Research report Nicergoline, a drug used for age-dependent cognitive impairment, protects cultured neurons against h-amyloid toxicity Filippo Caraci a,1 , Mariangela Chisari b,1 , Giuseppina Frasca b , Pier Luigi Canonico c , Angelo Battaglia d , Marco Calafiore a , Giuseppe Battaglia e , Paolo Bosco f , Ferdinando Nicoletti e,g , Agata Copani a,h , Maria Angela Sortino b, * a Department of Pharmaceutical Sciences, University of Catania, Viale A. Doria, 6, 95125, Catania, Italy b Department of Experimental and Clinical Pharmacology, University of Catania, Viale A. Doria, 6, 95125, Catania, Italy c DISCAFF, University of Piemonte Orientale, Italy d CNS Medical Department, Pfizer Italia, Rome, Italy e I.N.M. Neuromed, Pozzilli, Italy f IRCCS Oasi Maria SS, Troina, Italy g Department of Human Physiology and Pharmacology, University of Rome ‘‘La Sapienza’’, Italy h IBB, CNR, Catania, Italy Accepted 1 April 2005 Available online 10 May 2005 Abstract Nicergoline, a drug used for the treatment of Alzheimer’s disease and other types of dementia, was tested for its ability to protect neurons against h-amyloid toxicity. Pure cultures of rat cortical neurons were challenged with a toxic fragment of h-amyloid peptide (hAP 25 – 35 ) and toxicity was assessed after 24 h. Micromolar concentrations of nicergoline or its metabolite, MDL, attenuated hAP 25 – 35 -induced neuronal death, whereas MMDL (another metabolite of nicergoline), the a 1 -adrenergic receptor antagonist, prazosin, or the serotonin 5HT-2 receptor antagonist, methysergide, were inactive. Nicergoline increased the basal levels of Bcl-2 and reduced the increase in Bax levels induced by h-amyloid, indicating that the drug inhibits the execution of an apoptotic program in cortical neurons. In mixed cultures of rat cortical cells containing both neurons and astrocytes, nicergoline and MDL were more efficacious than in pure neuronal cultures in reducing h-amyloid neurotoxicity. Experiments carried out in pure cultures of astrocytes showed that a component of neuroprotection was mediated by a mechanism of glial – neuronal interaction. The conditioned medium of cultured astrocytes treated with nicergoline or MDL for 72 – 96 h (collected 24 h after drug withdrawal) was neuroprotective when transferred to pure neuronal cultures challenged with h-amyloid. In cultured astrocytes, nicergoline increased the intracellular levels of transforming-growth factor-h and glial-derived neurotrophic factor, two trophic factors that are known to protect neurons against h-amyloid toxicity. These results raise the possibility that nicergoline reduces neurodegeneration in the Alzheimer’s brain. D 2005 Elsevier B.V. All rights reserved. Theme: Disorders of the nervous system Topic: Degenerative disease: Alzheimer’s—neuropharmacology and neurotransmitters Keywords: h-Amyloid toxicity; Cortical neurons; Nicergoline; Glial-neuronal interaction 1. Introduction Nicergoline (10a-methoxy-1,6-dimethylergoline-8h- methanol-5-bromonicotinate) is an ergot alkaloid derivative that is clinically used for the treatment of cognitive impair- ment associated with various types of dementia, including 0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2005.04.004 * Corresponding author. Fax: +39 095 7384228. E-mail address: [email protected] (M.A. Sortino). 1 These authors contributed equally to this work. Brain Research 1047 (2005) 30 – 37 www.elsevier.com/locate/brainres

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/brainres
Brain Research 1047
Research report
Nicergoline, a drug used for age-dependent cognitive impairment,
protects cultured neurons against h-amyloid toxicity
Filippo Caracia,1, Mariangela Chisarib,1, Giuseppina Frascab, Pier Luigi Canonicoc,
Angelo Battagliad, Marco Calafiorea, Giuseppe Battagliae, Paolo Boscof,
Ferdinando Nicolettie,g, Agata Copania,h, Maria Angela Sortinob,*
aDepartment of Pharmaceutical Sciences, University of Catania, Viale A. Doria, 6, 95125, Catania, ItalybDepartment of Experimental and Clinical Pharmacology, University of Catania, Viale A. Doria, 6, 95125, Catania, Italy
cDISCAFF, University of Piemonte Orientale, ItalydCNS Medical Department, Pfizer Italia, Rome, Italy
eI.N.M. Neuromed, Pozzilli, ItalyfIRCCS Oasi Maria SS, Troina, Italy
gDepartment of Human Physiology and Pharmacology, University of Rome ‘‘La Sapienza’’, ItalyhIBB, CNR, Catania, Italy
Accepted 1 April 2005
Available online 10 May 2005
Abstract
Nicergoline, a drug used for the treatment of Alzheimer’s disease and other types of dementia, was tested for its ability to protect neurons
against h-amyloid toxicity. Pure cultures of rat cortical neurons were challenged with a toxic fragment of h-amyloid peptide (hAP25–35) andtoxicity was assessed after 24 h. Micromolar concentrations of nicergoline or its metabolite, MDL, attenuated hAP25–35-induced neuronal
death, whereas MMDL (another metabolite of nicergoline), the a1-adrenergic receptor antagonist, prazosin, or the serotonin 5HT-2 receptor
antagonist, methysergide, were inactive. Nicergoline increased the basal levels of Bcl-2 and reduced the increase in Bax levels induced by
h-amyloid, indicating that the drug inhibits the execution of an apoptotic program in cortical neurons. In mixed cultures of rat cortical cells
containing both neurons and astrocytes, nicergoline and MDL were more efficacious than in pure neuronal cultures in reducing h-amyloid
neurotoxicity. Experiments carried out in pure cultures of astrocytes showed that a component of neuroprotection was mediated by a
mechanism of glial–neuronal interaction. The conditioned medium of cultured astrocytes treated with nicergoline or MDL for 72–96 h
(collected 24 h after drug withdrawal) was neuroprotective when transferred to pure neuronal cultures challenged with h-amyloid. In cultured
astrocytes, nicergoline increased the intracellular levels of transforming-growth factor-h and glial-derived neurotrophic factor, two trophic
factors that are known to protect neurons against h-amyloid toxicity. These results raise the possibility that nicergoline reduces
neurodegeneration in the Alzheimer’s brain.
D 2005 Elsevier B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Degenerative disease: Alzheimer’s—neuropharmacology and neurotransmitters
Keywords: h-Amyloid toxicity; Cortical neurons; Nicergoline; Glial-neuronal interaction
0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2005.04.004
* Corresponding author. Fax: +39 095 7384228.
E-mail address: [email protected] (M.A. Sortino).1 These authors contributed equally to this work.
1. Introduction
Nicergoline (10a-methoxy-1,6-dimethylergoline-8h-methanol-5-bromonicotinate) is an ergot alkaloid derivative
that is clinically used for the treatment of cognitive impair-
ment associated with various types of dementia, including
(2005) 30 – 37

F. Caraci et al. / Brain Research 1047 (2005) 30–37 31
primary degenerative, vascular, and mixed dementia
[13,22,30,40]. Similarly to other ergoline derivatives, nicer-
goline improves brain hemodynamics and metabolism
[24,28,37], increases energy production in injured mitochon-
dria [31], and blocks a1-adrenergic receptors [21]. In
addition, nicergoline enhances cholinergic transmission [7]
and polyphosphoinositide turnover [8], two effects that
support the use of the drug in Alzheimer’s disease (AD).
The evidence that nicergoline stimulates protein kinase C-
mediated a-secretase processing of the amyloid precursor
protein (APP) [10] suggests that the drug limits the formation
of pathological cleavage products of APP. It becomes
particularly interesting to establish whether nicergoline is
also able to protect neurons against the degeneration cascade
occurring in the AD brain. In vivo studies have shown that a
nicergoline treatment supports the survival of forebrain
cholinergic neurons, preventing the age- and colchicine-
induced reduction in choline acetyl transferase mRNA and
increasing the content of nerve growth factor and brain-
derived neurotrophic factor in the brain of aged rats [17].
Although these effects are relevant to the AD pathology, no
evidence is yet provided that nicergoline affects neurotoxicity
induced by h-amyloid peptide (hAP), the main constituent of
the amyloid plaques in the AD brain. hAP is recognized
among the major factors involved in neurodegeneration
associated with AD and is largely used to induce neuronal
death in vitro [20]. We now report that nicergoline protects
cultured cortical neurons against hAP toxicity and that part ofthis effect is mediated by a mechanism of glial–neuronal
interaction.
2. Materials and methods
All animal experimental procedures were carried out in
accordance with the directives of the Italian and EU
regulations for care and use of experimental animals
(DL116/92) and approved by the Italian Ministry of Health.
2.1. Drugs
Nicergoline, 1-methyl-10-alpha-methoxy-9,10-dihydro-
lysergol (MMDL), and 10-alpha-methoxy-9,10-dihydroly-
sergol (MDL) were a kind gift of Pharmacia Italia, Nerviano,
Italy. The hAP fragment, hAP25–35, was purchased from
Bachem Feinchemikalien AG (Bubendorf, Switzerland).
2.2. Cultures of cortical neurons
Cultures of pure cortical neurons were obtained from rats
at embryonic day 15 (Morini, s.a.s., Reggio Emilia, Italy).
Briefly, cortices were dissected in Ca++/Mg++-free buffer and
mechanically dissociated. Cortical cells were plated at a
density of 2 � 106/dish on 35 mm dishes (Nunc, Rochester,
NY) precoated with 0.1 mg/ml poly-d-lysine (Sigma-Aldrich
S.r.l., Milan, Italy) in DMEM/Ham’s F12 (1:1) medium
supplemented with the following components: 10 mg/ml bo-
vine serum albumin, 10 Ag/ml insulin, 100 Ag/ml transferrin,
100 AM putrescine, 20 nM progesterone, 30 nM selenium,
2 mM glutamine, 6 mg/ml glucose, 100 U/ml penicillin, and
100 Ag/ml streptomycin. Cytosine-d-arabinofuranoside
(10 AM) was added to the cultures 18 h after plating to avoid
the proliferation of non-neuronal elements and was kept for
3 days before medium replacement. This method yields
>99% pure neuronal cultures, as judged by immunocytoche-
mistry for glial fibrillary acidic protein and flow cytometry
for neuron-specific microtubule-associated protein 2 [11].
For mixed cortical cultures, cells were grown into
DMEM/F12 (1:1) supplemented with 10% horse serum,
10% fetal calf serum (FCS), 2 mM glutamine, 6 mg/ml
glucose. After 7–10 days in vitro, glial cell division was
halted by exposure to 10 AM cytosine-d-arabinoside for 3
days and cells were shifted into a maintenance serum-free
medium. Mature cultures contained about 35–40% neurons.
2.3. Pure cultures of cortical astrocytes
Cortical glial cells were prepared from 1- to 3-day-old
Sprague–Dowley rats (Morini, s.a.s.). After removal of
meninges and isolation of cortices, cells were dispersed by
mechanical and enzymatic dissociation using a 0.25%
solution of trypsin (Invitrogen, S.r.l., Milan, Italy). Cells
were plated onto 75 mm2 flasks and maintained in DMEM
culture medium, supplemented with 10% FCS, penicillin/
streptomycin (100 U/ml–100 Ag/ml), and glutamine
(2 mM). All medium constituents were from Invitrogen,
and all plastic materials were from Corning Life Sciences
(Acton, MA). Confluent cultures at 8–10 DIV were
shaked overnight at 37 -C to remove microglia and
oligodendrocytes. Astrocytes were collected by trypsin
digestion, seeded onto 35 or 100 mm dishes and used for
experiments 6–8 days after re-plating.
2.4. Handling of bAP
Different lots of hAP25–35 were tested and the same
batch was used throughout the entire study to rely on a
consistent profile of toxicity. Peptides were solubilized in
sterile, double distilled water at an initial concentration of
2.5 mM, and stored frozen at �20 -C. hAP(25–35) was usedat a final concentration of 25 AM in the presence of the
glutamate receptor antagonists MK-801 (10 AM) and
DNQX (30 AM) to avoid the potentiation of endogenous
glutamate toxicity.
2.5. MTT assay
Cells were incubated with MTT (0.9 mg/ml final
concentration, Sigma) for 2 h at 37 -C. A solubilization
solution containing 20% SDS was then added for an
additional 1 h, and formazan production was evaluated in
a plate reader (absorbance = 560 nm).

F. Caraci et al. / Brain Research 1047 (2005) 30–3732
2.6. Evaluation of apoptotic neuronal death
Apoptotic death was assessed by FACS analysis of
prediploid DNA. Mature cortical neurons at 7–8 DIV were
harvested with 0.25% trypsin, fixed in 70% ethanol, and
stored at �20 -C until use. After repeated washing, cells
were incubated for 1 h with 100 Ag/ml RNase and stained
with 50 Ag/ml propidium iodide. DNA content and ploidy
was assessed by a Coulter Elite Flow Cytometer. The
prediploid peak was evaluated as an index of apoptosis.
2.7. Western blot analysis
Cells were harvested in lysis buffer containing a cocktail
of protease inhibitors including bestatin, aprotinin, leupep-
tin, and sodium EDTA. After sonication, an aliquot of the
sample was processed for protein concentrations according
to the method of Bradford. Samples were concentrated and
boiled for 5 min. Proteins were separated electrophoretically
on polyacrylamide gel (30 mA/h) using 60–80 Ag of cell
proteins per lane. Proteins were transferred to nitrocellulose
membranes (Hybond ECL, Amersham Biosciences Europe
GmbH, Milan, Italy) at room temperature using a transblot
semidry transfer cell. After blocking, the membranes were
incubated with rabbit anti-Bax or anti-Bcl-2 (1:200, Santa
Cruz Bioctechnology, Santa Cruz, CA) overnight at 4 -C.Membranes were then thoroughly washed and incubated
with HRP-conjugated secondary antibodies. Specific bands
were visualized using the SuperSignal chemiluminescent
detection system (Pierce Biotechnology, Rockford, IL).
2.8. RT-PCR
Astrocytes were treated with 10 AM nicergoline for 18 h.
Total RNA was extracted with the Tryzol reagent (Sigma).
Two micrograms of total RNA was used for cDNA
synthesis, using oligo(dT) primers and 10 mM deoxynu-
cleotide triphosphate mix (Invitrogen) and M-MLV Reverse
Transcriptase (RT, 200 U/reaction; Promega, Madison, WI)
at 37 -C for 50 min. Two microliters of cDNA was used in
each subsequent PCR amplification, in an automatic
thermocycler, using 2.5 U/reaction of Platinum Taq DNA
polymerase and corresponding primers. Reaction conditions
were 95 -C for 5 min followed by 33 cycles of 95 -C, 56 -C,and 72 -C each for 1 min. The final extension step was 7
min at 72 -C. PCR product were analyzed electrophoreti-
cally on a 1.8% agarose gel run in Tris–acetate/EDTA
buffer.
Primers used were as follows: TGF-h1 (f 5V-TGGACCG-CAACAACGCCATCTATGAGAAAACC-3V; r 5V-TGGAG-CTGAAGCAATAGTTGGTATCCAGGGCT-3V); IL-6(f 5V-CAAGAGACTTCCAGCCAGTTGC-3V; r 5V-TTGCCGAG-TAGACCTCATAGTGACC-3V); GDNF (f 5V-ATGAAGTT-ATGGGATGTCGTGGCTG-3V; r 5V-CCTTTTACGCGGA-ATGCTTTCTTAG-3V); GAPDH (f 5V-GCCGCCTGGTCAC-CAGGGCTG-3V; r 5V-ATGGACTGTGGTCATGAGCCC-3V).
3. Results
Initially, we examined the neuroprotective activity of
nicergoline against hAP toxicity using pure cultures of rat
cortical neurons, which were virtually devoid of astrocytes
or other contaminating cells [11]. Cultures were treated
with hAP25–35 in the presence of ionotropic glutamate
receptor antagonists (1 AM MK-801 + 10 AM DNQX) to
limit endogenous excitotoxicity. Under these conditions,
neurons exposed to hAP die showing an apoptotic
phenotype [26]. Neurotoxicity was assessed by combining
the MTT assay and FACS analysis of pre-diploid DNA.
Exposure to hAP25–35 (25 AM) was toxic in a time-
dependent manner, inducing apoptotic death in about 40–
50% of neuronal population at 24 h, and 60–70% at 48 h.
All experiments with nicergoline were carried out in
cultures exposed to hAP for 24 h. Nicergoline co-applied
with hAP reduced neuronal toxicity in a concentration-
dependent fashion and was maximally protective (about
60% of neuronal rescue) at 10 AM (Figs. 1a and b).
Nicergoline alone did not affect neuronal viability in a
concentration range of up to 10 AM (not shown). We
compared the action of nicergoline to that of its major
metabolites, MDL and MMDL, which contribute to the
overall therapeutic efficacy of nicergoline in humans [1].
MDL, applied at equimolar concentrations, mimicked the
neuroprotective action of nicergoline, whereas MMDL was
virtually inactive (Figs. 1c and d). As nicergoline is known
to interact with serotonin 5-HT2 and a-1 adrenergic
receptors, specific antagonist were tested to rule out the
possibility that the protective effect of nicergoline was due
to its interaction with these receptors. However, the mixed
5-HT2 receptor antagonist, methysergide (0.1 AM), and the
selective a1-receptor antagonist, prazosin (1 AM), did not
affect hAP toxicity (Fig. 1c).
When applied to cortical neurons challenged with the
apoptotic agent staurosporine (2 AM for 24 h) nicergoline
failed to exert any neuroprotective activity (Table 1).
Searching for a molecular correlate of neuroprotection, we
examined the levels of the pro-apoptotic protein Bax by
immunoblotting. As expected [9,12], a 24-h exposure to
hAP increased the intracellular levels of Bax more than 3-
fold in cortical neurons. This effect was substantially
reduced in cultures treated with 5 AM nicergoline or MDL
(Fig. 2a), whereas MMDL was inactive (not shown).
Neither nicergoline nor its metabolites induced changes in
Bax levels when applied alone (not shown). In contrast,
nicergoline alone increased the expression of the anti-
apoptotic protein Bcl-2 after 24 h of exposure. This is
consistent with the evidence that nicergoline activates
protein kinase C (PKC) in brain tissue [6], and that activated
PKC induces the expression of the anti-apoptotic protein,
Bcl-2 [25]. Exposure to hAP for 24 h induced a 50%
reduction in Bcl-2 levels. When hAP was combined with
nicergoline, Bcl-2 expression was still higher than in
untreated cultures (Fig. 2b).

Fig. 1. Protection by nicergoline and its metabolite MDL against h-amyloid toxicity in pure cultures of cortical neurons is shown by the MTT assay (a and c) or
by cytofluorometric analysis of pre-diploid DNA (b and d). In b, nicer = nicergoline; In c, nicergoline (nicer), MDL, and MMDL were applied at concentrations
of 5 AM; prazosin (praz) = 1 AM; methysergide (met) = 0.1 AM. Cultures were exposed to hAP25 – 35 for 24 h. Values are expressed as percent of h-amyloid
toxicity in panels a and c, or as percent of apoptotic neurons (neurons with pre-diploid DNA) in panels b and d. Values were calculated from 6 to 9
determinations from 2 to 3 independent experiments. *P < 0.05 (one-way ANOVA + Fisher’s PLSD) vs. hAP alone.
F. Caraci et al. / Brain Research 1047 (2005) 30–37 33
We extended the study to mixed cultures of cortical cells
containing both neurons and astrocytes, as detected by
immunocytochemistry with antibodies for the neuronal
marker, MAP-2, and for the astrocyte marker, GFAP (not
shown). Here, hAP toxicity was assessed by counting the
neurons stained with trypan blue. Using this model, neuro-
toxicity showed a slower kinetics, with a substantial neuronal
death (about 60%) being detected after 72 h of exposure to
hAP. Nicergoline (co-applied with hAP and maintained in
the medium for the following 72 h) completely prevented
Table 1
Effect of nicergoline on neuronal death induced by a 24 h-exposure to 2 AMstaurosporine
Neuronal survival (% of control)
Control 100.0 T 2.8
Staurosporine 71.5 T 4.2*
Nicergoline 102.2 T 4.7
Staurosporine + nicergoline 62.6 T 3.7*
Pure cortical cultures were treated with 5 AM nicergoline and 2 AMstaurosporine for 24 h prior to evaluation of neuronal viability by the MTT
assay.
Values are mean T SE of 8 values obtained in two independent experiments.
* P < 0.01 vs. respective control.
hAP neurotoxicity even at concentrations (3 AM) that
showed only a partial protection in pure neuronal cultures
(Fig. 3). The action of nicergoline was mimicked by MDL,
whereas MMDL showed only a partial neuroprotective
activity (Fig. 3). This suggested that the presence of
astrocytes enabled a full neuroprotective activity of nicer-
goline and MDL. To assess whether nicergoline could have a
direct action on glial cells, we treated pure cultures of rat
cortical astrocytes (GFAP+ cells > 95%) with nicergoline or
its metabolites, and the collected medium was transferred to
cultures of pure cortical neurons challenged with hAP.Experiments were carried out as follows: glial cultures were
exposed to nicergoline, MDL, or MMDL for 24 or 72 h. At
the end of this treatment the medium was replaced with a
fresh medium lacking nicergoline or metabolites, and this
new medium was collected after additional 24 h and
transferred to pure cultures of cortical neurons. This strategy
was adopted to minimize the amount of nicergoline (or
metabolites) present in the glial medium. Addition of
conditioned medium from untreated glial cultures did not
affect hAP toxicity in cultured cortical neurons. Similar
results were obtained by transferring the medium collected
from glial cultures 24 h after a 24-h exposure to nicergoline.
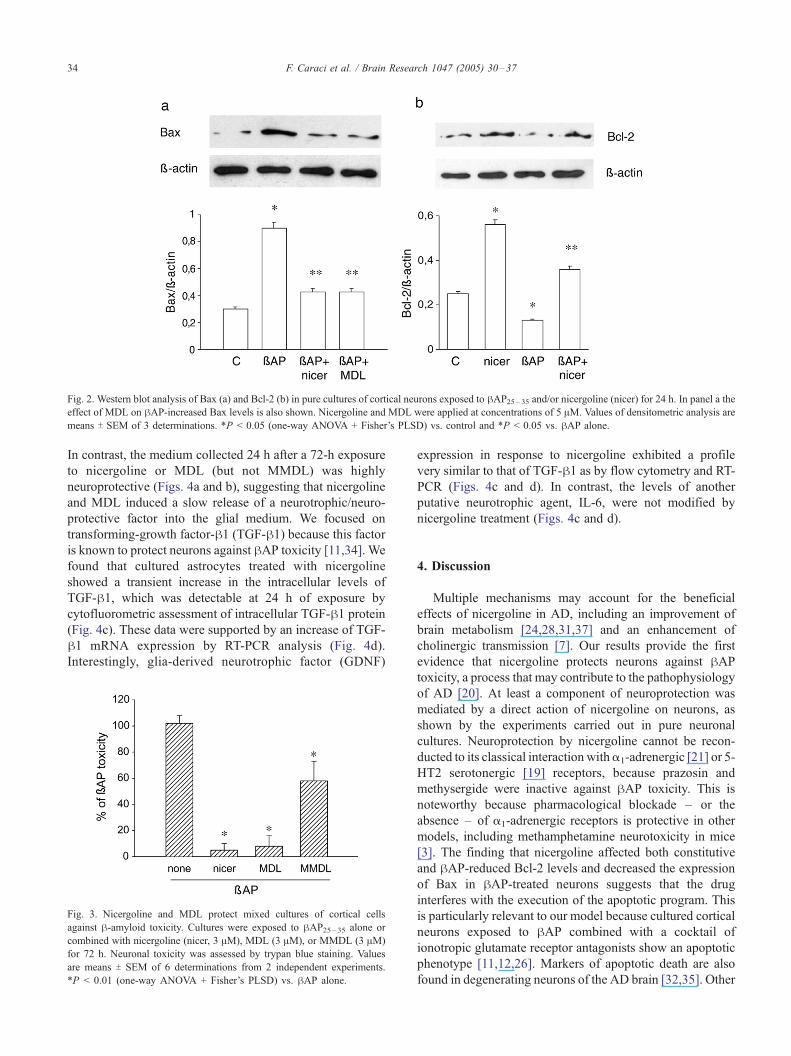
Fig. 2. Western blot analysis of Bax (a) and Bcl-2 (b) in pure cultures of cortical neurons exposed to hAP25 – 35 and/or nicergoline (nicer) for 24 h. In panel a theeffect of MDL on hAP-increased Bax levels is also shown. Nicergoline and MDL were applied at concentrations of 5 AM. Values of densitometric analysis are
means T SEM of 3 determinations. *P < 0.05 (one-way ANOVA + Fisher’s PLSD) vs. control and *P < 0.05 vs. hAP alone.
F. Caraci et al. / Brain Research 1047 (2005) 30–3734
In contrast, the medium collected 24 h after a 72-h exposure
to nicergoline or MDL (but not MMDL) was highly
neuroprotective (Figs. 4a and b), suggesting that nicergoline
and MDL induced a slow release of a neurotrophic/neuro-
protective factor into the glial medium. We focused on
transforming-growth factor-h1 (TGF-h1) because this factoris known to protect neurons against hAP toxicity [11,34]. We
found that cultured astrocytes treated with nicergoline
showed a transient increase in the intracellular levels of
TGF-h1, which was detectable at 24 h of exposure by
cytofluorometric assessment of intracellular TGF-h1 protein
(Fig. 4c). These data were supported by an increase of TGF-
h1 mRNA expression by RT-PCR analysis (Fig. 4d).
Interestingly, glia-derived neurotrophic factor (GDNF)
Fig. 3. Nicergoline and MDL protect mixed cultures of cortical cells
against h-amyloid toxicity. Cultures were exposed to hAP25 – 35 alone or
combined with nicergoline (nicer, 3 AM), MDL (3 AM), or MMDL (3 AM)
for 72 h. Neuronal toxicity was assessed by trypan blue staining. Values
are means T SEM of 6 determinations from 2 independent experiments.
*P < 0.01 (one-way ANOVA + Fisher’s PLSD) vs. hAP alone.
expression in response to nicergoline exhibited a profile
very similar to that of TGF-h1 as by flow cytometry and RT-
PCR (Figs. 4c and d). In contrast, the levels of another
putative neurotrophic agent, IL-6, were not modified by
nicergoline treatment (Figs. 4c and d).
4. Discussion
Multiple mechanisms may account for the beneficial
effects of nicergoline in AD, including an improvement of
brain metabolism [24,28,31,37] and an enhancement of
cholinergic transmission [7]. Our results provide the first
evidence that nicergoline protects neurons against hAPtoxicity, a process that may contribute to the pathophysiology
of AD [20]. At least a component of neuroprotection was
mediated by a direct action of nicergoline on neurons, as
shown by the experiments carried out in pure neuronal
cultures. Neuroprotection by nicergoline cannot be recon-
ducted to its classical interaction witha1-adrenergic [21] or 5-
HT2 serotonergic [19] receptors, because prazosin and
methysergide were inactive against hAP toxicity. This is
noteworthy because pharmacological blockade – or the
absence – of a1-adrenergic receptors is protective in other
models, including methamphetamine neurotoxicity in mice
[3]. The finding that nicergoline affected both constitutive
and hAP-reduced Bcl-2 levels and decreased the expression
of Bax in hAP-treated neurons suggests that the drug
interferes with the execution of the apoptotic program. This
is particularly relevant to our model because cultured cortical
neurons exposed to hAP combined with a cocktail of
ionotropic glutamate receptor antagonists show an apoptotic
phenotype [11,12,26]. Markers of apoptotic death are also
found in degenerating neurons of the AD brain [32,35]. Other

Fig. 4. (a and b) The conditioned medium collected from cultured astrocytes exposed to nicergoline or MDL for >24 h protects cultured neurons against
h-amyloid toxicity. Cultured astrocytes were treated with drugs for 24–96 h and then extensively washed. The medium was collected 24 h after drug
withdrawal. Values are means T SEM of 6–9 determinations from 2 to 3 independent experiments. *P < 0.05 (one-way ANOVA + Fisher’s PLSD) vs. hAPalone. Intracellular levels of TGF-h1, GDNF, or IL-6 in cultured astrocytes exposed to 5 AM nicergoline are shown in panel c, where values are means T SEM
of 3 determinations. RT-PCR of TGF-h1, GDNF, and IL-6 in cultured astrocytes treated with hAP for 24 h is shown in panel d. *P < 0.05 (one-way ANOVA +
Fisher’s PLSD) vs. control values.
F. Caraci et al. / Brain Research 1047 (2005) 30–37 35
studies have shown a protective role of nicergoline against
apoptosis in PC12 cells deprived of nerve growth factor [5],
GT1-7 cells depleted of glutathione [33], or B50 cells
challenged with hydrogen peroxide [23]. An antioxidant
activity of nicergoline may contribute to its protective effects
because perturbation of Ca2+ homeostasis and oxidative
stress are central to the neurotoxic cascade induced by hAP inneurons [2,18,27]. Nicergoline is known to inhibit lipid
peroxidation and free radical generation and reduces auto-
oxidation processes in brain tissue homogenates [36].
However, the evidence that both MDL and MMDL share
the antioxidant properties of nicergoline [38] – whereas only
MDL was substantially active in our model – suggests that
additional mechanisms are involved in the neuroprotective
activity of nicergoline against hAP toxicity. Among these,
activation of PKC [10] might be particularly relevant because
PKC iota protects neural cells against apoptosis induced by
hAP [39], and activated PKC enhances Bcl-2 expression in
neuroblastoma cells [25]. Studies with isoform-selective
inhibitors of PKC are necessary to address this issue. We
were particularly intrigued by the finding that nicergoline
showed a higher neuroprotective activity in mixed cortical
cultures, and that the medium collected from cultured
astrocytes exposed to nicergoline or MDL was protective
when transferred to cultured neurons challenged with hAP. Itis unlikely that the conditioned glial medium contained an
amount of nicergoline sufficient to account for neuroprotec-
tion because (i) astrocytes were extensively washed 24 h
before collecting the medium, and (ii) we observed protection
when astrocytes were exposed to nicergoline for 72 h, but not
for 24 h, although nicergoline levels in the medium are
expected to be higher after 24 than 72 h. The long exposure
time required for the protective activity of the glial medium
suggested that nicergoline or MDL induced the synthesis and
release of neurotrophic/neuroprotective factors in cultured
astrocytes. We focused on TGF-h and GDNF because both
factors are known to protect neurons against hAP toxicity
[11,16,29,34] and activation of glial metabotropic glutamate
receptors protects neighbor neurons against excitotoxic death
through the formation of TGF-h1 and -h2 [4,14]. Exposure tonicergoline was found to increase the intracellular levels of
both TGF-h1 and GDNF. In our experience [4,14,34] an
increase in intracellular levels of at least TGF-h is always
associated with its secretion in cultured astrocytes. We could

F. Caraci et al. / Brain Research 1047 (2005) 30–3736
not detect extracellular TGF-h or GDNF because levels were
too low to be quantified by ELISA. Although the relative
contribution of TGF-h or GDNF to the overall neuro-
protection remains to be determined, this finding suggests
an entirely novel mechanism of action of nicergoline based
on glial–neuronal interaction. Interestingly, an increased
production of TGF-h1 in astrocytes has recently been
implicated in the neuroprotective activity of h-estradiolagainst hAP toxicity [34]. More intriguing is the recent
observation that GDNF protects neurons against hAP by
reducing the activation of ERK kinases [16], an intracellular
pathway that we have shown is involved in the induction of
cell cycle by hAP [15].
In conclusion, the evidence that nicergoline protects
neurons against hAP toxicity raises the interesting possi-
bility that the drug slows the progression of AD by
limiting the death of neurons surrounded by amyloid
deposits.
References
[1] F. Arcamone, A.G. Glasser, J. Grafnetterova, A. Minghetti, V.
Nicolella, Studies on the metabolism of ergoline derivatives. Metab-
olism of nicergoline in man and in animals, Biochem. Pharmacol. 21
(1972) 2205–2213.
[2] S.W. Barger, D. Horster, K. Furukawa, Y. Goodman, J. Krieglstein,
M.P. Mattson, Tumor necrosis factors alpha and beta protect neurons
against amyloid beta-peptide toxicity: evidence for involvement of a
kappa B-binding factor and attenuation of peroxide and Ca2+
accumulation, Proc. Natl. Acad. Sci. U. S. A. 92 (1995) 9328–9332.
[3] G. Battaglia, F. Fornai, C.L. Busceti, G. Lembo, F. Nicoletti, A. De
Blasi, Alpha-1B adrenergic receptor knockout mice are protected
against methamphetamine toxicity, J. Neurochem. 86 (2003) 413–421.
[4] V. Bruno, G. Battaglia, G. Casabona, A. Copani, F. Caciagli, F.
Nicoletti, Neuroprotection by glial metabotropic glutamate receptors is
mediated by transforming growth factor-beta, J. Neurosci. 18 (1998)
9594–9600.
[5] P.L. Canonico, M.A. Sortino, N. Carfagna, S. Cavallaro, F. Pampa-
rana, K. Annoni, E. Wong, C. Post, Pharmacological basis for the
clinical effects of nicergoline in dementia, Geriatria 5 (Suppl VIII)
(1998) 24–48.
[6] A. Caputi, M. Di Luca, L. Pastorino, F. Colciaghi, N. Carfagna, E.
Wong, C. Post, F. Cattabeni, Nicergoline and its metabolite induce
translocation of PKC isoforms in selective rat brain areas, Neurosci.
Res. Commun. 23 (1998) 159–167.
[7] N. Carfagna, A. Di Clemente, S. Cavanus, D. Damiani, M. Gerna, P.
Salmoiraghi, B. Cattaneo, C. Post, Modulation of hippocampal ACh
release by chronic nicergoline treatment in freely moving young and
aged rats, Neurosci. Lett. 197 (1995) 195–198.
[8] N. Carfagna, S. Cavanus, D. Damiani, P. Salmoiraghi, R. Fariello, C.
Post, Modulation of phosphoinositide turnover by chronic nicergoline
in rat brain, Neurosci. Lett. 209 (1996) 189–192.
[9] A. Caricasole, A. Copani, F. Caraci, E. Aronica, A.J. Rozemuller, A.
Caruso, M. Storto, G. Gaviraghi, G.C. Terstappen, F. Nicoletti,
Induction of Dickkopf-1, a negative modulator of the Wnt pathway,
is associated with neuronal degeneration in Alzheimer’s brain,
J. Neurosci. 24 (2004) 6021–6027.
[10] A. Cedano-Minguez, L. Bonecchi, B. Winblad, C. Post, E.H.F. Wong,
R.F. Cowborn, L. Benfatti, Nicergoline stimulates protein kinase C-
mediated alpha-secretase processing of the amyloid precursor protein
in cultured human neuroblastoma SH-SY5Y cells, Neurochem. Int. 35
(1999) 307–315.
[11] A. Copani, F. Condorelli, A. Caruso, C. Vancheri, A. Sala, A.M.
Giuffrida Stella, P.L. Canonico, F. Nicoletti, M.A. Sortino, Mitotic
signaling by beta-amyloid causes neuronal death, FASEB J. 13 (1999)
2225–2234.
[12] A. Copani, M.A. Sortino, A. Caricasole, S. Chiechio, M. Chisari, G.
Battaglia, A.M. Giuffrida-Stella, C. Vancheri, F. Nicoletti, Erratic
expression of DNA polymerases by beta-amyloid causes neuronal
death, FASEB J. 16 (2002) 2006–2008.
[13] T.H. Crook, Nicergoline: parallel evolution of clinical trial method-
ology and drug development in dementias, Dement. Geriatr. Cogn.
Disord. 8 (Suppl 1) (1998) 22–26.
[14] M. D’Onofrio, L. Cuomo, G. Battaglia, R.T. Ngomba, M. Storto, A.E.
Kingston, F. Orzi, A. De Blasi, P. Di Iorio, F. Nicoletti, V. Bruno,
Neuroprotection mediated by glial group-II metabotropic glutamate reQ
ceptors requires the activation of the MAP kinase and the phospha-
tidylinositol-3-kinase pathways, J. Neurochem. 78 (2001) 435–445.
[15] G. Frasca, S. Chiechio, C. Vancheri, F. Nicoletti, A. Copani, M.A.
Sortino, Beta-amyloid-activated cell cycle in SH-SY5Y neuroblas-
toma cells: correlation with the MAP kinase pathway, J. Mol.
Neurosci. 22 (2004) 231–236.
[16] O. Ghribi, M.M. Herman, P. Pramoonjago, N.K. Spaulding, J.
Savory, GDNF regulates the A beta-induced endoplasmic reticulum
stress response in rabbit hippocampus by inhibiting the activation of
gadd 153 and the JNK and ERK kinases, Neurobiol. Dis. 16 (2004)
417–427.
[17] L. Giardino, A. Giuliani, A. Battaglia, N. Carfagna, L. Aloe, L. Calza’,
Neuroprotection and aging of the cholinergic system: a role for the
ergoline derivative nicergoline (Sermion), Neuroscience 109 (2002)
487–497.
[18] Q. Guo, B.L. Sopher, K. Furukawa, D.G. Pham, N. Robinson,
G.M. Martin, M.P. Mattson, Alzheimer’s presenilin mutation
sensitizes neural cells to apoptosis induced by trophic factor
withdrawal and amyloid beta-peptide: involvement of calcium and
oxyradicals, J. Neurosci. 17 (1997) 4212–4222.
[19] J.D. Hagen, P.A. Pierce, S.J. Peroutka, Differential binding of ergot
compounds to human versus rat 5-HT2 cortical receptors, Biol.
Signals 3 (1994) 223–229.
[20] J. Hardy, D.J. Selkoe, The amyloid hypothesis of Alzheimer’s disease:
progress and problems on the road to therapeutics, Science 297 (2002)
353–356.
[21] C. Heitz, J.J. Descombes, R.C. Miller, J.C. Stoclet, Alpha-adrenocep-
tor antagonistic and calcium antagonistic effects of nicergoline in the
rat isolated aorta, Eur. J. Pharmacol. 123 (1986) 279–285.
[22] W.M. Herrmann, K. Stephan, K. Gaede, M.A. Apeceche, A multi-
center randomized double-blind study on the efficacy and safety of
nicergoline in patients with multi-infarct dementia, Dement. Geriatr.
Cogn. Disord. 8 (1987) 9–17.
[23] E. Iwata, I. Miyazaki, M. Asanuma, A. Iida, N. Ogawa, Protective
effects of nicergoline against hydrogen peroxide toxicity in rat
neuronal cell line, Neurosci. Lett. 251 (1998) 49–52.
[24] M. Le Poncin-Lafitte, C. Grosdemouge, D. Duterte, J.R. Rapin,
Simultaneous study of haemodynamic, metabolic and behavioural
sequelae in a model of cerebral ischaemia in aged rats: effects of
nicergoline, Gerontology 30 (1984) 109–119.
[25] A. Lombet, V. Zujovic, M. Kandouz, C. Billardon, S. Carvajal-
Gonzalez, A. Gompel, W. Rostene, Resistance to induced apoptosis in
the human neuroblastoma cell line SK-N-SH in relation to neuronal
differentiation. Role of Bcl-2 protein family, Eur. J. Biochem. 268
(2001) 1352–1362.
[26] D.T. Loo, A. Copani, C.J. Pike, E.R. Whittemore, A.J. Walencewicz,
C.W. Cotman, Apoptosis is induced by beta-amyloid in cultured
central nervous system neurons, Proc. Natl. Acad. Sci. U. S. A. 90
(1993) 7951–7955.
[27] R.J. Mark, M.A. Lovell, W.R. Markesbery, K. Uchida, M.P. Mattson,
A role for 4-hydroxynonenal, an aldehydic product of lipid perox-
idation, in disruption of ion homeostasis and neuronal death induced
by amyloid beta-peptide, J. Neurochem. 68 (1997) 255–264.

F. Caraci et al. / Brain Research 1047 (2005) 30–37 37
[28] A. Miccheli, C. Puccetti, G. Capuani, M.E. Di Cocco, L. Giardino, L.
Calza, A. Battaglia, L. Battistin, F. Conti, [1-13C]Glucose entry in
neuronal and astrocytic intermediary metabolism of aged rats. A study
of the effects of nicergoline treatment by 13C NMR spectroscopy,
Brain Res. 966 (2003) 116–125.
[29] J.H. Prehn, V.P. Bindokas, J. Jordan, M.F. Galindo, G.D. Ghadge, R.P.
Roos, L.H. Boise, C.B. Thompson, S. Krajewski, J.C. Reed, R.J.
Miller, Protective effect of transforming growth factor-beta 1 on beta-
amyloid neurotoxicity in rat hippocampal neurons, Mol. Pharmacol.
49 (1996) 319–328.
[30] B. Saletu, E. Paulus, L. Linzmayer, P. Anderer, H.V. Semlitsch, J.
Grunberger, L. Wicke, A. Neuhold, I. Podreka, Nicergoline in senile
dementia of Alzheimer type and multi-infarct dementia: a double-
blind, placebo-controlled, clinical and EEG/ERP mapping study,
Psychopharmacology (Berl) 117 (1995) 385–395.
[31] K. Shintomi, Pharmacological study of nicergoline; effects on
regional cerebral blood flows and arterial carbon dioxide and oxygen
pressure and pH in rats under cyanide-induced histotoxic anoxia,
Arzheim.-Forsch. Drug Res. 41 (1991) 885–890.
[32] G. Smale, N.R. Nichols, D.R. Brady, C.E. Finch, W.E. Horton Jr,
Evidence for apoptotic cell death in Alzheimer’s disease, Exp. Neurol.
133 (1995) 225–230.
[33] M.A. Sortino, A. Battaglia, F. Pamparana, N. Carfagna, C. Post, P.L.
Canonico, Neuroprotective effects of nicergoline in immortalized
neurons, Eur. J. Pharmacol. 368 (1999) 285–290.
[34] M.A. Sortino, M. Chisari, S. Merlo, C. Vancheri, M. Caruso, F.
Nicoletti, P.L. Canonico, A. Copani, Glia mediates the neuroprotective
action of estradiol on beta-amyloid-induced neuronal death, Endo-
crinology 145 (2004) 5080–5086.
[35] J.H. Su, A.J. Anderson, B.J. Cummings, C.W. Cotman, Immunohis-
tochemical evidence for apoptosis in Alzheimer’s disease, Neuro-
Report 5 (1994) 2529–2533.
[36] M. Tanaka, T. Yoshida, K. Okamoto, S. Hirai, Antioxidant properties
of nicergoline; inhibition of brain auto-oxidation and superoxide
production of neutrophils in rats, Neurosci. Lett. 248 (1998) 68–72.
[37] M. Vairetti, F. Feletti, A. Battaglia, F. Pamparana, P.L. Canonico, P.
Richelmi, F. Berte, Haloperidol-induced changes in glutathione and
energy metabolism: effect of nicergoline, Eur. J. Pharmacol. 367
(1999) 67–72.
[38] M. Vairetti, A. Battaglia, N. Carfagna, P.L. Canonico, F. Berte, P.
Richelmi, Antioxidant properties of MDL and MMDL, two nicergo-
line metabolites, during chronic administration of haloperidol, Eur. J.
Pharmacol. 453 (2002) 69–73.
[39] J. Xie, Q. Guo, H. Zhu, M.W. Wooten, M.P. Mattson, Protein kinase C
iota protects neural cells against apoptosis induced by amyloid beta-
peptide, Brain Res. Mol. Brain Res. 82 (2000) 107–113.
[40] B. Winblad, N. Carfagna, L. Bonura, B.M. Rossini, E.H.F. Wong,
A. Battaglia, Nicergoline in dementia. A review of its pharmaco-
logical properties and therapeutic potential, CNS Drugs 14 (2000)
1–22.
Related Documents











