NOS inhib-itor (L-NAME) and an NO donor (SNP) were supplied separatelyto cut leaves Light microscopy showed that as compared withcontrols treating leaves with any of the test chemicals had noeffects on spore germination or subsequent germling develop-ment up to the differentiation of mature (hooked) appressoria(data not shown) Furthermore incident fluorescence microscopyindicated that none of the chemicals affected the timing frequency
or intensity of localized autofluorescent host cell responses asso-ciated with appressoria Detailed data for these responses are notgiven but in all cases approximately 10 of appressorial contactsites were associated with faint localized autofluorescent hostcell responses by 12 hai and by 24 hai approximately 60 ofsites showed autofluorescence which in most cases was judgedfaint or moderately intense (class 1 or 2 on the 0ndash3 subjectivescale) Other effects of each test chemical are considered in turn
Effects of C-PTIOA few very small haustoria (swollen apices of penetration pegs)indicating successful penetration were evident in epidermalcells of both barley lines by 12 hai and in both there was littleincrease in penetration frequency after 15ndash18 hai (Fig 8) Thisindicates that the outcome of attacks and therefore efficacy ofpenetration resistance was determined by this time Comparedwith controls overall analysis showed that C-PTIO treatmentsignificantly increased (P lt 0001) the frequency of successfulpenetration ie suppressed penetration resistance in bothbarley lines (Fig 8) Further analyses showed that this effectwas significant (P lt 005) at most individual sampling times Inboth Pallas and P01 successful penetration was associatedwith approximately 30 of appressoria in C-PTIO-treated leaveswhereas control values were around 15 In Pallas some cells diedfollowing successful penetration but this percentage remainedrelatively low throughout (only c 20ndash40 died Fig 8) andtreatment had no significant effect By contrast in P01 virtually allpenetrated haustorium-containing cells were dead by 18 hai(Fig 8) However at 12ndash15 hai some penetrated P01 cells showed
Fig 8 C-PTIO suppression of B graminis penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO scavenger C-PTIO (triangles) or water (circles) on percentages of B graminis appressoria with successful penetrations (haustoria) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (indicated by whole-cell autofluorescence) 12ndash30 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001
72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

66
E PRATS
et al
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Blumeria graminis
is an obligate biotroph whose asexual conidiagerminate and develop reasonably synchronously through a highlyordered morphogenetic sequence (reviewed by Green
et al
2002)Emergence of a short primary germ tube (30ndash60 min) is followedby that of the second appressorial germ tube that elongates anddifferentiates a hooked apical appressorium by around 10 hafter inoculation (hai) A penetration peg emerging beneath theappressorium (10ndash12 hai) attempts to breach the plant epidermalcell wall probably combining physical force and enzymaticdegradation If successful it enters the cell lumen where its tipswells (12ndash15 hai) and differentiates (15ndash20 hai) into a maturehaustorium that develops numerous digitate processes overthe next 4ndash5 days This absorbs nutrient from the epidermal cellto support growth of ectophytic secondary hyphae from whichsubsequent generations of haustoria (from 3 days) and conidio-phores (from 4 days) are formed
Two of the mechanisms by which cereals defend against powderymildew are by forming papillaemdashcell wall appositions depositedon the inner surface of epidermal cell walls directly beneathappressoriamdashand by the death of attacked cells Attacked cellsthat form an effective papilla survive and much evidence (reviewedby Zeyen
et al
2002) indicates that papillae present a physicalandor chemical penetration barrier Papillae form where appres-sorium contact attempted penetration causes focused cytologicalresponses including reorganization of the cell cytoskeletonand cytoplasmic streaming leading to formation of a cytoplasmicaggregate from which papilla components are depositedPapillae have a callose matrix containing various inorganic andorganic constituents including proteins and autofluorescentphenolic compounds (Zeyen
et al
2002) A halo of altered stainingaffinity in the epidermal cell wall often surrounds the papilla sitePapillae are formed even in susceptible host genotypes suchas the barley cv Pallas (Lyngkjaeligr
et al
2001) used in the presentstudies but often these papillae are ineffective and the fungalpenetration peg passes through the deposit which remains asthe haustorial neck collar that seals the site of invasion Papillaeffectiveness may relate to the rate of deposition to quantitativeor qualitative characteristics or to a combination of these factorsEarly H
2
O
2
accumulation indicated by DAB (33
prime
-diaminobenzidine)staining (Thordal-Christensen
et al
1997) may be important in cellsignalling to initiate papilla formation and it appears particularlyimportant in effective papilla-based resistance either through itsantimicrobial toxicity or by contributing to papilla-strengtheningthrough the oxidative cross-linking and immobilization of structuralcompounds (reviewed by Huumlckelhoven and Kogel 2003)
In cerealndashpowdery mildew interactions barley alleles at the
Mla
locus which condition HR have been relatively well characterized(Chelkowski
et al
2003 Panstruga and Schulze-Lefert 2002)When avirulent isolates attack
Mla1
barley such as the nearisogenic line P01 derived from cv Pallas (Koslashlster
et al
1986) usedhere HR occurs rapidly before or soon after visible haustoria form
leading to death and collapse of the single attacked epidermalcell Death is preceded by cessation of cytoplasmic streamingfollowed by loss of ability to take up vital dyes or to plasmolyse(Koga
et al
1988) The onset of whole-cell autofluorescence is areliable and convenient indicator of cell death (Bushnell 1981Koga
et al
1988 Lyngkjaeligr
et al
2001 Zeyen
et al
1995) One ofthe first signs of incompatibility conditioned by
Mla1
(Vanacker
et al
2000) or
Mla3
(Thordal-Christensen
et al
1997) is the transientappearance (at 14ndash15 hai) of H
2
O
2
at attachment sites betweenattacked epidermal cells and underlying mesophyll This precedesthe appearance of H
2
O
2
throughout the epidermal cell (at around16 h) which is in turn followed by death (Vanacker
et al
2000)Previous work suggested the possibility that the rapid signalling
events occurring in the challenged epidermis and underlyingmesophyll may arise from NO generation (Vanacker
et al
2000)Here we used confocal laser scanning electron microscopy (CLSM)to visualize NO generation associated with both papilla formationand HR during the interaction of
B graminis
f sp
hordei
andbarley by applying the stain DAF-2DA (45-diaminofluorescein-2-diacetate) which forms fluorescent DAF-2T (diaminotriazoloflu-orescein) in the presence of NO (Kojima
et al
1998ab) ReducingNO levels by treatment with the NO scavenger C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxidepotassium salt) (Delledonne
et al
1998 Garces
et al
2001Neill
et al
2003) suppressed penetration resistance and slightlydelayed HR Conversely when additional NO was providedby sodium nitroprusside (SNP Clarke
et al
2000 Garces
et al
2001 Pedroso
et al
2000) HR was slightly accelerated althoughpenetration resistance was unaffected Our data indicate roles forNO in both papilla defence and cell death
RESULTS
CLSM visualization of NO generation in barley attacked by
B graminis
CSLM after DAF-2DA staining of both Pallas (susceptible) and P01(
Mla1
resistance) revealed intense localized fluorescence indica-tive of NO generation at barley epidermal cell contact sites with
B graminis
f sp
hordei
appressoria Between 6 and 24 hai imagesof 157 and 120 germlings on Pallas and P01 respectively werecaptured before and after staining Visual inspection indicatedthat NO generation was an early response that became abruptlyevident as appressoria matured in both barley lines Thus up to9 hai there was no evidence for NO generation associated withimmature appressorial germ tubes However at 10ndash12 hai beforestrong autofluorescence indicative of phenolic compound accumula-tion was evident (Fig 1A) staining often led to localized pronouncedfluorescence beneath appressoria (Fig 1B) An intense fluorescentfocus subtending appressoria suggested NO accumulation in plantcell cytoplasmic aggregates and papillae and this focus was often
NO and resistanceresponse in barley
67
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
encircled by a less intense zone corresponding to the cell wallhalo (Fig 1B) The absence of DAF-2T fluorescence within thehook-shaped appressoria indicated that NO was primarily localizedto the plant cell However restricted NO generation appearedtransient because intensified fluorescence after staining becameincreasingly difficult to discern as localized autofluorescencedeveloped By 24 hai when intense autofluorescence wasevident (Fig 1C) staining did not appear to increase in intensity(Fig 1D)
Overview images (Fig 1) of localized fluorescence responsesdid not show the depth of NO generation within response sitesand thus three-dimensional models were created from transverseoptical sections made through such sites Figure 2(A) shows arepresentative example of an appressorial attack site examined14 hai As was common at this time (Fig 1) before stainingthe site was non-fluorescent indicating the absence of phenolicautofluorogens (data not shown) but after staining the overviewimage showed a characteristically circular zone of intensefluorescence indicative of NO generation in the region of papillaformation and the halo (as in Fig 1B) The transverse opticalsection suggested that NO generation was integral to the devel-oping papilla with little detectable DAF-2T fluorescence in thesurrounding cell wall and cytoplasm The transmitted light imagewas insufficiently detailed to resolve details of fungal structurebut it seems reasonable to suppose that the thickened region ofNO generation subtended the site of penetration peg emergencefrom the appressorial lobe Figure 2(B) shows before and afterDAF-2DA staining of a representative site of response to appres-sorial attack at 18 hai when deposition of the mature papillawould have been completed As shown previously (Fig 1) inoverview the only fluorescence detected was associated withautofluorescent phenolic compounds which accumulated in thepapilla and halo region and transverse optical sections of the site
were also virtually indistinguishable before and after stainingIt is also notable that the distribution of DAF-2T fluorescence inFig 2(A) and that of autofluorogens in Fig 2(B) was strikinglysimilar in shape and size This provides good evidence that NOdistribution at sites of early response to penetration is closelycorrelated with the subcellular locality in which components ofpapillae and haloes accumulate at maturity
Fluorescence intensity values (corrected for background) fromcontact sites that met the stringent criteria of acceptability (seeExperimental procedures) were obtained before and after DAF-2DA staining These provided objective data on the developmentof autofluorescence and fluorescence due to DAF-2DA stainingwithin separate time intervals over the period 6ndash24 hai (Fig 3)Thus for each individual contact site the fluorescence intensityvalue before and after staining was obtained and the differencebetween these values was calculated for each site Within theseparate time intervals the data for difference were subjected tostatistical analysis (see Experimental procedures) which showedvery similar trends in Pallas and P01 Appressorial germ tubecontact sites showed no localized fluorescence before 10 hai(before appressoria matured) but at 10ndash12 hai when autoflu-orescence was barely detectable in either barley line contactsites showed substantial elevated fluorescence after stainingindicative of NO generation Significant increases in fluorescenceafter staining were also evident at 13ndash14 and at 15ndash16 hai inP01 indicating continued NO production throughout this periodHowever in later samples there was no significant increase influorescence after DAF-2DA staining The data therefore suggesta burst of NO generation commencing very soon after appressorialmaturation but declining to undetectable levels from around 14 haionwards in Pallas and from 16 hai in P01
Interestingly at 17ndash19 hai staining led to a slight decreasein fluorescence intensity in both lines and in Pallas this effect was
Fig 1 Localized NO generation in Pallas (susceptible) barley epidermal cells at the B graminis appressorial germ tube contact site CLSM images of localized responses to B graminis attack 12 and 24 h after inoculation (hai) before and after DAF-2DA staining to reveal NO generation Ap = appressorium contact site C = conidium Ha = halo P = primary germ tube contact site (A) At 12 hai before staining a faint image of the fungus is evident and the P and Ap contact sites show faint autofluorescence (B) As in A after staining NO generation in the region of papilla deposition is indicated by intense fluorescence at the Ap site and fainter fluorescence is present in the halo region (C) At 24 hai before staining the P and Ap sites show strong autofluorescence in papillae and fainter autofluorescent haloes (D) As in C after staining there is no intensification of fluorescence indicating absence of NO Scale bars = 20 microM
68
E PRATS
et al
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
significant This suggests that components of the staining solu-tion either caused quenching or removed autofluorogens fromresponse sites To test this comparisons were made between thefluorescence intensity of individual papilla sites sampled in theperiods 15ndash16 and 17ndash19 hai before and after treatment withbuffer alone or treatment with buffer plus DMSO but omittingDAF-2DA Treatment with buffer alone had no effect on fluores-cence intensity but treatment with buffer plus DMSO led to asignificant decrease in intensity (
P
lt 0001) in papilla sites treated15ndash16 hai (data not shown) Because the data for intensityafter staining (Fig 3) represent the sum of fluorescence due toformation of DAF-2T and autofluorescence any reduction ofautofluorescence due to quenching or removal would decreasethe post-staining intensity value Thus the data probably under-estimate the contribution resulting from DAF-2T formation due to
Fig 3 Quantification of localized fluorescence responses at B graminis appressorium contact sites in Pallas (susceptible) and P01 (Mla1-resistant) barley epidermal cells Localized fluorescence intensities at appressorial contact sites on Pallas and P01 were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
Fig 2 Three-dimensional models of early localized NO generation and mature papilla structure at B graminis appressorial germ tube contact sites in Pallas barley (A) Overview image of an appressorium and host response site at 14 hai stained with DAF-2DA (no autofluorescence was present before staining image not shown) Intense green fluorescence indicates the presence of NO in the central region underlying the appressorial lobe (AL) at the tip of the appressorial germ tube (AGT) This is surrounded by a less intensely stained halo The red line marks the plane of a transverse optical section shown in the lower pane This shows a vertically thickened fluorescent region within the site of papilla deposition and a thinner region of peripheral staining corresponding to the halo (B) Upper images show overviews captured at 18 hai of an appressorium overlying a response site with a mature papilla Autofluorescence in the unstained specimen (left) indicates the presence of phenolic compounds and (right) no further fluorescence after DAF-2DA treatment The red lines mark the position of the transverse optical sections shown in the corresponding lower images which also show that staining did not add to fluorescence The dimensions and shape of autofluorescence associated with the mature papilla and halo correspond closely to the distribution of NO seen earlier (compare with A)
NO and resistanceresponse in barley
69
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
NO generation This does not however invalidate the conclusionthat NO was generated in an early but transient burst
In
Mla1
barley whole-cell autofluorescence indicating celldeath due to HR is first evident around 15ndash18 hai (Koga
et al
1990 Vanacker
et al
2000) but in susceptible Pallas cell deathis very infrequent Here cell death was seen so rarely in Pallasthat no attempt was made to study it In the current study visualinspection of P01 images also showed little evidence of whole-cell autofluorescence until after 15 hai but before this DAF-2DA staining often led to whole-cell fluorescence (Fig 4B)suggesting NO generation throughout some cells under attackfrom appressoria (Fig 4AB) These data suggested that whole-cell NO generation preceded autofluorogen accumulation in cellsundergoing HR (Fig 4A) As for localized responses this whole-cell generation of NO in P01 also appeared transient becomingincreasingly difficult to discern as autofluorescence developedThus by 24 hai when intense whole-cell autofluorescence wasevident (Fig 4C) staining added little if anything to the fluores-cence (Fig 4D)
Fluorescence intensity data from individual whole cells examinedbefore and after staining supported the conclusion that transientwhole-cell NO generation preceded cell death Figure 5 showsthat whole-cell autofluorescence intensities were very low insamples examined before 15 hai Nevertheless the
t
-test ofdifferences between whole-cell fluorescence intensity before and
Fig 4 Whole-cell fluorescence in P01 (Mla1-resistant) barley epidermis following challenge with B graminis CLSM images of whole-cell responses to B graminis attack 14 and 23 hai before and after DAF-2DA staining Ap = appressorium contact site P = primary germ tube contact site (A) At 14 hai before staining the P site shows distinct localized autofluorescence but the Ap site shows little and there is no whole-cell autofluorescence of the attacked cell (arrowhead) A second cell also without whole-cell autofluorescence is marked (double arrowhead) (B) As in A after staining the cell attacked by the appressorium (which was displaced slightly during staining) shows whole-cell NO generation indicated by its whole-cell fluorescence this is also evident in the second cell (double arrowhead) that was attacked out of view (C) At 23 hai before staining the P site shows distinct localized autofluorescence and the cell attacked by the appressorium shows whole-cell autofluorescence (D) As in C after staining there is no apparent intensification of fluorescence indicating absence of NO Scale bars = 50 microm
Fig 5 Quantification of whole-cell fluorescence responses to B graminis attack in P01 (Mla1-resistant) barley epidermal cells Whole-cell fluorescence intensities of attacked epidermal cells were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
70 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
after staining showed significantly increased whole-cell fluores-cence intensity following DAF-2DA at 10ndash12 hai and an evenmore marked and significant effect in the period 13ndash14 haiThis suggested a substantial burst of NO generation at a veryearly stage when the low intensity of whole-cell autofluorescenceindicated that cells remained alive From 15 hai howeverstatistical analysis showed no significant effect of DAF-2DAstaining indicating cessation of NO generation in cells that weredying or dead as indicated by their relatively intense whole-cellautofluorescence
C-PTIO suppresses NO accumulation as visualized by CLSM
The work above assumed that intensified fluorescence afterDAF-2DA staining was due to NO To test this CLSM was used ina second study of P01 over the critical period 10ndash19 hai buthere leaf segments were first bathed in the NO scavenger C-PTIObefore applying DAF-2DA
As previously (Fig 3) Fig 6 shows that the fluorescence intensityof papilla sites obtained before treatment indicating autofluores-cence intensity was low at 10ndash12 hai but increased thereafterHowever when treated with C-PTIO there was no detectableincrease in intensity of localized fluorescence following subse-quent DAF-2DA staining even over the period 10ndash14 hai And
as previously (Fig 3) staining samples 17ndash19 hai caused a lossof fluorescence
When C-PTIO was applied before staining no cells showed whole-cell fluorescence after C-PTIODAF-2DA treatment unless theyshowed whole-cell autofluorescence before treatment Over theperiod 10ndash19 hai 20 cells showing faint or obvious whole-cellautofluorescence were imaged and there was no statisticallysignificant increase in whole-cell fluorescence after treatment evenin the 10ndash14 hai period when the previous experiment showedmarked intensification (Fig 5) Together these observationsindicate that C-PTIO effectively removed NO from P01 barley cellsundergoing HR
L-NAME does not suppress NO accumulation as visualized by CLSM
Mammalian NOS is effectively inhibited by L-NAME which alsoinhibits some NO-related defensive responses in certain dicotyle-donous plants (Chandok et al 2003 Duumlrner et al 1998 Guo et al2003) In a similar experiment to that performed with C-PTIO(above) L-NAME was applied for 30 min prior to applying DAF-2DAin samples examined over the critical period 10ndash19 hai Figure 7shows data obtained from papilla sites demonstrating that thetemporal pattern of autofluorescence development was verysimilar to that observed in earlier experiments (Figs 3 and 6)intensifying from low to much higher levels over this period
Fig 6 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of the NO scavenger C-PTIO and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before C-PTIODAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
Fig 7 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of L-NAME and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before L-NAMEDAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
NO and resistanceresponse in barley 71
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Patterns of NO generation were also very similar to those seenin untreated leaves (Fig 3) again indicating a transient burst ofgeneration Hence DAF-2DA staining led to significant increasesin fluorescence intensity indicative of NO generation in papillasites over the period 10ndash14 hai a near-significant increase wasfound in samples examined 15ndash16 hai but thereafter therewas no evidence for NO generation Therefore unlike C-PTIO therewas no evidence for an immediate effect of L-NAME treatment onNO generation at sites of papilla formation Furthermore althoughthe number of cells undergoing HR was too small for reliablestatistical analyses there was no indication that L-NAME affectedwhole-cell fluorescence responses Thus as in untreated leaves(Fig 5) treatment did not diminish the early whole-cell fluores-cence revealed by DAF-2DA staining
Light and fluorescence microscope analyses of effects of C-PTIO L-NAME and SNP on B graminis development and barley epidermal cell responses to attack
To investigate the role(s) of NO generated in the interaction ofB graminis with barley an NO scavenger (C-PTIO) a NOS inhib-itor (L-NAME) and an NO donor (SNP) were supplied separatelyto cut leaves Light microscopy showed that as compared withcontrols treating leaves with any of the test chemicals had noeffects on spore germination or subsequent germling develop-ment up to the differentiation of mature (hooked) appressoria(data not shown) Furthermore incident fluorescence microscopyindicated that none of the chemicals affected the timing frequency
or intensity of localized autofluorescent host cell responses asso-ciated with appressoria Detailed data for these responses are notgiven but in all cases approximately 10 of appressorial contactsites were associated with faint localized autofluorescent hostcell responses by 12 hai and by 24 hai approximately 60 ofsites showed autofluorescence which in most cases was judgedfaint or moderately intense (class 1 or 2 on the 0ndash3 subjectivescale) Other effects of each test chemical are considered in turn
Effects of C-PTIOA few very small haustoria (swollen apices of penetration pegs)indicating successful penetration were evident in epidermalcells of both barley lines by 12 hai and in both there was littleincrease in penetration frequency after 15ndash18 hai (Fig 8) Thisindicates that the outcome of attacks and therefore efficacy ofpenetration resistance was determined by this time Comparedwith controls overall analysis showed that C-PTIO treatmentsignificantly increased (P lt 0001) the frequency of successfulpenetration ie suppressed penetration resistance in bothbarley lines (Fig 8) Further analyses showed that this effectwas significant (P lt 005) at most individual sampling times Inboth Pallas and P01 successful penetration was associatedwith approximately 30 of appressoria in C-PTIO-treated leaveswhereas control values were around 15 In Pallas some cells diedfollowing successful penetration but this percentage remainedrelatively low throughout (only c 20ndash40 died Fig 8) andtreatment had no significant effect By contrast in P01 virtually allpenetrated haustorium-containing cells were dead by 18 hai(Fig 8) However at 12ndash15 hai some penetrated P01 cells showed
Fig 8 C-PTIO suppression of B graminis penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO scavenger C-PTIO (triangles) or water (circles) on percentages of B graminis appressoria with successful penetrations (haustoria) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (indicated by whole-cell autofluorescence) 12ndash30 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001
72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

NO and resistanceresponse in barley
67
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
encircled by a less intense zone corresponding to the cell wallhalo (Fig 1B) The absence of DAF-2T fluorescence within thehook-shaped appressoria indicated that NO was primarily localizedto the plant cell However restricted NO generation appearedtransient because intensified fluorescence after staining becameincreasingly difficult to discern as localized autofluorescencedeveloped By 24 hai when intense autofluorescence wasevident (Fig 1C) staining did not appear to increase in intensity(Fig 1D)
Overview images (Fig 1) of localized fluorescence responsesdid not show the depth of NO generation within response sitesand thus three-dimensional models were created from transverseoptical sections made through such sites Figure 2(A) shows arepresentative example of an appressorial attack site examined14 hai As was common at this time (Fig 1) before stainingthe site was non-fluorescent indicating the absence of phenolicautofluorogens (data not shown) but after staining the overviewimage showed a characteristically circular zone of intensefluorescence indicative of NO generation in the region of papillaformation and the halo (as in Fig 1B) The transverse opticalsection suggested that NO generation was integral to the devel-oping papilla with little detectable DAF-2T fluorescence in thesurrounding cell wall and cytoplasm The transmitted light imagewas insufficiently detailed to resolve details of fungal structurebut it seems reasonable to suppose that the thickened region ofNO generation subtended the site of penetration peg emergencefrom the appressorial lobe Figure 2(B) shows before and afterDAF-2DA staining of a representative site of response to appres-sorial attack at 18 hai when deposition of the mature papillawould have been completed As shown previously (Fig 1) inoverview the only fluorescence detected was associated withautofluorescent phenolic compounds which accumulated in thepapilla and halo region and transverse optical sections of the site
were also virtually indistinguishable before and after stainingIt is also notable that the distribution of DAF-2T fluorescence inFig 2(A) and that of autofluorogens in Fig 2(B) was strikinglysimilar in shape and size This provides good evidence that NOdistribution at sites of early response to penetration is closelycorrelated with the subcellular locality in which components ofpapillae and haloes accumulate at maturity
Fluorescence intensity values (corrected for background) fromcontact sites that met the stringent criteria of acceptability (seeExperimental procedures) were obtained before and after DAF-2DA staining These provided objective data on the developmentof autofluorescence and fluorescence due to DAF-2DA stainingwithin separate time intervals over the period 6ndash24 hai (Fig 3)Thus for each individual contact site the fluorescence intensityvalue before and after staining was obtained and the differencebetween these values was calculated for each site Within theseparate time intervals the data for difference were subjected tostatistical analysis (see Experimental procedures) which showedvery similar trends in Pallas and P01 Appressorial germ tubecontact sites showed no localized fluorescence before 10 hai(before appressoria matured) but at 10ndash12 hai when autoflu-orescence was barely detectable in either barley line contactsites showed substantial elevated fluorescence after stainingindicative of NO generation Significant increases in fluorescenceafter staining were also evident at 13ndash14 and at 15ndash16 hai inP01 indicating continued NO production throughout this periodHowever in later samples there was no significant increase influorescence after DAF-2DA staining The data therefore suggesta burst of NO generation commencing very soon after appressorialmaturation but declining to undetectable levels from around 14 haionwards in Pallas and from 16 hai in P01
Interestingly at 17ndash19 hai staining led to a slight decreasein fluorescence intensity in both lines and in Pallas this effect was
Fig 1 Localized NO generation in Pallas (susceptible) barley epidermal cells at the B graminis appressorial germ tube contact site CLSM images of localized responses to B graminis attack 12 and 24 h after inoculation (hai) before and after DAF-2DA staining to reveal NO generation Ap = appressorium contact site C = conidium Ha = halo P = primary germ tube contact site (A) At 12 hai before staining a faint image of the fungus is evident and the P and Ap contact sites show faint autofluorescence (B) As in A after staining NO generation in the region of papilla deposition is indicated by intense fluorescence at the Ap site and fainter fluorescence is present in the halo region (C) At 24 hai before staining the P and Ap sites show strong autofluorescence in papillae and fainter autofluorescent haloes (D) As in C after staining there is no intensification of fluorescence indicating absence of NO Scale bars = 20 microM
68
E PRATS
et al
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
significant This suggests that components of the staining solu-tion either caused quenching or removed autofluorogens fromresponse sites To test this comparisons were made between thefluorescence intensity of individual papilla sites sampled in theperiods 15ndash16 and 17ndash19 hai before and after treatment withbuffer alone or treatment with buffer plus DMSO but omittingDAF-2DA Treatment with buffer alone had no effect on fluores-cence intensity but treatment with buffer plus DMSO led to asignificant decrease in intensity (
P
lt 0001) in papilla sites treated15ndash16 hai (data not shown) Because the data for intensityafter staining (Fig 3) represent the sum of fluorescence due toformation of DAF-2T and autofluorescence any reduction ofautofluorescence due to quenching or removal would decreasethe post-staining intensity value Thus the data probably under-estimate the contribution resulting from DAF-2T formation due to
Fig 3 Quantification of localized fluorescence responses at B graminis appressorium contact sites in Pallas (susceptible) and P01 (Mla1-resistant) barley epidermal cells Localized fluorescence intensities at appressorial contact sites on Pallas and P01 were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
Fig 2 Three-dimensional models of early localized NO generation and mature papilla structure at B graminis appressorial germ tube contact sites in Pallas barley (A) Overview image of an appressorium and host response site at 14 hai stained with DAF-2DA (no autofluorescence was present before staining image not shown) Intense green fluorescence indicates the presence of NO in the central region underlying the appressorial lobe (AL) at the tip of the appressorial germ tube (AGT) This is surrounded by a less intensely stained halo The red line marks the plane of a transverse optical section shown in the lower pane This shows a vertically thickened fluorescent region within the site of papilla deposition and a thinner region of peripheral staining corresponding to the halo (B) Upper images show overviews captured at 18 hai of an appressorium overlying a response site with a mature papilla Autofluorescence in the unstained specimen (left) indicates the presence of phenolic compounds and (right) no further fluorescence after DAF-2DA treatment The red lines mark the position of the transverse optical sections shown in the corresponding lower images which also show that staining did not add to fluorescence The dimensions and shape of autofluorescence associated with the mature papilla and halo correspond closely to the distribution of NO seen earlier (compare with A)
NO and resistanceresponse in barley
69
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
NO generation This does not however invalidate the conclusionthat NO was generated in an early but transient burst
In
Mla1
barley whole-cell autofluorescence indicating celldeath due to HR is first evident around 15ndash18 hai (Koga
et al
1990 Vanacker
et al
2000) but in susceptible Pallas cell deathis very infrequent Here cell death was seen so rarely in Pallasthat no attempt was made to study it In the current study visualinspection of P01 images also showed little evidence of whole-cell autofluorescence until after 15 hai but before this DAF-2DA staining often led to whole-cell fluorescence (Fig 4B)suggesting NO generation throughout some cells under attackfrom appressoria (Fig 4AB) These data suggested that whole-cell NO generation preceded autofluorogen accumulation in cellsundergoing HR (Fig 4A) As for localized responses this whole-cell generation of NO in P01 also appeared transient becomingincreasingly difficult to discern as autofluorescence developedThus by 24 hai when intense whole-cell autofluorescence wasevident (Fig 4C) staining added little if anything to the fluores-cence (Fig 4D)
Fluorescence intensity data from individual whole cells examinedbefore and after staining supported the conclusion that transientwhole-cell NO generation preceded cell death Figure 5 showsthat whole-cell autofluorescence intensities were very low insamples examined before 15 hai Nevertheless the
t
-test ofdifferences between whole-cell fluorescence intensity before and
Fig 4 Whole-cell fluorescence in P01 (Mla1-resistant) barley epidermis following challenge with B graminis CLSM images of whole-cell responses to B graminis attack 14 and 23 hai before and after DAF-2DA staining Ap = appressorium contact site P = primary germ tube contact site (A) At 14 hai before staining the P site shows distinct localized autofluorescence but the Ap site shows little and there is no whole-cell autofluorescence of the attacked cell (arrowhead) A second cell also without whole-cell autofluorescence is marked (double arrowhead) (B) As in A after staining the cell attacked by the appressorium (which was displaced slightly during staining) shows whole-cell NO generation indicated by its whole-cell fluorescence this is also evident in the second cell (double arrowhead) that was attacked out of view (C) At 23 hai before staining the P site shows distinct localized autofluorescence and the cell attacked by the appressorium shows whole-cell autofluorescence (D) As in C after staining there is no apparent intensification of fluorescence indicating absence of NO Scale bars = 50 microm
Fig 5 Quantification of whole-cell fluorescence responses to B graminis attack in P01 (Mla1-resistant) barley epidermal cells Whole-cell fluorescence intensities of attacked epidermal cells were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
70 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
after staining showed significantly increased whole-cell fluores-cence intensity following DAF-2DA at 10ndash12 hai and an evenmore marked and significant effect in the period 13ndash14 haiThis suggested a substantial burst of NO generation at a veryearly stage when the low intensity of whole-cell autofluorescenceindicated that cells remained alive From 15 hai howeverstatistical analysis showed no significant effect of DAF-2DAstaining indicating cessation of NO generation in cells that weredying or dead as indicated by their relatively intense whole-cellautofluorescence
C-PTIO suppresses NO accumulation as visualized by CLSM
The work above assumed that intensified fluorescence afterDAF-2DA staining was due to NO To test this CLSM was used ina second study of P01 over the critical period 10ndash19 hai buthere leaf segments were first bathed in the NO scavenger C-PTIObefore applying DAF-2DA
As previously (Fig 3) Fig 6 shows that the fluorescence intensityof papilla sites obtained before treatment indicating autofluores-cence intensity was low at 10ndash12 hai but increased thereafterHowever when treated with C-PTIO there was no detectableincrease in intensity of localized fluorescence following subse-quent DAF-2DA staining even over the period 10ndash14 hai And
as previously (Fig 3) staining samples 17ndash19 hai caused a lossof fluorescence
When C-PTIO was applied before staining no cells showed whole-cell fluorescence after C-PTIODAF-2DA treatment unless theyshowed whole-cell autofluorescence before treatment Over theperiod 10ndash19 hai 20 cells showing faint or obvious whole-cellautofluorescence were imaged and there was no statisticallysignificant increase in whole-cell fluorescence after treatment evenin the 10ndash14 hai period when the previous experiment showedmarked intensification (Fig 5) Together these observationsindicate that C-PTIO effectively removed NO from P01 barley cellsundergoing HR
L-NAME does not suppress NO accumulation as visualized by CLSM
Mammalian NOS is effectively inhibited by L-NAME which alsoinhibits some NO-related defensive responses in certain dicotyle-donous plants (Chandok et al 2003 Duumlrner et al 1998 Guo et al2003) In a similar experiment to that performed with C-PTIO(above) L-NAME was applied for 30 min prior to applying DAF-2DAin samples examined over the critical period 10ndash19 hai Figure 7shows data obtained from papilla sites demonstrating that thetemporal pattern of autofluorescence development was verysimilar to that observed in earlier experiments (Figs 3 and 6)intensifying from low to much higher levels over this period
Fig 6 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of the NO scavenger C-PTIO and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before C-PTIODAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
Fig 7 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of L-NAME and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before L-NAMEDAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
NO and resistanceresponse in barley 71
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Patterns of NO generation were also very similar to those seenin untreated leaves (Fig 3) again indicating a transient burst ofgeneration Hence DAF-2DA staining led to significant increasesin fluorescence intensity indicative of NO generation in papillasites over the period 10ndash14 hai a near-significant increase wasfound in samples examined 15ndash16 hai but thereafter therewas no evidence for NO generation Therefore unlike C-PTIO therewas no evidence for an immediate effect of L-NAME treatment onNO generation at sites of papilla formation Furthermore althoughthe number of cells undergoing HR was too small for reliablestatistical analyses there was no indication that L-NAME affectedwhole-cell fluorescence responses Thus as in untreated leaves(Fig 5) treatment did not diminish the early whole-cell fluores-cence revealed by DAF-2DA staining
Light and fluorescence microscope analyses of effects of C-PTIO L-NAME and SNP on B graminis development and barley epidermal cell responses to attack
To investigate the role(s) of NO generated in the interaction ofB graminis with barley an NO scavenger (C-PTIO) a NOS inhib-itor (L-NAME) and an NO donor (SNP) were supplied separatelyto cut leaves Light microscopy showed that as compared withcontrols treating leaves with any of the test chemicals had noeffects on spore germination or subsequent germling develop-ment up to the differentiation of mature (hooked) appressoria(data not shown) Furthermore incident fluorescence microscopyindicated that none of the chemicals affected the timing frequency
or intensity of localized autofluorescent host cell responses asso-ciated with appressoria Detailed data for these responses are notgiven but in all cases approximately 10 of appressorial contactsites were associated with faint localized autofluorescent hostcell responses by 12 hai and by 24 hai approximately 60 ofsites showed autofluorescence which in most cases was judgedfaint or moderately intense (class 1 or 2 on the 0ndash3 subjectivescale) Other effects of each test chemical are considered in turn
Effects of C-PTIOA few very small haustoria (swollen apices of penetration pegs)indicating successful penetration were evident in epidermalcells of both barley lines by 12 hai and in both there was littleincrease in penetration frequency after 15ndash18 hai (Fig 8) Thisindicates that the outcome of attacks and therefore efficacy ofpenetration resistance was determined by this time Comparedwith controls overall analysis showed that C-PTIO treatmentsignificantly increased (P lt 0001) the frequency of successfulpenetration ie suppressed penetration resistance in bothbarley lines (Fig 8) Further analyses showed that this effectwas significant (P lt 005) at most individual sampling times Inboth Pallas and P01 successful penetration was associatedwith approximately 30 of appressoria in C-PTIO-treated leaveswhereas control values were around 15 In Pallas some cells diedfollowing successful penetration but this percentage remainedrelatively low throughout (only c 20ndash40 died Fig 8) andtreatment had no significant effect By contrast in P01 virtually allpenetrated haustorium-containing cells were dead by 18 hai(Fig 8) However at 12ndash15 hai some penetrated P01 cells showed
Fig 8 C-PTIO suppression of B graminis penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO scavenger C-PTIO (triangles) or water (circles) on percentages of B graminis appressoria with successful penetrations (haustoria) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (indicated by whole-cell autofluorescence) 12ndash30 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001
72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972
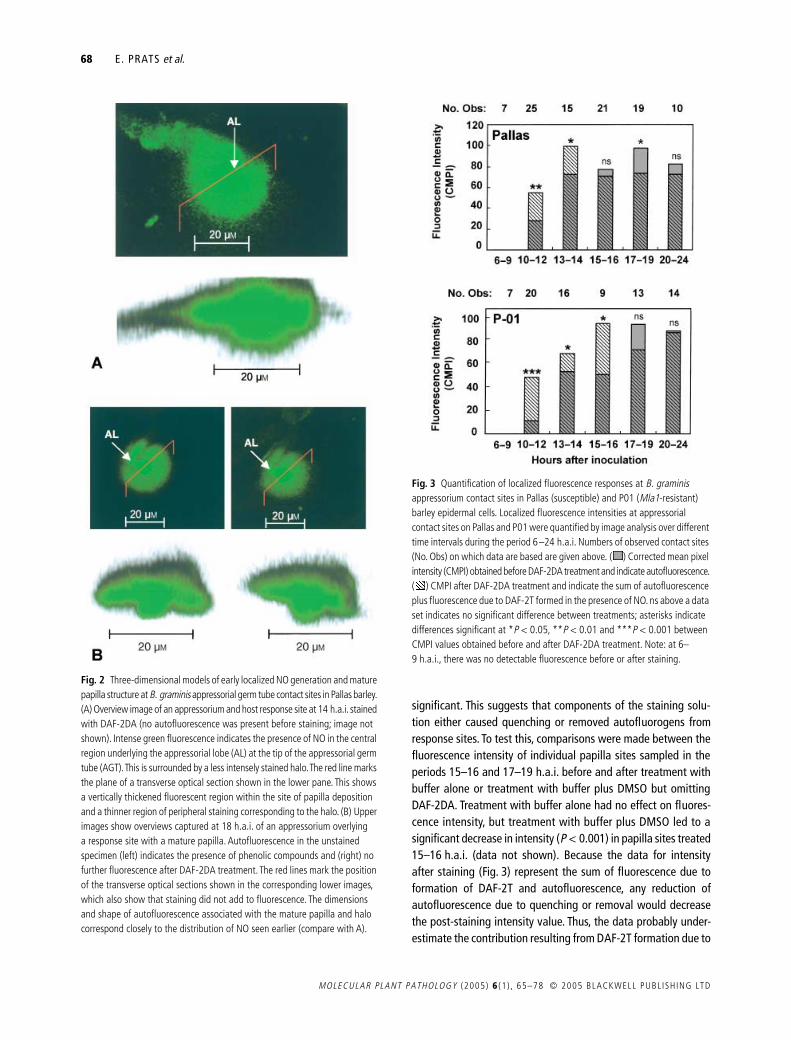
68
E PRATS
et al
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
significant This suggests that components of the staining solu-tion either caused quenching or removed autofluorogens fromresponse sites To test this comparisons were made between thefluorescence intensity of individual papilla sites sampled in theperiods 15ndash16 and 17ndash19 hai before and after treatment withbuffer alone or treatment with buffer plus DMSO but omittingDAF-2DA Treatment with buffer alone had no effect on fluores-cence intensity but treatment with buffer plus DMSO led to asignificant decrease in intensity (
P
lt 0001) in papilla sites treated15ndash16 hai (data not shown) Because the data for intensityafter staining (Fig 3) represent the sum of fluorescence due toformation of DAF-2T and autofluorescence any reduction ofautofluorescence due to quenching or removal would decreasethe post-staining intensity value Thus the data probably under-estimate the contribution resulting from DAF-2T formation due to
Fig 3 Quantification of localized fluorescence responses at B graminis appressorium contact sites in Pallas (susceptible) and P01 (Mla1-resistant) barley epidermal cells Localized fluorescence intensities at appressorial contact sites on Pallas and P01 were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
Fig 2 Three-dimensional models of early localized NO generation and mature papilla structure at B graminis appressorial germ tube contact sites in Pallas barley (A) Overview image of an appressorium and host response site at 14 hai stained with DAF-2DA (no autofluorescence was present before staining image not shown) Intense green fluorescence indicates the presence of NO in the central region underlying the appressorial lobe (AL) at the tip of the appressorial germ tube (AGT) This is surrounded by a less intensely stained halo The red line marks the plane of a transverse optical section shown in the lower pane This shows a vertically thickened fluorescent region within the site of papilla deposition and a thinner region of peripheral staining corresponding to the halo (B) Upper images show overviews captured at 18 hai of an appressorium overlying a response site with a mature papilla Autofluorescence in the unstained specimen (left) indicates the presence of phenolic compounds and (right) no further fluorescence after DAF-2DA treatment The red lines mark the position of the transverse optical sections shown in the corresponding lower images which also show that staining did not add to fluorescence The dimensions and shape of autofluorescence associated with the mature papilla and halo correspond closely to the distribution of NO seen earlier (compare with A)
NO and resistanceresponse in barley
69
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
NO generation This does not however invalidate the conclusionthat NO was generated in an early but transient burst
In
Mla1
barley whole-cell autofluorescence indicating celldeath due to HR is first evident around 15ndash18 hai (Koga
et al
1990 Vanacker
et al
2000) but in susceptible Pallas cell deathis very infrequent Here cell death was seen so rarely in Pallasthat no attempt was made to study it In the current study visualinspection of P01 images also showed little evidence of whole-cell autofluorescence until after 15 hai but before this DAF-2DA staining often led to whole-cell fluorescence (Fig 4B)suggesting NO generation throughout some cells under attackfrom appressoria (Fig 4AB) These data suggested that whole-cell NO generation preceded autofluorogen accumulation in cellsundergoing HR (Fig 4A) As for localized responses this whole-cell generation of NO in P01 also appeared transient becomingincreasingly difficult to discern as autofluorescence developedThus by 24 hai when intense whole-cell autofluorescence wasevident (Fig 4C) staining added little if anything to the fluores-cence (Fig 4D)
Fluorescence intensity data from individual whole cells examinedbefore and after staining supported the conclusion that transientwhole-cell NO generation preceded cell death Figure 5 showsthat whole-cell autofluorescence intensities were very low insamples examined before 15 hai Nevertheless the
t
-test ofdifferences between whole-cell fluorescence intensity before and
Fig 4 Whole-cell fluorescence in P01 (Mla1-resistant) barley epidermis following challenge with B graminis CLSM images of whole-cell responses to B graminis attack 14 and 23 hai before and after DAF-2DA staining Ap = appressorium contact site P = primary germ tube contact site (A) At 14 hai before staining the P site shows distinct localized autofluorescence but the Ap site shows little and there is no whole-cell autofluorescence of the attacked cell (arrowhead) A second cell also without whole-cell autofluorescence is marked (double arrowhead) (B) As in A after staining the cell attacked by the appressorium (which was displaced slightly during staining) shows whole-cell NO generation indicated by its whole-cell fluorescence this is also evident in the second cell (double arrowhead) that was attacked out of view (C) At 23 hai before staining the P site shows distinct localized autofluorescence and the cell attacked by the appressorium shows whole-cell autofluorescence (D) As in C after staining there is no apparent intensification of fluorescence indicating absence of NO Scale bars = 50 microm
Fig 5 Quantification of whole-cell fluorescence responses to B graminis attack in P01 (Mla1-resistant) barley epidermal cells Whole-cell fluorescence intensities of attacked epidermal cells were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
70 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
after staining showed significantly increased whole-cell fluores-cence intensity following DAF-2DA at 10ndash12 hai and an evenmore marked and significant effect in the period 13ndash14 haiThis suggested a substantial burst of NO generation at a veryearly stage when the low intensity of whole-cell autofluorescenceindicated that cells remained alive From 15 hai howeverstatistical analysis showed no significant effect of DAF-2DAstaining indicating cessation of NO generation in cells that weredying or dead as indicated by their relatively intense whole-cellautofluorescence
C-PTIO suppresses NO accumulation as visualized by CLSM
The work above assumed that intensified fluorescence afterDAF-2DA staining was due to NO To test this CLSM was used ina second study of P01 over the critical period 10ndash19 hai buthere leaf segments were first bathed in the NO scavenger C-PTIObefore applying DAF-2DA
As previously (Fig 3) Fig 6 shows that the fluorescence intensityof papilla sites obtained before treatment indicating autofluores-cence intensity was low at 10ndash12 hai but increased thereafterHowever when treated with C-PTIO there was no detectableincrease in intensity of localized fluorescence following subse-quent DAF-2DA staining even over the period 10ndash14 hai And
as previously (Fig 3) staining samples 17ndash19 hai caused a lossof fluorescence
When C-PTIO was applied before staining no cells showed whole-cell fluorescence after C-PTIODAF-2DA treatment unless theyshowed whole-cell autofluorescence before treatment Over theperiod 10ndash19 hai 20 cells showing faint or obvious whole-cellautofluorescence were imaged and there was no statisticallysignificant increase in whole-cell fluorescence after treatment evenin the 10ndash14 hai period when the previous experiment showedmarked intensification (Fig 5) Together these observationsindicate that C-PTIO effectively removed NO from P01 barley cellsundergoing HR
L-NAME does not suppress NO accumulation as visualized by CLSM
Mammalian NOS is effectively inhibited by L-NAME which alsoinhibits some NO-related defensive responses in certain dicotyle-donous plants (Chandok et al 2003 Duumlrner et al 1998 Guo et al2003) In a similar experiment to that performed with C-PTIO(above) L-NAME was applied for 30 min prior to applying DAF-2DAin samples examined over the critical period 10ndash19 hai Figure 7shows data obtained from papilla sites demonstrating that thetemporal pattern of autofluorescence development was verysimilar to that observed in earlier experiments (Figs 3 and 6)intensifying from low to much higher levels over this period
Fig 6 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of the NO scavenger C-PTIO and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before C-PTIODAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
Fig 7 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of L-NAME and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before L-NAMEDAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
NO and resistanceresponse in barley 71
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Patterns of NO generation were also very similar to those seenin untreated leaves (Fig 3) again indicating a transient burst ofgeneration Hence DAF-2DA staining led to significant increasesin fluorescence intensity indicative of NO generation in papillasites over the period 10ndash14 hai a near-significant increase wasfound in samples examined 15ndash16 hai but thereafter therewas no evidence for NO generation Therefore unlike C-PTIO therewas no evidence for an immediate effect of L-NAME treatment onNO generation at sites of papilla formation Furthermore althoughthe number of cells undergoing HR was too small for reliablestatistical analyses there was no indication that L-NAME affectedwhole-cell fluorescence responses Thus as in untreated leaves(Fig 5) treatment did not diminish the early whole-cell fluores-cence revealed by DAF-2DA staining
Light and fluorescence microscope analyses of effects of C-PTIO L-NAME and SNP on B graminis development and barley epidermal cell responses to attack
To investigate the role(s) of NO generated in the interaction ofB graminis with barley an NO scavenger (C-PTIO) a NOS inhib-itor (L-NAME) and an NO donor (SNP) were supplied separatelyto cut leaves Light microscopy showed that as compared withcontrols treating leaves with any of the test chemicals had noeffects on spore germination or subsequent germling develop-ment up to the differentiation of mature (hooked) appressoria(data not shown) Furthermore incident fluorescence microscopyindicated that none of the chemicals affected the timing frequency
or intensity of localized autofluorescent host cell responses asso-ciated with appressoria Detailed data for these responses are notgiven but in all cases approximately 10 of appressorial contactsites were associated with faint localized autofluorescent hostcell responses by 12 hai and by 24 hai approximately 60 ofsites showed autofluorescence which in most cases was judgedfaint or moderately intense (class 1 or 2 on the 0ndash3 subjectivescale) Other effects of each test chemical are considered in turn
Effects of C-PTIOA few very small haustoria (swollen apices of penetration pegs)indicating successful penetration were evident in epidermalcells of both barley lines by 12 hai and in both there was littleincrease in penetration frequency after 15ndash18 hai (Fig 8) Thisindicates that the outcome of attacks and therefore efficacy ofpenetration resistance was determined by this time Comparedwith controls overall analysis showed that C-PTIO treatmentsignificantly increased (P lt 0001) the frequency of successfulpenetration ie suppressed penetration resistance in bothbarley lines (Fig 8) Further analyses showed that this effectwas significant (P lt 005) at most individual sampling times Inboth Pallas and P01 successful penetration was associatedwith approximately 30 of appressoria in C-PTIO-treated leaveswhereas control values were around 15 In Pallas some cells diedfollowing successful penetration but this percentage remainedrelatively low throughout (only c 20ndash40 died Fig 8) andtreatment had no significant effect By contrast in P01 virtually allpenetrated haustorium-containing cells were dead by 18 hai(Fig 8) However at 12ndash15 hai some penetrated P01 cells showed
Fig 8 C-PTIO suppression of B graminis penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO scavenger C-PTIO (triangles) or water (circles) on percentages of B graminis appressoria with successful penetrations (haustoria) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (indicated by whole-cell autofluorescence) 12ndash30 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001
72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972
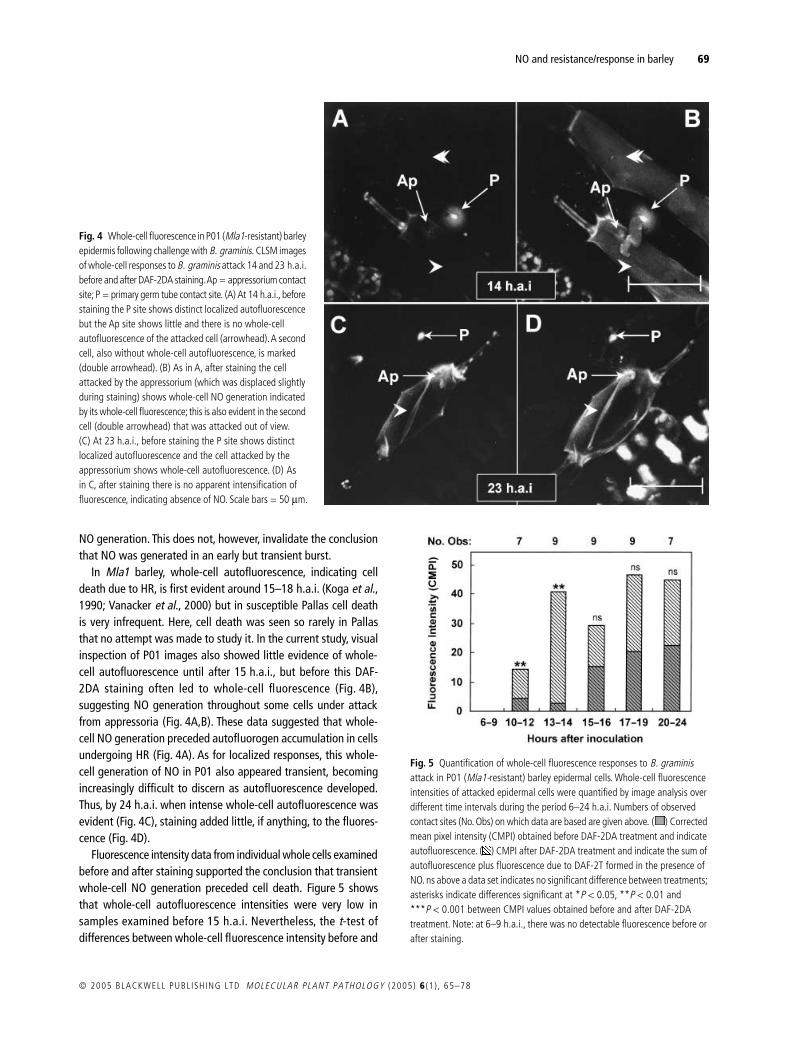
NO and resistanceresponse in barley
69
copy 2005 BLACKWELL PUBL ISH ING LTD
MOLECULAR PLANT PATHOLOGY
(2005)
6
(1 ) 65ndash78
NO generation This does not however invalidate the conclusionthat NO was generated in an early but transient burst
In
Mla1
barley whole-cell autofluorescence indicating celldeath due to HR is first evident around 15ndash18 hai (Koga
et al
1990 Vanacker
et al
2000) but in susceptible Pallas cell deathis very infrequent Here cell death was seen so rarely in Pallasthat no attempt was made to study it In the current study visualinspection of P01 images also showed little evidence of whole-cell autofluorescence until after 15 hai but before this DAF-2DA staining often led to whole-cell fluorescence (Fig 4B)suggesting NO generation throughout some cells under attackfrom appressoria (Fig 4AB) These data suggested that whole-cell NO generation preceded autofluorogen accumulation in cellsundergoing HR (Fig 4A) As for localized responses this whole-cell generation of NO in P01 also appeared transient becomingincreasingly difficult to discern as autofluorescence developedThus by 24 hai when intense whole-cell autofluorescence wasevident (Fig 4C) staining added little if anything to the fluores-cence (Fig 4D)
Fluorescence intensity data from individual whole cells examinedbefore and after staining supported the conclusion that transientwhole-cell NO generation preceded cell death Figure 5 showsthat whole-cell autofluorescence intensities were very low insamples examined before 15 hai Nevertheless the
t
-test ofdifferences between whole-cell fluorescence intensity before and
Fig 4 Whole-cell fluorescence in P01 (Mla1-resistant) barley epidermis following challenge with B graminis CLSM images of whole-cell responses to B graminis attack 14 and 23 hai before and after DAF-2DA staining Ap = appressorium contact site P = primary germ tube contact site (A) At 14 hai before staining the P site shows distinct localized autofluorescence but the Ap site shows little and there is no whole-cell autofluorescence of the attacked cell (arrowhead) A second cell also without whole-cell autofluorescence is marked (double arrowhead) (B) As in A after staining the cell attacked by the appressorium (which was displaced slightly during staining) shows whole-cell NO generation indicated by its whole-cell fluorescence this is also evident in the second cell (double arrowhead) that was attacked out of view (C) At 23 hai before staining the P site shows distinct localized autofluorescence and the cell attacked by the appressorium shows whole-cell autofluorescence (D) As in C after staining there is no apparent intensification of fluorescence indicating absence of NO Scale bars = 50 microm
Fig 5 Quantification of whole-cell fluorescence responses to B graminis attack in P01 (Mla1-resistant) barley epidermal cells Whole-cell fluorescence intensities of attacked epidermal cells were quantified by image analysis over different time intervals during the period 6ndash24 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before DAF-2DA treatment and indicate autofluorescence ( ) CMPI after DAF-2DA treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 between CMPI values obtained before and after DAF-2DA treatment Note at 6ndash9 hai there was no detectable fluorescence before or after staining
70 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
after staining showed significantly increased whole-cell fluores-cence intensity following DAF-2DA at 10ndash12 hai and an evenmore marked and significant effect in the period 13ndash14 haiThis suggested a substantial burst of NO generation at a veryearly stage when the low intensity of whole-cell autofluorescenceindicated that cells remained alive From 15 hai howeverstatistical analysis showed no significant effect of DAF-2DAstaining indicating cessation of NO generation in cells that weredying or dead as indicated by their relatively intense whole-cellautofluorescence
C-PTIO suppresses NO accumulation as visualized by CLSM
The work above assumed that intensified fluorescence afterDAF-2DA staining was due to NO To test this CLSM was used ina second study of P01 over the critical period 10ndash19 hai buthere leaf segments were first bathed in the NO scavenger C-PTIObefore applying DAF-2DA
As previously (Fig 3) Fig 6 shows that the fluorescence intensityof papilla sites obtained before treatment indicating autofluores-cence intensity was low at 10ndash12 hai but increased thereafterHowever when treated with C-PTIO there was no detectableincrease in intensity of localized fluorescence following subse-quent DAF-2DA staining even over the period 10ndash14 hai And
as previously (Fig 3) staining samples 17ndash19 hai caused a lossof fluorescence
When C-PTIO was applied before staining no cells showed whole-cell fluorescence after C-PTIODAF-2DA treatment unless theyshowed whole-cell autofluorescence before treatment Over theperiod 10ndash19 hai 20 cells showing faint or obvious whole-cellautofluorescence were imaged and there was no statisticallysignificant increase in whole-cell fluorescence after treatment evenin the 10ndash14 hai period when the previous experiment showedmarked intensification (Fig 5) Together these observationsindicate that C-PTIO effectively removed NO from P01 barley cellsundergoing HR
L-NAME does not suppress NO accumulation as visualized by CLSM
Mammalian NOS is effectively inhibited by L-NAME which alsoinhibits some NO-related defensive responses in certain dicotyle-donous plants (Chandok et al 2003 Duumlrner et al 1998 Guo et al2003) In a similar experiment to that performed with C-PTIO(above) L-NAME was applied for 30 min prior to applying DAF-2DAin samples examined over the critical period 10ndash19 hai Figure 7shows data obtained from papilla sites demonstrating that thetemporal pattern of autofluorescence development was verysimilar to that observed in earlier experiments (Figs 3 and 6)intensifying from low to much higher levels over this period
Fig 6 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of the NO scavenger C-PTIO and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before C-PTIODAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
Fig 7 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of L-NAME and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before L-NAMEDAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
NO and resistanceresponse in barley 71
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Patterns of NO generation were also very similar to those seenin untreated leaves (Fig 3) again indicating a transient burst ofgeneration Hence DAF-2DA staining led to significant increasesin fluorescence intensity indicative of NO generation in papillasites over the period 10ndash14 hai a near-significant increase wasfound in samples examined 15ndash16 hai but thereafter therewas no evidence for NO generation Therefore unlike C-PTIO therewas no evidence for an immediate effect of L-NAME treatment onNO generation at sites of papilla formation Furthermore althoughthe number of cells undergoing HR was too small for reliablestatistical analyses there was no indication that L-NAME affectedwhole-cell fluorescence responses Thus as in untreated leaves(Fig 5) treatment did not diminish the early whole-cell fluores-cence revealed by DAF-2DA staining
Light and fluorescence microscope analyses of effects of C-PTIO L-NAME and SNP on B graminis development and barley epidermal cell responses to attack
To investigate the role(s) of NO generated in the interaction ofB graminis with barley an NO scavenger (C-PTIO) a NOS inhib-itor (L-NAME) and an NO donor (SNP) were supplied separatelyto cut leaves Light microscopy showed that as compared withcontrols treating leaves with any of the test chemicals had noeffects on spore germination or subsequent germling develop-ment up to the differentiation of mature (hooked) appressoria(data not shown) Furthermore incident fluorescence microscopyindicated that none of the chemicals affected the timing frequency
or intensity of localized autofluorescent host cell responses asso-ciated with appressoria Detailed data for these responses are notgiven but in all cases approximately 10 of appressorial contactsites were associated with faint localized autofluorescent hostcell responses by 12 hai and by 24 hai approximately 60 ofsites showed autofluorescence which in most cases was judgedfaint or moderately intense (class 1 or 2 on the 0ndash3 subjectivescale) Other effects of each test chemical are considered in turn
Effects of C-PTIOA few very small haustoria (swollen apices of penetration pegs)indicating successful penetration were evident in epidermalcells of both barley lines by 12 hai and in both there was littleincrease in penetration frequency after 15ndash18 hai (Fig 8) Thisindicates that the outcome of attacks and therefore efficacy ofpenetration resistance was determined by this time Comparedwith controls overall analysis showed that C-PTIO treatmentsignificantly increased (P lt 0001) the frequency of successfulpenetration ie suppressed penetration resistance in bothbarley lines (Fig 8) Further analyses showed that this effectwas significant (P lt 005) at most individual sampling times Inboth Pallas and P01 successful penetration was associatedwith approximately 30 of appressoria in C-PTIO-treated leaveswhereas control values were around 15 In Pallas some cells diedfollowing successful penetration but this percentage remainedrelatively low throughout (only c 20ndash40 died Fig 8) andtreatment had no significant effect By contrast in P01 virtually allpenetrated haustorium-containing cells were dead by 18 hai(Fig 8) However at 12ndash15 hai some penetrated P01 cells showed
Fig 8 C-PTIO suppression of B graminis penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO scavenger C-PTIO (triangles) or water (circles) on percentages of B graminis appressoria with successful penetrations (haustoria) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (indicated by whole-cell autofluorescence) 12ndash30 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001
72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972
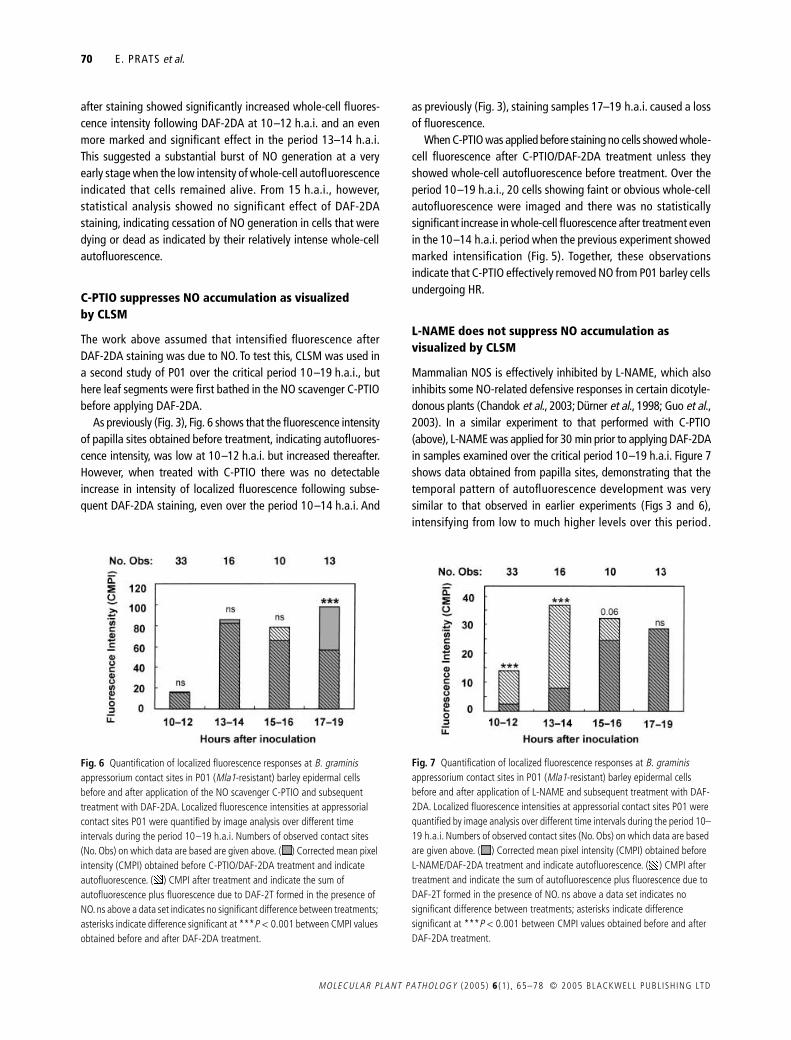
70 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
after staining showed significantly increased whole-cell fluores-cence intensity following DAF-2DA at 10ndash12 hai and an evenmore marked and significant effect in the period 13ndash14 haiThis suggested a substantial burst of NO generation at a veryearly stage when the low intensity of whole-cell autofluorescenceindicated that cells remained alive From 15 hai howeverstatistical analysis showed no significant effect of DAF-2DAstaining indicating cessation of NO generation in cells that weredying or dead as indicated by their relatively intense whole-cellautofluorescence
C-PTIO suppresses NO accumulation as visualized by CLSM
The work above assumed that intensified fluorescence afterDAF-2DA staining was due to NO To test this CLSM was used ina second study of P01 over the critical period 10ndash19 hai buthere leaf segments were first bathed in the NO scavenger C-PTIObefore applying DAF-2DA
As previously (Fig 3) Fig 6 shows that the fluorescence intensityof papilla sites obtained before treatment indicating autofluores-cence intensity was low at 10ndash12 hai but increased thereafterHowever when treated with C-PTIO there was no detectableincrease in intensity of localized fluorescence following subse-quent DAF-2DA staining even over the period 10ndash14 hai And
as previously (Fig 3) staining samples 17ndash19 hai caused a lossof fluorescence
When C-PTIO was applied before staining no cells showed whole-cell fluorescence after C-PTIODAF-2DA treatment unless theyshowed whole-cell autofluorescence before treatment Over theperiod 10ndash19 hai 20 cells showing faint or obvious whole-cellautofluorescence were imaged and there was no statisticallysignificant increase in whole-cell fluorescence after treatment evenin the 10ndash14 hai period when the previous experiment showedmarked intensification (Fig 5) Together these observationsindicate that C-PTIO effectively removed NO from P01 barley cellsundergoing HR
L-NAME does not suppress NO accumulation as visualized by CLSM
Mammalian NOS is effectively inhibited by L-NAME which alsoinhibits some NO-related defensive responses in certain dicotyle-donous plants (Chandok et al 2003 Duumlrner et al 1998 Guo et al2003) In a similar experiment to that performed with C-PTIO(above) L-NAME was applied for 30 min prior to applying DAF-2DAin samples examined over the critical period 10ndash19 hai Figure 7shows data obtained from papilla sites demonstrating that thetemporal pattern of autofluorescence development was verysimilar to that observed in earlier experiments (Figs 3 and 6)intensifying from low to much higher levels over this period
Fig 6 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of the NO scavenger C-PTIO and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before C-PTIODAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
Fig 7 Quantification of localized fluorescence responses at B graminis appressorium contact sites in P01 (Mla1-resistant) barley epidermal cells before and after application of L-NAME and subsequent treatment with DAF-2DA Localized fluorescence intensities at appressorial contact sites P01 were quantified by image analysis over different time intervals during the period 10ndash19 hai Numbers of observed contact sites (No Obs) on which data are based are given above ( ) Corrected mean pixel intensity (CMPI) obtained before L-NAMEDAF-2DA treatment and indicate autofluorescence ( ) CMPI after treatment and indicate the sum of autofluorescence plus fluorescence due to DAF-2T formed in the presence of NO ns above a data set indicates no significant difference between treatments asterisks indicate difference significant at P lt 0001 between CMPI values obtained before and after DAF-2DA treatment
NO and resistanceresponse in barley 71
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Patterns of NO generation were also very similar to those seenin untreated leaves (Fig 3) again indicating a transient burst ofgeneration Hence DAF-2DA staining led to significant increasesin fluorescence intensity indicative of NO generation in papillasites over the period 10ndash14 hai a near-significant increase wasfound in samples examined 15ndash16 hai but thereafter therewas no evidence for NO generation Therefore unlike C-PTIO therewas no evidence for an immediate effect of L-NAME treatment onNO generation at sites of papilla formation Furthermore althoughthe number of cells undergoing HR was too small for reliablestatistical analyses there was no indication that L-NAME affectedwhole-cell fluorescence responses Thus as in untreated leaves(Fig 5) treatment did not diminish the early whole-cell fluores-cence revealed by DAF-2DA staining
Light and fluorescence microscope analyses of effects of C-PTIO L-NAME and SNP on B graminis development and barley epidermal cell responses to attack
To investigate the role(s) of NO generated in the interaction ofB graminis with barley an NO scavenger (C-PTIO) a NOS inhib-itor (L-NAME) and an NO donor (SNP) were supplied separatelyto cut leaves Light microscopy showed that as compared withcontrols treating leaves with any of the test chemicals had noeffects on spore germination or subsequent germling develop-ment up to the differentiation of mature (hooked) appressoria(data not shown) Furthermore incident fluorescence microscopyindicated that none of the chemicals affected the timing frequency
or intensity of localized autofluorescent host cell responses asso-ciated with appressoria Detailed data for these responses are notgiven but in all cases approximately 10 of appressorial contactsites were associated with faint localized autofluorescent hostcell responses by 12 hai and by 24 hai approximately 60 ofsites showed autofluorescence which in most cases was judgedfaint or moderately intense (class 1 or 2 on the 0ndash3 subjectivescale) Other effects of each test chemical are considered in turn
Effects of C-PTIOA few very small haustoria (swollen apices of penetration pegs)indicating successful penetration were evident in epidermalcells of both barley lines by 12 hai and in both there was littleincrease in penetration frequency after 15ndash18 hai (Fig 8) Thisindicates that the outcome of attacks and therefore efficacy ofpenetration resistance was determined by this time Comparedwith controls overall analysis showed that C-PTIO treatmentsignificantly increased (P lt 0001) the frequency of successfulpenetration ie suppressed penetration resistance in bothbarley lines (Fig 8) Further analyses showed that this effectwas significant (P lt 005) at most individual sampling times Inboth Pallas and P01 successful penetration was associatedwith approximately 30 of appressoria in C-PTIO-treated leaveswhereas control values were around 15 In Pallas some cells diedfollowing successful penetration but this percentage remainedrelatively low throughout (only c 20ndash40 died Fig 8) andtreatment had no significant effect By contrast in P01 virtually allpenetrated haustorium-containing cells were dead by 18 hai(Fig 8) However at 12ndash15 hai some penetrated P01 cells showed
Fig 8 C-PTIO suppression of B graminis penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO scavenger C-PTIO (triangles) or water (circles) on percentages of B graminis appressoria with successful penetrations (haustoria) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (indicated by whole-cell autofluorescence) 12ndash30 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001
72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972
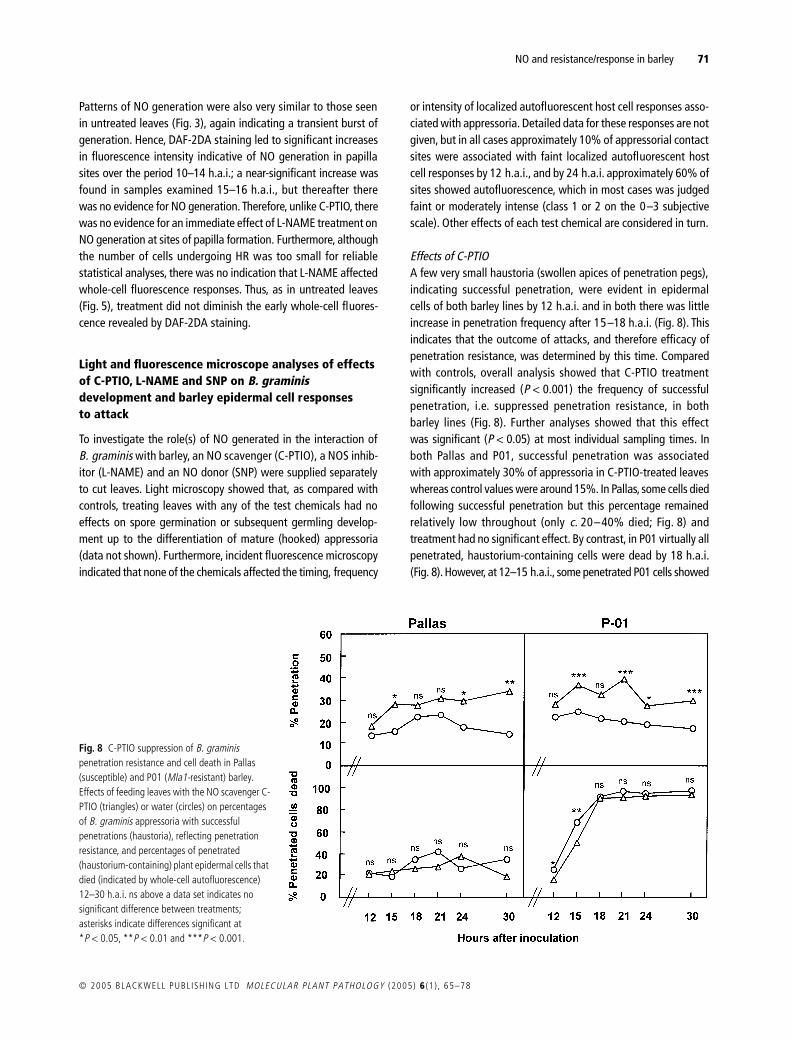
NO and resistanceresponse in barley 71
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Patterns of NO generation were also very similar to those seenin untreated leaves (Fig 3) again indicating a transient burst ofgeneration Hence DAF-2DA staining led to significant increasesin fluorescence intensity indicative of NO generation in papillasites over the period 10ndash14 hai a near-significant increase wasfound in samples examined 15ndash16 hai but thereafter therewas no evidence for NO generation Therefore unlike C-PTIO therewas no evidence for an immediate effect of L-NAME treatment onNO generation at sites of papilla formation Furthermore althoughthe number of cells undergoing HR was too small for reliablestatistical analyses there was no indication that L-NAME affectedwhole-cell fluorescence responses Thus as in untreated leaves(Fig 5) treatment did not diminish the early whole-cell fluores-cence revealed by DAF-2DA staining
Light and fluorescence microscope analyses of effects of C-PTIO L-NAME and SNP on B graminis development and barley epidermal cell responses to attack
To investigate the role(s) of NO generated in the interaction ofB graminis with barley an NO scavenger (C-PTIO) a NOS inhib-itor (L-NAME) and an NO donor (SNP) were supplied separatelyto cut leaves Light microscopy showed that as compared withcontrols treating leaves with any of the test chemicals had noeffects on spore germination or subsequent germling develop-ment up to the differentiation of mature (hooked) appressoria(data not shown) Furthermore incident fluorescence microscopyindicated that none of the chemicals affected the timing frequency
or intensity of localized autofluorescent host cell responses asso-ciated with appressoria Detailed data for these responses are notgiven but in all cases approximately 10 of appressorial contactsites were associated with faint localized autofluorescent hostcell responses by 12 hai and by 24 hai approximately 60 ofsites showed autofluorescence which in most cases was judgedfaint or moderately intense (class 1 or 2 on the 0ndash3 subjectivescale) Other effects of each test chemical are considered in turn
Effects of C-PTIOA few very small haustoria (swollen apices of penetration pegs)indicating successful penetration were evident in epidermalcells of both barley lines by 12 hai and in both there was littleincrease in penetration frequency after 15ndash18 hai (Fig 8) Thisindicates that the outcome of attacks and therefore efficacy ofpenetration resistance was determined by this time Comparedwith controls overall analysis showed that C-PTIO treatmentsignificantly increased (P lt 0001) the frequency of successfulpenetration ie suppressed penetration resistance in bothbarley lines (Fig 8) Further analyses showed that this effectwas significant (P lt 005) at most individual sampling times Inboth Pallas and P01 successful penetration was associatedwith approximately 30 of appressoria in C-PTIO-treated leaveswhereas control values were around 15 In Pallas some cells diedfollowing successful penetration but this percentage remainedrelatively low throughout (only c 20ndash40 died Fig 8) andtreatment had no significant effect By contrast in P01 virtually allpenetrated haustorium-containing cells were dead by 18 hai(Fig 8) However at 12ndash15 hai some penetrated P01 cells showed
Fig 8 C-PTIO suppression of B graminis penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO scavenger C-PTIO (triangles) or water (circles) on percentages of B graminis appressoria with successful penetrations (haustoria) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (indicated by whole-cell autofluorescence) 12ndash30 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001
72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

72 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
no whole-cell autofluorescence or cytoplasmic disorganizationindicating that they were alive when fixed Analysing data for the12 and 15 hai samples showed that overall the frequency ofcells killed by penetration at these early stages was significantlyhigher (P = 0015) in controls than in those with C-PTIO treatmentThe data suggest therefore that NO scavenging by C-PTIO delayedHR in P01 but did not prevent its eventual execution
Effects of L-NAMEIn P01 barley treatment with 10 mM L-NAME had no detectableeffects compared with treatment with the biologically inactive isomerD-NAME Irrespective of treatment some rudimentary haustoriawere evident by 12 hai and a maximum of approximately 16of appressoria produced successful penetration by 18 hai Treat-ment did not affect the timing of cell death approximately 30of penetrated cells showed some whole-cell autofluorescenceat 12 hai and this proportion increased to more than 95 by30 hai Because treatment had no effects in P01 barley Pallasleaves fixed only at 30 hai were examined and again therewere no effects of treatment
Explanations for a lack of L-NAME treatment effects include thepossibilities that the chemical either failed to reach barley epidermalcells or that it was present but at below a physiologically effectiveconcentration The former at least does not appear to be correctAlthough prepenetration fungus development was unaffectedL-NAME but not D-NAME greatly suppressed subsequent hyphaldevelopment in Pallas (Fig 9) Irrespective of treatment by 30 haimany haustoria were relatively well developed having formedapproximately six digitate processes On D-NAME-treated leavesyoung colonies had also formed several secondary hyphae witha total length of 100 microm or more (Fig 9A) By contrast on L-NAME-treated leaves hyphal growth was greatly suppressed andnormally only one stunted hypha had formed (Fig 9B) Thisdemonstrated that L-NAME was present within infected cellsat sufficient concentration to affect postinfection colony develop-ment severely
Effects of SNPCompared with controls treating leaves with 005 mM SNP did notaffect the frequency of successful penetration from appressoria ineither barley line (Fig 10) As in previous experiments in Pallaspenetration frequencies increased to about 20 by 15ndash18 haibut changed little thereafter In P01 the penetration frequency alsopeaked at this time but then apparently declined presumablybecause some rudimentary haustoria were obscured by cytoplasmicdisorganization following plant cell death (Koga et al 1990)The data again therefore indicate that the outcome of attacksand thus efficacy of penetration resistance was determined by15ndash18 hai Although SNP treatment did not affect penetrationresistance Fig 10 shows that it increased the rapidity of HR in P01Thus from 15 hai onwards the proportion of dead epidermal cellswas significantly (P lt 005) higher in SNP-treated leaves althoughby 24 hai almost all penetrated cells were dead irrespective oftreatment This suggests that NO liberated from SNP acceleratedHR in P01 Interestingly SNP treatment also increased the frequencyof cell death in Pallas so that by 18 hai and thereafter approx-imately 60 of haustorium-containing cells had died in treatedleaves of this compatible barley line
DISCUSSION
Prior to the current work only three studies have considered the roleof NO in cerealndashpathogen interactions but these have indicated itspotential importance in governing the outcomes In rice suspen-sion cultures treated with a Magnaporthe grisea-derived elicitorNO generation was associated with response-gene induction andHR (Hu et al 2003) In the oatndashcrown rust interaction simultaneousNO and H2O2 generation appeared to be associated with deathof cells adjacent to those attacked by an avirulent fungal isolate(Tada et al 2004) In the wheatndashstripe rust interaction analysis ofthe NO content of inoculated leaf extracts and pharmacologicalintervention indicated a general correlation of NO formation withexpression of race-specific resistance (Guo et al 2004)
Fig 9 Blumeria graminis hyphal development in L-NAME-treated and control barley leaves Light micrographs of B graminis colonies 30 hai on Pallas (susceptible) barley leaves treated either with (A) D-NAME as control or (B) L-NAME a mammalian NOS inhibitor In both cases conidia (C) have formed an appressorium (Ap) with a haustorium (H) with digitate processes but secondary hyphae (unlabelled arrows) are far more abundant on the D-NAME- than on the L-NAME-treated leaf where only a single stunted hypha is evident
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972
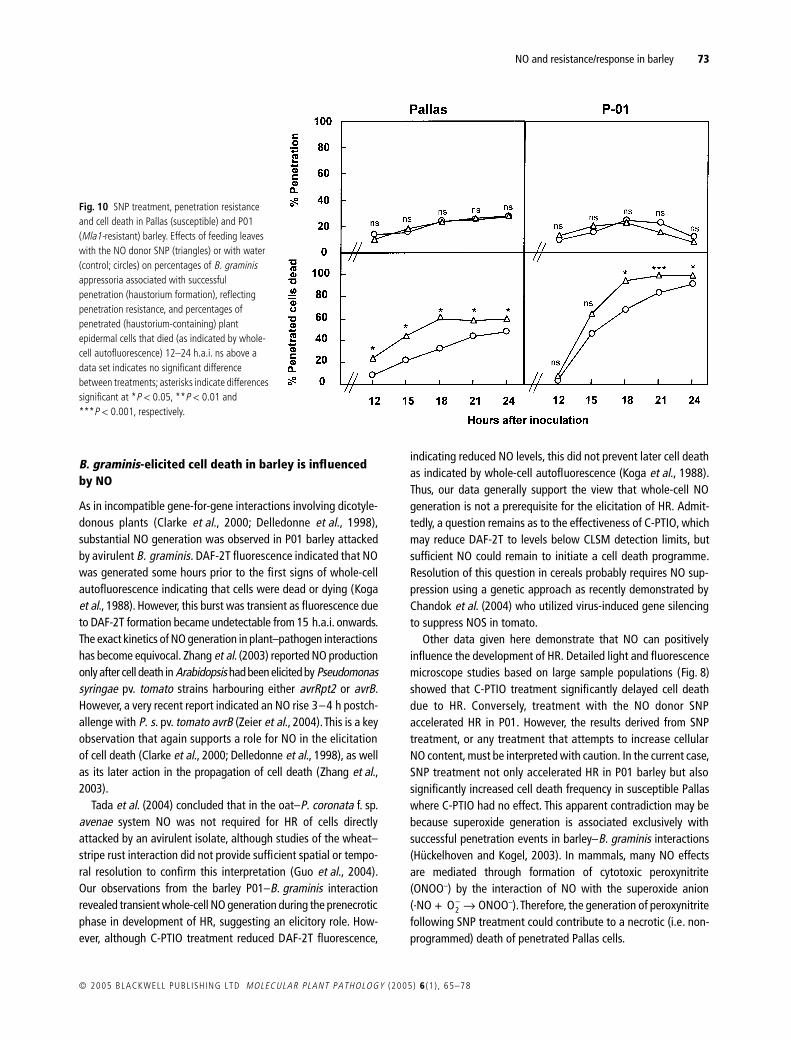
NO and resistanceresponse in barley 73
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
B graminis-elicited cell death in barley is influenced by NO
As in incompatible gene-for-gene interactions involving dicotyle-donous plants (Clarke et al 2000 Delledonne et al 1998)substantial NO generation was observed in P01 barley attackedby avirulent B graminis DAF-2T fluorescence indicated that NOwas generated some hours prior to the first signs of whole-cellautofluorescence indicating that cells were dead or dying (Kogaet al 1988) However this burst was transient as fluorescence dueto DAF-2T formation became undetectable from 15 hai onwardsThe exact kinetics of NO generation in plantndashpathogen interactionshas become equivocal Zhang et al (2003) reported NO productiononly after cell death in Arabidopsis had been elicited by Pseudomonassyringae pv tomato strains harbouring either avrRpt2 or avrBHowever a very recent report indicated an NO rise 3ndash4 h postch-allenge with P s pv tomato avrB (Zeier et al 2004) This is a keyobservation that again supports a role for NO in the elicitationof cell death (Clarke et al 2000 Delledonne et al 1998) as wellas its later action in the propagation of cell death (Zhang et al2003)
Tada et al (2004) concluded that in the oatndashP coronata f spavenae system NO was not required for HR of cells directlyattacked by an avirulent isolate although studies of the wheatndashstripe rust interaction did not provide sufficient spatial or tempo-ral resolution to confirm this interpretation (Guo et al 2004)Our observations from the barley P01ndashB graminis interactionrevealed transient whole-cell NO generation during the prenecroticphase in development of HR suggesting an elicitory role How-ever although C-PTIO treatment reduced DAF-2T fluorescence
indicating reduced NO levels this did not prevent later cell deathas indicated by whole-cell autofluorescence (Koga et al 1988)Thus our data generally support the view that whole-cell NOgeneration is not a prerequisite for the elicitation of HR Admit-tedly a question remains as to the effectiveness of C-PTIO whichmay reduce DAF-2T to levels below CLSM detection limits butsufficient NO could remain to initiate a cell death programmeResolution of this question in cereals probably requires NO sup-pression using a genetic approach as recently demonstrated byChandok et al (2004) who utilized virus-induced gene silencingto suppress NOS in tomato
Other data given here demonstrate that NO can positivelyinfluence the development of HR Detailed light and fluorescencemicroscope studies based on large sample populations (Fig 8)showed that C-PTIO treatment significantly delayed cell deathdue to HR Conversely treatment with the NO donor SNPaccelerated HR in P01 However the results derived from SNPtreatment or any treatment that attempts to increase cellularNO content must be interpreted with caution In the current caseSNP treatment not only accelerated HR in P01 barley but alsosignificantly increased cell death frequency in susceptible Pallaswhere C-PTIO had no effect This apparent contradiction may bebecause superoxide generation is associated exclusively withsuccessful penetration events in barleyndashB graminis interactions(Huumlckelhoven and Kogel 2003) In mammals many NO effectsare mediated through formation of cytotoxic peroxynitrite(ONOOndash) by the interaction of NO with the superoxide anion(middotNO + rarr ONOOndash) Therefore the generation of peroxynitritefollowing SNP treatment could contribute to a necrotic (ie non-programmed) death of penetrated Pallas cells
Fig 10 SNP treatment penetration resistance and cell death in Pallas (susceptible) and P01 (Mla1-resistant) barley Effects of feeding leaves with the NO donor SNP (triangles) or with water (control circles) on percentages of B graminis appressoria associated with successful penetration (haustorium formation) reflecting penetration resistance and percentages of penetrated (haustorium-containing) plant epidermal cells that died (as indicated by whole-cell autofluorescence) 12ndash24 hai ns above a data set indicates no significant difference between treatments asterisks indicate differences significant at P lt 005 P lt 001 and P lt 0001 respectively
O2minus
74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

74 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
The relatively minor modulation in the kinetics of the barley HRwhen NO levels were reduced by C-PTIO suggests that otherfactors influence the elicitation and elaboration of the HR It maybe that NO effects are exerted by its synergistic interaction withH2O2 In both soybean (Delledonne et al 2001) and tobacco (dePinto et al 2002) suspension cultures a balanced stoichiometricinteraction of NO with H2O2 appears important in influencingantioxidant levels (de Pinto et al 2002) and cell death (Delledonneet al 2001) In incompatible barleyndashB graminis interactionsH2O2 production at the epidermalndashmesophyll cell interface isdetected by DAB staining around 14ndash15 hai (Thordal-Christensen et al 1997 Vanacker et al 2000) ie shortly afterwe detected whole-cell NO generation However DAB stainingdepends on visualization of browning by transmitted light micro-scopy and this may be too insensitive to detect earlier lower levelH2O2 generation more closely coincident with onset of the whole-cell NO burst Thus in the incompatible barleyndashB graminis inter-action NO and H2O2 could be generated contemporaneously andtheir interaction could promote HR This would suggest thatfollowing C-PTIO treatment AOS might eventually compensatefor NO deficiency so that HR was only delayed Data from the oatndashcrown rust system (Tada et al 2004) support this view becauseC-PTIO treatment increased H2O2 production in HR lesions
Interestingly unlike in Arabidopsis (Chandok et al 2003 Guoet al 2003) and tobacco (Duumlrner et al 1998) an inhibitor (L-NAME) of mammalian NOS apparently failed to influence HR inP01 barley as determined by light microscopy CLSM also sug-gested that L-NAME failed to suppress whole-cell NO generationthat preceded the onset of whole-cell autofluorescence This find-ing could be explained by the short time cells were exposed to theinhibitor although data from tobacco (L A J Mur et al unpub-lished data) show that L-NAME can suppress NO productionwithin a similar time-frame In the light-microscope experimentthe possibility that L-NAME was not penetrating the plant tissuecan be discounted because of the observed inhibition of postin-fection fungal hyphal development Although our pharmaceuticalapproach suggests the enzyme may be present within the funguswe failed to find evidence for NOS activity in barley Howeverexpressed sequence tags (ESTs) exhibiting homology to Arabi-dopsis NOS have been identified in barley (accession numbersTC108982 BG309532 BG309532 Barley Gene Indices httpwwwtigrorg) so that further analysis is required to prove theexistence of NOS and its role in barleyndashB graminis interaction
Nitric oxide plays a role in papilla-based resistance to B graminis
Although there is good broadly based evidence for a role of NOin orchestrating HR no previous studies have investigated thepossible involvement of NO in papilla formation Here our CLSMimages clearly show that NO is generated precisely at sites of
papilla deposition commencing abruptly at around 10 hai ievery soon after B graminis appressoria mature and persistinguntil around 14ndash16 hai (Fig 3) when papilla effectiveness wasdetermined (Figs 8 and 10) Given the mobility of NO such highlylocalized DAF-2T fluorescence argues for a significant focusedgeneration probably coupled with efficient scavenging to producethe images shown in Fig 1 Furthermore application of C-PTIOhad the dual effects of suppressing localized NO accumulation(visualized by CLSM) and increasing the frequency of successfulfungal penetration (assessed by light microscopy of large popu-lations) The failure of SNP treatment to influence penetration(Fig 10) cannot be explained by its failure to reach and influencethe epidermis and probably indicates therefore that in untreatedcontrols NO levels were not limiting to papilla-based resistance
The first steps governing papilla formation are unknown butour data indicate that along with proton and calcium fluxes(Felle et al 2004) NO generation is among the earliest of cerealepidermal cell responses to appressorium contact Furthermorelocalized NO generation persisted until at least 14 hai when theefficacy of papillae was largely determined Therefore NO mayalso be involved in the complex second step that involves synthesismarshalling and assembly of papilla components We also notewith interest the more prolonged generation of NO (to 15ndash16 hai)in papillae of P01 compared with Pallas Thus it may be that Mla1could via NO affect the later stages of papilla elaboration
If NO acts as an early mediator of papilla formationthe increased fungal penetration following C-PTIO treatment(c 15) may appear relatively minor but it is in line with effectsobtained with certain other papilla-focused experimental manip-ulations For instance a similar level of increased fungal penetra-tion followed inhibition of enzymes involved in biosynthesis ofphenolic compounds in oat and barley mildew interactions(Carver et al 1994) and catalase application to suppress H2O2
production in the cowpeandashplantain powdery mildew interaction(Mellersh et al 2002) Thus NO appears to influence papillaeffectiveness in a quantitative manner Such a quantitative effectwould be explained if the rapidity or quantity of NO generationaffects the timing or expression of later events for examplecytoskeletal reorganization redirected cytoplasmic streamingcytoplasmic aggregate formation the synthesis of callose andphenolic compounds or the accumulation of inorganic papillacomponents such as silicon Such effects would favour theattempt of the fungus to penetrate the response site but it maynot prevent for example H2O2 generation and the subsequenterection of defences Alternatively removal of NO by C-PTIOapplication may lead to a compensatory accumulation of otherpapilla components Such a compensatory effect is seen forexample in oats where silicon deprivation leads to increasedPAL (phenylalanine ammonia lyase) activity and enhances theaccumulation of autofluorogenic papilla constituents that appar-ently compensate for the absence of silicon (Carver et al 1998)
NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

NO and resistanceresponse in barley 75
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Furthermore there is strong evidence that H2O2 is implicatedin effective papilla formation (Huumlckelhoven and Kogel 2003) andit may be that as in HR NO interacts synergistically with H2O2 tofacilitate the initiation of papillae andor their construction DABstaining has indicated H2O2 generation in papillae as early as12 hai (Vanacker et al 2000) or 15ndash16 hai (Huumlckelhovenet al 1999 2000 2001 Thordal-Christensen et al 1997) and itis possible that due to insensitivity of this stain H2O2 productionat earlier times is not detected Thus a role for H2O2 as an earlysignal for papilla initiation perhaps acting in concert with NOcannot be excluded at present
Using actin-polymerization inhibitors in Arabidopsis (Yun et al2003) and cowpea (Mellersh et al 2002) both H2O2 accumulationand callose deposition during assembly of the papilla was provento be influenced by cytoskeletal rearrangement It may be that aswith H2O2 the site of NO generation during papilla assembly isinfluenced by cytoskeletal organization In mammalian phagocytesNOS is associated with vesicles that colocalize with the corticalactin lying immediately adjacent to the peripheral cell membrane(Webb et al 2001) In such systems vesicle targeting and fusionfollowing the dimerization of vesicle and target membrane formsof SNARE proteins can be sufficient for endo- or exocytosis (Bockand Scheller 1999) In some instances vesicle formation is influ-enced by NO (Di Stasi et al 2002 Matsushita et al 2003) partic-ularly in vesicle-mediated neurotransmitter release in synapses(Sullivan 2003) It is now recognized in the barleyndashB graminisinteraction that some H2O2 at least is delivered to sites of papillaassembly within vesicles (Huumlckelhoven et al 1999) In Arabidop-sis and barley respectively the SNARE protein homologues PEN1and ROR1 are involved in targeting these vesicles to the plasmamembrane (Collins et al 2003) Thus analogies with mamma-lian systems could indicate that NO may affect the distribution ofH2O2 and the delivery of other papilla components possibly byinfluencing vesicle formation
Most studies into the role of NO in plant defence have been basedon well-characterized but relatively unsubtle model pathogenicinteractions Taken together our data suggest complex and variousroles of NO that are not only associated with the elicitation of celldeath but also with papilla initiation and construction and possiblywith in planta fungal development The challenge now is to developa molecular understanding of NO action controlling these processes
EXPERIMENTAL PROCEDURES
Plants pathogen inoculation and incubation
Pallas and its near-isogenic derivative P01 (Mla1) were grown incompost (Levington Horticulture Ipswich Suffolk UK) in a spore-proof glasshouse (natural light minimum temperature 12 degC)Fully expanded first leaves were excised at soil level and cut endswere trimmed under water to remove air embolisms Adaxial
surfaces were inoculated with 30 conidiamm2 of B graminis f sphordei isolate CC1 (grown on Pallas) by settling tower Cut endswere then trimmed again under water or the test solution inwhich they were immersed throughout incubation at 20 degC withlight when required at a photon flux density of 250 micromolm2s
Test chemicals
C-PTIO (2-(4-carboxyphenyl)-4455-tetramethylimidazoline-1-oxyl-3-oxide potassium salt)For light microscopy inoculated cut leaves incubated in darknesswere supplied continuously with 025 mM C-PTIO (Sigma-Aldrich)or water (controls) until four from each barley linetreatmentcombination were fixed every 3 h from 12 to 24 hai and againat 30 hai For CLSM 2-mm square segments cut from leavessupplied with water until 10ndash19 hai were bathed in C-PTIO for30 min before observation
L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride)For light microscopy inoculated cut leaves were supplied continu-ously with 10 mM L-NAME or D-NAME (Sigma-Aldrich) incubationwas under a 12-h12-h darkndashlight cycle and four leaves from eachbarley linetreatment combination were fixed for light microscopyevery 3 h from 12 to 24 hai and also at 30 hai For CLSM 2-mmsquare segments cut from leaves supplied with water until 10ndash19 haiwere bathed in 10 mM L-NAME for 30 min before observation
SNP (sodium nitroprusside)SNP (Fisher Scientific UK Ltd) releases NO when exposed to light butnot in darkness (Megson 2000) Unless stated otherwise treatedleaves and controls were incubated under a 12-h12-h darkndashlightcycle so that NO was released from 12 to 24 hai In preliminarytests leaves supplied with 10 or 05 mM SNP were necrotic by30 hai However 01 or 005 mM SNP caused no apparent leafdamage or effect on appressorium maturation Other testscompared various control treatments (i) de-ionized water (ii) 01 or005 mM SNP but incubating in continuous darkness and (iii) pro-vision of 01 or 005 mM SNP exhausted by continuous exposureto bright light for 5 days before application Data from lightmicroscopy showed no significant differences between thesetreatments For the experiment inoculated leaves were providedcontinuously with 005 mM SNP or water (controls) and fourfrom each barley linetreatment combination were fixed for lightmicroscopy every 3 h from 12 to 24 hai
Microscopy
Confocal laser scanning microscopy (CLSM)In the presence of NO DAF-2DA (Calbiochem USA formulation1 mg in 045 mL DMSO) is converted to fluorescent DAF-2T and thiswas visualized by CLSM (Bio-Rad MRC1024) using an argon laser
76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

76 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
(488-nm excitation515ndash530-nm band pass filter) laser microscopeand photomultiplier settings were standardized throughout Imageswere collected before and after DAF-2DA staining to enablediscrimination between autofluorescence (also excited by theargon laser) and fluorescence due to NO generation
Living segments (c 2 times 2 mm) taken from inoculated cut leavesprovided with water and incubated for between 6 and 24 haiwere mounted in MES buffer (MES 10 microMKCl 50 microM pH 615) ona glass slide with coverslip Laser images were captured from opticalsections stepped at 10-microM intervals from above germlings to belowplant cell response sites Specimens were then immersed in 10 microM
DAF-2DA in MES buffer (10 min) rinsed in pure MES buffer toremove excess dye (10 min) and laser images of the same interactionsites were captured As controls specimens were immersed in MESbuffer containing the equivalent quantity of DMSO present in the dye
To quantify fluorescence images were analysed (ImageJ software)to measure mean pixel intensity within localized and whole-cellresponses before and after DAF-2DA staining Subtracting the valuefor background fluorescence of an adjacent cell without fungalcontact gave corrected mean pixel intensity (CMPI) values CMPIvalues before staining indicated autofluorescence intensity whereasvalues after staining indicated additional fluorescence due toformation of DAF-2T Localized response sites were rejected ifobscured by the fungus if responses in adjacent cells interferedor if attacked cells showed whole-cell fluorescence Whole-cellresponses were analysed if there was evidence of whole-cellfluorescence before or after staining but only if there was nopossibility of interference from nearby responses
Statistical analysis of data for fluorescence intensity obtainedby analysis of CLSM images was complicated by the nature of theexperiment which involved observation of the living barley leafndashB graminis system For an individual sample it took up to 15 minto collect images of autofluorescence from a small number ofinteraction sites some of these were lost during subsequent handlingand others failed to meet the stringent criteria of acceptability fordetailed image analysis Demounting and staining took a further20 min and relocation and image collection of the original inter-action sites took another 15 min Thus it took several weeks tocollect data for the entire time-course study (6ndash24 hai) Overalltherefore experiments could not be organized in a conventionalreplicated and randomized design and could not be analysed assuch (eg by standard analysis of variance) Nevertheless a primeobjective of the experiment was to determine whether DAF-2DAstaining at a particular stage of the plantndashfungus interactionincreased the intensity of localized fluorescence in appressoriumcontact sites or whole-cell fluorescence in attacked epidermalcells thus indicating NO generation and its spatial distribution atthat stage For this purpose our data were of high quality con-sisting of paired observations where the datum for autofluores-cence intensity (before staining) at a particular site could becompared directly to the intensity datum after staining that indicated
NO generation in exactly the same site Thus for each individualcontact site the fluorescence intensity value before and afterstaining was obtained and the difference between these valueswas calculated for each site Given the relatively small number ofobservations from each time interval the variance was estimatedusing the KERNEL DENSITY procedure (Payne and Arnold 2002)in GenStat (Baird et al 2002) Within the separate time intervalsthe data for difference were subjected to a t-test assuming thedifference was equal to zero ie that staining had no influenceon intensity In this way the statistic indicated the probabilitythat staining led to a significant change in fluorescence intensityIt must be noted that although this allows comparisons of effectsof staining within a time interval (eg to investigate whetherstaining increased fluorescence at 10ndash12 hai) it does not allowcomparison between intervals (eg to investigate whetherfluorescence was greater at 20ndash24 hai than at 10ndash12 hai)
Light and incident fluorescence microscopyLeaves were fixed and cleared on pads and viewed unstainedand without a coverslip (Carver et al 1994) Transmitted lightmicroscopy determined fungal development and responses toappressoria in short epidermal cells (Types A and B Koga et al1990) One hundred conidia on each of the four replicate leavessubjected to every treatment were examined to determinewhether they had germinated and formed a mature (hooked)appressorium and a further 100 with mature appressoria wereexamined for successful penetration (haustorium formation)Autofluorescent responses of epidermal cells were observed byincident fluorescence microscopy (blue excitation filter maxi-mum transmittance 400 nm dichroic mirror and barrier filtertransmittance gt 530 nm) Localized autofluorescent host cellresponses were as described previously and their intensity wasrecorded on a 0ndash3 scale (Carver et al 1994) Death of attackedepidermal cells was recognized by whole-cell autofluorescence(Koga et al 1988 Zeyen et al 1995)
Percentage data calculated for leaf replicate values weretransformed to angles to normalize data and to stabilizevariances and were subjected to analysis of variance (Baird et al2002) Data sets containing high numbers of zero values were notanalysed Mean differences were indicated by calculating theleast significant difference (LSD) at P lt 005
ACKNOWLEDGEMENTS
EP was supported by a Marie Curie Individual Fellowship andTLWC by Defra project AR0712
REFERENCES
Alderton WK Cooper CE and Knowles RG (2001) Nitric oxidesynthases structure function and inhibition Biochem J 357 593ndash615
NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

NO and resistanceresponse in barley 77
copy 2005 BLACKWELL PUBL ISH ING LTD MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78
Baird DB Harding SA Lane PW Murray DA Payne RW andSoutar DM (2002) Genstat for Windows 6th edn Oxford VSN International
Bock JB and Scheller RH (1999) SNARE proteins mediate lipid bilayerfusion Proc Natl Acad Sci USA 96 12227ndash12229
Bushnell WR (1981) Incompatibility conditioned by the Mla gene in powderymildew of barley the halt in cytoplasmic streaming Phytopathology71 1062ndash1066
Carver TLW Robbins MP Thomas BJ Troth K Raistrick N andZeyen RJ (1998) Silicon deprivation enhances localized autofluores-cent responses and phenylalanine ammonia-lyase activity in oat attackedby Blumeria graminis Physiol Mol Plant Pathol 52 245ndash257
Carver TLW Zeyen RJ Bushnell WR and Robbins MP (1994)Inhibition of phenylalanine ammonia lyase and cinnamyl alcoholdehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC) Physiol Mol Plant Pathol 44 261ndash272
Chamnongpol SH Willekens H Moeder W Langbartels CSandermann H Van Montagu M Inze D and Van Camp W(1997) Defense activation and enhanced pathogen tolerance induced byH2O2 in transgenic tobacco Proc Natl Acad Sci USA 95 5818ndash5823
Chandok MR Ekengren SK Martin GB and Klessig DF (2004)Suppression of pathogen-inducible NO synthase (iNOS) activity in tomatoincreases susceptibility to Pseudomonas syringae Proc Natl Acad SciUSA 25 8239ndash8244
Chandok MR Ytterberg AJ van Wijk KJ and Klessig DF (2003)The pathogen inducible nitric oxide synthase (iNOS) in plants is a variantof the P protein of the glycine decarboxylase complex Cell 113 469ndash482
Chelkowski J Tyrka M and Sobkiewicz A (2003) Resistance genesin barley (Hordeum vulgare L) and their identification with molecularmarkers J Appl Genet 44 291ndash309
Clarke A Desikan R Hurst RD Hancock JT and Neill SJ (2000)NO way back nitric oxide and programmed cell death in Arabidopsisthaliana suspension cultures Plant J 24 667ndash677
Collins NC Thordal-Christensen H Lipka V Bau S Kombrink EQiu JL Huumlckelhoven R Stein M Freialdenhoven A SomervilleSC and Schulze-Lefert P (2003) SNARE-protein-mediated diseaseresistance at the plant cell wall Nature 425 973ndash977
Delledonne M Xia Y Dixon RA and Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance Nature 394 585ndash588
Delledonne M Zeier J Marocco A and Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygen intermediates inthe plant hypersensitive disease resistance response Proc Natl AcadSci USA 98 3454ndash13459
Di Stasi AM Mallozzi C Macchia G Maura G Petrucci TC andMinetti M (2002) Peroxynitrite affects exocytosis and SNARE complexformation and induces tyrosine nitration of synaptic proteins J Neuro-chem 82 420ndash429
Dorey S Kopp M Geoffroy P Fritig B and Kauffmann S (1999)Hydrogen peroxide from the oxidative burst is neither necessary norsufficient for hypersensitive cell death induction phenylalanine ammonialyase stimulation salicylic acid accumulation or scopoletin consumptionin cultured tobacco cells treated with elicitin Plant Physiol 121 163ndash171
Duumlrner J Wendehenne D and Klessig DF (1998) Defense geneinduction in tobacco by nitric oxide cyclic GMP and cyclic ADP-riboseProc Natl Acad Sci USA 95 10328ndash10333
Felle HH Herrmann A Hanstein S Huumlckelhoven R and Kogel KH(2004) Apoplastic pH signalling in barley leaves attacked by the powderymildew fungus Blumeria graminis f sp hordei Mol PlantndashMicrobe Interact17 118ndash123
Foissner ID Wendehenne D Langebartels C and Ducircrner J (2000)In vivo imaging of an elicitor-induced nitric oxide burst in tobacco PlantJ 23 817ndash824
Garces H Durzan D and Pedroso MC (2001) Mechanical stresselicits nitric oxide formation and DNA fragmentation in Arabidopsis thal-iana Ann Bot 87 567ndash574
Glazener JA Orlandi EW and Baker CJ (1996) The active oxygenresponse of cell suspensions to incompatible bacteria is not sufficient tocause hypersensitive cell death Plant Physiol 110 759ndash763
Green JR Carver TLW and Gurr SJ (2002) The formation andfunction of infection and feeding structures In The Powdery Mildews aComprehensive Treatise (Belanger RR Bushnell WR Dik AJ andCarver TLW eds) St Paul MN APS Press pp 66ndash82
Guo P Cao Y Li Z and Zhao B (2004) Role of an endogenous nitricoxide burst in the resistance of wheat to stripe rust Plant Cell Environ27 473ndash477
Guo FQ Okamoto M and Crawford NM (2003) Identification of aplant nitric oxide synthase gene involved in hormonal signalling Science302 100ndash103
Hu XY Neill SJ Cai WM and Tang ZC (2003) NO-mediated hyper-sensitive responses of rice suspension cultures induced by incompatibleelicitorChin Sci Bull 48 358ndash363
Huumlckelhoven R Dechert C Trujillo M and Kogel KH (2001)Differential expression of putative cell death regulator genes in near-isogenic resistant and susceptible barley lines during interaction withthe powdery mildew fungus Plant Mol Biol 47 739ndash748
Huumlckelhoven R Fodor J Preis C and Kogel KH (1999) Hypersen-sitive cell death and papilla formation in barley attacked by the powderymildew fungus are associated with hydrogen peroxide but not withsalicylic acid accumulation Plant Physiol 119 1251ndash1260
Huumlckelhoven R Fodor J Trujillo M and Kogel KH (2000) BarleyMla and Rar mutants compromised in the hypersensitive cell deathresponse against Blumeria graminis fsp hordei are modified in theirability to accumulate reactive oxygen intermediates at sites of fungalinvasion Planta 212 16ndash24
Huumlckelhoven R and Kogel KH (2003) Reactive oxygen intermediates inplantndashmicrobe interactions who is who in powdery mildew resistancePlanta 216 891ndash902
Jabs T Tschoumlpe M Colling C Hahlbrock K and Scheel D (1997)Elicitor-stimulated ion fluxes and from the oxidative burst are essen-tial components in triggering defense gene activation and phytoalexinsynthesis in parsley Proc Natl Acad Sci USA 94 4800ndash4805
Koga H Bushnell WR and Zeyen RJ (1990) Specificity of cell typeand timing of events associated with papilla formation and the hyper-sensitive reaction in leaves of Hordeum vulgare attacked by Erysiphegraminis f sp hordei Can J Bot 68 2344ndash2352
Koga H Zeyen RJ Bushnell WR and Ahlstrand GG (1988) Hyper-sensitive cell death autofluorescence and insoluble silicon accumulationin barley leaf epidermal cells under attack by Erysiphe graminis f sphordei Physiol Mol Plant Pathol 32 395409
Kojima H Nakatsubo N Kikuchi K Kawahara S Kirino YNagoshi H Hirata Y and Nagano T (1998a) Detection and imagingof nitric oxide with novel fluorescent indicators diaminofluoresceinsAnal Chem 70 2446ndash2453
Kojima H Sakurai K Kikuchi K Kawahara S Kirino Y Nagoshi HHirata Y and Nagano T (1998b) Development of a fluorescentindicator for nitric oxide based on the fluorescent chromophore ChemPhar Bull 46 373ndash375
O2minus
78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972

78 E PRATS et al
MOLECULAR PLANT PATHOLOGY (2005) 6 (1 ) 65ndash78 copy 2005 BLACKWELL PUBL ISH ING LTD
Koslashlster P Munk L Stolen O and Lohde J (1986) Near-isogenic barleylines with genes for resistance to powdery mildew Crop Sci 26 903ndash907
Levine AR Tenhaken R Dixon R and Lamb C (1994) H2O2 from theoxidative burst orchestrates the plant hypersensitive disease resistanceresponse Cell 79 583ndash593
Lyngkjaeligr MF Carver TLW and Zeyen RJ (2001) Virulent Blumeriagraminis infection induces penetration susceptibility and suppressesrace-specific hypersensitive resistance against avirulent attack in Mla1-barley Physiol Mol Plant Pathol 59 243ndash256
Matsushita K Morrell CN Cambien B Yang SX Yamakuchi MBao C Hara MR Quick RA Cao W OrsquoRourke B Lowenstein JMPevsner J Wagner DD and Lowenstein CJ (2003) Nitric oxideregulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitivefactor Cell 115 139ndash150
Megson IL (2000) Nitric oxide donor drugs Drugs Future 25 701ndash715Mellersh DG Foulds IV Higgins VJ and Heath MC (2002) H2O2
plays different roles in determining penetration failure in three diverseplantndashfungal interactions Plant J 29 257ndash268
Neill SJ Desikan R and Hankock JT (2003) Nitric oxide signalling inplants New Phytologist 159 11ndash35
Panstruga R and Schulze-Lefert P (2002) Live and let live insightsinto powdery mildew disease and resistance Mol Plant Pathol 3 495ndash502
Payne RW and Arnold GM (2002) Genstat Release 61 ReferenceManual Part 3 Procedure Library PL14 Oxford VSN International
Pedroso MC Magalhaes JR and Durzan D (2000) A nitric oxideburst precedes apoptosis in angiosperm and gymnosperm callus cellsand foliar tissues J Exp Bot 51 1027ndash1036
de Pinto MC Tommasi F and De Gara L (2002) Changes in theantioxidant systems as part of the signalling pathway responsible for theprogrammed cell death activated by nitric oxide and reactive oxygenspecies in tobacco Bright-Yellow 2 cells Plant Physiol 130 698ndash708
Sullivan J (2003) NO going back Nat Neurosci 6 905ndash906Tada Y Mori T Shinogi T Yao N Takahashi S Betsuyaku S
Sakamoto M Park P Nakayashiki H Tosa Y and Mayama S(2004) Nitric oxide and reactive oxygen species do not elicit hypersensi-tive cell death but induce apoptosis in the adjacent cells during thedefense response of oats Mol PlantndashMicrobe Interact 17 245ndash253
Thordal-Christensen H Zhang Z Wei YD and Collinge DB (1997)Subcellular localization of H2O2 in plants H2O2 accumulation in papillaeand hypersensitive response during the barleyndashpowdery mildew interac-tion Plant J 11 1187ndash1194
Vanacker H Carver TLW and Foyer CH (2000) Early H2O2 accumu-lation in mesophyll cells leads to induction of glutathione during thehypersensitive response in the barleyndashpowdery mildew interaction PlantPhysiol 123 1289ndash1300
Webb JL Harvey MW Holden DW and Evans TJ (2001) Macro-phage nitric oxide synthase associates with cortical actin but is notrecruited to phagosomes Infect Immun 69 6391ndash6400
Young RJ Beams RM Carter K Clark HA Coe DMChambers CL Davies PI Dawson J Drysdale MJ Franzman KWFrench C Hodgson ST Hodson HF Kleanthous S Rider PSanders D Sawyer DA Scott KJ Shearer BG Stocker RSmith S Tackley MC and Knowles RG (2000) Inhibition ofinducible nitric oxide synthase by acetamidine derivatives of hetero-substituted lysine and homolysine Bioorg Med Chem Lett 10 597ndash600
Yun BW Atkinson HA Gaborit C Greenland A Read NDPallas JA and Loake GJ (2003) Loss of actin cytoskeletal functionand EDS1 activity in combination severely compromises non-host resistancein Arabidopsis against wheat powdery mildew Plant J 34 768ndash777
Zeier J Delledonne M Mishna T Severi E Sonoda M andLamb C (2004) Genetic elucidation of nitric oxide signalling in incom-patible plantndashpathogen interactions Plant Physiol 136 2875ndash2886
Zeyen RJ Bushnell WR Carver TLW Robbins MP Clark TABoyles DA and Vance CP (1995) Inhibiting phenylalanine ammonia-lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR) butnot mlo5 (non-HR) barley powdery mildew resistances Physiol MolPlant Pathol 47 119ndash140
Zeyen RJ Carver TLW and Lyngkjaeligr MF (2002) Epidermal cellpapillae In Powdery Mildews a Comprehensive Treatise (Belanger RRBushnell WR Dik AJ and Carver TLW eds) St Paul MN APS Presspp 107ndash125
Zhang C Czymmek KJ and Shapiro AD (2003) Nitric oxide does nottrigger early programmed cell death events but may contribute to cell-to-cell signalling governing progression of the Arabidopsis hypersensitiveresponse Mol PlantndashMicrobe Interact 16 962ndash972
Related Documents











